Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO CEARÁDEPARTAMENTO DE ENGENHARIA QUÍMICA
PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
MARIA CRISTIANE MARTINS DE SOUZA
IMOBILIZAÇÃO DE LIPASE DE CANDIDA ANTARCTICA DO TIPO BEM NANOPARTÍCULAS MAGNÉTICAS VISANDO A APLICAÇÃO
NA SÍNTESE DE ÉSTERES
FORTALEZA, CEARÁ
2013

MARIA CRISTIANE MARTINS DE SOUZA
IMOBILIZAÇÃO DE LIPASE DE CANDIDA ANTARCTICA DO TIPO BEM NANOPARTÍCULAS MAGNÉTICAS VISANDO A APLICAÇÃO
NA SÍNTESE DE ÉSTERES
Tese de doutorado apresentada ao Programade Pós-Graduação em Engenharia Química daUniversidade Federal do Ceará, como requisitoparcial à obtenção do título de Doutor em En-genharia Química.
Área de concentração: Desenvolvimento deprocessos químicos ebioquímicos
Orientador: Profa. Dra. Luciana Rocha BarrosGonçalves
FORTALEZA, CEARÁ
2013

Dados Internacionais de Catalogação na Publicação Universidade Federal do Ceará
Biblioteca de Ciências e Tecnologia
S176i Souza, Maria Cristiane Martins de.
Imobilização de lipase de Candida antarctica do tipo B em nanopartículas magnéticas visando a aplicação na síntese de ésteres / Maria Cristiane Martins de Souza. – 2013.
87 f. : il., enc. ; 30 cm. Tese (doutorado) – Universidade Federal do Ceará, Centro de Tecnologia, Departamento de
Engenharia Química, Programa de Pós-Graduação em Engenharia Química, Fortaleza, 2013. Área de concentração: Desenvolvimento de Processos Químicos e Bioquímicos. Orientação: Profa. Dra. Luciana Rocha Barros Gonçalves. 1. Enzimas. 2. Nanoestruturas. 3. Lipase. 4. Nanopartículas. I. Título.
CDD 660


A minha família.

AGRADECIMENTOS
Agradeço a Deus, a minha família e amigos. Principalmente ao meu marido AlbertEinstein Fernandes Muritiba pelo companheirismo e amor diários.
À minha orientadora professora Luciana Rocha Barros Gonçalves, por todas asoportunidades e confiança depositada e por ser um exemplo de profissional e pessoa a qualadmirarei e me espelharei sempre.
Às amigas e amigos: Jocélia Sousa, Maria Valderez Ponte Rocha, Tigressa Helena,Álvaro Daniel, Bete, Larissa Pinto, Anayla Sousa e James Almada por toda a ajuda dada aomeu desenvolvimento pessoal e profissional desde a graduação.
Com carinho aos amigos do Laboratório de Processos Enzimáticos: Kimberle,Bruna, Nathalia, Lucas, Felipe, Leonardo, Ulisses, Cleiton, Bete, Marylane, Camilla e Fátimaque diariamente estiveram comigo e contribuíram para a minha tese. A todos os colegas dogrupo GPBIO pelo companheirismo, e principalmente a Jéssica por ter aceitado a cota.
Aos amigos de Campinas que são uma segunda família: Iara, Igor, Gisele, Rafael,Juliana, Martinha.
Aos amigos Jorge, Marianna, Dayvison, Érica, Michael e Carol que me acompa-nham sempre.
Aos colegas dos laboratórios vizinhos: Karine Bastos, Diego Romão, Karina, Fe-lipe, Francisca Maria, Márcia, Gabi e Regiane.
Ao grupo GPTA por sempre ajudar a todos.
Ao grupo GPSA, especialmente a Janne, Socorro, Carol, Ada e Rosângela peladisponibilidade e paciência de sempre ajudar com as minhas reações.
Ao grupo de Química de Materiais Avançados (GQMAT) pela colaboração na pes-quisa relacionada as nanopartículas magnéticas, especialmente ao Professor Pierre, Rafael eCésar.
A todos os professores e funcionários do Departamento de Engenharia Química-UFC.
A Universidade Federal do Ceará (UFC) e ao departamento de pós-graduação deEngenharía Química-UFC pelo primordial apoio ao desenvolvimento da tese.
A Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), fun-dação do Ministério da Educação (MEC), pelo apoio financeiro.
Aos professores Ivanildo e Pierre pelas contribuições no exame de qualificação.Aos professores James, Dasciana, Pierre, Talita pelas contribuições na defesa de doutorado.
E especialmente ao meu pai (em memória) que me deixou a melhor herança, aeducação.

“Talvez não tenhamos conseguido fazer omelhor, mas lutamos para que o melhor fossefeito. Não somos o que deveríamos ser, nãosomos o que iremos ser.. mas Graças a Deus,não somos o que éramos. ”
(Martin Luther King)

RESUMO
Neste trabalho, nanopartículas magnéticas de ferro (Fe3O4) (NPM) foram avalia-das como suporte para a imobilização de lipase de Candida antarctica do tipo B (CALB). Obiocatalisador (CALB-NPM) foi analisado na catálise dos ésteres: oleato de etila (biodiesel),butirato de metila e etila. Nanopartículas magnéticas são particularmente interessantes paraimobilização enzimática devido as suas propriedades magnéticas favorecerem a fácil separaçãoda mistura reacional através do uso de magnetismo. A enzima CALB é uma enzima capaz deatuar em diversas reações, como, hidrólises e transesterificações. Contudo, um dos problemasdo uso de enzimas como catalisadores homogêneos é a sua recuperação. Assim, é necessárioo uso de suportes que retenham a enzima, mantendo suas características catalíticas. As na-nopartículas foram produzidas pelo método de co-precipitação. Determinou-se o tamanho dasnanopartículas (11 nm) através da técnica de difração de raios-X (DRX) com posterior refi-namento das fases obtidas pelo método Rietveld. Espectros de infravermelho foram obtidospara análise de presença de hidroxilas usando pastilhas de KBr das ferritas magnéticas. Oespectro foi medido na região entre 400 e 4000 cm−1. Modificações foram realizadas na su-perfície das mesmas com γ-aminopropiltrietoxissilano (APTS) e glutaraldeído. No processo deimobilização, a influência da velocidade de agitação (20-250 rpm), carga enzimática (45-200UpNPB.g−1), tempo de contato enzima-suporte (0,5-5 h), concentração de glutaraldeído (2,5 e25 % (m/v)), aditivo dodecil sulfato de sódio (SDS 0,23 %) e reutilização do biocatalisador fo-ram avaliadas. A imobilização foi realizada na presença de 100 mM de tampão bicarbonato desódio, pH 10, a 25 °C. Após a imobilização, a enzima imobilizada exibiu melhor estabilidadetérmica e operacional do que na forma solúvel. As condições ótimas de imobilização foram:velocidade de agitação de 45 rpm, carga enzimática (80 UpNPB.g−1), tempo de imobilizaçãode 1 h, solução de glutaraldeído (25 % (m/v)), possibilitando um rendimento de imobilizaçãode 41,8 % e atividade enzimática do derivado de 29,1 UpNPB/g. Além disso, o biocatalisadormanteve aproximadamente, 53% de atividade catalítica inicial após cinco ciclos consecutivosde reação hidrolítica. Após a imobilização, a estabilidade térmica dos derivados foi realizadaa partir da reação de hidrólise com 0,01 g de CALB-NPM. atividade catalítica da enzima livree imobilizada foi analisada a 60 °C. A produção de esteres foi realizada com o biocatalisadorna melhor condição catalítica. Além das nanopartículas foram analisadas a bioconversão deesteres por resinas acrílicas comerciais (CALB imobilizada). A conversão de oleato de etila foide aproximadamente 90% para os biocatalisadores testados. Os ciclos de reação consecutivos(14) mostram a manutenção da produção de biodiesel. A máxima conversão de buitrato de etila(96,8%) e metila (93,9%) foram obtidos após 8 h de reação a 25 °C com CALB imobilizadaem nanopartículas magnéticas. Os ciclos de reação consecutivos (12) mostram a manutençãoda produção dos ésteres (aproximadamente 76% para as nanopartículas e 79% para a resinaacrílica).
Palavras-chave: Lipase. Nanopartículas. Biodiesel.

ABSTRACT
In this work, magnetic nanoparticles of iron (Fe3O4) (NPM) were evaluated as asupport for the immobilization of lipase Candida antarctica B (CALB). The biocatalyst (CALB-NPM) was analyzed in the catalysis of esters: ethyl oleate (biodiesel), methyl and ethyl buty-rate. Magnetic nanoparticles are particularly interesting for enzyme immobilization due to theirmagnetic properties favoring the easy separation from the reaction mixture by use of magne-tism. The CALB enzyme is an enzyme capable of acting in various reactions, such as hydrolysisand transesterifications. However, one problem of using enzymes as homogeneous catalysts istheir recovery. Thus, it is necessary to use brackets that retain the enzyme while maintainingits catalytic characteristics. Nanoparticles were produced by co-precipitation method. We de-termined the size of the nanoparticles (11 nm) using the technique of X-ray diffraction (XRD)with subsequent refining of the phases obtained by the Rietveld method. Infrared spectra wereobtained for analysis of the presence of hydroxyls using KBr pellets of magnetic ferrites. Thespectrum was measured in the region between 400 and 4000 cm −1. Modifications were car-ried out on the nanoparticles’ surfaces with γ-aminopropyltriethoxysilane (APTS) and glutaral-dehyde. The influence of stirring speed (20-250 rpm), enzyme load (45-200 UpNPB/gsupport),immobilization time (0.5-5 h), glutaraldehyde solution (2.5 and 25%), additive (SDS 0.23%)and reuse of the biocatalyst (six hydrolytic cycles reactions) were evaluated. The immobiliza-tion was performed in the presence of 100mM sodium bicarbonate buffer, pH 10, at 25 °C. Afterimmobilization, CALB exhibited improved thermal and operational stabilities. The best result(Immobilization yield: 53% and immobilized enzyme activity: 29.1 UpNPB/gsupport) wasobtained at 45 rpm, using 200 UpNPB/gsupport and 1h of immobilization. Furthermore, immo-bilized Calb maintained approximately 41.8 % of initial activity after five cycles of hydrolysis.The ethyl oleate production was analyzed with the best condition and compared to commercialacrylic resins (CALB immobilized). The ethyl oleate conversion was approximately 90 % forthe two biocatalyst at 48 h. The consecutive reaction cycles (14) show the maintenance in theproduction of biodiesel. Maximum conversion of methyl butyrate (93.9 %) and ethyl butyrate(96.8 %) were achieved after 8 h of reaction at 25 °C for CALB immobilized onto magneticnanoparticles. The consecutive reaction cycles (12) show the maintenance in the production ofesters (approximately 76 % for nanoparticles and 79 % for acrylic resin).
Keywords: Lipase. Nanoparticles. Biodiesel.

LISTA DE FIGURAS
Figura 2.1 Processo de formação da ligação peptídea das proteínas. . . . . . . . . . . . . . . . . . . 23
Figura 2.2 Estrutura química dos aminoácidos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
Figura 2.3 Mecanismo reacional de lipases. [1] Ligação de lipídeos: ativação da serinanucleofílica pela histidina e ataque nucleofílico do átomo de carbono da carbo-nila pelo Ser O−. [2] Intermediário tetraédrico transiente: com O− estabilizadopor interações com dois grupos NH de peptídeos. A histidina doa um prótonpara liberar o componente álcool do substrato. [3] O intermediário covalente(acil enzima), em que o componente ácido do substrato é esterificado a resí-duo serina da enzima. A entrada da molécula de água é ativada pelo resíduode histidinha, e os íons hidroxila resultantes atuam no ataque nucleofílico aoátomo de carbono da carbonila do intermediário covalente. [4] O resíduo dehistidina doa um próton para o átomo de oxigênio do resíduo de serina ativo,a ligação éster entre a serina e acil componentes é quebrada, e o produto acil éliberado. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
Figura 2.4 Sítio ativo CALB. O sítio ativo pode ser dividido em duas partes, um ladopara ligações acila e a outra para ligações a álcoois, essa restrição é necessáriadevido a transferência de um próton entre o oxigênio do álcool e a histidina dosítio ativo durante a catálise. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
Figura 2.5 Síntese de nanopartículas magnéticas (Fe3O4 ) e funcionalização por APTS(NETTO; ANDRADE; TOMA, 2009) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
Figura 2.6 Modificação da superfície de nanopartículas magnéticas, reticulação por gluta-raldeído e imobilização da enzima CALB (TANG et al., 2011). . . . . . . . . . . . 32
Figura 3.1 Espectro de infravermelho: Nanopartículas magnéticas de ferro. . . . . . . . . . . . 41
Figura 4.1 Efeitos da velocidade de agitação (rpm) em CALB imobilizada em NPM. Ati-vidade do derivado (U.g−1): 25 rpm (Att = 300 U.g−1) e 45 rpm (Att = 200U.g−1) (agitação rotacional) (�); 25 rpm (Att = 211,3 U.g−1), 45 rpm (Att =185,3 U.g−1), 90 rpm (Att = 116,4 U.g−1), 180 rpm (Att = 115,7 U.g−1) 250rpm (Att = 148,2 U.g−1) (agitação orbital) (�). . . . . . . . . . . . . . . . . . . . . . . . . . . 51

Figura 4.2 Efeito da carga enzimática (Att , U.g−1) na imobilização de CALB em IMNs.Atividade derivada (U.g−1) (�) e Rendimento da imobilização (%) (�). Aslinhas representam a tendência dos dados experimentais. . . . . . . . . . . . . . . . . . 52
Figura 4.3 Efeito do tempo de contato (0,5 - 5h) em CALB imobilizada em NPM. Ati-vidade teórica: 0,5 h (Att= 86,2 U.g−1), 1h (Att= 80 U.g−1), 2h (Att= 124,4U.g−1), 3h (Att= 75 U.g−1), 4h (Att= 84,8 U.g−1), 5h (Att= 82,4 U.g−1). Ativi-dade do derivado (U.g−1) (�) e quantidade de proteína (mg.g−1 (�). As linhasrepresentam a tendência dos dados experimentais. . . . . . . . . . . . . . . . . . . . . . . . 53
Figura 4.4 Estabilidade operacional na hidrólise de butirato de p-nitrophenila para CALB-NPM. Atividade relativa (%): 30 min (�), 1 h (�), 2 h (4) , 3 h (N), 4 h (◦) e 5h (•). As linhas representam a tendência dos dados experimentais. . . . . . . . . 53
Figura 4.5 Efeito de aditivos (0,23%, SDS) e concentração de glutaraldeído (2,5 e 25%(m/v)) em CALB imobilizada em NPM. Atividade teórica: Glut. 2,5 % (Att=62 U.g−1), Glut. 25 % (Att= 80 U.g−1), Glut. 25 % + SDS 0,23 % (Att= 55,8U.g−1). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
Figura 4.6 Estabilidade operacional em hidrolise de butirato de p-nitrofenila para CALB-NPM. Aditivos: 25% glutaraldeído (�) e (25% glutaraldeído + 0,23%, SDS)(�). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
Figura 4.7 Estabilidade térmica de CALB livre e CALB-IMNs a 60°C e pH 7. Enzimasolúvel Ati= 93 U.mL−1 e CALB-IMNs Att= 80 U.g−1. Atividade relativa:CALB imobilizada (•) e CALB livre (N). AS linhas representam a tendenciado modelo de Sadana e Henley. (FERNANDEZ-LAFUENTE et al., 1993) . 56
Figura 4.8 Dinâmica da síntese de oleato de etila usando CALB-NPM Att= 80 U.g−1, a37°C e 96 horas em agitação orbital. CALB− IMNs (�) e Novozym® 435(�). Reagentes: ácido oleico (0,67 mL) e etanol (0,15 mL) a mol.L−1 (livre desolvente). Volume reacional = 0,82 mL. Massa derivada = 0,01g) . . . . . . . . . 57
Figura 4.9 Estabilidade operacional da síntese de oleato de etila usando CALB-NPM Att=80 U.g−1. Cada ciclo teve duração de 48 h, agitação orbital, a 37°C. CALB−IMNs (�), Novozym® 435 (•). Reagentes: ácido oleico (0,67 mL) e etanol(0,15 mL) (livre de solvente). Volume reacional = 0,82 mL. Massa derivada =0,01g) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58

Figura 5.1 Efeito da temperatura na síntese dos butiratos de metila e etila. Substratos: eta-nol (�) e metanol (•). As reações foram realizadas utilizando CALB-NPM Att=80 U.g−1, 150 rpm, heptano, 02 mol/L de ácido butírico, 1:1 (ácido butírico:(metanol and etanol)) e 8 h de reação. Volume reacional = 1,0 mL. Massa decatalisador CALB-NPM = 0,01g. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
Figura 5.2 Efeito da concentração de substratos na conversão de butirato de etila e metila.Substrato álcoois: etanol (�) e metanol (•). As reações foram realizadas utili-zando CALB-NPM Att= 80 U.g−1, 25 °C, 150 rpm, heptano, 0,1-1,0 mol/L deácido butírico, 1:1 (ácido butírico: (metanol e etanol)) e 8 h de reação. Volumereacional de 1,0 mL. Massa de biocatalisador CALB-NPM de 0,01 g. . . . . . 65
Figura 5.3 Efeito da razão molar álcool:ácido na reação de esterificação para a produçãodos ésteres butirato de etila e metila. Perfil da variação da concentração deálcool (�) e ácido (�) para butirato de etila. Perfil da variação da concentraçãode álcool (•) e ácido (◦) para butirato de metila. As reações foram conduzidaspor uso de CALB-NPM Att= 80 U.g−1, 25 °C, 150 rpm, heptano, 0.5 mol/L(metanol) and 0.4 mol/L (etanol), 1:1-1:4 (ácido butírico: (metanol e etanol)) e8 h de reação. Volume reacional de 1,0 mL. Massa de catalisador CALB-NPMde 0,01 g. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66
Figura 5.4 Efeito da velocidade de agitação na síntese de buriato de etila e metila. Os álco-ois usados foram: etanol (�) e metanol (•). Reações foram realizadas usandoCALB-NPM Att= 80 U.g−1, 25 °C, 150-250 rpm, heptano, 0,5 (metanol) e 0,4(etanol) mol/L ácido butírico, 1:1 (ácido butírico: (metanol e etanol)) e 8 htempo de reação. Volume reacional = 1,0 mL. Massa do catalisador = 0,01 g 67
Figura 5.5 Dinâmica da síntese de butirato de etila e metila. Os álcoois usados foram:etanol (�) e metanol (•). Reações foram realizadas usando CALB-NPM Att=80 U.g−1, at 25°C, 8 h, 150 rpm, heptano, 0,5 (metanol) e 0,4 (etanol) mol/Lácido butírico, 1:1 (ácido butírico: (metanol e etanol)). Volume reacional = 1,0mL. Massa do catalisador = 0,01 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
Figura 5.6 Estabilidade operacional de CALB-IMNs(�) e CALB-resina acrílica(•) em sín-tese de buriato de etila. Reações foram executadas a 25°C, 8 h, 150 rpm, 0,4mol/L ácido butírico, 1:1 (ácido butírico: (metanol e etanol)). Volume reacio-nal = 1,0 mL. Massa do catalisador = 0,01 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
Figura 5.7 Estabilidade operacional de CALB-NPM (�), resina acrílica (•) em síntese de

buriato de etila. Reações foram executadas a 25°C, 8 h, 150 rpm, 0,4 mol/Lácido butírico, 1:1 (ácido butírico: (metanol e etanol)). Volume reacional = 1,0mL. Massa do catalisador = 0,01 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71

LISTA DE TABELAS
Tabela 2.1 Valores de pKa de aminoácidos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
Tabela 2.2 Enzimas imobilizadas em nanopartículas magnéticas e suas aplicações biotec-nológicas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
Tabela 3.1 Parâmetros de imobilização . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
Tabela 3.2 Parâmetros de imobilização . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
Tabela 4.1 Desativação térmica a 60°C e pH 7, de CALB solúvel e CALB-NPM . . . . . . 56
Tabela 5.1 Efeito de solvente orgânico em síntese de butirato de metila e etila. Reaçõesforam realizadas usando CALB-NPM Att= 80 U.g−1, 25 °C, 150 rpm, 0,5(metanol) e 0,4 (etanol) mol/L ácido butírico, 1:1 (ácido butírico: (metanole etanol)) e 8 h tempo de reação. Volume de reação = 1,0 mL. Massa docatalisador = 0,01 g . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68

SUMÁRIO
LISTA DE SIGLAS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1 INTRODUÇÃO . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
1.1 Contextualização . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
1.2 Justificativa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
1.3 Descrição dos capítulos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
1.4 Objetivo geral . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
1.5 Objetivos específicos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
2 REVISÃO BIBLIOGRÁFICA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
2.1 Conceitos básicos sobre proteínas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
2.1.1 Propriedades químicas de proteínas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
2.2 Enzimas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
2.3 Lipases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
2.4 Lipase de Candida antarctica do tipo B (CALB) . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
2.5 Nanopartículas magnéticas de ferro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
2.5.1 Produção de nanopartículas magnéticas de ferro Fe3O4 . . . . . . . . . . . . . . . . . . . . . . . 30
2.6 Imobilização de enzimas em nanopartículas magnéticas de ferro . . . . . . . . . . . . 30
2.7 Enzimas como catalisadores heterogêneos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
2.7.1 Atores do Processo catalisado . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
2.7.1.1 Água . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
2.7.1.2 Solventes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
2.7.1.3 Temperatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
2.8 Ésteres . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
3 ESTUDOS PRELIMINARES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
3.1 Materiais . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
3.2 Métodos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
3.2.1 Preparação de nanopartículas de quitosana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
3.2.2 Preparação de nanopartículas magnéticas de ferro . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
3.2.3 Tratamento do suporte com aminopropiltrietoxisilano (APTS) . . . . . . . . . . . . . . . . . 39

3.2.4 Ativação do suporte com Glutaraldeído . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.2.5 Imobilização Enzimática . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.2.6 Medida da atividade enzimática . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.2.7 Parâmetros de imobilização . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.2.7.1 Rendimento de imobilização . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.2.7.2 Atividade recuperada (Atr) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.3 Resultados e discussões . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.3.1 Nanopartículas de quitosana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.3.2 Caracterização das nanopartículas magnéticas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.3.3 Imobilização em nanopartículas magnéticas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.3.4 Influência de aditivos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.4 Resultados e discussão . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.4.1 Influência do suporte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.4.2 Influência do surfactante . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
3.5 Produção de ésteres . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
4 PRODUÇÃO DE OLEATO DE ETILA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
Síntese de oleato de etila catalisada por lipase do tipo B de Candida antarctica imo-bilizada em nanopartículas magnéticas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
Resumo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
4.1 Introdução . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
4.2 Materiais e métodos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
4.2.1 Materiais . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
4.2.2 Síntese de (Fe3O4) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
4.2.3 Tratamento do suporte com γ-aminopropiltrietoxisilano (APTS) . . . . . . . . . . . . . . . 48
4.2.4 Reticulação com solução de glutaraldeído . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
4.2.5 Imobilização . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
4.2.6 Concentração de proteínas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
4.2.7 Estabilidade térmica da CALB solúvel e imobilizada . . . . . . . . . . . . . . . . . . . . . . . . . 49
4.2.8 Síntese de oleato de etila . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
4.2.9 Estabilidade operacional de CALB imobilizada . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50

4.2.9.1 Hidrólise de butirato de p-nitrofenila . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
4.2.9.2 Síntese . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
4.3 Resultados e discussões . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
4.3.1 Efeito da velocidade de agitação na imobilização de CALB . . . . . . . . . . . . . . . . . . . . 50
4.3.2 Efeito da carga enzimática . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
4.3.3 Efeito do tempo de contato . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
4.3.4 Efeito de diferentes concetrações de glutaraldeído . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
4.3.5 Estabilidade térmica de CALB solúvel e imobilizada . . . . . . . . . . . . . . . . . . . . . . . . . 55
4.3.6 Esterificação catalítica . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56
4.3.7 Estabilidade operacional do derivado . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
4.4 Conclusões . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
Agradecimentos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
5 PRODUÇÃO DE BUTIRATO DE METILA E ETILA . . . . . . . . . . . . . . . . . . . . . . . 59
Produção de ésteres por lipase de Candida antarctica tipo B imobilizadas em nano-partículas magnéticas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
Resumo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
5.1 Introdução . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61
5.2 Métodos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.2.1 Materiais . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.2.2 Produção de nanopartículas magnéticas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.2.3 Modificação por APTS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.2.4 Glutaraldeído crosslinking . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.2.5 Imobilização de lipase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
5.2.6 Síntese . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
5.2.7 Reuso do biocatalisador . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
5.3 Resultados e discussões . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
5.3.1 Efeito da temperatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
5.3.2 Efeito da concentração de substratos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
5.3.3 Efeito da razão molar álcool:ácido . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66
5.3.4 Efeito da velocidade de agitação . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67

5.3.5 Produção de ésteres na presença de solventes orgânicos . . . . . . . . . . . . . . . . . . . . . . 68
5.3.6 Tempo de reação . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
5.3.7 Estabilidade operacional . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
5.4 Conclusões . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71
Agradecimentos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72
6 CONCLUSÃO E PROPOSTAS PARA TRABALHOS FUTUROS . . . . . . . . . . . . 73
REFERÊNCIAS BIBLIOGRÁFICAS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74

LISTA DE SIGLAS
NPM Nanopartículas magnéticas de ferroCALB Lipase de Candida antarctica do tipo BDRX Difração de raios-XAPTS γ-aminopropiltrietoxissilanoSDS Dodecil sulfato de sódioGQMAT Grupo de química de materiais avançadospNPB p-Nitrophenil ButiratopNP p-nitrophenoiTEM Transmissão eletrônica de varreduraSF Fator de estabilização

19
1 INTRODUÇÃO
1.1 Contextualização
O desenvolvimento de processos químicos considerados ambientalmente corretos éum dos grandes desafios para pesquisadores e indústria, tal fato requer o aprimoramento dosprocessos, com o objetivo fundamental de reduzir os resíduos tóxicos indesejáveis ao ambiente.Dentro deste cenário, o uso de catalisadores químicos tóxicos são um dos principais prolemas aserem contornados, já que são os principais mediadores de reações usadas industrialmente. Emcontrapartida aos catalisadores químicos, as enzimas são consideradas catalisadores "verdes",sendo altamente seletivas reduzindo a produção de reações indesejáveis. As condições brandasde reação aplicadas aos processos enzimáticos também favorecem sua demanda industrial comocatalisador (HASAN; SHAH; HAMEED, 2006).
Assim, a procura por catalisadores "ideais"que incorporem as vantagens dos cata-lisadores homogêneos (elevada atividade, seletividade e reprodutibilidade), heterogêneos (altoperíodo de reuso e facilidade de recuperação) e não danosos ao ambiente são os grandes moti-vadores das pesquisas atuais (END; SCHÖNING, 2004).
1.2 Justificativa
O emprego de suportes sólidos para a imobilização enzimática é necessário devidoa possibilidade de reuso do biocatalisador e para favorecer a estabilidade da enzima. A escolhado biocatalisador é essencial nesse processo, pois apresenta grande influência no processo deseparação, recuperação e reutilização do catalisador (CLARK; RHODES, 2000). Nesse con-texto, o uso de nanopartículas magnéticas como suporte sólido confere ao biocatalisador umdos requisitos essenciais, o de fácil recuperação por simples aplicação de campo magnético.O superparamagnetismo facultam a esses materiais grande facilidade de isolamento e sepa-ração de meios multifásicos complexos pelo simples uso de um ímã permanente, e sua fácildispersão depois de cessada a aplicação do campo. A preparação de nanomateriais com pro-priedades magnéticas e superfícies reativas permite a imobilização de enzimas, possibilitandouma maior recuperação e reutilização do biocatalisador, atendendo aos princípios da química"verde” (BRUNO et al., 2005).
No presente trabalho, será abordado o desenvolvimento das melhores condiçõespara o uso eficiente de biocatalisadores enzimáticos suportados em nanopartículas magnéticase aplicação em reações de interesse industrial. Especificamente, serão discutidas sobre as ques-tões relacionadas a manutenção da eficiência catalítica após a imobilização em nanopatículasmegnéticas e a eficiência catalítica frente a reações de produção de ésteres, como biodiesel earomas.
A contribuição deste trabalho é o estudo da biocatálise heterogênea na produção deésteres por enzima imobilizada em nanopartículas magnéticas. A produção industrial dessescompostos decorre principalmente por catálise química homogênea, sendo considerados esses

20
processos danosos ao ambiente. Como exposto anteriormente, a biocatálise heterogênea apre-senta vantagens em relação a catálise química homogênea e heterogênea, neste trabalho apli-camos a biossíntese de esteres por lipase imobilizada em nanopartículas magnéticas, obtendoresultados competitivos em relação a produção de ésteres, a reutilização do biocatalisador efacilidade de sepação do meio reacional.
1.3 Descrição dos capítulos
A tese é dividida em seis capítulos. O capítulo 2 introduz conceitos básicos sobrea estrutura química das proteínas e suas características funcionais. O uso e eficiência catalíticade lipases é apresentado, sendo discutido também os parâmetros que influenciam na biocatá-lise heterogênea, sendo enfatizada a produção de ésteres. O capítulo 3 relata os experimentospreliminares realizados com o objetivo de estudar o sistema de imobilização e os materiaisdisponíveis como suportes e escolher a melhor metodologia de imobilização.
O capítulo 4 é reservado a discussão da produção de biodiesel por lipase imobilizadaem nanopartículas magnéticas, apresentando-se na forma de artigo a ser publicado em periódicointernacional, consistindo do estudo detalhado dos parâmetros que afetam a imobilização da en-zima lipase de Candida antarctica do tipo B (CALB) em nanopartículas magnéticas (NPM). Amelhor condição de imobilização foi utilizada para a produção de biodiesel, comparando-se aprodução com a enzima CALB imobilizada em resina acrílica comercial (Novozyme® 435).Ao longo do capítulo 5, apresentando-se na forma de artigo a ser publicado em periódico in-ternacional, foi apresentado o estudo dos parâmetros que influenciam a biocatálise de aromaspor CALB-IMNs. Os resultados são comparados com Novozyme® 435. Finalmente, no capí-tulo 6, foram propostos caminhos para trabalhos futuros e são sintetizamos as contribuições dopresente trabalho.
1.4 Objetivo geral
O objetivo geral deste trabalho é estudar a imobilização de lipase comercial, lipasede Candida antartica do tipo B (CALB) em nanopartículas magnéticas (NPM) e sua aplicaçãoem reações de interesse industrial.
1.5 Objetivos específicos
• Estudar a modificação da superfície das nanopartículas magnéticas de ferro com amino-propiltrietoxissilano (APTS) e glutaraldeído para posterior imobilização de CALB;
• Caracterizar a lipase CALB solúvel e imobilizada em relação a estabilidade térmica, ati-vidade enzimática utilizando como substrato para-nitrofenolbutirato pNPB e quantidadede proteína por volume de solução e massa de suporte;
• Imobilizar a CALB em nanoestruturas magnéticas por imobilização covalente.

21
• Estudar o processo de imobilização da lipase em nanopartículas magnéticas (tempo deimobilização, velocidade de imobilização, tipo de agitação, concentração de reticulante(glutaraldeído), detergente, quantidade de enzima solúvel por grama de suporte e reusodo biocatalisador na reação de hidrólise do pNPB);
• Estudar a síntese de ésteres oleato de etila (biodiesel), butirato de etila e metila por enzimaimobilizada em nanopartículas magnéticas;
• Estudar os parâmetros de reação para a produção dos aromas butirato de etila e metila:temperatura, velocidade de reação, concentração de substratos, solventes, tempo de rea-ção e reuso do biocatalisador na reação de esterificação;
• Comparar a síntese dos ésteres utilizando a CALB-NPM com a síntese utilizando CALBimobilizada em resina acrílica (Novozyme® 435).

22
2 REVISÃO BIBLIOGRÁFICA
2.1 Conceitos básicos sobre proteínas
"Quais seriam as propriedades dos materiais se nós pudéssemos arrumar os átomosda forma que nós queremos? Não posso ver exatamente o que aconteceria, mas não duvido quequando tivermos algum controle sobre o arranjo das coisas em pequena escala nós teremos umavariedade enormemente maior de propriedades que as substâncias podem ter e das diferentescoisas que podemos fazer".
Richard P. Feynman, 29 de dezembro de 1959 no encontro anual da sociedade ame-ricana de Física.
O estudo sobre as propriedades das proteínas começou no século XVIII, sendo queno início do século XX os químicos passaram a analisar minuciosamente essas substâncias,descobrindo que a sua degradação liberava aminoácidos. Por volta de 1900, já haviam sidoidentificados doze aminoácidos diferentes liberados pela degradação de proteínas. Em 1906,já haviam sido identificados quinze tipos de aminoácidos; em 1935, esse número aumentoupara dezoito e, em 1940, chegou a vinte, completando a lista de aminoácidos que ocorremnaturalmente nas proteínas dos seres vivos (AMABIS; MARTHO, 1997).
2.1.1 Propriedades químicas de proteínas
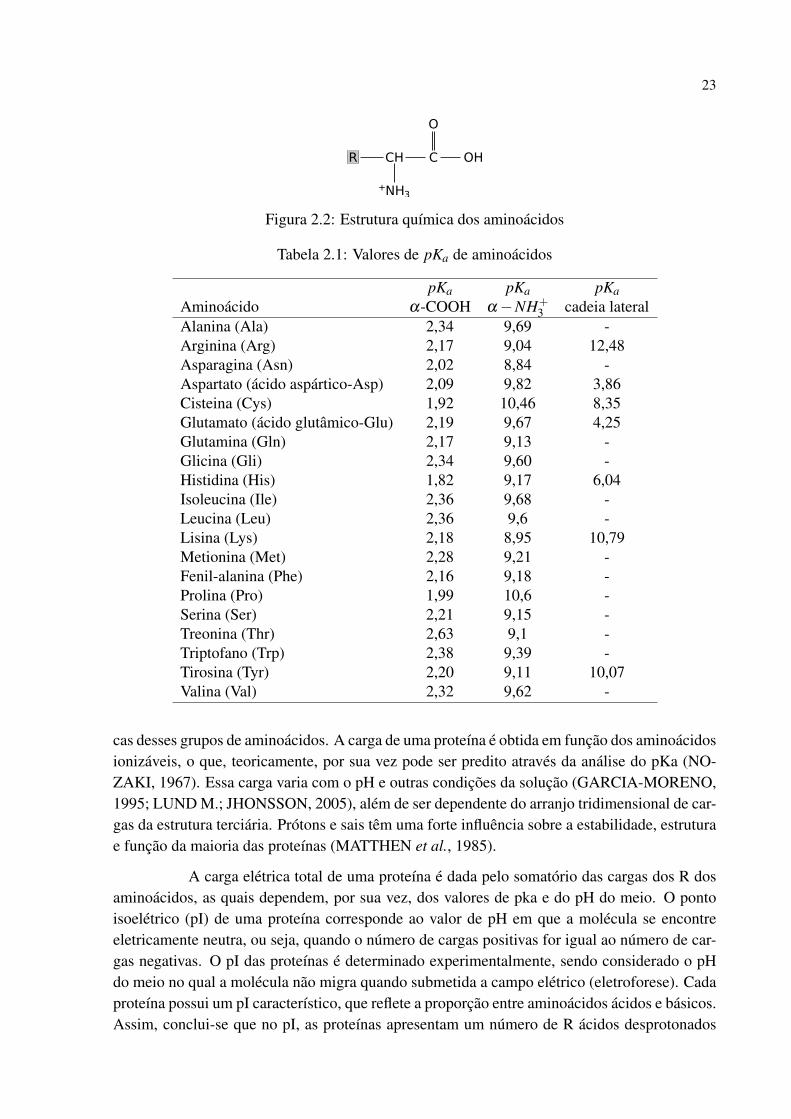
Proteínas são biomoléculas constituídas por polímeros de aminoácidos unidos porligações peptídicas. As unidades que se repetem são denominadas resíduos de aminoácidos.O grupo carboxílico de um aminoácido se une ao grupo amina de outro aminoácido formandouma ligação peptídica (C-N) e liberando uma molécula de água, ver Figura 2.1, adaptada deSouza, 2010 (SOUZA, 2010). Os aminoácidos (ácidos α-aminocarboxílicos) representados naFigura 2.2 diferem somente no substituinte (R) ligado ao carbono α .
R
R
R
R
Aminoácido (1)
Aminoácido (2)
Dipeptídeo
1
1
2
2
Água
Figura 2.1: Processo de formação da ligação peptídea das proteínas.
As macromoléculas biológicas contêm grupos de aminoácidos ionizáveis que fazemcom que sua carga líquida varie em função das condições da solução (JHONSSON; LUND;SILVA, 2007). A Tabela 2.1, adaptada de Bruice, 2006 (BRUICE, 2006), mostra as característi-
23
R CH C
O
OH
NH+3
Figura 2.2: Estrutura química dos aminoácidos
Tabela 2.1: Valores de pKa de aminoácidos
pKa pKa pKaAminoácido α-COOH α−NH+
3 cadeia lateralAlanina (Ala) 2,34 9,69 -Arginina (Arg) 2,17 9,04 12,48Asparagina (Asn) 2,02 8,84 -Aspartato (ácido aspártico-Asp) 2,09 9,82 3,86Cisteina (Cys) 1,92 10,46 8,35Glutamato (ácido glutâmico-Glu) 2,19 9,67 4,25Glutamina (Gln) 2,17 9,13 -Glicina (Gli) 2,34 9,60 -Histidina (His) 1,82 9,17 6,04Isoleucina (Ile) 2,36 9,68 -Leucina (Leu) 2,36 9,6 -Lisina (Lys) 2,18 8,95 10,79Metionina (Met) 2,28 9,21 -Fenil-alanina (Phe) 2,16 9,18 -Prolina (Pro) 1,99 10,6 -Serina (Ser) 2,21 9,15 -Treonina (Thr) 2,63 9,1 -Triptofano (Trp) 2,38 9,39 -Tirosina (Tyr) 2,20 9,11 10,07Valina (Val) 2,32 9,62 -
cas desses grupos de aminoácidos. A carga de uma proteína é obtida em função dos aminoácidosionizáveis, o que, teoricamente, por sua vez pode ser predito através da análise do pKa (NO-ZAKI, 1967). Essa carga varia com o pH e outras condições da solução (GARCIA-MORENO,1995; LUND M.; JHONSSON, 2005), além de ser dependente do arranjo tridimensional de car-gas da estrutura terciária. Prótons e sais têm uma forte influência sobre a estabilidade, estruturae função da maioria das proteínas (MATTHEN et al., 1985).
A carga elétrica total de uma proteína é dada pelo somatório das cargas dos R dosaminoácidos, as quais dependem, por sua vez, dos valores de pka e do pH do meio. O pontoisoelétrico (pI) de uma proteína corresponde ao valor de pH em que a molécula se encontreeletricamente neutra, ou seja, quando o número de cargas positivas for igual ao número de car-gas negativas. O pI das proteínas é determinado experimentalmente, sendo considerado o pHdo meio no qual a molécula não migra quando submetida a campo elétrico (eletroforese). Cadaproteína possui um pI característico, que reflete a proporção entre aminoácidos ácidos e básicos.Assim, conclui-se que no pI, as proteínas apresentam um número de R ácidos desprotonados

24
(COO−) igual ao número de R básicos protonados (NH3+). Desta forma, as proteínas apresen-
tarão carga líquida positiva em pH menor que o pI e carga líquida negativa em pH maior que opI (BRUICE, 2006).
A ampla variação dos substituintes (denominados cadeias laterais) é o que dá às pro-teínas sua grande diversidade estrutural e, consequentemente, sua grande diversidade funcional.Algumas protegem os organismos de seu ambiente ou fortalecem certas estruturas biológicas.Um grupo de proteínas denominadas enzimas catalisa as reações químicas que ocorrem nossistemas vivos (BRUICE, 2006).
2.2 Enzimas
Uma importante classe de proteínas são as enzimas. Enzimas são consideradas cata-lisadores naturais que aceleram a velocidade das reações, sendo altamente versáteis na catálisede vários tipos de reações que ocorrem sob condições brandas, normalmente à temperatura am-biente e em pH próximo da neutralidade. As velocidades de algumas reações podem alcançarvalores 1012 vezes maiores do que as não catalisadas (VOET; VOET, 1995).
Enzimas são proteínas formadas por subunidades de aminoácidos (entre 62 a 2500resíduos de aminoácidos), combinadas por ligações peptídicas. Os resíduos de aminoácidosformam ligações covalentes entre si, pelo grupo amino de um aminoácido com o grupamentocarboxílico de outro aminoácido, constituindo cadeias polipeptídicas extensas, que assumemum arranjo espacial e estrutural complexo (VOET; VOET, 1995).
A função catalítica das enzimas está relacionada à capacidade de interação enzima-substrato, essas interações são possíveis devido a orientações favoráveis na formação dos com-plexos enzima-substrato. As enzimas possuem um região ativa, uma parte da estrutura proteicaque possibilita a catálise enzimática. O substrato liga-se a região ativa da enzima e parte daespecificidade catalítica da mesma depende também da natureza das ligações envolvidas. Essasligações dependem do arranjo precisamente definido de átomos no sítio ativo (PALMER, 1995).
Em relação aos catalisadores químicos convencionais, as enzimas possuem muitasvantagens que tornam seu uso viável em aplicações industriais, uma dessas vantagens é a for-mação de produtos considerados "verdes” devido a alta especificidade das enzimas, evitando aformação de produtos indesejáveis e prejudiciais ao ambiente. As condições brandas de reaçãoaplicadas aos processos enzimáticos também favorecem sua demanda industrial como catalisa-dor (HASAN; SHAH; HAMEED, 2006).
O mercado mundial de enzimas industriais é estimado em US $ 4,0 bilhões (NO-VOZYMES, 2011). O segmento industrial é dividido em enzimas para limpeza doméstica,alimentos e bebidas, bioenergia, ração animal e outras enzimas técnicas, micro-organismos ebiofarmacêuticos. Diversas enzimas, como, proteases, amilases, lipases, celulases, xilanases efitases são utilizadas industrialmente, sendo os maiores produtores as empresas Gist-Brocades(Holanda), Genencor International (Finlândia) e Novozyme (Dinamarca) (MUSSATTO; FER-NANDES; MILAGRES, 2007).

25
Dentre os grupos de enzimas destacam-se as lipases, devido à gama de reações ca-talisadas em sistemas orgânicos com baixo teor de água (esterificação, interesterificação, ami-nólise, lactonização), à alta estabilidade nestes ambientes e à possibilidade de utilização emcondições brandas de reação. Essa flexibilidade aliada às diferentes lipases confere a essas en-zimas um potencial enorme de aplicações (KIM; YOUN; SHIN, 2006; YIN; LIU; TAN, 2006).
2.3 Lipases
Lipases (glicerol éster hidrolases, EC 3.1.1.3) catalisam a hidrólise de cadeias lon-gas de triglicerídeos insolúveis em meios aquosos e outros ésteres insolúves de ácidos graxos(SCHøNHEYDER; VOLQVARTZ, 1946; TRODLER; PLEISS, 2008). A cavidade catalítica(sítio ativo) das lipases consiste de três resíduos de aminoácidos: um resíduo nucleofílico (cis-teína, serina, ou aspartato), um resíduo ácido catalítico (aspartato ou glutamato), e um resíduode histidina (JAEGER; DIJKSTRA; REETZ, 1999).
A hidrólise de ésteres ocorre em duas etapas, como representado na Figura 2.3,adaptado de Jaeger et al. 1999 (JAEGER; DIJKSTRA; REETZ, 1999). A reação tem iníciocom o ataque nucleofílico do átomo de oxigênio presente no grupo hidroxila da serina ao car-bono do grupo carbonila do éster. Um intermediário tetraédrico é formado, que é caracterizadopela carga negativa do oxigênio da carbonila e o arranjo tetraédrico do átomo de carbono. Ointermediário é estabilizado pelos resíduos catalíticos de histidina e aspartato. Um próton dogrupo hidroxila da serina é transferido. A transferência deste próton é facilitada pela presençado ácido catalítico, que orienta o anel imidazol da histidina e neutraliza as cargas formadas. Opróton é doado ao oxigênio do éster, assim quebrando a ligação. Nesta etapa o componenteácido do substrato é esterificado a serina nucleofílica ("intermediário covalente"), enquanto oálcool é difundido (JAEGER; DIJKSTRA; REETZ, 1999).
A próxima etapa é a desacetilação, em que uma molécula de água hidrolisa (águana hidrólise e álcool na esterificação) o intermediário covalente. O sítio ativo da histidina ativaa molécula de água. Os íons OH− resultantes atacam o átomo de carbono da carbonila do grupoacila covalentemente ligado a serina. Novamente, um intermediário tetraédrico negativamentecarregado é formado. A histidina doa um próton para o átomo de oxigênio do resíduo de serinaativo, que então libera o componente acila. Depois da difusão do produto a enzima é recuperadanas mesmas condições iniciais, podendo o ciclo catalítico recomeçar (JAEGER; DIJKSTRA;REETZ, 1999).
Um grande número de lipases tem sido caracterizadas pela ampla variação na efi-ciência e especificidade por substratos (UPPENBERG et al., 1994; ROGALSKA et al., 2004).Triglicéridos podem ser clivado em todas as três ligações de éster ou especificamente em apenasuma ou duas posições. Lipases também podem mostrar diferentes especificidades em relaçãoao comprimentos dos ácidos graxos. Em meios orgânicos, o comportamento enzimático apre-senta alterações e estas enzimas podem ser utilizados para a transesterificação e outras reaçõesde síntese para produzir novos tipos de lipídios (UPPENBERG et al., 1994).
Enzimas apresentam diversas vantagens em relação à catálise química para uso in-

26
dústrial (WATANABE et al., 2001; SOO et al., 2004; VILLENEUVE, 2007). Dentre elas,pode-se citar elevada seletividade, condições reacionais brandas, que permitem um menor con-sumo energético e evitam degradação ou decomposição de produtos e reagentes. Além disso,lipases são aceitas ecologicamente, tornando o produto de fácil aceitação. Destacam-se entreas lipases, as lipases produzidas por Candida sp., que são enzimas bem estabelecidas para finsde biocatálise, especialmente lipase de Candida antarctica do tipo B (CALB) (MARIA et al.,2005).
O
O
N
O
H
N
O
H
His
Asp
HN N
HO
−O
O
Ser
1
Ser
His
Asp
HNH
O
O
O
H
O
NH
N
O
N+−O
O
2
His
Asp
−O
O
Ser
3 4
OH
HN NH
−OH
O
O
N
HN
O
O
H
O
O
N
O
H
N
O
H
H
His
Asp
HN N
HO
−O
O
Ser
Figura 2.3: Mecanismo reacional de lipases. [1] Ligação de lipídeos: ativação da serina nucle-ofílica pela histidina e ataque nucleofílico do átomo de carbono da carbonila pelo Ser O−. [2]Intermediário tetraédrico transiente: com O− estabilizado por interações com dois grupos NHde peptídeos. A histidina doa um próton para liberar o componente álcool do substrato. [3] Ointermediário covalente (acil enzima), em que o componente ácido do substrato é esterificado aresíduo serina da enzima. A entrada da molécula de água é ativada pelo resíduo de histidinha, eos íons hidroxila resultantes atuam no ataque nucleofílico ao átomo de carbono da carbonila dointermediário covalente. [4] O resíduo de histidina doa um próton para o átomo de oxigênio doresíduo de serina ativo, a ligação éster entre a serina e acil componentes é quebrada, e o produtoacil é liberado.

27
2.4 Lipase de Candida antarctica do tipo B (CALB)
A lipase CALB tem sido estudada para potenciais aplicações na indústria alimen-tícia, de detergente, farmacêutica, têxtil, cosméticos, papel e oleoquímica (HASAN; SHAH;HAMEED, 2006). Estas aplicações são possíveis, devido à sua ampla gama de especificidadepara substratos, alta atividade para várias reações sob condições brandas, resistência a solventesorgânicos, estabilidade térmica, atuação em uma faixa ampla de pH e elevada estéreo espe-cificidade (MCCABE; RODGER; TAYLOR, 2005; RODRIGUES et al., 2008; DENG et al.,2011).
CALB pertence à família das α/β hidrolases com uma tríade catalítica conservadaque consiste em Ser, His, Asp/Glu, tendo o mesmo mecanismo reacional de outras lipases, veritem 2.3, adaptado de Jaeger et al. 1999 (JAEGER; DIJKSTRA; REETZ, 1999). É compostapor 317 resíduos de aminoácidos com um peso molecular de 33 kDa e pI de 6,0 (UPPENBERGet al., 1994). A cavidade de ligação para o substrato ("sítio ativo") consiste em uma cavidadepara ligações acila e outra porção para ligações com álcoois. Em contraste com a maioria daslipases, que apresentam um impedimento maior ("tampa") na entrada do sítio ativo, na CALBessa tampa é reduzida e não prejudica a entrada de substratos e saída de produto(UPPENBERGet al., 1994).
No caso específico da CALB, a cavidade ligante do substrato é elíptica, ver Figura2.4, com o formato de um funil ingrime (TRODLER; PLEISS, 2008). As paredes desse canalsão bastante hidrofóbicas. Contudo, na parte superior do sítio ativo há apenas uma pequenaárea hidrofóbica, o que favorece a não formação de camadas sobrepostas de moléculas por estaenzima (UPPENBERG et al., 1994; TRODLER; PLEISS, 2008).
O uso de CALB em processos industriais, apresentam-se na produção de compos-tos opticamente ativos na indústria farmacêutica (ARROYO; SÁNCHEZ-MONTERO; SINIS-TERRA, 1999; FERNANDEZ-LORENTE et al., 2001; STRAATHOF et al., 2002); produçãode ésteres usados na indústria de aromas (LOZANO et al., 2002; BRÍGIDA et al., 2008); sín-tese de oleato de cetila, um análogo do óleo de cachalote que possui importantes aplicações emindústrias de cosméticos, biodiesel (SILVA et al., 2012), alimentos e fármacos (CASTRO et al.,2004); síntese de lactato de butila, éster bastante utilizado em indústria de alimentos, cosméticose farmacêutica por possuir propriedades higroscópicas, emulsificantes e esfoliantes (PIROZZI;GRECO, 2004); síntese de ésteres de vitaminas, objetivando tornar estas últimas solúveis emóleo para adição em produtos alimentícios (MAUGARD; LEGOY, 2000; ADAMCZAK; BED-NARSKI, 2004); e outras como esterificação de açúcares (FERNANDEZ-LORENTE et al.,2003) e glicidol (PALOMO et al., 2005).
As aplicações citadas anteriormente são frequentemente limitadas em termos deestabilidade, e também por dificuldades na recuperação e reuso do biocatalisador. A imobiliza-ção de enzimas oferece algumas vantagens operacionais em relação a enzimas solúveis, comoa escolha do tipo de processo: batelada ou contínuo, tempo reduzido de reações, controle daformação de produtos, fácil remoção da mistura reacional e adaptação de vários desenhos dereatores (ADRIANO et al., 2005). A imobilização também aumenta a estabilidade operacional,

28
O−
O
O
H
N
NH
lado acila lado álcool
Ser105
His224
Leu278
Ile285Ile189Leu140
Leu144
Figura 2.4: Sítio ativo CALB. O sítio ativo pode ser dividido em duas partes, um lado paraligações acila e a outra para ligações a álcoois, essa restrição é necessária devido a transferênciade um próton entre o oxigênio do álcool e a histidina do sítio ativo durante a catálise.
reduzindo assim custos. As técnicas de imobilização e as condições empregadas durante a imo-bilização, tipo de suporte, natureza do solvente e variações de reatores podem influenciar naspropriedades das enzimas imobilizadas. Essas condições podem ser moldadas para modificaras propriedades da enzima imobilizada para uso em determinada reação (IDRIS; BUKHARI,2011).
Existem vários métodos para imobilização de lipases, incluindo adsorção, ligaçãocovalente, aprisionamento, encapsulamento e ligaç ao cruzada (CAO, 2005). Esses métodosde imobilização têm sido empregados para melhorar a estabilidade de lipases (MATEO et al.,2007). Diversos tipos de suportes para imobilização de enzimas têm sido estudados nos últimosanos, destacando-se nanopartículas magnéticas (DUGUET et al., 2006).
2.5 Nanopartículas magnéticas de ferro
Neste contexto as nanoestruturas surgem como uma nova alternativa aos suportesconvencionais, possibilitando novas propriedades, como elevada área superficial, maior tolerân-cia a temperatura, boa reatividade química e interações fortes com as enzimas (YONG et al.,2008; LEI et al., 2009).
Magnetita (Fe3O4) é um importante minério de ferro responsável pelas proprie-dades magnéticas das rochas, sendo que muitos organismos desenvolvem esse mineral paraauxiliar na orientação geográfica. É um material ferrimagnético contendo ferro com estado deoxidação Fe2+ e Fe3+ e sua estrutura é do tipo espinélio inverso (CORNELL; SCHWERT-

29
MANN, 2003; JUNIOR, 2009).
A estrutura da magnetita Fe3O4 é formada por 32 íons de O2− regularmente orga-nizados formando uma célula unitária cúbica compacta de face centrada. Na estrutura espinélioinversa os 08 sítios tetraédricos são ocupados por Fe3+ e os 16 sítios octaédricos são ocupadospor íons Fe2+ e Fe3+; é o caso do Fe3O4. A principal diferença do Fe3O4 em relação aosdemais óxidos de ferro é a presença de ferro com diferentes valências Fe3+ e Fe2+. O Fe3O4
geralmente apresenta vacâncias, não apresentando estequiometria, e sua rigorosa formulaçãoFe3+[ Fe2+ Fe3+] O4 é reduzida para Fe3O4. O óxido é ferrimagnético à temperatura ambi-ente e paramagnético acima da TC = 850 K. Os dois diferentes íons de ferro, Fe3+ e Fe2+,ocupando os sítios octaédricos e tetraédricos formam a base de duas redes magnéticas interpe-netradas. Abaixo da TC os spins dos sítios A e B são antiparalelos, porém com magnitudesdiferentes, levando ao estado ferrimagnético (JUNIOR, 2009).
2.5.1 Produção de nanopartículas magnéticas de ferro Fe3O4
Diversos métodos tem sido utilizados para a produção de nanopartículas magnéti-cas, como o método de sonicação, pirólise e sol-gel, técnica hidrotermal, decomposição térmicaredutiva e liga mecânica. No entanto, o método de co-precipitação é um dos mais difundidose de baixo custo, devido à sua simplicidade e eficiência química para a preparação de váriosmateriais sólidos (SHI et al., 1999; KAVAS et al., 2009; BARRETO et al., 2012).
Quando Fe3O4 é produzido a partir da redução do alfa-Fe2O3 a área superficialespecífica fica em torno de 4 a 7 m2/g. Esse valor cresce para 100 m2/g para partículas de 50nm produzidas por precipitação (CORNELL; SCHWERTMANN, 2003; JUNIOR, 2009).
A presença de grupos hidroxila, tais como Fe-OH, sobre a superfícies de nano-partículas magnéticas pode permitir a ligação a compostos funcionais (XU; LIU; FINCH, 1997;NETTO; ANDRADE; TOMA, 2009), ver Figura 2.5 (adaptada de (NETTO; ANDRADE; TOMA,2009) ) (BOYER et al., 2010). A modificação da superfície das nanopartículas por silanos podeproporcionar pontos de ligação (grupos amina) para diferentes moléculas (XU; LIU; FINCH,1997). O tratamento, por exemplo com aminopropiltrietoxisilano - APTS, conduz a um reves-timento de silicato que ajuda a estabilizar as nanopartículas magnéticas contra a oxidação peloar. E os grupos amina são importantes para evitar a agregação das nanopartículas, a pH 7, epromover as interações moleculares. Uma dessas9 interações se forma com o composto glu-taraldeído, um reticulante muito utilizado para a imobilização enzimática (XU; LIU; FINCH,1997; NETTO; ANDRADE; TOMA, 2009).
2.6 Imobilização de enzimas em nanopartículas magnéticas de ferro
A elevada área superficial em relação ao volume apresentado por nanopartículasmagnéticas de ferro aumenta a capacidade de ligação e especificidade catalítica das enzimas(JOHNSON et al., 2008). O campo magnético característico dessas nanopartículas torna pos-sível uma eficiente recuperação do complexo enzimático, impedindo assim a contaminação do

30
OH
OH
OH
O iS O
O
NH2
Fe2+
2Fe3+
SOH-
O
O
O
SiNH2
+
Figura 2.5: Síntese de nanopartículas magnéticas (Fe3O4 ) e funcionalização por APTS(NETTO; ANDRADE; TOMA, 2009)
Tabela 2.2: Enzimas imobilizadas em nanopartículas magnéticas e suas aplicações biotecnoló-gicas
Enzimas Nanopartículas Aplicações ReferênciasLipase Fe3O4 Hidrólise (HUANG; LIAO; CHEN, 2008)Lipase Fe3O4 Produção de biodiesel (XIE; MA, 2009)Lipase Fe3O4 recoberta - polímero Produção de butirato de etila (CHEN; LIN, 2003)Colesterol oxidase Fe3O4 Análise de colesterol (KOUASSI et al., 2005)Queratinase Fe3O4 Síntese de queratina (KONWARH et al., 2009)Amilase Fe3O4 recoberta - celulose Degradação de amido (NAMDEO; BAJPAI, 2009)
produto final da enzima. A estabilidade enzimática é maximizada com a utilização de supor-tes em escala nanométrica, com a modulação da especificidade catalítica, baixa resistência atransferência de massa, melhorando assim a difusão e reduzindo o custo operacional (BRUNOet al., 2005). A Tabela 2.2 apresenta diversos usos de enzimas imobilizadas em nanopartículasmagnéticas (ANSARI; HUSAIN, 2012).
Diversas metodologias e suportes foram usados para a imobilização de diferentestipos de lipases em nanopartículas magnéticas. Em geral, a modificação de superfície das na-nopartículas magnéticas é necessária para o processo de imobilização (NETTO; TOMA; AN-DRADE, 2012). A Figura 2.6 mostra esquematicamente uma modificação da superfície porAPTS e reticulação com glutaraldeído, adaptada de Tang et al. 2011 (TANG et al., 2011).
Uma interessante aplicação para lipase imobilizada em nanopartículas magnéticasna indústria é a produção de biodiesel (NETTO; TOMA; ANDRADE, 2012). Dois trabalhosde pesquisa utilizaram lipase imobilizada em nanopartículas magnéticas para este propósito.Os pesquisadores Xie et al. (XIE; MA, 2009) utilizaram nanopartículas magnéticas modifi-cadas com aminopropiltrimetoxisilano (APTS) para imobilizar uma lipase (Lipozime-TL) deThermomyces lanuginosa utilizando glutaraldeído. Este biocatalisador foi aplicado para a tran-sesterificação de óleo de soja com metanol. Aproximadamente 89 % de atividade residual podeser observada quando a lipase foi imobilizada, sendo utilizada consecutivamente em reações porquatro ciclos, após isso houve um acentuado decréscimo da atividade, possivelmente devido adesnaturação da enzima imobilizada.
Outros estudos envolvendo reações catalisadas por enzimas imobilizadas em na-nopartículas magnéticas foi a reação de transesterificação de óleo de soja estudada por Xieet al. (XIE; MA, 2010). A enzima lipase (Lipozime-TL) foi ligada covalentemente a Fe3O4
utilizando 1-etil-3-(3-dimetilaminopropil) carbodiimida. A conversão observada para a transes-terificação enzimática de óleo de soja foi de 90 %. Em comparação com o outro estudo dos

31
3-aminopropiltrietoxisilano
Fe3O4 +
Aquecimento
sob atmosfera
N2
Silanização
Glutaraldeido
Imobilização
CALB
CALB
Figura 2.6: Modificação da superfície de nanopartículas magnéticas, reticulação por glutaral-deído e imobilização da enzima CALB (TANG et al., 2011).
mesmo autores já citados, após 4 ciclos, este sistema reteve 70 % da sua atividade inicial.
Um trabalho comparativo na imobilização da lipase de Burkholderia cepacia emnanopartículas magnéticas por meio de adsorção, reticulação com glutaraldeído e ativação porcarbodiimida foi realizada por Rebelo et al. (REBELO et al., 2010) e Andrade et al. (AN-DRADE et al., 2010). O melhor desempenho da lipase B. cepacia foi obtida quando foi imobi-lizada por reticulação com glutaraldeído. A enzima manteve 88 % da sua atividade inicial após10 ciclos sucessivos, para a resolução cinética de (RS)-1-(fenil)-etanol.

32
2.7 Enzimas como catalisadores heterogêneos
A catálise heterogênea em meio orgânico tem como principal vantagem a possibi-lidade de realizar reações que utilizam substratos pouco solúveis em água, como os exemploscitados na seção 2.6. Como vantagens das reações realizadas em solventes temos: a simplifica-ção dos procedimentos de recuperação do produto e do biocatalisador, a diminuição do númerode reações indesejáveis, a diminuição do risco de contaminação microbiana, a redução de pos-síveis inibições de substratos e produtos, e o aumento da estabilidade da enzima (KRIEGER etal., 2004). Contudo, solventes orgânicos possuem a tendência de remover moléculas de água dasuperfície da enzima, principalmente em sítios ativos, o que ocasionalmente as tornam inativas(AHMED; RAGHAVENDRA; MADAMWAR, 2010).
Várias abordagens têm sido propostas na literatura para contornar este problema.Entre elas se destacam a modificação química dos aminoácidos da superfície da enzima, enge-nharia de proteínas, engenharia do meio reacional (AHMED; RAGHAVENDRA; MADAMWAR,2010). Uma outra estratégia é, ao invés de se alterar a enzima aumentando sua estabilidade emsolvente, selecionar enzimas que sejam naturalmente mais resistentes a solventes para a utiliza-ção em reações de síntese em meios não aquosos (DANDAVATE et al., 2009).
As características hidrofílicas dos solventes orgânicos desempenham papel rele-vante no comportamento das enzimas. Quando hidrofóbicos, os solventes são menos nocivosàs enzimas, pois não interferem na água que matem a enzima cataliticamente ativa. Quandohidrofílicos, os solventes retêm água em seus grupos polares resultando na diminuição da ati-vidade enzimática (FABER, 2011). Por último, os solventes podem influenciar na constante deequilíbrio das reações. Esta influência pode ser utilizada para manipular o equilíbrio na dire-ção desejada. Note que o catalisador apenas alterará a velocidade na qual o sistema alcança oequilíbrio.
Em sistemas livres de solventes, onde o meio reacional é composto pelo substrato dareação, a precisão dos processos biocatalíticos se combina aos altos níveis de produtividade re-sultando em processos comercialmente competitivos (GIRALDO et al., 2007; FORESTI; FER-REIRA, 2005). Entre as vantagens das reações livres de solventes se destacam a eliminação deproblemas relacionadas a manipulação de solventes orgânicos como toxicidade e inflamabili-dade além de seus custos econômicos. Além disto, a recuperação do produto não exigem etapasde purificação ou evaporação (HE et al., 2008; SELMI et al., 1997).
Como desvantagens das reações livres de solventes encontram-se o aumento daviscosidade do meio racional (HALLING, 1990) e a redução da atividade enzimática devida adesnaturação da enzima pelo substrato presente em altas concentrações.

33
2.7.1 Atores do Processo catalisado
2.7.1.1 Água
A síntese de ésteres é um dos principais processos catalisados por lipase em meionão aquoso. Neste processo, a água desempenha forte papel na atividade catalítica e estabili-dade enzimática (HALLING, 1984). Embora a quantidade de água necessária para manter aenzima ativa em solvente orgânico seja mínima, a água pode influenciar bastante deslocando oequilíbrio nas reações de esterificação como na transesterificação (KAIEDA et al., 1999).
As lipases atuam na interface entre a fase orgânica e a fase aquosa, portanto suaatividade esta geralmente ligada a área interfacial. Neste ponto, a água atua como facilitadorada expansão da área de contato, mantendo a atividade da lipase. Por outro lado, reações lateraisindesejadas, como a hidrólise de produtos, podem resultar do excesso de água (MADALOZZO,2011). A atividade termodinâmica de água (aw) frequentemente determina a quantidade deágua em sistemas aquo-restritos. O valor ótimo para o aw varia com fatores como o tipo deenzima utilizado, o tipo de suporte, hidrofilia do suporte onde a enzima fora imobilizada (SA-LIS; SOLINAS; MONDUZZI, 2003), o solvente e cossolvente, e a polaridade e quantidade desubstratos (LU et al., 2008; JEGANNATHAN et al., 2008).
2.7.1.2 Solventes
A utilização de meios orgânicos permite a ocorrência de reações de sínteses queraramente ocorreriam em altas quantidades de água. Como já foi dito, a atividade enzimáticaé fortemente influenciada pela polaridade do solvente orgânico, que pode levar a desnaturaçãoda enzima resultando na perda de sua atividade catalítica. O principal parâmetro utilizado paramedir a polaridade do solvente e seus efeitos sobre a atividade da enzima é valor logP, ondeo coeficiente de partição P corrensponde a um padrão octanol/água do sistema de duas fases(LAANE et al., 1987).
Os efeitos de solventes orgânicos de diferentes logP na atividade de lipases foiextensivamente explorado na literatura (ESSAMRI; DEYRIS; COMEAU, 1998). Os autoresEssamri et al. estudaram o comportamento de dez solventes de polaridades distintas, comvalores de logP no intervalo de 0,91 a 3,5 na atividade da lipase R. oryzae. A lipase mantevea maior atividade naqueles solventes de menor polaridade (n-hexano, n-heptano, ciclohexanoe pentano). Outro trabalhos também mostram a vantagem dos solventes apolares (ADNANI etal., 2010; SÁNCHEZ-OTERO et al., 2010; PIRES-CABRAL; FONSECA; FERREIRA-DIAS,2010; LIU et al., 2010; LIMA et al., 2004).
A inativação da enzima pelo álcool pode decorrer da inibição competitiva, onde mo-léculas de álcool se ligam no sítio ativo da enzima (AL-ZUHAIR; LING; JUN, 2007; YADAV;DEVI, 2004). Outro fator é a desidratação ocasionada pelo solvente no entorno da moléculaenzimática que a desnatura reduzindo sua atividade catalítica (SHIMADA et al., 2002). Umaalternativa para reduzir esse efeito é a utilização de cossolventes.
Alguns estudos estão voltados a utilização de cossolventes, como o tert-butanol, que

34
aumentem a solubilidade do metanol ou etanol no meio reacional, pois estes são pouco solúveisem sistemas orgânicos, ocasionando a inativação da lipase de forma mais aguda (TAN et al.,2010). Os autores Royon et al. (ROYON et al., 2007) estudaram a lipase imobilizada Candidaantarctica B (Novozyme® 435) como catalisador em solvente tert-butanol para produção debiodiesel a partir do óleo de algodão. Neste trabalho a inibição da lipase causada pelo metanolfoi anulada pela adição de tert-butanol no meio reacional aumentando a velocidade de reação erendimento em éster (97%), enquanto que no meio livre de solvente não houve conversão eméster, resultado da total inibição da lipase pelo metanol.
Outros solventes como o 1,4-dioxano e líquidos iônicos foram pesquisados comoco-solventes para a produção de biodiesel catalisada por lipase reduzindo significativamente ainativação da lipase (HA et al., 2007; ISO et al., 2001). Porém, a dificuldade de se recuperar obiocatalisador do meio reacional torna o emprego destes solventes menos competitivos (HA etal., 2007).
2.7.1.3 Temperatura
A temperatura pode influenciar na velocidade da reação catalisada por dois meca-nismos: o mecanismo químico, onde o aumento da temperatura e o consequente aumento daenergia cinética dos reagentes resultam na aceleração do processo, e o mecanismo de desnatura-ção da enzima pelo aumento da temperatura (AL-ZUHAIR; LING; JUN, 2007; NOUREDDINI;GAO; PHILKANA, 2004; FUKUDA; KONDO; NODA, 2001; MACRAE, 1983). A tempera-tura ótima seria aquela onde a razão entre a desnaturação das enzimas e a energia cinética é amaior possível, ou seja, a temperatura é baixa o suficiente para não desnaturar um quantidadesignificativa de enzimas, ao mesmo tempo em que é alta o suficiente para acelerar o processoreacional. Esta temperatura ótima foi determinada pelos trabalhos acima citados e é em tornodos 40 °C variando de acordo com a enzima utilizada e de suas características de atividade eestabilidade em solventes orgânicos.
Estudos apontam que a estabilidade térmica da lipase imobilizada é maior do que daenzima livre porque o suporte evita a transição da conformação da enzima em altas temperaturas(KIM et al., 2006). No estudo apresentado por Yiungitoglu et al. (YIGITOGLU; TEMOÇIN,2010) foi verificado o efeito da temperatura na atividade da lipase de Candida rugosa na formalivre e imobilizada, constatando que após 180 min na temperatura de 50 °C houve maior esta-bilidade por parte da enzima imobilizada que se manteve com 54% da atividade de hidrólise,sendo que a enzima na forma livre refere apenas 30% da atividade residual. Foi relatado que oaumento da estabilidade térmica foi uma das vantagens gerais das enzimas imobilizadas.
2.8 Ésteres
Os ésteres de ácido graxos são encontrados na indústria em produtos como aromas,sabões, medicamentos, perfumes e cosméticos, na produção de biocombustíveis, entre outros.Asíntese de ésteres baseia-se na esterificação de ácidos graxos com álcool na presença de ca-talisadores inorgânicos a elevadas temperaturas (+100°C). Estes processos demandam grande

35
quantidade de energia, além de serem lentas e não seletivas. Com o crescente aumento dabusca por ésteres para o emprego em aditivos e aromas na indústria de perfumes, torna-se ne-cessário encontrar novas formas mais eficientes de extração de aromas (KUMAR; MODAK;MADRAS, 2005; ABBAS; COMEAU, 2003). Neste contexto, o emprego de enzimas comobiocatalisadores permitem a obtenção de produtos de melhor qualidade, em temperaturas re-lativamente baixas e portanto sendo rotuladas de naturais pela indústria (KISS et al., 2004;WELSH; WILLIAMS; DAWSON, 2006).
As lipases também são empregadas como biocatalisadores em reações de modifica-ção de óleos e gorduras na produção de lipídeos estruturados (UNDURRAGA; MARKOVITS;ERAZO, 2001; PABAI; KERMASHA; MORIN, 1995). Lipídios estruturados são produtos damodificação de um triacilglicerol, obtidos da transesterificação química ou enzimática, com a fi-nalidade de alterar a composição em ácidos graxos e/ou sua distribuição posicional na moléculade glicerol. Estes compostos são sintetizados na intenção de se melhorar características físicascomo: polimorfismo, ponto de fusão, conteúdo de gordura sólida, viscosidade e consistência, epropriedades nutricionais como: quantidade de ácidos graxos saturados ou insaturados de fácilabsorção e digestão (D’AGOSTINI; GIOIELI, 2010).
Os ésteres metílicos e etílicos, de ácidos graxos de cadeia longa como os oleatos,palmitatos, ricinoleatos e linolenatos são os principais constituintes do biodiesel. O biodieselpode ser obtido por processos de esterificação ou transesterificação. O método mais comumpara a produção do biodiesel é a transesterificação de óleos vegetais com alcoóis de cadeiacurta, como o metanol, na presença de catalisadores (MARCHETTI; MIGUEL; ERRAZU,2007). Dificuldades nos procedimentos de remoção do catalisador, recuperação do glicerol,alto dispêndio de energia, reações laterais indesejáveis e alto custo de refino tornam o métodode síntese química de biodiesel dispendioso (OGNJANOVIC et al., 2009).
A produção biotecnológica do biodiesel com lipase tem estado no foco de váriosestudo recentes e tem tido um rápido desenvolvimento, dado ao interesse de se adotar proces-sos ecologicamente aceitáveis (RANGANATHAN; NARASIMHAN; MUTHUKUMAR, 2008;SHIMADA et al., 2002). Com o emprego de lipases como biocatalisadores nas reações de tran-sesterificação se pode ter um processo que ocorre em temperaturas mais brandas, com fácilrecuperação do glicerol, sem produção de resíduos químicos, sem produção de sabões, queocorre em processos químicos que utilizam catalizadores básicos. Além disto, se a lipase es-tiver na forma imobilizada, há a possibilidade de se reutilizar o biocatalisador (DIZGE et al.,2009; NOUREDDINI; GAO; PHILKANA, 2004).
As lipases podem promover diversos tipos de reações em condições brandas e sele-tivas, destacando-se a produção de ésters. No presente estudo, a síntese enzimática dos ésters:oleato de etila, butirato de etila e metila foram estudadas. Oleato de etila é um éster usadocomo biodiesel, aditivo de combustível diesel e nas indústrias de cosméticos e de alimentos(FORESTI; FERREIRA, 2005; BLOOMER; ADLERCREUTZ; MATTIASSON, 1992; KA-NASAWUD et al., 1992). Atualmente, a produção comercial de biodiesel é por via química, oque passa por várias etapas, encarecendo o produto final, assim a demanda por processos enzi-máticos é crescente devido a redução das etapas de produção e a possibilidade de obtenção deprodutos que causam menores riscos ao meio ambiente (ROSSET, 2011).

36
A lipase de Candida antartica do tipo B utilizada neste trabalho é uma das lipasesmais empregadas em estudos para a produção de biodiesel. Essa lipase de origem microbiana émuito usada em reações para produção de alcoóis secundários enantiomericamente puros e emtransformações de ácidos carboxílicos (KIRK; CHRISTENSEN, 2002).
Ésteres de baixo peso molecular de ácidos carboxílicos e álcoois também tem de-sempenhado uma importante função na indústria, sendo usados como constituintes de sabor earomas (GROSSO; FERREIRA-DIAS; PIRES-CABRAL, 2012; LIAQUAT; APENTEN, 2008).Um dos exemplos é o butirato de etila e metila, importantes componentes de aromas de mui-tas frutas, por exemplo: abacaxi, maracujá, morango e maça (GROSSO; FERREIRA-DIAS;PIRES-CABRAL, 2012; LIAQUAT; APENTEN, 2008; RODRIGUEZ-NOGALES; ROURA;CONTRERAS, 2005).

37
3 ESTUDOS PRELIMINARES
Experimentos preliminares foram realizados com o objetivo de estudar o sistema deimobilização e os materiais disponíveis como suportes e escolher a melhor metodologia de imo-bilização. Inicialmente avaliou-se a produção de nanopartículas de quitosana e posteriormentea imobilização de lipase do tipo B de Candida antarctica (CALB) em nanopartículas mag-néticas e hibridas (nanopartículas magnéticas recobertas com quitosana) para se estabelecer ametodologia e a forma de condução dos processos de imobilização.
3.1 Materiais
Nanopartículas magnéticas (Fe3O4) e hibridas foram gentilmente cedidas pelo Grupode Química de Materiais Avançados (GQMAT), Departamento de Química Analítica e Físico-Química, Universidade Federal do Ceará, UFC. As nanopartículas magnéticas foram produ-zidas pelo método de co-precipitação (BARRETO et al., 2012) com tamanho de partícula de11,0 nm. Quitosana em pó (grau de desacetilação de 85,9%) foi adquirida junto a POLYMARIND LTDA, Fortaleza, Ceará. Lipase do tipo B de Candida antarctica (CALB) referenciadacomo Lipozyme CALB L® foi obtida da Codexis (Redwood, USA). Resinas acrílicas comer-ciais de lipase de Candida antarctica, Novozym 435 (lipase do tipo B de Candida antarcticaimobilizada em resina acrílica), γ-aminopropiltriethoxisilano (APTS), solução de glutaraldeidode grau II 25% (m/v), p-nitrophenil butirato (pNPB) and p-nitrophenil (pNP)foram obtidos daSigma-Aldrich (St. Louis, USA). Os demais reagentes de grau analítico foram obtidos da Synth(São Paulo, Brasil) e Vetec (São Paulo, Brasil).
3.2 Métodos
3.2.1 Preparação de nanopartículas de quitosana
Nanopartículas de quitosana foram produzidas por adição de tripolifosfato de sódioà solução de quitosana, mantendo-se a agitação, de acordo com metodologia apresentada pelosautores (QI et al., 2004). As concentrações de quitosana (0,1; 0,25 e 0,5 % p/v) e tripolifosfatode sódio (0,25 e 0,7 mg/ml) foram analisadas.
3.2.2 Preparação de nanopartículas magnéticas de ferro
Nanopartículas magnéticas de ferro (Fe3O4) foram produzidas pelo método de co-precipitação (BARRETO et al., 2012). Determinou-se o tamanho (11 nm) das nanopartículasatravés da técnica de Difração de Raios-X (DRX) com posterior refinamento das fases obtidaspelo método Rietveld. Espectros de infravermelho foram obtidos para análise de presença dehidroxilas usando pastilhas de KBr das ferritas magnéticas. O espectro foi medido na regiãoentre 400 e 4000 cm−1 (BARRETO et al., 2012).

38
3.2.3 Tratamento do suporte com aminopropiltrietoxisilano (APTS)
O tratamento dos suportes (nanopartículas magnéticas e hibridas) consistiu da adi-ção de solução de APTS (0,5 %v/v) a pH 3,3, temperatura de 75°C, por 3 horas, na razãolíquido-sólido de 3,0 (ml/g de suporte), em agitador recíproco (banho-maria). Seguiu-se o pro-cedimento de lavagem: inicialmente com água destilada e depois com acetona (ZANIN, 1989).Utilizaram-se três soluções de acetona com diferentes concentrações, 20 %, 50 % e pura, paraacelerar o tempo de secagem. O material foi seco em estufa até que se obtivesse massa constante(PEREIRA, 1996).
3.2.4 Ativação do suporte com Glutaraldeído
Após o tratamento com APTS, a ativação dos suportes foi realizada com solução deglutaraldeído 0.6% (v/v) por 2h a 25°C. Após a ativação, as nanopartículas foram lavadas comtampão bicarbonato-carbonato de sódio, 100mM, pH 10,0, para retirada do excesso do agenteativador, de acordo com metodologia apresentada por (SILVA et al., 2012).
3.2.5 Imobilização Enzimática
A lipase do tipo B de Candida antarctica solúvel foi imobilizada em nanopartículasmagnéticas e nanopartículas magnéticas recobertas por quitosana, sendo tradadas por APTS eativadas por glutaraldeído 0,6% na proporção de 200,0 UpNPB de enzima por grama de suporte.A imobilização foi realizada em presença do tampão bicarbonato 100mM, pH 10,0 (razão m/vde 1/10) a 25 °C e tempo de incubação de 5 h, sob agitação controlada (20 rpm), de acordo commetodologia apresentada por (SILVA et al., 2012).
3.2.6 Medida da atividade enzimática
A atividade hidrolítica da CALB solúvel e imobilizada (derivado) (Atd) foi determi-nada usando butirato de p-nitrofenol (pNPB) como substrato, a pH 7,0 e 25°C, de acordo com ametodologia descrita na literatura (KORDEL et al., 1991), com modificações. Uma unidade deatividade (1 U) enzimática foi definida como sendo a quantidade de enzima capaz de hidrolisar1 µmol de pNPB por minuto, de acordo com metodologia descrita por (KORDEL et al., 1991),com modificações.
3.2.7 Parâmetros de imobilização
3.2.7.1 Rendimento de imobilização
Após a medida inicial de atividade (Ati) e da atividade da enzima final (At f ) nosobrenadante e no ensaio em branco, a atividade enzimática remanescente no sobrenadante

39
(Ati - At f ) e o rendimento de imobilização (IY) pode ser calculada, utilizando a equação 3.1(KORDEL et al., 1991; SILVA et al., 2012).
IYimob(%) =Ati−At f
Ati∗100 (3.1)
3.2.7.2 Atividade recuperada (Atr)
A atividade teórica (Att), equação 3.2, imobilizada no suporte pode ser calculadautilizando a quantidade de enzima oferecida por grama de suporte (Ato f f - U / g de suporte) e orendimento de imobilização, equação 3.1. Após a medição da atividade da enzima imobilizada(Atd - U / g de suporte), a atividade recuperada (MENDES et al., 2011; SILVA et al., 2012) foicalculada utilizando a equação 3.3.
Att = IY ∗Ato f f (3.2)
Atr =AtdAtt∗100 (3.3)
3.3 Resultados e discussões
3.3.1 Nanopartículas de quitosana
Nanopartículas de quitosana foram produzidas por adição de tripolifosfato de sódioà solução de quitosana, mantendo-se a agitação. As concentrações de quitosana (0,1; 0,25 e0,5 % p/v) e tripolifosfato de sódio (0,25 e 0,7 mg/ml) foram analisadas. Observou-se que namenor concentração de quitosana, obteve-se o melhor resultado em relação a precipitação denanopartículas. A concentração de 0,7 mg/ml de tripolifosfato de sódio apresentou a melhorcondição em relação a precipitação. Após o processo de produção, dificuldades foram veri-ficadas na separação das nanopartículas do meio reacional, tendo-se significativas perdas porcentrifugação, assim não foi possível recuperar nanopartículas de quitosana para posterior usona imobilização da enzima CALB no suporte. Deste modo, estudou-se o uso de outros suportesque permitissem uma melhor separação do suporte do meio reacional: nanopartículas magnéti-cas e nanopartículas magnéticas recobertas por quitosana foram analisadas por possibilitar umaeficiente separação magnética e modificação da superfície para a imobilização enzimática.

40
3.3.2 Caracterização das nanopartículas magnéticas
Nanopartículas magnéticas de ferro (NPM) com diâmetro de 11 nm foram obtidos(dados obtidos por DRX das nanopartículas). A Figura 3.1 mostra o espectro de infravermelhodas nanopartículas utilizadas neste trabalho. Observamos picos característicos da ligação Fe-O em 580 cm−1 para Fe3O4, que corresponde ao estiramento vibracional no sítio tetraédricodas ferritas (HRDINA et al., 2010). A banda em 1637 cm−1 foi observada e é referente àvibração angular da hidroxila (O-H). Estes grupos apareceram no espectro devido à síntesedas ferritas magnéticas serem realizadas em meio aquoso, logo a superfície destes materiaisficaram encobertas por estes grupos. Também a presença de hidroxilas foi observada em 3411cm−1 para Fe3O4, referente ao estiramento da ligação O-H.
Figura 3.1: Espectro de infravermelho: Nanopartículas magnéticas de ferro.
3.3.3 Imobilização em nanopartículas magnéticas
Os resultados obtidos com os dois suportes magnéticos (nanopartículas magnéticasde ferro e nanopartículas magnéticas recobertas por quitosana) estão apresentados na Tabela3.2. O suporte hibrido apresentou rendimento de imobilização de 3,1 % e atividade do derivadode 0,016 UpNPB/g, sendo a atividade recuperada de 0,27 %. A imobilização em nanopartículasmagnéticas possibilitou uma maior atividade da enzima imobilizada, 0,56 (± 0,01) UpNPB/g erendimento de imobilização de 19,6%. Este comportamento pode ser explicado pelo tratamentode silanização com aminotrietoxisilano proporcionar um revestimento de silicato estável for-mando uma fortes ligações Fe-O-Si e Si-O-Si, levando a grupos disponíveis para interagir comespécies externas, como glutaraldeído (ANDRADE et al., 2010). A função do glutaraldeído épara reagir com o grupo amino do APTS, formando uma ligação imina e deixando um terminalaldeido para reagir com os resíduos de aminoácidos da enzima (ANDRADE et al., 2010).
O recobrimento com quitosana pode ter comprometido a ativação por APTS devidoa elevada concentração de grupos amina em quitosana (grau de desacetilação, 85,9 %) permitiruma elevada reticulação com o glutaraldeído. As reações envolvidas na reticulação por agentesbifuncionais ocorrem entre os grupos amino e hidroxilas da quitosana. O grau de desacetilaçãoe a massa molecular da quitosana são importantes parâmetros, porque o aumento da massamolecular e do grau de desacetilação também resulta em um maior rendimento de reticulação.Este efeito também pode prejudicar a difusão de macromoléculas como as proteínas, devido àformação de poros de pequenos diâmetros (GUPTA; JABRAIL, 2006).

41
Avaliou-se ainda a possibilidade do suporte catalisar a hidrólise do pNPB, levando aum falso positivo. Para tanto, nanopartículas magnéticas modificadas por APTS na ausência deenzima imobilizada foram utilizadas para determinação da atividade enzimática, não se obtendonenhuma atividade catalítica. Este resultado mostra que as atividades catalíticas provenientesdos biocatalisadores com enzima imobilizada provêm apenas do sítio catalítico enzimático enão do metal presente nas nanopartículas.
A partir dos resultados iniciais obtidos (Tabela 3.2), observaram-se pontos a seremotimizados, como por exemplo, o uso de aditivos para obtenção de uma melhor conformaçãoenzima-suporte.
Tabela 3.1: Parâmetros de imobilização
Parâmetros de imobilização Nanopartículas magnéticas Nanopartículas magnéticas:quitosanaCarga oferecida (U/g) 200,0 ± 1,41 200,0 ± 1,41Atividade remanescente (U/g) 160,8 ± 1,01 193,8 ± 0,24Rendimento de imobilização R (%) 19,6 ± 0,83 3,1 ± 2,90Atividade teórica (U/g) 22,7 ± 0,08 6,2 ± 5,8Atividade aparente (U/g.gel) 0,56 ± 0,01 0,016 ± 0,08Atividade recuperada (%) 2,5 ± 0,13 0,27 ± 0,13
3.3.4 Influência de aditivos
Visando uma melhora nos parâmetros de imobilização, analisou-se a influência dosaditivos: triton-X (0,0015 % v/v) e biodiesel (0,015 % v/v) na imobilização enzimática. Apresença destes aditivos não possibilitou uma eficiente ligação da enzima ao suporte, assim osresultados foram semelhantes aos apresentados na tabela 3.2, para ambos os suportes. O efeitode surfactantes é mais acentuado em enzimas com estruturas que bloqueiam o sitio ativo, assimas moléculas de surfactantes, por terem caráter anfótero permitem a abertura da mesma, poréma enzima CALB estudada não tem esse impedimento acentuado, assim não sofre aumento dasua atividade catalítica por ativação interfacial (FERNANDEZ-LAFUENTE et al., 1998).
A continuação do estudo de otimização é iniciada com a modificação da metodolo-gia de ativação por APTS para nanopartículas magnéticas de ferro. A reação foi conduzida sobatmosfera de nitrogênio a fim de proteger as nanopartículas contra a oxidação pelo ar e reduzira possível interferência de hidróxidos de oxido de ferro no processo catalítico (ANDRADE etal., 2010).
3.4 Resultados e discussão
3.4.1 Influência do suporte
O suporte hibrido ativado com solução de glutaraldeído apresentou rendimento deimobilização de 23,7 % (± 2,01) e atividade recuperada de 0,91 % (± 0,36). Os resultados ob-tidos com o suporte híbrido e magnetita de ferro são apresentados na Tabela 3.2. Os resultados

42
Tabela 3.2: Parâmetros de imobilização
Parâmetros de imobilização Nanopartículas magnéticas Nanopartículas magnéticas:quitosanaCarga oferecida (U/g) 200,0 ± 1,41 200,0 ± 1,41Atividade remanescente (U/g) 160,8 ± 1,01 193,8 ± 0,24Rendimento de imobilização R (%) 19,6 ± 0,83 3,1 ± 2,90Atividade teórica (U/g) 22,7 ± 0,08 6,2 ± 5,8Atividade aparente (U/g.gel) 0,56 ± 0,01 0,016 ± 0,08Atividade recuperada (%) 2,48 ± 0,13 0,27 ± 0,13
apresentados demonstram que as nanopartículas magnéticas recobertas por quitosana (híbridas)não apresentaram bom desempenho, em termo de atividade recuperada 0,27 % (±0,13), quandocomparadas às nanopartículas não recobertas, 2,48 % (± 0,13). Este comportamento pode serexplicado pela menor quantidade de grupos hidroxila presentes no suporte hibrido, que podemreagir com APTS, devido ao recobrimento do suporte com quitosana.
3.4.2 Influência do surfactante
Visando uma melhora nos parâmetros de imobilização, analisou-se a influência dossurfactantes Triton-X (0,0015 % v/v) e Biodiesel (0,015 % v/v) na imobilização enzimática. Apresença destes surfactantes não possibilitou uma eficiente ligação da enzima ao suporte, assimos resultados foram semelhantes aos apresentados na tabela 3.2, para ambos os suportes.
A ativação com APTS foi modificada com o objetivo de evitar a influência do oxigê-nio nas propriedades magnéticas das nanopartículas. Para tal fim, utilizou-se uma metodologiade modificação por APTS em presença de atmosfera de nitrogênio. A metodologia consistiuna adição de solução de APTS (2 %) às nanopartículas magnéticas na proporção 0,2 mL/mgde suporte. A suspensão foi aquecida sob atmosfera de nitrogênio a 100 ºC por 10 horas. Asolução foi lavada com metanol e etanol, e então as nanopartículas magnéticas funcionalizadasforam secas em estufa a 30 °C por 24 horas. Após, o suporte foi ativado utilizando glutaral-deído 25 % (v/v) por 2 horas. Posteriormente, 20mg do suporte foi adicionado em 1,0 mL desolução enzimática (0,60mg lipase/mL), a mistura foi agitada a 20, 45 e 250 rpm por 1 hora(ANDRADE et al., 2010).
Avaliou-se ainda a possibilidade do suporte catalisar a hidrólise do pNPB, levando aum falso positivo. Para tanto, nanopartículas magnéticas modificadas por APTS na ausência deenzima imobilizada foram utilizadas para determinação da atividade enzimática, não se obtendonenhuma atividade catalítica. Este resultado mostra que as atividades catalíticas provenientesdos biocatalisadores com enzima imobilizada provêm apenas do sítio catalítico enzimático enão do metal presente nas nanopartículas.
Analisou-se também a atividade catalítica das nanopartículas após a modificaçãopor APTS, não houve atividade do biocatalisador, mostrando assim a importância da ativaçãopor glutaraldeído.

43
3.5 Produção de ésteres
Os capítulos que seguem são compostos por dois artigos a serem submetidos a pe-riódicos internacionais. O primeiro é a respeito da produção de oleato de etila e o segundo é aprodução de butirato de metila e etila catalisada pela enzima CALB imobilizada em nanopartí-culas magnéticas de ferro.

44
4 PRODUÇÃO DE OLEATO DE ETILA
Síntese de oleato de etila catalisada por lipase do tipo B de Candida antarcticaimobilizada em nanopartículas magnéticas
Maria Cristiane Martins de Souza, Kímberle Paiva dos Santos, Rafael MeloFreire, Antonio César Honorato Barreto, Pierre Basílio Almeida Fechine e Luciana RochaBarros Gonçalves
Abstract
The iron magnetic nanoparticles (Fe3O4) were evaluated as support in the immo-bilization of Candida antarctica lipase type B (CALB). The biocatalyst was analyzed in theesterification reaction for the ethyl oleate production. The nanoparticles were produced by theco-precipitation method. Modifications were carried out on the nanoparticles’ surfaces withaminopropyltriethoxysilane (APTS) and glutaraldehyde. The influence of stirring speed (20-250 rpm), enzyme load (50-650 UpNPB/gsupport), contact time (0.5-5 h), glutaraldehyde con-centration (2.5 and 25%) and presence of additive (SDS 0.23%) during immobilization wereevaluated. The immobilization was performed on the presence of 100mM bicarbonate buffer,pH 10, at 25 °C. After immobilization, CALB exhibited improved thermal and operational sta-bilities. The best result (Immobilization yield: 53% and immobilized enzyme activity: 29.1UpNPB/gsupport) was obtained at 45 rpm, using 200 UpNPB/gsupport and 1h of immobiliza-tion. The ethyl oleate production was analyzed with the best condition of immobilization andcompared to the results obtained when reaction was catalyzed by CALB immobilized on acrylicresin (commercial). The conversion of oleic acid in ethyl oleate was approximately 90 % for thethree biocatalysts evaluated at 48 h. The consecutive reaction cycles (14) show the maintenancein the production of biodiesel (approximately 74 % for nanoparticles and 99 % for acrylic resin).
Keywords: Lipase , Magnetic nanoparticles , Immobilization , Biodiesel

45
Resumo
Nanopartículas magnéticas de ferro (Fe3O4) foram avaliadas como suporte na imo-bilização de lipase de Candida antarctica do tipo B (CALB). A catálise foi analisada na reaçãode esterificação para a produção de oleato de etila. O catalisador foi testado em sistema cata-lítico livre de solvente para a catálise de oleato de etila a partir de ácido oleico e etanol. Asnanopartículas foram produzida pelo método de co-precipitação. Modificações foram realiza-das na superfície das nanopartículas com γ-aminopropiltrietoxisilano (APTS) e glutaraldeído.A influência da velocidade de agitação (20-250 rpm), carga enzimática (45-200 UpNPB.g−1
suporte), tempo de contato (0,5 - 5h), concentração de glutaraldeído (2,5 e 25% (m/v)) e pre-sença do aditivo dodecil sulfato de sódio (SDS, 0,23%) foi observada durante a imobilização. Aimobilização foi realizada na presença de tampão bicarbonato de sódio 100mM, pH 10, a 25 °C.Após a imobilização, CALB imobilisada em nanopartículas magnéticas (CALB-NPM) mostroumelhor estabilidade térmica e operacional do que a forma solúvel da enzima. O melhor resul-tado (Rendimento de imobilização: 53% e atividade da enzima imobilizada: 29,1 .g−1 suporte)foi obtida a 45 rpm, usando 80 UpNPB/g suporte e 1 hora de imobilização. A produção deoleato de etila foi analisada sob a melhor condição de imobilização e comparada com os resul-tados obtidos quando a reação foi catalisada por CALB imobilizada em resila acrílica comercial(Novozym® 435). A conversão de ácido oleico em oleato de etila foi de aproximadamente 90% para os dois catalisadores (CALB-NPM e Novozym® 435) utilizados a 48 horas. Os ciclosde reação consecutivos (14) mostram a manutenção da produção de biodiesel (oleato de etila)(aproximadamente 74 % para CALB-NPM e 99 % para Novozym® 435).
Palavra-chave: Lipase , nanopartículas Magnéticas , imobilização , Biodiesel

46
4.1 Introdução
Os esteres de ácidos graxos, especialmente os ésteres de metila e etila, como olea-tos, palmitatos, ricinoleatos e linolenatos são os principais componentes do biodiesel. Oleato deetila, por exemplo,é um éster graxo utilizado como biodiesel, aditivo para diesel combustível ena industria cosmética e alimentícia (MARCHETTI; ERRAZU, 2008). Biodiesel pode ser ob-tido por meio de processos de esterificação e transesterificação (XIE; MA, 2009). Atualmente,a produção industrial de biodiesel é realizada por meio de catalise quimicamente homogênica.Algumas bases, tais como potássio e hidróxido de sódio ou seus metóxidos, são comumente em-pregados na transesterificação de óleos vegetais com metanol para produzir ésteres de ácido gra-xos metílicos (XIE; MA, 2009). A catálise química não é aceita como “verde” devido a sériosproblemas relacionados a remoção do catalisador do meio reacional, uso excessivo de energia,dificuldade na recuperação do glicerol, reações laterais indesejáveis e alto custo para a purifi-cação do produto (OGNJANOVIC et al., 2009). Portanto, a produção por catálise enzimáticatem recebido significante atenção e tem se desenvolvido rapidamente devido a necessidade dese adotar processos ambientalmente sustentáveis (SHIMADA et al., 1999; RANGANATHAN;NARASIMHAN; MUTHUKUMAR, 2008). A produção mono-ésteres de ácidos graxos (bio-diesel) tem sido amplamente estudado por diversos autores, principalmente utilizando lipase deCandida antartica do tipo B imobilizada em rezina acrílica (Novozym® 435) (ADACHI et al.,2012; FEDOSOV et al., 2012; TAN et al., 2010; DU et al., 2004; SAMUKAWA et al., 2000;SHIMADA et al., 2002; TALUKDER et al., 2009).
Lipase produzidas por Candida sp. são enzimas bem estabelecidas para propósitode catálise, especialmente lipase de Candida antarctica do tipo B (CALB) (MARIA et al.,2005). A enzima CALB tem sido estuda para potenciais aplicações na industria de alimentos,detergentes, fármacos, tecidos, cosmético, papel e óleo-química (HASAN; SHAH; HAMEED,2006). Essas aplicações são possíveis devido a ampla gama de especificidade para substratos,alta atividade sob condições reacionais brandas, estabilidade em solventes orgânicos, alta es-tabilidade térmica e estereoespecificidade (RODRIGUES et al., 2008; MCCABE; RODGER;TAYLOR, 2005; DENG et al., 2011).
Materiais alternativos aos suportes comerciais estão sendo estudados para atribui-ção de novas propriedades que permitem maior resistência, magnetismo, elevada superfíciede contato e redução de impactos ambientais. Dentre estes materiais destacam-se os mate-riais nano-estruturados. As dimensões destas estruturas são comparadas as de vírus (20-500nm), a proteínas (5-50 nm) ou a genes (2 nm largura e 10-100 nm comprimento) (TARTAJet al., 2005). A ampla superfície das nanopartículas é propícia a modificação para ligaçõescom enzimas (TARTAJ et al., 2005). Os grupos presentes na superfície da mesma, grupos hi-droxilas, podem definir pontos de ligações para agentes funcionalizantes (BARRETO et al.,2012; BOYER et al., 2010). Além disso, nanopartículas magnéticas apresentam o fenômenode superparamagnetismo, ou seja, eles não permanecem magnetizados após a ação do campomagnético, tendo-se assim a vantagem da redução do risco de agregação das partículas (RUIZ-HERNANDEZ et al., 2008). As nanopartículas magnéticas que são usadas em bio-aplicaçõessão usualmente fabricadas de materiais biocompatíveis, como magnetita (Fe3O4) (SHAW et

47
al., 2006). Estas propriedades permitem o uso eficiente como suporte para imobilização en-zimática (ASHTARI et al., 2012; TRAN; CHEN; CHANG, 2012; ANDRADE et al., 2010;NETTO; ANDRADE; TOMA, 2009; KIM; JIA; WANG, 2006; BAI et al., 2006). O catalisador(nanopartícula-enzima) pode ser facilmente recuperado por campo magnético, sendo possível adiminuição de custos operacionais e o aumento da pureza dos produtos obtidos (JIANG et al.,2009).
Neste trabalho, a imobilização da enzima CALB em nanopartículas magnéticas deferro foi avaliada. A influência das condições operacionais como: velocidade de agitação, cargaenzimática, tempo de contato e presença de aditivos durante a imobilização foram analisados.O catalisador foi testado em sistema catalítico livre de solvente para a catálise de oleato deetila a partir de ácido oleico e etanol. Etanol apresentou mais vantagens do que o metanol, porser considerado um álcool “verde”, enquanto o metanol pode ser considerado potencialmentetóxico (AKOH et al., 2007). Os resultados obtidos com CALB imobilizados em nanopartículasmagnéticas de ferro (CALB-NPM) foram comparadas com a CALB imobilizada em resinaacrílica (Novozym® 435).
4.2 Materiais e métodos
4.2.1 Materiais
Nanopartículas magnéticas de ferro (NPM) (Fe3O4) foram produzidas pelo mé-todo de co-precipitação (BARRETO et al., 2012). Lipase de Candida antarctica do tipo B(CALB) foi adquirida da Codexis (Redwood, USA). A CALB imobilizada em resina acrílica(Novozym® 435), γ-aminopropiltrietoxisilano (APTS), solução de glutaraldeído grau II 25%(m/v), butirato de p-nitrofenila (pNPB) e p-nitrofenila (pNP) foram obtidos da Sigma-Aldrich(St. Louis, USA). Os demais reagentes de grau analítico foram obtidos da Synth (São Paulo,Brasil) e Vetec (São Paulo, Brasil).
4.2.2 Síntese de (Fe3O4)
Nanopartículas magnéticas de ferro (Fe3O4) foram produzidas pelo método de co-precipitação (BARRETO et al., 2012). Difração de raio-X (XRD) e transmissão eletrônica devarredura (TEM) foram realizadas para determinação do tamanho das nanopartículas (11,0 nm)(BARRETO et al., 2012).
4.2.3 Tratamento do suporte com γ-aminopropiltrietoxisilano (APTS)
A reação foi iniciada com a adição de solução de APTS (2,0% v/v) ao suportena proporção de 0,2 (ml/mg de suporte). A solução foi aquecida a 100°C por 10 horas sobatmosfera de nitrogênio. As nanopartículas foram lavadas com 100 ml de metanol e 100 ml deetanol, separadas por magnetismo, em seguida o material foi seco a 30°C por 24 horas (NETTO;ANDRADE; TOMA, 2009).

48
4.2.4 Reticulação com solução de glutaraldeído
NPM (0,01g) foram suspendidas em solução 25% (m/v) de glutaraldeído na propor-ção de 2,5 (solução (ml)/suporte (g)). A mistura foi mantida sob agitação (45 rpm) por 2 horasà 25 C. Finalmente o suporte foi lavado com tampão bicarbonato de sódio 100 mM, pH 10,0para remover o excesso de glutaraldeído (ANDRADE et al., 2010).
4.2.5 Imobilização
O processo de imobilização foi realizado pelo contato de 0,01 g de nanopartículas(tratada por APTS e reticulada com solução de glutaraldeído) com 0,5 ml de solução tampão debicarbonato de sódio, 100 mM, pH 10,0. O sistema foi mantido sob agitação controlada: 20-45rpm (agitação rotacional) e 20-250 rpm (agitação orbital). O tempo de contato enzima-suportefoi estudado de 0,5-5 h. A imobilização em presença de surfactante SDS também foi analisada(0,23 %). A enzima CALB imobilizada foi removida da solução por magnetismo. Medidas deatividade da enzima solúvel e imobilizada foram realizadas de acordo com procedimento des-crito a seguir. A quantidade de CALB imobilizada em NPM foi determinada através da mediçãoda concentração inicial e final do CALB no sobrenadante de imobilização. Atividades hidro-líticas também foram determinadas para permitir o cálculo dos parâmetros de imobilização:rendimento de imobilização (R), atividade teórica (Att) e atividade recuperada (Atr ) (SILVA etal., 2012).
As medidas de atividade hidrolítica da CALB solúvel e imobilizada Atd foram de-terminadas utilizando pNPB como substrato, em 2-propanol, pH 7,0 e 25°C, de acordo com ametodologia descrita em literatura (KORDEL et al., 1991), com algumas modificações. Umaunidade da atividade da lipase foi definida como a quantidade de enzima capaz de liberar 1µmol de p-nitrofenila por minuto.
4.2.6 Concentração de proteínas
O método de Bradford (BRADFORDMM, 1976) foi utilizado para a análise daconcentração de proteínas.
4.2.7 Estabilidade térmica da CALB solúvel e imobilizada
A estabilidade térmica a 60°C da CALB imobilizada e livre foi examinada pela me-dição da atividade residual pela hidrólise de (pNPB) previamente descrita. Solúvel ou CALBimobilizada foi incubada em tampão fosfato de sódio 25 mM, pH 7, 60°C, por 48 horas. Amos-tras foram periodicamente analisadas e suas atividades residuais foram examinadas. A constantede desativação e tempo de meia-vida (t1/2) foram calculadas de acordo com o modelo de Sa-dana e Henley Fernandez-Lafuente et al. (1993) (FERNANDEZ-LAFUENTE et al., 1993). Osfatores de estabilização foram obtidos como a razão entre o tempo de meia-vida dos derivadose a CALB solúvel (SILVA et al., 2012).

49
4.2.8 Síntese de oleato de etila
Os experimentos de esterificação foram executados em microtubos plásticos (2,0mL) contendo 0,6 de ácido oleico e 151 µL de etanol (razão molar 1:1), bem como 0,01 g debiocatalisador CALB-NPM (1,7% de massa reacional). A reação foi conduzida em agitação or-bital (150 rpm), a 37°C. O índice de acidez foi analisado pelo método 5-40 AOCS (MOSSOBAet al., 2003).
4.2.9 Estabilidade operacional de CALB imobilizada
4.2.9.1 Hidrólise de butirato de p-nitrofenila
A estabilidade operacional de hidrólise da lipase imobilizada foi determinada porseis ciclos subsequentes de reações de hidrólise (pNPB). Após cada ciclo, as nanopartículasforam separadas por um campo magnético, lavadas com 25 mM de tampão de fosfato de sódio,pH 7 e reciclado.
4.2.9.2 Síntese
A estabilidade operacional por reação de esterificação foi realizada através da sín-tese consecutiva de oleato de etila, usando 0,01 g de CALB-NPM. Antes de cada ciclo, oderivado foi separado por magnetização e lavado com hexano para a remoção de produtos esubstrato não reagido. As reações foram executadas como descritas na Seção 4.2.8.
4.3 Resultados e discussões
4.3.1 Efeito da velocidade de agitação na imobilização de CALB
O efeito da velocidade de agitação na imobilização da enzima em NPM é mostradana Figura 4.1. O tipo de agitação (orbital ou rotacional) foi analisado. Devido a restrições comequipamentos, a influência da agitação rotacional foi investigada entre 20 e 45 rpm e velocida-des de 20 até 250 rpm foram investigados para agitação orbital. Em ambos os casos, o sistemafoi agitado por uma hora. Pode-se observar que a atividade do derivado se mantem quase cons-tante até 180 rpm, quando a agitação orbital é empregada. Um decréscimo na atividade doderivado foi notada quando a velocidade de agitação foi aumentada para 250 rpm. Em relaçãoà agitação rotacional, a atividade do derivado aumentou quando a velocidade de agitação foiaumentada de 20 até 45 rpm. De fato, o melhor resultado de atividade (17,5 ± 0,5 U.g−1) foiatingida nestas condições, 45 rpm em agitação rotacional. A interferência da agitação pode serrelacionada diretamente a taxa de cisalhamento, a qual, inicialmente promove mais interaçõesentre a enzima e o suporte (KAYA; HEITMANN; JOYCE, 1994; KIM et al., 1982; MOZHAEV;MARTINEK, 1982). Entretanto, o aumento nesta taxa pode resultar na redução da atividade,devido a desnaturação da enzima. A 250 rpm uma queda na atividade foi registrada, isto pro-

50
vavelmente se deve a desnaturação da enzima, ver Figura 4.1. Baseado nestes resultados, todosos experimentos seguintes foram conduzidos a 45 rpm em agitação orbital.
0
5
10
15
20
2501809045
Ativ
idad
e do
der
ivad
o (U
.g-1
)
Velocidade de agitação (rpm)
20
Figura 4.1: Efeitos da velocidade de agitação (rpm) em CALB imobilizada em NPM. Atividadedo derivado (U.g−1): 25 rpm (Att = 300 U.g−1) e 45 rpm (Att = 200 U.g−1) (agitação rotacional)(�); 25 rpm (Att = 211,3 U.g−1), 45 rpm (Att = 185,3 U.g−1), 90 rpm (Att = 116,4 U.g−1), 180rpm (Att = 115,7 U.g−1) 250 rpm (Att = 148,2 U.g−1) (agitação orbital) (�).
4.3.2 Efeito da carga enzimática
O efeito da carga enzimática foi conduzida de 45 até 200 U.g−1 (Att). Resulta-dos da atividade do derivado e estabilidade operacional são apresentados na Figura 4.2. Comoobserva-se na Figura 4.2, aumentando a carga enzimática para 80 U.g−1 de suporte promoveuum aumento na atividade do derivado, sendo 29,1 U.g−1 ± 0,9 o valor mais alto atingido. Mai-ores valores na carga enzimática resultaram em baixa atividade. No tocante ao rendimento daimobilização, é possível observar que este decresce com o aumento da carga enzimática. Xie eMa (2009) (XIE; MA, 2009) estudaram a imobilização de lipase (lipozyme-TL) de Thermomy-ces lanuginosa em nanopartículas magnéticas reticuladas por glutaraldeído e funcionalizadascom APTS Fe3O4 e relataram o efeito da carga enzimática na eficiência de imobilização, mos-trando perfil semelhante ao apresentado na Figura 4.2.
O comportamento apresentando com a variação da carga enzimática pode ser expli-cado pela análise das interações entre a enzima e o suporte. O aumento da carga permite maisinteração proteína-proteína, assim uma quantidade ótima de enzima é requerida. O aumento dacarga enzimática leva a limitações difusionais, uma barreira de difusão de produto e substrato,e impedimento estéreo, o qual pode explicar a baixa atividade do derivado observada quando200 U.g−1 (Att) foi utilizada (SILVA et al., 2012).
4.3.3 Efeito do tempo de contato
O efeito do tempo de contato foi investigado e os resultados são expostos na Fi-gura 4.3. A quantidade de proteína imobilizada manteve-se constante e a atividade derivada

51
0 50 100 150 200 2500
10
20
30
40
Carga enzimática (U.g-1)
Ativ
idad
e do
der
ivad
o (U
.g-1
)
0
20
40
60
80
100
Ren
dim
ento
(%)
Figura 4.2: Efeito da carga enzimática (Att , U.g−1) na imobilização de CALB em IMNs. Ati-vidade derivada (U.g−1) (�) e Rendimento da imobilização (%) (�). As linhas representam atendência dos dados experimentais.
máxima foi atingida em 1 hora, indicando que o equilíbrio de interação enzima-suporte foi ra-pidamente alcançada. Após 4 horas, houve uma queda de atividade catalítica, possivelmentedevido ao tempo de incubação mais longo que implica em mais interações enzima-suporte. Porconsequência, a queda na atividade enzimática devido a mudanças conformacionais e estrutu-rais da enzima seria esperada devido a maior rigidez ter ocasionado desnaturação da enzima(RODRIGUES et al., 2008). É relatada na literatura (HUANG; CHENG, 2008) que um tempocurto de contato enzima-nanopartículas é necessário para atingir uma imobilização rápida, aqual pode ser atribuída a ampla superfície de contato disponível em nanopartículas que reduz aresistência a transferência de massa. Além disto, a modificação do suporte com APTS e gluta-raldeído facilitou a ligação rápida da enzima por interações covalentes entre os grupos aldeídodo glutaraldeído e os grupos amina da enzima. Uma concentração alta de grupos aldeídos nasuperfície do suporte pode causar numerosos elos covalentes multipontuais com a enzima, bemcomo obtendo uma rigidez na estrutura da enzima, favorecendo uma alta estabilidade e altaatividade intrínseca do derivado (BETANCOR et al., 2006).
Os autores Hu et al. (2009) (HU et al., 2009) estudaram a imobilização de lipaseSerratia marcescens em nanopartículas magnéticas reticuladas por glutaraldeído e funcionali-zadas com APTS Fe3O4 e relataram um rápido processo de imobilização, 10 minutos. Temposde contato superiores a 120 minutos não apresentaram qualquer mudança significativa na imo-bilização enzimática, estando em consonância com os resultados obtidos neste trabalho.
Outros estudos Xie e Ma (2009) (XIE; MA, 2009), também estudaram a influên-cia do tempo de imobilização como parâmetro na imobilização de lipase (lipozyme-TL) deThermomyces lanuginosa em nanopartículas magnéticas reticuladas por glutaraldeído e funcio-nalizadas com APTS Fe3O4 e relataram que o tempo de imobilização ótimo foi de 2 horas, commanutenção da atividade por períodos mais longos, perfis similares aos apresentados na Figura4.3. Estes autores sugerem que tal comportamento se deve a imobilização covalente e, nano-partículas magnéticas oferecer elevados pontos de ligaç ao enzima-suporte. Longos períodosde contato também facilitam mais pontos de ligação da enzima ao suporte, tornando as mesmas

52
0 1 2 3 4 50
10
20
30
40
Tempo (h)
Ativ
idad
e do
der
ivad
o (U
.g-1
)
0
5
10
15
Con
cent
raçã
o de
pro
teín
as m
g.g-1
Figura 4.3: Efeito do tempo de contato (0,5 - 5h) em CALB imobilizada em NPM. Atividadeteórica: 0,5 h (Att= 86,2 U.g−1), 1h (Att= 80 U.g−1), 2h (Att= 124,4 U.g−1), 3h (Att= 75 U.g−1),4h (Att= 84,8 U.g−1), 5h (Att= 82,4 U.g−1). Atividade do derivado (U.g−1) (�) e quantidade deproteína (mg.g−1 (�). As linhas representam a tendência dos dados experimentais.
inativas.
A Figura 4.4 mostra os ciclos consecutivos de hidrólise catalisadas por diferentesderivados preparados pelo aumento do tempo de contato. Podem ser observado os melhoresresultados para a atividade da hidrólise após 6 ciclos para o tempo de contato de 1 h, aproxi-madamente 50%. Atividades relativas entre 30-40% foram obtidas para os demais catalisadoresutilizados.
0 1 2 3 4 5 6 70
20
40
60
80
100
120
0.5 h 1 h 2 h 3 h 4 h 5 h
Ativ
idad
e re
lativ
a (%
)
Ciclos
Figura 4.4: Estabilidade operacional na hidrólise de butirato de p-nitrophenila para CALB-NPM. Atividade relativa (%): 30 min (�), 1 h (�), 2 h (4) , 3 h (N), 4 h (◦) e 5 h (•). As linhasrepresentam a tendência dos dados experimentais.
Baseados nestes resultados, elevada atividade de CALB-NPM após o reuso, todosos seguintes catalisadores foram preparados com tempo de contato de 1 h.

53
4.3.4 Efeito de diferentes concetrações de glutaraldeído
Para a ligação covalente entre a enzima e o suporte, grupos aldeídos foram intro-duzidos nas nanopartículas magnéticas por reticulação com glutaraldeído (LÓPEZ-GALLEGOet al., 2005). A Figura 4.5 mostra a influência da concentração de glutaraldeído (2,5 e 25%(m/v)) e a presença de aditivo (dodecil sufato de sódio, 0,23%, SDS) na atividade da enzimaimobilizada. Há uma forte dependência da atividade da enzima em respeito à concentraçãode glutaraldeído: a atividade do derivado aumenta aproximadamente 13 vezes usando 25% daconcentração. Geralmente, a área superficial especifica de pequenas partículas é maior do quepartículas maiores (YESILOGLU, 2005; MENDES et al., 2012). Assim, uma maior superfícierequer o uso de altas concentrações de glutaraldeído para uma reticulação eficiente. Diversosautores discutiram a ativação em soluções com altas concentrações de glutaraldeído como agen-tes de ligação (BETANCOR et al., 2006; XIE; MA, 2009; ANDRADE et al., 2010; ZHU; SUN,2012; BARBOSA et al., 2012). Uma alta concentração é importante para promover a formaçãode numerosas interações enzima-suporte, bem como para obter a rigidez na estrutura de enzima,favorecendo alta estabilidade e alta atividade intrínseca do derivado (BETANCOR et al., 2006).
2.5% Glut. 25% Glut. 25% Glut. + 0.23% SDS0
10
20
30
40
Ativ
idad
e do
der
ivad
o (U
.g-1
)
Concentração de glutaraldeído (Glut.) e SDS
Figura 4.5: Efeito de aditivos (0,23%, SDS) e concentração de glutaraldeído (2,5 e 25% (m/v))em CALB imobilizada em NPM. Atividade teórica: Glut. 2,5 % (Att= 62 U.g−1), Glut. 25 %(Att= 80 U.g−1), Glut. 25 % + SDS 0,23 % (Att= 55,8 U.g−1).
A ativação da lipase por aditivos, tais como surfactantes, é bem documentada e ge-ralmente atribuída a uma melhor conformação ativa da enzima (THEIL, 2000; GUISAN, 2006).O processo de crosslinking na presença de surfactante favorece uma melhor conformação enzi-mática, resultando no aumento da atividade e seletividade. Ademais, o aditivo pode ser facil-mente removido, pois este não é ligado de forma covalente à enzima (GUISAN, 2006). Comobjetivo de melhorar os parâmetros de imobilização, a influência da adição de SDS ao meio deimobilização foi investigada. O surfactante anionico SDS tem sido estudado por vários autores(MOGENSEN; SEHGAL; OTZEN, 2005; A; WIEGEL, 2010) para aumentar a atividade ca-talítica. Infelizmente, como pode ser visto na Figura 4.5, a presença de SDS não permite umligação eficiente da enzima ao substrato, pois a atividade do derivado (15,8 U.g−1) foi de apro-ximadamente metade do valor obtido na ausência deste (29,1 U.g−1). O efeito do surfactante éatribuído à ativação interfacial da lipase, mas a enzima CALB apresenta uma tampa menor que

54
outras lipases, assim reduzindo os efeitos do detergente SDS (UPPENBERG et al., 1994).
Ciclos consecutivos de reação de hidrólise foram realizados, ver Figura 4.6. Osresultados confirmam uma melhor estabilidade catalítica da enzima na ausência de surfactante,o derivado preparado na ausência de SDS reteve mais de 50% de sua atividade após 5 ciclos.Derivados preparados na presença de SDS mantiveram menos de 30% da atividade inicial sobas mesmas condições.
0 1 2 3 4 50
20
40
60
80
100
120
A
tivid
ade
rela
tiva
(%)
Ciclos
Figura 4.6: Estabilidade operacional em hidrolise de butirato de p-nitrofenila para CALB-NPM.Aditivos: 25% glutaraldeído (�) e (25% glutaraldeído + 0,23%, SDS) (�).
4.3.5 Estabilidade térmica de CALB solúvel e imobilizada
A estabilidade térmica da lipase solúvel e imobilizada foi estudada a 60°C em pH7 (25 mM de tampão de fosfato de sódio) por 48 horas. A Figura 4.7 expõe a inativação dasenzimas solúvel e imobilizada. O processo de inativação é muito rápido para a CALB solúvel,perdendo mais de 70% da atividade catalítica em 6 minutos, enquanto que para a imobilizada háapenas uma perda de 20% de sua atividade inicial. Mesmo após 20 horas, a lipase imobilizadaainda mantem 20% de sua atividade inicial.
A Tabela 4.1 mostra os resultados da desativação térmica a 60°C de CALB solúvel eCALB-NPM. Para cada sistema, foram analisadas a estabilidade térmica, o tempo de meia-vida(t1/2) e o fator de estabilização (SF).O tempo de meia-vida (t1/2) aumentou de 3 para 60 minutoscom o processo de imobilização. A enzima imobilizada é 20 vezes mais estável que a enzimasolúvel a 60°C (maior fator de estabilização, veja Tabela 4.1). A enzima na forma solúvel apre-senta certa flexibilidade, o que significa que seu sítio ativo sofre mudanças conformacionais quesão frequentemente irreversíveis, causando inatividade. Quando imobilizada, uma forma maisrígida é adquirida devido a ligações ao suporte. Esta rigidez diminui a flexibilidade da enzima,mantendo a forma do sítio ativo, o qual é responsável por sua atividade (TREVAN, 1980). Osefeitos na estabilidade resultantes do processo de imobilização com APTS e glutaraldeído po-dem ser consequência de ambos: ligação cruzada intermolecular e modificação intramolecular(FERNANDEZ-LAFUENTE et al., 1998).

55
0 10 20 30 40 500.0
0.2
0.4
0.6
0.8
1.0
Ativ
idad
e re
lativ
a
Tempo(h)
0.0 0.5 1.0 1.5 2.0 2.5 3.00.0
0.2
0.4
0.6
0.8
1.0
Ativ
idad
e re
lativ
a
Tempo (h)
Figura 4.7: Estabilidade térmica de CALB livre e CALB-IMNs a 60°C e pH 7. Enzima so-lúvel Ati= 93 U.mL−1 e CALB-IMNs Att= 80 U.g−1. Atividade relativa: CALB imobilizada(•) e CALB livre (N). AS linhas representam a tendencia do modelo de Sadana e Henley.(FERNANDEZ-LAFUENTE et al., 1993)
Tabela 4.1: Desativação térmica a 60°C e pH 7, de CALB solúvel e CALB-NPM
Catálise Kd(h−1) t1/2(h) SFCALB solúvel 15,3±366 0,051 −CALB-NPM 0,94±18 1,0 20,0
Os autores Silva et al. (2012) (SILVA et al., 2012) mostraram a desativação térmicaa 60°C de CALB solúvel e CALB-quitosana modificada por diferentes métodos.O tempo demeia-vida dos catalisadores foi relatado entre 1-3,3 horas, sendo a enzima imobilizada maisestável do que a solúvel, entre (12-33 vezes). Outros autores Hu et al. (2009) (HU et al., 2009)estudaram os efeitos da temperatura na lipase imobilizada e solúvel (em aldeído - nanopartícu-las magnéticas funcionalizadas (Fe3O4)). A atividade máxima da lipase solúvel e imobilizadaforam observadas a 50°C e 45°C, respectivamente, mas a lipase imobilizada foi mais estávela altas temperaturas (entre 45°C e 55°C), estando em consonância com os resultados obtidosneste trabalho.
4.3.6 Esterificação catalítica
Biodiesel foi produzido por esterificação e a Figura 4.8 mostra, para 48 horas dereação, a conversão de ácido oleico em oleato de etila. Foi observado que aproximadamente90% de ambos os derivados CALB-NPM e Novozym® 435 foram obtidos. Inicialmente aconversão é mais rápida para reação catalizada por resina CALB comercial, mas a 48 horas, asduas catálises atingiram o equilíbrio reacional.
Estudos na literatura (XIE; MA, 2009; TRAN; CHEN; CHANG, 2012) reportama produção de biodiesel com lipase imobilizada em nanopartículas magnéticas de ferro. A li-pase lipozyme-TL de Thermomyces lanuginose imobilizada em Fe3O4 foi estudado por Xie eMa (2010) (XIE; MA, 2010; TRAN; CHEN; CHANG, 2012). A conversão em biodiesel atin-

56
giu 90% em 35 horas, após 5 ciclos, este sistema reteve 70% de sua atividade inicial. Estesresultados estão de acordo com as Figuras 4.8 e 4.9. Tran, Chen e Chang (2012) (XIE; MA,2009; TRAN; CHEN; CHANG, 2012) estudaram a produção de biodiesel por Burkholderia sp.imobilizada em alquil-Fe3O4−SiO2, obtendo uma conversão de 92% em 30 horas. Foresti eFerreira (2005) (FORESTI; FERREIRA, 2005) estudaram a imobilização de Candida antarc-tica do tipo B em pó de polipropileno usando na produção de biodiesel. Os autores obtiveramconversão de 78,5% após 7 horas. A Novozym® 435 foi estudada por Shimada et al. (1999)(SHIMADA et al., 1999) na produção de biodiesel, a conversão após 72 horas foi de 98%. Osresultados obtidos em nosso estudo são coerentes com aqueles relatados na literatura.
0 20 40 60 80 1000
20
40
60
80
100
CALB-NPM CALB-Resina acrilica
Con
vers
ão d
e ác
ido
olei
co (%
)
Tempo (h)
Figura 4.8: Dinâmica da síntese de oleato de etila usando CALB-NPM Att= 80 U.g−1, a 37°Ce 96 horas em agitação orbital. CALB− IMNs (�) e Novozym® 435 (�). Reagentes: ácidooleico (0,67 mL) e etanol (0,15 mL) a mol.L−1 (livre de solvente). Volume reacional = 0,82mL. Massa derivada = 0,01g)
4.3.7 Estabilidade operacional do derivado
A Figura 4.9 ilustra a manutenção da atividade catalítica após 14 ciclos para ambos,Novozym® 435 e CALB-NPM. Estes resultados são os melhores resultados encontrados naliteratura (XIE; MA, 2010; FORESTI; FERREIRA, 2005), a manutenção da atividade catalíticapor 5 ciclos em estudos conduzidos por Xie e Ma (2010) e 3 ciclos em Foresti e Ferreira (2005).Apesar de que a atividade catalítica para CALB-NPM reduzir-se de 25% após 14 ciclos, estaqueda pode se está relacionada à perda de massa catalítica após o procedimento (26% perda demassa catalítica).
A manutenção da conversão para biodiesel em nosso trabalho foi de 90% para aNovozym® 435 por 14 ciclos e CALB-NPM manteve 85% de atividade catalítica por 9 ciclosde 48 horas, alcançando 70% no 14º ciclo. A estabilidade mostrada pelos catalisadores emnosso estudo ressaltam a importância da catálise enzimática para a produção de biodiesel, apre-sentando uma potencial alternativa à catálise química que, por sua vez, é prejudicial ao meioambiente.

57
0 2 4 6 8 10 12 14 1650
60
70
80
90
100
Con
vers
ão d
e ác
ido
olei
co(
%)
Ciclos
CALB-NPM CALB-Resina acrílica
Figura 4.9: Estabilidade operacional da síntese de oleato de etila usando CALB-NPM Att= 80U.g−1. Cada ciclo teve duração de 48 h, agitação orbital, a 37°C. CALB− IMNs (�), Novozym®435 (•). Reagentes: ácido oleico (0,67 mL) e etanol (0,15 mL) (livre de solvente). Volumereacional = 0,82 mL. Massa derivada = 0,01g)
4.4 Conclusões
A análise experimental das condições de imobilização mostrou uma influência sig-nificativa da taxa de agitação, carga enzimática, concentração de glutaraldeído e tempo de imo-bilização de CALB em nanopartículas magnéticas de ferro. O melhor valor de atividade doderivado foi 29,1± 0,9, preparado de acordo com as melhores condições (45 rpm, 80 U.g−1,concentração de glutaraldeído: 25% e tempo de contato: 1 hora). A CALB-NPM foi 20 ve-zes nas estável do que a enzima solúvel, indicando um suporte promissor para imobilização deenzimas. Ainda, estudos de reuso mostraram que a produção de biodiesel em etanol reagidocom ácido oleico (taxa molar 1:1) misturado com catalisador 1,7% a 37°C por 48 horas, obteveuma produção de 90% para os dois derivados estudados. Os ciclos de reação operacional mos-tram manutenção da conversão por CALB-NPM e resina acrílica, assim o uso de CALB-NPMé particularmente vantajoso para a produção de ésteres.
Agradecimentos
Os autores gostariam de agradecer as agências brasileiras de fomento a pesquisaCNPq e CAPES.

58
5 PRODUÇÃO DE BUTIRATO DE METILA E ETILA
Produção de ésteres por lipase de Candida antarctica tipo B imobilizadas emnanopartículas magnéticas
Maria Cristiane Martins de Souza, Kímberle Paiva dos Santos, Rafael MeloFreire, Antonio César Honorato Barreto, Pierre Basílio Almeida Fechine e Luciana RochaBarros Gonçalves
Abstract
The enzyme Candida antarctica lipase B (CALB) immobilized onto iron magneticnanoparticles was evaluated as biocatalyst for synthesis of flavor esters. Methyl and ethyl buty-rate were biosynthesized by esterification of butyric acid with methanol and ethanol in heptanoas solvent. The nanoparticles were produced by the co-precipitation method. Modificationswere carried out on the nanoparticles’ surfaces with γ-aminopropyltriethoxysilane and glutaral-dehyde. The immobilization was performed in the presence of 100mM bicarbonate buffer, pH10, at 25 °C. The process parameters (solvents, temperature, substrate concentrations, molarratio, amount of biocatalyst, stirring speed and reaction time) were studied. The optimum con-ditions for both esters were heptano solvent, 25 °C, 0,5 mol/L (methyl butyrate) and 0,4 mol/L(ethyl butyrate), molar ratio of 1:1, amount of biocatalyst: 10 mg, 150 rpm and 8 h of reaction.Maximum conversion of methyl butyrate (93,9 %) and ethyl butyrate (96,8 %) were achievedafter 8 h of reaction at 25 °C for CALB immobilized onto magnetic nanoparticles. The synthe-sis of flavor esters were also evaluated by using novozymes 435, a commercial biocatalyst, forcomparison. Opearational stability (consecutive cycles of synthesis of esters) were performedand CALB immobilized onto magnetic nanoparticles retained 76 % of its initial activity after12 cycles, while novozym 435 etained 79 %.
Keywords: Lipase , Magnetic nanoparticles , Immobilization , Flavor esters

59
Resumo
A enzima lipase de Candida antarctica do tipo B (CALB) imobilizada em nano-partículas magnéticas de ferro foi avaliada como catalisador para a síntese de ésteres utilizadoscomo aromas. Butirato de metila e etila foram sintetizado por esterificação de ácido butíricocom o metanol e etanol em presença de heptano como solvente. As nanopartículas foram pro-duzidas pelo método de co-precipitação. Modificações foram realizadas na superfície das na-nopartículas com γ-aminopropiltrietoxissilano e glutaraldeído. A imobilização foi realizada napresença de 100 mM de tampão bicarbonato de sódio, pH 10, 25 °C. Os parâmetros de processo(temperatura, solventes, as concentrações de substrato, relação molar, a quantidade de biocata-lisador, velocidade de agitação e tempo de reacção) foram analisados. As condições óptimaspara ambos os ésteres foram heptano solvente, 25°C, 0,5 mol/L (metanol:ácido butírico) e 0,4volume mol/L (etanol:ácido butírico), razão molar de 1:1, massa de catalisador: 0,01 g, 150 rpme 8 h de reação. Conversão máxima para butirato de metila (93,9%) e butirato de etila (96,8%)foram obtidos após 8 h de reação a 25°C para CALB imobilizada em nanopartículas magnéticas(CALB-NPM). A síntese de ésteres também foi avaliada usando Novozym® 435, um catalisa-dor enzimático comercial, para comparação. Estabilidade operacional (ciclos consecutivos desíntese de ésteres) foram realizadas e CALB imobilizada em nanopartículas magnéticas reteve76% da sua atividade inicial após 12 ciclos, enquanto que a Novozym® 435 reteve 79%.
Palavra-chave: Lipase , nanopartícula magnética, imobilização, ésteres.

60
5.1 Introdução
Os esteres de ácidos graxos de cadeia curta são um importante grupo de compo-nentes de aromas amplamente empregados nas indústrias de alimento, bebidas, cosméticos efarmacêutica (XU et al., 2002; JIN et al., 2012). Estes aromas são tipicamente extraídos de fon-tes naturais, mas alguns problemas surgem com respeito à disponibilidade devido a problemasambientais (BJÖRKLING; GODTFREDSEN; KIRK, 1991; XU et al., 2002). A síntese destescompostos é por métodos químicos, utilizando catalisadores químicos poluentes, requerendopós-tratamento do produto obtido (Ben Salah et al., 2007). Assim a catálise natural é requerida,a síntese enzimática é considerada um rota “verde” (GILLIES; YAMAZAKI; ARMSTRONG,1987). A enzima lipase (glicerol éster hidrolases, EC 3.1.1.3) constitui um grupo diverso deenzimas que atuam na interface orgânico-aquosa, catalizando a hidrólise de ligações ésteres.Em meios orgânicos o comportamento enzimático é modificado e a enzima pode ser usadapara reações de esterificação e outras sínteses (UPPENBERG et al., 1994; IDRIS; BUKHARI,2011). Lipases produzidas por Candida sp. são enzimas bem estabelecidas para propósitos decatálise,especialmente a lipase de Candida antarctica do tipo B (CALB) (MARIA et al., 2005).A CALB tem sido estudada para potenciais aplicações em indústria de alimentos, aromas, de-tergentes, fármacos, tecidos, cosméticos, papel e óleo-químicos (HASAN; SHAH; HAMEED,2006; HUNG et al., 2003). Estas aplicações são possíveis devido a sua ampla faixa de espe-cificidade para substratos, alta atividade para várias reações sob condições moderadas, uso desolventes e alta estabilidade térmica e uso em diversas condições de pH, e estéreo-especificidade(RODRIGUES et al., 2008; MCCABE; RODGER; TAYLOR, 2005; DENG et al., 2011).
O uso de lipase imobilizada é recomendado para redução de custos e aumento daestabilidade da forma solúvel, estas vantagens incluem também a facilidade de separação deprodutos, aprimoramento das propriedades da lipase solúvel como estabilidade térmica e ati-vidades, o reuso e regeneração do suporte permitir maior flexibilidade da enzima-substrato dediversas configurações de reatores (SILVA et al., 2012; GOMES; PEREIRA; CASTRO, 2004).Materiais alternativos aos suportes comerciais (resinas acrílicas) são estudados para obtenção denovas propriedades que favoreçam maior resistência, alta superfície e reduzido impacto ambien-tal. Dentre estes materiais destacam-se as nanopartículas magnéticas. A grande superfícies des-tes materiais pode permitir modificações para imobilização de enzimas (TARTAJ et al., 2005).Os grupos presentes na superfície das nanopartículas, como grupos hidroxilas, podem defi-nir pontos para ligação com agentes funcionalizantes (BARRETO et al., 2012; BOYER et al.,2010). As aplicações industriais de nanopartículas magnéticas são as mais diversas, destacando-se as aplicações biomédicas (ressonância magnética e agentes terapêuticos no tratamento decâncer) (TARTAJ et al., 2003; MERBACH; TÓTH, 2001; CHARLES; POPPLEWELL, 1982).
No presente estudo, CALB imobilizada em nanopartículas magnéticas (CALB-NPM) foi usada sucessivamente na síntese de dois importantes aromas por esterificação. Osésteres butirato de etila e butirato de metila produzidos são amplamente utilizados na indús-tria de aromas e possuem uma alta demanda na indústria alimentícia como aromas de bananae abacaxi (GILLIES; YAMAZAKI; ARMSTRONG, 1987). A influência da temperatura, con-centração de substrato, razão molar ácido-álcool, velocidade de agitação, tempo de reação e

61
tipos de solventes na síntese dos ésteres de butirato de metila e butirato de metila foram inves-tigados. Os resultados obtidos foram comparados com a CALB imobilizada em resina acrílica(Novozym® 435). Ciclos consecutivos de esterificação foram realizados para a CALB imobili-zadas em nanopartículas magnéticas e resina acrílica (Novozym® 435).
5.2 Métodos
5.2.1 Materiais
Nanopartículas magnéticas de ferro (NPM) (Fe3O4) foram produzidas pelo métodode co-precipitação (BARRETO et al., 2012). Lipase de Candida antartica do tipo B (CALB) foiadquirida da Codexis (Redwood, USA). A CALB imobilizada em resina acrílica (Novozym®435), γ-aminopropiltrietoxisilano (APTS), solução de glutaraldeído grau II 25% (w/v), butiratode p-nitrofenila (pNPB) e p-nitrofenol (pNP) foram obtidos da Sigma-Aldrich (St. Louis,USA). Os demais reagentes de grau analítico foram obtidos da Synth (São Paulo, Brasil) eVetec (São Paulo, Brasil).
5.2.2 Produção de nanopartículas magnéticas
Nanopartículas magnéticas de ferro (Fe3O4) foram produzidas pelo método de co-precipitação (BARRETO et al., 2012). Difração de raio-X (XRD) e transmissão eletrônica devarredura (TEM) foram realizadas para determinação do tamanho das nanopartículas (11,0 nm)(BARRETO et al., 2012).
5.2.3 Modificação por APTS
A reação foi iniciada com a adição de solução de APTS (2,0% v/v) ao suportena proporção de 0,2 (ml/mg de suporte). A solução foi aquecida a 100°C por 10 horas sobatmosfera de nitrogênio. As nanopartículas foram lavadas com 100 ml de metanol e 100 ml deetanol, separadas por magnetismo, em seguida o material foi seco a 30°C por 24 horas (NETTO;ANDRADE; TOMA, 2009).
5.2.4 Glutaraldeído crosslinking
NPM (0,01g) foram suspendidas em solução 25% de glutaraldeído na proporção de2,5 (solução (ml)/suporte (g)). A mistura foi mantida sob agitação (45 rpm) por 2 horas à 25C. Finalmente o suporte foi lavado com tampão bicarbonato 100 mM, pH 10,0 para remover oexcesso de glutaraldeído (ANDRADE et al., 2010).

62
5.2.5 Imobilização de lipase
O processo de imobilização foi realizado pelo contato de 0,01 g de nanopartículas(tratada por APTS e reticulada com solução de glutaraldeído) com 0,5 ml de solução tampãode bicarbonato de sódio, 100 mM, pH 10,0. O sistema foi mantido sob agitação controlada: 45rpm . O tempo de contato enzima-suporte foi de 1 hora. A enzima CALB imobilizada foi remo-vida da solução por magnetismo. Medidas de atividade da enzima solúvel e imobilizada foramrealizadas de acordo com procedimento descrito a seguir. A quantidade de CALB imobilizadaem NPM foi determinada através da medição da concentração inicial e final do CALB no so-brenadante de imobilização. Atividades hidrolíticas também foram determinadas para permitiro cálculo dos parâmetros de imobilização: rendimento de imobilização (R), atividade teórica(Att) e atividade recuperada (Atr ) (SILVA et al., 2012).
5.2.6 Síntese
A produção de éstres por reação de esterificação foi composta de 1 mL de solventes(heptano, hexano, ciclohexano e 1,4-dioxano) como meio reacional, ácido butilico e os álcoois(metanol e etanol) em diferentes concentrações (0,2 - 1,0 moL/L) e razão molar (1:1 - 1:4). Obiocatalisador (0,01 g), denominado CALB-NPMfoi adicionado a uma atividade específica (80U.g−1). A reação foi conduzida sob agitação orbital (50 - 250 rpm), a 25 - 55°C. O índice deacidez foi avaliado pelo método Ca 5-40 AOCS (MOSSOBA et al., 2003). A melhor condiçãoreacional foi usada com CALB imobilizada em resina acrílica (Novozym® 435).
5.2.7 Reuso do biocatalisador
A estabilidade operacional foi realizada por reações consecutivas da síntese dos és-teres butirato de metila e butirato etila, usando 0,01 g de CALB-NPM. Antes de cada novo ciclo,CALB-NPM foi separada por magnetização e lavada com hexano para remoção de produtos esubstratos não reagidos. As reações foram conduzidas como descrito na seção 5.2.6.
5.3 Resultados e discussões
5.3.1 Efeito da temperatura
O efeito da temperatura sobre a conversão de ácido butírico por CALB-IMNs (Atd= 29,1 U. g−1, 0,01 g) é mostrado na Figura 5.1. A conversão de butirato de metila mantém-se constante de 15 até 40 °C e apresenta uma forte queda acima da 40°C. A conversão debutirato de etila permanece constante para 15-25°C, apresenta uma redução suave até 45°Ce depois se estabiliza. Este comportamento deve-se provavelmente ao fato de que o metanol(ponto de ebulição: 65°C) é mais volátil do que o etanol (ponto de ebulição: 78,4 °C). Umamaior evaporação da solução de reação com metanol foi observada. Sob condições de solventes

63
orgânicos, o ponto de ebulição do solvente pode definir o limite superior de conversão, bemcomo a estabilidade da enzima (HALLING, 2000).
Cambon et al. (2009) (CAMBON et al., 2009) estudaram a síntese de ésteres dealquiloade óleos vegetais e descobriram que as taxas de conversão e de inibição de metanol sãomuito dependentes da temperatura. Esta dependência foi atribuída ao estado termodinâmico domeio. O diagrama de equilíbrio de fases líquido-líquido e o coeficiente de partição líquido-vapordo metanol são parâmetros importantes para se descrever o estado termodinâmico do meio. Emresumo, o aumento da temperatura aumenta a concentração molar do metanol na fase de vapor,diminuindo a concentração molar em fase líquida em que a reação ocorre (CAMBON et al.,2009).
0 10 20 30 40 50 600
10
20
30
80
90
100
Con
vers
ão d
e ác
ido
butír
ico
(%)
Temperatura (°C)
Figura 5.1: Efeito da temperatura na síntese dos butiratos de metila e etila. Substratos: etanol(�) e metanol (•). As reações foram realizadas utilizando CALB-NPM Att= 80 U.g−1, 150 rpm,heptano, 02 mol/L de ácido butírico, 1:1 (ácido butírico: (metanol and etanol)) e 8 h de reação.Volume reacional = 1,0 mL. Massa de catalisador CALB-NPM = 0,01g.
Os mesmos autores Cambon et al. (2009) (CAMBON et al., 2009) observaram umaredução de 15% na conversão de ésteres de alquila no intervalo de 30 à 55 °C, alcançandoum máximo de conversão à 30 °C, esses resultados estão de acordo com os obtidos no nossotrabalho, em que ocorreu uma redução na conversão de ésteres com o aumento da temperatura,a máxima conversão foi obtida a 25 °C.
5.3.2 Efeito da concentração de substratos
A influência da concentração de substratos foi investigada utilizando CALB-NPM.O efeito da concentração de substratos e razão molar tem sido reportada como um impor-tante parâmetro nas reações de síntese (GUILLÉN; BENAIGES; VALERO, 2012). Os estudosapresentados na literatura mostram um significativo efeito no aumento das concentrações deácido e álcool na atividade enzimática (Ben Salah et al., 2007; GUILLÉN; BENAIGES; VA-LERO, 2012; GHAMGUI; KARRA-CHAÂBOUNI; GARGOURI, 2004; CHEN, 1996; AH-MED; RAGHAVENDRA; MADAMWAR, 2010; GUILLÉN; BENAIGES; VALERO, 2012).A Figura 5.2 apresenta o perfil para a conversão do éster butirato de etila e butirato de metila,

64
a maior conversão foi obtida para o éster butirato de etila (97 %) nas condições de 0,4 mol/Lde substratos (1:1, razão molar) e 8 horas de reação, tendo-se uma redução a 86.5 % com oaumento da concentração de substratos (1,0 mol/L). A conversão para o butirato de metila é deaproximadamente 93 %, após 8 horas de reação, 0,5 mol/L de substratos (1:1, razão molar),ocorrendo um decréscimo a 87.9 % na maior concentração utilizada 1,0 mol/L. Os resultadosestão de acordo com os apresentados na literatura (GUILLÉN; BENAIGES; VALERO, 2012),esses autores apresentaram conversão de 99% para butirato de etila produzido por lipase imo-bilizada em resina polimérica, porém com um tempo três vezes superior aos demostrados nonosso trabalho.
0.0 0.2 0.4 0.6 0.8 1.070
75
80
85
90
95
100
Con
vers
ão d
e ác
ido
butír
ico
(%)
Concentração de substrato (mol/L)
Figura 5.2: Efeito da concentração de substratos na conversão de butirato de etila e metila.Substrato álcoois: etanol (�) e metanol (•). As reações foram realizadas utilizando CALB-NPMAtt= 80 U.g−1, 25 °C, 150 rpm, heptano, 0,1-1,0 mol/L de ácido butírico, 1:1 (ácido butírico:(metanol e etanol)) e 8 h de reação. Volume reacional de 1,0 mL. Massa de biocatalisadorCALB-NPM de 0,01 g.
A maior concentração de metanol no meio reacional proporcionou inicialmente umaumento na conversão do ácido butírico para butirato de metila, porém os valores obtidos fo-ram menores dos que utilizando etanol como substrato, como explicado na seção anterior Seção5.3.1 este comportamento pode ser relacionado a maior evaporação do mesmo do meio reacio-nal.
Outros autores (ABBAS; COMEAU, 2003; GANDHI; SAWANT; JOSHI, 2008;SUN; XU; WANG, 2009) descreveram que a inibição por metanol está relacionada a menorenergia liberada por está molécula para a modificar a configuração do sítio ativo e facilitar ainteração com enzima-substratos. O metanol também é considerado tóxico, podendo desnaturara enzima (SUN; XU; WANG, 2009). Os álcoois tem a capacidade de ligação com proteínas einduzir a desidratação das mesmas, resultando em uma drástica perda da atividade enzimática.Assim, no nosso trabalho observa-se que o maior efeito de inibição ocorreu nos extremos deconcentração analisados, podendo ser correlacionado com a maior solubilização do álcool nomeio reacional.
A estrategia de usar concentrações iguais de álcool e ácido também foram estudadaspor Dave (2005) (DAVE, 2005) e Dias et al. (1991) (DIAS et al., 1991). Resultados similares

65
foram reportados quando foi utilizada a lipase de Candida rugosa imobilizada em sílica gelutilizando hexano como solvente e concentrações de ácido butírico e etanol de 0,2 mol/L, apre-sentando uma conversão a butirato de etila de 95%. A lipase de Candida rugosa imobilizadaem poliuretano utilizando hexano como solvente e concentrações de ácido butírico e etanol de0,3 mol/L apresentou uma conversão a butirato de etila de 70%. No nosso trabalho a melhorcondição foi obtida na concentração de 0.4 mol/L com conversão a butirato de etila de 97%.
5.3.3 Efeito da razão molar álcool:ácido
O efeito da razão molar álcool:ácido na reação de esterificação foi investigada. Ini-cialmente, manteve-se a concentração de ácido constante e após a concentração de álcool. AFigura 5.3 apresenta o perfil para a influência das diferentes razões molares na conversão de bu-tirato de etila e metila, o máximo de conversão obtida é alcançado na razão molar álcool:ácido1:1. O excesso de álcool não alterou drasticamente a síntese dos dois ésteres estudados. Esteresultado está de acordo com o descrito na literatura para a síntese de ésteres utilizando CALB(NORDBLAD; ADLERCREUTZ, 2008).
1:1 1:1.5 1:2 1:3 1:40
20
40
60
80
100
Con
vers
ão á
cido
but
írico
(%)
Razão molar [álcool :ácido]
Figura 5.3: Efeito da razão molar álcool:ácido na reação de esterificação para a produção dosésteres butirato de etila e metila. Perfil da variação da concentração de álcool (�) e ácido (�)para butirato de etila. Perfil da variação da concentração de álcool (•) e ácido (◦) para butiratode metila. As reações foram conduzidas por uso de CALB-NPM Att= 80 U.g−1, 25 °C, 150rpm, heptano, 0.5 mol/L (metanol) and 0.4 mol/L (etanol), 1:1-1:4 (ácido butírico: (metanol eetanol)) e 8 h de reação. Volume reacional de 1,0 mL. Massa de catalisador CALB-NPM de0,01 g.
O excesso de ácido reduziu a taxa de conversão de 93,4% para a 7,4 % para aprodução de butirato de metila. A produção de butirato de etila obteve redução de 96,8% para24,7 %. A rápida redução da taxa de conversão indicaram que o ácido butírico promoveuuma inibição pelo substrato, o que é consistente com vários autores (CLAON; AKOH, 1994;NORDBLAD; ADLERCREUTZ, 2008; JIN et al., 2012; ROMERO et al., 2005). Os estudosna literatura têm sugerido que o ácido e álcool podem inibir lipases por meio de mecanismossemelhantes de inibição competitiva (NORDBLAD; ADLERCREUTZ, 2008). Para o ácido,isto significaria que uma segunda molécula de ácido pode interferir com a desacetilação da

66
enzima, bloqueando o acesso ao sítio ativo (Hari Krishna; KARANTH, 2001). No entanto,as concentrações cada vez maiores de ácido podem aumentar a concentração de prótons nosistema e reduzindo assim a atividade enzimática por protonação prejudicial (NORDBLAD;ADLERCREUTZ, 2008). A maior eficiência de reação foi atingida quando a concentração deácido butírico foi de 0,4 e 0,5 mol/L (proporção molar de 1:1) para a esterificação de butiratode metila e etila, respectivamente.
5.3.4 Efeito da velocidade de agitação
A velocidade de agitação é um parâmetro essencial devido ao seu efeito positivona transferência de massa. Figura 5.4 mostra a influência da velocidade de agitação sobre asíntese de butirato metila e etila. O perfil apresentado indica que as velocidades de agitaçãoentre 50 e 200 rpm promovem as taxas de conversão de 80% até 90% para ambos os ésteres.Os autores Mahapatra et al. (2009) investigaram a influência da velocidade de agitação sobrea transferência de massa e observaram que um aumento da velocidade de agitação produz umaredução na espessura da película em torno das partículas sólidas, e em seguida, a resistênciade transferência de massa diminui. Como é mostrado na Figura 5.4, os melhores resultados seencontram a 150 rpm para ambos os ésteres produzidos.
0 50 100 150 200 250 3000
20
40
60
80
100
Butirato de etila Butirato de metila
Con
vers
ão d
e ác
ido
butír
ico
(%
)
Velocidade de agitação (rpm)
Figura 5.4: Efeito da velocidade de agitação na síntese de buriato de etila e metila. Os álcooisusados foram: etanol (�) e metanol (•). Reações foram realizadas usando CALB-NPM Att=80 U.g−1, 25 °C, 150-250 rpm, heptano, 0,5 (metanol) e 0,4 (etanol) mol/L ácido butírico, 1:1(ácido butírico: (metanol e etanol)) e 8 h tempo de reação. Volume reacional = 1,0 mL. Massado catalisador = 0,01 g
A menor influência de transferência de massa em nanopartículas, tal como apre-sentada na Figura 5.4, para velocidades entre 50 e 200 rpm, pode ser explicada pelo fato deque as nanopartículas apresentam maior área de superfície que causa uma maior dispersão daenzima, evitando assim a sua agregação e melhorando a transferência de massa (??DANDA-VATE; KEHARIA; MADAMWAR, 2009). A queda acentuada verificada da taxa de conversãodo metanol como substrato sobre a maior velocidade (250 rpm) pode ter sido favorecida poruma maior passagem para a fase de vapor de álcool, reduzindo a sua concentração molar nomeio reacional tal como explicado na secção 5.3.1.

67
5.3.5 Produção de ésteres na presença de solventes orgânicos
A produção de butirato de metila e etila por CALB-NPM também foi estudadana presença de solventes orgânicos (por ordem de aumento de hidrofobicidade): 1,4-dioxano,ciclo-hexano, hexano e heptano. O comportamento hidrofóbico é um dos factores importantespara a estabilidade dos catalisadores em um ambiente não aquoso (SUN; XU; WANG, 2009).O parâmetro logP, o coeficiente de partição do solvente, entre 1-octanol e água, é uma medidaquantitativa da polaridade do solvente, é frequentemente utilizado para predizer os possíveisefeitos sobre a atividade catalítica da enzima em ambientes orgânicos (LAANE et al., 1987;XU et al., 2002).
Tabela 5.1: Efeito de solvente orgânico em síntese de butirato de metila e etila. Reações foramrealizadas usando CALB-NPM Att= 80 U.g−1, 25 °C, 150 rpm, 0,5 (metanol) e 0,4 (etanol)mol/L ácido butírico, 1:1 (ácido butírico: (metanol e etanol)) e 8 h tempo de reação. Volume dereação = 1,0 mL. Massa do catalisador = 0,01 g
Solvente logPConversão de ácido butírico
Etila MetilaHeptano 4,0 96,8±0,3 93,9±0,67Hexano 3,5 83,7±1,4 84,3±1,3Ciclohexano 3,2 84,7±2,4 78,4±2,51,4-Dioxano -1,14 37,3±5,5 49±2,8
A Tabela 5.1 mostra a relação entre logP e a taxa de conversão de ésteres. O au-mento da hidrofobicidade de solvente orgânico (crescimento de logP) conduz a um aumentosignificativo da taxa de conversão para os butiratos estudados, o que indica a dependência emrelação ao tipo de solvente utilizado. A conversão de substrato máxima foi conseguida na pre-sença de heptano. 96,8% para butirato de etila e 93,9% para butirato de metila. Estudos têmmostrado que a produção da reação é melhor com solventes não polares, atendo-se ao fato deque os solventes polares poderiam distorcer a camada de água em torno de lipase (Ben Salahet al., 2007). Uma pequena quantidade de água em torno das moléculas de enzima é essencialpara manter a atividade da enzima (LAANE et al., 1987). Os resultados de nosso estudo sãoconsistentes com os descritos na literatura, em que solventes com logP > 2 suportar os níveismais elevados de atividade enzimática (GUILLÉN; BENAIGES; VALERO, 2012; XU et al.,2002; SUN; XU; WANG, 2009; NORDBLAD; ADLERCREUTZ, 2008).
Outro aspecto importante na análise de solventes é a sua constante dielétrica. Forçaseletrostáticas são importantes para a catálise enzimática e eles são responsáveis pela estabiliza-ção do estado de transição (WARSHEL et al., 1989; PARK; CHI, 2001).
As constantes dielétricas estudadas são: hexano 1,88, heptano 1,92, ciclo-hexano2,02 e 1,4-dioxano 2,25. É previsível que a energia eletrostática para a formação do estadode transição tetraédrico da catálise enzimática é estabilizada por baixa constante dielétrica dosolvente (PARK; CHI, 2001; WARSHEL et al., 1989). Este comportamento foi observado parao 1,4-dioxano e ciclo-hexano, o que a constante dielétrica mais baixa produziu um aumentode 50% de a taxa de conversão, consulte a Tabela 5.1. No entanto, o solvente hexano apre-sentou comportamento diferente do esperado, apesar da análise de estabilidade indicar que o

68
uso de hexano deveria proporcionar uma maior estabilização do sítio catalítico, a complexidadeda dependência da atividade em solventes resulta em melhores resultados para a utilização deheptano.
A esterificação de etanol com ácido butírico em presença de hexano e heptano érelatada por muitos autores (DIAS et al., 1991; MANJON; IBORRA; AROCAS, 1991; DAVE,2005; RODRIGUEZ-NOGALES; ROURA; CONTRERAS, 2005; ARAGÃO et al., 2011). Omelhor resultado obtido entre esses estudos foi por Manjon, Iborra e Arocas (1991) com ouso de lipase de R. miehei imobilizada em Celite e esterificação em presença de hexano. Osresultados apresentados na Tabela 5.1 estão condizentes com os comportamentos apresentadosna literatura.
5.3.6 Tempo de reação
O perfil de tempo de conversão é apresentado na Figura 5.5. Para o mesmo períodode tempo (1 hora), a conversão de substratos variaram significativamente com a natureza dossubstratos de álcool. As conversões mais elevada de 96,8% e 93,9% foram observadas com buti-rato de etila e metila, respectivamente, no tempo de incubação de 8 horas, passado este período,os valores aproximadamente constantes foram observados. O mesmo comportamento foi obser-vado pelos autores Abbas e Comeau (2003) (ABBAS; COMEAU, 2003), em que as primeiras 2horas de reação, as taxas de reação diferentes obtidos para três diferentes álcoois usados, mos-trando que a velocidade da reação é dependente da quantidade de átomos de carbono do ácidousado e da álcool.
0 2 4 6 8 100
20
40
60
80
100
Butirato de etilaButirato de metila
Cov
ersã
o ác
ido
butír
ico
(%)
Tempo (h)
Figura 5.5: Dinâmica da síntese de butirato de etila e metila. Os álcoois usados foram: etanol(�) e metanol (•). Reações foram realizadas usando CALB-NPM Att= 80 U.g−1, at 25°C, 8h, 150 rpm, heptano, 0,5 (metanol) e 0,4 (etanol) mol/L ácido butírico, 1:1 (ácido butírico:(metanol e etanol)). Volume reacional = 1,0 mL. Massa do catalisador = 0,01
Os estudos na literatura (DIAS et al., 1991; MANJON; IBORRA; AROCAS, 1991;DAVE, 2005; RODRIGUEZ-NOGALES; ROURA; CONTRERAS, 2005; ARAGÃO et al.,2011; GROSSO; FERREIRA-DIAS; PIRES-CABRAL, 2012) apresentam a utilização de vá-rias lipases imobilizadas para a produção de butirato de etila. Rodriguez-Nogales, Roura e

69
Contreras (2005) (RODRIGUEZ-NOGALES; ROURA; CONTRERAS, 2005) realizaram a ca-tálise de ácido butírico e etanol por resina acrílica Novozym® 435 em 10 ml de n-heptano, aconcentração de 0,04 mol/L e 0,52 mol/L álcool, obtendo-se uma taxa de conversão de 75% em96 horas. A taxa de conversão de 88% de butirato de etila foi obtida por Aragão et al. (2011),em 3 h de reação, 40 ml de volume reacional, utilizando lipase de Mucor miehei imobilizadaem pérolas de resina comerciais.
O melhor valor de produção volumétrica para o butirato de etila apresentados porDias et al. (1991), Manjon, Iborra e Arocas (1991), Grosso, Ferreira-Dias e Pires-Cabral (2012),Dave (2005), Rodriguez-Nogales, Roura e Contreras (2005), Aragão et al. (2011) (DIAS et al.,1991; MANJON; IBORRA; AROCAS, 1991; GROSSO; FERREIRA-DIAS; PIRES-CABRAL,2012; DAVE, 2005; RODRIGUEZ-NOGALES; ROURA; CONTRERAS, 2005; ARAGÃO etal., 2011) foi de 26,4 µmol/(mL.h), apresentado por Aragão et al. (2011) (ARAGÃO et al.,2011) com lipase de Mucor miehei imobilizadas em pérolas de resina comerciais. O resultadoobtido no nosso trabalho para a produção de butirato de etila por CALB imobilizada em nano-partículas magnéticas foi de 53,7 µmol/(mL.h), duas vezes aquele apresentado em Aragão et al.(2011) (ARAGÃO et al., 2011).
5.3.7 Estabilidade operacional
A estabilidade operacional do sistema imobilizado foi investigada para a síntese debutirato etila e metila, em que os ciclos de reação consecutivos, foram realizados, com reutili-zação do biocatalisador, usando 0,01 g de CALB-NPM e 8 h, 25°C, 150 rpm, 0,4 e 0,5 mol/Lde ácido butírico, 1:1 (ácido butírico: (metanol e etanol)). Volume reacional = 1,0 ml (heptano).As mesmas condições foram utilizadas com lipase B imobilizado na resina acrílica.
0 2 4 6 8 10 1260
65
70
75
80
85
90
95
100
Con
vers
ão d
e ác
ido
butír
ico
(%)
Ciclos
Figura 5.6: Estabilidade operacional de CALB-IMNs(�) e CALB-resina acrílica(•) em síntesede buriato de etila. Reações foram executadas a 25°C, 8 h, 150 rpm, 0,4 mol/L ácido butírico,1:1 (ácido butírico: (metanol e etanol)). Volume reacional = 1,0 mL. Massa do catalisador =0,01
A partir da Figura 5.6, a produção de butirato de metila de conversão foi de 89,6%para a resina de Candida antartica. Como mostrado na Figura 5.7, nas experiências do primeiro

70
ciclo para butirato de etila, a conversão de resinas acrílicas comerciais foi de 95,6% . As me-lhores conversões de substratos obtidos por butirato de metila e etila foram com CALB-NPM,96,8% e 93,9% foram observadas com butirato de etila e metila, respectivamente.
0 2 4 6 8 10 1250
60
70
80
90
100
CALB-NPM CALB-Resina acrílica
Con
vers
ão d
e ác
ido
butír
ico
(%
)
Ciclos
Figura 5.7: Estabilidade operacional de CALB-NPM (�), resina acrílica (•) em síntese de bu-riato de etila. Reações foram executadas a 25°C, 8 h, 150 rpm, 0,4 mol/L ácido butírico, 1:1(ácido butírico: (metanol e etanol)). Volume reacional = 1,0 mL. Massa do catalisador = 0,01
Os estudos da reutilização para os dois catalisadores mostram que a enzima imobi-lizada foi estável durante 9 ciclos de esterificação, ver Figura 5.7 e 5.6. No final do último ciclo(12), CALB-NPM manteve mais do que 74% da sua atividade catalítica e 81% para a resinaacrílica na conversão de butirato de etila, ver Figura 5.7.
A comparação dos resultados com os obtidos na literatura, (GUILLÉN; BENAI-GES; VALERO, 2012; PIRES-CABRAL; FONSECA; FERREIRA-DIAS, 2010; MANJON;IBORRA; AROCAS, 1991; RODRIGUEZ-NOGALES; ROURA; CONTRERAS, 2005), poresterificação do ácido butírico com etanol catalisada por lipases imobilizadas mostraram que asmelhores conversões obtidas nos estudos citados acima, foram obtidos por Guillén, Benaiges eValero (2012) em lipase Rhizopus oryzae imobilizada em octadecil sepabeads, 99,1 % em 24horas, esta conversão foi mantida durante três ciclos consecutivos com a diminuição de 40%após seis ciclos, enquanto que, no presente estudo, a atividade catalítica foi mantida a 93,4%durante 9 ciclos consecutivos de 8 horas cada um mostrando o uso potencial de CALB-NPMem reações de interesse industrial.
5.4 Conclusões
O estudo mostrou que as nanopartículas magnéticas modificadas por APTS e glu-taraldeído promove um suporte capaz de suportar a enzima ativa CALB, mantendo a atividadecatalítica por ciclos de reação consecutivos. Analisando os resultados dos diferentes efeitossobre a bioconversão de ésteres, há uma forte influência do álcool e do ácido e a razão molar desubstratos. A elevada volatilidade do metanol não permite a utilização de temperaturas superio-res. O aumento da razão molar de ácido tem um efeito sobre a atividade enzimática, ocorrendo

71
a desnaturação. O efeito da hidrofobicidade do solvente usado foi muito pronunciada, solven-tes não-polares são mais adequados para a produção de ésteres de reações por CALB-NPM. Ocatalisador estudado neste trabalho, CALB-NPM, promoveu uma síntese eficiente do buriatode etila e metila. As condições ótimas para a síntese de buriato de metila e etila por CALB-NPM resultou na conversão de substrato de 96,8% . O catalisador CALB-NPM mostrou boamanutenção da atividade catalítica durante 12 ciclos consecutivos de reação, estes resultadossão satisfatórios quando comparados com os obtidos com derivado comercial. Assim, o uso deCALB-NPM é particularmente vantajoso para a produção de ésteres.
Agradecimentos
Os autores gostariam de agradecer as agências brasileiras de fomento a pesquisaCNPq e CAPES.

72
6 CONCLUSÃO E PROPOSTAS PARA TRABALHOS FUTUROS
A superfície das nanopartículas magnéticas de ferro foram modificadas por gama-aminopropiltrietoxissilano (APTS) e glutaraldeído. Com base nos resultados obtidos, concluiu-se que o recobrimento de silicato proporcionado pelo APTS garantiu uma proteção a oxidaçãodas nanopartículas pelo ar. Além de evitar a agregação das nanopartículas e permitir a interaçãocom os grupos aldeídos presentes no glutaraldeído. A concentração de glutaraldeído apresentouum ponto importante a ser considerado no processo de reticulação de nanopartículas tratadaspor APTS, devido a grande área superficial do suporte. A concentração de 25 % promoveu umaeficiente imobilização da enzima CALB em nanopartículas magnéticas.
A lipase CALB solúvel e imobilizada foram caracterizadas em relação a estabili-dade térmica, atividade enzimática utilizando como substrato butirato de para-nitrofenila pNPBe quantidade de proteína por volume de solução e massa de suporte. Esses estudos permiti-ram o domínio do processo de imobilização, tendo-se obtido uma estabilidade 20 vezes maiorpara a enzima CALB após imobilização em nanopartículas magnéticas de ferro modificadas porAPTS e glutaraldeído. A melhor condição catalítica foi obtida com 1 hora de imobilização evelocidade de imobilização de 45 rpm (agitação rotacional).
O biocatalisador foi utilizado na produção dos ésteres oleato de etila, butirato demetila e etila, apresentando conversões compatíveis com os presentes na literatura, mostrando-se excelentes no uso em reações consecutivas (reuso). A estabilidade operacional apresentadamostra a potencial aplicação da CALB imobilizada em nanopartículas magnéticas como bioca-talisador heterogêneo em substituição a catalisadores químicos. Outro tópico de interesse estárelacionado à fácil separação e recuperação do biocatalisador por simples aplicação de campomagnético, essa característica é fundamental para a aplicação em processos que visem a dimi-nuição de poluentes.
O estudos apresentados neste trabalho podem ser aprimorados para a obtenção denovas possibilidades, abrangendo a imobilização de outras enzimas com diferentes especifici-dades, novos desenhos de reatores, como por exemplo, reatores com micro canais que permitemo acoplamento da enzima imobilizada em nanopartículas magnéticas, permitindo um fluxo con-tínuo de reagentes e produtos.
Portanto, concluí-se que existe um vasto campo de possibilidades a ser exploradoem trabalhos futuros, seja diretamente ou aplicando a presente proposta em problemas correla-tos.

73
REFERÊNCIAS BIBLIOGRÁFICAS
A, S. M.; WIEGEL, J. Effects of detergents on activity, thermostability and aggregation of twoalkalithermophilic lipases from thermosyntropha lipolytica. The open biochemistry journal,Bentham Science Publishers, v. 4, p. 22, 2010.
ABBAS, H.; COMEAU, L. Aroma synthesis by immobilized lipase from Mucor sp. Enzymeand microbial technology, Elsevier, v. 32, n. 5, p. 589–595, 2003.
ADACHI, D.; HAMA, S.; NAKASHIMA, K.; BOGAKI, T.; OGINO, C.;KONDO, A. Production of biodiesel from plant oil hydrolysates using an asper-gillus oryzae whole-cell biocatalyst highly expressing candida antarctica lipaseb. Bioresource Technology, n. 0, p. –, 2012. ISSN 0960-8524. Disponível em:<http://www.sciencedirect.com/science/article/pii/S0960852412010061>.
ADAMCZAK, M.; BEDNARSKI, W. Enhanced activity of intracellular lipases fromRhizomucor miehei and Yarrowia lipolytica by immobilization on biomass support particles.Process Biochemistry, Elsevier, v. 39, n. 11, p. 1347–1361, 2004.
ADNANI, A.; BASRI, M.; MALEK, E.; SALLEH, A.; RAHMAN, M. A.; CHAIBAKHSH,N.; RAHMAN, R. Optimization of lipase-catalyzed synthesis of xylitol ester by taguchi robustdesign method. Industrial Crops and Products, Elsevier, v. 31, n. 2, p. 350–356, 2010.
ADRIANO, W.; SILVA, J.; GIORDANO, R.; GONÇALVES, L. et al. Stabilization ofpenicillin g acylase by immobilization on glutaraldehyde-activated chitosan. Brazilian Journalof Chemical Engineering, SciELO Brasil, v. 22, n. 4, p. 529–538, 2005.
AHMED, E.; RAGHAVENDRA, T.; MADAMWAR, D. An alkaline lipase from organicsolvent tolerant Acinetobacter sp. eh28: Application for ethyl caprylate synthesis. Bioresourcetechnology, Elsevier, v. 101, n. 10, p. 3628–3634, 2010.
AKOH, C.; CHANG, S.; LEE, G.; SHAW, J. Enzymatic approach to biodiesel production.Journal of Agricultural and Food Chemistry, ACS Publications, v. 55, n. 22, p. 8995–9005,2007.
AL-ZUHAIR, S.; LING, F.; JUN, L. Proposed kinetic mechanism of the production ofbiodiesel from palm oil using lipase. Process Biochemistry, Elsevier, v. 42, n. 6, p. 951–960,2007.
AMABIS, J. M.; MARTHO, G. R. Do gene a proteína. Atualidades biológicas, EditoraModerna, v. 6, p. 1–8, 1997.
ANDRADE, L.; REBELO, L.; NETTO, C.; TOMA, H. Kinetic resolution of a drug precursorby Burkholderia cepacia lipase immobilized by different methodologies on superparamagneticnanoparticles. Journal of Molecular Catalysis B: Enzymatic, Elsevier, v. 66, n. 1, p. 55–62,2010.
ANSARI, S.; HUSAIN, Q. Potential applications of enzymes immobilized on/in nanomaterials: A review. Biotechnology advances, Elsevier, v. 30, n. 3, p. 512–523, 2012.

74
ARAGÃO, V. C.; PORTO, M. R. A.; BURKERT, C. A. V.; KALIL, S. J.; BURKERT, J. F.d. M. Response surface methodology approach for the synthesis of ethyl butyrate. 2011.
ARROYO, M.; SÁNCHEZ-MONTERO, J.; SINISTERRA, J. Thermal stabilization ofimmobilized lipase b from Candida antarctica on different supports: Effect of water activityon enzymatic activity in organic media. Enzyme and Microbial Technology, Elsevier, v. 24,n. 1, p. 3–12, 1999.
ASHTARI, K.; KHAJEH, K.; FASIHI, J.; ASHTARI, P.; RAMAZANI, A.; VALI, H.Silica-encapsulated magnetic nanoparticles: Enzyme immobilization and cytotoxic study.International Journal of Biological Macromolecules, Elsevier, 2012.
BAI, S.; GUO, Z.; LIU, W.; SUN, Y. Resolution of (±)-menthol by immobilized Candidarugosa lipase on superparamagnetic nanoparticles. Food chemistry, Elsevier, v. 96, n. 1, p. 1–7,2006.
BARBOSA, O.; TORRES, R.; ORTIZ, C.; FERNANDEZ-LAFUENTE, R. Ver-satility of glutaraldehyde to immobilize lipases: Effect of the immobilizationprotocol on the properties of lipase B from Candida antarctica. Process Bio-chemistry, v. 47, n. 8, p. 1220–1227, 2012. ISSN 1359-5113. Disponível em:<http://www.sciencedirect.com/science/article/pii/S1359511312001687>.
BARRETO, A.; MAIA, F.; SANTIAGO, V.; RIBEIRO, V.; DENARDIN, J.; MELE, G.;CARBONE, L.; LOMONACO, D.; MAZZETTO, S.; FECHINE, P. Novel ferrofluidscoated with a renewable material obtained from cashew nut shell liquid. Microfluidics andnanofluidics, Springer, p. 1–10, 2012.
Ben Salah, R.; GHAMGHUI, H.; MILED, N.; MEJDOUB, H.; GARGOURI, Y. Production ofbutyl acetate ester by lipase from novel strain of Rhizopus oryzae. Journal of bioscience andbioengineering, J-STAGE, v. 103, n. 4, p. 368–372, 2007.
BETANCOR, L.; LÓPEZ-GALLEGO, F.; HIDALGO, A.; ALONSO-MORALES, N.;MATEO, G. D.-O. C.; FERNÁNDEZ-LAFUENTE, R.; GUISÁN, J. M. Different mechanismsof protein immobilization on glutaraldehyde activated supports: effect of support activationand immobilization conditions. Enzyme and microbial technology, Elsevier, v. 39, n. 4, p.877–882, 2006.
BJÖRKLING, F.; GODTFREDSEN, S.; KIRK, O. The future impact of industrial lipases.Trends in Biotechnology, Elsevier, v. 9, n. 1, p. 360–363, 1991.
BLOOMER, S.; ADLERCREUTZ, P.; MATTIASSON, B. Facile synthesis of fatty acid estersin high yields. Enzyme and microbial technology, Elsevier, v. 14, n. 7, p. 546–552, 1992.
BOYER, C.; WHITTAKER, M.; BULMUS, V.; LIU, J.; DAVIS, T. The design and utilityof polymer-stabilized iron-oxide nanoparticles for nanomedicine applications. NPG AsiaMaterials, Nature Publishing Group, v. 2, n. 1, p. 23–30, 2010.
BRADFORDMM, A. rapid and sensitive method for the quantitation of microgram quantitiesof protein utilizing the principle of protein–dye binding. Anal Biochem, v. 72, n. 7, p. 248–254,1976.

75
BRÍGIDA, A.; PINHEIRO, Á.; FERREIRA, A.; GONÇALVES, L. Immobilization ofcandida antarctica lipase b by adsorption to green coconut fiber. Biotechnology for Fuels andChemicals, Springer, p. 293–307, 2008.
BRUICE, P. Y. Química orgânica. Prentice Hall Brasil, v. 2, 2006.
BRUNO, L.; COELHO, J.; MELO, E.; LIMA-FILHO, J. Characterization of mucor mieheilipase immobilized on polysiloxane-polyvinyl alcohol magnetic particles. World Journal ofMicrobiology and Biotechnology, Springer, v. 21, n. 2, p. 189–192, 2005.
CAMBON, E.; BOURLIEU, C.; SALUM, T.; PIOMBO, G.; DUBREUCQ, E.; VILLENEUVE,P. Ability of Vasconcellea x heilbornii lipase to catalyse the synthesis of alkyl esters fromvegetable oils. Process Biochemistry, Elsevier, v. 44, n. 11, p. 1265–1269, 2009.
CAO, L. Immobilised enzymes: science or art? Current Opinion in Chemical Biology, Elsevier,v. 9, n. 2, p. 217–226, 2005.
CASTRO, H. de; MENDES, A.; SANTOS, J. dos; AGUIAR, C. de. Modificação de óleos egorduras por biotransformação. Química Nova, SciELO Brasil, v. 27, n. 1, p. 146–156, 2004.
CHARLES, S.; POPPLEWELL, J. Properties and applications of magnetic liquids. Endeavour,Elsevier, v. 6, n. 4, p. 153–161, 1982.
CHEN, J. Production of ethyl butyrate using gel-entrapped Candida cylindracea lipase.Journal of fermentation and bioengineering, Elsevier, v. 82, n. 4, p. 404–407, 1996.
CHEN, J.; LIN, W. Sol-gel powders and supported sol-gel polymers for immobilization oflipase in ester synthesis. Enzyme and microbial technology, Elsevier, v. 32, n. 7, p. 801–811,2003.
CLAON, P.; AKOH, C. Effect of reaction parameters on SP435 lipase-catalyzed synthesis ofcitronellyl acetate in organic solvent. Enzyme and Microbial Technology, Elsevier, v. 16, n. 10,p. 835–838, 1994.
CLARK, J. H.; RHODES, C. N. Clean synthesis using porous inorganic solid catalysts andsupported reagents. [S.l.]: Royal Society of Chemistry, 2000.
CORNELL, R.; SCHWERTMANN, U. The iron oxides: structure, properties, reactions,occurrences and uses. [S.l.]: Wiley-vch, 2003.
D’AGOSTINI, D.; GIOIELI, L. Distribuição estereoespecífica de lipídios estruturados a partirde gordura de palma, palmiste e triacilgliceróis de cadeia media. [S.l.]: SciELO Brasil, 2010.
DANDAVATE, V.; JINJALA, J.; KEHARIA, H.; MADAMWAR, D. Production, partialpurification and characterization of organic solvent tolerant lipase from Burkholderiamultivorans v2 and its application for ester synthesis. Bioresource Technology, Elsevier, v. 100,n. 13, p. 3374–3381, 2009.
DANDAVATE, V.; KEHARIA, H.; MADAMWAR, D. Ethyl isovalerate synthesis usingCandida rugosa lipase immobilized on silica nanoparticles prepared in nonionic reversemicelles. Process Biochemistry, Elsevier, v. 44, n. 3, p. 349–352, 2009.

76
DAVE, D. M. R. Esterification in organic solvents by silica gel immobilized candida rugosalipase. In: Larroche, C., Pandey, A., Dussap, C.G. (Eds.), Current Topics on Bioprocesses inFood Industry, Asiatech Press, p. 71–80, 2005.
DENG, X.; FANG, Z.; LIU, Y.; YU, C. Production of biodiesel from Jatropha oil catalyzed bynanosized solid basic catalyst. Energy, Elsevier, v. 36, n. 2, p. 777–784, 2011.
DIAS, S. F.; VILAS-BOAS, L.; CABRAL, J.; FONSECA, M. Production of ethyl butyrateby candida rugosa lipase immobilized in polyurethane. Biocatalysis and Biotransformation,Informa UK Ltd UK, v. 5, n. 1, p. 21–34, 1991.
DIZGE, N.; AYDINER, C.; IMER, D.; BAYRAMOGLU, M.; TANRISEVEN, A.;KESKINLER, B. Biodiesel production from sunflower, soybean, and waste cooking oils bytransesterification using lipase immobilized onto a novel microporous polymer. Bioresourcetechnology, Elsevier, v. 100, n. 6, p. 1983–1991, 2009.
DU, W.; XU, Y.; LIU, D.; ZENG, J. Comparative study on lipase-catalyzed transformationof soybean oil for biodiesel production with different acyl acceptors. Journal of MolecularCatalysis B: Enzymatic, Elsevier, v. 30, n. 3, p. 125–129, 2004.
DUGUET, E.; VASSEUR, S.; MORNET, S.; DEVOISSELLE, J. Magnetic nanoparticles andtheir applications in medicine. Nanomedicine, Future Medicine, v. 1, n. 2, p. 157–168, 2006.
END, N.; SCHÖNING, K.-U. Immobilized catalysts in industrial research and application.Immobilized Catalysts, Springer, p. 241–271, 2004.
ESSAMRI, M.; DEYRIS, V.; COMEAU, L. Optimization of lipase production by Rhizopusoryzae and study on the stability of lipase activity in organic solvents. Journal of Biotechnology,Elsevier, v. 60, n. 1, p. 97–103, 1998.
FABER, K. Biotransformations in organic chemistry: a textbook. [S.l.]: Springer, 2011.
FEDOSOV, S.; BRASK, J.; PEDERSEN, A.; NORDBLAD, M.; WOODLEY, J.; XU, X.Kinetic model of biodiesel production using immobilized lipase Candida antarctica lipase B.Journal of Molecular Catalysis B: Enzymatic, Elsevier, 2012.
FERNANDEZ-LAFUENTE, R.; ARMISÉN, P.; SABUQUILLO, P.; FERNÁNDEZ-LORENTE, G.; M Guisán, J. Immobilization of lipases by selective adsorption on hydrophobicsupports. Chemistry and physics of lipids, Elsevier, v. 93, n. 1, p. 185–197, 1998.
FERNANDEZ-LAFUENTE, R.; ROSELL, C.; RODRIGUEZ, V.; SANTANA, C.; SOLER,G.; BASTIDA, A.; GUISAN, J. Preparation of activated supports containing low pK aminogroups. A new tool for protein immobilization via the carboxyl coupling method. Enzyme andmicrobial technology, Elsevier, v. 15, n. 7, p. 546–550, 1993.
FERNANDEZ-LORENTE, G.; FERNÁNDEZ-LAFUENTE, R.; PALOMO, J.; MATEO, C.;BASTIDA, A.; COCA, J.; HARAMBOURE, T.; HERNANDEZ-JUSTIZ, O.; TERRENI,M.; GUISÁN, J. Biocatalyst engineering exerts a dramatic effect on selectivity of hydrolysiscatalyzed by immobilized lipases in aqueous medium. Journal of Molecular Catalysis B:Enzymatic, Elsevier, v. 11, n. 4, p. 649–656, 2001.

77
FERNANDEZ-LORENTE, G.; PALOMO, J.; COCCA, J.; MATEO, C.; MORO, P.;TERRENI, M.; FERNANDEZ-LAFUENTE, R.; GUISAN, J. Regio-selective deprotection ofperacetylated sugars via lipase hydrolysis. Tetrahedron, Elsevier, v. 59, n. 30, p. 5705–5711,2003.
FORESTI, M.; FERREIRA, M. Solvent-free ethyl oleate synthesis mediated by lipase fromCandida antarctica B adsorbed on polypropylene powder. Catalysis today, Elsevier, v. 107, p.23–30, 2005.
FUKUDA, H.; KONDO, A.; NODA, H. Biodiesel fuel production by transesterification of oils.Journal of bioscience and bioengineering, Elsevier, v. 92, n. 5, p. 405–416, 2001.
GANDHI, N.; SAWANT, S.; JOSHI, J. Specificity of a lipase in ester synthesis: effect ofalcohol. Biotechnology progress, Wiley Online Library, v. 11, n. 3, p. 282–287, 2008.
GARCIA-MORENO, B. Probing structural and physical basis of protein energetics linked toprotons and salt. Methods in Enzymology, v. 259, p. 512–538, 1995.
GHAMGUI, H.; KARRA-CHAÂBOUNI, M.; GARGOURI, Y. 1-Butyl oleate synthesisby immobilized lipase from Rhizopus oryzae: a comparative study between n-hexane andsolvent-free system. Enzyme and Microbial Technology, Elsevier, v. 35, n. 4, p. 355–363, 2004.
GILLIES, B.; YAMAZAKI, H.; ARMSTRONG, D. Production of flavor esters by immobilizedlipase. Biotechnology Letters, Springer, v. 9, n. 10, p. 709–714, 1987.
GIRALDO, L. L.; LAGUERRE, M.; LECOMTE, J.; FIGUEROA-ESPINOZA, M.; BAROUH,N.; BARÉA, B.; VILLENEUVE, P. Lipase-catalyzed synthesis of chlorogenate fatty esters insolvent-free medium. Enzyme and Microbial Technology, Elsevier, v. 41, n. 6, p. 721–726,2007.
GOMES, F.; PEREIRA, E.; CASTRO, H. de. Immobilization of lipase on chitin and its usein nonconventional biocatalysis. Biomacromolecules, ACS Publications, v. 5, n. 1, p. 17–23,2004.
GROSSO, C.; FERREIRA-DIAS, S.; PIRES-CABRAL, P. Modelling and optimization ofethyl butyrate production catalysed by Rhizopus oryzae lipase. Journal of Food Engineering,Elsevier, 2012.
GUILLÉN, M.; BENAIGES, M.; VALERO, F. Biosynthesis of ethyl butyrate by immobilizedrecombinant Rhizopus oryzae lipase expressed in Pichia pastoris. Biochemical EngineeringJournal, Elsevier, 2012.
GUISAN, J. M. Immobilization of enzymes and cells. [S.l.]: Springer, 2006.
GUPTA, K.; JABRAIL, F. Effects of degree of deacetylation and cross-linking on physicalcharacteristics, swelling and release behavior of chitosan microspheres. Carbohydratepolymers, Elsevier, v. 66, n. 1, p. 43–54, 2006.
HA, S.; LAN, M.; LEE, S.; HWANG, S.; KOO, Y. Lipase-catalyzed biodiesel productionfrom soybean oil in ionic liquids. Enzyme and Microbial Technology, Elsevier, v. 41, n. 4, p.480–483, 2007.

78
HALLING, P. Effects of water on equilibria catalysed by hydrolytic enzymes in biphasicreaction systems. Enzyme and Microbial Technology, Elsevier, v. 6, n. 11, p. 513–516, 1984.
HALLING, P. High-affinity binding of water by proteins is similar in air and in organicsolvents. Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular Enzymology,Elsevier, v. 1040, n. 2, p. 225–228, 1990.
HALLING, P. J. Biocatalysis in low-water media: understanding effects of reaction conditions.Current opinion in chemical biology, Elsevier, v. 4, n. 1, p. 74–80, 2000.
Hari Krishna, S.; KARANTH, N. Lipase-catalyzed synthesis of isoamyl butyrate: a kineticstudy. Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular Enzymology,Elsevier, v. 1547, n. 2, p. 262–267, 2001.
HASAN, F.; SHAH, A.; HAMEED, A. Industrial applications of microbial lipases. Enzymeand Microbial Technology, Elsevier, v. 39, n. 2, p. 235–251, 2006.
HE, Q.; XU, Y.; TENG, Y.; WANG, D. Biodiesel production catalyzed by whole-cell lipasefrom Rhizopus chinensis. Chinese Journal of Catalysis, Elsevier, v. 29, n. 1, p. 41–46, 2008.
HRDINA, A.; LAI, E.; LI, C.; SADI, B.; KRAMER, G. A comparative study of magnetictransferability of superparamagnetic nanoparticles. Journal of Magnetism and MagneticMaterials, Elsevier, v. 322, n. 17, p. 2622–2627, 2010.
HU, B.; PAN, J.; YU, H.; LIU, J.; XU, J. Immobilization of Serratia marcescens lipaseonto amino-functionalized magnetic nanoparticles for repeated use in enzymatic synthesis ofDiltiazem intermediate. Process Biochemistry, Elsevier, v. 44, n. 9, p. 1019–1024, 2009.
HUANG, L.; CHENG, Z. Immobilization of lipase on chemically modified bimodal ceramicfoams for olive oil hydrolysis. Chemical Engineering Journal, Elsevier, v. 144, n. 1, p.103–109, 2008.
HUANG, S.; LIAO, M.; CHEN, D. Direct binding and characterization of lipase onto magneticnanoparticles. Biotechnology progress, Wiley Online Library, v. 19, n. 3, p. 1095–1100, 2008.
HUNG, T.; GIRIDHAR, R.; CHIOU, S.; WU, W. Binary immobilization of Candida rugosalipase on chitosan. Journal of Molecular Catalysis B: Enzymatic, Elsevier, v. 26, n. 1, p. 69–78,2003.
IDRIS, A.; BUKHARI, A. Immobilized candida antarctica lipase b: Hydration, stripping offand application in ring opening polyester synthesis. Biotechnology advances, Elsevier, 2011.
ISO, M.; CHEN, B.; EGUCHI, M.; KUDO, T.; SHRESTHA, S. Production of biodiesel fuelfrom triglycerides and alcohol using immobilized lipase. Journal of Molecular Catalysis B:Enzymatic, Elsevier, v. 16, n. 1, p. 53–58, 2001.
JAEGER, K.; DIJKSTRA, B.; REETZ, M. Bacterial biocatalysts: molecular biology,three-dimensional structures, and biotechnological applications of lipases. Annual Reviewsin Microbiology, Annual Reviews 4139 El Camino Way, PO Box 10139, Palo Alto, CA94303-0139, USA, v. 53, n. 1, p. 315–351, 1999.

79
JEGANNATHAN, K.; ABANG, S.; PONCELET, D.; CHAN, E.; RAVINDRA, P. Productionof biodiesel using immobilized lipase-a critical review. Critical reviews in biotechnology,Informa UK Ltd London, UK, v. 28, n. 4, p. 253–264, 2008.
JHONSSON, B.; LUND, M.; SILVA, F. L. B. da. Electrostatics in macromolecular solutions. .Food Colloids: Self-assembly and Material Science, v. 302, p. 129–154, 2007.
JIANG, Y.; GUO, C.; XIA, H.; MAHMOOD, I.; LIU, C.; LIU, H. Magnetic nanoparticlessupported ionic liquids for lipase immobilization: Enzyme activity in catalyzing esterification.Journal of Molecular Catalysis B: Enzymatic, Elsevier, v. 58, n. 1, p. 103–109, 2009.
JIN, Z.; NTWALI, J.; HAN, S.; ZHENG, S.; LIN, Y. Production of flavor esters catalyzed byCALB-displaying Pichia pastoris whole-cells in a batch reactor. Journal of Biotechnology,Elsevier, 2012.
JOHNSON, A.; ZAWADZKA, A.; DEOBALD, L.; CRAWFORD, R.; PASZCZYNSKI,A. Novel method for immobilization of enzymes to magnetic nanoparticles. Journal ofNanoparticle Research, Springer, v. 10, n. 6, p. 1009–1025, 2008.
JUNIOR, M. A. S. Obtenção e caracterização de nanopartículas magnéticas inseridas emmateriais carbonosos porosos a partir da decomposição do pentacarbonil ferro. Programa dePósGradução em Física, Universidade Federal do Espírito Santo, Centro de Ciências Exatas,2009.
KAIEDA, M.; SAMUKAWA, T.; MATSUMOTO, T.; BAN, K.; KONDO, A.; SHIMADA, Y.;NODA, H.; NOMOTO, F.; OHTSUKA, K.; IZUMOTO, E. et al. Biodiesel fuel productionfrom plant oil catalyzed by Rhizopus oryzae lipase in a water-containing system without anorganic solvent. Journal of bioscience and bioengineering, Elsevier, v. 88, n. 6, p. 627–631,1999.
KANASAWUD, P.; PHUTRAKUL, S.; BLOOMER, S.; ADLERCREUTZ, P.; MATTIASSON,B. Triglyceride interesterification by lipases. 3. Alcoholysis of pure triglycerides. Enzyme andmicrobial technology, Elsevier, v. 14, n. 12, p. 959–965, 1992.
KAVAS, H.; KASAPOGLU, N.; BAYKAL, A.; KÖSEOGLU, Y. Characterization of NiFe2O4nanoparticles synthesized by various methods. Chemical Papers, Springer, v. 63, n. 4, p.450–455, 2009.
KAYA, F.; HEITMANN, J.; JOYCE, T. Cellulase binding to cellulose fibers in high shearfields. Journal of biotechnology, Elsevier, v. 36, n. 1, p. 1–10, 1994.
KIM, H.; YOUN, S.; SHIN, C. Lipase-catalyzed synthesis of sorbitol–fatty acid esters atextremely high substrate concentrations. Journal of biotechnology, Elsevier, v. 123, n. 2, p.174–184, 2006.
KIM, J.; JIA, H.; WANG, P. Challenges in biocatalysis for enzyme-based biofuel cells.Biotechnology advances, Elsevier, v. 24, n. 3, p. 296–308, 2006.
KIM, M.; HAM, H.; OH, S.; PARK, H.; CHANG, H.; CHOI, S. Immobilization of Mucorjavanicus lipase on effectively functionalized silica nanoparticles. Journal of MolecularCatalysis B: Enzymatic, Elsevier, v. 39, n. 1, p. 62–68, 2006.

80
KIM, M.; LEE, S.; RYU, D.; REESE, E. Surface deactivation of cellulase and its prevention.Enzyme and Microbial Technology, Elsevier, v. 4, n. 2, p. 99–103, 1982.
KIRK, O.; CHRISTENSEN, M. Lipases from candida a ntarctica: Unique biocatalysts froma unique origin. Organic Process Research & Development, ACS Publications, v. 6, n. 4, p.446–451, 2002.
KISS, M.; SEFANOVITS-BÁNYAI, É.; TOTH, A.; BOROSS, L. Extractive synthesis ofethyl-oleate using alginate gel co-entrapped yeast cells and lipase enzyme. Engineering in lifesciences, Wiley Online Library, v. 4, n. 5, p. 460–464, 2004.
KONWARH, R.; KARAK, N.; RAI, S.; MUKHERJEE, A. Polymer-assisted iron oxidemagnetic nanoparticle immobilized keratinase. Nanotechnology, IOP Publishing, v. 20, n. 22,p. 225107, 2009.
KORDEL, M.; HOFMANN, B.; SCHOMBURG, D.; SCHMID, R. Extracellular lipase ofPseudomonas sp. strain ATCC 21808: purification, characterization, crystallization, andpreliminary X-ray diffraction data. Journal of bacteriology, Am Soc Microbiol, v. 173, n. 15,p. 4836–4841, 1991.
KOUASSI, G.; IRUDAYARAJ, J.; MCCARTY, G. et al. Examination of cholesterol oxidaseattachment to magnetic nanoparticles. J Nanobiotechnology, v. 3, n. 1, p. 1, 2005.
KRIEGER, N.; BHATNAGAR, T.; BARATTI, J.; BARON, A.; LIMA, V. de; MITCHELL,D. Non-aqueous biocatalysis in heterogeneous solvent systems. Food Technology andBiotechnology, v. 42, n. 4, p. 279–286, 2004.
KUMAR, R.; MODAK, J.; MADRAS, G. Effect of the chain length of the acid on theenzymatic synthesis of flavors in supercritical carbon dioxide. Biochemical engineeringjournal, Elsevier, v. 23, n. 3, p. 199–202, 2005.
LAANE, C.; BOEREN, S.; VOS, K.; VEEGER, C. Rules for optimization of biocatalysisin organic solvents. Biotechnology and Bioengineering, Wiley Online Library, v. 30, n. 1, p.81–87, 1987.
LEI, L.; BAI, Y.; LI, Y.; YI, L.; YANG, Y.; XIA, C. Study on immobilization of lipase ontomagnetic microspheres with epoxy groups. Journal of Magnetism and Magnetic Materials,Elsevier, v. 321, n. 4, p. 252–258, 2009.
LIAQUAT, M.; APENTEN, R. Synthesis of low molecular weight flavor esters using plantseedling lipases in organic media. Journal of food science, Wiley Online Library, v. 65, n. 2, p.295–299, 2008.
LIMA, V.; KRIEGER, N.; MITCHELL, D.; FONTANA, J. Activity and stability of a crudelipase from Penicillium aurantiogriseum in aqueous media and organic solvents. Biochemicalengineering journal, Elsevier, v. 18, n. 1, p. 65–71, 2004.
LIU, Y.; ZHANG, X.; TAN, H.; YAN, Y.; HAMEED, B. Effect of pretreatment by differentorganic solvents on esterification activity and conformation of immobilized Pseudomonascepacia lipase. Process Biochemistry, Elsevier, v. 45, n. 7, p. 1176–1180, 2010.

81
LÓPEZ-GALLEGO, F.; BETANCOR, L.; MATEO, C.; HIDALGO, A.; ALONSO-MORALES, N.; DELLAMORA-ORTIZ, G.; GUISÁN, J.; FERNÁNDEZ-LAFUENTE,R. Enzyme stabilization by glutaraldehyde crosslinking of adsorbed proteins on aminatedsupports. Journal of biotechnology, Elsevier, v. 119, n. 1, p. 70–75, 2005.
LOZANO, P.; PÉREZ-MARIN, A.; DIEGO, T. D.; GÓMEZ, D.; PAOLUCCI-JEANJEAN,D.; BELLEVILLE, M.; RIOS, G.; IBORRA, J. Active membranes coated with immobilizedCandida antarctica lipase b: preparation and application for continuous butyl butyratesynthesis in organic media. Journal of membrane science, Elsevier, v. 201, n. 1, p. 55–64, 2002.
LU, J.; NIE, K.; WANG, F.; TAN, T. Immobilized lipase Candida sp. 99-125 catalyzedmethanolysis of glycerol trioleate: Solvent effect. Bioresource technology, Elsevier, v. 99,n. 14, p. 6070–6074, 2008.
LUND M.; JHONSSON, B. On the charge regulation of proteins. Biochemistry, v. 44, p.5722–5727, 2005.
MACRAE, A. Lipase-catalyzed interesterification of oils and fats. Journal of the American OilChemists’ Society, Springer, v. 60, n. 2, p. 291–294, 1983.
MADALOZZO, A. Síntese de ésteres etílicos utilizando uma lipase recombinante de rhizopusoryzae. 2011.
MAHAPATRA, P.; KUMARI, A.; Kumar Garlapati, V.; BANERJEE, R.; NAG, A. Enzymaticsynthesis of fruit flavor esters by immobilized lipase from Rhizopus oligosporus optimizedwith response surface methodology. Journal of Molecular Catalysis B: Enzymatic, Elsevier,v. 60, n. 1, p. 57–63, 2009.
MANJON, A.; IBORRA, J.; AROCAS, A. Short-chain flavour ester synthesis by immobilizedlipase in organic media. Biotechnology letters, Springer, v. 13, n. 5, p. 339–344, 1991.
MARCHETTI, J.; ERRAZU, A. Comparison of different heterogeneous catalysts and differentalcohols for the esterification reaction of oleic acid. Fuel, Elsevier, v. 87, n. 15, p. 3477–3480,2008.
MARCHETTI, J.; MIGUEL, V.; ERRAZU, A. Possible methods for biodiesel production.Renewable and Sustainable Energy Reviews, Elsevier, v. 11, n. 6, p. 1300–1311, 2007.
MARIA, P. Dominguez de; CARBONI-OERLEMANS, C.; TUIN, B.; BARGEMAN, G.;MEER, A. van der; GEMERT, R. van. Biotechnological applications of Candida antarcticalipase A: State-of-the-art. Journal of molecular catalysis b: enzymatic, Elsevier, v. 37, n. 1, p.36–46, 2005.
MATEO, C.; PALOMO, J.; FERNANDEZ-LORENTE, G.; GUISAN, J.; FERNANDEZ-LAFUENTE, R. Improvement of enzyme activity, stability and selectivity via immobilizationtechniques. Enzyme and Microbial Technology, Elsevier, v. 40, n. 6, p. 1451–1463, 2007.
MATTHEN, J.; GURD, F. R. N.; E., B.; GARCIA-MORENO, M. A.; FLANAGAN, K. L.;MARCH; SHIRE, S. J. pH-dependent processes in proteins. CRC Critical Reviews inBiochemistry. Methods in Enzymology, v. 18, p. 91–197, 1985.
MAUGARD, T.; LEGOY, M. Enzymatic synthesis of derivatives of vitamin a in organic media.Journal of Molecular Catalysis B: Enzymatic, Elsevier, v. 8, n. 4, p. 275–280, 2000.

82
MCCABE, R.; RODGER, A.; TAYLOR, A. A study of the secondary structure of Candidaantarctica lipase B using synchrotron radiation circular dichroism measurements. Enzyme andmicrobial technology, Elsevier, v. 36, n. 1, p. 70–74, 2005.
MENDES, A.; CASTRO, H. de; RODRIGUES, D. de S.; ADRIANO, W.; TARDIOLI, P.;MAMMARELLA, E.; GIORDANO, R. de C.; GIORDANO, R. de L. Multipoint covalentimmobilization of lipase on chitosan hybrid hydrogels: influence of the polyelectrolytecomplex type and chemical modification on the catalytic properties of the biocatalysts. Journalof industrial microbiology & biotechnology, Springer, v. 38, n. 8, p. 1055–1066, 2011.
MENDES, A. A.; OLIVEIRA, P. C.; VÉLEZ, A. M.; GIORDANO, R. C.; GIORDANO, R.d. L.; CASTRO, H. F. de. Evaluation of immobilized lipases on poly-hydroxybutyrate beadsto catalyze biodiesel synthesis. International Journal of Biological Macromolecules, Elsevier,v. 50, n. 3, p. 503–511, 2012.
MERBACH, A.; TÓTH, É. The chemistry of contrast agents in medical magnetic resonanceimaging. [S.l.]: Wiley Chichester (W. Sx.) etc., 2001.
MOGENSEN, J. E.; SEHGAL, P.; OTZEN, D. E. Activation, inhibition, and destabilization ofthermomyces lanuginosus lipase by detergents. Biochemistry, ACS Publications, v. 44, n. 5, p.1719–1730, 2005.
MOSSOBA, M.; KRAMER, J.; DELMONTE, P.; YURAWECZ, M.; RADER, J. Officialmethods for the determination of trans fat. [S.l.]: AOCS Press Champaign, IL, 2003.
MOZHAEV, V.; MARTINEK, K. Inactivation and reactivation of proteins (enzymes). Enzymeand Microbial Technology, Elsevier, v. 4, n. 5, p. 299–309, 1982.
MUSSATTO, S.; FERNANDES, M.; MILAGRES, A. Enzimas: poderosa ferramenta naindustria. Ciência Hoje, SOCIEDADE BRASILEIRA PARA O PROGRESSO DA CIENCIA,v. 242, p. 28, 2007.
NAMDEO, M.; BAJPAI, S. Immobilization of α-amylase onto cellulose-coated magnetite(ccm) nanoparticles and preliminary starch degradation study. Journal of Molecular CatalysisB: Enzymatic, Elsevier, v. 59, n. 1, p. 134–139, 2009.
NETTO, C.; ANDRADE, L.; TOMA, H. Enantioselective transesterification catalysis byCandida antarctica lipase immobilized on superparamagnetic nanoparticles. Tetrahedron:Asymmetry, Elsevier, v. 20, n. 19, p. 2299–2304, 2009.
NETTO, C.; TOMA, H.; ANDRADE, L. Superparamagnetic nanoparticles as versatile carriersand supporting materials for enzymes. Journal of Molecular Catalysis B: Enzymatic, Elsevier,2012.
NORDBLAD, M.; ADLERCREUTZ, P. Effects of acid concentration and solvent choice onenzymatic acrylation by Candida antarctica lipase B. Journal of biotechnology, Elsevier,v. 133, n. 1, p. 127–133, 2008.
NOUREDDINI, H.; GAO, X.; PHILKANA, R. Immobilized pseudornonas cepacia lipase forbiodiesel fuel production from soybean oil. Papers in Biomaterials, p. 19, 2004.
NOVOZYMES. Enzymes. jun. 2011. Disponível em:<http://www.novozymes.com/en/Pages/default.aspx>.

83
NOZAKI, Y. T. Examination of titration behavior. Methods in Enzymology, v. 11, p. 715–734,1967.
OGNJANOVIC, N.; BEZBRADICA, D.; KNEZEVIC-JUGOVIC, Z. et al. Enzymaticconversion of sunflower oil to biodiesel in a solvent-free system: process optimization and theimmobilized system stability. Bioresource technology, v. 100, n. 21, p. 5146, 2009.
PABAI, F.; KERMASHA, S.; MORIN, A. Lipase from pseudomonas fragi crda 323: partialpurification, characterization and interesterification of butter fat. Applied microbiology andbiotechnology, Springer, v. 43, n. 1, p. 42–51, 1995.
PALMER, T. Understanding enzymes. 4. ed. [S.l.]: Oxford: Prentice Hall, 1995.
PALOMO, J.; SEGURA, R.; MATEO, C.; TERRENI, M.; GUISAN, J.; FERNÁNDEZ-LAFUENTE, R. Synthesis of enantiomerically pure glycidol via a fully enantioselectivelipase-catalyzed resolution. Tetrahedron: Asymmetry, Elsevier, v. 16, n. 4, p. 869–874, 2005.
PARK, H.; CHI, Y. M. The enhancement of electrostriction caused by lowering the solventdielectric constant leads to the decrease of activation energy in trypsin catalysis. Biochimica etBiophysica Acta (BBA)-General Subjects, Elsevier, v. 1568, n. 1, p. 53–59, 2001.
PEREIRA, G. Estudo da imobilização multipontual da penicilina g acilase em sílica ativadacom grupos glioxil. Master’s Thesis, Universidade Federal de São Carlos, 1996.
PIRES-CABRAL, P.; FONSECA, M. da; FERREIRA-DIAS, S. Esterification activity andoperational stability of Candida rugosa lipase immobilized in polyurethane foams in theproduction of ethyl butyrate. Biochemical Engineering Journal, Elsevier, v. 48, n. 2, p.246–252, 2010.
PIROZZI, D.; GRECO, G. Activity and stability of lipases in the synthesis of butyl lactate.Enzyme and microbial technology, Elsevier, v. 34, n. 2, p. 94–100, 2004.
QI, L.; XU, Z.; JIANG, X.; HU, C.; ZOU, X. Preparation and antibacterial activity of chitosannanoparticles. Carbohydrate Research, Elsevier, v. 339, n. 16, p. 2693–2700, 2004.
RANGANATHAN, S.; NARASIMHAN, S.; MUTHUKUMAR, K. An overview of enzymaticproduction of biodiesel. Bioresource Technology, Elsevier, v. 99, n. 10, p. 3975–3981, 2008.
REBELO, L.; NETTO, C.; TOMA, H.; ANDRADE, L. et al. Enzymatic kinetic resolution of(rs)-1-(phenyl) ethanols by burkholderia cepacia lipase immobilized on magnetic nanoparticles.J. Braz. Chem. Soc, SciELO Brasil, v. 21, n. 8, p. 1537–1542, 2010.
RODRIGUES, D.; MENDES, A.; ADRIANO, W.; GONÇALVES, L.; GIORDANO, R.Multipoint covalent immobilization of microbial lipase on chitosan and agarose activatedby different methods. Journal of Molecular Catalysis B: Enzymatic, Elsevier, v. 51, n. 3, p.100–109, 2008.
RODRIGUEZ-NOGALES, J.; ROURA, E.; CONTRERAS, E. Biosynthesis of ethyl butyrateusing immobilized lipase: a statistical approach. Process Biochemistry, Elsevier, v. 40, n. 1, p.63–68, 2005.

84
ROGALSKA, E.; CUDREY, C.; FERRATO, F.; VERGER, R. Stereoselective hydrolysis oftriglycerides by animal and microbial lipases. Chirality, Wiley Online Library, v. 5, n. 1, p.24–30, 2004.
ROMERO, M.; CALVO, L.; ALBA, C.; HABULIN, M.; PRIMOŽIC, M.; KNEZ, ž. Enzymaticsynthesis of isoamyl acetate with immobilized Candida antarctica lipase in supercriticalcarbon dioxide. The Journal of supercritical fluids, Elsevier, v. 33, n. 1, p. 77–84, 2005.
ROSSET, I. Produção de biodiesel empregando biocatálise via reaçõeses de esterificação etransesterificação. Biblioteca Digital de Teses e Dissertações da USP, 2011.
ROYON, D.; DAZ, M.; ELLENRIEDER, G.; LOCATELLI, S. Enzymatic production ofbiodiesel from cotton seed oil using t-butanol as a solvent. Bioresource Technology, Elsevier,v. 98, n. 3, p. 648–653, 2007.
RUIZ-HERNANDEZ, E.; LOPEZ-NORIEGA, A.; ARCOS, D.; VALLET-REGÍ, M.Mesoporous magnetic microspheres for drug targeting. Solid State Sciences, Elsevier, v. 10,n. 4, p. 421–426, 2008.
SALIS, A.; SOLINAS, V.; MONDUZZI, M. Wax esters synthesis from heavy fraction ofsheep milk fat and cetyl alcohol by immobilised lipases. Journal of Molecular Catalysis B:Enzymatic, Elsevier, v. 21, n. 4, p. 167–174, 2003.
SAMUKAWA, T.; KAIEDA, M.; MATSUMOTO, T.; BAN, K.; KONDO, A.; SHIMADA, Y.;NODA, H.; FUKUDA, H. Pretreatment of immobilized Candida antarctica lipase for biodieselfuel production from plant oil. Journal of bioscience and bioengineering, Elsevier, v. 90, n. 2,p. 180–183, 2000.
SÁNCHEZ-OTERO, M.; QUINTANA-CASTRO, R.; MORA-GONZÁLEZ, P.; MÁRQUEZ-MOLINA, O.; VALERIO-ALFARO, G.; OLIART-ROS, R. Enzymatic reactions and synthesisof n-butyl caproate: esterification, transesterification and aminolysis using a recombinantlipase from geobacillus thermoleovorans ccr11. Environmental technology, Taylor & Francis,v. 31, n. 10, p. 1101–1106, 2010.
SCHøNHEYDER, F.; VOLQVARTZ, K. The gastric lipase in man. Acta PhysiologicaScandinavica, Blackwell Publishing Ltd, v. 11, n. 4, p. 349–360, 1946. ISSN 1365-201X.Disponível em: <http://dx.doi.org/10.1111/j.1748-1716.1946.tb00356.x>.
SELMI, B.; GONTIER, E.; ERGAN, F.; THOMAS, D. Enzymatic synthesis of tricaprylin in asolvent-free system: lipase regiospecificity as controlled by glycerol adsorption on silica gel.Biotechnology techniques, Springer, v. 11, n. 8, p. 543–547, 1997.
SHAW, S.-Y.; CHEN, Y.-J.; OU, J.-J.; HO, L. Preparation and characterization of Pseudomonasputida esterase immobilized on magnetic nanoparticles. Enzyme and microbial technology,Elsevier, v. 39, n. 5, p. 1089–1095, 2006.
SHI, Y.; DING, J.; LIU, X.; WANG, J. NiFe2O4 ultrafine particles prepared by co-precipitation/ mechanical alloying. Journal of Magnetism and Magnetic Materials, Elsevier, v. 205, n. 2, p.249–254, 1999.

85
SHIMADA, Y.; WATANABE, Y.; SAMUKAWA, T.; SUGIHARA, A.; NODA, H.; FUKUDA,H.; TOMINAGA, Y. Conversion of vegetable oil to biodiesel using immobilized candidaantarctica lipase. Journal of the American Oil Chemists’ Society, Springer, v. 76, n. 7, p.789–793, 1999.
SHIMADA, Y.; WATANABE, Y.; SUGIHARA, A.; TOMINAGA, Y. Enzymatic alcoholysisfor biodiesel fuel production and application of the reaction to oil processing. Journal ofMolecular Catalysis B: Enzymatic, Elsevier, v. 17, n. 3, p. 133–142, 2002.
SILVA, J.; MACEDO, G.; RODRIGUES, D.; GIORDANO, R.; GONÇALVES, L.Immobilization of Candida antarctica lipase B by covalent attachment on chitosan-basedhydrogels using different support activation strategies. Biochemical Engineering Journal,Elsevier, v. 60, p. 16 – 24, 2012. ISSN 1369-703X.
SOO, E.; SALLEH, A.; BASRI, M.; RAHMAN, R.; KAMARUDDIN, K. Response surfacemethodological study on lipase-catalyzed synthesis of amino acid surfactants. ProcessBiochemistry, Elsevier, v. 39, n. 11, p. 1511–1518, 2004.
SOUZA, M. F. de. Suavização Hiperbólica Aplicada à Otimização de Geometria Molecular. J/COPPE/ Programa de Engenharia de Sistemas e Computação, p. 1–99, 2010.
STRAATHOF, A.; PANKE, S.; SCHMID, A. et al. The production of fine chemicals bybiotransformations. Current opinion in biotechnology, London, UK: Current Biology, c1990-,v. 13, n. 6, p. 548–556, 2002.
SUN, S.; XU, Y.; WANG, D. Novel minor lipase from Rhizopus chinensis during solid-statefermentation: Biochemical characterization and its esterification potential for ester synthesis.Bioresource technology, Elsevier, v. 100, n. 9, p. 2607–2612, 2009.
TALUKDER, M.; WU, J.; Van Nguyen, T.; FEN, N.; MELISSA, Y. Novozym 435 forproduction of biodiesel from unrefined palm oil: comparison of methanolysis methods. Journalof Molecular Catalysis B: Enzymatic, Elsevier, v. 60, n. 3, p. 106–112, 2009.
TAN, T.; LU, J.; NIE, K.; DENG, L.; WANG, F. Biodiesel production with immobilized lipase:a review. Biotechnology Advances, Elsevier, v. 28, n. 5, p. 628–634, 2010.
TANG, T.; FAN, H.; AI, S.; HAN, R.; QIU, Y. Hemoglobin (hb) immobilized on amino-modified magnetic nanoparticles for the catalytic removal of bisphenol a. Chemosphere,Elsevier, v. 83, n. 3, p. 255–264, 2011.
TARTAJ, P.; MORALES, M.; GONZALEZ-CARRENO, T.; VEINTEMILLAS-VERDAGUER, S.; SERNA, C. Advances in magnetic nanoparticles for biotechnologyapplications. Journal of magnetism and magnetic materials, Elsevier, v. 290, p. 28–34, 2005.
TARTAJ, P.; MORALES, M. del P.; VEINTEMILLAS-VERDAGUER, S.; GONZÁLEZ-CARRENO, T.; SERNA, C. The preparation of magnetic nanoparticles for applications inbiomedicine. Journal of Physics D: Applied Physics, IOP Publishing, v. 36, n. 13, p. R182,2003.
THEIL, F. Enhancement of selectivity and reactivity of lipases by additives. Tetrahedron,[Oxford; New York: Pergamon Press, 1957-, v. 56, n. 19, p. 2905–2920, 2000.

86
TRAN, D.; CHEN, C.; CHANG, J. Immobilization of Burkholderia sp. lipase on a ferric silicananocomposite for biodiesel production. Journal of Biotechnology, Elsevier, 2012.
TREVAN, M. D. Immobilized enzymes. John Wiley & Sons Ltd., first ed., 1980.
TRODLER, P.; PLEISS, J. Modeling structure and flexibility of candida antarctica lipase b inorganic solvents. BMC structural biology, BioMed Central Ltd, v. 8, n. 1, p. 9, 2008.
UNDURRAGA, D.; MARKOVITS, A.; ERAZO, S. Cocoa butter equivalent through enzymicinteresterification of palm oil midfraction. Process Biochemistry, Elsevier, v. 36, n. 10, p.933–939, 2001.
UPPENBERG, J.; HANSEN, M.; PATKAR, S.; JONES, T. The sequence, crystal structuredetermination and refinement of two crystal forms of lipase b from Candida antarctica.Structure, Elsevier, v. 2, n. 4, p. 293–308, 1994.
VILLENEUVE, P. Lipases in lipophilization reactions. Biotechnology advances, Elsevier,v. 25, n. 6, p. 515–536, 2007.
VOET, D.; VOET, J. Biochemistry. 2. ed. [S.l.]: Wiley, 1995.
WARSHEL, A.; NARAY-SZABO, G.; SUSSMAN, F.; HWANG, J. How do serine proteasesreally work? Biochemistry, ACS Publications, v. 28, n. 9, p. 3629–3637, 1989.
WATANABE, Y.; MIYAWAKI, Y.; ADACHI, S.; NAKANISHI, K.; MATSUNO, R.Equilibrium constant for lipase-catalyzed condensation of mannose and lauric acid inwater-miscible organic solvents. Enzyme and microbial technology, Elsevier, v. 29, n. 8, p.494–498, 2001.
WELSH, F.; WILLIAMS, R.; DAWSON, K. Lipase mediated synthesis of low molecularweight flavor esters. Journal of food science, Wiley Online Library, v. 55, n. 6, p. 1679–1682,2006.
XIE, W.; MA, N. Immobilized lipase on fe3o4 nanoparticles as biocatalyst for biodieselproduction. Energy & Fuels, ACS Publications, v. 23, n. 3, p. 1347–1353, 2009.
XIE, W.; MA, N. Enzymatic transesterification of soybean oil by using immobilized lipase onmagnetic nano-particles. Biomass and Bioenergy, Elsevier, v. 34, n. 6, p. 890–896, 2010.
XU, Y.; WANG, D.; MU, X.; ZHAO, G.; ZHANG, K. Biosynthesis of ethyl esters ofshort-chain fatty acids using whole-cell lipase from Rhizopus chinesis CCTCC M201021 innon-aqueous phase. Journal of Molecular Catalysis B: Enzymatic, Elsevier, v. 18, n. 1, p.29–37, 2002.
XU, Z.; LIU, Q.; FINCH, J. Silanation and stability of 3-aminopropyl triethoxy silane onnanosized superparamagnetic particles: I. direct silanation. Applied surface science, Elsevier,v. 120, n. 3, p. 269–278, 1997.
YADAV, G.; DEVI, K. Immobilized lipase-catalysed esterification and transesterificationreactions in non-aqueous media for the synthesis of tetrahydrofurfuryl butyrate: comparisonand kinetic modeling. Chemical engineering science, Elsevier, v. 59, n. 2, p. 373–383, 2004.

87
YESILOGLU, Y. Utilization of bentonite as a support material for immobilization of< i>candida rugosa</i> lipase. Process Biochemistry, Elsevier, v. 40, n. 6, p. 2155–2159, 2005.
YIGITOGLU, M.; TEMOÇIN, Z. Immobilization of Candida rugosa lipase on glutaraldehyde-activated polyester fiber and its application for hydrolysis of some vegetable oils. Journal ofMolecular Catalysis B: Enzymatic, Elsevier, v. 66, n. 1, p. 130–135, 2010.
YIN, C.; LIU, T.; TAN, T. Synthesis of vitamin a esters by immobilized Candida sp. lipase inorganic media. Chinese Journal of Chemical Engineering, Elsevier, v. 14, n. 1, p. 81–86, 2006.
YONG, Y.; BAI, Y.; LI, Y.; LIN, L.; CUI, Y.; XIA, C. Characterization of Candida rugosalipase immobilized onto magnetic microspheres with hydrophilicity. Process Biochemistry,Elsevier, v. 43, n. 11, p. 1179–1185, 2008.
ZANIN, G. Sacarificaçãoo de amido em reator de leito fluidizado com enzima amiloglicosidaseimobilizada. Campinas, 1989. Tese (Doutorado) — Tese (Doutorado)-Faculdade de Engenhariade Alimentos da Universidade Estadual de Campinas, 1989.
ZHU, J.; SUN, G. Lipase immobilization on glutaraldehyde-activated nanofibrous membranesfor improved enzyme stabilities and activities. Reactive and Functional Polymers, Elsevier,2012.
![[T] Análise da estabilidade postural e propriocepção de ... · A estabilidade postural foi analisada utilizando uma plataforma de força no ... diminuir a incidência de quedas](https://img.pdfslide.us/doc/110x75/5c60477009d3f28b648b461f/t-analise-da-estabilidade-postural-e-propriocepcao-de-a-estabilidade.jpg)




























