Embed Size (px)
Citation preview

UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl)
UvA-DARE (Digital Academic Repository)
Inflammatory response in obstructive jaundice and peritonitis
Sewnath, M.E.
Link to publication
Citation for published version (APA):Sewnath, M. E. (2003). Inflammatory response in obstructive jaundice and peritonitis.
General rightsIt is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s),other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons).
Disclaimer/Complaints regulationsIf you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, statingyour reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Askthe Library: https://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam,The Netherlands. You will be contacted as soon as possible.
Download date: 25 Jun 2020

Interleukin- 11 receptor type I gene-deficient bile ductt ligated mice are partiall y protected
againstt endotoxin
Miguell E. Sewnath1, Tom van der Poll2'3, Fiebo J.W. ten Kate4, Cornellss J.F. van Noorden\ and Dirk J. Gouma'
Departmentss of Surgery, "experimental Internal Medicine. 'Infectious Diseases. Tropical
Medicinee and AIDS. 4Pathology. and 'Cell Biology and Histology. Academic Medical
Center.. University of Amsterdam. Amsterdam. The Netherlands
HEMATOLOGYHEMATOLOGY2002;2002; 35:149-15H

EnüohhxemiaEnüohhxemia in IL-IR-/- cholestatic mice
ABSTRACT T
Cholestaticc liver injtirv is associated with an increased susceptibility toward endotoxin-induced toxicity To determine
thee role of inierleukin (II.)- ! herein, exlrahepahc cholestasis was induced hv bile dud ligation (bdl) in II.-1 receptor type
1-gcnee deficient (II.-1R- -l mice, which arc unresponsive to II.-l u and 11.-1(1 and normal II -1 R - mice. Bdl elicited
increasess m hepatic II -lu and II .-1 r m R \ .\ and protein Hepatocellular injury at 2 weeks after bdl was similar m II -1R-
/-- and I1.-IR+/+ mice as demonstrated bv clinical chemistry and hislopathologv. Administration of endotoxin to
cholestaticc mice at 2 weeks alter 1x11 was associated with enhanced cytokine release, more severe liver damage, and
occurrencee of death when compared io sham operated mice. Imdotoxin effects m sham operated IL-I R-/- and I1.-1R+/+
micee were largelv similar, but cholestatic 1L-1R-/- mice were better protected against toxic effects of endotoxin, as
reflectedd bv low ered cvtokine release, less profound liver injury, and reduced mortality. These data indicate that II -1 u
andd II -1 b' are produced in the liver alter bdl. but that these cytokines do not play a significant role in cholestatic liver
damage:: however, endogenous II.-I aclivitv is an important denominator of enhanced endotoxin sensitivitv thai is
observedd during cholestasis induced hv bdl.
INTRODUCTIO N N
Surgeryy in jaundiced patients with periampullary tumors carries an increased risk of
postoperativee complications.' Whereas in experienced centers, postoperative mortality has been
reducedd from 20 to 5c/c morbidity remains as high as 5 0 1 ." Most complications have a septic
etiologyy and are considered to be related with translocation of endotoxin from the intestinal
lumenn into the portal and systemic circulation where an inflammatory cascade is triggered. '
Potentiall causes of translocation of endotoxin include lack of bile salts in the intestinal lumen,
resultingg in increased bacterial translocation,'1' decreased function of the resident macrophages
off the liver, the Kupffer cells, leading to inadequate interception of endotoxin.'1"* and changes in
plasmaa concentrations of lipoproteins which bind endotoxin.'' In addition, cholestasis is
associatedd with enhanced susceptibility toward endotoxin-induced toxicity.
Endotoxinn exerts its biological effects by activating immunocompetent cells such as monocytes
andd macrophages. After exposure to endotoxin, these cells readily release various inflammatory
mediatorss such as the cytokines tumor necrosis factor (TNF) and interleukin-1 (IL-1 ). which
leadss to an inflammatory response. The Kupffer cells are also capable of releasing these
cytokiness in response to endotoxin.",J IL-l a and IL-1 p are pleiotropic proinflammatory
cytokiness that are considered to play a proximal role in initiation and regulation of the
inflammatoryy cascade triggered in response to endotoxins or bacteremia. " ' Both IL-l a and IL-
11 p can interact with two specific cell surface receptors, the type I and type II IL-1 receptor (IL-
1R).. of which only the former is involved in intracellular signaling of IL-1 effects \ As a
consequence,, mice that lack the gene encoding the type I IL-1R (IL-1R-/- mice) can not respond
to IL - laorr IL-ip .
Thee role of IL-I in pathophysiological consequences of cholestasis and the ensuing increased
100 0

ChapterChapter 6
susceptibilityy to septic complications is still not understood. Therefore, we used the model of extrapehaticc cholestasis induced by bile duct ligation (bdl) in IL-1R-/- mice to study whether (a) IL-11 contributes to hepatic pathology associated with cholestasis, and (b) endogenous IL-1 is involvedd in the enhanced susceptibility to endotoxin that accompanies cholestasis.
MATERIAL SS AND METHOD S
Animal s s
IL-11 R-/- mice hackcrossed 6 times to a C57BL/6 background (kindh provided by Immunex. Seattle. WA) and their
normall C57BI/6 wild type counterpart (Il.- I R+/+. Harlan. Zeist. The Netherlands) were used. Fach experimental group
consistedd of S-12 mice! sex and age matched: X-10 weeks old) per time point. IL-IR-/ - mice were normal in size, wt and
fertility ,, and displayed no abnormalities in leukocyte subsets.: In addition. IL-IR-/ - mice have normal serum Ig levels
andd generate normal levels of primary and sccondar) Ahs. All mice were housed in the same animal room, maintained
underr a 1 2-hour light/dark cycle, and provided regular mouse chow (SRM-A; Hope Partus. Woerden. The Netherlands)
andd water ail lihitum.
Surgicall procedures
Thee Institutional Animal Care and L'se Committee of the Academic Medical Center, Amsterdam, the Netherlands
approvedd the experiments. After 5 days of acclimatization, surgery was performed under sterile conditions (t = 0). Mice
weree divided random!) into a sham-operated (shamI control group and a bile duct ligated (bdl) group. Thev were
anaesthetizedd by intraperitoneal injection of 0.07 ml/ kg FFM mixture (Fentanyl {0.3 15 mg/ml). Fluanisone (10 mg/mll
(bothh Janssen. Beersen. Belgium I. and Midazolam (5 mg/ml) (Roche. Mijdrecht. The Netherlands)). A midabdominal
incisionn was made, and the common bile duct was ligated and divided as previous!) described." In mice undergoing sham
operation,, the bile duct was identified and ligatures were placed (but not tiedl and removed again. Postoperative
analgesiaa was achieved with subcutaneous injection of 0.1 |j,L/kg Temgesic (Rcckiu. Kingston-L'pon-Hill. L'nited
Kingdom). .
Material s s
Endotoxin,, lipopolysaccharide E. coli serotype O l l l : B4. was purchased as a lyophilized powder from Sigma (St. Louis.
MO),, resuspended in a 0.5 ml sterile pyrogen-free isotonic saline and injected intraperitoneal!) (4 mg/kg). Control mice
receivedd sterile pyrogen-lree isotonic saline.
Bloodd sampling and organ preparation
Att the designated time points, mice were anesthetized h_v intraperitoneal FFM (0.07 ml/kg) anesthesia and blood was
drawnn from the inferior caval vein into a sterile syringe. One part of the blood was transferred to tubes containing
heparin,, and plasma was prepared by centri ('ligation (3000 i>. 20 min. 4CC) tor measurement of levels of cytokines, total
bilirubin,, aspartate aminotransferase (AST), alanine aminotransferase (ALT) , alkaline phosphatase, and -/-glutamyl
transferasee (y-GT). All samples were aliquoted and stored immediately at - 2 0C until analysis.
Liverss were immediately dissected and homogenized at 4 'C in 4 volumes of sterile saline using a tissue homoiieni/er
101 1

EndotoxemiaEndotoxemia in IL-IR-/- cholestatic mice
iBiospec.. Mar t le t i l k \ L'K). I. ivcr homogcnales were diluted 1:2 in lysis huffci containing 300 m \ l NaCI. 30 m M Tris.
22 m.M M g C k \'< Tr i ton X-100. and pepstatm A. leupeptin and aproptinin ILII I 20 ng/ml : pH 7.4: G ihco /BRL. Ber l in.
Germainn ) and incubated at 4 C lor 30 min. Homogcnatcs were ccnlril 'uged at 190 \ .e at 4 "C lor 10 min to remove
eelll debris. Litter which supeniaianis were stored at -21) C'.
K N AA preparat io n ant i revers e transcriptas e PCR
Tissuee samples were homogenized in I nil of T r i / o l i ( ' n lvo) according to the manufacturer's instructions and total RNA
wass isolated using chloroform extraction and isopropanol precipitation. R N A was dissolved in dielhy l-pyrocarbonaat-
treatedd water and quanti f ied by spectrophotometry. c D N A was synthesized by mix ing 2 kig o\ R N A wi th 0.5 | iü
o l igo idT)) (Gibco i . and h\ incubating the solti l ion dotal \o lume i 2 CLLI tor 10 min al 72 C. Subsequently. S \il. o f a
solutionn containing 5x first strand buffer (Gibco). 1.25 m M each o f d N T P s (Amersham Pharmacia. Amersham. UK) .
100 m M D T T iG ihco i . and the Superscript Pre-ampli l ical ion system (Gibco i was added, and the final solution was
incubatedd lor 60 min at 37 C'. I or RT-PCR of II.-1 a and 1L-1|1 m R N A , equivalent amounts of c D N A (5 [i\.\ were
ampli f iedd using a solution (20 LiL) containing 4 'Y D M S O . 12.5 p,g BSA (l)i l 'eo, Detroit. M l ) . 1.25 mM of each dNTPs.
lOxx PCR buffer (0.67 M Tris-HC'i i pH S.Si. 67 m M MgCI . 0.1 M ;)-mercapioethanol. 67 ^ 1 I d ) T A . 0.166 M
i N H p - S O ; ) .. 0.5 U o l 'Ampl iTaq D N A polymerase iPerkin Elmer. Bianchburg. NJ). and the forward (Pi and the reverse
(R ll primers I 100 m M eaehi.
Thee PGR reactions were carried out in the thermocycler Gene A M P ® PCR S\ stem 9700 \ Perk in Plmer. Norwalk. CT t .
Porr 11.-I re. IL-1[J. and |3-aetin mRNAs . the number of cycles thai resulted in linear amplif ication was determined (data
nott shown). Cyc l ing conditions for PCR ampli f icat ion o( 1L- \<x. 11.1 | i . and (i-actin mRNAs were: 94 C for 5 m i n d
cvclc).. before 30 (11.-1 a). 30 ( I I . - I (J) or 24 i|i-actin) cycles of 95 C for 60 s. 5S "C for 60 sand 72 C for 6 0 s fol lowed
bvv a final extension phase at 72 C for 10 mm. The primers used were: IL- l rc . (P) :5 ' -
C T C A G A G C A C C A T G C 1 ' A C A G A - 3 ' .. iR) : 5' T G G A A f C C A G G G G A A A C A t l 'G-3": I L - I | i . (1): 5 -
T C A T G G G A T G A T G A II ' A A C C T G C T - 3 ' . (R l : 5 - C C C A 1 A C T I T A G G A A G A C A C C i G A T - . V ; [Uactin. (P): 5 -
G T C A G A A G G A C T C C T AA I C K . - 3 ' . (R): 5 - G C T C G T T G C C A A T A G T G A T G - 3 ' .The PCR products were separated
onn 1.5'r agarose gels containing 0,5 x TUI - (50 m M Tr is. 45 m M boric acid. 0.5 m M l .DT.V pH N.3i and 0.5 cig/ml
ethid iumm bromide.
Assay s s
dota ll plasma bi l i rub in. AST. \ I . T . alkaline phosphatase, and y-G'l were determined with commercially available kits
iS iümai .. usinü a Hitachi analv/er (Boehnnger. Mannheim. Germany ) according to the manufacturer's instructions.
Cvtokiness were measured in duplicate by Id , ISA according to the instructions of the manufacturer (with detection l imits
inn pg/ml): I l . - l ix (S2). IL-1 |i 1156)- T N I -(M31.3) (all from R & D Systems. Abingdon. UK) and II -6 (9.S) (Pharmingen.
Sann Diego. C A I.
Histolog yy and histochemistr y
Sampless from each l iver lobe, and other parenchymal organs, were removed rapidly alter k i l l ing the animals, l ived in
oo formaldehyde, and embedded in paral l in for routine histology (3-5 uni thick sections, stained wi th haemaloxylm-
eostnn i l k U : ) ) . Histologic examination was performed on coded samples by 2 independent investigators, blinded tor
treatmentt groups and cholestatic parameters, f i ve histopathologic lealures were scored semiquantaiivelv in 4 grades ol
102 2

ChapterChapter 6
severit)) (0-3): portal inflammation, ductular proliferation, fibrosis, mitosis and necrosis. The 5 features were equally
weighedd and summed. This scoring has been descnhed previous!},' Periportal and pericentral regions of the liver lobule
weree compared, with the latter defined as concentric layers of 5 hepatocyles surrounding the terminal branches of the
hepaticc \ein. for each li\er lobe. 1 2 sections were used and 3 periportal and pericentral /ones were randomly selected
inn each section.
h>rr endotoxin-induced liver injury. 2 parameters were scored: the area of necrosis and hepatic neutrophil sequestration.
Neutrophilss were stained with the chloroacetatc esterase technique." Neutrophils were identified on the basis of positive
stainingg and morphologv and were counted in 50 high-power fields (400x). Images of the sections were captured using
ann Vano\-T microscope (Oh mpus. Tokyo. Japan) attached to a charge-coup!ed-de\ ice camera with an X-bit resolution
(Cohuu 4910: San Diego. CA). a frame grabber (LG-3; Scion. Frederick. MDl . and a Power Macintosh 8100/1 10
computerr (Apple. Cupertino. CA) using the public domain NIH Imaging software program (version 1,57: written bv
Waynee Rassband at the National Institutes of Health and available via internet from http://rsb.info.nih.go\). Sellings of
thee camera and frame grabber were adjusted according to Jonker et al."' Cell damage was quantitated in parallel sections
stainedd with H&I i by measuring pixel intensities bv blinded investigators. The percentage of necrosis was calculated bv
evaluatingg the number of microscopic fields with necrosis compared with the entire histological section (x100) SH.
Quantativee analysis with the use of digital microscopy was performed as described bv Jonker et al.:
Statisticall analysis
Statisticss were performed using the SPSS Base 9.0 for Windows Statistical Software Package (SPSS. Chicago. IL) .
Al ll results are given as mean SE. Sun ival curves were compared with the log-rank test. Differences in biochemical,
immunological,, immunohistochemical and histological data were analyzed using the Mann-Whitnev I ' lest. Changes
inn time were analyzed bv one-way analvsis of variance. Differences at indicated time points were assessed by Dunnett
t-lestt for multiple comparisons where appropriate. A 2-tailed /> value < (1.05 was considered significant.
RESULTS S
Rolee of IL- 1 in cholestatic liver injur y
RT-PCRR was performed on liver homogenates to determine whether 1L-6 is produced after bdl.
IL-66 mRNA was detectable in livers ofIL-6+/+ mice after induction of extrahepatie cholestasis
andd in sham-operated animals (Fig. 1). However. mRNA levels were distinctly higher in the bdl
group.. Similarly. IL-6 protein levels were higher in liver and plasma of bdl IL-6+/+ mice (Fig.
2). .
Neitherr sham surgery nor bdl were associated with mortality during 1 -month follow-up. Bdl mice
becamee slightly ill (ruffled fur. anorexia) from 3 weeks onwards, but mortality did not occur up
too 4 weeks after bdl, and differences between both mouse strains were not observed. Bdl mice
showedd biochemical evidence of hepatocellular injury as reflected by elevated plasma levels of
ALTT and AST. and significant cholestasis as reflected by elevated plasma levels of bilirubin,
alkalinee phosphatase, and y-GT levels (Fig. 3). However, significant differences between IL-
6+/++ and IL-6-/- mice were not found. Sham operation did not affect normal liver parenchyma
103 3

EndotoxemiaEndotoxemia in IL-IR-/- cholestatic mice
off IL-6+/+ and IL-6-/- mice up to 14 days
afterr operation as determined bv
histopathologicall examination (Fig. 4), In
contrast,, bell lead to typical cholestatic
changess in liver parenchyma which were
similarr in both mice strains (Fig. 4).
Inn conclusion, our data demonstrate that
IL-66 was produced in the liver after bdl.
butt did not play an important role in
cholestaticc liver injury.
Bdll IL-6-/ - mice are protected against endotoxinn induced lethality Too determine the role of endogenously
producedd IL-6 in the immune response to
endotoxin.. IL-16+/+ and IL-6-/- mice were
challengedd with endotoxin at 2 weeks after
bdll or sham operation and monitored
duringg 24 h. All mice that survived the
endotoxinn challenge during the first 24 h
provedd to be permanent survivors. As
shownn in Fig. 5. during the first 6 h after
endotoxinn challenge, lethality was not
IL-1R+/++ IL-1R-/-
303 3
502 2
c c 617 7
IL-1( (
IL-1p p
. . « « . . .
p-actin n
v v & &
"Q Q .6X X
F F & &
s j --6X X
observedd in any group. Moreover, endotoxi Figuree 1. 11.-la (A), [L-lp (B). and P-actin (C) mRNA expressionn in livers of 1L-1R+/+ and IL-1R-/- mice as
challengee in sham operated mice was not determined bv RT-PCR ai 2 weeks after bile duel ligation associatedd with mortality at all at the (bdl) or sham operation (n = 8).
endotoxinn dose given. However, after bdl. both mouse strains demonstrated significant mortality
afterr administration of endotoxin (both /> < 0.05 vs. sham). Importantly, a significant difference
wass found in mortality between both mouse strains. A 15% mortality was observed in IL-6-/-
micee and only 449r mortality in IL-6+/+ mice (p = 0.014).
Increasedd production of proinflammator y cytokines in bdl IL-6-/ - mice after endotoxin injection n IL-6+/++ and IL-6-/- mice were challenged intraperitoneally with endotoxin at 2 weeks after sham
surgeryy or bdl. and IL-1 a. IL-1 p. IL-6. and TNF levels were measured in plasma at different time
intervalss during the first 6 h. i.e. before any animal died. In response to endotoxin.
proinflammatoryy cytokine levels were significantly higher in bdl than in sham mice (all/) values
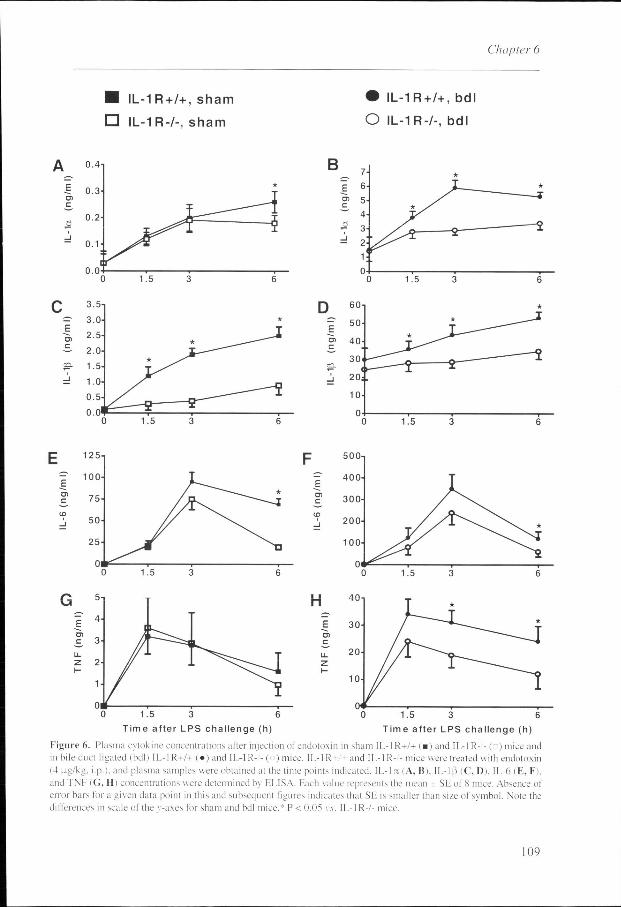
<< 0.05: Fig. 6). Compared with endotoxemic sham mice. IL-1 a levels were 20-fold higher. IL-1 P
levelss were up to 40-fold higher. IL-6 levels were 4-fold higher, and TNF levels were 10-fold
104 4

ChapterChapter 6
higherr in endotoxemic cholestatic mice.
Overall.. IL-6-/- mice demonstrated a relatively higher release of cytokines upon exposure to
endotoxinn as compared with IL-6+/+ mice. The difference in cytokine release between the 2
mousee strains was especially apparent in bdl mice (Fig. 6).
Endotoxinn induced liver damage is increased in bdl IL-6-/ - mice
Endotoxinn challenge resulted in influx of neutrophils in the liver, development of hepatocellular
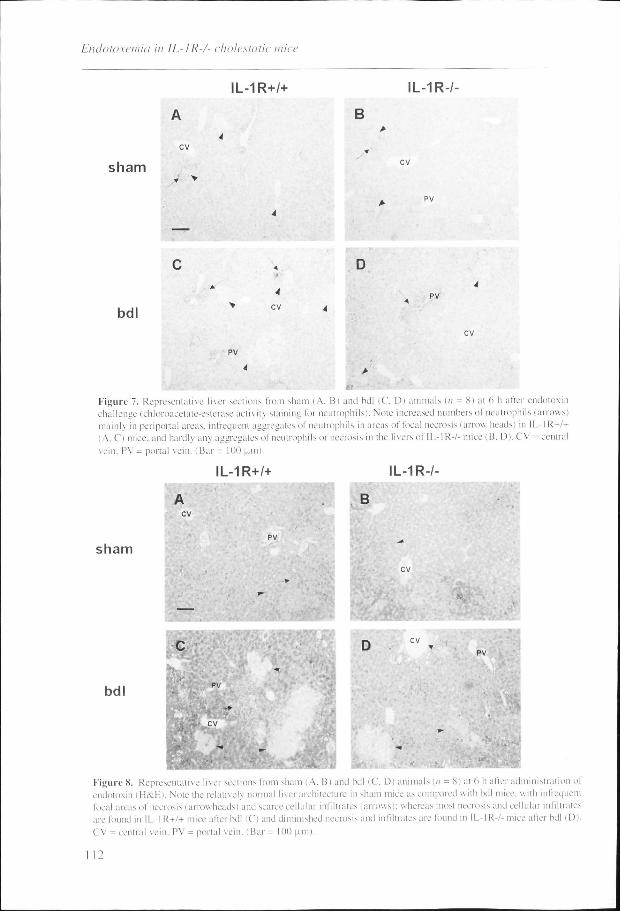
necrosis,, and an increase in plasma ALT levels (Tabic 1; Figs. 7. 8). which were more
pronouncedd in bdl mice as compared with sham mice (p < 0.001 for all responses). After bdl and
subsequentt challenge to endotoxin, neutrophil influx, hepatocellular necrosis, and the rise in
plasmaa ALT levels were all significantly more pronounced in IL-6-/- than in IL-6+/+ mice (p <
0.055 for all responses), whereas in sham animals these responses were not significantly different
inn IL-6+/+ and in IL-6-/- mice.
B B
> >
£ £ c c
ca a
11 week
60 0
50--
40--
30 0
20 0
10H H
0 0
P<P< 0.001
22 weeks
P<P< 0.001 P<P< 0.001 r
1 1 i i
X X
JSSLJJS_ _ 11 week 2 weeks
Timee after surgery
MM IL-1R+/+, sham
IL-1R-/-, sham
IL-1 R+/+, bdl
IL-1 R-/-, bdl
Figuree 2. Mean i SF. levels of IL-l aa (A) and IL-1 P (B) in livers off 1L-1R7- and IL-IR+/+ mice afterr bile duct ligation (hdl) or shamm operation (n = 8). P values indicatee differences between bdl andd sham operated mice (either IL-1R-/-- or IL-IR+/+). Differences betweenn 1L-1R-/- and [L-1R+/+ micee within bdl and sham operated groupss were not significant
105 5

EndotoxemiaEndotoxemia in IL-IR-/- cholestatic mice
JJ 300
E250H H
—— 200H c c
150-i n n m m
50H H o o
PP < 0.0001
II 1
Li i I I
11 week
11 week
QQ 900
P<P< 0.0001
r r
22 weeks
22 weeks
11 week 2 weeks Timee after surgery
üü IL-1R+/+, sham IL-1R-/-, sham IL-1R+/+. bdl
DD IL-1R-/-, bdl
1250 0
5"" 1000
HH 750 O O
^^ 500
250 250
0 0
EE 1250
3-1000 0
~~ 750 H H C/) ) << 500
11 week
250--
0 0
PP < 0.01
LI I
P<0.01 1 II I
„ J J
PP < 0.001
I I 22 weeks
pp < 0.001
11 week 2 weeks Timee after surgery
Figuree 3. Mean Sh lewis ol'bilirubm (A), alkaline phosphatase (B). y-GT (C). .ALT (D), and AST (E) in plasma oi IL-ll R+/+ and ll.-1 R-/- mice at I and 2 weeks after bile duct ligation (bdl) or sham operation t» = Si. P \allies indicate differencess between bdl and sham operated mice teither IL-l R-/- or II -l R+/+). Differences between II.-1 R-/- and II.-II R+/+ mice within bdl and sham operated groups were not significant.
DISCISSION N
Surgicall procedures in patients with obstructive jaundice arc more frequently associated with
complicationss than similar operations in patients without biliary obstruction, including the
occurrencee of sepsis syndrome. Endotoxin has been implicated as an etiologic factor in such
complicationss in jaundiced patients. In the present study, we sought to determine the role of
endogenouss IL- l in the development of hepatic inflammation and injury following biliary
obstruction,, and in the increased susceptibility toward endotoxin-induced toxicity associated with
106 6

ChapterChapter 6
obstructivee jaundice. The main findings of our study were that although IL-1 does not play an
importantt role in hepatic pathology resulting from biliary obstruction, this cytokine contributes
too a significant extent to the pathogenesis of endotoxin-induced liver injury and death in
jaundicedd mice.
II.-l uu and [L-i p arc encoded b} separate genes. ~" Both 1L-1 forms can interact with 2 types of
receptors.. Type 1 IL-1 R is expressed on virtually all cell types and is responsible for transducing
cellularr effects of IL-1 </ and IL-1 [1 In contrast, biological effects generated by IL-l a or If-1 (3 are
thuss far not known to be mediated by type 11 IL-1 R. which has led to the concept that this
receptorr type is inactive and serves as a negative regulator of IL-1 activity. Therefore, mice that
lackk type 1 II.-1 R are functionally deficient for both II,-1 a and IL-i p responses.-1 As far as we
know,, the present study is the first to examine the role of IL-1 during inflammation during biliary
obstruction. .
IL-1R+/++ IL-1R-/-
AA B
sham m
PVV c v
CC D
bd ll pv
PV V
Figur ee 4. Representative liver sections from sham and bdl animals (n = 8) at 14 days alter operation (H&E). Note thee normal liver architecture in sham mice (A. B) compared to bdl mice ( C D ). The latter exhibit infrequent local areas off necrosis t arrow heads) and scarce cellular infiltrates with marked bile duct prol iteration (arrows), comparable in both I1.-1R+/+IA.. B) and IL-1R-/-mice (C, D). CV = centra] vein. PV = portal vein. (Bar= 100 U-m).
Wee first documented that both II.-1 u and II.-1 |i are produced m the liver after biliary obstruction
Hepaticc IL-1 concentrations were already markedly elevated at I wk after bdl and did not
107 7

EndotoxemiaEndotoxemia in IL- IR-/- cholestatic mice
100» »
«« 75-> > > >
II 50-
25--
00 — 00 6 12 18 24
Hourss after endotoxin challenge Figuree 5. Endotoxin-induced lethalit) in bdl and sham II.-1R+/+ and [L-1R-/- mice. Sham IL-1R+/+ (a. n = I2). shamm II.-1R-/-Ian = 12). bdl IL-1R+/+ ) = 16). and Ixll II. 1 R-/- ( , n= 16) mice were injected with endotoxin (44 u.g/kg, i.p.l. and survival was assessed every hour during 24 h.
increasee further during the second wk of obstruction of the bile duct. We then evaluated the role
off this endogenously produced IL-I in hepatic inflammation and injury during bile duct
obstructionn by comparing several parameters in 1L-1 R+/+ and IL-1 R-/- mice. Each parameter
analysed,, i.e. mortality, levels of transaminases, alkaline phosphatase, and y-GT in plasma and
histologyy of the liver proved to be similar in both mouse strains. 1 lence, neither II,-1 a nor IL-i p
appearss to influence pathological consequences of biliary obstruction in the liver. It should be
noted,, however, that at least some extrahepatic effects of IL-1 may be exaggerated in jaundiced
animals.. Indeed, an increased responsiveness of the central nervous system to II.- ip has been
implicatedd in "sickness behavior" that is commonly observed in patients with cholestatic liver
disease,, characterized by lethargy, fatigue and malaise. This notion is derived from experiments
inn cholestatic rats, in which infusion of IL-1P into the central nervous system caused a marked
reductionn in their activity, whereas such decline was not observed in control rats/ However, in
anotherr investigation, it vv as found that the febrile response to 11.-1 p w as unaltered in cholestatic
rats.255 Recently, IL-6 was reported to be produced in livers of mice subjected to bdl,'' which is
inn line with earlier reports of elevated plasma levels of IL-6 in mice with experimentally induced
obstructivee jaundice.11'27'28 Similar to the absence of differences between IL-1 R+/+ and IL-1R.-/-
micee as described here. IL-6 gene-deficient mice did not demonstrate clear phenotypic
differencess after bdl when compared to wild type mice."
Experimentallyy induced obstruction of the bile duct was associated with a profoundly increased
susceptibilityy towards endotoxin, as indicated by the occurrence of mortality after administration
off endotoxin in a dose that did not cause mortality, enhanced cytokine release or more severe
liverr damage in sham-operated mice.
* I L -1R+ /+ +
-D-IL-1R-/-, ,
IL-1R+/+
0-IL-1R-/-, ,
,, sham
sham m
,, bdl
bdl l
PP < 0.01
P < 0 . 05 5
P << 0.001
108 8
ChapterChapter 6
IL-1R+/+, sham
IL-1R-/-, sham
IL-1R+/+, bdl
OO IL-1R-/-, bdl
E E "5> >
E E
c c
u. . z z h-h-
b --
4--
3--
2-2-
1--
ni i
H H
00 1.5 3 6 0 1.5 3 6
T imee af ter LPS chal lenge (h) T ime af ter LPS chal lenge (h)
Figuree 6. Plasma cytokine concentrations after injection of endotoxin in sham 11.-I R+/+ ) and IL-IR- - (n)mice and inn bile duct ligated (bdl) ) and IL-IR-/-(o) mice. II.-IR- and IL-IR-/-mice were treated with endotoxin (44 p.g/kg. i.p.l. and plasma samples were obtained at the time points indicated. IL-l a (A, Bi. ll.-l|3 (C, I)). IL-6 (E, F). andd TNF (G, H) concentrations were determined by ELISA. Each value represents the mean SE of S mice. Absence of errorr bars for a given data point in this and subsequent figures indicates that SE is smaller than size of symbol. Note the differencess in scale of the v-axes for sham and bdl mice.* P < 0.0? vs. ll.-l R-/- mice.
109 9

lindoto.u'iiiialindoto.u'iiiia in IL-IR-/- cholestatic mice
Thesee data confirm and extend previous studies in hdl rodents. In earlier studies in
rodents.'"11 we showed that endotoxin sensitivity is increased after bdl, and rises further when
endotoxinn transport to the cholestatic liver is increased by binding with (reconstituted) HDL.
Humanss with biliary obstruction also demonstrate evidence for enhanced susceptibility to
endotoxin/'0'11 Possible causes for this phenomenon include alterations in circulating levels of
endotoxin-bindingg proteins, a general activation state of neutrophils, and the fact that biliary
obstructionn influences the reticuloendothelial system in the liver as well as e.xtrahepatic
phagocyticc cells.
Althoughh 1L-1R-/- mice were indistinguishable from IL-IR+/+ mice with respect to their
responsee to bdl. thev were partial ly protected against the effects of systemic endotoxin exposure.
Thee role of endogenous IL-] in the pathogenesis of endotoxin shock has been studied before in
normall non-jaundiced animals. Repeated treatment with recombinant IL-1 receptor antagonist.
thee naturallv occurring inhibitor ofIL-1u and II.-1 |i conferred some protection in mice injected
withh a lethal dose of endotoxin.1 However. IL-I receptor antagonist did not inlluence
hemodynamic,, metabolic or inflammatory responses to a sublethal dose of endotoxin in baboons
orr healthy humans,l s>S l' ' and IL-1 R-/- mice have been found to generate responses to low and
highh doses of endotoxin that were similar to those registered in IL-1 R+/+ mice with respect to
fever,, anorexia, w t loss, and hepatic acute phase protein production," I .ikewise. IL-1 |V- mice
demonstratedd unaltered endotoxin responses when compared to IL-1 [W+ mice Furthermore,
neitherr II.-1R-/- nor II.-1|-W- mice were protected against lethality caused by intraperitoneal
injectionn of either high dose endotoxin or low dose endotoxin combined with D-
galaelosamine.211 4I Hence, the majority of studies argue against a significant role of IL-1 in
endotoxin-inducedd inflammation and lethality, although one study with a limited number of
animalss showed that D-galactosamine sensitized IL-1 R7- mice were protected against a lethal
intravenouss endotoxin challenge.4" Our data obtained in sham-operated mice showed that IL-1 R-
/-- mice had a modestly reduced endotoxin responsiveness, as indicated by diminished secretion
off IL- L/. IL-1 |L and IL-o. but not of TNL. whereas liver pathology was similar in sham-operated
IL-1R-/-- and IL-1R+/+ mice after administration of endotoxin. More importantly, the present
studvv demonstrates that endogenous IL-1 plavs an important role in endotoxin-induced toxicity
inn mice after biliary obstruction. Indeed. IL-1R-/- mice were less responsive to all endotoxin-
inducedd effects analysed, including cytokine release, liver injury and lethality. Thus, bdl results
inn an endotoxin hypersensitive state in which endogenous IL-1 activity is an important mediator
off endotoxin-induced damage. These data suggest that attempts to block IL-1 activity as an
adjunctivee treatment of septic complications in patients with obstructive jaundice may be more
successfull than anticipated from the negative data derived from clinical trials with recombinant
IL-11 receptor antagonist after general sepsis.
Too date, understanding of the role of inflammatory mediators in the increased endotoxin
sensitivityy during cholestasis is limited. TNF is known to be involved in the changes in body
temperaturee in response to endotoxin in cholestatic rats."" However, treatments aimed at
elinlinatiiiüü endogenous TNF activity were not able to reduce mortality in cholestatic mice
110 0

ChapterChapter 6
indicatingg that circulating TNF is not the ultimate cause of mortality observed in these mice.454''
1L-11 is a major proinflammatory cytokine that has been implicated in the pathogenesis of several
inflammatoryy diseases.1--1 We demonstrate here that production of II.-l a and ll.-lf- i occurs in
liverss of mice with bile duct obstruction, but that endogenous])' synthesized 1L-1 does not
contributee to hepatic inflammation and injury in this condition. However, exaggerated IL-1
releasee is found in bdl mice exposed to endotoxin, and eliminating this IL-1 response protects
animalss against the proinflammatory and lethal effects of endotoxin. These results suggest that
endogenouss IL-1 plays an important role in the hypersensitivity towards endotoxin during biliary
obstruction. .
Tablee 1. Endotoxin induced liver chanses
T„ T„
IL-1R+/+.. sham
IL-IR-/- .. sham
1L-1R+/+.. bdl
IL-1R-/-.. bdl
'' 90 min
1L-IR+/+.. sham
1L-1R-/-.. sham
IL-IR+/+ .. bdl
IL-IR-/- ,, bdl
T,sn T,sn
IL-1R+/+.. sham
IL-IR-/- .. sham
IL-1R+/+.. bdl
IL-IR-/- .. bdl
7\v,, ,„ IL-IR+/+ .. sham
IL-IR-/- .. sham
IL-1R+/+.. bdl IL-IR-/- .. bdl
Neutrophi i
1 1
1 1 ">") )
18 8
13 3
8 8
50 0
33 3
56 6
34 4
150 0 94 4
120 0
99 9
400 0
320 0
influx x
1 1
1 1
77 *
66 *
5 5
4 4
122 * v
77 *
12 2
9 9
333 * v
233 *
34 4
25 5 544 * -;-
344 *
Necrosiss (c/c)
0 0
0 0
23 3
17 7
5 5
3 3
288
20 0
77
5 5
35 5
25 5
15 5
8 8
40 0
299
0 0
0 0
55 *
77 =:=
3 3 -> >
66 *
44 *
4 4
3 3
55 ::: v
44 *
5 5
4 4
66 * v
44 *
ALT( U U
54 4
67 7
1254 4
1187 7
211 1
125 5
15233 -
13133 -.
326 6 too o
2020 0
1750 0
526 6
431 1
2560 0
2088 8
lits/L) )
13 3 15 5
1422 *
1377 *
34 4
24 4
11 13 :::
977 *
11 1 1
78 8
2133 :;: v
1899 *
97 7
78 8
1977 * v
2111 *
Dataa represent the mean SF from S animals per tmnip. at 9()-3oO min after endotoxin challenge. Abbreviations: bdl bilee duet limited: ALT. alanine aspartate transaminase.
I' 0.001 vs. sham: v />< 0.05 vs. IL-IR-/- . bdl
111 1
EndotoxemiaEndotoxemia in IL- IR-/- cholestatic mice
IL-1R+/+ + IL-1R-/--
B B
sham m
bdl l
Figuree 7. Representative liver sections from sham I A. Bi and bdl (C, D) animals in = Si al 6 li aller i challengee (chloroacetate-esterase activity staining lor neutrophils). Note increased numbers of neutrophil: mainhh in periportal areas, infrequent aggregates of neutrophils in areas of local necrosis i arrow heads) in (A,, C) mice, and hardly any aggregates of neutrophils or necrosis in the livers of IL-1R7- mice (B, D). CV vein,, PV = portal vein. I Bar = 100 pm).
indotoxin n (arrows) ) L-1R+/+ + == central
IL-1R+/+ + IL-1R-/--
sham m
bdl l
B B
Figuree 8. Representative liver sections from sham (A, Bi and bdl (C, D) animals!/; = 8) at 6 h after administration of endotoxinn (H&E). Note the relatively normal liver architecture in sham mice as compared with bdl mice, with infrequent locall areas of necrosis (arrowheads) and scarce cellular infiltrates (arrows); whereas most necrosis and cellular infiltrates aree found in IL-1R+/+ mice after bdl (C) and diminished necrosis and infiltrates are found in IL-IR-/- mice after bdl (D). CVV = central vein, PV = portal vein. (Bar = 100 pm).
112 2

ChapterChapter 6
REFERENCES S
1.. Armstrong CP. Dixon JM. Taylor TV. Da\ ies GC Surgical experience of deeply jaundiced patients with hue ductt obstruction. Br J Surg 1984: 7 1:234-238.
2.. Howe JR. Klinistra DS, Moccia RD. Conlon KC. Brcnnan MF. Factors predictive of survival in ampullar_v carcinoma.. Ann Surg 1998: 228:87-94.
3.. Yeo CI. Cameron JL. Sohn TA. I.illcmoe KD. Pitt HA. Talammi MA et al. Six hundred lift } consecutive pancreaticoduodenectomiess in the 1990s: pathology, complications, and outcomes. Ann Surg 1997; 226:248-257. .
4.. Pain JA. Bailey Ml: . Measurement of operative plasma endotoxin levels in jaundiced and non-jaundiced patients.. Fur Surg Res 1987: 19:207-216.
5.. Jacob AJ, Goldberg PK. Bloom N. Degenshein GA. Ko/inn PJ. Fndoioxin and bacteria in portal blood. Gastroenterologyy 1977: 72:1268-1 27()/
6.. Diamond T. Thompson RL. McGlone F. Rowlands BJ. Fndotoxin concentrations following internal and externall biliary drainage lor obstructive jaundice. Curr Surg 1989:46:31 1-313.
7.. Roughneen PT. Kumar SC. Pellis NR. Row lands BJ. F.ndoto.xemia and cholestasis. Surg Gynecol Obstel I 988: 167:205-210. .
8.. Pain JA. Reticulo-endothelial function in obstructive jaundice. Br J Surg 1987: 74:1091-1094. 9.. Sewnath Ml- , Levels JHM. Oude Fllennk RP. Van Noorden CJF. ten Kate JYVF. van Deventer SJH et al.
Fndotoxin-inducedd mortalitx in bile duct-ligatcd rats alter administration of reconstituted High-Density lipoprotein.. Hematology 2000: 32:1289-1299.
11 0 Kimmings AN. van Deventer SJ, Obcrlop H, Gou ma DJ. Treatment with recombinant hactericidal/permeahility-increasingg protein to prevent endotoxin-induced mortality in bile duct-ligated rats. .1 Am Coll Surg 1999: 189:374-379. .
11 I Lechner AJ. Velasquez A, Knudsen KR. Johanns CA. Tracy TFJr. Matuschak GM. Cholestatic li\cr injury increasess circulating 'I"NT-alpha and 1L-6 and mortality after Fscherichia coli endotoxemia. Am J RespirCrit Caree Med 1998: 157:1550-1558.
12.. Greve JW. Gouma DJ. Soeters PB, Buurman WA. Suppression of cellular immunity in obstructive jaundice' iss caused by endotoxins: a study with germ-free rats. Gastroenterology 1990: 98:478-485.
13.. Harry D. Anand R. Holt S. Dav ies S. Marie} R. Fernando B et al. Increased sensitiv ity to endotoxemia in the bilee duct-ligated cirrhotic Rat. Hematology l'999: 30:1 198-1205.
14.. O'Neil S. Html J. Filkins J. Gamelli R. Obstructive jaundice in rats results in exaggerated hepatic production off tumor necrosis factor-alpha and systemic and tissue tumor necrosis factor-alpha levels after endotoxin. Surgeryy 1997: 122:281-286.
15.. DmarelloCA. Biologic basis for interleukin-1 m disease. Blood 1996:87:2095-2147. 16.. Luster ML Gennolec DR. Yoshida 'I . Kayama P. Thompson M. Fndotoxin-induced cytokine gene expression
andd excretion in the liver. Hepatology 1994: 19:480-488. 17.. Karck I'. Peters T. Decker K. The release of tumor necrosis factor from endotoxm-stimulated rat Kupffer cells
iss regulated by prostaglandin F2 and dexamethasone. J Hepatol 1988; 7:352-361. 18.. Fischer F. Marano MA, Van Zee KJ, Rock CS, Hawes AS, Thompson WA et al. Interleukin-1 receptor
blockadee improves survival and hemodynamic performance in Fscherichia coli septic shock, but lails to alter hostt responses to sublethal endotoxemia. J Clin Invest 1992; 89:1551-1557.
19.. Zheng H, Fletcher D. Ko/ak W. Jiang M. Hofniann KJ. Conn CA et al. Resistance lo fever induction and impairedd acute-phase response in interleukin-1 beta-deficient mice. Immunity 1995: 3:9-19.
20.. Pantu/zi G. Dinarello CA. The inflammatory response in interleukin-1 beta-deficient mice: comparison with otherr cytokine-related knock-out mice. J Leukoc Biol 1996; 59:489-493.
21.. Glaccum MB. Stocking KL. Charrier K. Smith JL. Willi s CR. Malis/ewski C el al. Phenotypic and functional characterizationn of mice that lack the type I receptor for IL-1. J Immunol 1997; 159:3364-3371.
22.. Jaeschke H, Farhood A, Fisher MA, Smith CW. Sequestration of neutrophils in the hepatic vasculature during endotoxemiaa is independent of beta 2 integrins and intercellular adhesion molecule-1. Shock 1996: 6:351-356.
23.. Jonker A. Geerts W.I. Chieco P. Moorman AF. Lamers WH. Van Noorden CL Basic strategies for valid cviomelryy using image analv sis. Histochem.l 1997:29:347-364.
24.. Swain MG. Beck P. Rioux K. Le T. Augmented interleukin-1 beta-induced depression of locomotor activ ity in cholestaticc rats. Hepatology 1998: 28:1561-1565.
25.. McCullough LK. Takahashi Y. Le T. Pittman QJ. Swain MG. Attenuated febrile response to Iipopolysaccharide inn rats with biliary obstruction. Am J Physiol Gastrointest Liver Physiol 2000: 279:G 172-G 1 77.
26.. Liu Z. SakamotoT. Yokomuro S. F/ure T. Suhbotin V. Murase N el al. Acute obstructive cholangiopathv in
113 3

EndotoxemiaEndotoxemia in IL-1R-f- cholestatic mice
inieileukin-66 deficient mice: compensation by leukemia inhibitory factor (LIE) surest-- importance of gp-1 30 signalingg in the diictular reaction. Liver 2000: 20:114-1 24.
27.. Bemclmans MH. Gouma D.I. (ireve J\\'. Huurman WA. Cvtokmes tumor necrosis factor and intcrleukin-6 in experimentall biliary obslrnetion in mice. Hcpatology 1992: 15:1 132-1 13d.
28.. K/itre T. Sakamoto I'. Tsuji H. Lun/ JG. Murase N. lung .1.1 el al. The development and compensation of biliaryy cirrhosis in interlcukin-6-deficient mice. Am .1 Pathol 2000: I 56:1 6 2 7] 639.
29.. Kinmiings AN'. \an Deventer S.IH. Ohcrlop H. Rauus LAJ. Huibregtse K, (iounia DJ. Endotoxin, cytokines, andd endotoxin binding proteins in obstructive jaundice and after preoperative biliary drainage, (nil 2000: 46:725-731. .
30.. Puntis MC, Jiang WG. Plasma cvtokine levels and monoevte activation in patients with obstructive jaundice. .11 Gastroenterol Hepatol 1996; I 1:7-13.
33 I. Kunmmgs AN. van Deventer SJ. Mairuhu WM. van den Lnde A. Rauvvs LAJ. Goiima DJ. The abnormal lipid spectrumm in malignant obstructive jaundice in relation to endotoxin sensitivity and the result of preoperative biliaryy drainage. Surgery 2001: 129:282-29],
32.. Parks R\Y. Clements WD. Sinye MG. Pope C. Rowlands BJ. Diamond 'I'. Intestinal barrier dysfunction in clinicall and experimental obstructive jaundice and its reversal by internal biliary drainage. Br J Surg 1996; S3:1545-11 349.
33.. Ballinger AB. Woolley JA. Ahmed M. Miiicahy H. AKiead LM. Landon J et al. Persistent systemic inflammatoryy res|ionse aller stent insertion in patients with malignant bile duct obstruction. Gut I998; 42:555-559. .
34.. Ding JW, Andersson R. Norgien L. Stenram I' . Bengmark S. The influence of biliary obstruction and sepsis onn reticuloendothelial function in rats. Eur.I Surg 1992; 158:157-164.
35.. Ding JW. Andersson R. Stenram I' . Lunderquist A. Bengmark S. Effect of biliary decompression on reticuloendotheliall function in jaundiced rats. Br J Surg 1992: 79:648-652.
36.. Scon-Conner CE. Grogan JB. Scher KS. Bernstein JM. Bailey-Berk C. Impaired bacterial killin g in early obstructivee jaundice. Am J Surg 1993: 166:308-310.
37.. Alexander HR. Doherty ( i \ l . Buresh CM, \'en/on DJ. Norton JA. A recombinant human receptor antagonist too mierleiikin 1 improves survival after lethal endotoxemia in mice. J Exp Med 1991: 173:1029-1032.
3S.. Granowit/ E\'. Porat R. Mier JW. Orencole SF. Callahan MY. Cannon JG et al. Hematologic and immunomodulatoryy effects of an inlerleukin] receptor antagonist coinfusion during low-dose endoioxemia in healthyy humans. Blood 1993: S2:29S5-299().
39.. Van Zee K.I. Coyle SM. Calvano SE. Oldenburg HS. Stiles DM. Pnbble .1 et al. Influence of IL-1 receptor blockadee on the human response to endotoxemia. J Immunol 1995: 154:1499-1507.
40.. Leon LR. Conn CA. Glaccum MB. Kluger M.I. IL-1 type 1 receptor mediates acute phase response to turpentine. butt not lipopolvsaccharide, in mice. Am J Physiol 1996; 27 1 ;R 166S-RI675.
41.. Eaiitu/./i (i . Zheng H. laggioni R. Benigni I', Ghe//i P. Si|X' .ID et al. Effect of endotoxin in IL-1 beta-deficient mice.. J Immunol 1996: 157:291-296.
42.. Acton RD, Dahlberg PS, 1'knis ME!. Klaerner HG. Link GS. Norman JG et al. Differential sensitivity to Escherichiaa coh infection in mice lacking tumor necrosis factor p55 or uitei leukin-l pSO receptors. Arch Sing 1996:: 131:1216-1221.
43.. Opal SM, Fisher C,l, Jr., Dhainaut .IE, Vincent JL. Brase R. Loury SI'el al. Confirmatory inteileukin-l receptor antagonisii uial in severe sepsis: a phase III . randomi/ed. double-blind, placebo-controlled. multicenteriii.il. The Inlerleukin-11 Receptor Antagonist Sepsis Investigator Group, ('ril Care Med 1997: 25:1 I 15-1 124
44.. fisher C.I. Jr.. Dhainaut .IF. Opal SM. Pnbble .IP, Balk RA, Slotman G.I el al. Recombinant human interleukin II receptor antagonist in the treatment of patients with sepsis syndrome. Results from a randomi/ed. double-blind,, placebo-controlled trial. Phase 111 rhll.-lra Sepsis Syndrome Study Group. JAMA 1994: 27 1:1 S36-1S43.
45.. Bemclmans MH. Greve JW. Gou ma DJ. Buurman WA. Increased concentrations of tumour necrosis factor i fNF )) and soluble TNT' receptors in biliary obstruction in mice: soluble 'I'NF receptors as prognostic factors forr mortality. Gul 1996: 38:447-455.
46.. Bemclmans Mil . Gouma DJ. Greve JW. Buurman WA, Effect ot antitumour necrosis factor treatment on circulatingg tumour necrosis factor levels and mortality alter surgery in jaundiced mice. Br ,1 Surg 1993: 8(1:1055-1058. .
114 4





























