Embed Size (px)
Citation preview

Contents lists available at SciVerse ScienceDirect
Livestock Science
Livestock Science 149 (2012) 282–288
1871-14
http://d
n Corr
Institut
Carrete
Tel.: þ3
E-m
journal homepage: www.elsevier.com/locate/livsci
Use of different sources of information for the recovery and geneticmanagement of endangered populations: Example with the extremecase of Iberian pig Dorado strain
Jesus Fernandez a,n, Ignacio Clemente b, Carmen Amador a, Alberto Membrillo c, Pedro Azor c,Antonio Molina c
a Departamento de Mejora Genetica Animal, Instituto Nacional de Investigacion y Tecnologıa Agraria y Alimentaria (INIA), 28040 Madrid, Spainb Centro Tecnologico Andaluz del Sector Carnico (TEICA), Pol. Ind. El Ponton, parcela 136, 21230 Cortegana (Huelva), Spainc Departamento de Genetica, Facultad de Veterinaria, Universidad de Cordoba, 14071 Cordoba, Spain
a r t i c l e i n f o
Article history:
Received 17 November 2011
Received in revised form
17 July 2012
Accepted 23 July 2012
Keywords:
Ex-situ conservation
Genetic recovery
Rare breeds
Iberian pig
13/$ - see front matter & 2012 Elsevier B.V.
x.doi.org/10.1016/j.livsci.2012.07.019
espondence to: Departamento de Mejora
o Nacional de Investigacion y Tecnologıa Agr
ra de la Coruna Km. 7, 28040 Madrid, Spain.
4 913471487; fax: þ34 913478743.
ail address: [email protected] (J. Fernandez).
a b s t r a c t
Recovery and management of an endangered population requires the use of all available
tools. Besides the genealogical data (often scarce) molecular information should be used
to refine the genetic relationships between individuals within the programme. Pheno-
typic information on a diagnostic trait could also be integrated as a management
criterion, but this strategy could conflict with the objective of maintaining the highest
levels of genetic diversity. In this study we used data from 19 individuals, founders
of the recovery programme of the Iberian pig Dorado strain and genotyped for
19 microsatellites, to test the optimal way to combine all information (i.e. pedigree,
molecular markers and values for a morphological index) and the consequences of each
decision on the genetic structure of the population. Results show that it is possible to
find equilibrium solutions where, for example, 43% of increase in the average levels of
the trait of interest can be reached with just a 4% of loss in genetic diversity and almost
no increase in the inbreeding coefficient.
& 2012 Elsevier B.V. All rights reserved.
1. Introduction
Populations of animals (whether wild or domestic) witha reduced number of individuals should be managed withcare from the genetic point of view to avoid the problemsarising from genetic drift (Falconer and Mackay, 1996).Small effective population sizes lead to loss of geneticdiversity and increase inbreeding. Both processes maycompromise the survival of the population due to fitnessreduction of individuals and the loss of the ability to adapt
All rights reserved.
Genetica Animal,
aria y Alimentaria,
to a changing environment. Typical management methodsrely on the knowledge of the genetic relationship betweenmembers of the population, until recently based on genea-logies (Frankham et al., 2002). However, the rapid develop-ment of molecular genetics has enabled the substitution orcomplementation of pedigree information, often absent orincomplete, by molecular information.
In some situations, besides the general objectivesalready pointed out, a conservation programme may havethe additional aim of recovering a breed or strain that hasbeen mixed with other individuals of the same speciesdue to mismanagement or absent management. In thatcase, strategies should also be aimed to gradually recoverthe specific genetic composition of that population. Forsuch a task we may use the genealogical information (ifwe can trace back to the moment exogenous individuals

J. Fernandez et al. / Livestock Science 149 (2012) 282–288 283
entered the population) or molecular information (ifmarker data on the genetic constitution of populationbefore the undesired introgression took place are avail-able; Amador et al., 2011). However, these data are rarelyavailable and, therefore, ‘purification’ must be based onother information. One of the possibilities could be to relyon the phenotypic records of traits used to characterisethe population. This introduces an extra objective in theconservation programme next to the maintenance of thegenetic diversity: individuals should resemble the oneshistorically accepted as pure animals. These considera-tions imply the implementation of an explicit process ofselection, which is in conflict with the general objectiveof maintaining the highest levels of genetic diversity.Consequently, selection and maintenance of geneticdiversity must be balanced to optimise the conservationprogramme (Colleau et al., 2004; Grundy et al., 1998;Meuwissen, 1997).
The berian pig breed is characterised by a rich internaldiversity, which is organised in different strains and lines.In recent decades, the within-breed diversity has sufferedintense erosion due to the dominance of the moreproductive lines (mainly coming from the Retinta strain),thus making the recovery of other lines advisable. TheDorado Gaditano (Cadiz’s Golden Iberian Pig Strain) was avariety of Iberian pig (included in the Blond group ofstrains) which was officially declared extinct in the 90swhen the last existing animals were slaughtered forhealth reasons. However, in recent years, some feral pigshave been detected in numerous locations of the moun-tains around Campo de Gibraltar (Cadiz, south of Spain)showing a similar morphology to the one described forthe ancestral Dorado Gaditano. These animals have beencaptured to study the possibility of using them for therecovery of the strain.
Fig. 1. Genealogical structure of the 19 pigs used in t
The objective of the present study was to investigatethe consequences of applying different strategies anddifferent sources of information in the management andrecovery of a small population using the data from theDorado Gaditano pig as a case study. In particular, wetried to determine (i) the differences between usingmolecular or pedigree based coancestry in the geneticmanagement; (ii) the effect on inbreeding and geneticdiversity of excluding individuals because of their mor-phology; (iii) the consequences of managing for lessinbreeding or managing for less coancestry (morediversity).
2. Material and methods
2.1. Population
A total of 22 pigs showing a similar morphology to theone described for the ancestral Dorado Gaditano werecaptured from nature between 2008 and 2010 in severallocations of the mountains around Campo de Gibraltar(Los Alcornocales and Los Barrios area in Cadiz, southernSpain). The first analysis was performed to check for theabsence of crosses with wild boar through cytogeneticprobes (Sysa, 1980). Crosses with other pig breeds wereexcluded by searching for breed specific alleles in thegenes IGF2 and MCR1 (Carrodeguas et al., 2005;Fernandez et al., 2004). After these processes, 19 pigs(12 females and 7 males) remained available as initialfounders in the recovery centre (Centro ExperimentalAgrıcola y Ganadero de Jerez de la Frontera, Cadiz, Spain).These consisted of two sows with their offspring, a coupleof presumed full-sibs and three other animals withoutknown relationship (see Fig. 1). Results presented in thispaper were based on these 19 founders. At the time of
he study and their morphological index value.

J. Fernandez et al. / Livestock Science 149 (2012) 282–288284
writing the population kept at the Dorado GaditanoRecovery Centre comprises more than 100 pigs.
Animals were genotyped for a panel of 19 microsatel-lites from the set recommended by FAO for genotypingpig populations (FAO, 2004). Two other groups of Iberianpigs, one from the Torbiscal strain and another from otherIberian lines, were also genotyped for the same micro-satellites. The Torbiscal strain was founded in 1944 usinga small number of breeding animals and, in this sense, hasa similar history to the population of this study. Themolecular information was used to perform a ‘cluster’analysis to ascertain the relationship of the studiedanimals with other strains using STRUCTURE (Pritchardet al., 2000).
Genealogical relationships (known or assumed) werechecked for agreement with microsatellite information.Parent-offspring compatibility was tested by checkingthat they share one allele at every loci. For full sibs familycongruence each loci was studied to determine if itcomplied with the Mendelian transmission rules, i.e.(i) not more than four different alleles and genotypescan exist in a family; (ii) a particular allele cannot be inheterozygosity with two other alleles at most; (iii) if thereare four alleles, no homozygotes can be found; (iv) if thereare three alleles, only one type of homozygote can exist.In all comparisons performed with our data genealogyand molecular information were in agreement.
2.2. Morphological characterisation
All animals included in the recovery nucleus weremorphologically scored according to the standard definedfor Dorado Gaditano from previous morphological char-acterisation studies (Pardo et al., 1998) and compiledfrom old photographic materials. The body regions eval-uated were head (head profile, shape of the ear and nose),coat-colour, hair, rump, back, limbs, number of teats andgeneral conformation. These scores were summarised in aglobal assessment index, which ranged from one (i.e.completely conforming to breed standard) to zero. Valuesof the index for each individual are also shown in Fig. 1.The mean value in the founders was 0.5270.23.
2.3. Management procedures
2.3.1. Objective function
Objectives for the production of offspring in the nextgeneration were minimisation of the global coancestry ofactual breeding animals (in order to maximise theamount of genetic diversity transmitted) and the mini-misation of the coancestry between each mating pair (tominimise inbreeding in the offspring). The mixed objec-tive function to optimise (minimise) was the one pro-posed by Fernandez et al. (2001)
XN
i ¼ 1
XN
j ¼ 1
Xk
xik
! Xl
xjl
!f ij
" #þl
XNm
i ¼ 1
XN
j ¼ Nmþ1
xijf ij
0@
1A,
where xij is a binary variable with a value 1 if the matingbetween male i and female j is to be performed and 0otherwise, N is the total number of available breeding
animals (the Nm first are males and the rest females), andfij is the coancestry between two individuals. The firstterm of the equation represents the expected coancestryin the next generation (which is inversely related to thetransmitted diversity) as it is the average coancestrybetween potential parents weighted by the number ofmatings in which they take part. The second term deter-mines the inbreeding in the offspring because it is theaverage coancestry between the male and female of theparticular couples to be mated. The factor l is the relativeimportance given to the avoidance of inbreeding in theshort-term versus the maintenance of genetic diversity.Different values of the weighting factor (l¼0, 0.5 or 1)were simulated to test for the consequences of puttingmore importance on inbreeding or genetic variability. Lowor null values of l imply prioritising the transmission ofthe highest levels of diversity even if it leads to inbredmatings while high l yield a minimum coancestry matingdesign although the contributions from each parent aresuboptimal.
2.3.2. Demographic considerations
As in any other conservation programme, one of theinitial priorities is to quickly enlarge the population toavoid deleterious genetic consequences arising from smallcensus sizes, as well as minimising the risk from demo-graphical stochasticity. Consequently, the policy is to keepall the born offspring from each pair. Apart from differ-ences in litter size (which are not predictable), contribu-tions of potential parents (i.e. individuals available toreproduce), therefore, can only vary by changing thenumber of matings in which each animal participates.This is the reason why variables are binary instead ofnatural numbers as in Fernandez et al. (2001). Addition-ally, all females were made to contribute with one litterand, thus, just optimising their partners; males couldparticipate in several, one or none matings. Notwith-standing this, another scenario was simulated by remov-ing the restriction on the use of females to enlarge thefeasible solution space and to allow the algorithm toexclude highly related females (carrying redundantinformation) or those showing a low morphological index(see below).
2.3.3. Morphological traits
An additional restriction was imposed on the optimi-sation limiting the minimum expected value for themorphological index in the offspring. In this way, thepopulation will evolve towards the morphological defini-tion of the breed. The index was assumed as an additivetrait and, therefore, the expected value was calculated bymultiplying the value of each breeder by the proportion ofmatings in which it participates. A trade-off was con-structed by changing the value of the restriction to testthe consequences of index levels on other variables.
If the main criteria for breed definition are morpholo-gical, as in the studied breed, it might not be enough torestrict the mean value of the index in the next genera-tion. A particular average population level for the indexcan be obtained by creating families with very highmean values and other families with low values. However,

J. Fernandez et al. / Livestock Science 149 (2012) 282–288 285
offspring from the latter will be useless for the recoveryprogramme, since they will show very low values for theindex. To study the possibility of avoiding this drawback,another scenario was simulated where not only theexpected mean value of the population was restrictedbut the mean value for the index of each couple wasforced to be above 0.5 as well.
2.4. Genetic relationships
The genealogical coancestry (fG) matrix was con-structed from the known relationships (see above). Indi-viduals in the top of the genealogy were assumedunrelated and non-inbred, although some of the ‘foun-ders’ might be relatives considering that the wild popula-tions were rather small.
The molecular coancestry (fM; Eding and Meuwissen,2001) was also calculated from the microsatellite infor-mation. Pedigree coancestry corresponds to the expectedgenetic relationship between individuals while molecularcoancestry is the particular realisation of this expectationin the marker loci (taking into account the Mendeliansampling; Baumung and Solkner, 2003; Engelsma et al.,2011; Wang, 2001). The correlation between both coan-cestry matrices was high (40.80) reflecting the con-gruency between the assumed pedigree relationshipsand the markers genotypes mentioned before. Using acomparable set of microsatellites, Leroy et al. (2009)found a much lower correlation between genealogicaland molecular based measures of genetic relationship indog breeds.
Optimisations involved in the management procedurewere performed in all simulated scenarios using in theobjective function the genealogical coancestry, the mole-cular one or a linear combination of both coancestries(namely fGþ0.1� fM). In this way (i.e. with a small weighton molecular coancestry) main decisions were driven bypedigree relationships but it was possible to detectdifferent degrees of similarity within equal genealogicalrelationships.
Summarising, simulated scenarios were the combina-tion of the following factors: (i) type of coancestry(genealogical, molecular or combined); (ii) weightingfactor (l) for the avoidance of inbred matings (0, 0.5 or 1);(iii) restriction only on the average expected index valueor on the particular value of each family; (iv) all femalesmade to mate or exclusion of females possible. In allcases, optimisations were performed using a simulatedannealing algorithm, as described by Fernandez et al.(2001).
2.5. Evaluation criteria
To compare the consequences of the different scenar-ios and the particular levels for each of the restrictions,several variables were monitored. Expected average off-spring inbreeding (F), coancestry (f) and morphologicalindex value were calculated as measures directly relatedto the objectives of the conservation and recovery pro-gramme. The first two variables were calculated with thesame coancestry matrix used in the optimisation (either
genealogical, molecular or mixed coancestry). Number ofcontributing males and the maximum number of matingsper male were also recorded as a measure of the varianceof the contributions which is related to the effectivepopulation size (Falconer and Mackay, 1996).
3. Results and discussion
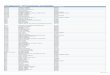
Clustering analysis showed that the most probablestructure comprised five groups (Fig. 2) The three originalsets of individuals were clearly separated (i.e. Dorado,Torbiscal and rest of Iberian pigs) and both within theDorado and Torbiscal strain two subpopulations weredistinguished. Individuals are assigned almost always tosingle groups, presenting little signature of admixture.
Two different groups of Dorado pigs were detected,closely in agreement with the geographical origin of theanimals, captured in two completely isolated zones. It is alsoremarkable to find a clear genetic differentiation in theTorbiscal strain. This probably reflects the effects of geneticdrift in small almost isolated populations although markedfamily structure (e.g. non related families as is the casewithin the studied population) may bias results fromclustering methods detecting this family structure insteadof higher levels of structuring (lines/breeds) as might bedesired (Anderson and Dunham, 2008).
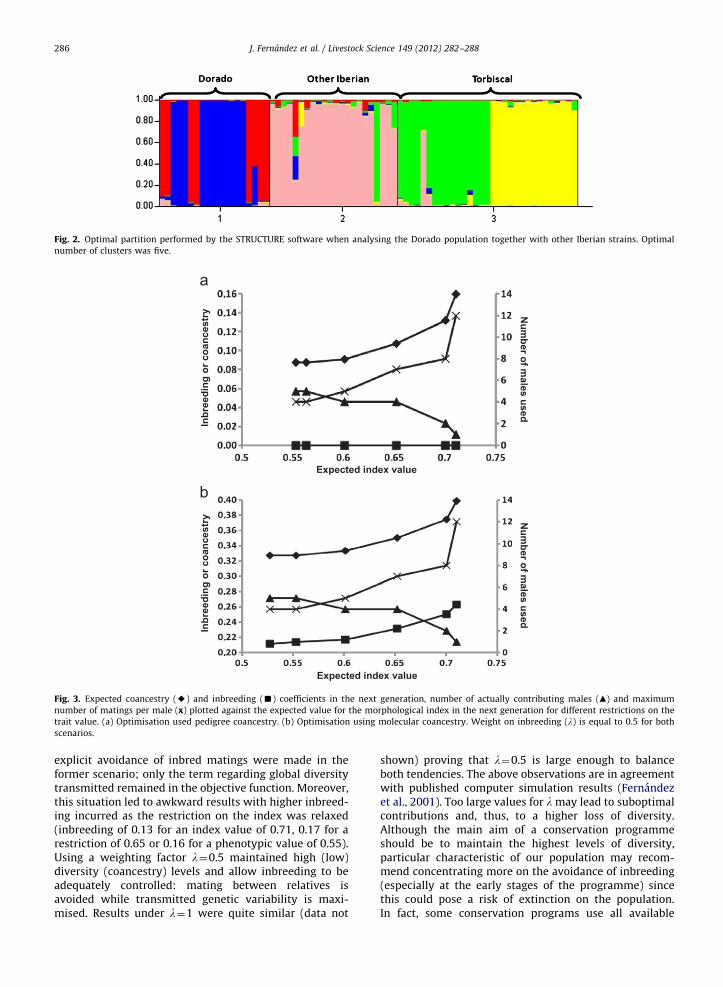
Trends obtained by applying different restrictions onthe mean expected value for the morphological index areshown in Fig. 3 for scenarios using exclusively genealogi-cal or molecular information, respectively (with a weighton the expected inbreeding l¼0.5). Due to the highcorrelation between coancestry matrices calculated fromboth sources of information (E80%) tendencies were verysimilar, as happened with the mixed coancestry matrix(data not shown).
Expected coancestry in the next generation was higherwith stronger restrictions on morphology (Fig. 3). The mostprominent difference between scenarios was between theinbreeding levels. When relying on the genealogical infor-mation and because of the ‘shallow’ available pedigree, itwas possible to avoid inbred litters for all levels of restric-tion on phenotype, as there were males unrelated to everyfemale. Remember that relationships are equal for allmembers of a full sib family and that candidate parentsbelong to unrelated families. Contrarily, molecular coances-try allowed to distinguish degrees of similarity even withinfamilies, selecting less inbred matings as the restriction onthe index was relaxed.
In populations with ‘deeper’ genealogies (i.e. moregenerations of known pedigree) the different degrees ofgenealogical relationship will increase allowing a finercontrol of both inbreeding and diversity. Anyway, the useof the molecular coancestry is still advisable in order tochoose between different individuals from the samefamily. The higher the number of markers the moreprecise the differentiation between sibs.
Regarding the optimal value of l, it was observed thatvalues close to 0.5 could be a good choice. Under l¼0average inbreeding ranged from 0.13 to 0.17 dependingon the restriction on the morphological index while itvaried between 0.11 and 0.13 for l¼0.5 This is because no
Expected index value
Expected index value
Inbr
eedi
ng o
r coa
nces
try
Inbr
eedi
ng o
r coa
nces
try
Num
ber of males used
Num
ber of males used
Fig. 3. Expected coancestry (E) and inbreeding (’) coefficients in the next generation, number of actually contributing males (m) and maximum
number of matings per male (x) plotted against the expected value for the morphological index in the next generation for different restrictions on the
trait value. (a) Optimisation used pedigree coancestry. (b) Optimisation using molecular coancestry. Weight on inbreeding (l) is equal to 0.5 for both
scenarios.
Fig. 2. Optimal partition performed by the STRUCTURE software when analysing the Dorado population together with other Iberian strains. Optimal
number of clusters was five.
J. Fernandez et al. / Livestock Science 149 (2012) 282–288286
explicit avoidance of inbred matings were made in theformer scenario; only the term regarding global diversitytransmitted remained in the objective function. Moreover,this situation led to awkward results with higher inbreed-ing incurred as the restriction on the index was relaxed(inbreeding of 0.13 for an index value of 0.71, 0.17 for arestriction of 0.65 or 0.16 for a phenotypic value of 0.55).Using a weighting factor l¼0.5 maintained high (low)diversity (coancestry) levels and allow inbreeding to beadequately controlled: mating between relatives isavoided while transmitted genetic variability is maxi-mised. Results under l¼1 were quite similar (data not
shown) proving that l¼0.5 is large enough to balanceboth tendencies. The above observations are in agreementwith published computer simulation results (Fernandezet al., 2001). Too large values for l may lead to suboptimalcontributions and, thus, to a higher loss of diversity.Although the main aim of a conservation programmeshould be to maintain the highest levels of diversity,particular characteristic of our population may recom-mend concentrating more on the avoidance of inbreeding(especially at the early stages of the programme) sincethis could pose a risk of extinction on the population.In fact, some conservation programs use all available

J. Fernandez et al. / Livestock Science 149 (2012) 282–288 287
animals as parents in the early generations and justoptimise the mating scheme.
Obviously, the number of males actually contributingdecreased as the restriction on the value of the morpholo-gical index was more stringent. In the extreme, just one male(the one with the highest morphological score) is used tomate with all the females. However, not all males are usedeven in the case of no restriction on morphology, becausethe method avoids using males from the same litter sincethis would increase the average coancestry of the offspring.
Following the same reasoning and realising that therewere females from the same litter (i.e. being full-sibs) ascenario where females could be excluded from reproduc-tion was also tested. In Fig. 4 it can be observed that therelaxation of this restriction enlarged the feasible space ofsolutions with higher and lower values for the indexobtained compared to the fixed contribution scenario were.
The main advantage of this strategy is that, for thesame value of the index, the relaxed scheme alwaysyielded lower levels of inbreeding and coancestry. Conse-quently, from the pure genetic point of view the use of allfemales should not be forced. However, from the demo-graphic point of view and looking for a rapid populationsize increase, not reproducing some females can be lessadvisable since fewer females will contribute (only fiveout of twelve in the present example). Limiting thenumber of females (litters) may increase the risk to theprogramme if there is a mating failure or any of the littersis not viable. This is especially crucial in species with lowprolificacy (e.g. cattle, horses) where reproductive ratesdo not allow for a rapid increase in population size and,therefore, requires the reproduction of all available indi-viduals. This means that the avoidance of inbreedingthrough the design of matings will automatically becomethe main objective in those scenarios.
When the mean value for the index of any particular pairwas restricted (i.e. banning solutions with pairs havingmean morphological index below 0.5) the lowest average
Inbr
eedi
ng o
r coa
nces
try
Expected index value
Fig. 4. Expected coancestry (E) and inbreeding (’) in the next gen-
eration plotted against the expected value for the morphological index
in the next generation for different restrictions on the trait value when
imposing all females have to contribute offspring (solid line) or when
some females can be excluded from breeding (dashed line). Molecular
coancestry matrix used in the optimisation. Weight on inbreeding (l) is
equal to 0.5.
value obtained was 0.6 for any single litter. However, resultsfor any of the measured variables coincided with thoseobtained in the situation where only the mean offspringindex value was restricted. For example, mean offspringinbreeding (coancestry) was 0.22 (0.33) when indexrestricted to 0.6 in both scenarios. These observations provethat it is possible to limit the minimum expression for a traitwithin the conservation programme without changing thelevels of the rest of variables.
Although in conservation programs the main objectiveis the maintenance of diversity, the levels of performancefor some traits cannot be ignored, especially when dealingwith livestock species. Therefore, dual objective functionsare to be optimised, trying to obtain gain on the traitbut limiting the loss of diversity (Grundy et al., 1998;Meuwissen, 1997) or, as it was done in this study,maximising the maintained diversity but at a minimumlevel for the trait. Although conceptually different, the twoapproaches (i.e. maximising gain or maximising diversity)are mathematically equivalent and the same trade-off isobtained by moving the restriction on inbreeding or gain,respectively. Besides morphological characteristics, itwill be desirable to use another productive trait in themanagement criteria, such as the number of weanedpiglets: the Dorado Gaditano strain animals have beenclaimed to have higher prolificacy and maternal charac-teristics than other Iberian lines. If this population can beassociated with a superior performance for a productiveor reproductive trait it will provide another reason tocontinue with the conservation programme, promoting itssustainable recovery. Unless dealing with ornamentalor leisure species or when the morphological character-istics may confer special adaptations to the animals, itseems little advisable to base recovery programs on thesekind of traits.
The present study shows that, even under adverseconditions (i.e. an extremely small population size withscarce information), the use of the adequate methodologyand the sensible restrictions can provide acceptable solu-tions by balancing the needs for maintaining high levels ofdiversity and the increase in the expression of a trait ofinterest. The management of a population within a con-servation programme must take advantage of every possiblesource of information that helps to reach the goals. There-fore, it is important to use the molecular information next tothe available pedigree, especially among founders to deter-mine their unknown genetic relationships.
Conflict of interest statement
Jesus Fernandez Martın declares that no conflict ofinterest exists for any of the authors.
Acknowledgements
We want to thank the two anonymous referees and theeditor for their comments on the initial version of themanuscript. This work was supported by Grants RZ2008-00015 (INIA, Spain) and CGL2009-13278-C02-02 (Minis-terio de Ciencia e Innovacion, Spain) and Fondos FEDER.

J. Fernandez et al. / Livestock Science 149 (2012) 282–288288
Acknowledgements to the Agricultural and LivestockExperimental Centre of Jerez de la Frontera Cadiz Spainwhere the Cadiz’s Golden Iberian Pig recovery Centre hasbeen established.
References
Amador, C., Toro, M.A., Fernandez, J., 2011. Removing exogenous informationusing pedigree data. Conserv. Genet. 12, 1565–1573.
Anderson, E.C., Dunham, K.K., 2008. The influence of family groups oninferences made with the programme structure. Mol. Ecol. Res. 8,1219–1229.
Baumung, R., Solkner, J., 2003. Pedigree and marker information require-ments to monitor genetic variability. Genet. Sel. Evol. 35, 369–383.
Carrodeguas, J.A., Burgos, C., Moreno, C., Sanchez, A.C., Ventanas, S.,Tarrafeta, L., Barcelona, J.A., Lopez, M.A., Oria, R., Lopez-Buesa, P.,2005. Incidence in diverse pig populations of an IGF2 mutation withpotential influence on meat quality and quantity: an assay based onreal time PCR (RT-PCR). Meat Sci. 71, 577–582.
Colleau, J.J., Moureaux, S., Briend, M., Bechu, J., 2004. A method for thedynamic management of genetic variability in dairy cattle. Genet.Sel. Evol. 36, 373–394.
Eding, H., Meuwissen, T.H.E., 2001. Marker-based estimates of betweenand within population kinships for the conservation of geneticdiversity. J. Anim. Breed. Genet. 118, 141–159.
Engelsma, K.A., Veerkamp, R.F., Calus, M.P.L., Windig, J.J., 2011. Con-sequences for diversity when prioritizing animals for conservationwith pedigree or genomic information. J. Anim. Breed. Genet. 128,473–481.
Falconer, D.S., Mackay, T.F.C., 1996. In: An Introduction to QuantitativeGenetics4th ed. Longman, Harlow, England.
FAO, 2004. Secondary Guidelines for Development of National FarmAnimal Genetic Resources Management Plans. Measurement ofDomestic Animal Diversity (MoDAD): Recommended MicrosatelliteMarkers. Rome, Italy.
Fernandez, A., Fabuel, E., Alves, E., Rodrıguez, C., Silio, L., Ovilo, C., 2004.DNA tests based on coat colour genes for authentication of the rawmaterial of meat products from Iberian pigs. J. Sci. Food Agric. 84,1855–1860.
Fernandez, J., Toro, M.A., Caballero, A., 2001. Practical implementation ofoptimal management strategies in conservation programmes: amate selection method. Anim. Biodivers. Conserv. 24, 17–24.
Frankham, R., Ballou, J.D., Briscoe, D.A., 2002. Introduction to Conserva-tion Genetics. Cambridge University Press, Cambridge, UK.
Grundy, B., Villanueva, B., Woolliams, J.A., 1998. Dynamic selectionprocedures for constrained inbreeding and their consequences forpedigree development. Genet. Res. 72, 159–168.
Leroy, G., Verrier, E., Meriaux, J.C., Rognon, X., 2009. Genetic diversity ofdog breeds: within-breed diversity comparing genealogical andmolecular data. Anim. Genet. 40, 323–332.
Meuwissen, T.H.E., 1997. Maximizing the response of selection with apredefined rate of inbreeding. J. Anim. Sci. 75, 934–940.
Pardo, J., Mata, C., Barba, C., Rodero, A., Delgado, J.V., Molina, A., Dieguez,E., Canuelo, P., 1998. Morphometric study in red varieties of Iberianpig breed and Jabugo spotted. Arch. Zootec. 47, 287–290.
Pritchard, J.K., Stephens, M., Donnelly, P., 2000. Inference of populationstructure using multilocus genotype data. Genetics 155, 945–959.
Sysa, P., 1980. Polymorphism of metaphase chromosomes in swine (Susscrofa L.). Genetica 52-53, 213–315.
Wang, J., 2001. Optimal marker-assisted selection to increase theeffective size of small populations. Genetics 157, 867–874.