Embed Size (px)
Citation preview

University of Groningen
Dengue and Chikungunya virusvan Duijl-Richter, Mareike
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2016
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):van Duijl-Richter, M. (2016). Dengue and Chikungunya virus: Cell entry mechanisms and the impact ofantibodies on infectivity. [Groningen]: University of Groningen.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 28-08-2019

Chapter
Dengue Virus Life Cycle and Pathogenesis
2

Chapter 2
2
18

DENV Life Cycle and Pathogenesis
2
19
Dengue Virus Life Cycle and Pathogenesis
Introduction
Dengue virus (DENV) – mainly transmitted by the Aedes aegypti mosquito – is the most common mosquito-borne viral infection worldwide. DENV causes an estimated 390 million infections per year, of which about 100 million are symptomatic [1]. Each of the four serotypes of DENV (designated DENV 1-4) can cause disease with symptoms including fever, rash, headache, and musculoskeletal pain [2]. Approximately 500,000 to 1 million individuals however develop more severe disease including abdominal pain, bleedings, organ impairment, hematological aberrations, and plasma leakage. Severe dengue can be fatal if not properly treated, and each year about 25,000 patients die from dengue infection [3,4]. Severe dengue is almost exclusively seen during secondary infection with a heterotypic DENV serotype and during primary infection of infants with declining levels of maternal antibodies [5-7]. This observation points towards a pathogenic role of the immune system in severe disease development. In contrast, upon homotypic re-infection, the newly infecting DENV is effectively neutralized and individuals do not develop disease [8]. Also, third and tertiary infections usually do not lead to disease symptoms [9]. In this chapter, the viral structure and life cycle of DENV will be described, followed by a review of the host cell tropism and the mechanisms controlling disease outcome.
Viral structure & life cycle
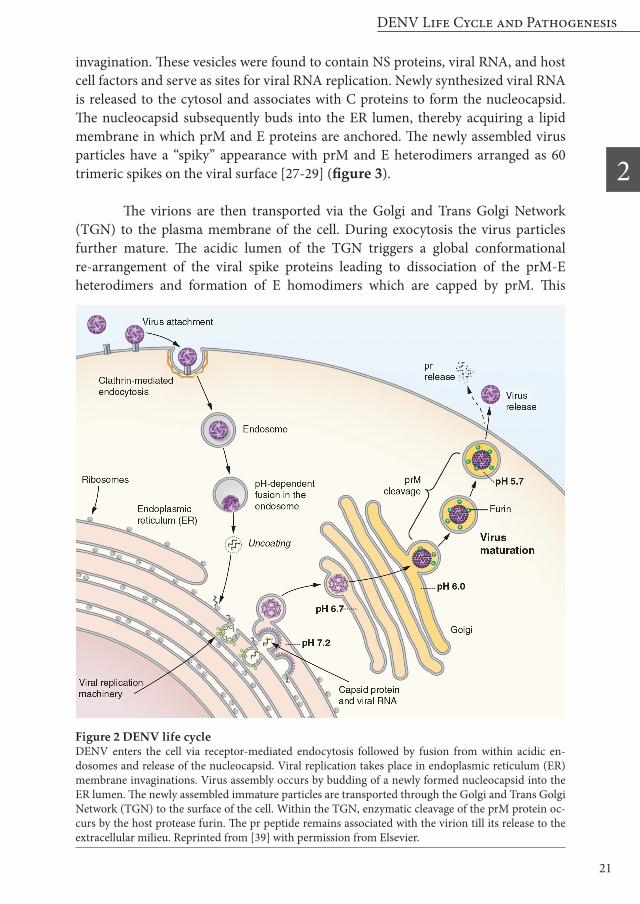
Dengue virus (DENV) belongs to the flavivirus genus within the Flaviviridae family. Other flaviviruses are for example West Nile virus (WNV), Yellow Fever virus (YFV), Japanese encephalitis virus (JEV), Tick-borne encephalitis virus (TBEV), and the emerging Zika virus (ZIKV) [10,11]. DENV virions are small spherical particles with a diameter of approximately 50 nm. Each virion contains a single-stranded, positive-sensed RNA genome of 10.8kB in length, which is packaged by the capsid (C) protein to form the nucleocapsid. The nucleocapsid is surrounded by a host-cell derived lipid bilayer in which 180 copies of the membrane protein (M) and envelope protein (E) are anchored. The E ectodomain consists of three structural domains (DI, DII and DIII). The E proteins are arranged as homodimers in a head-to-tail orientation. The homodimers are lying tangential to the viral membrane in a herringbone-like pattern, thereby giving the particle a “smooth” surface (figure 1 a,b) [12-15]. The first step in the viral life cycle (figure 2) involves binding of the E glycoprotein to a receptor or attachment factor on the host target cell. The putative

Chapter 2
2
20
receptor binding domain has been allocated to E DIII [16-19]. Upon binding, DENV particles enter the host cell via clathrin-mediated endocytosis. The internalized virions are delivered to Rab5-positive early endosomes, which further mature into Rab7-positive late endosomes [20,21]. Upon exposure of the virus to the acidic lumen of the endosome, the E protein homodimers dissociate, which results in the exposure of the fusion loop situated on DII (figure 1, green). The hydrophobic fusion peptide subsequently inserts into the endosomal membrane and E homotrimers are formed. Then, DIII folds back towards the fusion loop, which forces the opposing outer leaflets of the membranes together. The outer membranes subsequently merge, which is referred to as hemifusion. This short-lived intermediate is followed by complete fusion and expansion of the fusion pore. Finally, the nucleocapsid is released to the cytosol [12,22-24]. It has been suggested that fusion mainly occurs from within Rab7-positive late endosomes [20], although contrasting reports have been published considering the location of membrane fusion [25]. In addition to low pH, negatively charged lipids were shown to support membrane fusion of DENV. This observation strongly points towards fusion from within late endosomal compartments as negatively charged lipids are abundantly present within these compartments [26]. Upon the release of the nucleocapsid to the cytosol, the capsid proteins dissociate from the viral genome. The viral genome is translated at the rough endoplasmic reticulum (ER) into a single polyprotein and processed into 3 structural proteins (prM, E and C) and 7 non-structural (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) proteins. The ectodomains of prM and E, NS1 and parts of NS4A and NS4B are facing the ER lumen whereas the other proteins are present at the cytosolic side of the ER. NS4A, presumably together with other viral and host proteins, subsequently triggers membrane rearrangements and vesicle formation by ER membrane
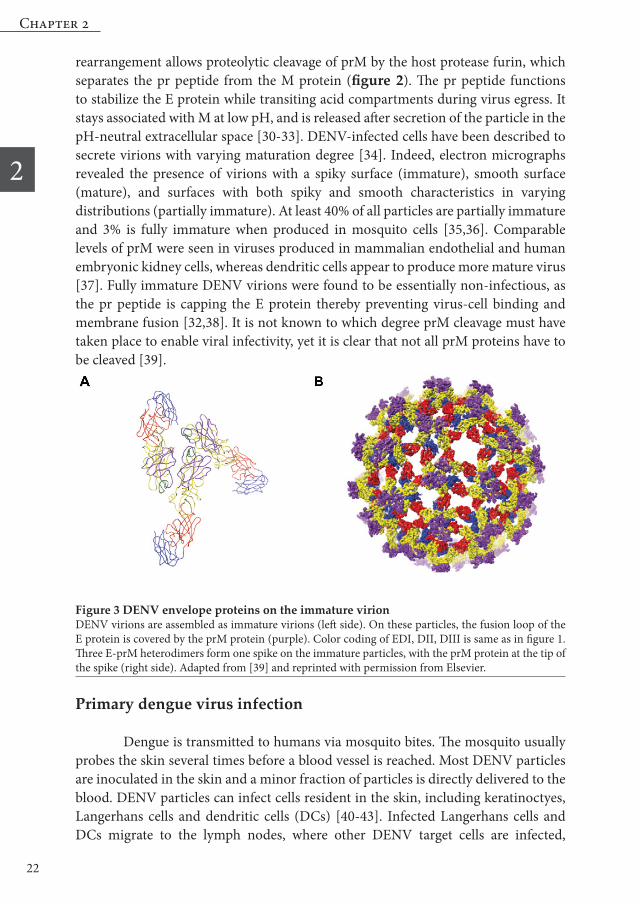
Figure 1 DENV envelope proteins on the mature virionThe E protein consists of three functional domains (DI, DII & DIII). Receptor-binding is presumably mediated by DIII (blue). The fusion loop (green) is located at the tip of DII (yellow). DI is depicted in red. On the mature virions, E homodimers lie flat on the membrane in a head-to-tail orientation, giv-ing the particle a smooth appearance. Adapted from [39] and reprinted with permission from Elsevier
DENV Life Cycle and Pathogenesis
2
21
invagination. These vesicles were found to contain NS proteins, viral RNA, and host cell factors and serve as sites for viral RNA replication. Newly synthesized viral RNA is released to the cytosol and associates with C proteins to form the nucleocapsid. The nucleocapsid subsequently buds into the ER lumen, thereby acquiring a lipid membrane in which prM and E proteins are anchored. The newly assembled virus particles have a “spiky” appearance with prM and E heterodimers arranged as 60 trimeric spikes on the viral surface [27-29] (figure 3).
The virions are then transported via the Golgi and Trans Golgi Network (TGN) to the plasma membrane of the cell. During exocytosis the virus particles further mature. The acidic lumen of the TGN triggers a global conformational re-arrangement of the viral spike proteins leading to dissociation of the prM-E heterodimers and formation of E homodimers which are capped by prM. This
Figure 2 DENV life cycle DENV enters the cell via receptor-mediated endocytosis followed by fusion from within acidic en-dosomes and release of the nucleocapsid. Viral replication takes place in endoplasmic reticulum (ER) membrane invaginations. Virus assembly occurs by budding of a newly formed nucleocapsid into the ER lumen. The newly assembled immature particles are transported through the Golgi and Trans Golgi Network (TGN) to the surface of the cell. Within the TGN, enzymatic cleavage of the prM protein oc-curs by the host protease furin. The pr peptide remains associated with the virion till its release to the extracellular milieu. Reprinted from [39] with permission from Elsevier.
Chapter 2
2
22
rearrangement allows proteolytic cleavage of prM by the host protease furin, which separates the pr peptide from the M protein (figure 2). The pr peptide functions to stabilize the E protein while transiting acid compartments during virus egress. It stays associated with M at low pH, and is released after secretion of the particle in the pH-neutral extracellular space [30-33]. DENV-infected cells have been described to secrete virions with varying maturation degree [34]. Indeed, electron micrographs revealed the presence of virions with a spiky surface (immature), smooth surface (mature), and surfaces with both spiky and smooth characteristics in varying distributions (partially immature). At least 40% of all particles are partially immature and 3% is fully immature when produced in mosquito cells [35,36]. Comparable levels of prM were seen in viruses produced in mammalian endothelial and human embryonic kidney cells, whereas dendritic cells appear to produce more mature virus [37]. Fully immature DENV virions were found to be essentially non-infectious, as the pr peptide is capping the E protein thereby preventing virus-cell binding and membrane fusion [32,38]. It is not known to which degree prM cleavage must have taken place to enable viral infectivity, yet it is clear that not all prM proteins have to be cleaved [39].
Primary dengue virus infection
Dengue is transmitted to humans via mosquito bites. The mosquito usually probes the skin several times before a blood vessel is reached. Most DENV particles are inoculated in the skin and a minor fraction of particles is directly delivered to the blood. DENV particles can infect cells resident in the skin, including keratinoctyes, Langerhans cells and dendritic cells (DCs) [40-43]. Infected Langerhans cells and DCs migrate to the lymph nodes, where other DENV target cells are infected,
Figure 3 DENV envelope proteins on the immature virion DENV virions are assembled as immature virions (left side). On these particles, the fusion loop of the E protein is covered by the prM protein (purple). Color coding of EDI, DII, DIII is same as in figure 1. Three E-prM heterodimers form one spike on the immature particles, with the prM protein at the tip of the spike (right side). Adapted from [39] and reprinted with permission from Elsevier.

DENV Life Cycle and Pathogenesis
2
23
including monocytes (which can also be directly infected by virus injected in the blood vessels) and macrophages [40,44,45]. Moreover, monocytes are recruited to the dermis were they differentiate into DCs that can be infected as well [46]. The virus then spreads to other parts of the body and becomes a systemic infection [2]. Although cells derived from the mononuclear lineage are the main target cells of DENV, also a variety of other cell types is permissive to infection, like for example cells of hepatic and endothelial origin or fibroblasts [47,48]. Such a wide cell tropism suggests that DENV uses several receptors or attachment factors. Indeed, molecules that were described to interact with DENV are (amongst others) heat shock protein 70, heat shock protein 90, GRP78/BiP, a 37/67kDa high-affinity laminin receptor, heparin sulphate, CD14, and C-type lectin receptors [45,49-58]. C-type lectin receptors are expressed on cells of the myeloid lineage. For example, Dendritic Cell Specific Intercellular adhesion molecule-3-Grabbing Non-integrin (DC-SIGN/CD209) is abundantly expressed on immature dendritic cells (imDC) and on certain types of macrophages [59]; the mannose receptor is mainly expressed on imDCs and macrophages, but also on dermal fibroblasts and keratinocytes [60]; and C-type lectin domain family 5, member A (CLEC5A/MDL-1) is expressed on monocytes and macrophages [58]. The infected cells produce large amounts of progeny virus, and approximately three days after the mosquito bite, viremia can be detected in the blood of infected patients. Peak titers are usually measured three days after the appearance of fever. Viral load can reach 106-108 viral copies/ml, though in patients with severe disease, higher titers are observed as well. DENV viremia is cleared on average 5 days after onset of fever [61,62]. Early in DENV infection the innate immune response is activated, which is important for restricting the infection and viral clearance [63,64]. The expression of IFNs and other pro-inflammatory cytokines is triggered by activation of pattern recognition receptors, including Toll-like receptor (TLR) 3, TLR 7 and TLR 8; but also the C-type lectins DC-SIGN, CLEC-5, and MR [58,65-68]. Subsequent binding of IFNs to infected and neighboring cells leads to activation of the JAK/STAT pathway and induces an antiviral state of the cell. Moreover, activation of natural killer (NK) cells leads to secretion of type-II IFNγ, which is likewise important in the antiviral defense [63,69]. DENV can however counteract the antiviral effects of IFNα/β binding by downregulating the JAK/STAT pathway [70]. Therefore, activation of the adaptive immune system is essential for complete clearance of infection. IFN production not only induces an antiviral state, it also stimulates the adaptive immune system by inducing imDC maturation and activation of lymphocytes [71]. Approximately 6 days after infection, DENV specific B and T cells appear. The majority of antibodies against DENV are generated against E and prM, which suggests that the secreted virus particles indeed consist of a mixture of virions with varying maturation state [72-74]. Serotype-specific, strongly neutralizing antibodies are often targeting the E protein and can efficiently prevent infection [75]. NS1 antibodies are produced as the NS1 protein is expressed on the surface of infected

Chapter 2
2
24
cells and secreted in the extracellular space [76,77]. NS1 antibodies were shown to mediate complement-mediated lysis of DENV infected cells [78,79]. Activated CD4+ and CD8+ T cells also induce lysis of DENV-infected target cells and produce a range of antiviral cytokines. Thus, infection is cleared through natural killer cells and macrophages, assisted by the presence of neutralizing antibodies and activated T cells [8,80].
Re-infection: Neutralization versus enhancement of infection Upon re-infection with DENV, the adaptive immune response is rapidly activated and anti-DENV antibodies are secreted. In case of a homologous re-infection, the pre-existing and newly produced antibodies efficiently prevent infection and together with a rapid activation of memory T-cells, the individual is protected from disease [81]. If heterotypic re-infection occurs shortly after primary infection (estimations range from six months up three years), cross-reactive antibodies are present in high titers thereby protecting the individual from disease [82,83]. However, if re-infection occurs after a longer time-span, the pre-existing cross-reactive antibodies are no longer protective against heterologous infection and may even enhance the severity of disease. Indeed, severe disease development is strongly associated with a heterologous secondary infection [62,84]. In contrast, third and tertiary DENV infections only rarely lead to disease as cross-immunity is then sufficient to prevent infection [9].
Role of antibodies in homologous re-infection
During homologous re-infection, antibodies play an important role in neutralization. NS1 antibodies have been described to induce complement-dependent lysis of infected cells [76] and lysis of antibody-opsonized viruses [85]. Antibodies directed against the viral spike proteins can directly neutralize viral infectivity. The dogma is that antibody-bound virions are taken up by Fc-receptor expressing cells and are delivered to the endocytic pathway for degradation. Importantly, Fc-receptors are expressed on myeloid cells, which are the main natural target cells of DENV. Therefore, in case of dengue, neutralization has to occur at a post-attachment step [86,87]. Indeed, multiple high-affinity antibodies were identified that neutralized infection by prevention of membrane fusion [88-90]. Most neutralizing antibodies were found to target the E protein. In murine studies, antibodies binding to the DIII domain were most potent, whereas in humans, antibodies targeting quaternary structures and cross-linking multiple E proteins have been described as strongly neutralizing. These antibodies likely act by preventing the essential conformational changes required for membrane fusion [37,91-95]. Neutralization of DENV can be described as a “multiple hit” phenomenon, in which a certain amount of available epitopes must have bound to prevent infection. For strongly neutralizing antibodies, only a fraction of epitopes has

DENV Life Cycle and Pathogenesis
2
25
to be covered to efficiently prevent infection, whereas weakly neutralizing antibodies have to bind more available sites to mediate neutralization [96]. Thus, in homotypic re-infection, type-specific antibodies displaying high affinity to the virus are present in high titers and prevent infection.
Enhanced disease during heterologous secondary infection
Approximately 2.5% of the individuals experiencing a secondary heterologous re-infection develop severe disease [2,3]. It is generally believed that antibodies play an important role in the development of severe disease as infants with declining levels of maternal antibodies were found to have an increased risk for severe disease during primary infection [7,84,97]. During a secondary heterologous re-infection, pre-existing cross-reactive antibodies can recognize newly infecting virus serotype. Functional studies revealed that cross-reactive antibodies generally have weakly neutralizing properties [72,73,98,99]. The presence of weakly-neutralizing cross-reactive antibodies is further increased through original antigenic sin, a phenomenon in which cross-reactive memory B-cells are expanded, but the number of immune cells against the currently remaining serotype remains low [8,73,98,100]. The cross-reactive antibodies bind to the virions, which are then internalized by Fc-receptor bearing DENV target cells. Intriguingly, these particles escape from degradation and fuse from within acidic endosomes [87]. This phenomenon is called antibody-dependent enhancement of infection (ADE). In vitro, all antibodies that can neutralize flavivirus infectivity also show enhancement of infection when antibody concentrations are low enough. Important factors in ADE are: antibody affinity, epitope accessibility, and antibody concentration. High affinity antibodies that bind to accessible epitopes are more prone to neutralization than low affinity antibodies that bind to cryptic sites. ADE occurs when the occupancy of viral particle by a neutralizing antibody is below the threshold of neutralization [39,74,96,101]. At ADE conditions, antibody-opsonized virions not only escape from degradation, these particles also have a higher chance to induce membrane fusion and productive infection [102]. This might be related to the alternative entry route by which antibody-opsonized DENV is taken up into the cell [103]. In addition, we and others showed that virtually non-infectious fully immature particles are rendered infectious by antibodies [104-106]. Upon entry of antibody-opsonized immature particles in Fc-receptor bearing target cells, furin cleavage and maturation occurs within the endosome, followed by infection of the cell [105,107]. ADE of infection of both mature and immature DENV particles leads to an enhanced infected cell mass and viral load early in infection. Epidemiological studies showed that an increased viral load is associated with a higher risk of developing severe dengue [61]. Furthermore, not only memory B cells but also memory T cells specific for the primary infecting serotype are preferentially expanded. Original antigenic sin of T cells leads to the generation of low avidity T cells that are less efficient in clearing

Chapter 2
2
26
the infection. This results in the secretion of a vast amount of pro-inflammatory cytokine and modulators which can cause capillary leakage as seen in severe dengue [2,8,73,98,108,109]. Though these and other aspects like virulence of the infecting strain and host genetic factors are clearly associated with severe disease, the complete picture is not understood given the small percentage of individuals with a heterologous secondary infection that develop severe disease [2,3].
Concluding remarks
DENV has a tremendous clinical impact, yet no antiviral treatment or vaccine has been approved. The development of an efficient and – most importantly – safe vaccine is challenging, as it is of upmost importance to prevent vaccine enhanced disease through ADE and original antigenic sin [110]. Therefore, an optimal vaccine has to neutralize all four DENV serotypes and elicit a long-term antibody response. The most promising candidate vaccine so far is from Sanofi Pasteur. Their tetravalent chimeric DENV/Yellow Fever (YF) vaccine (prM and E proteins of DENV and the replicative backbone of the YF vaccine 17D) reduced hospitalization by 67%-80% and dengue hemorrhagic fever by 80%-90% [111,112]. The efficacy to protect against natural DENV infection was surprisingly low (30%-60%, depending on the study population) [111-114]. The low protection against disease was rather unexpected, as antibody titers towards all DENV serotypes were found using the in vitro plaque reduction neutralization test (PRNT) [114,115]. The “golden standard” PRNT involves cells of endothelial origin [116]. Infection of these cells by DENV can be prevented by neutralizing antibodies. However, as endothelial cells do not express Fc-receptors, the enhancing properties of antibodies by Fc-receptor mediated entry cannot be assessed. Therefore, neutralization in DENV target cells might require higher titers than the ones found in standard PRNTs. Indeed, several groups have shown that Fc-receptor bearing target cells often require higher antibody titers for neutralization, especially when cross-reactive antibodies are tested [117-119]. For this, assays employing Fc-receptor bearing cells might represent a better predicting neutralization test for DENV, yet this approach is challenging in an industrial setting. Alternatively, the definition of the correlate of protection should be re-defined in retro perspective based on the results of the recent clinical trials, since the current correlate of protection of DENV is based on early vaccine-related studies on the flaviviruses JEV, YFV, and TBEV; but does not take the genetic variability between and within the different serotypes of DENV into account [110]. In summary, dengue is a complex disease and multiple factors are important in controlling disease outcome. More fundamental research into the virus life cycle and host immune response is imperative to fully understand disease pathogenesis and should guide the rational design of antiviral therapies and vaccines.

DENV Life Cycle and Pathogenesis
2
27
1. Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature. 2013.
2. Martina BE, Koraka P, Osterhaus AD. Dengue virus pathogenesis: an integrated view. Clin Microbiol Rev. 2009;22: 564-581.
3. World Health Organization. WHO Fact Sheet: Dengue and severe dengue. 2015. Available: http://www.who.int/mediacentre/factsheets/fs117/en/
4. Horstick O, Tozan Y, Wilder-Smith A. Reviewing dengue: still a neglected tropical disease? PLoS Negl Trop Dis. 2015;9: e0003632.
5. Guzman MG, Kouri G, Valdes L, Bravo J, Vazquez S, Halstead SB. Enhanced severity of secondary dengue-2 infections: death rates in 1981 and 1997 Cuban outbreaks. Rev Panam Salud Publica. 2002;11: 223-227.
6. Halstead SB. Neutralization and antibody-dependent enhancement of dengue viruses. Adv Virus Res. 2003;60: 421-467.
7. Kliks SC, Nimmanitya S, Nisalak A, Burke DS. Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. Am J Trop Med Hyg. 1988;38: 411-419.
8. Rothman AL. Immunity to dengue virus: a tale of original antigenic sin and tropical cytokine storms. Nat Rev Immunol. 2011.
9. Gibbons RV, Kalanarooj S, Jarman RG, Nisalak A, Vaughn DW, Endy TP, et al. Analysis of repeat hospital admissions for dengue to estimate the frequency of third or fourth dengue infections resulting in admissions and dengue hemorrhagic fever, and serotype sequences. Am J Trop Med Hyg. 2007;77: 910-913.
10. Musso D, Cao-Lormeau VM, Gubler DJ. Zika virus: following the path of dengue and chikungunya? Lancet. 2015;386: 243-244.
11. Kuno G, Chang GJ, Tsuchiya KR, Karabatsos N, Cropp CB. Phylogeny of the genus Flavivirus. J Virol. 1998;72: 73-83.
12. Mukhopadhyay S, Kuhn RJ, Rossmann MG. A structural perspective of the flavivirus life cycle. Nat Rev Microbiol. 2005;3: 13-22.
13. Heinz FX, Stiasny K. Flaviviruses and flavivirus vaccines. Vaccine. 2012;30: 4301-4306.
14. Zhang Y, Zhang W, Ogata S, Clements D, Strauss JH, Baker TS, et al. Conformational changes of the flavivirus E glycoprotein. Structure. 2004;12: 1607-1618.
15. Kuhn RJ, Zhang W, Rossmann MG, Pletnev SV, Corver J, Lenches E, et al. Structure of dengue virus: implications for flavivirus organization, maturation, and fusion. Cell. 2002;108: 717-725.
16. Hung JJ, Hsieh MT, Young MJ, Kao CL, King CC, Chang W. An external loop region of domain III of dengue virus type 2 envelope protein is involved in serotype-specific binding to mosquito but not mammalian cells. J Virol. 2004;78: 378-388.
17. Hung SL, Lee PL, Chen HW, Chen LK, Kao CL, King CC. Analysis of the steps involved in Dengue virus entry into host cells. Virology. 1999;257: 156-167.
18. Yu S, Wuu A, Basu R, Holbrook MR, Barrett AD, Lee JC. Solution structure and structural dynamics of envelope protein domain III of mosquito- and tick-borne flaviviruses. Biochemistry. 2004;43: 9168-9176.
19. Crill WD, Roehrig JT. Monoclonal antibodies that bind to domain III of dengue virus E glycoprotein are the most efficient blockers of virus adsorption to Vero cells. J Virol. 2001;75: 7769-7773.
20. van der Schaar HM, Rust MJ, Chen C, van der Ende-Metselaar H, Wilschut J, Zhuang X, et al. Dissecting the cell entry pathway of dengue virus by single-particle tracking in living cells. PLoS Pathog. 2008;4: e1000244.
21. Acosta EG, Castilla V, Damonte EB. Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-mediated endocytosis. J Gen Virol. 2008;89: 474-484.
22. Heinz FX, Allison SL. The machinery for flavivirus fusion with host cell membranes. Curr Opin Microbiol. 2001;4: 450-455.
23. Kielian M, Rey FA. Virus membrane-fusion proteins: more than one way to make a hairpin. Nat Rev Microbiol. 2006;4: 67-76.
24. Modis Y, Ogata S, Clements D, Harrison
References

Chapter 2
2
28
SC. Structure of the dengue virus envelope protein after membrane fusion. Nature. 2004;427: 313-319.
25. Krishnan MN, Sukumaran B, Pal U, Agaisse H, Murray JL, Hodge TW, et al. Rab 5 is required for the cellular entry of dengue and West Nile viruses. J Virol. 2007;81: 4881-4885.
26. Zaitseva E, Yang ST, Melikov K, Pourmal S, Chernomordik LV. Dengue virus ensures its fusion in late endosomes using compartment-specific lipids. PLoS Pathog. 2010;6: e1001131.
27. Welsch S, Miller S, Romero-Brey I, Merz A, Bleck CK, Walther P, et al. Composition and three-dimensional architecture of the dengue virus replication and assembly sites. Cell Host Microbe. 2009;5: 365-375.
28. Clyde K, Kyle JL, Harris E. Recent advances in deciphering viral and host determinants of dengue virus replication and pathogenesis J Virol. 2006;80: 11418-11431.
29. Apte-Sengupta S, Sirohi D, Kuhn RJ. Coupling of replication and assembly in flaviviruses. Curr Opin Virol. 2014;9: 134-142.
30. Li L, Lok SM, Yu IM, Zhang Y, Kuhn RJ, Chen J, et al. The flavivirus precursor membrane-envelope protein complex: structure and maturation. Science. 2008;319: 1830-1834.
31. Yu IM, Zhang W, Holdaway HA, Li L, Kostyuchenko VA, Chipman PR, et al. Structure of the immature dengue virus at low pH primes proteolytic maturation. Science. 2008;319: 1834-1837.
32. Yu IM, Holdaway HA, Chipman PR, Kuhn RJ, Rossmann MG, Chen J. Association of the pr peptides with dengue virus at acidic pH blocks membrane fusion. J Virol. 2009;83: 12101-12107.
33. Perera R, Kuhn RJ. Structural proteomics of dengue virus. Curr Opin Microbiol. 2008;11: 369-377.
34. Junjhon J, Lausumpao M, Supasa S, Noisakran S, Songjaeng A, Saraithong P, et al. Differential modulation of prM cleavage, extracellular particle distribution, and virus infectivity by conserved residues at nonfurin consensus positions of the dengue virus pr-M junction. J Virol. 2008;82: 10776-10791.
35. Plevka P, Battisti AJ, Junjhon J, Winkler
DC, Holdaway HA, Keelapang P, et al. Maturation of flaviviruses starts from one or more icosahedrally independent nucleation centres. EMBO Rep. 2011;12: 602-606.
36. Junjhon J, Edwards TJ, Utaipat U, Bowman VD, Holdaway HA, Zhang W, et al. Influence of pr-M cleavage on the heterogeneity of extracellular dengue virus particles J Virol. 2010;84: 8353-8358.
37. Dejnirattisai W, Wongwiwat W, Supasa S, Zhang X, Dai X, Rouvinski A, et al. A new class of highly potent, broadly neutralizing antibodies isolated from viremic patients infected with dengue virus. Nat Immunol. 2015;16: 170-177.
38. Zybert IA, van der Ende-Metselaar H, Wilschut J, Smit JM. Functional importance of dengue virus maturation: infectious properties of immature virions J Gen Virol. 2008;89: 3047-3051.
39. Pierson TC, Diamond MS. Degrees of maturity: the complex structure and biology of flaviviruses. Curr Opin Virol. 2012;2: 168-175.
40. Jessie K, Fong MY, Devi S, Lam SK, Wong KT. Localization of dengue virus in naturally infected human tissues, by immunohistochemistry and in situ hybridization. J Infect Dis. 2004;189: 1411-1418.
41. Limon-Flores AY, Perez-Tapia M, Estrada-Garcia I, Vaughan G, Escobar-Gutierrez A, Calderon-Amador J, et al. Dengue virus inoculation to human skin explants: an effective approach to assess in situ the early infection and the effects on cutaneous dendritic cells. Int J Exp Pathol. 2005;86: 323-334.
42. Wu SJ, Grouard-Vogel G, Sun W, Mascola JR, Brachtel E, Putvatana R, et al. Human skin Langerhans cells are targets of dengue virus infection. Nat Med. 2000;6: 816-820.
43. Marovich M, Grouard-Vogel G, Louder M, Eller M, Sun W, Wu SJ, et al. Human dendritic cells as targets of dengue virus infection. J Investig Dermatol Symp Proc. 2001;6: 219-224.
44. Durbin AP, Vargas MJ, Wanionek K, Hammond SN, Gordon A, Rocha C, et al. Phenotyping of peripheral blood mononuclear cells during acute dengue

DENV Life Cycle and Pathogenesis
2
29
illness demonstrates infection and increased activation of monocytes in severe cases compared to classic dengue fever. Virology. 2008;376: 429-435.
45. Miller JL, de Wet BJ, Martinez-Pomares L, Radcliffe CM, Dwek RA, Rudd PM, et al. The mannose receptor mediates dengue virus infection of macrophages. PLoS Pathog. 2008;4: e17.
46. Schmid MA, Harris E. Monocyte recruitment to the dermis and differentiation to dendritic cells increases the targets for dengue virus replication. PLoS Pathog. 2014;10: e1004541.
47. Suksanpaisan L, Cabrera-Hernandez A, Smith DR. Infection of human primary hepatocytes with dengue virus serotype 2. J Med Virol. 2007;79: 300-307.
48. Avirutnan P, Malasit P, Seliger B, Bhakdi S, Husmann M. Dengue virus infection of human endothelial cells leads to chemokine production, complement activation, and apoptosis. J Immunol. 1998;161: 6338-6346.
49. Reyes-Del Valle J, Chavez-Salinas S, Medina F, Del Angel RM. Heat shock protein 90 and heat shock protein 70 are components of dengue virus receptor complex in human cells. J Virol. 2005;79: 4557-4567.
50. Chen Y, Maguire T, Hileman RE, Fromm JR, Esko JD, Linhardt RJ, et al. Dengue virus infectivity depends on envelope protein binding to target cell heparan sulfate. Nat Med. 1997;3: 866-871.
51. Germi R, Crance JM, Garin D, Guimet J, Lortat-Jacob H, Ruigrok RW, et al. Heparan sulfate-mediated binding of infectious dengue virus type 2 and yellow fever virus. Virology. 2002;292: 162-168.
52. Chen YC, Wang SY, King CC. Bacterial lipopolysaccharide inhibits dengue virus infection of primary human monocytes/macrophages by blockade of virus entry via a CD14-dependent mechanism. J Virol. 1999;73: 2650-2657.
53. Jindadamrongwech S, Thepparit C, Smith DR. Identification of GRP 78 (BiP) as a liver cell expressed receptor element for dengue virus serotype 2. Arch Virol. 2004;149: 915-927.
54. Thepparit C, Smith DR. Serotype-specific entry of dengue virus into liver cells: identification of the 37-kilodalton/67-
kilodalton high-affinity laminin receptor as a dengue virus serotype 1 receptor. J Virol. 2004;78: 12647-12656.
55. Lozach PY, Burleigh L, Staropoli I, Navarro-Sanchez E, Harriague J, Virelizier JL, et al. Dendritic cell-specific intercellular adhesion molecule 3-grabbing non-integrin (DC-SIGN)-mediated enhancement of dengue virus infection is independent of DC-SIGN internalization signals. J Biol Chem. 2005;280: 23698-23708.
56. Dejnirattisai W, Webb AI, Chan V, Jumnainsong A, Davidson A, Mongkolsapaya J, et al. Lectin switching during dengue virus infection. J Infect Dis. 2011;203: 1775-1783.
57. Navarro-Sanchez E, Altmeyer R, Amara A, Schwartz O, Fieschi F, Virelizier JL, et al. Dendritic-cell-specific ICAM3-grabbing non-integrin is essential for the productive infection of human dendritic cells by mosquito-cell-derived dengue viruses. EMBO Rep. 2003;4: 723-728.
58. Chen ST, Lin YL, Huang MT, Wu MF, Cheng SC, Lei HY, et al. CLEC5A is critical for dengue-virus-induced lethal disease. Nature. 2008;453: 672-676.
59. Geijtenbeek TB, Gringhuis SI. Signalling through C-type lectin receptors: shaping immune responses. Nat Rev Immunol. 2009;9: 465-479.
60. Sheikh H, Yarwood H, Ashworth A, Isacke CM. Endo180, an endocytic recycling glycoprotein related to the macrophage mannose receptor is expressed on fibroblasts, endothelial cells and macrophages and functions as a lectin receptor. J Cell Sci. 2000;113 ( Pt 6): 1021-1032.
61. Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, Suntayakorn S, et al. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J Infect Dis. 2000;181: 2-9.
62. Halstead SB (2007) Dengue. The Lancet 370: 1644–1652.
63. Shresta S, Kyle JL, Snider HM, Basavapatna M, Beatty PR, Harris E. Interferon-dependent immunity is essential for resistance to primary dengue virus infection in mice, whereas T- and B-cell-dependent immunity are less critical. J Virol. 2004;78: 2701-2710.
64. Diamond MS, Roberts TG, Edgil D, Lu B,

Chapter 2
2
30
Ernst J, Harris E. Modulation of Dengue virus infection in human cells by alpha, beta, and gamma interferons. J Virol. 2000;74: 4957-4966.
65. Fernandez-Garcia MD, Mazzon M, Jacobs M, Amara A. Pathogenesis of flavivirus infections: using and abusing the host cell Cell Host Microbe. 2009;5: 318-328.
66. de Kruif MD, Setiati TE, Mairuhu AT, Koraka P, Aberson HA, Spek CA, et al. Differential gene expression changes in children with severe dengue virus infections. PLoS Negl Trop Dis. 2008;2: e215.
67. Tsai YT, Chang SY, Lee CN, Kao CL. Human TLR3 recognizes dengue virus and modulates viral replication in vitro. Cell Microbiol. 2009;11: 604-615.
68. Akira S. TLR signaling. Curr Top Microbiol Immunol. 2006;311: 1-16.
69. Azeredo EL, De Oliveira-Pinto LM, Zagne SM, Cerqueira DI, Nogueira RM, Kubelka CF. NK cells, displaying early activation, cytotoxicity and adhesion molecules, are associated with mild dengue disease. Clin Exp Immunol. 2006;143: 345-356.
70. Munoz-Jordan JL, Fredericksen BL. How flaviviruses activate and suppress the interferon response. Viruses. 2010;2: 676-691.
71. Erickson AK, Gale M,Jr. Regulation of interferon production and innate antiviral immunity through translational control of IRF-7. Cell Res. 2008;18: 433-435.
72. Beltramello M, Williams KL, Simmons CP, Macagno A, Simonelli L, Quyen NT, et al. The human immune response to Dengue virus is dominated by highly cross-reactive antibodies endowed with neutralizing and enhancing activity. Cell Host Microbe. 2010;8: 271-283.
73. Mathew A, West K, Kalayanarooj S, Gibbons RV, Srikiatkhachorn A, Green S, et al. B-cell responses during primary and secondary dengue virus infections in humans. J Infect Dis. 2011;204: 1514-1522.
74. Rodenhuis-Zybert IA, Wilschut J, Smit JM. Dengue virus life cycle: viral and host factors modulating infectivity Cell Mol Life Sci. 2010;67: 2773-2786.
75. Wahala WM, Silva AM. The human antibody response to dengue virus infection. Viruses.
2011;3: 2374-2395.
76. Muller DA, Young PR. The flavivirus NS1 protein: molecular and structural biology, immunology, role in pathogenesis and application as a diagnostic biomarker. Antiviral Res. 2013;98: 192-208.
77. Amorim JH, Alves RP, Boscardin SB, Ferreira LC. The dengue virus non-structural 1 protein: risks and benefits. Virus Res. 2014;181: 53-60.
78. Henchal EA, Henchal LS, Schlesinger JJ. Synergistic interactions of anti-NS1 monoclonal antibodies protect passively immunized mice from lethal challenge with dengue 2 virus. J Gen Virol. 1988;69 ( Pt 8): 2101-2107.
79. Kurosu T, Chaichana P, Yamate M, Anantapreecha S, Ikuta K. Secreted complement regulatory protein clusterin interacts with dengue virus nonstructural protein 1. Biochem Biophys Res Commun. 2007;362: 1051-1056.
80. St John AL, Rathore AP, Yap H, Ng ML, Metcalfe DD, Vasudevan SG, et al. Immune surveillance by mast cells during dengue infection promotes natural killer (NK) and NKT-cell recruitment and viral clearance. Proc Natl Acad Sci U S A. 2011;108: 9190-9195.
81. Rothman AL, Medin CL, Friberg H, Currier JR. Immunopathogenesis Versus Protection in Dengue Virus Infections. Curr Trop Med Rep. 2014;1: 13-20.
82. Reich NG, Shrestha S, King AA, Rohani P, Lessler J, Kalayanarooj S, et al. Interactions between serotypes of dengue highlight epidemiological impact of cross-immunity. J R Soc Interface. 2013;10: 20130414.
83. SABIN AB. Research on dengue during World War II. Am J Trop Med Hyg. 1952;1: 30-50.
84. Halstead SB, O’Rourke EJ. Dengue viruses and mononuclear phagocytes. I. Infection enhancement by non-neutralizing antibody. J Exp Med. 1977;146: 201-217.
85. Shresta S. Role of complement in dengue virus infection: protection or pathogenesis? MBio. 2012;3: 10.1128/mBio.00003-12. Print 2012.
86. Fischl W, Bartenschlager R. Exploitation of cellular pathways by Dengue virus. Curr

DENV Life Cycle and Pathogenesis
2
31
Opin Microbiol. 2011;14: 470-475.
87. van der Schaar HM, Wilschut JC, Smit JM. Role of antibodies in controlling dengue virus infection. Immunobiology. 2009;214: 613-629.
88. Cockburn JJ, Navarro Sanchez ME, Goncalvez AP, Zaitseva E, Stura EA, Kikuti CM, et al. Structural insights into the neutralization mechanism of a higher primate antibody against dengue virus. EMBO J. 2011.
89. Lok SM. Human antibodies stop dengue virus by jamming its mechanics. Proc Natl Acad Sci U S A. 2014;111: 1670-1671.
90. Costin JM, Zaitseva E, Kahle KM, Nicholson CO, Rowe DK, Graham AS, et al. Mechanistic study of broadly neutralizing human monoclonal antibodies against dengue virus that target the fusion loop. J Virol. 2013;87: 52-66.
91. de Alwis R, Smith SA, Olivarez NP, Messer WB, Huynh JP, Wahala WM, et al. Identification of human neutralizing antibodies that bind to complex epitopes on dengue virions. Proc Natl Acad Sci U S A. 2012;109: 7439-7444.
92. Crill WD, Chang GJ. Localization and characterization of flavivirus envelope glycoprotein cross-reactive epitopes. J Virol. 2004;78: 13975-13986.
93. Smith SA, de Alwis AR, Kose N, Jadi RS, de Silva AM, Crowe JE,Jr. Isolation of dengue virus-specific memory B cells with live virus antigen from human subjects following natural infection reveals the presence of diverse novel functional groups of antibody clones. J Virol. 2014.
94. Fibriansah G, Tan JL, Smith SA, de Alwis AR, Ng TS, Kostyuchenko VA, et al. A potent anti-dengue human antibody preferentially recognizes the conformation of E protein monomers assembled on the virus surface. EMBO Mol Med. 2014;6: 358-371.
95. Fibriansah G, Ibarra KD, Ng TS, Smith SA, Tan JL, Lim XN, et al. DENGUE VIRUS. Cryo-EM structure of an antibody that neutralizes dengue virus type 2 by locking E protein dimers. Science. 2015;349: 88-91.
96. Pierson TC, Xu Q, Nelson S, Oliphant T, Nybakken GE, Fremont DH, et al. The stoichiometry of antibody-mediated neutralization and enhancement of West
Nile virus infection. Cell Host Microbe. 2007;1: 135-145.
97. Halstead SB. Antibodies determine virulence in dengue Ann N Y Acad Sci. 2009;1171 Suppl 1: E48-56.
98. Zompi S, Montoya M, Pohl MO, Balmaseda A, Harris E. Dominant Cross-Reactive B Cell Response during Secondary Acute Dengue Virus Infection in Humans. PLoS Negl Trop Dis. 2012;6: e1568.
99. de Alwis R, Beltramello M, Messer WB, Sukupolvi-Petty S, Wahala WM, Kraus A, et al. In-depth analysis of the antibody response of individuals exposed to primary dengue virus infection. PLoS Negl Trop Dis. 2011;5: e1188.
100. Halstead SB, Rojanasuphot S, Sangkawibha N. Original antigenic sin in dengue. Am J Trop Med Hyg. 1983;32: 154-156.
101. Dowd KA, Jost CA, Durbin AP, Whitehead SS, Pierson TC. A dynamic landscape for antibody binding modulates antibody-mediated neutralization of West Nile virus. PLoS Pathog. 2011;7: e1002111.
102. Flipse J, Diosa-Toro MA, Hoornweg TE, Ayala-Núñez NV, van de Pol DPI, Urcuqui-Inchima S, et al. Antibody-dependent enhancement of dengue virus infection in primary human macrophages; balancing higher fusion against antiviral responses. In: Molecular Mechanisms of Dengue Virus Infection - Cell Tropism, Antibody-Dependent Enhancement, and Cytokines. 2015.
103. Ayala-Nunez NV, Hoornwega TE, van de Pol DPI, Sjollema KA, van der Schaar H, Smit JM. Seeing is believing: how antibodies alter the cell entry pathway of dengue virus particles in living macrophages. In: Dengue virus cell entry - unraveling the role of antibodies, maturation status, and antiviral drugs. 2014.
104. Dejnirattisai W, Jumnainsong A, Onsirisakul N, Fitton P, Vasanawathana S, Limpitikul W, et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science. 2010;328: 745-748.
105. Rodenhuis-Zybert IA, van der Schaar HM, da Silva Voorham JM, van der Ende-Metselaar H, Lei HY, Wilschut J, et al. Immature dengue virus: a veiled pathogen?

Chapter 2
2
32
PLoS Pathog. 2010;6: e1000718.
106. da Silva Voorham JM, Rodenhuis-Zybert IA, Ayala Nunez NV, Colpitts TM, van der Ende-Metselaar H, Fikrig E, et al. Antibodies against the envelope glycoprotein promote infectivity of immature dengue virus serotype 2. PLoS One. 2012;7: e29957.
107. Zheng A, Umashankar M, Kielian M. In vitro and in vivo studies identify important features of dengue virus pr-E protein interactions. PLoS Pathog. 2010;6: e1001157.
108. Friberg H, Bashyam H, Toyosaki-Maeda T, Potts JA, Greenough T, Kalayanarooj S, et al. Cross-reactivity and expansion of dengue-specific T cells during acute primary and secondary infections in humans. Sci Rep. 2011;1: 51.
109. Cardier JE, Marino E, Romano E, Taylor P, Liprandi F, Bosch N, et al. Proinflammatory factors present in sera from patients with acute dengue infection induce activation and apoptosis of human microvascular endothelial cells: possible role of TNF-alpha in endothelial cell damage in dengue. Cytokine. 2005;30: 359-365.
110. Flipse J, Smit JM. The Complexity of a Dengue Vaccine: A Review of the Human Antibody Response. PLoS Negl Trop Dis. 2015;9: e0003749.
111. Villar L, Dayan GH, Arredondo-Garcia JL, Rivera DM, Cunha R, Deseda C, et al. Efficacy of a tetravalent dengue vaccine in children in Latin America. N Engl J Med. 2015;372: 113-123.
112. Villar LA, Rivera-Medina DM, Arredondo-Garcia JL, Boaz M, Starr-Spires L, Thakur M, et al. Safety and immunogenicity of a recombinant tetravalent dengue vaccine in 9-16 year olds: a randomized, controlled,
phase II trial in Latin America. Pediatr Infect Dis J. 2013;32: 1102-1109.
113. Capeding MR, Tran NH, Hadinegoro SR, Ismail HI, Chotpitayasunondh T, Chua MN, et al. Clinical efficacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: a phase 3, randomised, observer-masked, placebo-controlled trial. Lancet. 2014;384: 1358-1365.
114. Sabchareon A, Wallace D, Sirivichayakul C, Limkittikul K, Chanthavanich P, Suvannadabba S, et al. Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in Thai schoolchildren: a randomised, controlled phase 2b trial. Lancet. 2012.
115. Wilder-Smith A. Dengue vaccines: dawning at last? Lancet. 2014;384: 1327-1329.
116. Roehrig JT, Hombach J, Barrett AD. Guidelines for Plaque-Reduction Neutralization Testing of Human Antibodies to Dengue Viruses. Viral Immunol. 2008;21: 123-132.
117. Wu RS, Chan KR, Tan HC, Chow A, Allen JC,Jr, Ooi EE. Neutralization of dengue virus in the presence of Fc receptor-mediated phagocytosis distinguishes serotype-specific from cross-neutralizing antibodies. Antiviral Res. 2012;96: 340-343.
118. Moi ML, Lim CK, Chua KB, Takasaki T, Kurane I. Dengue virus infection-enhancing activity in serum samples with neutralizing activity as determined by using FcgammaR-expressing cells. PLoS Negl Trop Dis. 2012;6: e1536.
119. Chawla T, Chan KR, Zhang SL, Tan HC, Lim AP, Hanson BJ, et al. Dengue virus neutralization in cells expressing Fc gamma receptors. PLoS One. 2013;8: e65231.

DENV Life Cycle and Pathogenesis
2
33






























