Embed Size (px)
DESCRIPTION
V11 Splice varitants in TRP channels. Review of lecture V10 .. Introduction of TRP channels. RNA splicing. Splicing is a modification of an RNA after transcription, in which introns are removed and exons are joined. - PowerPoint PPT Presentation
Citation preview

SS 2008 lecture 11
Biological Sequence Analysis1
V11 Splice varitants in TRP channels
Review of lecture V10 .. Introduction of TRP channels

SS 2008 lecture 11
Biological Sequence Analysis2
RNA splicing
Simple illustration of exons and introns in pre-mRNA and the formation of mature mRNA by splicing. The UTRs are non-coding parts of exons at the ends of the mRNA.
www.wikipedia.org
Splicing is a modification of an RNA after transcription, in which introns are
removed and exons are joined.
This is needed for the typical eukaryotic messenger RNA before it can be used
to produce a correct protein through translation.
For many eukaryotic introns, splicing is done in a series of reactions which are
catalyzed by the spliceosome, a complex of small nuclear ribonucleoproteins
(snRNPs), but there are also self-splicing introns.

SS 2008 lecture 11
Biological Sequence Analysis3
alternative splicing
www.wikipedia.org
Alternative splicing is the RNA splicing variation mechanism in which the
exons of the primary gene transcript, the pre-mRNA, are separated and
reconnected so as to produce alternative RNA arrangements.
Via translation, these then give different (isoform) proteins.
In this way, alternative splicing uses genetic expression to facilitate the
synthesis of a greater variety of proteins.
Alternative splicing is of great importance to genetics - it invalidates the old
"one-gene-one-protein" hypothesis. External information is needed in order to
decide which polypeptide is produced, given a DNA sequence and pre-mRNA.
The amount of alternative splicing is comparable, with no large differences
between humans and other animals.
The "record-holder" for alternative splicing is a Drosophila gene called Dscam,
which has 38 016 splice variants.

SS 2008 lecture 11
Biological Sequence Analysis4
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
TRPV4 channels
The non-selective cation channel TRPV4 is a member of the transient
receptor potential (TRP) family of channels.
TRPV4 shows multiple modes of activation and regulatory sites, enabling it to
respond to various stimuli, including osmotic cell swelling,
mechanical stress,
heat,
acidic pH,
endogenous ligands,
high viscous solutions, and
synthetic agonists such as 4-phorbol 12,13-didecanoate.
TRPV4 mRNA is expressed in a broad range of tissues, although functional tests
have only been carried out in a few:
endothelial, epithelial, smooth muscle, keratinocytes, and DRG neurons.

SS 2008 lecture 11
Biological Sequence Analysis5
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Topology of TRPV4 channels
The general topology of a TRP subunit consists of 6 predicted TM domains with a
putative pore loop between TMD5 and TMD6 and intracellular N- and C-terminal
regions of variable length, the former containing multiple ankyrin (ANK) repeats
in the TRPC, TRPA, TRPN, and TRPV subfamilies.
ANK repeats are modular protein interaction domains, each composed by 33
amino acids with a highly conserved helix turn helix motif that determines ist
interaction properties.
Functional TRP channels are supposed to result following the assembly of 4 TRP
subunits.
The rules governing subunit assembly and the protein domains implied in this
oligomerization process are just starting to emerge and may involve the cytosolic
N-terminal region, the ANK domains, transmembrane domains, and the
cytoplasmic C terminus.

SS 2008 lecture 11
Biological Sequence Analysis6
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Cloning of TRPV4 variants from human airway epithelial cells
A reverse transcriptase-PCR-based cloning process identified 5 variants of the
TRPV4 channel in human tracheal epithelial cells.
2 of the cloned cDNAs corresponded to the already described - TRPV4 isoform A (fulllength cDNA) and - TRPV4 isoform B (lacking exon number 7, 384–444 amino acids).
We also identified 3 new splice variants affecting the cytoplasmic N-terminal region.
TRPV4-C lacks exon 5 (237–284 amino acids),
TRPV4-D presents a short deletion inside exon 2 (27–61 amino acids), and TRPV4-
E (237–284 and 384–444 amino acids) is produced by a double alternative splicing
lacking exons 5 and 7.

SS 2008 lecture 11
Biological Sequence Analysis7
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Different splice variants of TRPV4
A, schematic diagram showing the
intracellular N-terminal region of
the human TRPV4 channel
(amino acids 1–471).
Exons and the corresponding
amino acids lost in each TRPV4
isoform are indicated by numbers.

SS 2008 lecture 11
Biological Sequence Analysis8
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Functional analysis of TRPV4 variants: intracellular [Ca2+]
The TRPV4-A channel responds to
a wide variety of stimuli.
Here, HeLa cells were transiently
transfected and intracellular Calcium
concentration was determined via
Fura-2 ratios as reponse to 3 well
known activators of TRPV4-A: 30%
hypotonic solution, 1 M 4-PDD, or
10 M arachidonic acid
Only TRPV4-A and TRPV4-D
show channel activity.

SS 2008 lecture 11
Biological Sequence Analysis9
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
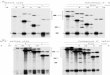
TRPV4-A and D produce functional channels
TRPV4-A and TRPV4-D isoforms
produce functional channels with
similar properties when expressed
in HEK-293 cells.
A, current traces obtained from
TRPV4-A and TRPV4-D-expressing
HEK-293 cells at the indicated
voltages in the presence of 1M
4-PDD. Dashed lines indicate the
zero current level.
B, I–V relationship of 4-PDD-
activated TRPV4-A (open circle) and
TRPV4-D (closed circle) channels in
inside-out patches.

SS 2008 lecture 11
Biological Sequence Analysis10
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Retention in ER
Co-localization experiments (not shown):
TRPV4-B, C and E are trapped in the ER and not translocated to the plasma
membrane.

SS 2008 lecture 11
Biological Sequence Analysis11
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Homomerization of TRPV4 variants
FRET efficiencies determined between
identical CFP- and YFP-fused TRPV4
variants (A–E) transiently cotransfected
in HEK-293 cells.
High FRET efficiencies corresponding to
homomultimer formation could only be
demonstrated for TRPV4-A and TRPV4-
D variants.

SS 2008 lecture 11
Biological Sequence Analysis12
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Heteromerization of TRPV4 variants
B, FRET efficiencies
determined between different
TRPV4 variants showed
heterooligomerization only for A
and D proteins.

SS 2008 lecture 11
Biological Sequence Analysis13
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Summary
This study of oligomerization, localization, and channel activity of human TRPV4
splice variant identified the N-terminal ANK repeats as key molecular
determinants of subunit assembly and subsequent processing of the assembled
channel.
Five TRPV4 variants (TRPV4-A–E) cloned from human airway epithelial cells
were grouped into two classes:
group I: TRPV4-A and TRPV4-D
group II: TRPV4-B, TRPV4-C, and TRPV4-E.
Group I variants are correctly processed and targeted to the plasma membrane
where they form functional channels with similar electrophysiological properties.
Variants from group II, which are lacking parts of the ANK domains are unable to
oligomerize and were retained intracellularly, in the ER.

SS 2008 lecture 11
Biological Sequence Analysis14
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Summary II
Discovery of three important traits of TRPV4 biogenesis.
1) Glycosylation of TRPV4 channel involves ER to Golgi transport with the
corresponding change in the N-linked oligosaccharides from the high mannose type
characteristic of the ER to the complex type characteristic of the Golgi apparatus,
without apparent O-glycosylation.
2) TRPV4-A subunits oligomerize in the ER.
3) Impaired subunit assembly of type II variants is because of the lack of N-terminal
ANK domains and causes protein retention in the ER.

SS 2008 lecture 11
Biological Sequence Analysis15
Arniges et al. J. Biol. Chem. 281, 1580 (2006)
Summary III
Ion channel functional diversity is greatly enlarged by both the presence
of splice variants and heteromerization of different pore-forming and regulatory
subunits. Alternative splicing is a major contributor to protein diversity.
Within the TRP family of ion channels several splice variants have been identified,
some of them resulting in lack of responses to typical stimuli, others modifying the
pore properties, and those exerting dominant negative effects.
Group II TRPV4 splice variants have been identified in two unrelated, human airway
epithelial cell lines. Considering the relevance of TRPV4 channels in epithelial
physiology, a change in the expressed ratio of group I to group II variants, favoring
the later, may modify normal epithelial functioning.
Splicing can be regulated by several stressing stimuli including pH, osmotic, and
temperature shocks, all of them being also activating stimuli of the TRPV4.

SS 2008 lecture 11
Biological Sequence Analysis16
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Next: Identification of TRMP3 splice variants from mouse brain
A, schematic diagram of the mouse Trpm3 gene, comprising 28 exons.

SS 2008 lecture 11
Biological Sequence Analysis17
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
TRPM3 splice variants
C, schematic presentation of TRPM3 with transmembrane domains 1–6, coiled coil
region (cc), and TRP homology domain (Trp).
Novel mouse TRPM3 protein variants shown as thick black lines are compared with
the human variants hTRPM3a–f and hTRPM31325. The numbers of amino acid
residues of each variant are indicated in parentheses.
Starting from residue 156, mouse and human TRPM3 have 97% sequence identity.

SS 2008 lecture 11
Biological Sequence Analysis18
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Pore regions of splice variants
D, putative pore regions of TRPM31 and TRPM32 compared with the
corresponding mouse sequences of TRPM6, TRPM7, TRPV5, and TRPV6.
The 12 additional amino acid residues present in TRPM31 are indicated.
Identical residues are boxed in black, conserved in gray.
An aspartate residue that determines Ca2+ permeation of the TRPV5/TRPV6
pore is marked by an asterisk. Residues proposed to build the selectivity filter of
TRPV6 are underlined.

SS 2008 lecture 11
Biological Sequence Analysis19
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
TRPM3 functions as cation channel
Heterologous expression of TRPM31 induces outwardly rectifying cation
currents inhibited by intracellular Mg2+.
A,current-voltage relationship of a TRPM31-expressing cell in standard Ringer
or NMDG solution within 60 s after establishing the whole cell patch clamp
configuration.

SS 2008 lecture 11
Biological Sequence Analysis20
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Permeability for divalent cations
TRPM31 and TRPM32 display large differences in their relative
permeability ratios for divalent cations.
A, comparison of TRPM31 and TRPM32 currents at 80 mV and 80 mV in
extracellular solutions containing indicated amounts of Ca2+.
B, reversal potential during the experiment shown in panel A.

SS 2008 lecture 11
Biological Sequence Analysis21
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Identification of TRMP3 variants from mouse brain
C and D, statistical analysis of reversal
potential measurements in experiments
similar to that shown in panel B during the
application of solutions containing the
indicated concentration of Ca2+ (C) or Mg2+
(D) as the only permeable ion.
Continuous thin lines show the expected
reversal potential calculated from
Goldman-Hodgkin-Katz theory for the
indicated relative permeability ratios.
Each point represents the mean of 3–15
independent measurements (at a divalent
concentration of 10 mM p < 0.001,
otherwise at least p < 0.05).

SS 2008 lecture 11
Biological Sequence Analysis22
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Effect of extracellular cations
Inhibition of TRPM3-dependent currents
by extracellular cations.
A, comparison of TRPM31 and TRPM32
currents at 80 mV and 80 mV in extra-
cellular solutions containing indicated
amounts of Na+.
Outward currents through TRPM31 are
unaffected by extracellular Na+, whereas
outward currents through TRPM32 are
inhibited in a dose-dependent manner by
these ions.
B, statistical analysis of recordings with
varying concentrations of Na+, K+, Ca2+, and
Mg2+.
TRPM32 is inhibited byall cations tested on the extra-cellular side.

SS 2008 lecture 11
Biological Sequence Analysis23
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Summary
Alternative Splicing Switches the Ion Selectivity of TRPM3 Channels—
The selectivity of ion channels is thought to be determined by the geometry and
charge distribution of the selectivity filter, usually envisioned as the narrowest part
of the channel pore.
Typically, all members of an ion channel family, such as voltage-gated Na+, K+, or
Ca2+ channels, share common ionic selectivities.

SS 2008 lecture 11
Biological Sequence Analysis24
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Summary II
The TRP family of ion channels is already somewhat unusual in this respect as it
encompasses members with quite diverging cationic selectivity profiles.
The Trpm3 gene adds extra complexity to this picture, because two channels can
be expressed from this gene with entirely different ionic selectivities. One
channel, TRPM31, preferentially conducts monovalent cation influx, whereas
TRPM32 strongly favors divalent entry.
In vivo, such a change in ionic selectivity must be expected to have considerable
consequences for the function of the channel and the physiology of the cell that
expresses it.

SS 2008 lecture 11
Biological Sequence Analysis25
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Summary III
Locating the Ion-conducting Pore in TRPM Channels—The switch of ionic
selectivity in TRPM3 variants is brought about by removing a short stretch of 12
amino acid residues and exchanging 1 further residue within the linker domain
between the presumed fifth and sixth transmembrane regions (Fig. 1B).
The differences in ion selectivity seen for the TRPM3 splice variants strongly
indicate that this linker domain constitutes the pore of TRPM3.
Although this domain could already be suspected to be the ion-conducting pore,
due to direct evidence obtained for TRPV1, TRPV4, TRPV5, and TRPV6
channels, this prediction has not been confirmed up to now for any member of the
TRPM subfamily.

SS 2008 lecture 11
Biological Sequence Analysis26
Oberwinkler et al. J. Biol. Chem. 280, 22540 (2005)
Summary IV
Compared with the presumed pore regions of other members of the TRP family,
the pore loop of TRPM3 is considerably longer by 8 (TRPM32) and 20
(TRPM31) additional amino acid residues.
The domains that build the proposed selectivity filter of the Ca2+-selective
TRPV5/V6 channels are conserved in TRPM3 proteins. The splicing within the
TRPM3 channel pore introduces additional, positively charged amino acid
residues into this domain.
This might decrease the Ca2+ permeability of TRPM31 compared with
TRPM32, perhaps simply because of increased electrostatic repulsion.
Block of TRPM3 Channels by Intra- and Extracellular Cations — Both TRPM31
and TRPM32 are regulated by physiological concentrations of intracellular Mg2+,
similar to related members of the TRPM family such as TRPM6 and TRPM7.