Embed Size (px)
Citation preview

Journal of Virological Methods, 12 (1985) 59-70
Elsevier
JVM 00439
59
USE OF CHICKEN AND RABBIT ANTIBODIES IN A SOLID PHASE
PROTEIN A RADIOIMMUNOASSAY FOR VIRUS DETECTION*
DAVID KATZ, SHOSHANA LEHRER and ALEXANDER KOHN
Israel Institute for Biological Research, P.O.B. 19, Ness-Ziona, Israel
(Accepted 17 June 1985)
A new rapid indirect solid phase radioimmunoassay was developed for the detection of Sindbis virus.
Chicken antibodies were adsorbed onto wells In microplates to serve as ‘capture antibodies* and rabbn
antlbodies were used as the second antibody. 12SI-labelled protein A that does not bind to chicken
antibodles, but binds firmly to rabbit antibodies was used as the tracer.
All the steps necessary for the development of the assay are described. The minimal amount of Sindbis
virus detectable was around 3 X lo5 PFU/ml and the interassay reproducibility was about f 30%.
solid phase radioimmunoassay Sindbis virus
INTRODUCTION
Protein A, a cell wall constituent of Staphylococcus UW~US (Cowan I), has a high
affinity for the Fc part of many mammalian immunoglobulins (Langone, 1982;
Lindmark et al., 1983; Richman et al., 1982; Richman, 1983). It can therefore replace
antispecies antibodies in indirect immunoassays, and serve as a broad reacting tracer
reagent. This feature of protein A was successfully applied for the detection of
antibodies in solid phase protein A radioimmunoassay (SPA-RIA) and enzyme immu-
noassay (Engvall, 1978; Jahrling et al., 1978; Anastasiu and Perrin, 1979; Colombati
and Hilgers, 1979; Marier et al., 1979; Mador et al., 1978; Potgieter et al., 1980; Sprouse
et al., 1981; Richman, 1983).
Detection of antigens using protein A, requires antigen adsorbtion directly onto a
solid phase (Goding, 1978; Enzman, 1978; Langone et al., 1977; Langone, 1978;
Langone, 1980; Richman et al., 1982; Richman, 1983; Butler et al., 1979). However,
such systems are not applicable to unpurified viruses or antigens in clincal specimens,
since irrelevant proteins and other materials present bind also, limiting the amount of
*In memory of the late Eva Gruber whose excellent technical assistance and contribution had a major
influence on this work.
0166.0934/85/$03.30 0 1985 Elsevier Science Publishers B.V. (Biomedical Division)

60
specific antigen bound and reducing the sensitivity of the detection. The problem is
even worse with serum samples, because background values increase due to reactivity
of the immunoglobulins with the labelled protein A. There has been limited use of a
sandwich assay with protein A as tracer. Yolken and Stopa (1980) used goat antiserum
as ‘capture antibodies’, and Yolken and Leister (1981), used protein A enzyme
immunoglobulin conjugates and goat or burrow antisera as ‘capture antibodies’.
Chicken antibodies do not bind protein A (Kronval, 1974). Thus, we devised an
indirect sandwich type of immunoassay for the detection of antigens using chicken
antibodies which were absorbed on microwells to serve as ‘capture antibodies’. The
antigen, bound by the chicken antibodies was then detected by rabbit antibodies
followed by ‘2SI-labelled protein A. This assay, named solid phase protein A radioim-
munoassay (Sandwich) or SPA-RIA(S) was used in our laboratory to detect a variety
of antigens.
We describe here the steps involved in the development of SPA-RIA(S) for the
detection and titration of Sindbis virus as a model. Results are presented to demon-
strate the performance parameters of the assay, i.e., specificity, sensitivity and repro-
ducibility.
MATERIALS AND METHODS
Viruses
Sindbis virus was grown in baby hamster kidney (BHK-21) cells, cultivated in
Eagle’s medium supplemented with 5% fetal calf serum (FCS). Three types of prepara-
tions were used:
(a) Unpurified virus stocks derived from the infected cells and medium by repeated
freezing and thawing.
(b) Semi-purified Sindbis virus, prepared from unpurified virus stocks by centrifuga-
tion in Beckman L2-65 ultracentrifuge, SW 50.1 rotor, 45,000 rpm, for 75 min and
resuspension of the pellet to the original volume in phosphate-buffered saline
(PBS), pH 7.2.
(c) Purified Sindbis virus stocks, prepared from unpurified stocks by gradient centri-
fugation (Pfefferkorn, 1963).
In addition, the following unpurified virus stocks were prepared: Sindbis virus grown
in chick embryo fibroblasts (CEF) in Eagle’s medium supplemented with 5% normal
chick serum; Semliki forest virus (SFV), cultured in BHK-21 cells, as described for
Sindbis virus; West Nile virus (WNV) was grown in Vero-cells, in Eagle’s medium
supplemented with 5% FCS. All preparations were subsequently stored in -7O”Cuntil
used. Infectivity titers were determined by plaque assays.
All virus preparations were kindly provided by colleagues from our department:
Sindbis viruses by Dr. S. Lustig, Dr. A. Shapira and Dr. P. Fuchs; SFV by Dr. U.
Olshevski, and WNV by Dr. Y. Akov.

61
Protein A iodina tion
Protein A (Pharmacia, Sweden) was labelled by the chloramine T method (Hunter,
1973); 10 ul of a 1 mg/ml Protein A solution were mixed with 10 ul containing 1 mCi of
125I (Amersham, England) and 10 ul of chloramin T (0.5 mg/ml) for 60 s. The reaction
was stopped by adding 10 pl of sodium bisulphite (1 mg/ml) and 100 ul of potassium
iodide (10 mg/ml). L251-labelled protein A (‘251-PA) was separated from free iodine by
gel chromatography on a 0.8 X 20 cm Sephadex G-50 (Pharmacia) column. The
specific activity of this preparation was approximately 43 pCi/mg. The batch was then
diluted in PBS containing 40% FCS, to a concentration of 13 X 10 dpm/ml and stored
at -20°C for up to 4 mth, aliquoted in 4 ml volumes.
Preparation of anti-Sindbis antibodies in rabbits and chickens
Purified Sindbis virus (lOi PFU/ml) was inoculated twice intravenously, at 24
day intervals, into 6-mth-old, albino rabbits.
Chicken antibodies were prepared in two, 8- to lo-mth-old, laying hens, by 2
inoculations of approximately 5 X log PFU per ml of the CEF grown Sindbis virus
stock. The 2 inoculations were given intravenously at 40-day intervals. Blood was
taken at 0,8, 19,40,48, 55, and 78 days after the first inoculation. Eggs were collected
and kept at 4°C until used. For the determination of antibody content in eggs, the yolks
were separated from the whites, mixed with an equal volume of PBS containing 0.02%
sodium azide (PBS-AZ) and centrifuged 20 min at 10,OOOrpm in a Sorvall refrigerated
(4°C) centrifuge (Bar-Joseph and Malkinson, 1980). The supernatant thus obtained
was kept at -20°C until tested for antibodies.
SPA-RIA for rabbit antibodies The procedure was essentially the same as that described by Marier et al. (1979).
Microplates (polyvinyl chloride, 96 flat bottom wells, Dynatech Laboratories Inc.)
were coated with 0.2 ml per well of 10’ PFU per ml semi-purified Sindbis virus in 50
mM carbonate buffer and 0.02% AZ, pH 9.6 (coating buffer) at 4°C overnight. The
wells were then washed 3 times with 0.1% bovine serum albumin (BSA) in PBS-AZ
(PBS-BSA = blocking buffer). Duplicates (0.15 ml) of 5-fold rabbit serum dilutions
in PBS-AZ containing 0.05% Tween 20 (PBS-T) were added to the cells and incubated
at 37°C for 1 h. A known antibody-negative normal rabbit serum (NRS) was similarly
diluted and incubated in each test. After 3 cycles of washing with PBS-T the stock
solution of 1251-PA was diluted in PBS-T, so as to give 50,000 cpm (67,000 dpm) per
0.15 ml. This volume was added to the wells and incubated at 37°C for 1 h. After 4
cycles of washing with PBS-T, wells were numbered, cut from the plate and counted in
a gamma counter (Gammatic I, Kontron, Switzerland). Titration curves were then
drawn by plotting on semi-log paper the meancpmvaluesversus thereciprocalofserum
dilutions. From the NRS control a ‘cut-off curve was obtained by plotting the mean
cpm values to which 3 times the SD was added, versus the reciprocal of the NRS
dilutions. The titer of each serum was defined as the reciprocal of the serum dilution at
the intersection of the titration curve with the ‘cut-off curve.

62
SPA-RIA inhibition test for chicken antibodies
In the SPA-RIA inhibition test (SPA-RIA(IN)), chicken antibodies, first incubated
with the solid phase attached antigens, inhibit the binding of subsequently added,
rabbit antibodies that are directed to the same antigen. The inhibition is recognized by
the addition of “‘I-PA that binds only to rabbit antibodies. A decrease in the amount
of bound ‘251-PA, as compared to normal chick serum (NChS) or normal yolk (NY)
antibody-negative controls, is an indirect indication of the presence of chicken anti-
bodies.
The SPA-RIA(IN) test was performed as follows: Sindbis virus-coated microplate
wells prepared as for SPA-RIA, were incubated with 0.15 ml duplicate serial dilutions
(in PBS-T) of chicken sera or yolks, at 37°C for 1 h. As controls, a series of dilutions of
NChS or NY, negative for Sindbis virus antibodies, were used. The wells were then
washed 3 times with PBS-BSA. A predetermined rabbit anti-Sindbis virus serum
optimal dilution (1 : 1,000 in this study) was incubated (0.15 ml per well) at 37°C for 1
h. After 3 cycles of washes, ‘251-PA (50 000 cpm) was added and incubation continued > at 37°C for 1 h. After 4 cycles of washings, wells were prepared for the gamma counter
as decribed for SPA-RIA. From the data obtained, the percent of inhibition of bound
‘251-PA compared to controls (0% inhibition) was calculated and plotted on a semi-log
paper as a function of the sample dilution. The titer was defined as the reciprocal of the
sample dilution at the intersection of the curve with a 20% inhibition, arbitrarily
chosen ‘cut-off’ line.
SPA-RIA(S) for virus titration
The following procedure was used after defining optimal conditions: microplate
wells (as for SPA-RIA) were filled with 0.2 ml of a 1 : 1,000 dilution in coating buffer of
the pooled chicken anti-Sindbis virus antiserum (A-Sind-ChS) and incubated at room
temperature overnight. The cells were then washed 3 times with PBS-BSA. The coated
plates were dried and kept in sealed plastic bags at -70°C up to 1 yr.
For use, 0.15 ml of duplicate serial virus dilutions in PBS-T were incubated in wells
at 37°C for 1 h. As controls, 2 or more wells were incubated with PBS-T only ((-)virus
control). The wells were then washed 3 times with PBS-T and incubated at 37°C for 1 h
with 0.15 ml of a 1 : 1,000 dilution (in PBS-T) of rabbit anti-Sindbis virus antiserum
(A-Sind-RS). After additional 3 cycles of PBS-T washes, 0.15 ml of the conjugate
(50,000 cpm) were added to the wells and incubated at 37°C for 1 h. The wells were then
washed 4 times with PBS-T and counted in the gamma counter as described for
SPA-RIA.
The amount of bound lz51-PA was plotted on a semi-log (or log/log) paper as a
function of the reciprocal of the sample dilution. The titer was defined as the reciprocal
of sample dilution at the intersection with a cut-off line that equalled the mean cpm
values of the ‘(-)virus’ controls to which 3 times the SD was added. Alternatively and
preferably, the titer was determined using a TI-55-11 of TI-59 calculator (Texas
Instruments, Inc.) for computing a regression line from the log cpm bound 12’I-PA,

63
versus the log of the reciprocals of the sample dilutions. The titers were obtained from
the calculator program by introducing the cut-off value and finding the corresponding
reciprocal dilution value. Apart from titer values, the correlation coefficients, and the
slopes of the curves were computed from the same data. As a rule, the number of points
used for the regression line was never less than 3. Care was taken that the lowest
dilution used gave at least 2 times higher cpm than the ‘(-)virus’ controls.
RESULTS
Antibody response in sera and egg yolk of chickens inoculated with Sindbis virus
Chicken antibodies in sera or yolks were titrated by the SPA-RIA(IN) test. In typical
titration curves obtained with pooled sera and pooled egg yolks the titers obtained
with the 20% inhibition cut-off line were 16,000 for the serum and 10,000 for the yolk.
Maximal cpm mean values of the NChS and NY controls (0% inhibition) from which
the percent of inhibition values were calculated, were around 2,000 cpm.
Blood and eggs collected from the immunized chicken at different times after the
first inoculation were similarly titrated. The rise of SPA-RIA(IN) antibody titers in the
sera and egg yolks of the 2 chickens is shown at different times after incubation in Fig.
1. Relatively few eggs were laid during the follow-up period. The titers of sera and eggs
were similar, although as a rule the titer in the yolk was somewhat lower. Peak titers
were obtained at 8 days after the second inoculation. Six chicken sera which had titers
of 9,000 or higher were pooled (A-Sind-ChS); a similar yolk pool was prepared from
the 3 eggs with titers equal or higher than 9,000. Only the pooled chicken sera were
further used in this study.
Antibody response in rabbits inoculated with Sindbis virus One of the rabbits died of diarrhea within several days after the first injection. The
SPA-RIA antibody titers in sera of the surviving rabbit at 0, 24 and 34 days after the
first inoculation were 20, 10,000, and 530,000 respectively. The high titered serum,
obtained 34 days after the first inoculation, was the ‘A-Sind-RS’ preparation used
throughout the present study.
,Determination of the optimal chicken antiserum dilution for use in SPA-RIA(S) A series of titrations of Sindbis virus (stock 2 = 1 X lo9 PFU/ml) were performed in
microplates coated with a series of IO-fold chicken antiserum dilution from 10m2 to
10W6. The dilution of the rabbit antiserum was kept constant at 1: 1,000. From the
results (not shown) it was evident that with the 10m2, 10m3 and 10e4 dilutions of the
A-Sind-ChS similar titration curves were obtained. Significantly lower responses were
obtained with the lower A-Sind-ChS dilutions. Practically no response was obtained
with a 1: 1,000 dilution of a NRS control (used instead of the rabbit antiserum) in wells
coated with a 10e2 dilution of A-Sind-ChS. Some response, however, was obtained
when the rabbit antiserum was used in wells coated with a 10m2 dilution of randomly
pooled NChS.
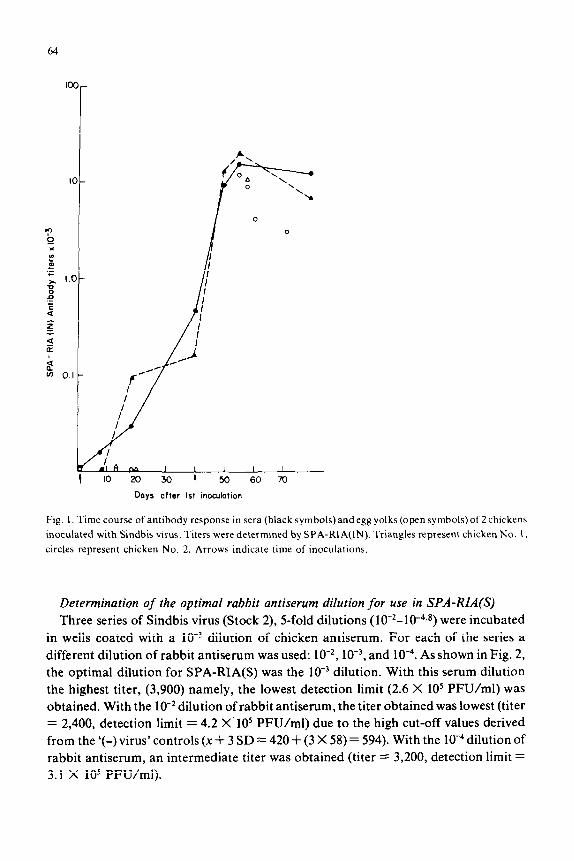
Days after 1st inowlation
Fig. 1. Time course of antibody response in sera (black symbols) and egg yolks (open symbols) of 2 chickens
inoculated with Sindbis virus. Titers were determined by SPA-RIA(IN). Triangles represent chicken No. I, circles represent chicken No. 2. Arrows indicate time of inoculations.
Determination of the optimal rabbit antiserum dilution for use in SPA-RIA(S) Three series of Sindbis virus (Stock 2), 5-fold dilutions ( 10-2-10-4.8) were incubated
in wells coated with a 10e3 dilution of chicken antiserum. For each of the series a
different dilution of rabbit antiserum was used: 10w2, 10m3, and 10m4. As shown in Fig. 2,
the optimal dilution for SPA-RIA(S) was the low3 dilution. With this serum dilution
the highest titer, (3,900) namely, the lowest detection limit (2.6 X lo5 PFU/ml) was
obtained. With the 10m2 dilution of rabbit antiserum, the titer obtained was lowest (titer
= 2,400, detection limit = 4.2 X lo5 PFU/ml) due to the high cut-off values derived
from the ‘(-)virus’ controls (x -I- 3 SD = 420 + (3 X 58)~ 594). With the low4 dilution of
rabbit antiserum, an intermediate titer was obtained (titer = 3,200, detection limit =
3.1 X lo5 PFU/ml).
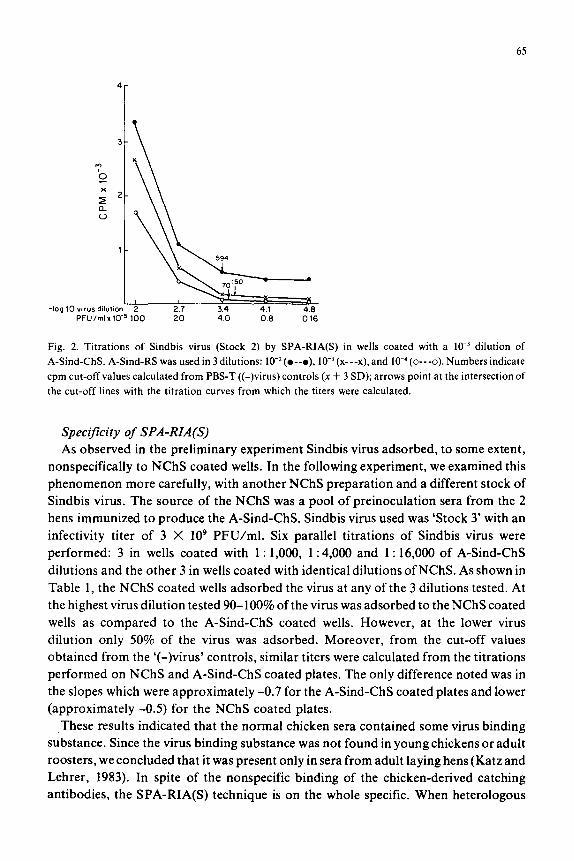
65
Fig. 2. Titrations of Sindbis virus (Stock 2) by SPA-RIA(S) in wells coated with a IO-’ dilution of
A-Sind-ChS. A-Sind-RS was used in 3 dilutions: 10-2(o--o), lo-’ (x---x), and 10e4 (o---o). Numbers indicate
cpm cut-off values calculated from PBS-T ((-) virus) controls (x + 3 SD); arrows point at the intersection of
the cut-off lines with the titration curves from which the titers were calculated.
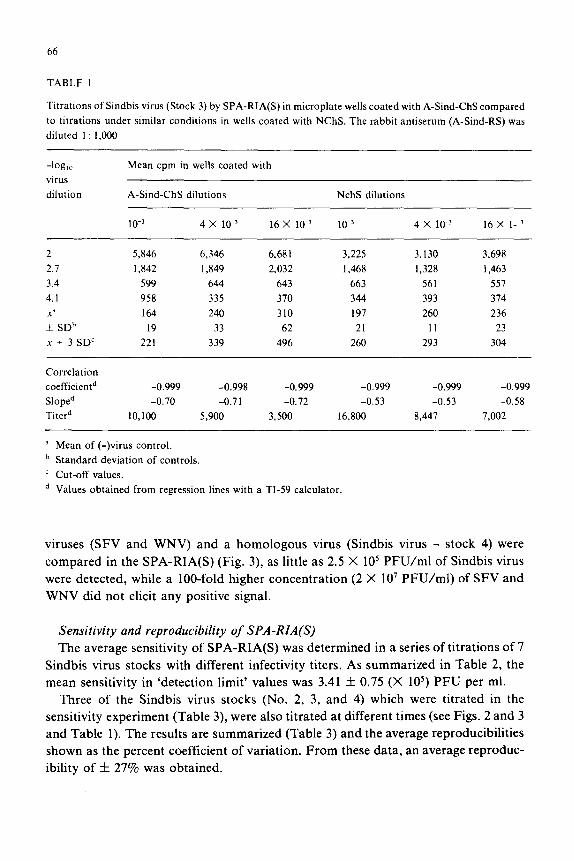
Specificity of SPA-RIA(S) As observed in the preliminary experiment Sindbis virus adsorbed, to some extent,
nonspecifically to NChS coated wells. In the following experiment, we examined this
phenomenon more carefully, with another NChS preparation and a different stock of
Sindbis virus. The source of the NChS was a pool of preinoculation sera from the 2
hens immunized to produce the A-Sind-ChS. Sindbis virus used was ‘Stock 3’ with an
infectivity titer of 3 X IO9 PFU/ml. Six parallel titrations of Sindbis virus were
performed: 3 in wells coated with 1 : 1,000, 1: 4,000 and 1: 16,000 of A-Sind-ChS
dilutions and the other 3 in wells coated with identical dilutions of NChS. As shown in
Table 1, the NChS coated wells adsorbed the virus at any of the 3 dilutions tested. At
the highest virus dilution tested 90-100% of the virus was adsorbed to the NChS coated
wells as compared to the A-Sind-ChS coated wells. However, at the lower virus
dilution only 50% of the virus was adsorbed. Moreover, from the cut-off values
obtained from the ‘(-)virus’ controls, similar titers were calculated from the titrations
performed on NChS and A-Sind-ChS coated plates. The only difference noted was in
the slopes which were approximately -0.7 for the A-Sind-ChS coated plates and lower
(approximately -0.5) for the NChS coated plates.
,These results indicated that the normal chicken sera contained some virus binding
substance. Since the virus binding substance was not found in young chickens or adult
roosters, we concluded that it was present only in sera from adult laying hens (Katz and
Lehrer, 1983). In spite of the nonspecific binding of the chicken-derived catching
antibodies, the SPA-RIA(S) technique is on the whole specific. When heterologous
66
TABLE 1
Titrations of Sindbis virus (Stock 3) by SPA-RIA(S) in microplate wells coated with A-Sind-ChS compared
to titrations under similar conditions in wells coated with NChS. The rabbit antiserum (A-Sind-RS) was
diluted 1 : 1,000
-log,, virus
dilution
Mean cpm in wells coated with
A-Sind-ChS dilutions NchS dilutions
10-r 4 x 10-r 16 X lo-’ 10-s 4 x 10-j 16 X I--’
2 5,846 6,346 6,681 3,225 3,130 3,698
2.1 1,842 1,849 2,032 1,468 1,328 1,463
3.4 599 644 643 663 561 557
4.1 958 335 370 344 393 374
x’ 164 240 310 197 260 236
+ SDb 19 33 62 21 11 23
x + 3 SDC 221 339 496 260 293 304
Correlation
coefficientd
Sloped
Titerd
-0.999 -0.998 -0.999 -0.999 -0.999 -0.999
-0.70 -0.71 -0.72 -0.53 -0.53 -0.58
10,100 5,900 3,500 16,800 8,447 7,002
a Mean of (-)virus control.
b Standard deviation of controls.
’ Cut-off values.
d Values obtained from regression lines with a TI-59 calculator.
viruses (SFV and WNV) and a homologous virus (Sindbis virus - stock 4) were
compared in the SPA-RIA(S) (Fig. 3), as little as 2.5 X lo5 PFU/ml of Sindbis virus
were detected, while a lOO-fold higher concentration (2 X 10’ PFU/ml) of SFV and
WNV did not elicit any positive signal.
Sensitivity and reproducibility of SPA-RIA(S) The average sensitivity of SPA-RIA(S) was determined in a series of titrations of 7
Sindbis virus stocks with different infectivity titers. As summarized in Table 2, the
mean sensitivity in ‘detection limit’ values was 3.41 f 0.75 (X 105) PFU per ml.
Three of the Sindbis virus stocks (No. 2, 3, and 4) which were titrated in the
sensitivity experiment (Table 3), were also titrated at different times (see Figs. 2 and 3
and Table 1). The results are summarized (Table 3) and the average reproducibilities
shown as the percent coefficient of variation. From these data, an average reproduc-
ibility of f 27% was obtained.
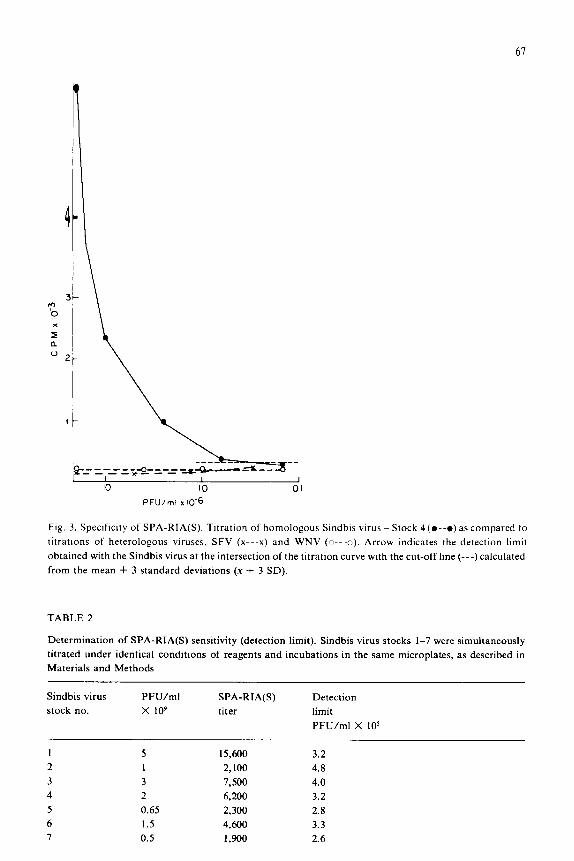
61
1
* h x
z a ”
3 c
-_=-___ 4 _=_-__ .&&-kL_ - -*- 1 I 1 IO IO 01
PFU/ml I 10‘6
Ftg. 3. Spectfictty of SPA-RIA(S). Titration of homologous Sindbis virus - Stock 4(0--o) as compared to
titrattons of heterologous viruses, SFV (x---x) and WNV (o---c). Arrow Indicates the detection limit
obtained with the Sindbis virus at the intersection of the titration curve with the cut-off line (---) calculated
from the mean + 3 standard deviations (x + 3 SD).
TABLE 2
Determination of SPA-RIA(S) sensitivity (detection limit). Sindbis virus stocks 1-7 were simultaneously
titrated under identical conditions of reagents and incubations in the same microplates, as described in
Materials and Methods
Sindbis virus PFU/ml
stock no. x 109
SPA-RIA(S)
titer
Detection
limit
PFU/ml X IO’
1 5 15,600 3.2
2 I 2,100 4.8
3 3 7,500 4.0
4 2 6,200 3.2
5 0.65 2,300 2.8
6 1.5 4,600 3.3
7 0.5 1,900 2.6

68
TABLE 3
Reproducibility of SPA-RIA(S) detection limit determinations. Each one of the tests of experiments 1 and 2
were performed at different days under similar conditions, as described in Materials and Methods
Sindbis virus
stock no.
Infectivity
titer
(PFU/ml
x 109)
Detection limit
(PFU/ml X 105)
Exp. 1 Exp. 2d Mean
f SD
CV (%)
2 1 2.6a 4.8 3.7 f 1.6 43
3 3 3.0b 4.0 3.5 f 0.7 20
4 2 2.5’ 3.2 2.8 i 0.5 32
Mean reproducibility (CV%) zlz SD = 31.6 * Il.
a Data from Fig. 2.
b Data from Table 1.
’ Data from Fig. 3.
d Data from Table 2.
CV = coefficient of variation [(SD/mean) X 1001.
DISCUSSION
Indirect solid phase immunoassays have several advantages over direct assays (Yol-
ken and Stopa, 1980; Devergne et al., 1981; Halonen and Meurman, 1982; Yolken,
1982; Richman et al., 1984). The tracer, which is usually a labelled anti-species IgG can
be used for the detection of many antigens, obviating the need of labelling individual
antibodies for each antigen. The indirect assays are potentially more sensitive than
direct assays because there are more layers of antibodies in the indirect tests. Indirect
assays are easier and quicker to apply, since the titer of the second antibody used may
be lower than the one needed for a sensitive direct test.
The use of protein A as a general tracer instead of anti-species antibodies makes
construction and application of indirect assays even easier. Protein A is commercially
available as a purified, highly active powder; it is comparable in its activity to affinity
purified anti-species antibodies. The use of protein A, therefore, saves time and effort
required for affinity chromatography. Moreover, the labelled protein A can be used
for the detection of antibodies from many animal species (Langone, 1982; Richman,
1983).
Another improvement which we advocate, is the use of laying hens as a source of
viral antibodies. Chickens are good antibody producers and antibodies can be derived
either from blood or egg yolks or both (Patterson et al., 1962; Scherer and Pancake,

69
in indirect assays using a chicken antibody in one layer and a mammalian antibody (Al
Moudallal et al., 1984; Yolken, 1982;‘Devergne et al., 1981) for the second layer.
We have described here all the steps necessary for the development of an indirect
solid phase protein A, sandwich type of radioimmunoassay for Sindbis virus (SPA-
RIA(S)). Only chicken antisera were used, but the possibility of using yolk antibodies
was confirmed as well (unpubl. results).
The versatility of the ‘Z51-labelled protein A was examplified by its use, not only for
the estimation of viral antigens, but also for the estimation of viral antibodies. We have
also demonstrated a method for titrating chicken antibodies, which do not bind protein
A directly, by an inhibition assay (SPA-RIA(IN)).
The average minimal amount of Sindbis virus, detectable with SPA-RIA(S), was
(3.41 f 0.75) X IO5 PFU/ml. Assuming each PFU contains 20-30 virions 3 X lo5
PFU/ml were equivalent to 0.3-0.45 ng/ml viral protein. This sensitivity was within
the sensitivity range obtained by solid phase immunoassays (Halonen and Meurman,
1982; Yolken, 1982; Richman, 1984). Replacing the iZ51-protein A of SPA-RIA(S) by
an alkaline-phosphatase labelled protein A conjugate, colorogenic (SPA-EIA(S)) and
fluorogenic (SPA-ELFA( enzyme immunoassays resulted. In our hands, SPA-
RIA(S) was approximately 6 times more sensitive than the correspondent colorogenic
SPA-EIA(S) (Katz and Lehrer, 1982; Katz et al., 1982), and about 2 times more
sensitive than the fluorogenic SPA-ELFA (Katz et al., unpubl. data). SPA-RIA(S)
was recently used by us for the detection of a variety of viral and non-viral antigens,
e.g. rabies virus (Katz et al., 1984), West Nile virus (Akov and Katz, unpubl. data),
bovine serum albumin (Shneerson-Porath and Katz, 1983), and phytohemagglutinin
(Rosenberg, Bino and Katz, unpubl. data). The use of protein A as the only general
tracer for all those antigens was shown to be both efficacious and economic.
An interesting, serendipitous finding was that normal sera from laying hens, used as
a control ‘capture’ layer, bound Sindbis virus nonspecifically. Preliminary findings
(Katz and Lehrer, 1983), on the nature of the virus bindings substance in normal sera
from laying hens, indicated that this substance is most probably vitellogenin (Deeley et
al., 1975). However, SPA-RIA(S) as a whole was specific due to the specificity of the
second layer of rabbit antibodies.
In this work, we have dealt only with Sindbis virus from tissue cultures as a model.
Whether SPA-RIA(S) will perform equally for the detection of viruses in clinical
specimens, remains to be determined.
REFERENCES
Al-Moudallal, Z., D. Altschuh, J.P. Briand and M.H.V. Van Regenmortel, 1984, J. Immunol. Methods 66,
35.
Altschuh, D., G. Hennache and M.H.V. Van Regenmortel, 1984, 69, 1.
Anastasiu, P. and P. Perrin, 1979, Ann. Microbial. (Paris), 130, 257.
Bade, H. and H. Stegemann, 1984, J. Immunol. Methods 72,421.
Bar-Joseph, M. and M. Malkinson, 1980, J. Virol. Methods 1. 179.

70
Butler, G.H., M.R. Proffit and B.C. Del Villano, 1979, J. Immunol. Methods 28, 293.
Colombatti, A. and J. Hilgers, 1979, J. Gen. Viral. 43, 395.
Deeley, R.G., K.P. Mullinix, W. Wetekam, H.M. Kronenberg, M. Meyers, J.D. Eldridge and R.F.
Goldberger, 1975, J. Biol. Chem. 250, 9060.
Devergne, J.C., L. Cardin, J. Burckard and M.H.V. Van Regenmortel, 1981, J. Viroi. Methods 3, 193.
Engvall, E. 1978, Stand. J. Immunol. (Suppl. 7) 8, 25.
Enzmann, P.J., 1978, J. Gen. Viral. 41, 641.
Gardner, P.S. and S. Kay, 1982, J. Viral. Methods 4, 257.
Goding, J.W., 1978, J. Immunol. Methods 20, 241.
Halonen, P. and 0. Meurman, 1982, in: New Developments in Practical Virology (Alan R. Liss, Inc., New
York) p. 83.
Hunter, W.M., 1973, in: Handbook of Experimental Immunology, ed. W. Weir (Blackwell Scientific
Publications, London) p. 1.
Jahrling, P.S., R.A. Hesse and J.F. Metzger, 1978, J. Clin. Microbial. 8, 54.
Katz, D. and A. Kohn, 1980, Abstract, 2nd International Conference on the Impact of Viral Diseases on the
Development of Africa and Middle East Countries, Nairobi, Kenya, p. 96.
Katz, D. and S. Lehrer, 1982, European Group for Rapid Viral Diagnosis, Meeting, St. Gallen, Switzerland,
Issue 4, p. 6.
Katz, D. and S. Lehrer, 1983, Israel J. Med. Sci. 19, 678.
Katz, D., E. Gruber and A. Kohn, 1981, European Group for Rapid Viral Diagnoses, Meeting, Vienna,
Austria, Issue 1, p. 8.
Katz, D., E. Gruber and S. Lehrer, 1981, Abstract, 5th International Congress of Virology, Strasbourg,
France, p. 169.
Katz, D., R. Oren, H. Ben-Moshe, 0. Galan and P. Fuchs, 1984, Abstract, 6th International Congress of
Virology, Sendai, Japan.
Katz, D., S. Lehrer, E. Gruber and A. Kohn, 1982, Israel J. Med. Sci. 18, 2.
Kronvall, G., U.S. Seal, S. Svensson and R.C. Williams, 1974, Acta Pathol. Microbial. Stand. Sect. B, 82,
12.
Langone, J.J., 1978, J. Immunol. Methods 24, 269.
Langone, J.J., 1980, J. Immunol. Methods 34, 93.
Langone, J.J., 1982, Adv. Immunol. 32, 157.
Langone, J.J., M.D.P. Boyle and T. Borsos, 1977, J. Immunol. Methods 18, 281.
Lindmark, R., K. Thoren-Tolling and J. Sjoquist, 1983, J. Immunol. Methods 62, 1.
Madore, P.H. and A. Baumgarten, 1979, J. Clin. Microbial. 4, 529.
Marier, R., M. Jansen and V.T. Andriole, 1979, J. Immunol. Methods 28, 41.
Patterson, R., J.S. Youngner, W.O. Weigle and F.J. Dixon, 1962, J. Immunol. 89, 772.
Pfefferkorn, E.R. and H.S. Hunter, 1963, Virology 20, 433.
Potgieter, L.N.D., B.T. Rouse and T.A. Webb-Martin, 1980, Am. J. Vet. Res. 41, 978.
Richman, D.D. 1983, Current Topics in Microbiology and Immunology, 104, 159.
Richman, D.D., P.H. Cleveland, M.N. Oxman and K.M. Johnson, 1982, J. Immunol. 128, 2300.
Richman, D.D., P.H. Cleveland, D.C. Redfield, M.N. Oxman and G.M. Wahl, 1984, J. Infect. Dis. 149,298.
Scherer, W.F. and B.A. Pancake, 1977, J. Clin. Microbial. 6, 578.
Shneerson-Porath, S. and D. Katz, 1983, Israel J. Med. Sci. 19, 678.
Sprouse, R.F., C.W. Caldwell and E.D. Everett, 1981, J. Clin. Microbial. 13, 66.
Van Regenmortel, M.H.V. and J. Burckard, 1980, Virology 106, 327.
Van Regenmortel, M.H.V., R. Yolken, G. Obert and J. Burckard, 1983, in: ImmunoenzymaticTechniques,
eds. S. Avrameas et al., p. 291.
Yolken, R.H., 1982, Rev. Infect. Dis. 4, 35.
Yolken, R.H. and F.J. Leister, 1981, J. Immunol. 43, 209.
Yolken, R.H. and P.J. Stopa, 1980, J. Clin. Microbial. 11, 546.





























