Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO
BIOLOGIA DE AGENTES INFECCIOSOS E PARASITÁRIOS
MORFOLOGIA E FILOGENIA DE MYXOZOA EM Rhamdia quelen (QUOY &
GAIMARD, 1824) (TELEOSTEI, PIMELODIDAE) DA ILHA DO MARAJÓ, REGIÃO
AMAZÔNICA
JACQUELINE POMPEU ABRUNHOSA
Belém-Pará
2018

JACQUELINE POMPEU ABRUNHOSA
MORFOLOGIA E FILOGENIA DE MYXOZOA EM Rhamdia quelen (QUOY &
GAIMARD, 1824) (TELEOSTEI, PIMELODIDAE) DA ILHA DO MARAJÓ, REGIÃO
AMAZÔNICA
Tese apresentada ao Programa de Pós-graduação
em Biologia de Agentes Infecciosos e
Parasitários do Instituto de Ciências Biológicas
da Universidade Federal do Pará como requisito
para obtenção do título de Doutor em Biologia
de Agentes Infecciosos e Parasitários.
Orientador: Prof. Dr. Edilson Matos
Co-orientador: Prof. Dr. Igor Hamoy
Belém – Pará
2018

2
JACQUELINE POMPEU ABRUNHOSA
MORFOLOGIA E FILOGENIA DE MYXOZOA EM Rhamdia quelen (QUOY &
GAIMARD, 1824) (TELEOSTEI, PIMELODIDAE) DA ILHA DE MARAJÓ, REGIÃO
AMAZÔNICA
Tese apresentada ao Programa de Pós-graduação em Biologia de Agentes Infecciosos e
Parasitários do Instituto de Ciências Biológicas da Universidade Federal do Pará, como
requisito para obtenção do grau de Doutor em Biologia de Agentes Infecciosos e Parasitários.
Orientador: Prof. Dr. Edilson Matos
Laboratório de Pesquisa Carlos Azevedo, ISPA-UFRA
Co-orientador: Prof. Dr. Igor Hamoy
Laboratório de Genética Aplicada, ISARH-UFRA
Banca Examinadora: Prof. Dr. Adriano Furtado
Laboratório de Biologia Celular e Helmintologia, ICB-UFPA
Profa. Dra. Cristiana Ramalho Maciel
Laboratório de Aquicultura, IECOS-UFPA, Campus Bragança
Profa. Dra. Elane Guerreiro Giese
Laboratório de Histologia e Embriologia Animal, ISPA-UFRA
Prof. Dr. Luís Fernando Rodrigues Filho
Universidade Federal Rural da Amazônia, Campus Capanema
Prof. Dr. Moacir Cerqueira (Suplente)
Universidade Federal Rural da Amazônia
Belém, 8 de Maio de 2018.

3
Dados Internacionais de Catalogação na Publicação (CIP)
Sistema de Bibliotecas da Universidade Federal do Pará
Gerada automaticamente pelo módulo Ficat, mediante os dados fornecidos pelo(a) autor(a)
P788m Pompeu Abrunhosa, Jacqueline
Morfologia e Filogenia de Myxozoa em Rhamdia quelen (Quoy & Gaimard, 1824) (Teleostei,
Pimelodidae) da Ilha do Marajó, Região Amazônica / Jacqueline Pompeu Abrunhosa. — 2018
99 f. : il. color
Tese (Doutorado) - Programa de Pós-graduação em Biologia de Agentes Infecciosos e Parasitários
(PPGBAIP), Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, 2018.
Orientação: Prof. Dr. Edilson Rodrigues Matos
Coorientação: Prof. Dr. Igor Guerreiro Hamoy.
1. microparasito, Henneguya, Myxobolus, 18S rDNA, jundiá. I. Rodrigues Matos, Edilson , orient. II. Título
CDD 571.999 Powered by TC PDF ( www.tcpdf.org)

4
Ofereço a Deus, que rege todo o universo, pois
nele foram criadas todas as coisas nos céus e na
terra, as visíveis e as invisíveis, e sem Ele nada
do que foi feito se fez! Toda honra, glória e
louvor!!
(João 1:3; Colossensses 1:16)

5
Dedico ao meu companheiro Fernando
e aos nossos príncipes Vinícius e Dário

6
AGRADECIMENTOS
Ao Prof. Dr. Edilson Matos por ter aceito orientar esta Tese. Agradeço a todos os
ensinamentos científicos, foi um grande aprendizado.
Ao Prof. Dr. Igor Hamoy por ter aceitado a co-orientação nas análises de biologia
molecular no Laboratório de Genética Aplicada (LGA/UFRA).
Aos Profs. Dr. Sidney Santos e Dra. Ândrea Kelly por concederem o do uso do
Laboratório de Genética Humana e Aplicada (UFPA) para sequenciamento de parte das
amostras.
Ao MSc. José Sindeaux Neto por todo apoio nas técnicas de laboratório, sempre
pronto com uma palavra de incentivo.
Aos parceiros Dra Michele Velasco, Patrícia Sacco, MSc. Márcia Sacco, Elideth
Pacheco, Weverton por toda ajuda na execução dos trabalhos de laboratório e grande amizade.
A equipe do LPCA pelos auxílios no laboratório Joyce, Débora, Diehgo, Lenize,
Danielle.
A equipe do Laboratório de Genética Aplicada (LGA) Paola, Sávio, Kaio, Renato,
Sayume por todos os auxílios no laboratório.
Ao MSc Marcos Antônio Trindade Amador do Laboratório de Genética Humana e
Aplicada (UFPA) por sua atenção e presteza no sequenciamento das amostras.
Prof. Dr. Osimar Sanches pelo auxílio nas análises histopatológicas.
Ao Sr Pedrinho pelo auxílio nas coletas de campo.
Ao Sr. Vavá (in memorian) e Euriana por cederem sua casa em Salvaterra para auxílio
a equipe do LPCA e a todas as pessoas que colaboraram com a coleta dos peixes em
Salvaterra e Cachoeira do Arari.

7
SUMÁRIO
LISTA DE SIGLAS .................................................................................................................. 8
RESUMO ................................................................................................................................... 9
ABSTRACT ............................................................................................................................ 10
1. INTRODUÇÃO GERAL .................................................................................. 11
1.1. HOSPEDEIRO .................................................................................................... 12
1.2. IMPORTÂNCIA DO ESTUDO DE ICTIOPARASITAS .................................. 15
1.3. FILO CNIDARIA (Hatschek, 1888) ................................................................... 15
1.3.1 Subfilo Myxozoa Grassé, 1970 .......................................................................... 17
1.3.2 Filogenia do Myxozoa ....................................................................................... 22
1.3.3 Myxobolus e Henneguya....................................................................................24
1.3.3.1 Gênero Myxobolus Butschli, 1882 ................................................................... 25
1.3.3.2 Gênero Henneguya Thélohan, 1892 ................................................................ 24
1.3.5 JUSTIFICATIVA................................................................................................26
2. OBJETIVOS ...................................................................................................26
3. APRESENTAÇÃO DOS ARTIGOS................................................................27
3.1 ARTIGO I .......................................................................................................... 27
3.2 ARTIGO II ......................................................................................................... 34
3.3 ARTIGO III ....................................................................................................... 42
3.4 ARTIGO IV ....................................................................................................... 62
4. CONCLUSÃO ................................................................................................... 82
REFERÊNCIAS BIBLIOGRÁFICAS .................................................................................. 83
ANEXO: Normas das revistas Zootaxa
Parasitology Research
Declaração da CEUA

8
LISTA DE SIGLAS
CEUA - Comissão de Ética no Uso Animal
CP - Cápsula Polar
ICB - Instituto de Ciências Biológicas
ISPA - Instituto da Saúde e Produção Animal
LGA - Laboratório de Genética Aplicada
LPCA - Laboratório de Pesquisa Carlos Azevedo
LPEM - Laboratório de Pesquisa Edilson Matos
UFPA - Universidade Federal do Pará
UFRA - Universidade Federal Rural da Amazônia

9
RESUMO
Rhamdia quelen (Quoy & Gaimard, 1824) é um bagre de água doce com ampla distribuição,
na América Central e Sul, ocorrendo desde o sul do México até a Argentina. No Brasil, esta
espécie ocorre em vários estados, apresentando relevada importância na pesca do norte e
amplo crescimento na aquicultura, notavelmente na região sul e sudeste. Apesar de sua
importância comercial, pouco se tem pesquisado sobre ações de parasitos nesta espécie que
podem ter como consequência danos à saúde dos peixes e de seus consumidores. Diante disto,
o presente estudo investigou a fauna microparasitária em 132 exemplares de Rhamdia quelen
coletados nos rios Paracauari (Salvaterra) e Arari (Cachoeira do Arari) entre julho de 2014 à
novembro de 2017. Foram observadas infecções por Myxobolus na camada muscular entre a
serosa e a mucosa do intestino, entre as camadas epi e hipo axial da musculatura, e pelo
mixosporídio Henneguya no rim. Para identificação destes microparasitos foram utilizadas
técnicas de microscopia de luz e histologia. As espécies foram identificadas por análises de
biologia molecular utilizando a subunidade ribossomal 18S rDNA, construção de banco de
dados utilizando sequências de outras espécies de mixosporídios disponíveis no GenBank,
alinhamento das sequências, construção de árvores filogenéticas por inferência bayesiana para
determinação do posicionamento filogenético dos taxons, além de serem realizadas
comparações morfológicas com outras espécies de mixosporídios. Diante dos resultados
obtidos constatou-se existência de três espécies novas, duas do gênero Myxobolus: Myxobolus
marajoensis infectando o trato intestinal e Myxobolus arariensis na musculatura, e uma
espécie do gênero Henneguya no rim, denominada Henneguya quelen.
Palavras-chave: microparasito, Henneguya, Myxobolus, 18S rDNA, jundiá.

10
ABSTRACT
Rhamdia quelen (Quoy & Gaimard, 1824) is a freshwater catfish widely distributed in the
Central and South America, occurring since Southern Mexico to Argentine. In Brazil, this
species occurs in several states, showing significant importance for both, fisheries and
aquaculture, mainly in the southern and southeastern regions. Despites of the commercial
importance, few investigations have been developed on the microparasitic infections on this
species in which, may have as consequence damages in the fish health as well as in their
consumers. This fact, suggest that further studies must be accomplished in order to obtain
further knowledge on the parasite fauna on this species. In this study, the microparasitic fauna
were investigated for 132 Rhamdia quelen collected in the Paracauari (Salvaterra) and Arari
(Cachoeira do Arari) rivers between July 2014 to November 2017. Infections by Myxobolus
were observed in the muscular layer between the intestinal serose and mucous layers, between
the epi and hipoaxial layers and also for the microparasite Henneguya disposed in the kidney.
For the identification of this parasites were used light microscopy and histological sections.
The species were also identificated by DNA extraction analysis, polymerase chain reaction
(PCR) and sequencing utilizing a subunit (18rDNA), construction of the database design
using sequences of other Myxosporidia species available in the GenBank, alignment of
sequences, phylogenetic tree constructions by Bayesian inference for the determination of the
taxon phylogenetic positioning, besides being performed morphological comparisons with
other myxosporid species. Based on these data, three new species were found, two belong to
the Myxobolus: Myxobolus marajoensis infecting the intestinal tract and M. arariensis the
musculature, and a new species of Henneguya in the kidney, denominated Henneguya quelen.
Key words: microparasite, Henneguya, Myxobolus, 18S rDNA, silver catfish.

11
1. INTRODUÇÃO GERAL
O Brasil apresenta uma grande diversidade de peixes de água doce, superior a 4.000
espécies. Somente a bacia amazônica abriga a maior e mais diversa ictiofauna do mundo,
tendo sido registrada mais de 1500 espécies (Goulding, 1980, 1989; Lowe-McConnell, 1999;
Reis et al., 2003). Cada espécie está susceptível à enfermidades causadas por inúmeros tipos
de parasitos, o que os tornam sujeitos à ações de doenças. No entanto, menos de 25% das
espécies de peixes foram objetos de investigações em relação à fauna parasitária, o que leva a
concluir que a biodiversidade desta fauna está longe de ser conhecida (Pavanelli et al., 2013).
Dentre os microparasitos em organismos aquáticos, destacam-se as espécies dos filos:
Cnidaria, Microspora e Apicomplexa, as quais pertencem aos reinos Animalia (sub-reino
Metazoa), Fungi e Protista, respectivamente. Estes contribuem com grande parte dos
microrganismos causadores de doenças em peixes de água doce e marinha, além de
microcrustáceos, nematódeos, entre outros (Lom & Diková, 1992; 2006; Matos et al., 2004;
2006; Eiras & Adriano, 2013).
O Subfilo Myxozoa abriga em torno de 2.400 espécies. São considerados importantes
microparasitos de peixes marinhos e de água doce, que, por muito tempo foram conhecidos
como protistas, atualmente classificados como cnidários (Fiala et al., 2015).
Os indivíduos afetados pelos parasitos podem ter sérias influências sobre os impactos
que envolvem interações ecológicas, mas também na saúde humana, pois várias destas
espécies são utilizadas como alimento. Muitas dessas espécies de peixes são reconhecidos
pela importância econômica, dentre estas podemos citar como exemplo o jandiá, Rhamdia
quelen (Quoy & Gaimard, 1824), que tem se destacado em duas áreas comerciais: a pesca
extrativa e na aquicultura. Obteve uma produção nacional próxima a 2.000 toneladas de
produto pescado em 2010 (MPA, 2011) e, apresentado um consolidado pacote tecnológico no
setor aquícola da região sul, obteve produção próxima de 2.600 toneladas em 2000, o que
corresponde a 1,4% do total produzido no Brasil (Baldisserotto & Radünz Neto, 2004;
Boscardim Borghetti et al., 2003; Piaia & Baldisseroto, 2000).
Contrariamente ao crescimento comercial dessa espécie, principalmente na
aquicultura, investigações voltadas às ações da fauna microparasitária, que pode ocasionar
infecções tanto para o indivíduo capturado no ambiente, como naqueles cultivados.
Resultando no comprometimento da sanidade dos peixes e também dos seus consumidores.

12
Sugerindo fortemente que estudos sobre microparasitismo sejam realizados. Neste sentido, o
presente estudo tem como objetivo investigar a fauna microparasitária de R. quelen, da região
Amazônica, utilizando a biologia molecular, comparações morfológicas e histológicas.
1.1. HOSPEDEIRO
A espécie R. quelen é um siluriforme que apresenta crescimento rápido nos meses
quentes e mais lento durante os meses mais frios do ano. É euritérmico e estenoalino, bem
adaptado a região amazônica. No Brasil é conhecimento por vários nomes comuns: jandiá,
jundiá, jundiá-tinga, jandiátinga, mandi e sapipoca; na Argentina, como bagre, bagre-negro,
bagre-sapo e bagre-sul-americana (Gomes et al., 2000; Baldisserotto & Radünz Neto, 2004).
Este bagre tem distribuição neotropical ocorrendo desde o sudeste do México, Bolivia,
Guianas, Venezuela, Peru, até o centro da Argentina. No Brasil se distribui em toda sua
extensão, na Amazonia, Rio de Janeiro, Sao Paulo, Rio Grande do Sul, e bacias do rio Sao
Francisco, Jequitinhonha, Mucuri, Paraiba e do Prata (Fowler, 1951; Silvergrip, 1996) (Fig.
1). Nestes locais há um complexo de espécies: 1. As que ocorrem somente na região norte -
Rhamdia foina, R. itacaiunas, R. laukidi, R. muelleri, R. poeyi; 2. Apenas na bacia do rio
Jequitinhonha, sudeste do Brasil – R. jequitinhonha; e 3. Aqueles que ocorrem em todo o
território brasileiro - R. quelen, com suas sinonímias R. branneri, R. voulezi, R. sapo, R.
hilarii e R. sebae (Baldisserotto & Randüz Neto, 2004).
O jandiá pode alcançar em média 3 kg e até 50 cm de comprimento, sua preferência é
por ambientes de águas paradas, com fundo arenoso e lamoso, próximo as margens e
vegetação, onde se abriga contra predadores, além de ter hábito noturno e viver em lagos e
poços fundos dos rios (Guedes, 1980).
A coloração do corpo varia de marrom-avermelhado claro a cinza ardósia
(acinzentado). A parte inferior da cabeça apresenta pigmentação variável. Possui como
característica crescimento alométrico negativo dos barbilhões, e esta relação é provavelmente
aumentada, devido à grande possibilidade de dano destas estruturas em exemplares grandes
(Silvergrip, 1996) (Fig. 2).

13
Figura 1- Mapa de distribuição geográfica de Rhamdia quelen.
Figura 2 - Exemplar de Rhamdia quelen confinado em laboratório.
Fonte: Arquivo LPCA, 2015.

14
No ambiente natural, a alimentação varia de acordo com a época do ano sendo
constituído, principalmente de crustáceos, molusco, restos de vegetais, e em cativeiro aceita
rações elaboradas (Baldisserotto & Randüz Neto, 2004).
Em relação à sua taxonomia, esta tem sido considerada controversa (Baldisserotto &
Randüz Neto, 2004) devido aos inúmeros caracteres morfológicos similares desta espécie e
outros do mesmo gênero, refletindo assim na dificuldade de uma correta classificação
(Scaranto, 2017). Uma ampla revisão taxonômica desta espécie, baseado em caracteres da
morfologia interna foi realizada por Silvergrip (1996) resultando na seguinte classificação:
CLASSE Actinopterygii
SUBCLASSE Neopterygii
INFRACLASSE Teleostei
SUPERORDEM Characiphysae
ORDEM Siluriforme
FAMÍLIA Pimelodidae
GÊNERO Rhamdia
ESPÉCIE Rhamdia quelen (Quoy & Gaimard, 1824)

15
1 IMPORTÂNCIA DO ESTUDO DE ICTIOPARASITAS
Estudos sobre microparasitos em peixes são importantes, pois podem elucidar aspectos
relacionados às patologias ocorrentes em seus hospedeiros, pelos impactos ecológicos e
econômicos causados por espécies invasoras e seus parasitas, e ainda, como bioindicadores na
qualidade da água (Lafferty, 1997).
As suas interações e ciclos de vida servem de base para uma série de investigações
sobre as relações entre espécies de parasitos e seus hospedeiros em áreas ainda não
investigadas. Neste sentido, pode-se considerar que a microparasitologia nos diferentes
grupos de animais tem constituído uma vertente biológica de grande interesse nas várias áreas
geográficas. Segundo Kubitza & Kubtiza (2004), vários são os mecanismos de transmissão de
parasitos entre os peixes, já que o ambiente aquático, por ser de certo modo bastante
homogêneo, facilita a propagação e distribuição desses organismos.
A parasitologia busca a compreensão das relações entre o parasito e seu hospedeiro,
assim como, o ambiente. Para a identificação de um parasita em seu hospedeiro é necessário
inicialmente realizar um procedimento de necropsia do peixe seguido de, observação em lupa
para detecção dos cistos e, por microscopia, no qual o cisto é detalhadamente observado entre
lâmina e lamínula. Esta sequência pode ser considerada o primeiro passo para entender e
estabelecer diagnósticos de doenças. Sua aplicação tem fundamental importância na
manutenção da sanidade de organismos tanto nos ambientes naturais, como naqueles
utilizados em cultivo (Schaperclaus, 1991; Pavanelli et al., 2013). Estudos de microparasitos
observados em microscopia de luz (ML) são observados nos trabalhos de Matos et al. 2004;
Abdel-Ghaffar et al., 2008; microscopia eletrônica de transmissão (MET) Rocha et al., 2011;
Casal et al., 2010, 2012; biologia molecular (BM) Carriero et al., 2013; Azevedo et al. 2014,
2016; Rocha et al. 2014b; Casal et al. 2017; Matos et al., 2018.

16
1.3 FILO CNIDARIA (Hatschek, 1888)
Os cnidários, também chamados de celenterados, são invertebrados típicos de ambientes
marinhos e de água doce e reconhecidos por sua importância ecológica e econômica global.
Estão entre os seres mais antigos do Reino Animal, tendo como representantes as meduzas
(águas vivas, anêmonas, corais e hidras) (Fiala et al., 2015).
Sabe-se atualmente que, na antiguidade, um clado de cnidários se divergiu dos demais
para se tornar endoparasitas, que hoje compõem o Myxozoa. Portanto, os myxozoários agora
merecem amplo reconhecimento como cnidário exibindo ampla riqueza de espécies, e que
convergiu pelas suas estratégias de microparasitas (protistas parasitas) para a exploração do
hospedeiro, em muitos casos, considerados altamentes problemáticos por causar parasitismo
em conhecidas espécies de peixes (Okamura et al., 2015).
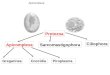
Em relação à sua taxonomia Fiala et al. (2015) classificou este filo como demonstrado
a seguir na Figura 3:
Figura 3 – Classificação do Filo Cnidaria (Subfilo Myxozoa).
Fonte: Fiala et al., 2015 (adaptado)
Os membros pertencentes a este grupo são endoparasitos, os quais podem ser
histozóicos (com formação dentro das células e tecidos), celozóicos (dentro das cavidades do
corpo ou de órgãos do corpo como na bexiga urinária e na vesícula biliar) ou ainda dentro de

17
vasos sanguíneos (El-Matbouli et al., 1992; Casal et al., 2009; Eiras & Adriano, 2013). Diante
desta grande variedade como agentes parasitários e patogênicos na ictiofauna, destaca-se
aquelas espécies de Myxozoa encontrados parasitando peixes de valor comercial (Matos et
al., 2004; Casal et al., 2009) e que podem causar mixosporidiose, doença que atua em
diversos tecidos e órgãos, como gônadas, rins, fígado, brânquias, intestino, pele e outros
(Azevedo et al., 2009; Casal, 2009).
1.3.1 Subfilo Myxozoa Grassé, 1970
Myxozoa é uma assembléia de organismos microscópicos, oligocelulares,
endoparasitas de vertebrados e invertebrados aquáticos. Os myxozoários foram
freqüentemente comparados aos cnidários, devido sua afinidade mútua (Stolc, 1899). Por este
motivo, o Myxozoa foi por muito tempo classificado no taxon protista, mas que atualmente é
classificado como cnidário (Fiala et al., 2015).
A origem metazoária do Myxozoa, bem como a sua relação próxima com o
polipoidozoário, Polypodium hydriforme (Ussov, 1885), descrito como parasita dos ovos
de esturjão e de peixes afins, constituindo um dos poucos cnidários que vive no interior
das células de outros animais. Esta semelhança corroborou com a colocação de Myxozoa
dentro do filo Cnidaria.
As análises filogenômicas reforçaram esta afirmação em agrupar o Myxozoa dentro do
subfilo Medusozoa, um grupo composto por pólipos, hydras e anêmonas (Jimenez-Guri et al.,
2007; Nesnidal et al., 2013; Feng et al., 2014). A sistemática dos Myxozoa passou então por
alterações tendo ainda influência de duas importantes descobertas: 1. O ciclo de vida é
dixeno, ou seja possui dois hospedeiros, um intermediário vertebrado e outro definitivo
invertebrado; 2. O fato de se reconhecer que o parasita de Briozoários, Buddenbrokia
plumatellae era um Myxozoa, o que criou a nova classe (Malacosporea).
O subfilo Myxozoa alberga 64 gêneros, 17 famílias e aproximadamente 2.400 espécies
descritas, representando cerca de 18% da diversidade das espécies cnidárias conhecidas (Lom
& Dyková, 2006; Matos et al., 2004; Zhang, 2011; Eiras & Adriano, 2013) (Fig. 4).
Disseminados nos mais diversos meios aquáticos de água doce e salgada, a maioria destes
parasitos são considerados patogênicos, atuando em diversos tecidos e órgãos, como fígado,
rim, brânquias, gônadas, intestino, pele e outros.

18
Figura 4 - Gráfico demonstrando as proporções (e os números) de espécies descritas no
Myxozoa e outros clados nos cnidários. Cubozoa foi agrupado com Scyphozoa devido ao
baixo número de espécies (42) neste grupo. Dados para o número de espécies descritas de
Zhang (2011). Fonte: Zhang, (2011).
Os myxozoas parasitam peixes na forma de esporos, podendo, no entanto, apresentar
diferentes formas, com um ou mais esporoplasmas amebóide, com tamanhos variáveis e
várias cápsulas polares (geralmente duas), variando de uma a quinze (Fig. 5). Essas estruturas
são semelhantes aos nematocistos dos Cnidaria contendo no interior um filamento geralmente
enrolado em espiral, sendo o número de voltas importante para a caracterização da espécie. A
parede do esporo é constituída por duas ou mais valvas, unidas por linhas de sutura. O
esporoplasma, é formado por uma célula binucleada ou por duas células uninucleadas, tem a
função de infectar o hospedeiro (Pavanelli et al., 2013).

19
Figura 5 – Representantes de mixosporídios apresentando diversidade mofológica e no
número de cápsulas polares em microscopia de luz. a) Henneguya nagelii (Azevedo et al.
2013) b) Myxobolus arariensis; c) (P) Plasmódio dispórico de Ellipsomyxa gobioides
(Azevedo et al. 2013) d) Ceratomyxa anko (Freeman; Yokoyama & Ogawae, 2008) e) Kudoa
inornata (Dykovà et al. 2009).
A maioria destes microparasitos apresenta especificidade parasitária, no entanto, uma
espécie de peixe pode albergar várias espécies de mixosporídios. Desde o final do século
XIX, especialistas estão pesquisando o ciclo evolutivo dos mixosporídios e as doenças que
eles provocam (Békési et al., 2002).
O ciclo de vida dos mixosporídios alterna entre hospedeiros vertebrados (peixes -
intermediário) e invertebrados (anelídeos - definitivo) (Fig. 6). Primeiramente ocorre à
extrusão do filamento polar ancorando o mixosporo no epitélio intestinal do anelídeo, em
seguida ocorre a abertura das valvas do esporo, passando por gametogonia, esporogonia do
estágio actinosporo até sua forma madura que é liberada na água. O actinosporo entra em
contato com a superfície da pele ou brânquias do peixe, libera o filamento polar, facilitando a
invasão do esporoplasma no novo hospedeiro, multiplica-se, ocorrendo esporogonia e
esporogonia dos mixósporos, que serão liberados no ambiente novamente e ingeridos pelos
anelídeos, recomeçando o ciclo (Yokoyama et al., 2012).

20
Figura 6 - Desenho esquemático do ciclo de vida de um mixosporídio: a) hospedeiro
definitivo; b) actinosporo; c) hospedeiro intermediário; d) mixosporo. Fonte: Adaptado de
Chang et al. (2015).
Na embriogênese dos mixosporídios, a esporogênese começa com o envolvimento de
uma célula esporogônica, desenvolvida e protegida por um fagócito, chamado de pericito.
Dentro deste, a célula esporogênica divide-se, sucessivamente, por meio de nucleocinese e
citocineses, originando células uninucleares, exceto uma célula, na qual ocorre apenas uma
nucleocinese, originando-se, assim, uma célula binucleada, a célula esporoplasmática. Dessa
divisão, nascem dois grupos de cinco células. Cada grupo delas se distingue por duas células
valvogênicas, diferenciam-se e englobando as outras três, formando-se, assim, um esporo.
Das três células que permanecem no interior, duas destas são células capsulogênicas, que no
citoplasma se diferenciam as cápsulas polares, possuindo cada uma delas no seu interior um
filamento polar enrolado em hélice. A terceira célula é referida como esporoplasmática
binucleada que se caracteriza por conter inúmeras vesículas eletrondensas designadas de

21
esporoplasmosomas, bem como partículas de glicogênio. As cápsulas são estruturas
específicas dos esporos dos mixosporídios (Matos et al., 2001) (Fig. 7).
As cápsulas polares resultam das células capsulogênicas que se diferenciam durante o
processo de maturação. No interior de cada uma das cápsulas localiza-se um tubo polar com
filamento enrolado em espiral. O filamento polar é uma estrutura responsável pela propagação
da espécie, no período pós-maturação. Durante o processo de formação do esporo foi
observado que cada uma das valvas desenvolve um complexo sistema microtubular que se
diferencia, em algumas espécies, nas caudas ou prolongamentos (Matos et al., 2004) (Fig. 7).
complexo sistema microtubular que se diferencia, em algumas espécies, nas caudas ou
prolongamentos (Matos et al ., 2004a).
A presença de esporos de mixosporídios (Phylum Myxozoa) são facilmente
observados em Microscopia de Contraste Interferencial (Nomarski - DIC) e
identificados pela sua morfologia (Lom & Dyková, 1992). Os esporos, apesar de se
Figura 4: Esquema do ciclo de vida dos mixosporidios (gênero Henneguya). Mostrando
a fase em que a célula pericítica (pericito - a) começa a englobar a célula germinativa
(b) (1). A célula germinativa fica completamente envolvida pelo pericito (2). A célula
germinativa dividi-se sucessivamente em duas células (3) e em 4 células (4). No final
da divisão, o esporo (monoespórico - 5) é constituído por 5 células diferenciadas em 2
valvogênicas (d), 2 células capsulogênicas (e) e uma célula binucleada – o
esporoplasma (c). Esporo (6).
Fonte: Matos et al . (2001).
Figura 7 - Esquema da embriogênese dos mixosporídios. Mostrando a fase em que a célula
pericítica (pericito – a) começa a englobar a célula germinativa (b) (1). A célula germinativa
fica completamente envolvida pelo pericito (2). A célula germinativa divide-se
sucessivamente em duas células (3) e em 4 células (4). No final da divisão, o esporo
(monoespórico – 5) é constituído por 5 células diferenciadas em 2 valvogênicas (d), 2 células
capsulogênicas (e) e uma célula binucleada – o esporoplasma (c). Esporos (6). Fonte: Matos
et al. (2001).

22
1.3.2 Filogenia do Myxozoa
De acordo com Fiala et al. (2015) a morfologia dos esporos era o principal critério
para a classificação e caracterização dos microparasitos. Neste contexto, o subfilo Myxozoa
era agrupado como Protista, tendo como base de classificação, em relação ao plano sutural
para caracterizar ordem e subordem, o numero e a forma das válvulas, assim como, o número
e a disposição das cápsulas polares (Lom & Dyková, 1992). A posição dos myxozoários como
protista foi aceita por muito tempo. No entanto, a classificação de qualquer grupo de
organismos não deveria ser baseada apenas com as características morfológicas, mas numa
combinação de vários fatores, como: habitat, especificidade do hospedeiro, local de infecção,
interação com as células hospedeiras e as características do ciclo de vida do parasito (Casal,
2009).
Com o avanço das técnicas de microscopia eletrônica de transmissão as
investigações ultraestruturais permitiram a caracterização da estrutura de esporos, de suas
cápsulas e filamentos polares. Este fato, possibilitou novos conhecimentos sobre biologia
funcional e características citológicas deste grupo (Lom & Puytorac, 1965; Lom, 1969) o que
levantou dúvidas sobre a antiga classificação.
Posteriormente, Siddall et al., 1995, apresentou o primeiro trabalho a combinar
dados ultraestruturais de microscopia eletrônica de transmissão com dados de sequência de
DNA. Este estudo revelou que os myxozoas não se incluíram no grupo protista como eram
aceito, mas se revelaram com características de origem metazoária, com estreita afinidade
com o polypoidozoário Polypodium hydriforme Ussov, 1885 inserindo então os Myxozoa
dentro do filo Cnidaria atualmente aceito. Siddal et al (1995) redesenhou a árvore filogenética
colocando Myxidium, Henneguya e Myxobolus em mesmo subclado com o Polypodium (Fig.
8)

23
Figura 8. Árvore filogenética redesenhada de Siddall et al. (1995) mostrando 3 espécies irmãs
de myxozoários próximo ao parasita narcomedusae Polypodium hydriforme e entre outros do
filo Cnidaria (ramos pretos) em oposição ao agrupamento com bilaterais (ramos cinzas
interrompidos). A árvore resultou da análise de uma combinação de caracteres morfológicos e
sequências do gene de rRNA 18S. Uma análise adicional com a dinâmica método de
homologia que considera os eventos de exclusão de inserção corroborando com a inclusão de
Myxozoa dentro do grupo monofilético Cnidaria (Siddall et al., 1995).
Através do aumento da amostragem de táxons para dados moleculares em
combinação com dados morfológicos e ontogenéticos, demonstraram que o grupo Myxozoa
com narcomedusans como esperado dentro da Cnidaria (Fig. 8). Além do resultado
filogenético, Siddall et al. (1995) deixou claro ultraestrutural evidência de características
metazoárias conhecidas, tais como células terminais de diferenciação, septado intercelular e
junções desmossômicas, comunicação citoplasmática intercelular e colágeno reticulado.

24
1.3.3 Myxobolus e Henneguya
Os parasitos destes gêneros podem gerar danos significativos, por podem levar seus
hospedeiros a morte tanto em ambiente natura e em sistemas de cultivo, isto por que há uma
grande diversidade encontrada e ao potencial patogênico de algumas espécies destes gêneros
(Woo, 2006; Ferguson et al 2008; Eiras et al 2009).
1.3.3.1 Gênero Myxobolus Bütschli, 1882
O gênero Myxobolus é o maior e mais expressivo dentro da classe Myxosporea, atingiu
quase 800 descrições de espécies (Mólnar, 2014). No Brasil, já foram descritas
aproximadamente 60 espécies de Myxobolus infectando peixes da nossa fauna (Azevedo et al.
2012; Eiras et al. 2014). Este mixosporídio pode causar a morte dos hospedeiros em ambiente
natural e de cultivo (Eiras et al. 2009).
A espécie conhecida como representante do gênero, é o Myxobolus cerebralis Hofer,
1903, tornou-se a espécie mais conhecida dos mixosporídios e considerada a espécie símbolo
do gênero por ser um agente causador da doença do rodopio ou doença da cauda que causa
alta mortalidade em trutas em ambientes naturais nos Estados Unidos (Rose et al. 2000) (Fig.
9). Outra espécie consideradas altamente patogênicas para seus hospedeiros, é o Myxobolus
buckei que parasita a coluna vertebral de ciprinídeos (Longshaw et al., 2003).
Figura 9 – Desenho esquemático de Myxobolus cerebralis a partir de péletes digeridos com
pepsina-tripsina da truta arco-íris Oncorhynchus mykiss e truta marrom Salmo trutta coletados
na Bacia do Rio Watauga, Carolina do Norte (EUA). Fonte: Ruiz et al., (2017).

25
1.3.3.2 Gênero Henneguya Thélohan, 1892
Entre os microparasitos encontrados em peixes da Amazônia destaca-se o gênero
Henneguya (Fig. 10), com características importantes tais como, valvas que são estruturas
comuns a todas as espécies, a existência de cauda e aspectos morfológicos, como tamanho e
forma do corpo do esporo. A ação destes parasitos frequentemente encontrados nas brânquias,
no fígado, na vesícula e nos músculos, já foram relatadas por Matos et al. (1999).
O gênero Henneguya além de ser o segundo mais descrito da família, possui
importância patogênica no ambiente natural e em sistemas de cultivo (Naldoni, 2009). Pagrus
major é um exemplo de hospedeiro de ambiente natural que foi infectado por Henneguya
pagri no coração causando cardiomiopatias degenerativas levando a mortalidade (Yokoyama,
2005). O exemplo de hospedeiro na piscicultura é o Pseudoplatystoma corruscans infectado
por Henneguya ictaluri que causa redução do epitélio respiratório e o Ictalurus punctatus
infectado por H. ictaluri ocasiona a doença proliferativa da brânquia” (PGD) (Fig. 10) (Pote
et al 2000; Naldoni et al 2009).
Figura 10 – Fotomicrografia de Henneguya ictaluri na forma Actinosporo do mixosporídio.
Bar = 50 μm. Fonte: Pote et al., (2012).

26
1.3.4 JUSTIFICATIVA
Sobre a espécie em estudo, pode-se considerar que R. quelen apesar de ser considerada
uma espécie de valor econômico e, como tem sido alvo de criação em cativeiro nos estados do
Rio Grande do Sul e Paraná (Gomes et al., 2000) e importante recurso na pesca extrativista
para região norte, possua uma fauna microparasitária pouco explorada.
Assim, verificamos ser de grande relevância o presente estudo que servirá de base para
o conhecimento sobre a fauna microparasitária de R. quelen, tanto de ambiente natural como
de cultivo, descrevendo os aspectos morfológicos e moleculares dos microparasitas que foram
identificados, contribuindo para o conhecimento do estado sanitário desta espécie.
2. OBJETIVOS
2.1 OBJETIVO GERAL
Este trabalho teve como objetivo investigar a ocorrência de mixosporídios, em jandiá,
Rhamdia quelen de ambiente natural, dos municípios de Salvaterra e Cachoeira do Arari da
Ilha de Marajó, Pará.
2.2 OBJETIVOS ESPECÍFICOS:
Realizar o estudo taxonômico através da filogenia clássica e molecular.
Identificar e classificar as espécies de microparasitos de R. quelen capturados na
Ilha de Marajó (Salvaterra e Cachoeira do Arari);
Descrever os aspectos morfológicos dos microparasitos;
Verificar a prevalência de cada grupo de microparasitos nos espécimes analisados;
Analisar a preferência parasitária em relação aos vários órgãos do hospedeiro;
Identificar e classificar as espécies de microparasitos de R. quelen capturados na
Ilha de Marajó (Salvaterra e Cachoeira do Arari);
Descrever os aspectos morfológicos dos microparasitos;
Verificar a prevalência de cada grupo de microparasitos nos espécimes analisados;
Analisar a preferência parasitária em relação aos vários órgãos do hospedeiro;
Avaliar as relações filogenéticas dos táxons em estudo.
Identificar a ocorrência de multiparasitismo.
Descrever a presença de lesões causadas por microparasitos nos tecidos e órgãos.

27
3. APRESENTAÇÃO DOS ARTIGOS
Os resultados e discussão desta Tese estão apresentados em quatro artigos científicos,
sendo 2 publicados e 2 submetidos à publicação.
Artigo I apresenta um estudo morfológico e histopatológico de um mixosporídio,
Myxobolus sp. infectando o intestino de seu hospedeiro, o siluriforme R. quelen (Quoy &
Gaimard, 1824) capturado na Ilha do Marajó, Pará.
Artigo II o mixosporídio Myxobolus sp. apresentado no trabalho anterior, foi
investigado neste estudo sob aspectos moleculares, utilizando gene nuclear (rDNA 18S) e
realizada análise filogenética cujo posicionamento do parasito em relação a outras espécies
mixosporídios previamente descritos infectando peixes das ordens: siluriformes,
characiformes, mugiliformes e cypriniformes obtidas do GenBank. Os resultados apontaram
uma nova espécie, Myxobolus marajoensis em R. quelen.
Artigo III apresenta uma nova espécie de Myxobolus, denominada, M. arariensis,
encontrada no músculo esquelético do siluriforme R. quelen, capturado no rio Arari, Ilha do
Marajó. Este trabalho usa como ferramentas técnicas histológicas e de biologia molecular.
Determina o posicionamento filogenético deste taxon, comparado com outras espécies de
mixosporídios parasitas de peixes de água doce e marinha.
Artigo IV descreve uma nova espécie de Henneguya, H. quelen nos rins de Rhamdia
quelen capturado no rio Paracauari analisado através das comparações morfológicas,
histológicas, moleculares e filogenéticos.

28
ARTIGO I
Título: Myxozoan infection in the muscle layer of the intestine of Rhamdia quelen from the
Amazon River Basin, Brazil.
Autores: Jacqueline Pompeu Abrunhosa, Michele Velasco Oliveira da Silva, José Ledamir
Sindeaux Neto, Patrícia de Fátima Sacco dos Santos, Patrícia Santos Matos, Osimar de
Carvalho Sanches, Edilson Rodrigues Matos.
Status: Publicado
Revista: Ciência Rural, Santa Maria, v.46, n.11, p.2024-2028, nov, 2016
ISSN 1678-4596
Fator de impacto: 0,26 (Qualis B5)
Doi: http:// dx.doi.org/10.1590/0103-8478cr20151652

29
Myxozoan infection in the muscle layer of the intestine of Rhamdia quelen from the Amazon River Basin, Brazil. 2024
Ciência Rural, v.46, n.11, nov, 2016.
ISSN 1678-4596 PARASITOLOGY
Myxozoan infection in the muscle layer of the intestine of Rhamdia quelen from the Amazon River Basin, Brazil
Infecção por Myxozoa na camada muscular intestinal do Rhamdia quelen
da Bacia do Rio Amazonas, Brasil
Jacqueline Pompeu AbrunhosaI Michele Velasco Oliveira da SilvaI José Ledamir Sindeaux NetoI
Patrícia de Fátima Sacco dos SantosI Patrícia Santos MatosI I Osimar de Carvalho SanchesI I I
Edilson Rodrigues MatosI *
ABSTRACT
The present study investigated the morphology and
pathology associated with the occurrence of cysts caused by
Myxobolus sp. in the intestine of the silver catfish Rhamdia quelen. Comparisons with the other Myxobolus species that
infect the intestines of cyprinids and siluriforms revealed that the pyriform spores of Myxobolus sp. are similar in shape to those
described previously in M. miyairii, M. duodenalis and M.
cunhai, but different in size. Morphometric analyses revealed that mature spores of Myxobolus sp. (10.9 x 5.1µm) are smaller than
those of most species except M. cunhai (10.0 x 5.0µm), which has been described infecting the Brazilian catfish Pimelodus clarias,
although the spores differ morphologically in relation to the shape of the polar capsules. Further research is necessary in order to
clarify the taxonomic and phylogenetic relationships among these congeneric species.
Key words: Myxobolus, histopathology, fish parasites, catfish,
neotropical.
RESUMO
O presente estudo investigou a morfologia
e patologia associada com a ocorrência de cistos de Myxobolus sp. no intestino do peixe siluriforme Rhamdia
quelen. Comparações com outras espécies de Myxobolus que têm infectado intestinos de carpas e bagres de água doce
demonstraram que os esporos piriformes de Myxobolus sp. são similares no formato dos anteriormente descritos para M.
miyairii, M. duodenalis e M. cunhai, diferindo, no entanto, quanto ao tamanho. As análises morfométricas revelaram
que os esporos maduros de Myxobolus sp. (10,9 x 5,1µm) são menores do que os da maioria das espécies, exceto para M.
cunhai (10,0 x 5,0µm), que foi descrito infectando o bagre
brasileiro Clarias pimelodus, embora os esporos sejam
diferentes morfologicamente em relação à forma das cápsulas
polares. Futuras pesquisas são necessárias para esclarecer as relações taxonômicas e filogenéticas entre espécies congêneres.
Palavras-chave: Myxobolus sp., histopatologia, parasitas de
peixe, bagres, neotropical.
INTRODUCTION
The Myxozoa Grassé, 1970 are a
heterogeneous group with approximately 60 genera
(KENT et al., 2001; LOM & DYKOVÁ, 2006),
which are widespread in aquatic environments,
both freshwater and marine. Most of these parasites
are considered pathogenic, infecting a variety of
tissue in different types of organ, such as liver,
kidneys, gills, gonads, intestines, and skin (LOM &
DYKOVÁ, 2006).
The known species of Myxobolus
BÜTSCHLI, 1882 (Myxobolidae) are important
pathogens in freshwater and marine fish (LOM &
DYKOVÁ, 1992; EIRAS et al., 2005). There are few
reports of Myxobolus cysts in the intestines of fish;
although, a number of histopathological descriptions
are available. These include MOLNÁR’s (2002)
description of M. cyprinicola parasitizing Cyprinus
carpio LINNAEUS, 1758 from Lake Balaton in
Hungary, M. nodulointestinalis in Barbus spp. from
ILaboratório de Pesquisa Carlos Azevedo, Universidade Federal Rural da Amazônia (UFRA), 66077-901, Belém, PA, Brasil. E-mail: [email protected]. *Corresponding author.
IIEdilson Matos Research Laboratory, Universidade Federal do Pará (UFPA), Belém, PA, Brasil. IIICentro de Diagnóstico Veterinário (CDAPVET), Presidente Prudente, SP, Brasil.
Received 12.23.15 Approved 06.23.16 Returned by the author 09.14.16 CR-
2015-1652.R2
Ciência Rural, Santa Maria, v.46, n.11, p.2024-2028, nov, 2016 http://dx.doi.org/10.1590/0103-8478cr20151652

30

31

32

33

34
ARTIGO II
Título: Myxobolus marajoensis n. sp (Myxosporea: Myxobolidae), parasite of the freshwater
catfish Rhamdia quelen (Siluriformes: Pimelodiade) from the Brazilian Amazon Region
Autores: Jacqueline Abrunhosa, José Ledamir Sindeaux-Neto, Ândrea Kely Santos, Igor
Hamoy, Edilson Matos
Status: Publicado
Revista: Revista Brasileira de Parasitologia Veterinária
ISSN: ISSN 1984-2961 (Electronic)
Fator de impacto: 1,139 (Qualis B4)
DOI: http://dx.doi.org/10.1590/S1984-29612017067

35

36

37

38

39

40

41

42
ARTIGO III
Título: A new species of myxozoa in the skeletal striated musculature of Rhamdia quelen
(Quoy & Gaimard) (Siluriforme: Pimelodidae) Amazonian fish, Marajó island, Brazil
Autores: Jacqueline Abrunhosa, José L. Sindeaux-Neto, Sidney dos Santos, Igor Hamoy,
Edilson Matos
Revista: Zootaxa
Status: Submetido
ISSN: 1175-5334
Fator de impacto: 0,972 (Qualis B4)

43
A new species of myxozoa in the skeletal striated musculature of Rhamdia quelen (Quoy
& Gaimard) (Siluriforme: Pimelodidae) Amazonian fish, Marajó island, Brazil
JACQUELINE ABRUNHOSA1,2,4
, JOSE L. SINDEAUX-NETO1,5
, SIDNEY DOS
SANTOS3, IGOR HAMOY
2 & EDILSON MATOS
1,*
¹Carlos Azevedo Research Laboratory, Institute of Animal Health and Production, Federal
Rural University of Amazonia, Avenida Presidente Tancredo Neves, 2501, Montese, 66.077-
901, Belém, PA, Brazil. E-mail: [email protected]; [email protected]
²Laboratory of Applied Genetics, Sociambiental Institute and Water Resources, Federal
Rural University of Amazonia, Avenida Presidente Tancredo Neves, 2501, Montese, 66.077-
901, Belém, PA, Brazil.
E-mail:[email protected]
³Laboratory of Human and Medical Genetics, Institute of Biological Scis, Federal University
of Pará, Rua Augusto Corrêa, 1 - Guamá, 66075-110, Belém, PA, Brazil. E-mail:
*Corresponding author. E-mail: [email protected]
Abstract
A new myxozoan was found parasitizing the freshwater catfish, Rhamdia quelen (Quoy &
Gaimard), in the Marajó Island, Amazon region, Brazil. The new species is described based
on the results of morphological and molecular analyses. The parasite is approximately 1.5 mm
in diameter and develops in the musculature of the host in the form of spherical, whitish cysts,
which are visible macroscopically between the epaxial and hypaxial layers. When ruptured,
these cysts produced ellipsoidal spores with a mean length of 11.4 μm (10.7-12.6) and width
of 7.2 μm (6.4-7.9). Anomalous spores with a caudal elongation, vesicles in the peripheral
portion of the spore and ornamentation of the valves were also observed. The results of the
phylogenetic analysis of sequences of the 18S rDNA gene using Bayesian Inference indicated
clear differences among the Myxobolus species that reinforced the taxonomic position of the
parasite, confirming its status as a new species, denominated Myxobolus arariensis n. sp.
Key words: parasite, freshwater fish, Myxobolidae, histology, SSU 18S rDNA gene
Running title: New parasite of an Amazonian fish …

44
Introduction
Catfish (Siluriformes) are widely consumed as food in Brazil, in particular in the northern
region, where commercial fisheries harvest approximately 60,000 tons each year (Brasil
2011). The most popular species include the mapará, Hypophthalmus marginatus
Valenciennes, 1840, the yellow mandi, Pimelodus maculatus, and the mandi, Pimelodus sp.,
the silver catfish (known locally as the jandiá), Rhamdia quelen (Quoy & Gaimard), and the
goliath catfishes, Brachyplatystoma filamentosum (Lichtenstein), Brachyplatystoma
rousseauxii (Castelnau), and Brachyplatystoma vaillantii (Valenciennes) (Barthem &
Goulding 2007, Brasil 2011).
The popularity of these fish species as sources of food reinforces the need for
parasitological studies, given that these organisms can cause a variety of diseases, alterations
in the behavior of the fish (Thompson et al. 2002), and impacts on the environment, in
addition to serious problems for human health (Boreham et al. 1998, Moncada et al. 2001,
Hessen & Zamzame 2004, Martinez de Velasco et al. 2008). The occurrence and seriousness
of the disease depends on a series of factors, including the characteristics of the environment,
the host, and the pathogen, and their interactions (Snieszko 1974, Hedrick 1998). In this
context, parasitological research in Brazil has focused primarily on a number of the catfish
species found in the Amazon region, including B. vaillantii (Silva et al. 2012), Pimelodus sp.
(Matos et al. 2014), and H. marginatus (Velasco et al. 2015a, 2015b, Rocha et al. 2015).
While many catfish species are important resources for extractive fisheries, R. quelen also
has considerable potential for fish farming, principally in southern Brazil, where the farming
of this species has been growing steadily, stimulated by the high levels of fry production
(Baldisseroto & Raduz Neto 2004, Brasil 2011).
However, few data are available on the parasite fauna of R. quelen. Matos et al. (2005)
described Henneguya rhamdia in gills of this catfish, while Abrunhosa et al. (2017) observed
infections by Myxobolus marojoensis in the intestine. The present study investigated the
presence of microparasites in R. quelen, which resulted in the description of a new species of
Myxobolus from the epaxial and hypaxial muscle layers. The morphological and phylogenetic
analyses (18 S rDNA sequences), confirmed the existence of a new species, which was
denominated Myxobolus arariensis n. sp.
Materials and Methods
Morphological analysis - Twenty five specimens of R. quelen (total length: 15–23 cm) were
captured from the Arari River in Cachoeira do Arari (01º00’S 48º57’W) on Marajó Island, in
the Brazilian state of Pará, June to November 2017.

45
The fish were transported live in aerated water, obtained from the natural habitat, to the
Carlos Azevedo Research Laboratory at the Federal Rural University of Amazonia (UFRA) in
Belém, Pará, Brazil. The specimens were maintained in aquaria until necropsy.
In the Laboratory, after being anesthetized with tricaine methanesulfonate (50 mg/L)
(MS222 SIGMA), the fish were euthanized and dissected in accordance with the procedures
approved by the UFRA ethics committee for animal experimentation (CEUA: 013/2014). The
collection and transportation of biological samples was authorized by the Chico Mendes
Institute for Biodiversity and Conservation (SISBIO/ICMBIO license 27119).
Necropsy was initiated with an incision in the abdominal cavity. The parasitized organs
were examined under a stereomicroscope and the observed cysts were analyzed by light
microscopy. The whitish cysts found in the epaxial and hypaxial layers of the musculature of
the host were removed and mounted on slides, covered with a drop of water and coverslip, for
observation under a light microscope (LM). This examination confirmed the presence of
spores with characteristics typical of the genus Myxobolus.
Fresh plasmodia with mature spores were examined morphologically and
morphometrically under a light microscope, following Kudo 1921. The spores were measured
(μm) and mounted on slides with a coverslip and photographed under a Zeiss Primo Star
optical microscope equipped with a Zeiss AxioCam ERc 5s photographic camera and the
AxioVision 5.1 software.
Histological examination - For histology, small fragments of the muscle tissue
containing cysts were removed and fixed in Davidson solution for 24 h, and then dehydrated
in an increasing series of ethyl alcohol concentrations, clarified in xylene, and then embedded
in paraffin. The tissue samples were sectioned in a microtome with a thickness of 5 µm and
then stained with Hematoxylin-Eosin (H-E), following the Gutierrez and Ziehl Neelsen
techniques (Luna 1968). The sections were observed and photographed under light
microscopy (Zeiss Primo Star).
Extraction of the DNA and sequencing - For the extraction of the DNA, the spores were
preserved in 80% ethanol as soon as they were retrieved, and then centrifuged and washed in
distilled water to remove the excess ethanol. The PureLink® Genomic DNA mini kit from
Invitrogen was then used, following the manufacturer’s protocol. The DNA concentration was
measured by a NanoDrop 1000 spectrophotometer (Thermo Scientific) at 260 nm, with the
samples diluted to 5ng/μL.
A partial sequence of the SSU rDNA gene was amplified using the primers MC5
(forward) 5’-CCTGAGAAACGGCTACCACATCCA-3’ and MC3 (reverse) 5’-
GATTAGCCTGACAGATCACTCCACGA-3 (Molnár 2002) in 25 μL of the Polymerase
Chain Reaction (PCR) mixture, containing 1μL of the DNA template, 2 mM MgCl2 of 4 mM
of dNTPs mix (Invitrogen), 5 pmol of each primer, and 1.2 units of Taq DNA polymerase
(Invitrogen®). The amplification profile comprised 35 cycles of 1 min at 95°C, 1 min at
66°C, and 2 min at 72°C, preceded by 5 min at 95°C for block temperature homogenization,
and followed by a final 30-min step at 72°C for the polymerization of any molecules that the
polymerase had dissociated before the end of the synthesis of the complete fragment.

46
The amplicons were electrophoresed in 1.0 % agarose gel purified using GFX PCR DNA
and a Gel Purification Kit (GE Healthcare), according to the manufacturer’s instructions. The
purified material was sequenced in an ABI 3130 automatic DNA analyzer (Applied
Biosystems TM) with BigDye® Terminator v3.1, following the manufacturer’s specifications.
Phylogenetic analysis - The nucleotide sequences obtained by the PCR were edited and
aligned using the BioEdit software (Hall 2007). A number of ambiguous bases were identified
using ABI chromatograms, and the consensus sequences were deposited in GenBank under
accession numbers MG572219, and verified as myxozoan by a BLAST search.
The phylogenetic relationships between the new species and related myxosporids were
based on the analysis of 41 sequences obtained from GenBank, which were aligned using the
Clustal X 1.8 program (Thompson et al. 1997). These species included Kudoa thyrsites
(AY941819) and K. alliaria (DQ182561) as the outgroup. The data used for the alignment
consisted of the best results obtained by the BLAST search, together with representatives of
neighboring clades, as determined in the previous analyses of Myxobolus (Ferguson et al.
2008, Liu et al. 2010, Molnár et al. 2006, Zhao et al. 2008). The alignment was corrected
manually using the BioEdit alignment editing software (Hall 2007).
The Genetic distances (p distance) were determinate using PAUP 4.0b10 (Swofford
2003), for the SSU rDNA gene sequences from the Myxobolus parasite of Fish.
The phylogenetic analysis ran by Bayesian Inference (BI) implemented in MrBayes 3.1.2
(Ronquist & Huelsenbeck 2003). The performance was accomplished using two parallel runs
of four synchronized Chain Monte Carlo (MCMC) searches of 5 million generations each,
and the results of the first 1,000 trees were discarded as “burn-in”. The remaining trees were
used by MrBayes to estimate the posterior probability of each node in our phylogenetic
reconstruction. Fig-Tree 1.3.1 software (Rambaut 2008) was used to create visuals of the
results of Bayesian analysis.
Results
Following dissection, cysts were observed in the epaxial and hypaxial regions of the
musculature of three of the 25 R. quelen specimens examined. The cysts were visible
macroscopically following dissection of the musculature, showed ovoid shape, with a whitish
coloration, and a diameter of approximately 1.37 mm (Fig. 1A). When ruptured, the cysts
revealed numerous ellipsoidal spores encased in a wall, with two equal polar capsules in the
anterior region of the spore, and sporoplasm, showing all characteristics of the genus
Myxobolus (Figs. 2A, 1B). A number of anomalous spores were also observed under light
microscopy (Figs. 2B, C).

47
TABLE 1. Comparison between Myxobolus arariensis (mean measurements in μm) and other
Myxobolus spp. spores, previously described, infecting the musculature of the
different freshwater fish species.
Characteristics of some Myxobolus species. Abbreviations: FC = Capsule Formate, SL, spore length, SW = Spore Width,
PCL = Polar Capsule Length, PCW= Polar Capsule Width. PC = relative size of the polar capsules (= = equal in size, # =
different in size, or equal and different); All measurements are given in micrometers.
Myxobolus species Host FC SL SW PCL PCW PC Country Order
M. terengganuensis Székely, et al.
(2009b)
Osteochilus
hasselti
(Valenciennes, 1842)
ellipsoidal 12.7
(12.0–13.4)
7.4
(6.7–8.3)
3.2 (2.9–
3.4)
6.8 (6.2–7.3)
2.3
(2.2–2.6)
3.2 (2.9–3.4)
# Malaysia Cypriniforme
M. tasikkenyirensis
Székely, et al. (2009b)
Osteochilus vittatus
(Valenciennes,
1842)
pyriform 12.8
(11.8–13.8)
9.2
(8.3–9.9)
6.4 (5.9–
7.2)
3.0
(2.7–3.4) = Malaysia Cypriniforme
M. tauricus
Miroshnichenko
(1979)
Luciobarbus bocagei
(Steindachner,
1864)
ellipsoidal 13.0
(11.5–14.5) 9–11
6
8.5
2.7
3.5 # Portugal Cypriniforme
M. groenlandicus
Buchmann et al.
(2012)
Reinhardtius
hippoglossoides
(Walbaum, 1792)
round 10.3
(8.5–11.0) 10.1
(9.1 – 11.2) 4.4 (4.0–
5.1) 2.5
(2.1–4.1) = Greenland
Pleuronectiformes
M. omari
Székely et al.(2009a)
Pangasianodon
hypophthalmus (Sauvage, 1878)
ellipsoidal 7.9
(7.2–8.8)
12.0
(11.0–13.9)
5.6 (4.0–
6.2)
5.9 (4.4–6.6)
4.3
(3.6–4.9)
4.7 (4.0–5.3)
# Malaysia Siluriforme
M. leptobarbi
Székely et al.
(2009a)
Leptobarbus
hoevenii
(Bleeker, 1851)
oval 16.0
(14.8 - 17) 8.9
(8.4 - 9.6)
9.9 (8.8 – 10.6)
10.5 (9.9 -
11.5)
3.0 (2.3- 3.6)
# Malaysia Cypriniforme
M. lentisuturalis
Dyková et al. (2002)
Carassius auratus
auratus (Linnaeus, 1758)
ellipsoidal 11.8
(11.2–12.4)
7.6
(7.2–8.4)
4.2 (4.0–
4.4)
2.5
(2.0–2.8) = Italy
Cypriniforme
s
Myxobolus
arariensis Present study
Rhamdia quelen
(Quoy & Gaimard, 1824)
ellipsoidal 11.4
(10.7-12.6)
7.2
(6.4-7.9)
4.0 (3.6-
4.3)
1.9
(1.7-2.2) = Brazil Siluriforme

48
Myxobolus arariensis n. sp. (Figs 1-3)
Morphological description - Mature spores are ellipsoidal in shape, with a mean
length of 11.4 μm (10.7-12.6) and mean width of 7.2 μm (6.4-7.9). Each spore contains
two polar capsules (PCs) of equal size 4.0 ± 0.7 μm (3.6-4.3) long and 1.9 ± 0.36 μm
(1.7-2.2) in width (Figs. 1B and 3) (Table I).
FIGURE 1: A. Musculature of Rhamdia quelen (Quoy and Gaimard 1824) infected by
Myxobolus arariensis n. sp. showing the whitish, ovoid pseudocyst (arrow) Scale bar: 0.6 mm.
B. Fresh spores, frontal view (arrow heads). Scale bar: 10 μm.
FIGURE 2: Fresh spores of Myxobolus arariensis n. sp. from Rhamdia quelen (Quoy and
Gaimard 1824). A – The frontal view of the spore. Scale bar: 5 μm. B - Lateral view of the
spore. Scale bar: 5 μm. C - Spores of Myxobolus arariensis n. sp. showing external
ornamentation (arrow head) and anomalous external morphology (*), with caudal filaments
(arrows), but no ornamentation of the external wall. Scale bar: 10 μm

49
Type host - Rhamdia quelen (Quoy & Gaimard 1824)
Site of infection - epaxial and hypaxial layers of the musculature, with plasmodia
containing numerous spores.
Type-locality - Arari River, Cachoeira do Arari on Marajó Island, northern Brazil.
Etymology - The species was named for the locality of origin, the Arari River in
northern Brazil.
Specimens deposited - Microscope slides containing spores from the muscle layer,
prepared using the paraffin technique, stained in Gutierrez and mounted in Entellan
were deposited in the International Protozoan Type Collection of the National Institute
for Amazonian Research (INPA) in Manaus, Amazonas state, Brazil (catalog number:
INPA/027). The partial 18S rDNA sequence was deposited in GenBank under accession
number MG572219.
Prevalence – Three of twenty-five R. quelen examined 12% (3/25) had plasmodia
of an unknown parasite from the genus Myxobolus.
FIGURE 3: Schematic drawing of Myxobolus arariensis n. sp. spore in frontal view found in
the muscle. Spore in valvular view, showing its internal organization.

50
Remarks - M. arariensis can be differentiated morphologically from all seven
Myxobolus species known to infect the muscle tissue of freshwater fish (Table I). The
new species can be distinguished from M. tasikkenyirensis (Székely et al. 2009a) and M.
groenlandicus (Buchmann et al. 2012) by the different shape of the anterior extremity
of the spores, and from M. leptobarbi (Székely et al. 2009b) by the same trait. The
length of M. arariensis (11.4 µm) is most similar to that of M. lentisuturalis (Dyková et
al. 2002), which is 11.8 µm long, whereas M. omari (Székely et al. 2009b) is the
shortest species, at 7.9 µm. Anomalous spores with a caudal filament and lack of
ornamentation on the external wall were also observed (Fig. 2C).
Histology - The histological analysis revealed the presence of cysts of M. arariensis
lodged in the fibers of the skeletal muscles (Fig. 4). Immature spores were observed in
the most external layer of the cyst, with mature spores being found more internally. The
cyst wall is thick and fibrous, and the adjacent musculature was compressed, with the
sarcoplasm frayed, and evidence of a necrotic reaction caused by this compression.
FIGURE 4: A. Histological section of the cyst and musculature of Rhamdia quelen stained with
Gutierrez, showing mature spores (MS) and imature spores (IS) of Myxobolus arariensis n. sp.
(*), causing bulging (arrow) and disorganization of the muscle fibers (M). B. thick fibrous (F)
wall , the adjacent musculature (M) and infiltrate inflamatory (IF).
Molecular data - In the molecular analysis, the specific pair of myxozoan primers
(MC5-MC3) amplified 974 bps of the 18S rDNA gene of the spores obtained from the
plasmodia found infecting the musculature of R. quelen. The BLAST search of the 18S
rDNA sequence data (974 bps) of the Myxobolus species parasitizing R. quelen found
no identical myxozoan sequence in GenBank, although a similarity of at least 85% was
found with four species: Myxobolus cordeiroi (KF296353, 90% similarity), Myxobolus

51
sp. GA2 (KU170935, 86%), Myxobolus lentisuturalis (AY278563, 85%), and
Myxobolus cultus (HQ613409, 85%).
The optimal evolutionary model for maximum likelihood (ML) and Bayesian
analysis were determined by jModelTest 3.0 (Posada, 2008) which identified the best
evolutionary model as the general time reversible model (GTR + I + G), using Akaike
information criteria. Nucleotide frequencies were estimated from the data (A = 0.2574,
C = 0.1848, G = 0.2625, T = 0.2326) and six rates of nucleotide substitution calculated
as AC = 0.8659, AG = 2.6388, AT = 1.7658, CG = 0.4883, CT = 3.4814, GT = 1.000.
The proportion of invariable site was 0.5565 and the alpha value of gamma distribution
parameter 0.3612. Two independent runs were conducted with 4 chains for 2 million
generations for Bayesian analysis. Ceratomyxa shasta (AF001579) e C. amazonensis
(KX236169) was designated as outgroup. Phylogenetic trees were sampled every 100-
generation.
In the phylogenetic analysis, trees generated by Bayesian Inference (BI) had similar
topologies, but with different support values at some nodes. A strong clustering
tendency was found according to phylogenetic affinities. The phylogram indicated the
existence of three clades, A, B and C the first paraphyletic, includes species of
Henneguya and Myxobolus formed by the freshwater and marine water (Mugiliformes).
The clade A subdivide into 2 subclades, A1 and A2. The subclade A1 shows M.
arariensis grouping on the same branch with M. cordeiroi and with adjacent subclade
with M. marajoensis species, having same host. The other subclade, A2 have the
presence of the Myxobolus and Henneguya, corroborating the characteristic of the
Myxobolus genus to be paraphyletic. On the other hand, the clades B and C have
agrouped species of various Myxobolus parasites of freshwater fish belonging to same
and different Order (Table III).
The type of host defines a well-supported freshwater and marine water clade of
Myxobolus and Henneguya (clade A) that infect fish of the orders Siluriformes,
Mugiliformes, Characiformes and Perciformes. However, M. arariensis, which infects
silurids, evolved independently from the Myxobolus subclade A1 that infects fish of the
families Pimelodidae and Ictaluridae. The tree presented a similar topology for the other
clades, clustering according to the taxonomic order of the host. The clade B groups
Myxobolus, of the orders Cypriniformes and Perciformes. The clade C is also
compounded by Myxobolus groups that parasitize hosts of the different orders
Salmoniformes, Siluriformes, Characiformes and Cypriniformes (Fig. 5).
The p distance found between M. arariensis and any other Myxobolus species that
had Siluriformes as host ranged from 11.6 to 22.1% (Table II), which reinforces the
definition of M. arariensis as a new species. The myxosporidian sequences analysed are
showing in the Table 3.

52
FIGURE 5: Phylogenetic tree generated by Bayesian Inference (BI) applied to the partial SSU rRNA gene sequences of Myxobolus arariensis sp. n.
and related myxosporeans, rooted at Kudoa thyrsites and K. alliaria. The GenBank accession numbers are shown adjacent to the species names. The
numbers at the nodes indicate the bootstrap support for the Bayesian Inference. M. arariensis is shown in bold type. The taxonomic order of all the
species selected for the phylogenetic analysis is shown. An asterisk indicates any values below 50%. GenBank Accession numbers follow the species
name.

47
TABLE 2. Pairwise distances based on fragments of the 18S rDNA gene of Myxobolus arariensis n. sp. and
those hosts of Siluriforms of freshwater species only infect by Myxobolus.
1 2 3 4 5 6 7 8
Myxobolus arariensis
Myxobolus marajoensis 0.158
Myxobolus cordeiroi 0.119 0.096
Myxobolus miyairii 0.134 0.072 0.116
Myxobolus flavus 0.119 0.051 0.093 0.099
Myxobolus hakyi 0.122 0.057 0.101 0.107 0.104
Myxobolus pangasii 0.125 0.060 0.104 0.110 0.107 0.119
Myxobolus omari 0.164 0.137 0.104 0.113 0.119 0.131 0.096
Myxobolus terengganuensis 0.221 0.185 0.170 0.149 0.158 0.134 0.134 0173
TABLE 3. The myxosporidian sequences included in the present analysis
Access
Number
Myxosporidian
species
Host Site of
infection
Locality Family Order
KF296353 Myxobolus
cordeiroi
Zungaro jahu visceral
cavity
Brazil Pimelodidae Siluriformes
KX857727 Myxobolus
marajoensis
Rhamdia quelen intestine Brazil Pimelodidae Siluriformes
AF021881 Henneguya exilis Ictalurus
punctatus
gill Brazil Ictaluridae Siluriformes
KP404438 Henneguya
mississipiensis
Ictalurus
punctatus
gill USA Ictaluridae Siluriformes
EU492929 Henneguya adiposa Ictalurus
punctatus
conjunctive
tissue
USA Ictaluridae Siluriformes
AY129318 Myxobolus bizerti Mugil cephalus gill Tunisia Mugilidae Mugiliformes
JF810537 Myxobolus
episquamalis
Mugil cephalus wild mullet South
Korea
Mugilidae Mugiliformes
AF378341 Myxobolus
spinacurvatura
Mugil sp brain
mesentery
Tunisia Mugilidae Mugiliformes

54
AF378337
Myxobolus
ickeulensis
Mugil sp - Tunisia Mugilidae Mugiliformes
AY129317 Myxobolus exiguus Liza ramada intestine Tunisia Mugilidae Mugiliformes
KF296345 Henneguya
maculosus
Pseudoplatysto
ma reticulatum
gill Brazil Pimelodidae Siluriformes
HQ655111 Henneguya eirasi Pseudoplatysto
ma spp
gill Brazil Pimelodidae Siluriformes
KT001495 Myxobolus miyairii Silurus asotus intestine China Siluridae Siluriformes
KF296346 Myxobolus flavus Pseudoplatysto
ma corruscans
gill Brazil Pimelodidae Siluriforme
FJ816269 Myxobolus hakyi Pangasianodon
hypophthalmus
serosa Thailandia Pangasiidae Siluriforme
FJ816270 Myxobolus pangasii Pangasius
hypophthalmus
spleen
Malasia Pangasiidae Siluriforme
KJ849240 Myxobolus
filamentum
Brycon
orthotaenia
gill filaments Brazil Bryconidae Characiformes
HM754633 Myxobolus oliveirai Brycon hilarii gill filaments Brazil Bryconidae Characiformes
KF296349 Myxobolus
pantanalis
Salminus
brasiliensis
gill Brazil Bryconidae Characiformes
KC771143 Henneguya visibilis Leporinus
obtusidens
connective
tissue
Brazil Anostomidae Characiformes
KF296352 Henneguya
pellucida
Piaractus
mesopotamicus
visceral
cavity / swim
bladder
Brazil Serrassalmidae Characiformes
EU732599 Henneguya sp ex
Perca
Perca fluviatilis gill Brazil Percidae Perciformes
AF378343 Myxobolus sp. KAB Catostomos
commersoni
- USA Catostomidae Cypriniformes
AY278563 Myxobolus
lentisuturalis
Carassius
auratus
muscle Italy Cyprinidae Cypriniformes
HQ613409 Myxobolus cultus Carassius
auratus
gill China Cyprinidae Cypriniformes

55
AB274267 Myxobolus
nagaraensis
Rhinogobius sp. body cavity Japan Gobiidae Perciformes
KF211436 Myxobolus sp HB Ophiocara
porocephala
gill lamellae Malaysia Eleotridae Perciformes
EF370479 Myxobolus
cerebralis
Oncorhynchus
mykiss
brain USA Salmonidae Salmoniformes
JX910362 Myxobolus
squamalis
Oncorhynchus
mykiss
skin USA Salmonidae Salmoniformes
AF085180 Myxobolus
neurobius
Oncorhynchus
mykiss
neural tissue USA Salmonidae Salmoniformes
EU643624 Myxobolus omari Pangasianodon
hypophthalmus
muscle Malaysia Pangasiidae Siluriformes
KP990667 Myxobolus sp1HS-2
015
Clarias
gariepinus
rim Egito Clariidae Siluriformes
KF296348 Myxobolus aureus Salminus
brasiliensis
kidney Brazil Bryconidae Characiforme
KF296351 Myxobolus
piraputangae
Brycon hilarii kidney Brazil Bryconidae Characiforme
KF296350 Myxobolus umidus Brycon hilarii spleen
Brazil Bryconidae Characiforme
KF296358 Myxobolus
macroplasmodialis
Salminus
brasiliensis
visceral
cavity
Brazil Bryconidae Characiforme
DQ439809 Myxobolus gayerae Leusciscus
cephalus
intestine Hungria Cyprinidae Cypriniformes
EU567312 Myxobolus
shaharomae
Alburnus
alburnus
blood vessels Hungary Cyprinidae Cypriniformes
DQ439805 Myxobolus
cyprinicola
Cyprinus carpio intestine Hungary Cyprinidae Cypriniformes
DQ439806 Myxobolus muelleri Leuciscus
cephalus
gill Hungary Cyprinidae Cypriniformes
EU643629 Myxobolus
terengganuensis
Pangasianodon
hypophthalmus
muscle Malasia Pangasiidae Siluriforme

56
Discussion
The histological analyses revealed the presence of Myxobolus infecting the musculature
of R. quelen. These myxosporidians are known to parasitize bony fishes (Osteichthyes),
infecting a variety of sites, including the gill of the characiform, Prochilodus lineatus
(Valenciennes 1836), which was infected by Myxobolus lomi (Azevedo et al. 2014), the
intestine of the siluriform R. quelen (Abrunhosa et al. 2017), infected by M.
marajoensis, and the nervous system of the gymnotiform Eignmannia sp., infected by
Myxobolus sp (Sindeaux-Neto et al. 2016).
In the case of the Myxobolus species that infect the skeletal musculature of fish,
Székely et al. (2009a) recorded infections in three cyprinid fishes, the silver shark
minnow, Osteochilus hasselti (Valenciennes 1842), the bonylip barb, Osteochilus
vittatus (Valenciennes 1842), and Hoven’s carp, Leptobarbus hoevenii (Bleeker 1851).
In Brazil, Manrique et al. (2015) recorded cases in the characiform the pacu Piaractus
mesopotamicus (Holmberg 1887) and the striped catfish Pangasianodon hypophthalmus
(Sauvage 1878), a siluriform.
Clear differences were found between the morphology of M. arariensis and other
freshwater Myxobolus species, in relation to the size and shape of the spore and the
polar capsules, and their distribution (Table I), reinforcing the identification of the new
species. M. arariensis is shorter than M. leptobarbi, M. terengganuensis, M.
tasikkenyirensis, M. tauricus and M. lentisuturalis, but larger than M. groenlandicus,
although M. omari is the shortest species yet found in a siluriform. Overall, the length of
M. arariensis was most similar to that of M. lentisuturalis.
In M. arariensis, the polar capsules are equal in size and relatively small in
comparison with those of M. tasikkenyirensis, M. tauricus, M. groenlandicus, and M.
lentisuturalis. The species M. leptobarbi, M. terengganuensis, and M. omari are distinct
because they have polar capsules of unequal size. The elipsoidal shape of the spores of
M. arariensis is similar to that of M. terengganuensis, M. tauricus, M. lentisuturalis and
M. omari (Table I), but with different lengths and ornamentation patterns.
Spherical cysts delimited by a fibrous layer, similar to those observed in the present
study, were also described in the genus Myxobolus by Chun-Li and Sato (2014).
Milanin et al. (2010) describe a Myxobolus cyst with immature spore stages in its
periphery, and mature spores in its central portion, similar to the configuration recorded
in the present study.
The phylogenetic analysis based on the comparisons of the 18S rDNA gene
sequences of 33 Myxobolus and 8 Henneguya species obtained from GenBank
reinforced the existence of a new species. Using this same gene, Adriano et al. (2009)
described M. cordeiroi infecting the gill, the serosa of the body cavity, the urinary
bladder, and the eye of the siluriform Zungaro jahu Humboldt, 1821 in the Pantanal
wetlands of western Brazil. Xi et al. (2013) also used this gene to describe M. cultus in
the gill of the Crucian carp, Carassius cuvieri Temminck and Schelegel, 1846.

57
In clade A, there was a tendency for the formation of groups strongly related to
their host species. The phylogenetic proximity found between Myxobolus and
Henneguya may reflect coevolutionary processes, ranging from the common ancestor of
the two genera to the relationships between parasite and host. Gleeson and Adlard et al.
2012, Hartigan et al. 2012 and Adriano et al. 2012 demonstrated that many factors
influence species clustering, such as the phylogenetic proximity of the host, tissue
tropism, geographic distribution and morphology characteristics.
The cladogram (Fig. 2) showed that these parasites were more closely related to the
orders of their hosts than to any morphological or biogeographic similarities. In the
specific case of clade A2, the fish parasitized by Myxobolus species group according to
the type of environment (freshwater or marine) and taxon (Fiala 2006, Ferguson et al.
2008, Adriano et al. 2012, Brougthon et al. 2013, Carriero et al. 2013, Moreira et al.
2014a).
Overall, the results of the morphological and phylogenetic analyses indicated
conclusively that M. arariensis n. sp. is a new myxosporidian species, providing new
insights into the characteristics of the microparasite fauna of R. quelen.
Bioethics and Biosecurity Committee Approval
The present study was approved by the Ethics Committee for Animal Experimentation
of the Federal Rural University of Amazonia, UFRA (CEUA approval no. 013/2014).
Acknowledgements
To the research funding agencies CAPES (Coordination of Higher Education Training)
through PVE 88881.064967/2014-01, CNPq (Brazilian National Council for Scientific
and Technological Development) through CNPq UNIVERSAL/2014, N. 441645/2014-
3, and a productivity fellowship to Edilson Rodrigues Matos (CNPq 300949/2012-0),
FAPESPA (Pará State Research Foundation) research notice 006/2014, icaaf
n°162/2014. We also thank SISBIO/ICMBIO-IBAMA (Brazilian Institute of the
Environment and Renewable Natural Resources), for authorizing the research through
scientific license no. 27119, and the Edilson Matos Research Laboratory (LPEM-
UFPA).

58
References
Abrunhosa, J., Sindeaux-Neto, J., Santos, A.K.D., Hamoy, I., Matos, E. (2017)
Myxobolus marajoensis sp. n. (Myxosporea: Myxobolidae), parasite of the
freshwater catfish Rhamdia quelen from the Brazilian Amazon region. Revista
Brasileira de Parasitologia Veterinária, 26(4): 465-471.
Adriano, E.A., Arana, S., Alves, A.L., Silva, M.R., Ceccarelli, P.S., Henrique-Silva, F.,
Maia, A.A. (2009) Myxobolus cordeiroi n. sp., a parasite of Zungaro jahu
(Siluriformes: Pimelodiade) from Brazilian Pantanal: Morphology, phylogeny and
histopathology. Veterinary Parasitology, 162:221–229.
Adriano, E.A., Carriero, M.M., Maia, A.A.M., Silva, M.R.M., Naldoni, J., Ceccarelli,
P.S., Arana, S. (2012) Phylogenetic and host-parasite relationship analysis of
Henneguya multiplasmodialis n. sp infecting Pseudoplatystoma spp. in Brazilian
Pantanal wetland. Veterinary Parasitology, 185: 110–120.
Azevedo, R.K., Vieira, D.H.M.D., Vieira, G.H., Silva, R.J., Matos, E., Abdallah, V.D.
(2014) Phylogeny, ultrastructure and histopathology of Myxobolus lomi sp. nov., a
parasite of Prochilodus lineatus (Valenciennes, 1836) (Characiformes:
Prochilodontidae) from the Peixes River, São Paulo State, Brazil. Parasitology
International, 63: 303–307.
Baldisseroto, B., Radunz Neto, J. (2004) Criação de Jundiá. Ed. UFSM, Santa Maria,
232 pp.
Barthem, R.B., Goulding, M. (2007) Um ecossistema inesperado: a Amazônia revelada
pela pesca. Amazon Conservation Association (ACA), Sociedade Civil Mamirauá,
Belém, 241 pp.
Boreham, R.E., Hendrick, S., O’Donoghue, P.J., Stenzel, D.J. (1998) Incidental finding
of Myxobolus spores (Protozoa: Myxozoa) in stool samples from patients with
gastrointestinal symptoms. Journal of Clinical Microbiology, 36: 3728-3730.
Brasil. Ministério da Pesca e Aquicultura. (2011) Boletim estatístico da pesca e
aquicultura 2011. Brasília: MPA. 60 pp.
Buchmann, K., Skovgaard, A., Kania, P.W. (2012) Myxobolus groenlandicus n. sp.
(Myxozoa) distorting skeletal structures and musculature of Greenland halibut
Reinhardtius hippoglossoides (Teleostei: Pleuronectidae). Diseases of Aquatic
Organisms, 98: 133–141.
Carriero, M.M., Adriano, E.A., Silva, M.R., Ceccarelli, P.S., Maia, A.A. (2013)
Molecular phylogeny of the Myxobolus and Henneguya genera with several new
South American species. Plos One, 8 (9):e73713.
Chun-Li, Y., Sato, H. (2014) Two novel myxosporean species (Myxosporea:
Bivalvulida), Myxobolus marumotoi n. sp. and Cardimyxobolus japonensis n. sp.,

59
from the dark sleeper, Odontobutis obscura, in Japan. Parasitology Research,
113:1371–1381.
Dyková, I., Fiala, I., Nie, P. (2002) Myxobolus lentisuturalis sp. n. (Myxozoa:
Myxobolidae), a new muscle-infecting species from the Prussian carp, Carassius
gibelio from China. Folia Parasitologica, 49: 253-258.
Ferguson, J.A., Atkinson, S.D., Whipps, C.M., Kent, M.L. (2008) Molecular and
Morphological analysis of Myxobolus spp. of salmonid fishes with the description
of a new Myxobolus species. Journal of Parasitology, 94(6): 1322–1334.
Fiala, I. (2006) The phylogeny of Myxosporea (Myxozoa) based on small subunit
ribosomal RNA gene analysis. International Journal for Parasitology, 36: 1521–
1534.
Gleeson, R.J., Adlard, R.D. (2012) Phylogenetic relationships amongst Chloromyxum
Mingazzini, 1890 (Myxozoa: Myxosporea), and the description of six novel species
from Australian elasmobranchs. Parasitology International, 61: 267–274.
Hall, T. (2007) BioEdit. Biological Sequence Alignment Editor for
Win95/98/NT/2K/XP. Carlsbad, CA: Ibis Biosciences.
Hartigan, A., Fiala, I., Dyková, I., Rose, K., Phalen, D.N., Šlapeta, J. (2012) New
species of Myxosporea from frogs and resurrection of the genus Cystodiscus Lutz,
1889 for species with myxospores in gallbladders of amphibians. Parasitology,
139(4): 478–496.
Hedrick, R.P. (2011) Relationships of the host, pathogen, and environment: implications
for diseases of cultured and wild fish populations. Journal of Aquatic Animal
Health, 10:107–111.
Hessen, E.M., Zamzame, M.L. (2004) Myxobolus sp: a possible new opportunistic
parasite in immunocompromized patients in Ismailia. Journal of the Egyptian
Society of Parasitology, 34(3): 925-930.
Kudo, R. (1921) On the effect of some fixatives upon myxosporidian spores.
Transactions of the American Microscopical Society, 40: 161-167.
Liu, Y., Gum, Z.M., Luo, Y.L. (2010) Some additional data to the occurrence,
morphology and validity of Myxobolus turpisrotundus Zhang, 2009 (Myxozoa:
Myxosporea). Parasitology Research, 107: 67-73.
Luna, L.G. (1968) Manual of histologic staining methods of the Armed Forces Institute
of Pathology. 3rd edn. McGraw- Hill, New York, NY, 258 pp.
Manrique, W., Figueiredo, G.M.P., Belo, M.A.A., Martins, M.L, Moraes, F.R. (2015)
First report of Myxobolus sp. infection in the skeletal muscle of Neotropical
freshwater fish Piaractus mesopotamicus. Parasitology Research, 114: 2041–2044.
Martinez de Velasco, G., Rodero, M., Cuellar, C., Chivato, T., Mateos, J.M., Laguna, R.
(2018) Skin prick test of Kudoa sp. antigens in patients with gastrointestinal and/or
allergic symptoms related to fish ingestion. Parasitology Research, 103: 713-715.

60
Matos, E., Tajdari, J., Azevedo, C. (2005) Ultrastructural studies of Henneguya rhamdia
n. sp. (Myxozoa) a parasite from the Amazon Teleost fish (Rhamdia quelen)
(Pimelodidae). Journal of Eukaryotic Microbiology, 52(6): 532-537.
Matos, E., Videira, M., Velasco, M., Sanches, O., São Clemente, S.C., Matos, P. (2014)
Infection of the heart of Pimelodus ornatus (Teleostei, Pimelodidae), by
Myxobolus sp. (Myxozoa, Myxobolidae). Revista Brasileira de Parasitolia
Veterinária, 23 (4): 543-546.
Milanin, T., Eiras, J.C., Arana, S., Maia, A.A.M., Alves, A.L., Silva, M.R.M., Carriero,
M.M., Ceccarelli, P. S., Adriano, E.A. (2010) Phylogeny, ultrastructure,
histopathology and prevalence of Myxobolus oliveirai sp. nov., a parasite of Brycon
hilarii (Characidae) in the Pantanal wetland, Brazil. Memórias do Instituto Oswaldo
Cruz, 105(6): 762-769.
Molnár, K., Eszterbauer, E., Székely, C., Dán, Á., Harrach, B., 2002. Morphological
and molecular biological studies on intramuscular Myxobolus spp. of cyprinid fish.
J. Fish Dis. 25, 643–652.
Molnár. K., Marton, S., Eszterbauer, E., Székely, C. (2006) Comparative morphological
and molecular studies on Myxobolus spp. infecting chub from the River Danube,
Hungary, and description of M. muellericus sp. n. Diseases of Aquatic Organisms,
73: 49–61.
Molnár, K.E., Marton, E., Székely, S.Z.C.S., Eiras, J.C. (2012) Comparison of the
Myxobolus fauna of common barbel from Hungary and Iberian barbel from Portugal.
Diseases of Aquatic Organisms, 100: 231–248.
Moncada, L.I., Lopez, M.C., Murcia, M.I., Nicholis, S.L.F., Guio, O.L., Corredor, A.
(2001) Myxobolus sp., another opportunistic parasite in immunosuppressed patients.
Journal of Clinical Microbiology, 39(5): 1938-1940.
Moreira, G.S.A., Adriano, E.A., Silva, M.R.M., Ceccarelli, P.S., Maia, A.A.M. (2014)
The morphological and molecular characterization of Henneguya rotunda n. sp., a
parasite of the gill arch and fins of Salminus brasiliensis from the Mogi Guaçu River,
Brazil. Parasitology Research, 113: 1703–1711.
Rambaut, A. (2008). FigTree v. 1.4.0. Rambaut. Available from
http://tree.bio.ed.ac.uk/software/figtree/. (Accessed 20 December 2017).
Rocha, S., Matos, E., Velasco, M., Casal, G., Alves, A., Azevedo, C. (2015)
Ultrastructure and phylogeny of Thelohanellus sp. (Myxozoa: Myxosporea) infecting
the gills of Hypophthalmus marginatus (Actinopterygii: Pimelodidae), a fish from
the Amazon River. Microscopy and Microanalysis, 21 (SupplementS6), 46-47.
Ronquist, M.A.F., Huelsenbeck, J.P. (2003) MrBayes 3: Bayesian phylogenetic
inference under mixed models. Bioinformatics, 19: 1572–1574.
Silva, M.C., São Clemente, S.C. Picanço Júnior, J.A., Silva, M.V.O., Matos, E.R.
(2012) Calyptospora sp. in Brachyplatystoma vaillantii trapped at the Vigia, State of
Pará, Brazil. Revista Brasileira de Parasitologia Veterinária, 21(2): 176-178.

61
Sindeaux Neto, J.L., Velasco, M., Silva, J.M.V, Santos, P.F.S., Sanches, O., Matos, P.,
Matos, E. (2016) Lymphocytic meningoencephalomyelitis associated with
Myxobolus sp. (Bivalvulidae: Myxozoa) infection in the Amazonian fish
Eigenmannia sp. (Sternopygidae: Gymnotiformes). Revista Brasileira de Parasitolia
Veteterinaria, 25(2): 158-162.
Székely, C.S., Shaharom-Harrison, F., Cech, G., Mohamed, K., Molnár, K. (2009a)
Myxozoan pathogens of Malaysian fishes cultured in ponds and net-cages. Diseases
of Aquatic Organisms, 83: 49–57.
Székely, C.S., Shaharom-Harrison, F., Cech, G., Ostoros, G., Molnár, K. (2009b)
Myxozoan infections in fishes of the Tasik Kenyir Water Reservoir, Terengganu,
Malaysia. Diseases of Aquatic Organisms, 83: 37–48.
Snieszko, S.F. (1974) The effects of environmental stress on outbreaks of infectious
diseases of fishes. Journal of Fish Biology, 6:197–208.
Swofford, D.L. (2003). PAUP*. Phylogenetic analysis using parsimony (*and other
methods). Version 4. Sinauer Associates, Sunderland.
Velasco, M., Videira, M., Silva, J.M.V., Sanches. O., Matos, P.S., São Clemente, S.C.,
Matos, E. (2015a) Esophageal infection due to Kudoa sp. (Myxozoa) in mapara
catfish, Hypophthalmus marginatus. Aquaculture Reports, 2:22-25.
Velasco, M., Videira, M., Sindeaux-Neto, J.L., Santos, P., Sanches, O., Matos, P.,
Matos, E. Infection by Henneguya sp. (Myxozoa) in the bone tissue of the gill
filaments of the Amazonian catfish Hypophthalmus marginatus (Siluriformes).
Revista Brasileira de Parasitologia Veterinária, 24:365-369.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G. (1997) The
CLUSTAL–X window interface: flexible strategies for multiple sequence alignment
aided by quality analysis tools. Nucleic Acids Research, 25: 4876–4882.
Thompson, K.G., Nehring, R.B., Bowden, D.C., Wygant, T. (2002) Response of
rainbow trout Oncorhynchus mykiss to exposure to Myxobolus cerebralis above and
below a point source of infectivity in the upper Colorado River. Diseases of Aquatic
Organisms, 49(3): 101-178.
Xi, B.W., Zhang, J.Y., Xie, J., Pan, L.K., Xu, P., Ge, P.G. (2013) Three actinosporean
types (Myxozoa) from the oligochaete Branchiura sowerbyi in China. Parasitology
Research, 112: 1575-1582.
Zhao, Y., Chunyan, S., Kent, M.L., Deng, J., Whipps, C.M. (2008) Description of a
New Species of Myxobolus (Myxozoa: Myxobolidae) Based on Morphological and
Molecular Data. Journal of Parasitology, 94(3): 737–742.

62
ARTIGO IV
Título: A new species of Henneguya from silver catfish in Amazonia
Autores: Jacqueline Abrunhosa, José Ledamir Sindeaux-Neto, Igor Hamoy, Edilson
Matos
Revista: Parasitology Research
Status: Submetido
ISSN: 0932-0113
Fator de impacto: 2,329 (Qualis B2)

63
A new species of Henneguya from silver catfish in Amazonia
Jacqueline ABRUNHOSA1,3
, José Ledamir SINDEAUX-NETO1,3
, Igor HAMOY2,
Edilson MATOS3*
1Programa de Pós-Graduação em Biologia de Agentes Infecciosos e Parasitários (BAIP)
Universidade Federal do Pará (UFPA), Belém, Brazil;
2 Laboratório de Genética Aplicada, Universidade Federal Rural da Amazônia (UFRA),
Belém, Brazil
3Laboratório de Pesquisa Carlos Azevedo, Universidade Federal Rural da Amazônia
(UFRA), Belém, Brazil;
*Corresponding autor: Edilson Matos. Avenida Presidente Tancredo Neves, 2501,
Montese Cep: 66.077-901, Belém, Pará, Brasil - e-mail: [email protected]
Abstract
Henneguya quelen n. sp. (Myxosporea) is described from the kidney of the silver
catfish, Rhamdia quelen, from Marajó Island, in the Brazilian Amazon region. In all,
40% of the 57 silver catfish collected from the Paracauari River in the municipality of
Salvaterra presented cysts in the kidney. The cysts were whitish, spherical, and 0.45 mm
in diameter. The histological analyses revealed tubular renal compression and cellular
degeneration in the area adjacent to the xenomas. The spores are ellipsoid, with well-
marked valves, caudal projection, and two elongated polar capsules of equal size. The
mature spores measure approximately 15.6 µm in length, with the caudal appendage
split posteriorly, 24.3 µm in length, for a total spore length of 40.0 µm, and a width of
4.1 µm. The polar capsules are pyriform, 5.5 µm long and 1.7 µm wide. A phylogenetic
analysis of the morphometric traits based on Bayesian Inference confirmed that the
specimens represented a new species, which was denominated Henneguya quelen n. sp.
Keywords: Myxozoa, microparasite, kidney, histology, 18S SSU rRNA
Introduction
The silver catfish Rhamdia quelen (Quoy & Gaimard, 1824) (Pimelodidae,
Siluriforme) is an important fishery resource in Brazilian waters. This species is
important not only for local fisheries in the Amazon region, but is also reared on fish
farms in the extreme south of Brazil (Baldisseroto, 2004; Brasil, 2011). Despite the
potential interest in this fish, few data are available on most aspects of its health, in
particular those that would be relevant to its use as a source of human food.
Myxosporan cnidarian parasites can cause grave damage to the health of their fish
hosts, and have been investigated extensively, with numerous cases of infection by

64
these parasites being reported in a diversity of siluriform species. Henneguya Thélohan,
1892 is the second most prominent and abundant genus of the class Myxosporea
worldwide (Eiras and Adriano 2012), and infections by some Henneguya species are
considered pathogenic. This implies potential economic impacts, especially for fish-
farming operations (Feist and Longshaw 2006; Lom and Dyková 2006; Velasco et al.
2015).
These parasites have been identified as important pathogens in freshwater,
estuarine and marine catfish (Hallett and Diamant 2001; Kalavati and Narasimhamurti
1985; Lom and Dyková 1995; Martins and Souza 1997; Matos et al. 2005). In catfish,
tropism of Henneguya has been recorded in the gills (Lin et al. 1999; Pote et al., 2000),
nervous system (Azevedo et al. 2011), adipose fin (Griffin et al. 2009), dorsal, pectoral
and caudal fins (Iwanowicz et al. 2008), skin (Griffin et al. 2009), and even in the
kidneys and intestine (Vidal et al. 2017).
Henneguya species are difficult to identify based only on the morphological
characteristics of the spores, and a number of authors have recommended the use of
sequences of the 18S rDNA gene to support the diagnosis of new species (Andree et al.
1999; Kent et al. 2001; Lom and Dyková 2006). This approach has contributed to the
description of a number of myxozosporean species in recent years, including forms
found in Brazilian catfish and other fish groups (Abrunhosa et al. 2017; Azevedo et al.
2016; 2014; Carriero et al. 2013; Casal et al. 2017; Matos et al. 2018; Rocha et al.
2014b;Velasco et al. 2017). The molecular 18S rDNA marker has been widely used for
the detection, identification, and phylogenetic analysis of myxosporeans (Holzer et al.,
2006).
In the present study, a myxosporid of the genus Henneguya was observed as a
pathogenic parasite in the kidney of the silver catfish, Rhamdia quelen. The
morphological and phylogenetic analyses of the specimens indicated that these parasites
represented a new species, which is described here as Henneguya quelen n. sp.
Materials and Methods
Fish sampling
Between January and September 2017, samples were obtained from 57 adult
specimens (body length 15–23 cm) of Rhamdia quelen caught by local fishers using
nylon gill nets on the Paracauari River, in the municipality of Salvaterra (00º45’ S,
48º31’ W) on Marajó Island, in the eastern Amazon region of northern Brazil. The fish
were kept alive in plastic bags containing aerated water from their natural habitat, and
transported to the UFRA laboratory in Belém, where they were transferred to aerated
aquariums until necropsy. For necropsy, the specimens were anesthetized with 50 mg/L
tricain methanesulfonate (MS222 SIGMA), following the recommendations of the
UFRA ethics committee for the use of animals in experiments (CUSA: 013/2014).
Sample preparation and histological analysis
Necropsy was initiated with a longitudinal incision from the anogenital opening to
the region of the operculum, made using a scalpel and scissors, to expose the celiac
cavity for the inspection of the internal organs under a stereomicroscope. This
examination revealed a number of rounded, whitish cysts in the kidney. Fragments of
the kidney containing cysts were removed, placed on a microscope slide in a drop of
water and covered with a coverslip for the confirmation of the diagnosis under light
microscopy. Samples of the organ were fixed in Davidson solution for 24 h, and
embedded in paraffin for histological analysis. Sections of 5µ were obtained, and

65
stained with Hematoxylin and Eosin (H&E) and Ziehl Neelsen (ZN) for the
morphological analyses (Luna 1968). The spores (n = 30) were measured, with the
values being presented in micrometers (μm), and photographed using a ZEISS
microscopic Primo Star AxioCam ERc 5s camera with AxioVision 5.1 software.
Molecular and phylogenetic analyses
The cysts were removed from the kidneys and preserved in 80% alcohol for
molecular and phylogenetic analyses. The DNA was extracted using the PureLink® Genomic DNA Mini kit (Invitrogen), following the maker’s protocol. The primers
MC5-MC3 (Molnár 2002) and MX5-MX3 (Andree et al. 1999) were used to amplify
the 18S SSU rDNA gene by Polymerase Chain Reaction (PCR) in the SimpliAmpli
Thermal Cycler (Applied Biosystems). The final volume of the PCR was 25 μL, which
contained approximately 5–10 ng of the DNA template, 2 mM MgCl2, 4 mM of dNTP
mix (Invitrogen), 5 pmol of each primer, and 1.2 units of Taq DNA polymerase
(Invitrogen®). The amplification protocol consisted of denaturation at 95°C for 1 min,
followed by 35 cycles of 66°C for 1 min, 72°C for 2 min and 95°C for 5 min, followed
by final extension at 72°C for 30 min.
Aliquots (3 μL) of the PCR products were visualized with Sybr® safe DNA gel
stain (Invitrogen) following electrophoresis in 1% agarose gel, and purified using GFX
PCR DNA and the Gel Purification kit (GE Healthcare), according to the
manufacturer’s instructions. The samples were sequenced in an ABI 3130 automatic
DNA analyzer (Applied Biosystems) with the BigDye® (Applied Biosystems)
Terminator v3.1 kit, following the manufacturer’s specifications. The MC5-MC3 and
MX5-MX3 primers used to obtain the amplicons were also used for sequencing. The
nucleotide sequences obtained through this process were edited and aligned using the
BioEdit software (Hall 2007).
The partial sequence of the 18S rRNA gene of the Henneguya specimens obtained
from the R. quelen samples was aligned using the BioEdit software (Hall 2007) for
comparisons with the 25 sequences of Myxobolus and Henneguya species available in
GenBank. The outgroup included Ceratomyxa shasta (Bartholomew et al 1997) and
Ceratomyxa amazonensis (Mathews et al 2016).
The phylogenetic relationships were determined through Bayesian inference (BI),
using Markov Chain Monte Carlo (MCMC) tree searches in MrBayes 3.1.2 (Ronquist
and Huelsenbeck 2003). The most appropriate evolutionary model was determined
using jModelTest 2.0.2 (Darriba et al. 2012), based on the Akaike Information Criterion
(AIC). We conducted two parallel runs of four simultaneous MCMC searches of 5
million generations each, sampling one tree every 500 generations, and discarding the
results of the first 1000 trees the burn-in. The remaining trees were used by MrBayes to
estimate the posterior probability of each node in the phylogenetic reconstruction. The
Tracer v1.4.1 program (Rambaut and Drummond 2008) was used to check the
stationarity of all the parameters sampled by the chains, according to the model
indicated by jModelTest 2.0.2 (Darriba et al. 2012). Genetic distances (p) in relation to
other Myxobolus species were determined using PAUP 4.0b (Swofford 1998).
Results
Henneguya quelen n. sp. was observed infecting the kidney of R. quelen from
Marajó Island, Pará, Brazil. The new species is described based on morphological and
molecular analyses. The morphological features and infection site of the new parasite
species were compared with those of other Henneguya species (see Table 1).

66
Overall, 23 (40%) of the 57 R. quelen specimens analyzed presented infection by
the myxosporidian parasite. In the kidney, whitish, spherical cysts were observed, with
a diameter of approximately 0.45 mm (Fig. 1a).
Fig. 1. Desenho esquemático de Henneguya quelen n. sp

67
TABLE 1. Comparative measurements (μm) and other characters of the spores of different species of the genus Henneguya infecting kidney of
brazilian fishes
Abbreviations are as follows: LSB = Length of Spore Body, WSB = Width of Spore Body, LPC = Length of Polar Capsule, WPC = Width of Polar Capsule, CPL = Caudal
Process Length, TSL = Total Spore Length, SI = Site of Infection, NA = data Not Available. When both a mean and a range are available, the range is given in parentheses.
Species LSB WSB CPL LPC WPC TSL Host References
Henneguya sp. 9.5
(8.9-10.1)
6.4
(5.9-6.9)
2.6 3.2
(2.9 –3.5)
1.4
(1.0-1.8)
12.1±
(10.9-13.4)
Piaractus
mesopotamicus
Manrique et
al. 2017
H. schizodon
13.1
(12-14)
3.3
(3-4)
16.3
(15-17)
5.4
(5-6)
1.3
(1-5)
28.9
(27-30)
Schizodon fasciatus Eiras et al.
2004
H. curimata 16.6
(16.0-17.4)
6.2
(5.8-6.6)
19.1
(18.3-19.9)
6.5
(6.2-6.8)
1.2
(1.0-1.4)
35.4
(34.2-36.1)
Curimata inornata Azevedo and
Matos 2002
H. friderici 12.8 ± 2.1
(7.4-14.8)
4.4 ± 0.4
(3.4-5.2)
19.6 ± 2.2
(16.1-24.4)
5.1 ± 0.5
(3.7-5.9)
1.5 ± 0.1
(1.2- 1.8)
32.4
(28.1-36.7)
Leporinus friderici Vidal and
Luque 2017
Henneguya
quelen n. sp.
15.6
(14.3-16.4)
4.1
(3.9-4.4)
24.3
(21-26.5)
5.5
(5.2-6)
1.7
(1.4-1.8)
40.0
(37-42.8)
Rhamdia quelen This study

68
Type host: Rhamdia quelen (Quoy &Gaimard, 1824) (Siluriforme: Pimelodidae)
Type locality: Paracauari River, Salvaterra, Marajó Island Pará, Brazil
Site of infection: kidney
Prevalence of infection: 40% (23/57)
Type material: Slides of spores from the kidney, sections embedded in paraffin and
stained with Hematoxylin & Eosin, mounted in low viscosity medium, deposited in the International Collection of Protozoan Type slides at the National Institute for
Amazonian Research (INPA) in Manaus, Amazonas, Brazil (Catalog number:
INPA/33).
Etymology: The new species was named for the host species: Rhamdia quelen.
Representative sequence: The partial 18S rDNA sequence was deposited in GenBank
under accession number MH230064
Histology: Parasitic cyst within fine, fibrous conjunctive capsule in the renal
parenchyma, causing tubular compression with cellular degeneration in the surrounding
area (Fig 1b).

69
Fig 1. Henneguya quelen n. sp. infecting the kidney of Rhamdia quelen. a. kidney with
whitish cysts (arrow head) macroscopically. Scale bar = 1500µm b. Histological section
of the kidney showing cyst of Henneguya quelen n. sp. (*) stained with H&E. Scale bar
= 50 µm c. Fresh spores. Inset: frontal view, highlighting the polar capsules (arrow).
Scale bar = 5 µm.
Molecular and phylogenetic analyses
A sequence of 1469 base pairs (bps) of the 18S SSU rDNA gene of the spores
found infecting the kidney of R. quelen was amplified successfully. The BLAST search
based on this 1469 bps 18S rDNA sequence found that no other myxozoan sequence

70
available in GenBank is more than 92% similar. These sequences included those of
Henneguya sutherlandi (EF191200), Henneguya ictaluri (AF195510), Henneguya
adiposa (EU492929), Henneguya bulbosus (KM000055), Henneguya pellis (FJ468488),
and the genus Raabeia (KF263539).
The tree generated by Bayesian Inference (BI) placed Henneguya quelen n. sp.
in the same subclade as a sister species of Myxobolus marajoensis, which also
parasitizes R. quelen, and was described by Abrunhosa et al. (2017). Henneguya quelen
n. sp. was paraphyletic, grouping in the same clade as Myxobolus, as often happens in
this genus (Carriero et al. 2013). The affinities of the groupings are arranged by the host
taxa, with the tree being divided into three clades (A, B and C). The major clade A is
divided into two subclades (A1 and A2): clade A1 is made up mainly of Henneguya and
Myxobolus species that infect siluriforms, with the exception of H. pellucida, which
infects a characiform host, and is grouped in a subclade of hosts from the same
geographic region (Brazil). Clade A2 also grouped myxosporidians of the genera
Henneguya and Myxobolus, but with characiform hosts, with the exception of
Henneguya lobosa that has an esociform host, of the family Esocidae (Fig.2). However,
no morphological or evolutionary factor influences this grouping clearly. Clade B
groups Myxobolus lepomis and Henneguya crepili, which have perciform hosts, while
clade C is formed by Henneguya piaractus, which has a characiform host (Fig. 2).
Fig. 2. Phylogenetic tree generated by Bayesian Inference (BI) of the partial sequence of
the 18S SSU rDNA gene of Henneguya quelen sp. n. and closely-related
myxosporidians, rooted by Ceratomyxa shasta and Ceramtomyxa amazonensis. The
GenBank access numbers are presented after the name of each species. The number at
each node represents the bootstrap support for the Bayesian Inference. The taxonomic
order of all the species selected for the phylogenetic analysis is shown. * and **
indicate the orders Characiformes and Esociformes, respectively.

71
The optimal evolutionary models for the maximum likelihood (ML) and
Bayesian analyses were determined by jModelTest 3.0 (Posada, 2008) which identified
the best evolutionary model as the general time reversible model (GTR + I + G), based
on the Akaike Information Criterion (AIC). Nucleotide frequencies were estimated (A =
0.2836, C = 0.1904, G = 0.2754, T = 0.2506) and six rates of nucleotide substitution
were calculated, with AC = 0.7241, AG = 2.9406, AT = 1.000, CG = 0.7241, CT =
5.0822, GT = 1.000. The proportion of invariable sites was 0.2500 and the alpha value
of the gamma distribution parameter was 0.4770. Two independent runs were conducted
with four chains of 2 million generations in the Bayesian analysis. Ceratomyxa shasta
(AF001579) and Ceratomyxa amazonensis (KX236169) was included as the outgroup.
The phylogenetic trees were sampled every 1000 generations. The smallest p distance
between any pair of species assigned to the same clade was 3.26%, between Henneguya
sutherlandi and H. bulbolsus (Tab. 2).

72
Table 2. Pairwise (p) distances between the 18S rDNA sequences of the myxosporidian species that formed a clade with Henneguya quelen n. sp.
in the present study.
Myxosporidians 1 2 3 4 5 6 7 8 9 10 11 12 13 14
1 ) Henneguya quelen
2) H. sutherlandi 0.0326
3) H. ictaluri 0.0371 0.283
4) H. bulbosos 0.0326 0.0132 0.0221
5) H. adiposa 0.0397 0.0291 0.0097 0.0265
6) H. exilis 0.0424 0.0336 0.0088 0.0256 0.0150
7) H. mississipiensis 0.0462 0.0338 0.0124 0.0293 0.0124 0.0133
8) H. lassea 0.0441 0.0289 0.0234 0.0216 0.0297 0.0234 0.0297
9) H. gurlei 0.0413 0.0214 0.0218 0.0156 0.0285 0.0247 0.0304 0.0177
10) M. marajoensis 0.0595 0.0857 0.0835 0.0873 0.0886 0.0876 0.0915 0.0923 0.0952
11) H. pellucida 0.0923 0.0859 0.0814 0.0861 0.0853 0.0805 0.0851 0.0851 0.0851 0.1155
12) Myxobolus miayrii 0.0605 0.384 0.0357 0.0354 0.0444 0.369 0.0412 0.0333 0.0378 0.1069 0.0852
13) H. corruscans 0.0745 0.0780 0.0698 0.0717 0.0753 0.0707 0.0760 0.0705 0.0776 0.1060 0.1114 0.0730
14) M. flavus 0.0514 0.0512 0.0416 0.0474 0.0475 0.0466 0.0464 0.0484 0.0482 0.0886 0.0882 0.0565 0.0713
15) H. eirasi 0.0531 0.0493 0.0440 0.0417 0.0526 0.0430 0.0484 0.0416 0.0440 0.0934 0.0878 0.0569 0.0632 0.0471

73
Table 3. The myxosporidian sequences included in the present analysis
Acess number Mixosporidian species Host Site of
infection
Environment Host Family Host Order Local REF
KF296352 Henneguya pellucida Piaractus
mesopotamicus
visceral
cavity
Freshwater Serrassalmidae Characiformes Brazil Carriero et al. (2013)
KX857727 Myxobolus
marajoensis
Rhamdia quelen intestine Freshwater Pimelodidae Siluriformes Brazil Abrunhosa et al. (2017)
EF191200 Henneguya sutherlandi Ictalurus puctatus skin Freshwater Ictaluridae Siluriformes EUA Griffin et al. (2008)
KM000055 Henneguya bulbosus Ictalurus puctatus gills Freshwater Ictaluridae Siluriformes EUA Rosser et al. (2014b)
KX354352 Henneguya laseeae Pylodictis olivaris gills Freshwater Ictaluridae Siluriformes EUA Leis et al. (2017)
DQ673465 Henneguya gurlei Ameiurus nebulosus caudal fin Freshwater Ictaluridae Siluriformes USA Iwanowicz et al. (2008)
EU492929 Henneguya adiposa Ictalurus puctatus adipose fin Freshwater Ictaluridae Siluriformes EUA Griffin et al. (2009b)
KP404438 Henneguya
mississipiensis
Ictalurus puctatus gills Freshwater Ictaluridae Siluriformes EUA Rosser et al. (2015)
AF195510 Henneguya ictaluri Ictalurus puctatus gills Freshwater Ictaluridae Siluriformes EUA Pote et al. (2000)
AF021881 Henneguya exilis Ictalurus puctatus gills Freshwater Ictaluridae Siluriformes EUA Lin et al. (1999)
KT001495 Myxobolus miyarii Silurus asotus intestine Freshwater Siluridae Siluriformes China Liu et al. (2016)
KF296356 Henneguya corruscans Pseudoplatystoma
reticulatum
gills Freshwater Pimelodidae Siluriformes Brazil Carriero et al. (2013)

74
HQ655111 Henneguya eirasi Pseudoplatystoma
spp
gills Freshwater Pimelodidae Siluriformes Brazil Naldoni et al. (2011)
KF296347
Myxobolus flavus Pseudoplatystoma
reticulatum
gills Freshwater Pimelodidae Siluriformes Brazil Carriero et al. (2013)
KF296349 Myxobolus pantanalis Pseudoplatystoma
corruscans
gills Freshwater Pimelodidae Siluriformes Brazil Carriero et al. (2013)
KJ416131 Henneguya rotunda Salminus brasiliensis fin
membrane
Freshwater Bryconidae Characiformes Brazil Moreira et al. (2014)
KJ849240 Myxobolus filamentum Brycon orthotaenia gill
filaments
Freshwater Bryconidae Characiformes Brazil Naldoni at al. (2015)
HM754633 Myxobolus oliveirai Brycon hilarii gills Freshwater Bryconidae Characiformes Brazil Milanin at al. (2010)
FJ468488 Henneguya pellis Ictalurus furcatus dermis Freshwater Ictaluridae Siluriformes EUA Griffin et al. (2009a)
EU732600 Henneguya lobosa Esox lucius gills Freshwater Esocidae Esociformes Hungary Eszterbauer et al.
(2006)
KY315824 Heneguya friederici Leporinus friderici
gills,liver,
intestine,
kidney
Freshwater Anostomidae Characiformes Brazil Vidal & Luque (2017)
KP980550 Henneguya
leporinicola
Leporinus
macrocephalus
gill
filaments
Freshwater Anostomidae Characiformes Brazil Capodifoglio et al.
(2015)
KY203391 Myxobolus lepomis Lepomis marginatus gill
filaments
Freshwater Centrarchidae Perciformes EUA Rosser at al. (2017)
EU732598 Henneguya creplini Sander lucioperca gills Freshwater Percidae Perciformes Hungary Eszterbauer et al.
(2006)
KF597016 Henneguya piaractus Piaractus
mesopotamicus
gills Freshwater Serrassalmidae Characiformes Brazil Müller et al. (2013)

75
Discussion The morphological characteristics of Henneguya quelen n. sp. are consistent with
those of the genus Henneguya, including spores with an elongated body, fusiform to
ellipsoidal, two elongated bifurcated caudal projections, the presence of striations and
ornamentation on the surface of the body, in addition to a flattening parallel to the suture line.
Straight suture line, formed by an external projection, and two elongated polar capsules (Eiras
et al. 2004; Lom and Dyková 2006).
No previous Brazilian studies have reported the infection of the siluriform kidney by
Henneguya. However, this parasite has been detected in the kidney of four species of
characiform (Table 1), the common aracu, Schizodon fasciatus Spix & Agassiz, 1829,
infected by Henneguya schizodon (Eiras et al. 2004), the pacu, Piaractus mesopotamicus
(Holmberg 1887), infected by Henneguya sp. (Manrique et al., 2017), and the three-spotted
piau, Leporinus friderici, Bloch 1794, infected by Henneguya friederici (Vidal and Luque
2017).
Morphometrically, the Henneguya species that infect these fish all present significant
differences in size in comparison with Henneguya quelen n. sp. The spore of Henneguya
quelen n. sp. is smaller than that of H. curimata, but larger than those of H. friederici, H.
schizodon, and Henneguya sp. The caudal projection of Henneguya quelen n. sp. is longer
than that of any of these species (Table 1). The histological analyses also revealed similarities,
related to the asynchronous development, in which the immature spores are arranged around
the periphery of the plasmodium (Fig. 1b), and the mature spores are found in the central zone
(Azevedo et al. 2002; Eiras et al. 2004; Manrique et al. 2017).
Based on the phylogenetic comparisons of H. friederici, H. pellis and H. sutherlandi
(Griffin et al. 2008), Vidal and Luque (2017) analyzed the 18S rDNA region, the same marker
investigated in the present study. The H. friederici sequence was included in the phylogenetic
tree of Henneguya quelen n. sp., and the two species were grouped in the same clade (clade
A), although they were not closely related, and H. friederici was assigned to the same
subclade as Henneguya leporinicola, which also parasitizes a characiform host.
The phylogenetic analysis of Henneguya and Myxobolus indicated an affinity with the
hosts, with a strong evolutionary signal, as reported by Carriero et al, (2013), Fiala (2006),
and Moreira et al. (2014). The arrangement of most of the species was determined primarily
by the order of the host, followed by the family.
The habitat, the host, and the infection site, determining factors that are reflected in the
order, family, and genus, influence the genetic evolution of these myxozoans. In the specific
case of the family Myxobolidae, the aquatic environment (fresh- or saltwater) and the host are
the primary evolutionary factors, which accounts for the lack of clades containing only
Myxobolus or Henneguya species (Andree et al. 1999; Fiala 2006; Holzer et al. 2004 ; Li et al.
2012 ; Rocha et al. 2014a). Henneguya quelen n. sp. was assigned to a clade of Myxobolus-
Henneguya species that infect siluriforms, which was well supported, sharing a subclade with
Myxobolus marajoensis, also a parasite of R. quelen.
The phylogenetic tree clearly distinguishes Henneguya quelen n. sp. from the other
Henneguya species deposited in GenBank, and places it closest to Myxobolus marajoensis, a
parasite of the same host, R. quelen, from the same geographic region, on Marajó Island
(Abrunhosa et al. 2017). This arrangement further corroborates the effectiveness of the 18S
rDNA gene for analyses of the phylogenetic relationships among myxozoans, given that
closely related species can be extremely variable, with their characteristics being determined
primarily by the process of coevolution with their respective hosts (Carriero et al. 2013; Kent
et al., 2001).

76
One other similarity between Henneguya quelen n. sp. and M. marajoensis is the
infection site, with both species presenting tropism through the excretory system of R. quelen,
in the kidney and intestine, respectively, with co-infection by both parasites being observed in
20% of the fish specimens, in their respective sites of infection. This may account for the
assignment of these two species to the same subclade. In southern Brazil, Manrique et al.
(2017) described a co-infection de Myxobolus sp. and Henneguya sp. in P. mesopotamicus,
with a prevalence of 70.91%.
The phylogenetic tree of the myxobolid species that included Henneguya quelen n. sp.
has three clades. The two minor clades include parasites that infect characiform hosts,
whereas the other, dominant clade encompasses parasites of silurmform hosts, with the
exception of H. pellucida, which parasitizes charciforms.
As in the present study, previous analyses of Brazilian myxobolids (Capodifoglio et al.
2016; Casa et al. 2017) have also identified three clades, two that include species that
parasitize characiform hosts, and a third that includes siluriform parasites.
The present study describes Henneguya quelen n. sp., a myxobolid parasite of a
siluriform host, R. quelen, from the Brazilian Amazon region, which is also an important
aquaculture resource. This is an important contribution to the understanding of the
microparasite fauna of the siluriform species of the Neotropical region.
Bioethics and Biosecurity Committee Approval
The present study was approved by the Ethics Committee for Animal Experimentation of the Federal
Rural University of Amazonia, UFRA (CEUA approval no. 013/2014).
Acknowledgements
To the research funding agencies CAPES (Coordination of Higher Education Training) through PVE
88881.064967/2014-01, CNPq (Brazilian National Council for Scientific and Technological
Development) through CNPq UNIVERSAL/2014, N. 441645/2014-3, and a productivity fellowship to
Edilson Rodrigues Matos (CNPq 300949/2012-0), FAPESPA (Pará State Research Foundation)
research notice 006/2014, icaaf n°162/2014. We also thank SISBIO/ICMBIO-IBAMA (Brazilian
Institute of the Environment and Renewable Natural Resources), for authorizing the research through
scientific license no. 27119, and the Edilson Matos Research Laboratory (LPEM-UFPA).

77
References
Abrunhosa J, Sindeaux-Neto J, Santos AKD, Hamoy I, Matos, E (2017) Myxobolus
marajoensis sp. n. (Myxosporea: Myxobolidae), parasite of the freshwater catfish
Rhamdia quelen from the Brazilian Amazon region. Rev Bras Parasitol Vet, 26(4): 465-
471. http://doi.org/10.1590/s1984-29612017067
Andree BK, Székely C, Molnár K, Gresoviac SJ, Hedrick RP (1999) Relationships among
members of the genus Myxobolus (Myxozoa: Bilvalvidae) based on small subunit
ribosomal DNA sequences. J. Parasitol 85:68–74. http://doi.org/10.2307/3285702
Azevedo C, Casal G, Matos P, Matos E (2011) Henneguya torpedo sp. nov. (Myxozoa), a
parasite from the nervous system of the Amazonian teleost Brachyhypopomus
pinnicaudatus (Hypopomidae). Diseases of Aquatic Organisms, 93(3):235-42.
https://doi.org/10.3354/dao02292
Azevedo C, Matos E (2002) Fine Structure of the Myxosporean, Henneguya curimata n. sp.,
Parasite of the Amazonian Fish, Curimata inormata (Teleostei, Curimatidae). Journal of
Eukaryotic Microbiology, 49: 197–200. http://doi.org/10.1111/j.1550-
7408.2002.tb00522.x
Azevedo C, Rocha S, Matos P, Matos E, Oliveira E, Al-Quraishy S, Casal G (2014)
Morphology and phylogeny of Henneguya jocu n. sp. (Myxosporea, Myxobolidae),
infecting the gills of the marine fish Lutjanus jocu. Europ J Protistol 50:185–193.
http://doi.org/10.1016/j.ejop.2013.12.002.
Azevedo C, Rocha S, Matos E, Oliveira E, Matos P, Al-Quraishy S, Casal G (2016)
Ultrastructural and Phylogenetic Description of Kudoa orbicularis n. sp. (Myxosporea:
Multivalvulida): A Parasite Infecting the Muscle of the Fish Chaetobranchopsis
orbicularis (Teleostei: Cichlidae) in the Amazon Region. The Journal of Eukaryotic
Microbiology, v. 63, p. 27-36. http://doi.org/10.1111/jeu.12244
Baldisseroto B, Radunz Neto J (2004) Criação de Jundiá. Ed. UFSM, Santa Maria, 232p.
Bartholomew JL, Whipple MJ, Stevens DG, Fryer JL (1997) The Life Cycle of Ceratomyxa
shasta, a Myxosporean Parasite of Salmonids, Requires a Freshwater Polychaete as an
Alternate Host. J Parasitol. 83(5):859-68. http://doi.org/10.2307/3284281
Brasil. Ministério da Pesca e Aquicultura (2011) Boletim estatístico da pesca e aquicultura
2011. Brasília: MPA, 60p.
Capodifoglio KR, Adriano EA, Milanin T, Silva MR, Maia AA (2016) Morphological,
ultrastructural and phylogenetic analyses of Myxobolus hilarii n. sp. (Myxozoa,
Myxosporea), a renal parasite of farmed Brycon hilarii in Brazil. Parasitol Int,
65(3):184–190. https://doi.org/10.1016/j.parint.2015.12.006
Capodifoglio KRH, Adriano EA, Silva MRM, Maia AAM (2015) Supplementary data of
Henneguya leporinicola (Myxozoa, Myxosporea) a parasite of Leporinus
macrocephalus from fish farms in the state of São Paulo, Brazil. Acta Parasitol, 60(3):
451-458. PMid:26204182. http://doi.org/10.1515/ap-2015-0062
Carriero MM, Adriano EA, Silva MR, Ceccarelli PS, Maia AA (2013) Molecular phylogeny
of the Myxobolus and Henneguya genera with several new South American species.
PLoS One, 8(9): e73713. PMid:24040037. http://doi.org/10.1371/journal.pone.0073713.
Casal G, São Clemente SC, Lopes L, Rocha S, Felizardo N, Oliveira E, Al-Quraishy S,
Azevedo C (2017) Ultrastructural morphology and phylogeny of Henneguya gilbert n. sp.
(Myxozoa) infecting the teleostean Cyphocharax gilbert (Characiformes: Curimatidae)
from Brazil Parasitol Res, 116: 2747. https://doi.org/10.1007/s00436-017-5585-9

78
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: more models, new
heuristics and parallel computing. Nat Methods, 9(8): 772. PMid:22847109.
http://doi.org/10.1038/nmeth.2109
Eiras JC, Adriano EA (2012) A checklist of new species of Henneguya Thélohan, 1892
(Myxozoa: Myxosporea, Myxobolidae) described between 2002 and 2012. Syst
Parasitol, 83: 95. https://doi.org/10.1007/s11230-012-9374-7
Eiras JC, Malta JC, Varela A, Pavanelli GC (2004) Henneguya schizodon n. sp. (Myxozoa,
Myxobolidae), a parasite of the Amazonian teleost fish Schizodon fasciatus
(Characiformes, Anostomidae). Parasite. 2004, 11(2):169-73.
https://doi.org/10.1051/parasite/2004112169
Eszterbauer E, Marton S, Rácz OZ, Letenyei M, Molnár K (2006) Morphological and genetic
differences among actinosporean stages of fish-parasitic myxosporeans (Myxozoa):
difficulties of species identification. Syst Parasitol, 65(2): 97-114. PMid:16676228.
http://doi.org/10.1007/s11230-006-9041-y
Feist SW, Longshaw M (2006) Phylum myxozoa. In PTK Woo (ed.), Fish diseases and
disorders. Protozoan and metazoan infections. Vol. 1, 2nd ed., CAB International,
Oxfordshire, p. 230-296.
Fiala I (2006) The phylogeny of Myxosporea (Myxozoa) based on small subunit ribosomal
RNA gene analysis. Int J Parasitol 36:1521–1534.
https://doi.org/10.1016/j.ijpara.2006.06.016
Griffin MJ, Khoo LH, Torrans L, Bosworth BG, Quiniou SM, Gaunt PS, Pote LM (2009a)
New data on Henneguya pellis (Myxozoa: Myxobolidae), a parasite of blue catfish
Ictalurus furcatus. J Parasitol, 95(6): 1455-1467. PMid:19575542.
http://doi.org/10.1645/GE-2106.1
Griffin MJ, Pote LM, Wise DJ, Greenway TE, Mauel MJ, Camus AC (2008) A
Novel Henneguya Species from Channel Catfish Described by Morphological,
Histological, and Molecular Characterization. Journal of Aquatic Animal Health, 20:
127–135. https://doi.org/doi:10.1577/H07-001.1
Griffin MJ, Wise DJ, Pote LM (2009) Morphology and small-subunit ribosomal DNA
sequence of Henneguya adiposa (Myxosporea) from Ictalurus punctatus (Siluriformes). J
Parasitol, 95(5):1076-85. https://doi.org/10.1645/GE-2012.1
Hall T (2007) BioEdit: biological sequence alignment editor for Win95/98/NT/2K/ XP.
Carlsbad, CA: Ibis Biosciences.
Hallett SL, Diamant A (2001) Ultrastructure and small-subunit ribosomal DNA sequence
of Henneguya lesteri n. sp. (Myxosporea), a parasite of sand whiting Sillago analis
(Sillaginidae) from the coast of Queensland, Australia. Dis Aquat Org 46:197–212.
https://doi.org/10.3354/dao046197
Holzer AS, Sommerville C, Wootten R (2004) Molecular relationships and phylogeny in a
community of myxosporeans and actinosporeans based on their 18S rDNA sequences. Int
J Parasitol 34:1099–1111. https://doi.org/10.1016/j.ijpara.2004.06.002
Holzer AS, Sommerville C, Wootten R (2006) Molecular studies on the seasonal occurrence
and development of five myxozoans in farmed Salmo trutta L. Parasitology 132, 193–
205. https://doi.org/10.1017/S0031182005008917
Iwanowicz LR, Iwanowicz DD, Pote LM, Blazer VS, Schill WB (2008) Morphology and
18S rDNA of Henneguya gurlei (myxosporea) from Ameiurus nebulosus (siluriformes)
in North Carolina. J Parasitol 94(1):46–57. https://doi.org/10.1645/GE-1092.1
Kalavati C, Narasimhamurti CC (1985) Histopathological changes in the gills
of Channapunctatus BL. infected with Henneguya waltairensis. Arch Protistenk 129:
199-202. https://doi.org/10.1016/S0003-9365(85)80023-3

79
Kent ML, Andree KB, Bartholomew JL, El-Matbouli M, Desser, SS, Devlin RH, Feist SW,
Hedrick RP, Hoffmann RW, Khattra J, Hallett SL, Lester RJG, Longshaw M, Palenzeula
O, Siddall ME, Xiao C (2001) Recent Advances in Our Knowledge of the Myxozoa.
Journal of Eukaryotic Microbiology, 48: 395–413. https://doi.org/10.1111/j.1550-
7408.2001.tb00173.x
Leis EM, Rosser TG, Baumgartner WA, Griffin MJ (2017) Henneguya laseeae n. sp. from
flathead catfish (Pylodictis olivaris) in the upper Mississippi River. Parasitol Res,
116(1):81-89. https://doi.org/10.1007/s00436-016-5264-2
Li Y, Sato H, Kamata Y, Ohnishi T, Sugita-Konishi Y (2012) Three novel myxobolid species
of genera Henneguya and Myxobolus (Myxosporea: Bivalvulida) from marine fish in
Japan. Parasitol Res 111:819–826. https://doi.org/10.1007/s00436-012-2904-z
Lin D, Hanson La, Pote Lm (1999) Small Subunit Ribosomal RNA Sequence of Henneguya
exilis (Class Myxosporea) Identifies the Actinosporean Stage from an Oligochaete Host.
Journal of Eukaryotic Microbiology, 46: 66–68. https://doi.org/10.1111/j.1550-
7408.1999.tb04585.x
Liu XH, Zhang JY, Batueva MD, Voronin VN (2016) Supplemental description and
molecular characterization of Myxobolus miyarii Kudo, 1919 (Myxosporea:
Myxobolidae) infecting intestine of Amur catfish (Silurus asotus). Parasitol Res,
115(4):1547-56. https://doi.org/10.1007/s00436-015-4889-x
Lom J, Dyková L (1995) Myxosporea (Phylum Myxozoa). In PTK Woo, Fish Diseases and
Disorders - Protozoan and Metazoan Infections, Vol. 1, Cab International, p. 87-147.
Lom J, Dyková L (2006) Myxozoan genera: definition and notes on taxonomy, life-cicle
terminology and pathogenic species. Folia Parasitologica v. 53, p. 1-36.
https://doi.org/10.14411/fp.2006.001
Luna LG (1968) Manual of histologic staining methods of the Armed Forces Institute of
Pathology. 3.ed. New York: McGraw-Hill. 258p.
Manrique WG, Figueiredo MAP, de Andrade Belo MA, Martins ML, Molnár K (2017)
Myxobolus sp. and Henneguya sp. (Cnidaria: Myxobolidae) natural co-infection in the
kidney of Piaractus mesopotamicus (Characiformes: Serrasalmidae). Parasitol Res, 116:
2853. https://doi.org/10.1007/s00436-017-5571-2
Martins ML, Souza VN (1997) Henneguya piaractus n. sp. (Myxozoa: Myxobolidae), a gill
parasite of Piaractus mesopotamicus Holmberg, 1887 (Osteichthyes: Characidae), in
Brazil. Rev Bras Biol, 57: 239-245.
Mathews PD, Naldoni J, Maia AA, Adriano EA (2016) Morphology and small subunit rDNA-
based phylogeny of Ceratomyxa amazonensis n. sp. parasite of Symphysodon discus, an
ornamental freshwater fish from Amazon. Parasitol Res, 115: 4021.
https://doi.org/10.1007/s00436-016-5173-4
Matos E, Tajdari J, Azevedo C (2005) Ultrastructural studies of Henneguya rhamdia n. sp.
(Myxozoa) a parasite from the Amazon Teleost fish (Rhamdia quelen) (Pimelodidae). J.
Eukariot. Microbiol, 52 (6): 532-537. https://doi.org/10.1111/j.1550-7408.2005.00063.x
Matos PS, Silva DT, Hamoy I, Matos E (2018) Morphological features and molecular
phylogeny of Hoferellus azevedoi n. sp. (Myxozoa: Myxobilatidae) found in
Chaetobranchus flavescens Heckel, 1840 (Teleostei: Cichlidae) from Marajó Island,
northern Brazil. Parasitology Research https://doi.org/10.1007/s00436-018-5785-y
Molnár, K., Eszterbauer, E., Székely, C., Dán, Á., Harrach, B., 2002. Morphological and
molecular biological studies on intramuscular Myxobolus spp. of cyprinid fish. J. Fish
Dis. 25, 643–652. https://doi.org/10.1046/j.1365-2761.2002.00409.x
Milanin T, Eiras JC, Arana S, Maia AA, Alves AL, Silva MR, Carriero MM, Ceccarelli PS,
Adriano EA. (2010) Phylogeny, ultrastructure, histopathology and prevalence of Myxobolus

80
oliveirai sp. nov., a parasite of Brycon hilarii (Characidae) in the Pantanal wetland, Brazil.
Mem. Inst. Oswaldo Cruz [online], 105(6): 762-769. http://doi.org/10.1590/S0074-
02762010000600006.
Moreira GSA, Adriano EA, Silva MRM, Ceccarelli PS, Maia AAM (2014) The
morphological and molecular characterization of Henneguya rotunda n. sp., a parasite of
the gill arch and fins of Salminus brasiliensis from the Mogi Guaçu River, Brazil.
Parasitol Res, 113: 1703–1711. https://doi.org/10.1007/s00436-014-3815-y
Müller MI, Adriano EA, Ceccarelli PS, Silva MRM, Maia AAM, Ueta M.T (2013)
Prevalence, intensity, and phylogenetic analysis of Henneguya piaractus and Myxobolus
cf. colossomatis from farmed Piaractus mesopotamicus in Brazil. Diseases of Aquatic
Organisms. Vol. 107: 129–139. https://doi.org/10.3354/dao02668
Naldoni J, Arana S, Maia AA, Silva MR, Carriero MM, Ceccarelli OS, Tavares, LER,
Adriano E.A. (2011) Host-parasite-environment relationship, morphology and molecular
analyses of Henneguya eirasi n. sp. parasite of two wild Pseudoplatystoma spp. in
Pantanal Wetland, Brazil. Vet Parasitol, 177: 247–255.
https://doi.org/10.1016/j.vetpar.2010.12.008
Naldoni J, Zatti SA, Capodifoglio, KRH, Milanin T, Maia AAM, Silva MRM, Adriano EA
(2015) Host-parasite and phylogenetic relationships of Myxobolus filamentum sp. n.
(Myxozoa: Myxosporea), a parasite of Brycon orthotaenia (Characiformes: Bryconidae)
in Brazil. Folia Parasitol, 1;62. http://doi.org/10.14411/fp.2015.014
Pote LM, Hanson LA, Shivaji R (2000) Small Subunit Ribosomal RNA Sequences Link the
Cause of Proliferative Gill Disease in Channel Catfish to Henneguya n. sp. (Myxozoa:
Myxosporea). Journal of Aquatic Animal Health, 12:3, 230-240.
https://doi.org/10.1577/1548-8667(2000)012<0230:SSRRSL>2.0.CO;2
Rambaut A, Suchard M, Drummond A (2008) Tracer v1.4.1 [online]. 2008 [cited 2017
August 25]. Available from: http://tree.bio.ed.ac.uk/software/tracer.
Rocha S, Casal G, Garcia P, Matos E, Al-Quraishy S, Azevedo C (2014a) Ultrastructure and
phylogeny of the parasite Henneguya carolina sp. nov. (Myxozoa), from the marine fish
Trachinotus carolinus in Brazil. Dis Aquat Org 112:139–148.
https://doi.org/10.3354/dao02794
Rocha S, Casal G, Velasco M, Alves A, Matos E, Al-Quraishy S, Azevedo C (2014b)
Morphology and phylogeny of Thelohanellus marginatus n. sp. (Myxozoa: Myxosporea),
a parasite infecting the gills of the fish Hypophthalmus marginatus (Teleostei:
Pimelodidae) in the Amazon River. J Eukaryot Microbiol 61:586–593.
https://doi.org/10.1111/jeu.12136
Ronquist F, Huelsenbeck JP (2003) MrBayes version 3.0: Bayesian phylogenetic inference
under mixed models. Bioinformatics, 19 (12):1572-1574, 2003.
https://doi.org/10.1093/bioinformatics/btg180
Rosser TG, Baumgartner WA, Barger MA, Griffin MJ (2017) Myxobolus lepomis n. sp.
(Cnidaria: Myxobolidae), a gill myxozoan infecting Lepomis marginatus Holbrook and
Lepomis miniatus Jordan (Perciformes: Centrarchidae), in the Big Thicket National
Preserve, Texas, USA. Syst Parasitol, 94(4):535-545. https://doi.org/10.1007/s11230-
017-9719-3
Rosser TG, Griffin MJ, Quiniou SM, Greenway TE, Khoo LH, Wise DJ, Pote LM (2014a)
Molecular and morphological characterization of myxozoan actinospore types from a
commercial catfish pond in the Mississippi delta. J Parasitol, 100(6):828-39.
https://doi.org/10.1645/13-446.1
Rosser TG, Griffin MJ, Quiniou SMA, Khoo LH, Greenway TE, Wise DJ, Pote LM (2015)
Small subunit ribosomal RNA sequence links the myxospore stage of Henneguya

81
mississippiensis n. sp. from channel catfish Ictalurus punctatus to an actinospore released
by the benthic oligochaete Dero digitata. Parasitol Res, 114(4): 1595-1602.
PMid:25716821. http://doi.org/10.1007/s00436-015-4345-y
Rosser TG, Griffin MJ, Quiniou SMA, Khoo LH, Pote, LM (2014b) 18S rRNA gene
sequencing identifies a novel species of Henneguya parasitizing the gills of the channel
catfish (Ictaluridae). Parasitol Res, 113:4651. https://doi.org/10.1007/s00436-014-4156-6
Swofford DL (1998) PAUP* Phylogenetic Analysis Using Parsimony (*and other methods).
Version 4. Sunderland: Sinauer Associates. 128 p.
Velasco M, Videira M ; Sindeaux-Neto JL, Santos P, Sanches, O, Matos P, Matos E (2015).
Infection by Henneguya sp. (Myxozoa) in the bone tissue of the gill filaments of the
Amazonian catfish Hypophthalmus marginatus (Siluriformes) Braz. J. Vet. Parasitol,
Jaboticabal. http://doi.org/10.1590/S1984-29612015021
Velasco M, Videira M, Nascimento LC, Matos P, Gonçalves EC, Matos E (2016) Henneguya
paraensis n. sp. (Myxozoa;Myxosporea), a new gill parasite of the Amazonian fish
Cichla temensis (Teleostei: Cichlidae): morphological and molecular aspects. Parasitol
Res 115:1779–1787. http://doi.org/doi:10.1007/s00436-016-4916-6
Vidal LPE, Luque JL (2017) New morphological data and molecular diagnostic of
Henneguya friderici (Myxozoa: Myxobolidae), a parasite of Leporinus friderici
(Osteichthyes: Anostomidae) from southeastern Brazil. Rev. Bras. Parasitol. Vet.[online],
vol.26, n.1, pp.81-88. Epub 16-Mar-2017. ISSN 0103-846X.
http://doi.org/10.1590/s1984-29612017010

82
4. CONCLUSÃO
O hospedeiro R. quelen apresentou suscetibilidade à ação de microparasitos em diferentes
órgãos: no intestino, no músculo esquelético e no rim infectados por Myxobolus marajoensis,
M. arariensis e Henneguya quelen respectivamente, sendo descritas neste trabalho três novas
espécies, através de análises morfológicas e moleculares.
As análises histológicas demonstraram inflamações nos órgãos afetados pelas três
espécies descritas, demonstrando doenças parasitárias, que pode afetar a saúde do hospedeiro.
Estudos sobre a reação destas parasitoses no organismo humano devem ser investigados,
visto que pode afetar a saúde das populações ribeirinhas que o consomem na região
Amazônica, e também poderá causar prejuízos na aquicultura, pois se trata de uma espécie
comercial com pacote tecnológico consolidado em outras regiões no Brasil e de ampla
distribuição na América do Sul.

83
REFERÊNCIAS
ABDEL-GHAFFAR, F., ALI, M.A., AL QURAISHY, S., AL RASHEID, K., AL FARRAJ,
S., ABDEL-BAKI, A.S., BASHTAR, A.R. Four new species of Ceratomyxa Thelohan
1892 (Myxozoa: Myxosporea: Ceratomyxidae) infecting the gallbladder of some Red
Sea fishes. Parasitol. Res. 103:559-565, 2008.
ABRUNHOSA J, SINDEAUX-NETO J, SANTOS AKD, HAMOY I, MATOS, E.
Myxobolus marajoensis sp. n. (Myxosporea: Myxobolidae), parasite of the freshwater
catfish Rhamdia quelen from the Brazilian Amazon region. Rev. Bras. Parasitol. Vet.
26 (4): 465-471, 2017.
AZEVEDO, C.; SÃO CLEMENTE, S. C.; CASAL, G.; MATOS, P.; ALVES, Â.; AL-
QURAISHY, S.; MATOS, E. Myxobolus myleus n. sp. infecting the bile of the
Amazonian freshwater fish Myleus rubripinnis (Teleostei: Serrasalmidae): morphology
and pathology. Syst. Parasitol. 82(3): 241-247. 2012.
AZEVEDO, C., CASAL, G., MATOS, P., FERREIRA, I., MATOS, E. Light and electron
microscopy of the spore of Myxobolus heckelii n. sp. (Myxozoa), parasite from the
Brazilian fish Centromochlus heckelii (Teleostei, Auchenipteridae). J. Eukaryot.
Microbiol. 56: 589–593. 2009.
AZEVEDO, C., VIDEIRA, M., CASAL, G., MATOS, P., OLIVEIRA, E., AL-QURAISHY,
S., MATOS, E. Fine structure of the Plasmodia and Myxospore of Ellipsomyxa
gobioides n. sp. (Myxozoa) found in the gallbladder of Gobioides broussonnetii
(Teleostei: Gobiidae) from the lower Amazon river. J. Eukaryot. Microbiol. 60
(5):490-6. 2013.
AZEVEDO, R.K., VIEIRA, D.H.M.D., VIEIRA, G.H., SILVA, R.J., MATOS, E.,
ABDALLAH, V.D. Phylogeny, ultrastructure and histopathology of Myxobolus lomi sp.
nov., a parasite of Prochilodus lineatus (Valenciennes, 1836) (Characiformes:
Prochilodontidae) from the Peixes River, São Paulo State, Brazil. Parasitol. Int. 63:
303–307. 2014.
AZEVEDO, R.K., ABDALLAH, V.D., PAES, J.V.K., SILVA, R.J, MATOS, P., VELASCO,
M., MATOS, E. Henneguya nagelii n. sp. (Myxozoa: Myxobolidae) in Cyphocharax
nagelii (Steindachner, 1881) (Teleostei: Characiformes: Curimatidae) from the Peixe’s
River, São Paulo State, Brazil. Parasitol. Res. 112: 3601-3605, 2013.
AZEVEDO, C., ROCHA, S., MATOS, E., OLIVEIRA, E., MATOS, P., AL-QURAISHY, S.,
CASAL, G. Ultrastructural and Phylogenetic Description of Kudoa orbicularis n. sp.
(Myxosporea: Multivalvulida): A Parasite Infecting the Muscle of the Fish
Chaetobranchopsis orbicularis (Teleostei: Cichlidae) in the Amazon Region. J.
Eukaryot. Microbiol. 63: 27–36, 2016.
BALDISSEROTTO, B., RADUNZ NETO, J. Biologia do jundiá. In: Criacao de Jundia.
Santa Maria: Editora UFSM, 2004. P. 67-72.

84
BÉKÉSI, L., SZÉKELY, C., MOLNÁR, K. Atuais conhecimentos sobre Myxosporea
(Myxozoa), parasitas de peixes. Um estágio alternativo dos parasitas no Brasil. Braz. J.
Vet. Res. An. Sci. 39 (5): 271-276, 2002.
BORGHETTI, N. R., OSTRENSKY, A., BORGHETTI, J. R. Aqüicultura: uma visão geral
sobre a produção de organismos aquáticos no Brasil e no mundo. Curitiba: Grupo
Integrado de Aqüicultura e Estudos Ambientais, 128 p. 2003.
CARRIERO, M.M., ADRIANO, E.A., SILVA, M.R., CECCARELLI, P.S., MAIA, A.A.
Molecular phylogeny of the Myxobolus and Henneguya genera with several new South
American species. PLoS One 8 (9): e73713, 2013.
CASAL, G., MATOS, E., AZEVEDO, C. Ultrastructural data on the spore of Myxobolus
maculatus n. sp. (Phylum Myxozoa), parasite from the Amazonian fish Metynnis
maculatus (Teleostei). Dis. Aquatic. Organ. 51: 107-112, 2002.
CASAL, G., MATOS, E., AZEVEDO, C. A new myxozoan parasite from the amazonian fish
Metynnis argenteus (Teleostei, Characidae): light and electron microscope observations.
J. Parasitol. 92 (4): 817–821, 2006.
CASAL, G. Microsporidioses e Mixosporidioses da ictiofauna portuguesa e brasileira:
Caracterização ultraestrutural e filogenética. Tese (Doutorado em Ciências
Biomédicas). Porto-Portugal. Instituto de Ciências Biomédicas Abel Salazar –
Universidade do Porto, 2009. 249 p.
CASAL, G., MATOS, E., TELES-GRILO, M. L., AZEVEDO, C. Morphological and
genetical description of Loma psittaca sp. n. isolated from the Amazonian fish species
Colomesus psittacus. Parasitol. Res. 105 (5):1261–1271, 2009.
CASAL, G., MATOS, E., TELES-GRILO, M.L., AZEVEDO, C. Ultrastructural and
Molecular Characterization of a New Microsporidium Parasite from the Amazonian
Fish, Gymnorhamphichthys rondoni (Rhamphichthyidae). J. Parasitol. 96: 1155-1163,
2010.
CASAL, G., CLEMENTE, S.C.S., MATOS, P., KNOFF, M., MATOS, E., ABDEL, A.,
ABDEL-BAKI, AZEVEDO, C. Redefining the genus Spraguea based on ultrastructural
and phylogenetic data from Spraguea gastrophysus n. sp. (Phylum Microsporidia), a
parasite found in Lophius gastrophysus (Teleostei) from Brazil. Parasitol. Res. 111:
79-88, 2012.
CASAL, G., SÃO CLEMENTE, S.C., LOPES, L., ROCHA, S., FELIZARDO, N.,
OLIVEIRA, E., AL-QURAISHY, S., AZEVEDO, C. Ultrastructural morphology and
phylogeny of Henneguya gilbert n. sp. (Myxozoa) infecting the teleostean Cyphocharax
gilbert (Characiformes: Curimatidae) from Brazil. Parasitol. Res. 116: 27-47, 2017.
CELLERE, E.F., CORDEIRO, N.S., ADRIANO, E.A. Myxobolus absonus sp. n. (Myxozoa:
Myxosporea) parasitizing Pimelodus maculatus (Siluriformes: Pimelodidae), a South
American freshwater fish. Mem. Inst. Oswaldo Cruz 97: 79-80, 2002.

85
CHANG, E.S., NEUHOF, M., RUBINSTEIN, N.D., DIAMANT, A., PHILIPPE, H.,
HUCHON, D., CARTWRIGHT, P. Genomic insights into the evolutionary origin of
Myxozoa within Cnidaria. Proc. Nati. Acad. Sci. USA. 112 (48): 14912-14917, 2015.
DYKOVÁ, I., BURON, I., FIALA, I., ROUMILLAT, W.A. Kudoa inornata sp. n.
(Myxosporea: Multivalvulida) from the skeletal muscles of Cynoscion nebulosus
(Teleostei: Sciaenidae). Folia Parasitol. 56 (2): 91–98, 2009.
EIRAS, J.C., MOLNÁR, L., LU, Y.S. Synopsis of the species of Myxobolus Butschli, 1882
(Myxozoa: Myxosporea: Myxobolidae). Syst. Parasitol. 61: 1-46, 2005.
EIRAS, J.C., ADRIANO, E.A. Myxozoa In: Parasitologia de peixes de água doce do
Brasil. PAVANELLI, G. C., TAKEMOTO, R. M., EIRAS, J. C. (Org.) Maringá,
Eduem, 2013. P. 249-272.
EIRAS, J. C., ZHANG, J.; MOLNÁR, K. Synopsis of the species of Myxobolus Bütschli,
1882 (Myxozoa: Myxosporea, Myxobolidae) described between 2005 and 2013. Syst.
Parasitol. 88:11–36, 2014.
EL-MATBOULI, M., FISCHER-SCHERL, T.; HOFFMANN, R.W. Present knowledge on
the life cycle, taxonomy, pathology, and therapy of some Myxosporea spp. important
for freshwater fish. Ann. Rev. Fish Dis. 3: 367-402, 1992.
FENG, J.M., XIONG, J., ZHANG, J.Y., YANG, Y.L., YAO, B., ZHOU, Z.G., MIAO, W.
New phylogenomic and comparative analyses provide corroborating evidence that
Myxozoa is Cnidaria. Mol. Phylogenet. Evol. 81 (2014) 10–18, 2014.
FIALA, I., BARTOŠOVÁ, P. History of myxozoan character evolution on the basis of rDNA
and EF-2 data. BMC Evol. Biol. 10:228, 2010.
FIALA, I., BARTOŠOVÁ-SOJKOVÁ, P., WHIPPS, C.M. Classification and Phylogenetics
of Myxozoa. In: Myxozoan Evolution, Ecology and Development. Springer
International Publishing. Switzerland, 2015. 441p.
FOWLER, H.W. Os peixes de água doce do Brasil. In: Arquivos de Zoologia do Estado de
São Paulo. São Paulo, 1951. P. 6: 405-625.
FREEMAN, M,A., YOKOYAMA, H., OGAWA, K. Description and phylogeny of
Ceratomyxa anko sp n. and Zschokkella lophii sp n. from the Japanese anglerfish,
Lophius litulon (Jordan). J. Fish Dis. 31 (12):921-30, 2008.
GOMES, L.C, GOLOMBIESKI, J. I., GOMES, A.R.C., BALDISSEROTTO, B. Biologia do
jundiá Rhamdia quelen (Teleostei, Pimelodidae). Ciênc. Rural 30 (1): 179-185, 2000.
GOULDING, M. The Fishes and the Forest. Exploration in Amazonian Natural History.
University of California Press. Berkeley, USA. 1980. 280p.
GOULDING, M. Amazon: the flooded forest: BBC Books, London. 1989. 208 p.
GUEDES, D.S. Contribuição ao estudo da sistemática e alimentação de jundiás

86
(Rhamdia spp) na região central do Rio Grande do Sul (Pisces, Pimelodidae). Santa
Maria – RS, 1980. 99p. Dissertação (Mestrado em Zootecnia) - Curso de Pós-graduação
em Zootecnia, Universidade Federal de Santa Maria, 1980.
HOLZER, A.S., SOMMERVILLE, C., WOOTTEN, R. Molecular studies on the seasonal
occurrence and development of five myxozoans in farmed Salmo trutta L. Parasitology
132: 193–205, 2006.
YOKOYAMA, H., GRABNER, D., SHIRAKASHI, S. Transmission Biology of the
Myxozoa. In: Carvalho E (Ed.) Health and Environment in Aquaculture. InTech,
Croatia. P. 3-42.
KUBITZA, F., KUBITZA, L.M.M. Principais parasitoses e doenças dos peixes cultivados.
4. ed. Jundiaí: F. Kubitza, 2004. 118p.
JIMÉNEZ-GURI, E., PHILIPPE, H., OKAMURA, B., HOLLAND, P.W.H. Buddenbrockia is
a cnidarian worm. Science 317:116–118, 2007.
LAFFERTY, KD. Environmental parasitology: what can parasites tell us about human impact
on the environment? Parasitol. Today 13: 251-255, 1997.
LOM, J. Notes on the ultrastructure and sporoblast development in fish paraziting
myxosporidian of the genus Sphaeromyxa. Z. Zellforsch. 97: 416-437, 1969.
LOM, J., PUYTORAC, P. Studies on the myxosporidian ultrastructure and polar capsule
development. Protistologica 1: 53-65, 1965.
LOM, J., ARTHUR, J.R. A guideline for the preparation of species descriptions in
Myxosporea. J. Fish Dis. 12 (2): 151–156, 1989.
LOM, J., DYKOVÁ, I. Protozoan Parasites of Fishes. Amsterdam, Elsevier, 1992. 315p.
LOM, J., DYKOVÁ, I. Myxozoan genera: definition and notes on taxonomy, life-cycle
terminology and pathogenic species. Fol. Parasitol. 43: 1-36, 2006.
LONGSHAW, M., FREAR, P., FEIST, S.W. Myxobolus buckei sp. m. (Myxozoa), a new
pathogenic parasite from the spinal column of three cyprinid fishes from the United
Kingdom. Fol. Parasitol. 50: 251-262, 2003.
LOWE-McCONNELL, R.H. Estudos ecológicos de comunidades de peixes tropicais:
EDUSP, São Paulo. 1999. 536 p.
MATOS, E., AZEVEDO, C., CORRAL, L., CASAL, G., MATOS, P. Ultraestrutura de
protozoários parasitas de peixes da região amazônica. Acta Amaz. 29 (4): 575-585,
1999.
MATOS, E., CORRAL, L., MATOS, P., CASAL, G. & AZEVEDO, C. Incidência de
parasitas do Phylum Myxozoa (Sub-reino Protozoa) em peixes da região amazônica,
com especial destaque para o gênero Henneguya. Revista de Ciências Agrárias 36:
83 - 99, 2001.

87
MATOS, E., CASAL, G., MATOS, P., CORRAL, L. & AZEVEDO, C. Microrganismos
Parasitos de Animais Aquáticos da Amazônia. In: Sanidade de organismos aquáticos.
Ranzani-Paiva, M.J.T., Takemoto, R.M., Lizama, M.A.P. Editora Varela, São Paulo,
Brasil, 2004. P. 158-178.
MATOS, E., TAJDARI, J. & AZEVEDO, C. Ultrastructural studies of Henneguya rhamdia n.
sp. (Myxozoa) a parasite from the Amazon Teleost fish (Rhamdia quelen)
(Pimelodidae). J. Eukariot. Microbiol. 52 (6): 532-537, 2005.
MATOS, E., MENDONÇA, I., AZEVEDO, C. Vavraia lutzomyiae n. sp. (Phylum
Microspora) infecting the sandfly Lutzomyia longipalpis (Psychodidae, Phlebotominae),
a vector of human visceral leishmaniasis. Eur. J. Protistol. 42: 21–28, 2006.
MATOS, P.S., SILVA, D.T., HAMOY, I., MATOS, E. Morphological features and molecular
phylogeny of Hoferellus azevedoi n. sp. (Myxozoa: Myxobilatidae) found in
Chaetobranchus flavescens Heckel, 1840 (Teleostei: Cichlidae) from Marajó Island,
northern Brazil. Parasitol. Research. 117: 1087, 2018.
MPA-MINISTÉRIO DA PESCA E AQUICULTURA. Boletim estatístico da pesca e
aquicultura 2011. Brasília: MPA. 2011. 60 p.
NESNIDAL, M.P., HELMKAMPF, M., BRUCHHAUS, I., EL-MATBOULI, M.,
HAUSFORF, B. Agent of whirling disease meetsorphan worm: phylogenomic analyses
firmly place Myxozoa in Cnidaria. PLoS ONE 8: (1):e54576. 2013.
OKAMURA, B., GRUHL, A., BARTHOLOMEW, J. Myxozoan Evolution, Ecology and
Development. Springer Eds. 2015. 435p.
PAVANELLI, G.C., TAKEMOTO, R.M., EIRAS, J.C. (Org.) Parasitologia de peixes de
água doce do Brasil. Maringá, Eduem, 2013. 452p.
PIAIA, R., BALDISSEROTO, B. Densidade de estocagem e crescimento de alevinos de
jundiá Rhamdia quelen (Quoy & Gaimard, 1824). Ciência Rural 30 (3): 509-513,
2000.
PINTO, C. Myxobolus noguchii, M. stokesi e Henneguya ihering, espécies novas de
myxosporídeos de peixes de água doce do Brasil. Boletim Biológico 12: 41-43, 1928.
REIS, R.E., KULLANDER, S.O., FERRARIS, J.R., C. J. Check list of the Freshwater Fishes
of South and Central America. Edipucrs, Porto Alegre. 2003. 729p.
POTE, L.M.W., KHOO, L., GRIFFIN, M. Henneguya ictaluri. In: Fish Parasites:
pathobiology and protection. edited by Patrick T.K. Woo, Kurt Buchmann, Cambridge,
EUA. p. 177-192, 2012.
ROCHA, S., CASAL, G., MATOS, P., MATOS, E., DKHIL, M.; AZEVEDO, C. 2011.
Description of Triangulamyxa psittaca sp. nov. (Myxozoa: Myxosporea), a new parasite
in the urinary bladder of Colomesus psittacus (Teleostei) from the Amazon River, with
emphasis on the ultrastructure of plasmodial stages. Acta Protozool. 50:327–338.

88
ROCHA, S., CASAL, G., VELASCO, M., ALVES, A., MATOS, E., AL-QURAISHY, S.,
AZEVEDO, C. Morphology and phylogeny of Thelohanellus marginatus n. sp.
(Myxozoa: Myxosporea), a parasite infecting the gills of the fish Hypophthalmus
marginatus (Teleostei: Pimelodidae) in the Amazon River. J. Eukaryot. Microbiol.
61:586–593, 2014.
RUIZ, C.F., RASH, J.M., ARIAS, C.R., BESLER, D.A. 2017. Morphological and molecular
confirmation of Myxobolus cerebralis myxospores infecting wild-caught and cultured
trout in North Carolina (SE USA). Dis. Aquat. Org. 126:185-198.
SCHAPERCLAUS, W. Diseases caused by ciliates. In: Fish diseases. Schaperclaus, E.,
Kulow, H. (eds) Published for the U.S. Department of the Interior and the National
Science Foundation, Washington, DC. Amerind, New Delhi, 1991. P. 702-725.
SCARANTO, B. M. S. Caracterização genética do jundiá (Rhamdia quelen) por meio do
DNA barcode e marcadores microssatélites. Dissertação de mestrado, Florianópolis,
Universidade Federal de Santa Catarina, 2017. 88p.
SILVERGRIP, A.M.C. A sistematic revision of the neotropical catfish genus Rhamdia
(Teleostei, Pimedodidae). Stockholm, Sweden, (PhD Thesis) - Department of Zoology,
Stockholm University and Department of Vertebrate Zoology, Swedish Museu of
Natural History, 1996. 156p.
SMOTHERS, J.F., VON DOHLEN, C.D., SMITH JR., L.H., SPALL, R.D., Molecular
evidence that the myxozoan protists are metazoans. Science 265 (5179): 1719–1721,
1994.
SIDDALL, M.E., MARTIN, D.S., BRIDGE, D., DESSER, S.S., CONE. D.K. The demise of
a phylum of protists: Phylogeny of Myxozoa and other parasitic cnidaria. J. Parasitol.
81 (6): 961–967, 1995.
ŠTOLC, A. Actinomyxidies, nouveau groupe de Mésozoaires parent des Myxosporidies. Bul.
Int. Acad. Sci. Bohème 22:1–12, 1899.
TAJDARI, J., MATOS, E., MENDONCA, I., AZEVEDO, C. Ultrastrutural Morphology of
Myxobolus testicularis sp. n., Parasite of the Testis of Hemiodopsis microlepis
(Teleostei: Hemiodontidae) from the NE of Brazil. Acta Protozool. 44:377– 384, 2005.
VELASCO, M.; VIDEIRA, M.; NASCIMENTO, L.C.S.; MATOS, P.; GONÇALVES, E.C.;
MATOS, E. Henneguya paraensis n. sp. (Myxozoa; Myxosporea), a new gill parasite of
the Amazonian fish Cichla temensis (Teleostei: Cichlidae): morphological and
molecular aspects. Parasitol. Res. 115: 1779-1787, 2016.
ZHANG, Z.Q. Animal biodiversity: an introduction to higher-level classification and
taxonomic richness. Zootaxa 3148:7–12, 2011.

89
ANEXO:
Email de Submissão :Revista Zootaxa referente ao Artigo III
Edilson Matos <[email protected]>
27 de mar
para sina.adl
Dear editor, We attach the manuscript entitled: " A new species of myxozoa in the skeletal striated musculature of Rhamdia quelen (Quoy & Gaimard) (Siluriforme: Pimelodidae) Amazonian fish, Marajó island, Brazil" for consideration and possible publication in the Zootaxa. This study describe as new species
of myxozoan, Myxobolus arariensis n. sp. based on the results of morphological and molecula
r analyses. These parasites were found in a Brazilian freshwater catfish captured in the Arari
River,eastern Amazon region. This manuscript is an original work and is not under considerat
ion by any other journal. All authors approved the manuscript and this submission. This article doesn´t violate any copyright or other proprietary right of any person or entity and
does not contain abusive,defamatory, obscene, or fraudulent statements or any other unlawful
statements in any way.
Sincerely, Prof. Dr. Edilson Matos Universidade Federal Rural da Amazônia (UFRA) Carlos Azevedo Research Laboratory Avenida Presidente Tancredo Neves, n◦ 2501- Montese Cep: 66.077-901, Belém, Pará - Brazil - Telephone: +55 (91) 3210-5207
Adl, Sina <[email protected]>
5 de abr
5 de abr
para mim Dear Dr Matos, Your manuscript has been sent out for review. Regards, Sina Sina Adl Professor Department of Soil Sciences College of Agriculture and Bioresources University of Saskatchewan (306) 966-6866 agbio.usask.ca Editor-in-Chief, Rhizosphere http://www.journals.elsevier.com/rhizosphere/

90
From: Edilson Matos <[email protected]> Sent: Tuesday, March 27, 2018 8:20 AM Normas da revista Zootaxa
Preparation of manuscripts
1) General. All papers must be in English. Authors whose native language is not English are
encouraged to have their manuscripts read by a native English-speaking colleague before
submission. Nomenclature must be in agreement with the International Code of Zoological
Nomenclature (4th edition 1999), which came into force on 1 January 2000. Author(s) of
species name must be provided when the scientific name of any animal species is first
mentioned (the year of publication needs not be given; if you give it, then provide a full
reference of this in the reference list). Authors of plant species names need not be given.
Metric systems should be used. If possible, use the common font Times New Roman and use
as little formatting as possible (use only bold and italics where necessary and indentions of
paragraphs except the first). Special symbols (e.g. male or female sign) should be avoided
because they are likely to be altered when files are read on different machines (Mac versus PC
with different language systems). You can code them as m# and f#, which can be replaced
during page setting. The style of each author is generally respected but they must follow the
following general guidelines.
2) The title should be concise and informative. The higher taxa containing the taxa dealt with
in the paper should be indicated in parentheses: e.g. A taxonomic revision of the
genus Aus (Order: family).
3) The name(s) of all authors of the paper must be given and should be typed in the upper
case (e.g. ADAM SMITH, BRIAN SMITH & CAROL SMITH). The address of each author
should be given in italics each starting a separate line. E-mail address(es) should be provided
if available.
4) The abstract should be concise and informative. Any new names or new combinations
proposed in the paper should be mentioned. Abstracts in other languages may also be
included in addition to English abstract. The abstract should be followed by a list ofkey
wordsthat are not present in the title. Abstract and key words are not needed in short
correspondence.
5) The arrangement of the main text varies with different types of papers (a taxonomic
revision, an analysis of characters and phylogeny, a catalogue etc.), but should usually start
with an introduction and end with a list of references. References should be cited in the text

91
as Smith (1999), Smith & Smith (2000) or Smith et al. (2001) (3 or more authors), or
alternatively in a parenthesis (Smith 1999; Smith & Smith 2000; Smith et al. 2001). All
literature cited in the text must be listed in the references in the following format (see
a sample page here in PDF).
A) Journal paper: Smith, A. (1999) Title of the paper. Title of the journal in full, volume
number, page range.
B) Book chapter: Smith, A. & Smith, B. (2000) Title of the Chapter. In: Smith, A, Smith, B.
& Smith, C. (Eds), Title of Book. Publisher name and location, pp. x–y.
C) Book: Smith, A., Smith, B. & Smith, C. (2001) Title of Book. Publisher name and location,
xyz pp.
D) Internet resources Author (2002) Title of website, database or other resources, Publisher
name and location (if indicated), number of pages (if known). Available from:
http://xxx.xxx.xxx/ (Date of access).
Dissertations resulting from graduate studies and non-serial proceedings of
conferences/symposia are to be treated as books and cited as such. Papers not cited must not
be listed in the references.
Please note that:
(1) journal titles must be written in full (not abbreviated)
(2) journal titles and volume numbers are followed by a ","
(3) page ranges are connected by "n dash", not hyphen "-", which is used to connect two
words.
For websites, it is important to include the last date when you see that site, as it can be moved
or deleted from that address in the future.
On the use of dashes: (1) Hyphens are used to link words such as personal names, some
prefixes and compound adjectives (the last of which vary depending on the style manual in
use). (2) En-dash or en-rule (the length of an ‘n’) is used to link spans. In the context of our
journal that means numerals mainly, most frequently sizes, dates and page numbers (e.g.
1977–1981; figs 5–7) and also geographic or name associations (Murray–Darling River; a
Federal–State agreement). (3) Em-dash or em-rule (the length of an ‘m’) are used far more
infrequently, and are used for breaks in the text or subject, often used much as we used
parentheses. In contrast to parentheses an em-dash can be used alone; e.g. What could these

92
results mean—that Niel had discovered the meaning of life? En-dashes and em-dashes should
not be spaced.
6) Legends of illustrations should be listed after the list of references. Small illustrations
should be grouped into plates. When preparing illustrations, authors should bear in mind that
the journal has a matter size of 25 cm by 17 cm and is printed on A4 paper. For species
illustration, line drawings are preferred, although good quality B&W or colour photographs
are also acceptable. See a guide here for detailed information on preparing plates for
publication.
7) Tables, if any, should be given at the end of the manuscript. Please use the table function
in your word processor to build tables so that the cells, rows and columns can remain aligned
when font size and width of the table are changed. Please do not use Tab key or space bar to
type tables.
8) Keys are not easy to typeset. In a typical dichotomous key, each lead of a couplet should be
typed simply as a paragraph as in the box below:
1 Seven setae present on tarsus I ; four setae present on tibia I; leg I longer than the body; legs
black in color ... Genus A - Six setae present on tarsus I; three setae present on tibia I; leg I
shorter than the body; legs brown in color ... 22 Leg II longer than leg I ... Genus B
- Leg II shorter than leg I ... Genus C
Our typesetters can easily convert this to a proper format as in this PDF file.
Deposition of specimens
Whenever possible, authors are advised to deposit type specimens in national or international
public museums or collections. Authors are also advised to request registration numbers of
deposited material in advance of the acceptance of papers to avoid unnecessary delay of
publication. Some countries (e.g. Australia) require that primary type specimens be deposited
in collections of the country of origin; authors are advised to take this into consideration.

93

94
Normas da Parasitology Research
Instructions for Authors
MANUSCRIPT SUBMISSION
Manuscript Submission
Submission of a manuscript implies: that the work described has not been published before;
that it is not under consideration for publication anywhere else; that its publication has been
approved by all co-authors, if any, as well as by the responsible authorities – tacitly or
explicitly – at the institute where the work has been carried out. The publisher will not be held
legally responsible should there be any claims for compensation.
Permissions
Authors wishing to include figures, tables, or text passages that have already been published
elsewhere are required to obtain permission from the copyright owner(s) for both the print and
online format and to include evidence that such permission has been granted when submitting
their papers. Any material received without such evidence will be assumed to originate from
the authors.
Online Submission
Please follow the hyperlink “Submit online” on the right and upload all of your manuscript
files following the instructions given on the screen.
Costs of Colour Illustrations
Online publication of color illustrations is always free of charge.
TITLE PAGE
Title Page
The title page should include:
The name(s) of the author(s) A concise and informative title
The affiliation(s) and address(es) of the author(s)
The e-mail address, and telephone number(s) of the corresponding author If available, the 16-
digit ORCID of the author(s)
Abstract
Please provide an abstract of 150 to 250 words. The abstract should not contain any undefined
abbreviations or unspecified references.
Keywords
Please provide 4 to 6 keywords which can be used for indexing purposes.

95
TEXT
Text Formatting
Manuscripts should be submitted in Word.
Use a normal, plain font (e.g., 10-point Times Roman) for text. Use italics for emphasis.
Use the automatic page numbering function to number the pages. Do not use field functions.
Use tab stops or other commands for indents, not the space bar. Use the table function, not
spreadsheets, to make tables.
Use the equation editor or MathType for equations.
Save your file in docx format (Word 2007 or higher) or doc format (older Word versions).
Manuscripts with mathematical content can also be submitted in LaTeX.
LaTeX macro package (zip, 181 kB)
Headings
Please use no more than three levels of displayed headings.
Abbreviations
Abbreviations should be defined at first mention and used consistently thereafter.
Footnotes
Footnotes can be used to give additional information, which may include the citation of a
reference included in the reference list. They should not consist solely of a reference citation,
and they should never include the bibliographic details of a reference. They should also not
contain any figures or tables.
Footnotes to the text are numbered consecutively; those to tables should be indicated by
superscript lower-case letters (or asterisks for significance values and other statistical data).
Footnotes to the title or the authors of the article are not given reference symbols.
Always use footnotes instead of endnotes.
Acknowledgments
Acknowledgments of people, grants, funds, etc. should be placed in a separate section on the
title page. The names of funding organizations should be written in full.
Important note:
Authors are requested to use automatic continuous line numbering throughout the manuscript
and in double space.
SCIENTIFIC STYLE
Please always use internationally accepted signs and symbols for units (SI units).

96
Nomenclature
The International Code of Zoological Nomenclature (ICZN) must be observed. Genus and
species names should be in italics. Authors of scientific names of the genus and species group
should not be italicized. At first mention, a specific name should be cited with nomenclatural
author and year, e.g. Catenula lemnae (in italics) Dugès, 1832. When three or more joint
authors have been responsible for a name, then the citation of the name of the authors may be
expressed by use of the term "et al." following the name of the first author, provided that all
authors of the name are cited in full elsewhere in the same work, either in the text or in a
bibliographic reference. Authors unfamiliar with the taxonomy of the group to which a
species belongs should consult an expert to ensure that it is properly identified and that the
correct name is used.
REFERENCES
Citation
Cite references in the text by name and year in parentheses. Some examples: Negotiation
research spans many disciplines (Thompson 1990).
This result was later contradicted by Becker and Seligman (1996).
This effect has been widely studied (Abbott 1991; Barakat et al. 1995a, b; Kelso and Smith
1998; Medvec et al. 1999, 2000).
Reference list
The list of references should only include works that are cited in the text and that have been
published or accepted for publication. Personal communications and unpublished works
should only be mentioned in the text. Do not use footnotes or endnotes as a substitute for a
reference list.
Reference list entries should be alphabetized by the last names of the first author of each
work. Order multi-author publications of the same first author alphabetically with respect to
second, third, etc. author. Publications of exactly the same author(s) must be ordered
chronologically.
Journal article
Gamelin FX, Baquet G, Berthoin S, Thevenet D, Nourry C, Nottin S, Bosquet L (2009) Effect
of high intensity intermittent training on heart rate variability in prepubescent children. Eur J
Appl Physiol 105:731-738. https://doi.org/10.1007/s00421-008-0955-8
Ideally, the names of all authors should be provided, but the usage of “et al” in long author
lists will also be accepted:

97
Smith J, Jones M Jr, Houghton L et al (1999) Future of health insurance. N Engl J Med
965:325–329
Article by DOI
Slifka MK, Whitton JL (2000) Clinical implications of dysregulated cytokine production. J
Mol Med. https://doi.org/10.1007/s001090000086
Book
South J, Blass B (2001) The future of modern genomics. Blackwell, London Book chapter
Brown B, Aaron M (2001) The politics of nature. In: Smith J (ed) The rise of modern
genomics, 3rd edn. Wiley, New York, pp 230-257
Online document
Cartwright J (2007) Big stars have weather too. IOP Publishing PhysicsWeb.
http://physicsweb.org/articles/news/11/6/16/1. Accessed 26 June 2007
Dissertation
Trent JW (1975) Experimental acute renal failure. Dissertation, University of California
Always use the standard abbreviation of a journal’s name according to the ISSN List of Title
Word Abbreviations, see
ISSN LTWA
If you are unsure, please use the full journal title.
For authors using EndNote, Springer provides an output style that supports the formatting of
in- text citations and reference list.
EndNote style (zip, 2 kB)
TABLES
All tables are to be numbered using Arabic numerals.
Tables should always be cited in text in consecutive numerical order.
For each table, please supply a table caption (title) explaining the components of the table.
Identify any previously published material by giving the original source in the form of a
reference at the end of the table caption.
Footnotes to tables should be indicated by superscript lower-case letters (or asterisks for
significance values and other statistical data) and included beneath the table body.