Embed Size (px)
Citation preview

UNIVERSIDADE DO VALE DO ITAJAÍ
CENTRO DE CIÊNCIAS DA SAÚDE
RODRIGO HERMES ZANDONAI
ANÁLISE DA ATIVIDADE LINFOPROLIFERATIVA DE CÉLULAS ESPLÊNICAS MURINAS FRENTE A EXTRATO DE SEIS PLANTAS
MEDICINAIS DA FLORA CATARINENSE
Itajaí - 2007

UNIVERSIDADE DO VALE DO ITAJAÍ
CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE MESTRADO ACADÊMICO EM CIÊNCIAS
FARMACÊUTICAS
ÁREA DE CONCENTRAÇÃO EM PRODUTOS NATURAIS E
SUBSTÂNCIAS SINTÉTICAS BIOATIVAS
RODRIGO HERMES ZANDONAI
ANÁLISE DA ATIVIDADE LINFOPROLIFERATIVA DE CÉLULAS ESPLÊNICAS MURINAS FRENTE A EXTRATO DE SEIS PLANTAS
MEDICINAIS DA FLORA CATARINENSE
Dissertação submetida à Universidade do Vale do Itajaí como parte dos requisitos para a obtenção do grau de Mestre em Ciências Farmacêuticas.
Orientador: Prof. Dr. Ednéia Casagranda
Bueno
Itajaí, Outubro de 2007.

ANÁLISE DA ATIVIDADE LINFOPROLIFERATIVA DE CÉLULAS ESPLÊNICAS MURINAS FRENTE A EXTRATO DE SEIS PLANTAS
MEDICINAIS DA FLORA CATARINENSE
Rodrigo Hermes Zandonai
‘Esta Dissertação foi julgada adequada para obtenção do Título de Mestre em Ciências Farmacêuticas, Área de Concentração Produtos Naturais e Substâncias Bioativas e aprovada em sua forma final pelo Programa de Mestrado em Ciências Farmacêuticas da Universidade do Vale do Itajaí.’
____________________________________ Ednéia Casagranda Bueno, Dra. Orientador
__________________________________________________________ Tânia Mari Bellé Bresolin, Dra.
Coordenador do Programa Mestrado em Ciências Farmacêuticas Apresentado perante a Banca Examinadora composta pelos Professores:
______________________________________
Dra. Ednéia Casagranda Bueno (UNIVALI) Presidente
______________________________________
Dra. Darlene Camati Persuhn Rolin de Moura (UNIVALI) Membro
______________________________________
Dra. Elaine Maria de Souza Fagundes (UFMG) Membro
Itajaí (SC), 05, outubro, de 2007.

3
Dedico este trabalho a todos que confiaram e me deram apoio
a Deus e aos meus Pais as pessoas que ficaram ao meu lado
e aos que duvidaram pois deles veio a força de provar que sou capaz.

4
AGRADECIMENTOS
Quero agradecer primeiramente a quem mais merece, a Professora Doutora Ednéia Casagranda Bueno, pois ela me ensinou o verdadeiro significado da palavra orientação, pois não basta apenas corrigir, deve-se guiar. Sabemos todos que grandes generais não são aqueles que se apóiam nas estrelas de em seus ombros, mais sim na confiança que seus soldados têm em seguir suas ordens, sabendo que ali está sua segurança da vitória, agradeço muito a ela por não só ter me guiado na pesquisa, mais ter me ensinado mais uma profissão e me ter proporcionado a grande chance de sentir no estágio de docência o quanto é maravilhoso participar da formação do conhecimento, tornando-me um apaixonado por essa profissão. Agradeço a ela pela confiança depositada, pela força nos momentos mais difíceis dessa jornada, enfim agradeço por tudo, pois sem ela nada disso teria sido feito, muito obrigada Professora, por ser nossa luz nos momentos mais escuros e nosso guia nas horas em que tudo pareceu perdido.
Agradeço ao Doutor Rilton pelo apoio e confiança depositados na nossa equipe
ao ceder espaço para realizarmos nosso trabalho, sempre lembrando-nos da importância e do cuidado ao manipularmos nossas culturas. A ele meu muito obrigado, pois sem sua ajuda nada teria feito.
Agradeço a Doutora Tânia pela confiança, de que conseguiria cumprir meus
prazos e o apoio de coordenadora desse ilustre Curso. Agradeço ao Doutor Rivaldo Niero, ao Doutor Cechinel Filho, a Doutora Maique,
pelo apoio e por cederem os extratos para este estudo. Agradeço a Rosélia e a Vânia por sempre estarem dispostas a me ajudar e por
serem muitas vezes amigas, parceiras e sempre me receberem bem. Agradeço a minha grande amiga Marina Elisa Felippe, minha ex-chefe,
companheira de pesquisa, colega que sempre esteve ao meu lado desde o início e foi quem me incentivou a fazer o mestrado. Ao meu grande amigo Luiz Fischer, pelas risadas, pela ajuda nas aulas e tudo. Ao Mário, pelo mesmo motivo, a Tereza , a Roseni, ao Humberto, e enfim a todos meus maravilhosos colegas que juntos enfrentamos essa jornada.
Aos meus pais pelo apoio financeiro, e por terem confiado na minha capacidade. Agradeço a Deus por ter estado comigo sempre, por ter me carregado no colo,
por estar ao meu lado mesmo quando toda esperança tinha ido embora e por ter mantido minha cabeça no lugar nas horas de desespero.

5
"Nós buscamos muito por algo que vá mudar por completo nossa existência,
quando o verdadeiro milagre é estarmos aqui e agora, aceitando-nos, os altos
e baixos, as fraquezas e os momentos fortes.
O futuro acaba prejudicado pela ansiedade em encontrá-lo. Os dias, minutos e segundos passam e muitas vezes passam até as
pessoas que amamos sem que percebamos. E não esqueçamos: se tentarmos escurecer os outros, jamais seremos iluminados."
(Ludovic W. M. Wind)

ANÁLISE DA ATIVIDADE CITOTÓXICA E LINFOPROLIFERATIVA DOS EXTRATOS METANÓLICOS DE PLANTAS MEDICINAI EM
CÉLULAS ESPLÊNICAS MURINAS Rodrigo Hermes Zandonai
Outubro/2007 Orientador: Ednéia Casagranda Bueno, Dra. Área de Concentração: Produtos naturais e substâncias bioativas Número de páginas: 68 Vários compostos têm sido utilizados como agentes imunomoduladores, permitindo modificar a função do sistema imune, estimulando ou suprimindo a mesma. O estudo de atividades terapêuticas de extratos de plantas tem mostrado aumento crescente nestes últimos anos, inclusive quanto à atividade imunomodulatória. O presente trabalho objetivou identificar e avaliar a ação de extratos das plantas medicinais Calophyllum brasiliensis, Ipomoea pes-caprae, Matayba elaegnoides, Maytenus robusta, Rubus imperialis e Vernonia scorpioides sobre a proliferação de células esplênicas de camundongos empregando o ensaio de proliferação celular in vitro. O ensaio foi realizado em microplacas de 96 cavidades utilizando-se meio Eagle Modificado por Dulbecco (D’MEM) suplementado com soro bovino fetal, L-glutamina, HEPES e piruvato de sódio. As células esplênicas murinas foram obtidas por ruptura mecânica do baço (5x103 células/cavidade). Os extratos metanólicos das plantas foram testados nas concentrações de 1, 5, 10 e 20 mg/cavidade, enquanto que a fitohemaglutinina (PHA) e o pokeweed mitogen (PWM) foram utilizados a 0,5mg/cavidade. A proliferação celular foi investigada incubando a suspensão celular com o extrato da planta, isolada ou simultaneamente à PHA e PWM. O controle negativo de proliferação foi constituído de células e D´MEM, enquanto que os controles positivos foram células e PHA e PWM, isoladamente. O cultivo celular foi mantido a 37 oC com 5% de CO2 durante 72 horas e a quantificação da proliferação celular foi realizada pelo ensaio de redução do azul de tetrazolium (MTT). Os resultados, expressos em percentagem de crescimento e analisados pelos testes de Kruskal-Wallis e Mann-Whitney, identificaram diferentes efeitos dependendo da planta estudada. O extrato de Calophyllum brasiliensis mostrou indução da proliferação celular de maneira dose-dependente, com aumento significativo na proliferação quando incubado isoladamente e em conjunto com a PHA (p<0,029), potencializando o efeito da PHA e sugerindo estímulo de linfócitos T. O extrato de Ipomoea pes-caprae induziu a proliferação dose-dependente das células esplênicas, com aumento significante da mesma quando incubado isoladamente e em conjunto com o PWM (p<0,043), indicando, em conjunto com outros dados, que tanto os linfócitos T quanto os linfócitos B são ativados. O extrato de Matayba elaegnoides mostrou indução dose-dependente na proliferação celular, tanto isolado quanto em conjunto com o PWM (p<0,030) sugerindo que o extrato tem ação estimuladora sobre a ativação e proliferação de linfócitos T e B. O extrato de Maytenus robusta mostrou não interferir na proliferação celular quando incubado isolado e simultâneo ao PWM (p>0,083), mas com efeito supressor quando incubado simultaneamente com PHA, apresentando assim uma diversidade de resultados que não permitiram fazer inferências quanto à atividade do extrato. Os extratos de Rubus imperialis e de Vernonia scorpioides apresentaram inibição comprovada da proliferação celular incubado isolado e simultaneamente ao PWM (p<0,043). Desta forma, este trabalho apresenta plantas imunomoduladoras e que merecem destaque e continuidade nos estudos, seja devido ao efeito estimulador, Calophyllum brasiliensis e Ipomoea pes-caprae, bem como pelo efeito inibidor, Rubus imperialis e Vernonia scorpioides. Palavras-chave: Calophyllum brasiliensis, imunomodulação, Ipomoea pes-caprae, Matayba elaegnoides, Maytenus robusta, resposta imune, Rubus imperialis, Vernonia scorpioides.

ANALYSIS OF THE CITOTOXIC AND LYMPHOPROLIFERATIVE ACTIVITY OF METHANOLIC EXCTRACTS OF MEDICINAL PLANTS
ON MURINE SPLEEN CELLS Rodrigo Hermes Zandonai
October/2007 Supervisor: Ednéia Casagranda Bueno, Dr. Area of Concentration: Natural Products and Bioactive Substances Number of Pages: 68 Some compounds have been used as immunomodulatory agents, enabling the function of the immune system to be modified, stimulating or suppressing it. There has been increasing interest in the study of therapeutic activities of plant extracts in recent years, including their immunomodulatory activity. The aim of the present work is to identify and evaluate the action of extracts of the medicinal plants Calophyllum brasiliensis, Ipomoea pes-caprae, Matayba elaegnoides, Maytenus robust, Rubus imperialis and Vernonia scorpioides on the development of spleen cells from mice using the in vitro cellular proliferation assay. The assay was carried out in 96-well microplates using Eagle Dulbecco’s Modified Medium (D’MEM) supplemented with fetal bovine serum, L-glutamin, HEPES and sodium pyruvate. The murine spleen cells were obtained by mechanical rupture of the spleen (5x103 cells/well). The methanol extracts were used at 1, 5, 10 and 20 mg/well, while the phytohemagglutinin (PHA) and pokeweed mitogen (PWM) were both used at 0.5 mg/well. The cell proliferation was performed by incubating the cellular suspension with the plant extract, isolated or simultaneously with PHA and PWM. The negative proliferation control consisted of cells and DMEM, while the positive controls consisted of cells and PHA and PWM in isolation. The cell culture was kept at 37 oC with 5% of CO2 for 72 hours, and the cell proliferation was quantified by means of the blue tetrazolium reduction assay (MTT assay). The results were expressed as percentage growth, and were analyzed by the Kruskal-Wallis and Mann-Whitney tests. The Calophyllum brasiliensis extract showed dose-dependent induction of the cell proliferation, with a significant increase in cell proliferation when incubated in isolation and together with the PHA (p<0.029), increasing the PHA effect and suggesting stimulation of the T lymphocytes. The Ipomoea pes-caprae extract induced dose-dependent cell proliferation, with a significant increase in proliferation when incubated in isolation and together with PWM (p<0.043), indicating, with other data, that T and B lymphocytes are activated. The Matayba elaegnoides extract showed dose-dependent induction of cell proliferation with significant induction of growth, both in isolation and with PWM (p<0.030), suggesting that the extract has a stimulatory effect on T and B lymphocyte activation and proliferation. The Maytenus robust extract showed no interference on cell proliferation when incubated in isolation and simultaneously with PWM (p>0.083), but showed a suppressant response when incubated simultaneously with PHA. The diversity of the results obtained did not allow us to make any inferences concerning the activity of this extract. The Rubus imperialis and Vernonia scorpioides extracts presented inhibition of the cellular proliferation both when incubated in isolation and simultaneously with PWM (p<0.043). As a result, this work presents plants with immunomodulatory activities that deserve to be highlighted and studied in further depth, due to their stimulatory effect, e.g. Calophyllum brasiliensis and Ipomoea pes-caprae, as well as their inhibitory effect, Rubus imperialis and Vernonia scorpioides. Key words: Keywords: Calophyllum brasiliensis, immune response, immunomodulation, Ipomoea pes-caprae, Matayba elaegnoides, Maytenus robusta, Rubus imperialis, Vernonia scorpioides

8
LISTA DE TABELAS E FIGURAS
Tabela 1. Mitógenos: características e células alvo............................................. 27
Figura 1 Avaliação da proliferação celular, em percentagem de crescimento, obtida no ensaio de proliferação de células esplênicas murinas em incubação de 72 horas a 37 oC com 5% de CO2 frente ao pokeweed mitogen (5 µg/mL)................................................................................. 37
Figura 2 Avaliação da proliferação celular, em percentagem de crescimento, na ausência de estímulo mitogênico, incubação de 72 horas a 37 ºC e 5% de CO2 frente a extrato metanólico de diferentes plantas medicinais............................................................................................. 43
Figura 3 Curva dose-resposta do efeito dos extratos no ensaio de proliferação com células esplênicas murinas e extrato metanólico de diferentes plantas medicinais durante 72 horas a 37 ºC e 5% CO2...................... 44
Figura 4 Avaliação da proliferação celular, em percentagem de crescimento, frente ao extrato metanólico de diferentes plantas medicinais na presença de estímulo mitogênico com fitohemaglutinina (PHA), em incubação de 72 horas a 37 ºC e 5% de CO2....................................... 48
Figura 5 Curva dose-resposta do efeito dos extratos no ensaio de proliferação com células esplênicas murinas e extrato metanólico de diferentes plantas medicinais e fitohemaglutinina, incubados simultaneamente no cultivo celular durante 72 horas a 37 ºC e 5% CO2......................... 49
Figura 6 Avaliação da proliferação celular, em percentagem de crescimento, frente ao extrato metanólico de diferentes plantas medicinais na presença de estímulo mitogênico com pokeweed mitogen (PWM), em incubação de 72 horas a 37 ºC e 5% de CO2................................. 54
Figura 7 Curva dose-resposta do efeito dos extratos no ensaio de proliferação com células esplênicas murinas e extrato metanólico de diferentes plantas medicinais e pokeweed mitogen, incubados simultaneamente no cultivo celular durante 72 horas a 37 ºC e 5% CO2....................................................................................................... 55

SUMÁRIO
1 INTRODUÇÃO...................................................................................................... 12
2 OBJETIVOS…………………………………………………………...……….............14
2.1 Objetivo Geral...................................................................................................14
2.2 Objetivos Específicos......................................................................................14
3 REVISÃO DA LITERATURA……………..………………………………….............15
3.1 Plantas medicinais de uso popular de interesse neste estudo ................. 18
3.1.1 Calophyllum brasiliensis............................................................................ 18
3.1.2 Ipomoea pes-caprae……………………………………….............................. 19
3.1.3 Matayba elaeagnoides................................................................................. 20
3.1.4 Maytenus robusta........................................................................................ 21
3.1.5 Rubus imperialis.......................................................................................... 22
3.1.6 Vernonia scorpioides .................................................................................. 23
3.2 O Sistema Imune..............................................................................................24
4 MATERIAL E MÉTODOS……………………………………………..................... 31
4.1 Material..............................................................................................................31
4.1.1 Reagentes.......................................................................................................31
4.1.2 Equipamentos.................................................................................................31
4.2 Métodos............................................................................................................32
4.2.1 Tipo de estudo................................................................................................32
4.2.2 Local de estudo...............................................................................................32
4.2.3 População/amostra.........................................................................................32
4.2.4 Obtenção material botânico............................................................................32
4.2.5 Obtenção das células esplênicas....................................................................33
4.2.6 Proliferação celular in vitro..............................................................................34
4.3 Análise e discussão dos dados......................................................................36
4.4 Procedimentos éticos......................................................................................36
5 RESULTADOS E DISCUSSÃO............................................................................37
6 CONCLUSÕES.....................................................................................................56
7 PERSPECTIVAS..................................................................................................59
REFERÊNCIAS.......................................................................................................60

12
1 INTRODUÇÃO
O uso de plantas medicinais tem se mostrado adequado para atender as
necessidades básicas de saúde, em função da facilidade de acesso, baixo custo e
compatibilidade com as tradições populares. O estudo de atividades terapêuticas de
extratos oriundos de plantas, gerado a partir de avanços científicos envolvendo
estudos químicos, farmacológicos e toxicológicos, tem mostrado aumento crescente
nestes últimos. Isso é refletido no grande número de medicamentos oriundos de
plantas já disponíveis comercialmente (WAGNER, 2007).
Os fármacos que manipulam o sistema imune eram conhecidos até
recentemente como imunossupressores. Estes fármacos tiveram o uso e o
desenvolvimento ampliado devido aos alo e xenotransplantes e ao avanço no
conhecimento da Imunologia Clínica, que tem revelado a fisiopatologia de doenças
causadas por exacerbação da resposta imune e por imunodeficiências em diversas
áreas da medicina (LIMA, 2007). O sistema imune constitui múltiplos mecanismos de
defesa que objetivam a proteção do organismo contra ação dos agentes infecciosos
e a manutenção da homeostase interna. A pele e as superfícies mucosas funcionam
como barreira à entrada de microorganismos, constituindo uma primeira linha de
defesa. Quando estas barreiras são rompidas, as etapas seqüenciais na defesa do
organismo são ativadas, incluindo neste eficiente processo as células do sistema
imune (HENRY, 2003).
A imunomodulação é o processo de regulação da função do sistema imune,
compreendendo resposta de estímulo à resposta de defesa e auxiliando no combate
aos agentes infecciosos ou, contrariamente, suprimindo esta resposta. Vários
compostos têm sido utilizados como agentes imunomoduladores, como adjuvantes
naturais, agentes sintéticos e anticorpos reagentes, mas existem limitações no uso
desses compostos devido ao risco de infecções e efeito generalizado através do
sistema imunológico (MAKARE; BODHANKAR; RANGARI, 2001).
A análise da atividade imunomodulatória de compostos naturais tem sido
determinada por diferentes metodologias e em diferentes modelos, tanto in vivo
quanto in vitro. O ensaio de proliferação de linfócitos é um dos modelos utilizados
para avaliar atividade imunomodulatória de compostos naturais (PANDIMA DEVI et
al., 2003). No ensaio são utilizados mitógenos como controle de positivo de

13
proliferação celular, como a fitohemaglutinina (PHA), que estimula linfócitos T, e o
pokeweed mitogen (PWM), que estimula linfócitos B dependentes de linfócitos T
(ROCHA; GORESCU; BELTRAME, 2007). Desta forma, o emprego destes
mitógenos permite avaliar comparativamente o efeito causado pelos extratos de
plantas na linfoproliferação, determinando a presença de indução ou inibição da
mesma e identificando o tipo celular da resposta imune adquirida envolvido no
processo.
O estudo aqui relatado apresenta dados da atividade biológica relacionada à
resposta imune adquirida, por meio do ensaio de proliferação de células esplênicas
de camundongos in vitro frente aos extratos metanólicos de seis plantas medicinais
já estudo quanto à caracterização fitoquímica e de atividade biológica em projetos
desenvolvidos pelos pesquisadores do Núcleo de Investigações Químico-
Farmacêuticas (NIQFAR). As plantas selecionadas encontradas em solo
Catarinense têm uso medicinal popular e apresentaram, anteriormente, resultados
promissores sob ponto de vista químico medicinal: a Calophyllum brasiliensis é
utilizada contra bronquites, distúrbios hepáticos e gástricos, dor, inflamação,
diabetes, varicoses, diarréia, herpes, reumatismo, hemorróidas e úlceras crônicas
(SILVA et al.,2001); a Ipomea pes-caprae é utilizada no tratamento de dermatite
causada por água-viva, cólicas, desordem diuréticas, gonorréia, inflamação e
processos dolorosos (PONGPRAYOON et al., 1989; SOUZA et al., 1998); a Matayba
eleagnoides é utilizada popularmente contra inflamações, dor e no combate ao
câncer de fígado (LORENZI, 2000; RIBEIRO, 2004), assim como a Maytenus
robusta (ANDRADE et al., 2007); a Rubus imperialis apresenta uso popular no
tratamento de úlcera péptica, diabetes e enfermidades relacionadas à dor
(CECHINEL FILHO, 2000); por fim, a Vernonia escorpioides apresenta uso popular
no tratamento de afecções cutâneas (MONTEIRO et al., 2001).

14
2 OBJETIVOS
2.1 Objetivo Geral
Estudar o efeito in vitro de seis extratos metanólicos obtidos a partir de
plantas medicinais de uso popular sobre a proliferação de células esplênicas de
camundongos, in vitro, com o objetivo de identificar extratos com potencial
imunomodulador.
2.2 Objetivos Específicos
· Avaliar os efeitos dos extratos metanólicos de Calophyllum brasiliensis, Matayba
elaegnoides e Maytenus robusta na proliferação de células esplênicas, in vitro,
na presença de PHA;
· Verificar o efeito dos extratos metanólicos de Calophyllum brasiliensis, Ipomoea
pes-caprae, Matayba elaegnoides, Maytenus robusta, Rubus imperialis e
Vernonia scorpioides na proliferação de células esplênicas, in vitro, na presença
de PWM;
· Avaliar o efeito imunomodulador dos extratos metanólicos de Calophyllum
brasiliensis, Ipomoea pes-caprae, Matayba elaegnoides, Maytenus robusta,
Rubus imperialis e Vernonia scorpioides em células esplênicas na ausência de
estímulo exógeno;
· Investigar o impacto dos extratos na proliferação celular.

15
3 REVISÃO DA LITERATURA
O uso de plantas para tratar enfermidades e doenças do ser humano é mais
antigo do que a história escrita, o que tem sido comprovado pelas descobertas de
restos de plantas medicinais utilizadas em sítios arqueológicos habitados por
civilizações pré-históricas. O estudo das drogas vegetais, seguramente uma das
ciências mais antigas, provavelmente surgiu como uma necessidade do homem em
separar as plantas comestíveis das tóxicas. Posteriormente, este estudo empírico
passou ao objetivo de proteção e cura contra enfermidades, formando um arsenal de
informações conhecido como conhecimento popular e que se transmitiu entre as
gerações (ALVES, 2001).
Através da história, diversas civilizações procuraram organizar esses
conhecimentos de forma útil e documentada. A obra escrita com descrição
adequada de plantas medicinais é conhecida como Herbário, e o mais antigo de que
se tem relato é o el Pen´t, escrito no tempo do imperador Shen Nung em 2700 a.C.,
contendo 365 plantas. Em 1700 a.C. foi escrito o famoso papiro de Ebers, no Egito
Antigo, com a descrição de vários compostos de plantas medicinais. Outras
descrições se encontram na literatura védica oriunda da Índia, entre 4000 a 1000
a.C com descrições da planta e suas ações, e ainda os babilônios e assírios, com
registros similares em 600 a.C. (ALVES, 2001).
As plantas representaram, durante séculos, a única fonte de agentes
terapêuticos para o homem. O desenvolvimento da Química Farmacêutica no início
do século XIX também estabeleceu o uso de plantas como fonte de escolha para a
obtenção de princípios ativos e para o desenvolvimento de medicamentos. Estas
plantas, cujas propriedades terapêuticas são amplamente utilizadas na medicina
popular contra diversas patologias, receberam a denominação de plantas
medicinais. Vários são os fatores que proporcionaram o crescimento gradativo do
uso de plantas medicinais ao longo dos últimos anos, como baixo custo e baixo
poder aquisitivo da população (WAGNER, 2007).
Até meados do século XX, a terapêutica medicamentosa tinha como base o
uso de produtos obtidos de espécies vegetais. A síntese de medicamentos teve
início no final do século XIX, grandemente impulsionada a partir da década de trinta,

16
com o desenvolvimento de drogas muito eficazes que acabaram por substituir o uso
de fitoterápicos em diversos ramos da medicina. Entretanto, na década de 70,
diversas empresas farmacêuticas passaram a desenvolver pesquisas na área da
fitoterapia. Com isso, já na década de 90, metade das 250 grandes empresas do
ramo farmacêutico mundial introduziu programas de pesquisa na área de produtos
naturais (CARVALHO, 2001).
O reino vegetal, maior reservatório de moléculas farmacologicamente ativas a
serem descobertas, tem sido alvo de investimentos de grandes companhias
farmacêuticas que visam à pesquisa de novas substâncias ativas em extratos de
plantas. Na Terra existem aproximadamente 350.000 espécies de plantas, mas
apenas uma pequena percentagem foi investigada fitoquímicamente, com um
número ainda menor analisado farmacologicamente. Considerando ainda que uma
planta possa conter milhares de diferentes metabólitos secundários, os estudos
fitoquímicos possibilitam apenas uma pequena fração de conhecimento acerca desta
complexidade (HOSTETTMANN; QUEIROZ; VIEIRA, 2003).
Atualmente, apesar do grande desenvolvimento da síntese orgânica e de
novos processos biotecnológicos, 25% dos medicamentos prescritos nos países
industrializados são originários de plantas. A terapia moderna utiliza
aproximadamente 120 compostos de origem natural e que são obtidos a partir de
cerca de 90 espécies de plantas (HOSTETTMANN; QUEIROZ; VIEIRA, 2003). Além
disso, 10 dentre os 20 medicamentos mais vendidos mundialmente no ano de 1998
também foram desenvolvidos a partir de produtos naturais. Ainda é necessário
considerar que muitas moléculas desenvolvidas a partir de produtos naturais estão
atualmente em fase de estudo clínico (CALIXTO, 2001).
As plantas respondem a estímulos ambientais bastante variáveis, de
natureza, física, química ou biológica. Fatores como fertilidade, tipo de solo,
umidade, radiação solar, vento, temperatura e poluição atmosférica, dentre outros,
podem influenciar e alterar a composição química dos vegetais. Além desses fatores
há interações e adaptações co-evolutivas complexas, que se produzem entre
diferentes plantas, plantas e animais, plantas e microrganismos e também entre
plantas e insetos de um dado ecossistema. Um exemplo clássico desta interação é a
que acontece entre planta e inseto, que depende da presença, tempo certo e
quantidade apropriada de determinados compostos voláteis para a formação de
aromas característicos. Essas substâncias voláteis se difundem com facilidade a

17
partir da evaporação e constituem uma importante comunicação entre fonte
produtora e o meio ambiente, implicando diretamente na atração ou repulsão destes
insetos (MARGIS et al., 1999)
Outros fatores intrínsecos referem-se ao metabolismo da planta. O
metabolismo primário é responsável pela produção de celulose, lignina, proteínas,
lipídeos, açúcares e outras substâncias que realizam as principais funções vitais,
enquanto que do metabolismo secundário resultam substâncias de baixo peso
moleculares, às vezes produzidas em pequenas quantidades. Os metabólitos
secundários produzidos pelas plantas são formados por vários caminhos
biossíntéticos e produzem moléculas dotadas de grande diversidade de esqueletos e
grupamentos funcionais, como ácidos graxos e seus ésteres, hidrocarbonetos,
álcoois, aldeídos e cetonas, compostos acetilênicos, alcalóides, terpenóides,
derivados de fenilpropanóides, compostos fenólicos e cumarinas. Estas substâncias
funcionam como agentes defensivos na luta contra predadores, a exemplo de
microrganismos patogênicos, insetos e animais herbívoros (ALVES, 2001).
Esse conhecimento etnofarmacológico culminou com desenvolvimento
substâncias com grande importância na terapêutica atual, tais como ácido salicílico,
a atropina, a pilocarpina, o quinino, a artemisina, o taxol, a digoxina e a morfina. Nos
anos 80, o desenvolvimento da pesquisa científica resultou na identificação de 120
compostos de origem vegetal provenientes de 95 espécies de plantas. No período
de 1983 a 1994, 6% dos medicamentos aprovados pelo Food and Drug
Administration (FDA) foram extraídos diretamente de espécies vegetais, outros 24%
são produtos derivados destas e 9% foram desenvolvidos através de modelagem
molecular, onde as estruturas moleculares dos compostos serviram como
precursores de processos de sínteses químicas (VEROTTA, et. al., 2000).
Somado à relevância do estudo de plantas com objetivo medicinal, é
indispensável considerar a existência de uma imensa biodiversidade no Brasil, muito
pouco conhecida e investigada quanto aos estudos químicos e farmacológicos. O
Brasil é o país com a maior diversidade genética tanto na fauna quanto na flora,
contando com mais de 55.000 espécies catalogadas em sua flora, de um total
estimado entre 350.000 e 550.000 espécies diferentes. Esta diversidade genética,
acrescida da diversidade química oferecida pelo grande número de espécies,
permite a utilização de plantas como fonte inestimável de compostos bioativos úteis
no desenvolvimento de novos fármacos. O desenvolvimento do conhecimento dessa

18
flora, bem como de novas tecnologias para obtenção de fitoterápicos com maior
eficácia, requer o estudo prévio relativo aos aspectos botânicos, agronômicos,
fitoquímicos, farmacológicos, de atividade biológica, toxicológica e finalmente o
desenvolvimento de tecnologia analítica e metodológica (OTTO; KAPLAN;
BORIN,1996).
A escolha das plantas a serem avaliadas em um estudo deve ser criteriosa,
observando o uso e forma de administração na medicina popular, bem como os
possíveis efeitos tóxicos observados. Aliado a isto, é necessário o conhecimento e
experiência da população e da cultural local, bem como informações sobre
composição química da planta em estudo (CHECINEL; YUNES, 1998). Neste
contexto, algumas espécies de uso popular tem sido objeto de estudos de vários
grupos de pesquisadores.
3.1 Plantas medicinais de uso popular de interesse neste estudo
3.1.1 Calophyllum brasiliensis
O gênero Calophyllum (da família Clusiaceae e Gutiiferae) é composto de 180
a 200 diferentes espécies confinadas no ambiente tropical úmido. A Calophyllum
brasiliensis está muito bem distribuída na América Latina, desde o Brasil até o
México, apresentando diferentes nomes populares, como Jacareuba, Guanandi e
Bari. No Brasil, a Calophyllum brasiliensis cresce principalmente em regiões de Mata
Atlântica (SILVA et al., 2001). A espécie apresenta folhas glabras e coriáciceas,
perenifólia, característica das florestas pluviais localizadas sobre os solos úmidos e
brejosos, vindo a florescer nos meses de setembro a novembro (LORENZI, 1998).
Esta planta medicinal é utilizada popularmente em bronquites, distúrbios
hepáticos e gástricos, dor, inflamação, diabetes, hipertensão, diarréia, herpes,
reumatismo, varicose, hemorróidas e úlceras crônicas (SILVA et al., 2001). O infuso
e banho preparados com as cascas da arvore é o uso mais popular empregado no
tratamento destas patologias (SARTORI et al., 1999).
Uma extensa investigação do gênero Calophyllum resultou no isolamento de
uma grande variedade de compostos, incluindo xantonas, cumarinas, biflavonoídes,
chalconas, benzofenonas e triterpenos (ISAIAS, et al., 2004). Os extratos e óleos de
várias espécies desse gênero mostraram comprovada atividade antibacteriana
(DHARMARATNE, WANIGASEKERA, 1996). Os flavonóides e xantonas isolados de

19
Calophyllum paciflorum demonstraram atividade preventiva anticancer (ITO et al.,
1999). Alguns compostos isolados da Calophyllum inophyllum, oriundos da casca,
foram testados em modelos antitumorais in vitro, destacando derivados 4-fenil-
cumarinas, aos quais são atribuídos atividades inibitórias para enzima transcriptase
reversa do HIV-1 (ITOGAWA et al., 2001).
As frações hexano e acetato de etila, bem como compostos isolados a partir
desta, quercitina, ácido gálico, ácido protocatético, hiperosideo (quercetin-3-O-
galactosideo) e amantoflavona, obtidos a partir da folha da Calophyllum brasiliensis,
apresentaram efeito analgésico similar aos fármacos de referência (SILVA et al.,
2001). Estudos fitoquímicos do caule e da resina desta espécie revelaram a
presença de diversas substâncias químicas, e algumas cumarinas isoladas
apresentaram atividade antimicrobiana (GASPAROTTO et al., 2005). Um estudo
recente revelou que o extrato hexano das folhas desta espécie não tem ação
relevante no desenvolvimento do tumor ascítico de Ehrlich, porém estimula a
produção de células pela medula óssea, mostrando ser um estimulador da
hematopoiese (BREHMER, 2005).
3.1.2 Ipomoea pes-caprae
As plantas pertencentes ao gênero Ipomoea, da família Convolvulaceae,
consistem em mais de 200 espécies amplamente distribuídas nos países tropicais e
subtropicais, sendo frequente na maioria das praias dos Oceanos Índico, Atlântico e
Pacífico (SOUZA et al., 1998). O gênero é oriundo das dunas costeiras, encontrado
com hábitos herbáceos em grande quantidade margeando o litoral brasileiro, desde
o litoral do Estado do Pará até o Estado do Rio Grande do Sul (SOUZA et al., 2000).
Uma pesquisa da flora litorânea específica de dunas menciona que algumas
plantas formam raízes adventícias em caules rastejantes e que são conhecidas
como formação pes-caprae. O representante mais importante é conhecido
popularmente como salsa da praia, a Ipomoea pes-caprae, formação essa que
permite retirar nutrientes em solo pobre e arenoso, em manter a mobilidade da
planta de acordo com a mudança da estrutura física do solo em que a mesma se
encontra (MARTINES; MORENO-CASASOLA, 1996).
A Ipomoea pes-caprae é uma espécie perene de vida longa, apresenta
sistema de estolões longos de 10 a 15 metros e ramos curtos de 25 a 30
centímetros, tanto estolões e ramos curtos podem apresentar inflorescência. Em

20
Santa Catarina, a floração e frutificação da Ipomoea pes-caprae ocorre no verão e
no outono (SANTOS; ARRUDA, 1995). Esta espécie tem sido utilizada em muitos
países como planta medicinal em casos de inflamação, cólica, desordens diuréticas,
gonorréia, processos dolorosos, entre outros (PONGPRAYOON et al., 1989; SOUZA
et al., 1998).
O extrato obtido de todas as partes desta espécie inibiu a ação proteolítica e
hemolítica da água-viva, fundamentando o uso popular da planta no tratamento
desta dermatite (PONGPRAYOON; BOHLIN; WASUWAT, 1991). Os estudos
realizados com esta espécie apresentam efeito analgésico em modelos
experimentais de dor, tanto na fase neurogênica quanto na fase inflamatória. A
presença de esteróides, terpenóides, alcalóides e flavonóides, entre outros
compostos identificados na análise fitoquímica, parecem justificar esses efeitos,
provavelmente pela inibição na síntese de prostaglandinas (SOUZA et al., 2000).
A inibição das contrações da musculatura lisa do intestino, em modelos
experimentais, atividade antiespasmódica de potência semelhante à papaverina e
propiciada por componentes isoprenóides isolados do extrato bruto da planta, é
outra comprovação de outro uso popular da planta (KROGH et al., 1999). O extrato
bruto desta espécie mostrou ainda aumento no desenvolvimento do tumor ascítico
de Ehrlich em camundongos, embora tenha apresentado estimulo na proliferação de
leucócitos na medula óssea dos animais (BREHMER, 2005).
3.1.3 Matayba elaeagnoides
A Matayba elaeagnoides é encontrada desde o Estado de Minas Gerais e São
Paulo até o Rio Grande do Sul, em matas de altitude do tipo semidecídua e mata
dos Pinhais. A planta, popularmente conhecida como camboatá ou cuvatã, é uma
árvore mediana 6 a14 metros de altura, com tronco tortuoso de 30 a 50 cm de
diâmetro, folhas compostas do tipo pinada de 10 a 20 cm e folíolos em número de 4
a 13, casca áspera cor cinza-claro ou mais escuro e com leve escamação. As flores
surgem em forma de inflorescência menores que as folhas e de cor branca,
florescendo durante os meses de setembro e novembro, enquanto que os frutos
amadurecem entre os meses de dezembro e janeiro. A planta produz sementes
viáveis anualmente com ciclo de reprodução ligados às aves. Ainda, a madeira desta
espécie tem sido amplamente empregada na construção civil (LORENZI, 2000).

21
Embora a espécie seja popularmente utilizada no tratamento de inflamações,
analgesia e combate ao câncer de fígado, são escassos os estudos científicos que
sugiram a comprovação destas atividades farmacológicas usadas na medicina
popular (LORENZI, 2000; RIBEIRO, 2004). Estudos realizados com extratos
oriundos das cascas de Matayba elaeagnoides demonstraram a presença de vários
compostos químicos, como amarinas, lupeol, sitosterol, escopoletina, umbeliferona e
betulina. Todos os extratos e frações avaliados pelo autor apresentaram atividade
antinociceptiva e antimicrobiana, somado ainda à atividade antifúngica da fração
clorofórmio e à ação antiparisitária da fração hexano. Ainda de acordo com este
estudo, a betulina mostrou-se 7 vezes mais potente do que o paracetamol e a
aspirina em testes de dor induzidos pelo ácido acético e pela formalina (SOUZA,
2006).
3.1.4 Maytenus robusta
A Maytenus robusta é popularmente conhecida como cafezinho, coração-
de-bugre e seca-ligeiro. A espécie é semidecídua, heliófita, seletiva hidrófita,
secundaria característica, e exclusiva da vegetação das restingas litorâneas e das
matas semidecídua de altitude possuindo, por tanto, de grande amplitude ecológica.
A planta produz sementes viáveis anualmente, em quantidade abundante e
amplamente disseminada pela avifauna. A florescência ocorre durante os meses de
setembro a novembro, e os frutos amadurecem a partir do mês de maio. A semente
de M. robusta é liberada do fruto após a abertura deste, que é do tipo cápsula
deiscente bivalvar (LORENZI, 1998).
Estudos fitoquímicos demonstraram que Maytenus robusta apresenta os
mesmos componentes da Maytenus ilicifolia, como terpenoides, flavonóides e
principalmente a classe de triterpenos fridelin, responsável pela atividade anti-
ulcerogênica. Considerando estes dois fatores, muitos fitoterápicos apresentam essa
composição em ambas as espécies (NIERO et al., 2001), e demonstram também a
importância dos estudos fitoquímicos e de atividade biológica com esta espécie
alternativa.
Conhecida pelos índios há muitos anos, a Maytenus ilicifolia, ou espinheira
santa, ganhou esse nome justamente pela aparência das folhas, que contém
espinhos nas margens. A maior ocorrência desta planta é em regiões de clima
ameno, de Minas Gerais ao Rio Grande de Sul, sendo mais abundante nas matas do

22
Sul. O clima preferido é o subtropical e temperado, com solos argilosos, bem
drenados e com alto teor de matéria orgânica (ROSE, 1997).
Na medicina popular a espinheira santa é famosa no combate a úlcera,
dado este já confirmado em modelo experimental de lesão estomacal em
camundongos (ANDRADE et al., 2007). A composição química da Maytenus ilicifolia
revelou a presença de triterpenos e compostos fenólicos com importantes
propriedades biológicas, como atividade citotóxica, antibaceriana e anti-ulcerogênica
(OLIVEIRA et al., 1991; CARLINI, 1988; SOUZA-FORMIGONI et al., 1991; ZHU,
1998).
Considerando que a população nativa de Maytenus ilicifolia quase foi extinta
devido à crescente exploração para uso medicinal e que a Maytenus robusta tem
sido referenciada como fonte alternativa para substituição da mesma devido à
grande incidência na Região Sul e à presença dos mesmos compostos em abas as
espécies (NIERO et al., 2001), no presente estudo foi utilizada a espécie Maytenus
robusta na identificação da presença de efeito imunomodulador.
3.1.5 Rubus imperialis
A família das Rosáceas é representada por um variável e complexo grupo de
plantas que crescem em todas as partes do mundo, especialmente em regiões
temperadas, compreendendo cerca de 100 gêneros e mais de 700 espécies.
Algumas espécies do gênero Rubus são cultivadas há décadas e conhecidas pela
variedade de frutos requintados, utilizados na fabricação de vários tipos de doces e
geléias (PATEL; ROJAS-VERA; DACKE, 2004).
A planta possui ramos longos, angulosos e com acúleos que se estendem de
4 a 5 metros, sustentados em outros vegetais. As folhas são compostas, palmadas e
dentadas, enquanto que as flores apresentam pétalas brancas, florescendo de
outubro a novembro e com frutos compostos por inúmeras drupas (SILVA; SANTOS;
CECHINEL FILHO, 1999). A Rubus imperialis, conhecida como amora-branca,
amora-do-mato ou amora-brava, bem como outras espécies, tem sido empregada
em comunidades rurais para diabetes e outras enfermidades, algumas delas
relacionadas à dor (CECHINEL FILHO, 2000).
A Rubus imperialis apresenta perspectivas terapêuticas desde que Niga-
ichigoside F1, um composto triterpeno isolado com acetato de etila, exibiu atividade
analgésica em ratos (LOCHER et al., 1996). Outros autores demonstraram que a

23
atividade analgésica da Rubus imperialis foi aproximadamente trinta vezes mais
potente no teste de contorções abdominais induzidas pelo ácido acético em
camundongos que duas drogas de referência, a aspirina e o paracetamol. No
mesmo estudo, o teste da formalina mostrou que esta espécie previne tanto a dor de
origem neurogênica quanto a de origem inflamatória (NIERO et al., 2002). Os relatos
da literatura incluem ainda estudos químicos e farmacológicos que confirmaram que
algumas espécies de Rubus sp possuem efeito hipoglicemiante, atividades anti-
alérgicas e antibacterianas (SWANSTON-FLATT et al., 1990).
A fração acetato de etila das raízes da planta apresentou atividade no
bioensaio de citotoxicidade frente Artemia salina, enquanto que o Niga-ichigosídeo
F1 desta fração não teve ação sobre o microcrustáceo, indicando que devem existir
outros compostos responsáveis pela atividade citotóxica ou ainda ocorrer um
sinergismo entre mais de um princípio ativo (KANEGUSUKU, 2001). Estudo
realizado sobre o mecanismo de ação desta substância mostrou resultados
importantes e sugerem que estão envolvidas tanto no sistema adrenérgico quanto
no serotoninérgico (ARDENGHI et al., 2006). A análise dos constituintes químicos
encontrados na fração acetato de etila de partes aéreas de Rubus imperialis mostrou
presença de triterpenos niga-ichigosídeo, ácido tormêntico e ácido 23-hidroxi-
tormentico (NIERO et al., 2002).
3.1.6 Vernonia scorpioides
Existem cerca de 200 espécies de Vernonia no Brasil, sendo muito comum
em todo Brasil e crescendo em solos pobres e desflorestados. A V. scorpioides é
conhecida popularmente como piracá, enxuga ou erva-de-São-Simão (CABRERA;
KLEIN, 1980). A planta apresenta característica de arbusto com ramos numerosos e
lanuginosos, com folhas pecoladas, ovaladas, agudas no ápice e atenuadas na
base, inteira ou creanada. As flores são do tipo corola purpúreas reunidas em
caítulo numerosos, síniles, unilaterais, dispostas em panículos alongados
escorpióides, com fruto aquênico piloso turbinado com papo brancacento (CORREA,
1978; REITZ, 1980).
Algumas das espécies do gênero Vernonia são tradicionalmente usadas para
tratar desordens gastrointestinais, como as folhas frescas maceradas de V.
condensata conhecida como fel-de-bugre, fel-de-índio ou alumã. O uso tópico do

24
extrato hidroalcoólico das folhas frescas é amplamente empregado pela população
para tratar uma variedade de afecções cutâneas, incluindo úlceras crônicas
(MONTEIRO et al., 2001). A literatura apresenta estudos focalizando atividades
antiinflamatórias (MAZUMDER et al., 2003; IWALEWA; IWALEWA; ADEBOYE,
2003), antipirética (GUPTA et al., 2003), anticâncer (IZEVBIGIE; BRYANT;
WALKER, 2004; LAMBERTINI et al., 2004; HOWARD et al., 2003) e antimalárica
(ABOSI; RASEROKA, 2003) de várias espécies de Vernonia.
O gênero Vernonia produz lactonas sesquiterpênicas, metabólitos típicos da
família Asteraceae com várias atividades biológicas descritas, como fungistáticas
(KRISHNA KUMARI et al., 2003), relaxantes do músculo liso (CAMPOS et al., 2003)
e citotóxicas (KUO; KUO, YU, 2003). Outras substâncias isoladas de Vernonia são
flavonóides (HUANG; DING; LIU, 2003), esteróides (TCHINDA et al., 2003) e
polissacarídeos (NERGARD; DIALLO ; MICHAELSEN, 2004).
A Vernonia scorpioides teve comprovada atividade fungicida (FREIRE et al.,
1996) e leve atividade cicatrizante (LEITE et al., 2002). Ainda, a fração
diclorometano desta espécie administrada via intra-venosa apresentou efeito
citotóxico sobre o tumor ascítico e sólido de Ehrlich em camundongos, devido
principalmente às lactonas sesquiterpenica e outras lactonas bioativas, enquanto
que o tratamento por via oral ou intraperitoneal não interferiu no volume do tumor
sólido, sugerindo ações enzimáticas que desativem a via sistêmica (BREHMER,
2005). Outra propriedade que pode exacerbar esta ação citotóxica é o aumento do
influxo de neutrófilos para a cavidade peritoneal a capacidade promovida pela
fração, ao contrário do convencional perfil de imunossupressão da terapia
convencional (PAGNO et al., 2006). Recentemente foi relatada a atividade citotóxica
das lactonas sesquiterpênicas e do poliacetileno obtido desta espécie sobre células
tumorais Hela, com indução de apoptose e aumento na expressão de caspase 3 por
este último composto (BUSKÜHL, 2007).
3.2 O Sistema Imune
O sistema imunológico tem por função proteger o organismo contra o
constante ataque de agentes infecciosos, como vírus, bactérias, fungos e

25
protozoários. O desenvolvimento fisiológico do sistema imune tem início ainda na
vida intra-uterina com a hematopoiese no saco vitelino, seguido do fígado fetal como
tecido hematopoiético e, finalmente, com a medula óssea assumindo a produção
das células. A divisão celular em precursores linfóides e mielóides constituem o
primeiro passo na diferenciação das células do sistema imune. Os precursores
linfóides originam os linfócitos T e B e as células natural killers, enquanto os
precursores mielóides originam os eritrócitos, plaquetas, monócitos, macrófagos,
células dendríticas, neutrófilos, basófilos e eosinófilos. Os linfócitos T e B são as
células relacionadas à resposta imune adquirida (específica), enquanto que as
demais populações celulares representam a resposta imune inata (inespecífica)
(WEISSMAN, 1994).
Os organismos extracelulares, como bactérias e protozoários, são combatidos
pela resposta imune inespecífica via ativação de células apresentadoras de
antígeno, que fagocitam os organismos extracelulares e os digerem pela ação de
mieloperoxidase, lizosimas, defensinas, lactoferrina e a produção dos reativos
intermediários de oxigênio e nitrogênio. Os microorganismos, por sua vez, ativam a
via alternativa do sistema complemento, que é capaz de estimular a fagocitose,
quimiotaxia das células imunes, produção e liberação de mediadores farmacológicos
dos mastócitos e lise direta do organismo invasor. Na imunidade específica, os
organismos extracelulares se defrontam com anticorpos produzidos pelos linfócitos
B, e esta ligação ativa a via clássica do sistema complemento, neutralizando o
organismo ou facilitando a fagocitose (HOWARD; GARRA; ISHIDA, 1992).
Os microorganismos intracelulares, como bactérias ou vírus, são destruídos
por inúmeros mecanismos. Na imunidade inespecífica, a infecção viral de uma célula
induz à produção de interferon, tornando as células vizinhas resistentes a infecção.
As células natural killer reconhecem uma célula infectada com vírus e liberam
perforinas, granzinas e linfotoxinas, destruindo a célula infectada. A imunidade
mediada por células neste caso é realizada pelos linfócitos T CD8, os linfócitos T
citotóxicos, cujos receptores reconhecem especificamente as proteínas virais
produzidas na célula infectada e em associação com moléculas de MHC de classe I
do hospedeiro. Este reconhecimento das proteínas vírais através do MHC de classe
I desencadeia a destruição da célula infectada (HOWARD; GARRA; ISHIDA, 1992).
Os microrganismos intracelulares, quando no interior de células apresentadoras de
antígeno como os macrófagos, também podem ser destruídos por linfócitos T CD4,

26
os linfócitos T auxiliares, cujos receptores reconhecem antígenos estranhos
apresentados pelas moléculas MHC de classe II das células de defesa do
hospedeiro. O reconhecimento estas proteínas estranhas através do MHC de classe
II induz a liberação de citocinas, incluindo o interferon, que ativam os macrófagos e
funções antimicrobianas (WEISSMAN, 1994).
A ativação dos linfócitos decore da interação entre um antígeno e o respectivo
receptor presente na superfície celular, o receptor de célula T (TCR). Nos linfócitos T
CD4, por exemplo, a proteína de membrana responsável por esta ligação da célula
ao antígeno é o receptor de célula T (TcR), e esta interação, culmina com a
proliferação, diferenciação e produção de citocinas que medeiam às funções
específicas de células T efetoras. Eventos subseqüentes que ocorrem em segundos
ou minutos após a interação entre ligante e receptor incluem mudanças no fluxo de
sódio e potássio, ativação de metiltransferases e síntese e utilização de fosfolipídios.
O influxo de cálcio é ponto central na ativação de linfócitos e ocorre em menos de
cinco minutos após a interação do antígeno com os receptores presentes na
superfície dos linfócitos. Após algumas horas dessa interação ocorre aumento na
síntese de várias proteínas (COOPER, 1972). O conteúdo de RNA mensageiro em
linfócitos dobra em cerca de 48 horas e reflete as alterações morfológicas distintas
da célula. Por fim, o aumento da síntese de DNA, ação inerente na proliferação
celular, tem inicio por volta de 36 horas após o contato, alcançando níveis elevados
entre 48 e 72 horas quando ocorre a mitose (GREAVES; JANOSSY, 1972).
Na replicação dos linfócitos T também deve ser considerada a importante
função da IL-2. A interação em níveis significativos de IL-2 com o respectivo receptor
(IL-2-R) presente na membrana das células T desencadeia a replicação do DNA. A
intensa expressão deste receptor na membrana celular ocorre somente após receber
estímulo por antígenos. Entretanto, anticorpos monoclonais específicos para células
T, lectinas mitogênicas ou forbol éster também exacerbam a expressão deste
receptor (SMITH; CANTRELL, 1985). Assim, estas substâncias possuem a
habilidade de ativar e, conseqüentemente, induzir a proliferação in vitro de linfócitos
T ou linfócitos B, ou ainda ambos, sendo genericamente denominados mitógenos
(GREAVES; JANOSSY, 1972). A Tabela 1 mostra uma seleção dessas substâncias
com a respectiva especificidade com referência ao tipo de célula que os mesmos
estimulam.

27
Tabela 1. Mitógenos: características e células alvo.
Fonte: Adptado de Myers (1995) e Rocha, Gorescu e Beltrame (2007).
Os mitógenos PHA, PWM e concanavalina A (ConA) são lectinas extraídas e
purificadas de plantas que estimulam sub populações de linfócitos T e/ou B,
apresentam ampla capacidade de utilização em estudos de proliferação dessas
populações in vitro e a resposta resultante pode ser utilizada para qualificar e
quantificar a imunocompetência dessas células (MYERS, 1995).
A ativação de um linfócito, in vivo, reside na interação entre o agente estranho e o
receptor de superfície das células T, iniciando eventos que envolvem o recrutamento
de cálcio e a ativação simultânea das enzimas tirosina quinases citossólicas
fosfolipases, proteína C quinase, proteína quinase ativada por mitógeno e
calcineurina. Estas enzimas, a partir da via de cascatas metabólicas específicas,
produzem sinais direcionados ao núcleo da célula, promovendo a transcrição de
fatores pré-mitóticos e conduzindo os eventos da proliferação e da síntese de IL-2,
sendo a mesma o principal mediador químico da amplificação da resposta
imunológica (RAO; CHANRABARTI, 2005).
As deficiências do sistema imunológico podem ser causadas por defeitos
enzimáticos, agentes infecciosos, drogas, entre outros. Em geral, os sintomas e
achados físicos da imunodeficiência estão relacionados com grau de deficiência e o
sistema particular cuja função está deficiente. No tratamento destas
imunodeficiências, além dos agentes antimicrobianos utilizados no tratamento das
Mitógeno Fonte Tamanho
molecular (kDa) Células Alvo Espécie(s)
Concanavalina A (Con A)
Canavalia einsformis 55.000 Linfócitos T
Humana e camundongo
Fitohemaglutinina (PHA)
Phaseolus vulgaris
140.000 Linfócitos T Humana e
camundongo
Lipopolissacarídeo (LPS)
Bactérias gram Negativas
1-24x106 Linfócitos B
Roedores
Pokeweed mitógeno (PWM)
Phytolacca americana 32.000
Linfócitos T e B
Humana e camundongo
Dextran sulfato 500.000 Linfócitos B

28
infecções oportunistas, também há novas formas de imunoterapia que visam auxiliar
ou controlar a deficiência através do estímulo da resposta imune. Esta abordagem
prevê o desenvolvimento de substâncias capazes de ativar as células do sistema
imune diretamente e sem dependência de células apresentadoras de antígeno
(STITES; TERR; PARSOLW, 2000).
Em contrapartida, a capacidade técnica de efetuar transplantes bem
sucedidos de muitos órgãos criou, em particular, um forte ímpeto pelo
desenvolvimento de esquemas imunossupressores eficazes e que permitam evitar a
rejeição ao enxerto. Além disso, mais de 40 doenças consideradas como
decorrentes de respostas imunológicas aberrantes são passíveis de tratamento com
agentes capazes de inibir a resposta imunológica. Estes progressos efetuados pela
Imunologia Clínica têm estimulado a pesquisa de fármacos capazes de inibir a
resposta imune (STITES; TERR; PARSOLW, 2000).
A modulação da resposta imune na eliminação e controle de doenças tem
sido área de grande interesse e de estudo. A imunomodulação é um mecanismo
fisiológico de regulação da resposta imunológica via mecanismos supressores e
estimuladores, que suprimem ou potencializam uma resposta imunológica. A
exacerbação das reações imunes é definida como imunoestimulação e implica
diretamente na estimulação do sistema imune e potencialização da resposta de
defesa. Ao contrário, a imunossupressão implica principalmente no decréscimo da
atividade do sistema imune, ocorrendo devido a uma gama de fatores genéticos,
ambientais e terapêuticos, favorecendo o estabelecimento de infecções
(PATWARDHAN et al., 1990; MAKARE; BODHANKAR; RANGARI, 2001).
Considerando a necessidade de controle e equilíbrio entre as duas atividades
para o funcionamento imunológico normal, desde a década de 80 a identificação e
caracterização de compostos naturais com atividade imunomodulatória tem se
apresentado como área de interesse científico (PHILLIPSON, 2003). Vários
compostos têm sido utilizados como agentes imunomodulatórios, incluindo
adjuvantes naturais, agentes sintéticos e anticorpos reagentes. Contudo, grande
parte destes compostos apresenta limitações quanto ao uso devido ao risco de
infecções e desenvolvimento de auto-imunidade (MAKARE; BODHANKAR;
RANGARI, 2001).
A análise da atividade imunomodulatória de compostos naturais tem sido
determinada por diferentes metodologias e em diferentes modelos, tanto in vivo

29
quanto in vitro. A linfoblastogênese, ou ensaio de proliferação de linfócios, é um dos
modelos in vitro empregados para avaliar o efeito destes compostos sobre a
proliferação celular. Este sistema permite medir a função normal de células B e T em
resposta a uma estimulação mitogênica, é um dos modelos empregados para avaliar
o efeito destes compostos sobre a proliferação celular. Neste modelo, o estudo do
efeito de compostos naturais pode ser avaliado in vitro na presença ou ausência de
estimuladores como PHA, PWM, lipopolissacáride, ConA, ou ainda antígenos
específicos obtidos de diferentes agentes agressores. Também são empregadas
outras populações celulares neste ensaio, como linfócitos murinos e humanos e
células mononucleares humanas (PANDIMA DEVI et al., 2003).
A identificação e caracterização de compostos naturais com atividade
imunomodulatória abrem uma possibilidade de uso para medicina moderna. Estudos
realizados quanto ao efeito imunomodulatório do extrato da Premma tomentosa,
planta da região sul da Índia popularmente utilizada como medicinal, mostraram que
o extrato da planta adicionado na cultura de linfócitos esplênicos promove
diminuição na citotoxicidade e da apoptose em comparação à indução tóxica do
cromo (Cr VI), apresentando característica de citoproteção (PANDIMA DEVI et al.,
2003).
Outro exemplo é a Acorus calamus, amplamente utilizada pela medicina
tradicional indiana no tratamento de insônia, melancolia, neuroses, epilepsia,
histeria, perda de memória e febre recorrente, onde a comprovação da atividade do
extrato etanólico do rizoma compreende a ação antiproliferativa e imunossupressora
em células humanas e de camundongos e células tumorais, estimuladas com
mitógenos PHA e LPS em cultura celular enriquecida com soro bovino fetal
(MEHROTA, 2003).
Em modelo de estudo semelhante, outros autores descreveram a propriedade
imunoestimulante dos polissacarídeos purificados de sementes de Phellodendri
cortex sobre os linfócitos B, espécie esta de uso popular na medicina oriental
chinesa e coreana (PARK et al.,1999). Ainda, a partir de atividade popularmente
conhecida na medicina tradicional indiana, o extrato alcoólico da casca da Mangifera
indica Linn, popularmente conhecida no Brasil como mangueira, mostrou atividade
imunomodulatória com estimulação dose dependente em linfócitos esplênicos de
camundongos (MAKARE; BODHANKAR; RANGARI, 2001).

30
O trabalho aqui apresentado mostra um estudo pioneiro acerca da atividade
biológica relacionada à resposta imune adquirida após estimulação in vitro com
extratos metanólicos de seis plantas medicinais presentes na flora do Estado de
Santa Catarina. A escolha das plantas Calophyllum brasiliensis, Ipomoea pes-
caprae, Matayba elaegnoidesMaytenus robusta, Rubus imperialis e Vernonia
scorpioides foi realizada de acordo com a disponibilidade dos extratos, tendo como
fator determinante os estudos a cerca da fitoquímica e atividade biológica realizadas
anteriormente, cujos resultados foram descritos na descrição de cada planta.

31
4 MATERIAL E MÉTODOS
4.1 Material
4.1.1 Reagentes
· Ácido clorídrico - HCl (VETEC Química Fina LTDA, Duque de Caxias, RJ);
· Azul de tetrazolium - MTT - Brometo de 3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazolio
(Amreco, catálago no 0793, Solon, Ohio, USA);
· Azul de tripan - C34H24Na4O16S4 (VETEC Química Fina LTDA, Duque de Caxias,
RJ);
· Bicarbonato de sódio P.A. - NaHCO3 (Dinâmica, São Paulo, SP);
· Dimetilsulfoxido P.A. - DMSO - (CH3)2SO (VETEC Química Fina LTDA, Duque de
Caxias, RJ);
· Dodecil sulfato de sódio - SDS, 99% de pureza (Amreco, catálago no 0227/00G,
Solon, Ohio, USA);
· Fitohemaglutinina - PHA (Sigma, catálogo no L9132, St. Louis, MO, USA);
· Hepes - Isento de Ácido (Sigma Aldrich, catálogo no H4034, St. Louis, MO, USA);
· L-glutamina (Sigma Aldrich, catálogo nº AB185, St. Louis, MO, USA);
· Meio de cultura Dulbelccos’s Modified Eagles Médium - DMEM (Sigma Inc.,
catálogo no D7777, St. Louis, MO, USA);
· Mesilato de pefloxacino (Peflacin, Aventis Pharma LTDA, São Paulo, SP);
· Piruvato de sódio - C3H3NaO (VETEC Quimica fina LTDA, Duque de Caxias, RJ);
· Pokeweed mitogen - PWM, lectina obtida de Phytolacca Americana (Sigma Aldrich,
catálogo nº L 9379, St. Louis, MO, USA);
· Soro bovino fetal estéril (Cultilab, catálago no 000072, Campinas, SP);
· Sulfato de Gentamicina (Gentamil, Green Pharma LTDA, Anápolis, GO).
4.1.2 Equipamentos
· Fluxo laminar (Veco, modelo VLFS-12, Campinas, SP);
· Centrifuga (Fanem, modelo, 206-R Exelsa Baby II, São Paulo, SP);
· Estufa de CO2 (Jouan, modelo IG150, Saint Herbain, França);

32
· Leitor de microlacas (Quick ELISA, Drake, São Paulo, SP).
4.2 Métodos
4.2.1 Tipo de estudo
Análise experimental laboratorial de amostras biológicas.
4.2.2 Local de estudo
Laboratórios 310 e 311 do Curso de Farmácia e Laboratório de Cultivo Celular
do Centro de Ciências da Saúde, Laboratório de Cultivo Vegetal do Centro de
Ciências Tecnológicas da Terra e do Mar.
4.2.3 População/amostra
Foram utilizados 60 camundongos Swiss machos com peso entre 20 e 30
gramas, obtidos junto ao Biotério da UNIVALI.
4.2.4 Obtenção do material botânico
Os extratos metanólicos foram fornecidos pelos colaboradores do presente
projeto, Prof. Dr. Valdir Cechinel Filho, Prof. Dr. Rivaldo Niero e Profa. Dra. Maique
Weber Biavatti. As raízes de Calophyllum brasiliensis foram coletadas nos jardins da
UFSC, em Florianópolis, em abril de 2001. O material foi identificado pelo Dr Ademir
Reis (Departamento de Botânica da UFSC) e uma exsicata foi depositada no
Herbário Barbosa Rodrigues em Itajaí-SC sob número VC Filho 007. A planta inteira
de Ipomoea pes-caprae foi coletada na Praia da Esplanada, em Jaguaruna-SC e
uma exsicata foi depositada no mesmo Herbário sob número V.C. Filho 012. As
cascas de Matayba elaegnoides foram coletadas no município de Caçador - SC no
mês de maio 2004 identificado pelo Dr Ademir Reis (Departamento de Botânica da
UFSC) e uma exsicata foi depositada no mesmo Herbário sob número MTS 001. As
partes aéreas de Maytenus robusta foram coletadas em outubro de 1997 no Parque
Ecológico do Morro do Baú, município de Ilhota, Santa Catarina, identificada pelo Dr
Ademir Reis (Departamento de Botânica da UFSC) e uma exsicata foi depositada no
mesmo Herbário sob número V.C. Filho 016. As partes aéreas de Rubus imperialis
foram coletadas em São Sebastião, Treze de Maio-SC e uma exsicata foi depositada

33
no mesmo Herbário sob número V.C. Filho 012. As folhas e inflorescências de
Vernonia scorpioides foram coletadas em Navegantes (SC) em local previamente
determinado, e identificadas pela Dra. Ana Claudia Araújo, sendo comparada a
exsicatas tombadas no Herbário Barbosa Rodrigues sob número HBR, M. Biavatti
11.
O extrato alcoólico das plantas citadas acima foi preparado a partir do
material vegetal seco e triturado, utilizando o metanol como líquido extrator mediante
maceração durante 10 dias em recipiente fechado à temperatura ambiente. Os
extratos metanólicos das plantas utilizados nos experimentos de proliferação celular
foram obtidos pela filtração e concentração do material por evaporação à pressão
negativa. Métodos espectroscópicos como infravermelho, ressonância magnética
nuclear de próton e carbono 13 e técnicas bidimensionais foram utilizadas em
estudos prévios na elucidação estrutural em estudos prévios de caracterização
fitoquímica de compostos isolados destes extratos (KROGH et al., 1999; NIERO et
al., 1999; NIERO et al., 2001; SILVA et al., 2000; SOUZA, 2006; BUSKÜHL, 2007).
Os extratos metanólicos foram conservados em dessecador com sílica para
retirada de umidade e solubilizados no momento do uso em 2% de DMSO
constituindo junto com meio de cultura os extratos concentrados (1 mg/mL). O
extrato concentrado foi submetido a processo de filtração em filtro com poro de 22 m
para garantia da esterilidade do mesmo, evitando assim a contaminação do cultivo
celular.
4.2.5 Obtenção das células esplênicas
Os animais foram sacrificados por deslocamento cervical, seguido da
retirada cirúrgica do baço. As células esplênicas foram obtidas assepticamente por
rompimento mecânico da cápsula do baço em solução salina tamponada estéril.
Após centrifugação 10 minutos a 1000 r.p.m. a suspensão celular foi submetida a
choque hipotônico. As hemácias presentes no sedimento foram lisadas com água
destilada estéril durante 30 segundos, seguidos da reconstituição da isotonicidade
para 0,9%. Após nova centrifugação, a concentração celular foi determinada pela
contagem em câmara de Neubauer, simultaneamente à verificação da viabilidade
celular pela exclusão com azul de Tripan (BOYUM, 1968), sendo utilizadas somente
suspensões celulares com viabilidade superior a 90%. Para a obtenção do número

34
suficiente de células para a realização dos procedimentos foram utilizadas células
obtidas de baço de diferentes animais.
4.2.6 Proliferação celular
O procedimento foi realizado em câmara de fluxo laminar, evitando assim a
contaminação com bactérias e fungos e a interferência nos resultados. A avaliação
da proliferação celular, previamente padronizado em nossa Instituição (KRUEGER et
al. 2005; PETTRES et al., 2006), foi adequado às condições dos laboratórios onde a
pesquisa foi desenvolvida (COELHO et al., 2007). Foi empregado no cultivo celular o
meio de cultura DMEM suplementado com 10% de soro bovino fetal, 2% de
bicarbonato de sódio a 10%, 1% de L-glutamina a 200 mM, 1% de HEPES a 10mM e
110 mg/mL de piruvato de sódio.
O ensaio foi realizado em microplacas de 96 poços, de fundo chato, estéreis e
com tampa (TTP - Techno Plastic Products, Trasadingen, Suíça). As células
esplênicas foram empregadas na concentração de 50.000 células/mL (5.000
células/cavidade) e o mitógeno PHA foi utilizado na concentração de 5 mg/mL (0,5
mg/cavidade) (COELHO et al., 2007). O ensaio de diferentes concentrações de
células (2.000 a 250.000 células/mL, respectivamente 200 a 25.000
células/cavidade) frente ao mitógeno PWM na concentração de 5 mg/mL (0,5
mg/cavidade) definiu, por meio da maior percentagem de crescimento encontrada, a
concentração celular ideal para a proliferação celular com o PWM como controle
positivo de crescimento. Os extratos metanólicos das plantas foram ensaiados nas
concentrações de 10 mg/mL, 50 mg/mL, 100 mg/mL e 200 mg/mL (respectivamente 1,
5, 10 e 20 mg/cavidade).
Somente foram utilizados os resultados dos experimentos cujos controles
internos mostraram não interferir no ensaio de proliferação celular, a saber: a)
controle do meio de cultura, com plaqueamento de meio de cultura apenas; b)
controle dos mitógenos, com incubação de PHA e PWM, separadamente, com meio
de cultura; c) controle do extrato, com incubação do extrato com meio de cultura; d)
controle das células esplênicas, com incubação de células e meio de cultura.
O ensaio de proliferação celular com os extratos das plantas foi realizado na
presença e na ausência dos mitógenos: a) ensaio de proliferação celular na
ausência dos mitógenos, com incubação de células e extrato com meio de cultura; b)

35
ensaio de proliferação celular na presença dos mitógenos, com incubação de células
e extrato com PHA ou PWM, isoladamente, em meio de cultura. Para avaliação do
ensaio de proliferação foram empregados os seguintes controles internos: a)
controle negativo de proliferação, com incubação de células e meio de cultura; b)
controle positivo de proliferação para PHA, com incubação de células e PHA com
meio de cultura; c) controle positivo de proliferação para PWM, com incubação de
células e PWM com meio de cultura.
Todos os ensaios, realizados em triplicata, foram incubados a 37 oC em
atmosfera com 5% de CO2 durante 72 horas, período este determinado de acordo
com a literatura para estudo em modelo murino (YASNI et al., 1993; ROSEGHINI et
al., 2006). A quantificação da proliferação celular foi realizada pelo ensaio de
redução MTT, que forma cristais de coloração azulada quando convertido em sal de
formazan pela atividade da desidrogenase mitocondrial das células vivas
(MOSMANN, 1983). Para este procedimento foram adicionados a cada poço da
microplaca, três horas antes do término do período de incubação, 10 mL de MTT a 5
mg/mL em NaCl 0,9%. Após o término do período de incubação foram adicionados a
cada poço 100 mL de SDS a 10% em HCl 0,001 N e deixados over night para a
dissolução dos cristais formados pela redução do MTT pelas células. A densidade
óptica de cada poço foi determinada em espectrofotômetro de microplacas a 540 nm
(DENIZOT; LANG, 1986).
Os resultados do ensaio de proliferação celular foram apresentados em
percentagem de crescimento, que elimina a proliferação residual de células ativadas
previamente in vivo, bem como as variações inter-testes. Este indicador foi obtido
pela seguinte fórmula:
% crescimento = 100 x [(DOt - DOc) / DOc]
onde DOt corresponde à densidade óptica do ensaio de proliferação e DOc refere-se
à densidade óptica do controle negativo de proliferação. A percentagem de
crescimento obtida no ensaio será proporcional à viabilidade celular e ao número de
células viáveis. Assim, um resultado de percentagem de crescimento positivo indica
proliferação celular induzida pelo composto adicionado ao cultivo celular
(MANOSROI; SARAPHANCHOTIWITTHAYA; MANOSROI, 2003; RISCO et al.,
2003). Todos os extratos avaliados mostraram, quando incubados com meio de

36
cultura apenas, densidade óptica semelhante àquela obtida com no controle
negativo, ou seja, sem background.
4.3 Análise e discussão dos dados
Para representação das observações encontradas foram utilizados gráficos,
enquanto que os dados encontrados foram avaliados estatisticamente pelos testes
de Kruskal-Wallis e Mann-Whitnhey. A correlação entre a concentração do extrato e
efeito sobre a proliferação celular foi avaliado pelo coeficiente de determinação,
obtido a partir da linha de tendência polinomial na ordem 3, com valor R2=1,0. A
determinação da concentração que promove 50% de crescimento celular foi
estimada pela estatística de Probitos (MicrocalTM OriginTM, Microcal Software Inc.,
Northampton, MA, EUA).
4.4 Procedimentos éticos
Este estudo foi realizado de acordo com as normas éticas para pesquisa e
conta com parecer favorável da Comissão de Ética em Pesquisa da UNIVALI
(parecer número 131/2006).
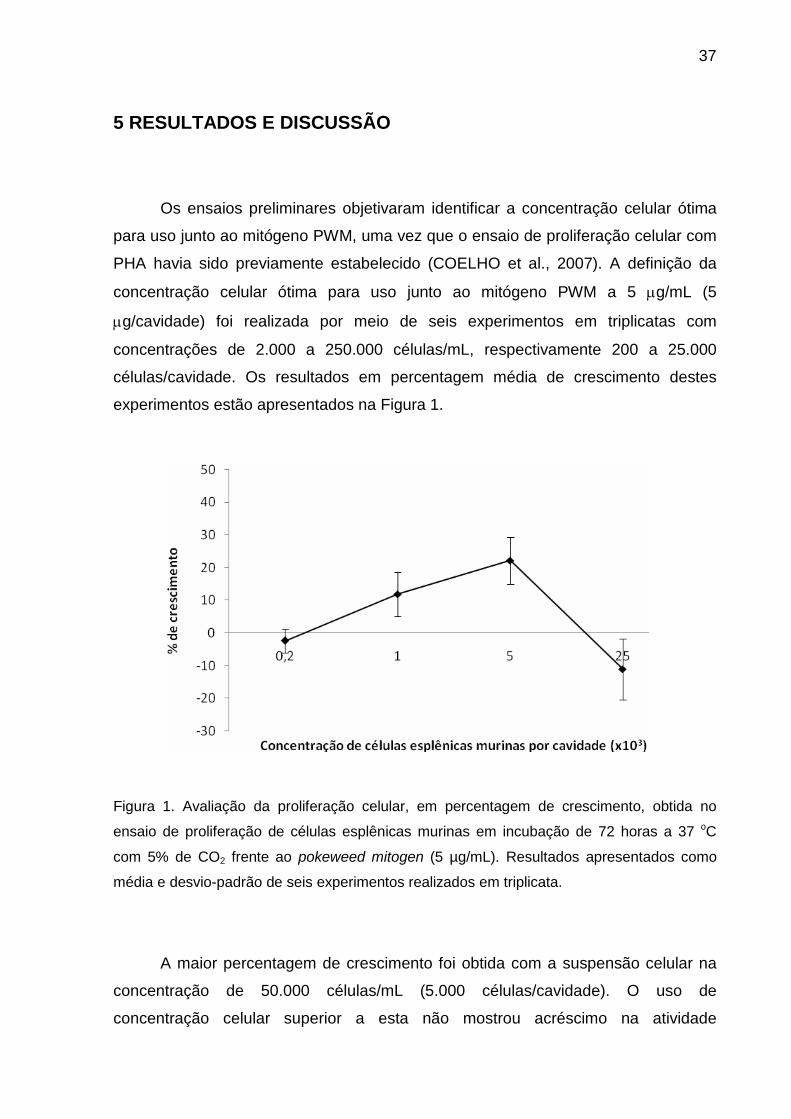
37
5 RESULTADOS E DISCUSSÃO
Os ensaios preliminares objetivaram identificar a concentração celular ótima
para uso junto ao mitógeno PWM, uma vez que o ensaio de proliferação celular com
PHA havia sido previamente estabelecido (COELHO et al., 2007). A definição da
concentração celular ótima para uso junto ao mitógeno PWM a 5 mg/mL (5
mg/cavidade) foi realizada por meio de seis experimentos em triplicatas com
concentrações de 2.000 a 250.000 células/mL, respectivamente 200 a 25.000
células/cavidade. Os resultados em percentagem média de crescimento destes
experimentos estão apresentados na Figura 1.
Figura 1. Avaliação da proliferação celular, em percentagem de crescimento, obtida no
ensaio de proliferação de células esplênicas murinas em incubação de 72 horas a 37 oC
com 5% de CO2 frente ao pokeweed mitogen (5 µg/mL). Resultados apresentados como
média e desvio-padrão de seis experimentos realizados em triplicata.
A maior percentagem de crescimento foi obtida com a suspensão celular na
concentração de 50.000 células/mL (5.000 células/cavidade). O uso de
concentração celular superior a esta não mostrou acréscimo na atividade

38
mitocondrial de células vivas, sugerindo maior consumo de nutrientes e que
provavelmente inviabilizaram o crescimento celular. Desta forma, todos os demais
experimentos foram realizados com esta concentração celular.
A Figura 2 apresenta os resultados obtidos com os extratos metanólicos de
Calophyllum brasiliensis, Ipomoea pes-caprae, Matayba elaegnoides, Maytenus
robusta, Rubus imperialis e Vernonia scorpioides incubados isoladamente no ensaio
de proliferação celular, com controles positivos PHA e PWM. Os resultados foram
apresentados como média e desvio padrão de quatro experimentos realizados em
triplicata.
Estudos com o objetivo de descrever novos agentes terapêuticos têm
revelado diferentes grupos de metabólitos secundários de natureza química variada,
como alcalóides, flavonóides, saponinas, taninos e lectinas como agentes
protagonistas de relevantes atividades biológicas (BALUNAS; KINGHOM, 2005;
BUSS; WAIGH, 1995). Baseado nesse conceito, o extrato metanólico de
Calophyllum brasiliensis, que possui em sua composição compostos como
flavonoídes, amentoflavonas, ácido gálico e outros, mostrou indução da proliferação
celular de maneira dose-dependente quando incubado isoladamente nas
concentrações de 10, 50, 100 e 200 µg/mL (R2=1,0) (Figura 2A e Figura 3A). A
percentagem média de crescimento do extrato variou de 5,8 a 88,2%, enquanto que
as dos controles positivos foram de 12,2% para PHA e de 14% para PWM. As
concentrações mais elevadas do extrato, de 100 e 200 µg/mL, mostraram indução
significativamente maior na proliferação celular em comparação com os controles
positivos PHA (p=0,021) e PWM (p<0,029). A concentração de 23,2 µg/mL foi
estimada como necessária para a indução de 50% do crescimento celular.
Um estudo anterior com extrato hexânico das folhas de Calophyllum
brasiliensis demonstrou atividade sobre a produção de células da medula óssea de
camundongos tratados por gavagem com o extrato, estimulando a hematopoiese
(BREHMER, 2005). O estímulo promovido pelo extrato metanólico de Calophyllum
brasiliensis na proliferação celular também pode estar relacionado a presença do
ácido gálico, que tem atividade antioxidante protetora nas células contra ação dos
reativos de oxigênios produzidos durante a fagocitose ou quando estimulados por
diferentes agentes (FORMAN; TORRES, 2002).
Os resultados obtidos com o extrato metanólico de Ipomoea pes-caprae, nas
concentrações de 10, 50, 100 e 200 µg/mL, isoladamente no cultivo celular estão

39
apresentados na Figura 2B. O extrato avaliado induziu a proliferação das células
esplênicas de maneira dose-dependente (R2=1,0) (Figura 3B), em concordância com
estudo realizado anteriormente (COELHO et al., 2007). A percentagem média de
crescimento observada com o extrato variou de 13,4 a 73,1%, com maior
crescimento na concentração de 200 µg/mL. Considerando as percentagens médias
de crescimento obtido nos controles positivos com PHA (12,2%) e com PWM
(10,1%), apenas o extrato a 200 µg/mL mostrou indução significativa da proliferação
celular (p=0,021). A indução de 50% do crescimento celular foi estimada com a
concentração de 40,7 µg/mL.
Um estudo realizado recentemente demonstrou que o extrato bruto de
Ipomoea pes-caprae, induziu aumento significativo no número total de células da
medula óssea dos camundongos a 200 mg/Kg, propondo que a planta tem efeito
estimulador sobre a hematopoiese (BREHMER, 2005). A presença de flavonóides
nesta espécie, como a quercitina, podem estar relacionados, com a indução no
aumento da proliferação celular devido à diminuição da peroxidação de lipídios em
nível de membrana celular. Os flavonóides também são conhecidos por atuarem
contra espécies reativas de oxigênio que tem como alvo biológico as proteínas
celulares. A oxidação destas proteínas por estas espécies reativas conduz à perda
de função, degradação prematura pelos proteossomas, alteração das propriedades
físicas da membrana celular e do DNA, tendo como conseqüência a morte celular
(CAI; RANHN; ZANG, 2004). Ainda, a presença de alcalóides em extrato de Uncaria
tomentosa (SHENG; BRYNGELSSON; PERO, 2000) apresentou potente efeito
imunoestimulante, alcalóides estes também encontrados na composição do extrato
de Ipomea pes-caprae.
Semelhantemente aos extratos metanólicos de Calophyllum brasiliensis e
Ipomoea pes-caprae, o extrato metanólico de Matayba elaegnoides também mostrou
indução dose-dependente da proliferação de células esplênicas nas concentrações
de 10, 50, 100 e 200 µg/mL (R2=1,0) (Figura 2C e Figura 3C). O extrato mostrou
percentagem média de crescimento de 3,4 a 52,7% sendo o maior crescimento
observado na concentração de 200 µg/mL. A concentração de 33,6 µg/mL foi
estimada como necessária para a indução de 50% do crescimento celular. O
crescimento celular com 10 µg/mL de extrato foi significativamente inferior ao
controle positivo PHA (12,5%, p=0,021), enquanto que a 200 µg/mL o crescimento

40
celular foi superior ao controle com PHA e com PWM (7,5%) (p<0,021). Estes dados
sugerem que ocorre ativação celular em concentrações maiores do extrato.
A variedade de compostos encontrados no extrato de Matayba elaegnoides,
como o lupeol, pode ter ação imunossupressora e também de inibição de migração
de células inflamatórias, reduzindo os fatores quimiotáticos pró-inflamatórios (PATO
KA, 2003). Este composto mostrou ainda efetividade na redução da produção de
prostaglandinas em tecidos inflamados (REYES et al., 2006). A betiluna, outro
composto isolado desta planta, além de indutor mitogênico, possui a habilidade de
induzir e modular a produção de citocinas em culturas de células humanas, com
produção de fator de necrose tumoral (ZDZSI et al., 2003). Entretanto, em
concentrações menores do extrato os compostos como o lupeol podem apresentar
maior atividade e, consequentemente, reduzir a característica estimulatória de outros
compostos.
Os resultados obtidos com o extrato metanólico de Maytenus robusta nas
concentrações de 10, 50, 100 e 200 µg/mL mostraram efeito dose-dependente
(R2=1,0) (Figura 3D) e com percentagem média de crescimento de -5,8 a 7,0%,
sendo o menor e o maior valor a 100 e 200 µg/mL, respectivamente (Figura 2D).Os
controles positivos de crescimento celular foram de 12,5% para PHA e de 7,5% para
PWM, sem diferença significativa com as quatro concentrações do extrato, sejam
elas com valores positivos ou negativos de percentagem de crescimento (p>0,075
para PHA e p>0,149 para PWM).
A presença de diferentes compostos no extrato pode ter promovido o estímulo
da proliferação celular e a supressão da mesma. Um dos compostos, a quercitina, é
um flavonóide que possui a capacidade de induzir aumento do crescimento celular
pela diminuição da peroxidação de lipídeos ao nível de membrana celular (LUNA et
al., 1996). Outros compostos, como os triterpenos, possuem atividade citotóxica em
linhagens de células tumorais sensíveis e resistentes a múltiplas drogas, atividade
esta promovida pela perda do potencial de membrana mitocondrial similar àquele
encontrada na apoptose celular (ROCHA et al., 2007).
Os resultados do crescimento celular apresentados no presente estudo foram
obtidos a partir dos valores do cultivo de células e extrato com meio de cultura frente
aos valores de células mantidas em cultivo apenas com meio de cultura, expresso
em percentagem (MANOSROI; SARAPHANCHOTIWITTHAYA; MANOSROI, 2003;
RISCO et al., 2003). Considerando esta premissa, um resultado negativo é inibidor

41
da proliferação celular, atividade esta que pode ser evidenciada pela supressão da
atividade proliferativa do mitógeno quando incubado simultaneamente ao extrato e
que, molecularmente, pode ser confirmada na imunocitoquímica pela ausência de
marcadores sinalizadores da ativação celular e proliferação, como CD69 (MAINO;
SUNI; RUITENBERG, 1995; CRASTON et al., 1997) e KI67 (STARBORG et al.,
1996; SCHOLZEN; GERDES, 2000; KAUSCH et al., 2003).
Outra hipótese a ser considerada é a lise celular promovida pelo composto
adicionado à cultura, ou seja, o efeito citotóxico do extrato sobre as células e cuja
avaliação emprega metodologias específicas para a identificação de necrose e
apoptose, como dosagem de LDH no sobrenadante do cultivo celular (CRUZ-
CHAMORRO et al., 2006) e de detecção de caspase 3 por imunocitoquímica
(KRAJEWSKA et al., 1997) ou pela liberação de p-nitroalinina (POSMANTUR;
WANG; GILBERTSEN, 1998).
O extrato metanólico de Rubus imperialis inibiu a proliferação celular nas
quatro concentrações testadas, sendo significativo nas concentrações de 10, 100 e
200 mg/mL frente aos controles positivos com PHA (15,9%) e com PWM (13,8%)
(p<0,030) (Figura 2E), com correlação com a dose (R2=1,0) (Figura 3E). A
percentagem média de crescimento observada com esta planta foi de -11,8% a -
4,4%, sendo a primeira correspondente à concentração de 10 mg/mL. A
concentração de extrato necessária para a indução de 50% do crescimento celular
não pôde ser determinada devido ao efeito supressor do mesmo. Existe uma grande
semelhança quanto às características fitoquímicas entre as várias espécies do
gênero Rubus, principalmente no que se refere à presença de triterpenos (TANAKA
et al., 1993). O mesmo extrato testado nesse estudo já apresentou atividade
citotóxica comprovada frente Artemia salina (KANEGUSUKU, 2001).
O extrato metanólico de Vernonia scorpioides apresentou percentagem média
de crescimento de -4,1 a 6,7%, estas correspondendo à concentração de 10 e 200
mg/mL, respectivamente (Figura 2F). A atividade, embora extremamente discreta, foi
dose-dependente (R2= 1,0) (Figura 3F) e com resultado significantemente inferior ao
controle positivo (15,9%, p=0,043) para o extrato na concentração de 100 mg/mL. A
concentração de 11,3 µg/mL foi estimada como necessária para a indução de 50%
do crescimento celular. Embora não seja possível avaliar a atividade citotóxica do

42
extrato no presente estudo, a literatura tem relatado esta atividade em diferentes
modelos de estudo (BREHMER, 2005; PAGNO et al., 2006; BUSKÜHL, 2007).
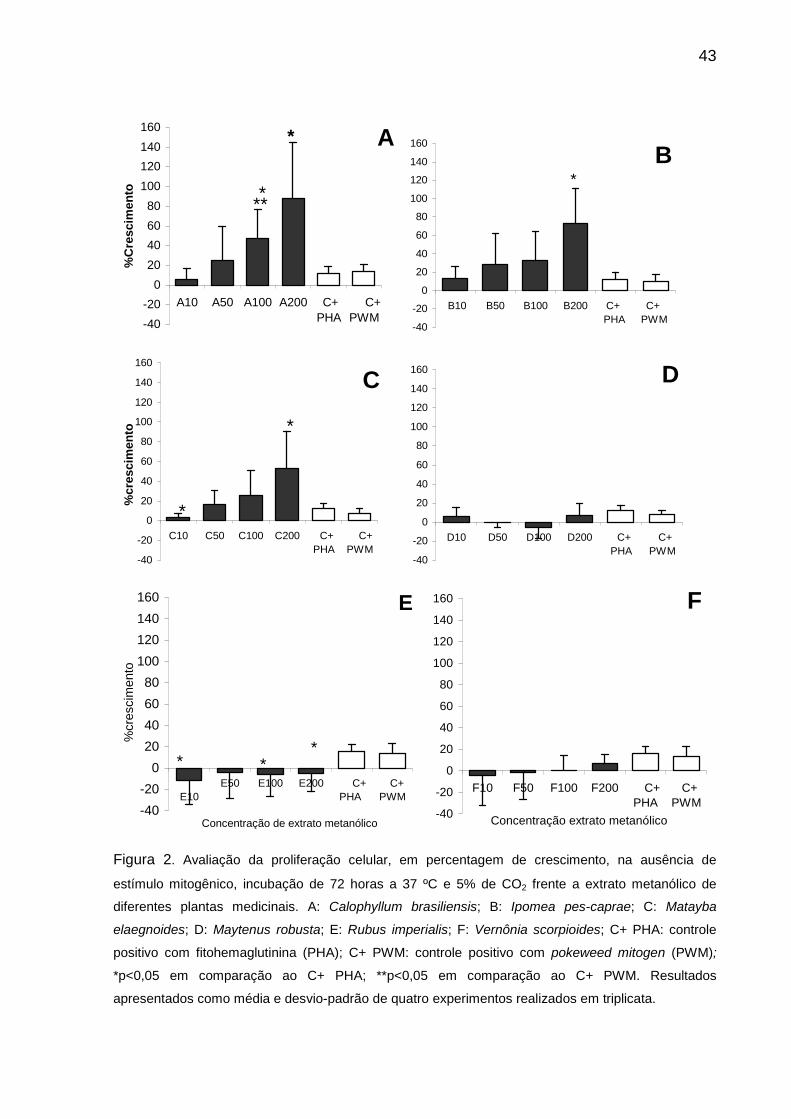
43
A
-40
-20
0
20
40
60
80
100
120
140
160
A10 A50 A100 A200 C+ PHA
C+PWM
%C
resc
imen
to
***
*B
-40
-20
0
20
40
60
80
100
120
140
160
B10 B50 B100 B200 C+ PHA
C+ PWM
*
C
-40
-20
0
20
40
60
80
100
120
140
160
C10 C50 C100 C200 C+ PHA
C+ PWM
%cr
esci
men
to *
*
D
-40
-20
0
20
40
60
80
100
120
140
160
D10 D50 D100 D200 C+ PHA
C+ PWM
E
-40
-20
0
20
40
60
80
100
120
140
160
E10
E50 E100 E200 C+ PHA
C+ PWM
Concentração de extrato metanólico
%cr
esci
men
to
* **
F
-40
-20
0
20
40
60
80
100
120
140
160
F10 F50 F100 F200 C+ PHA
C+ PWM
Concentração extrato metanólico
Figura 2. Avaliação da proliferação celular, em percentagem de crescimento, na ausência de
estímulo mitogênico, incubação de 72 horas a 37 ºC e 5% de CO2 frente a extrato metanólico de
diferentes plantas medicinais. A: Calophyllum brasiliensis; B: Ipomea pes-caprae; C: Matayba
elaegnoides; D: Maytenus robusta; E: Rubus imperialis; F: Vernônia scorpioides; C+ PHA: controle
positivo com fitohemaglutinina (PHA); C+ PWM: controle positivo com pokeweed mitogen (PWM);
*p<0,05 em comparação ao C+ PHA; **p<0,05 em comparação ao C+ PWM. Resultados
apresentados como média e desvio-padrão de quatro experimentos realizados em triplicata.
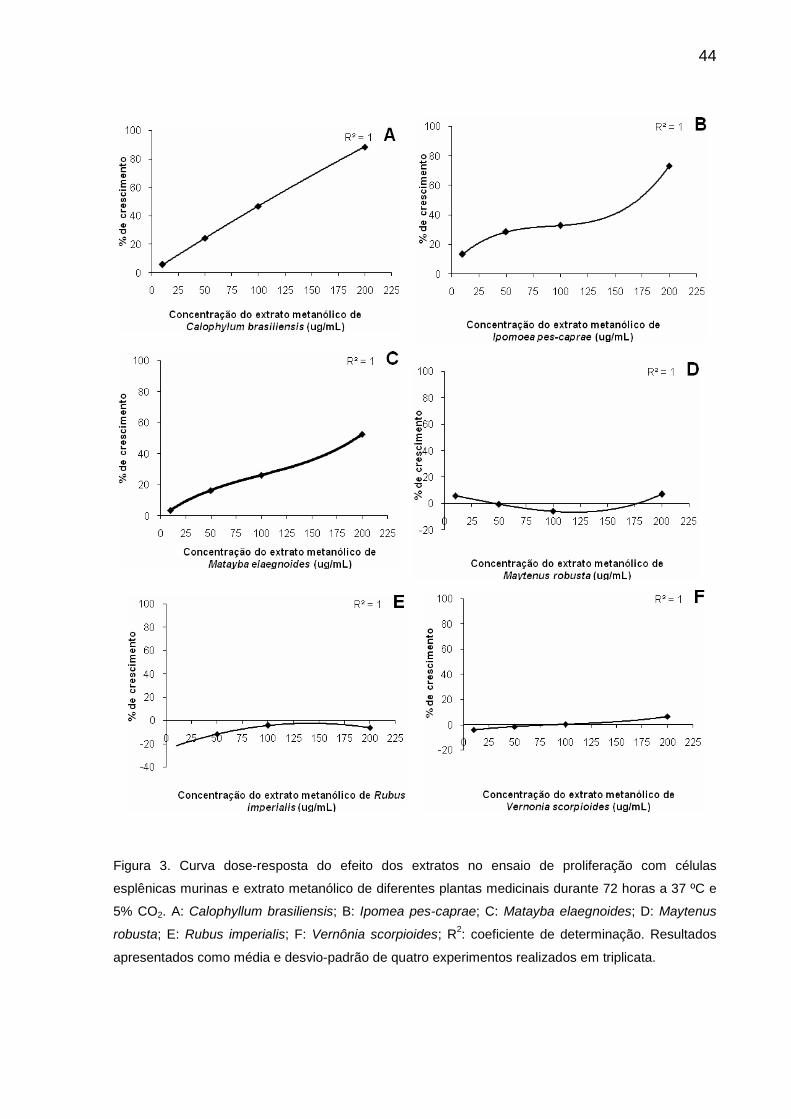
44
Figura 3. Curva dose-resposta do efeito dos extratos no ensaio de proliferação com células
esplênicas murinas e extrato metanólico de diferentes plantas medicinais durante 72 horas a 37 ºC e
5% CO2. A: Calophyllum brasiliensis; B: Ipomea pes-caprae; C: Matayba elaegnoides; D: Maytenus
robusta; E: Rubus imperialis; F: Vernônia scorpioides; R2: coeficiente de determinação. Resultados
apresentados como média e desvio-padrão de quatro experimentos realizados em triplicata.

45
A Figura 4 apresenta os resultados obtidos com os extratos metanólicos de
Calophyllum brasiliensis, Matayba elaegnoides e Maytenus robusta incubados na
presença de PHA no ensaio de proliferação celular, tendo como controle positivo o
próprio mitógeno incubado isoladamente no cultivo celular. Os resultados foram
apresentados como média e desvio padrão de quatro experimentos realizados em
triplicata.
O perfil na indução do crescimento celular com extrato metanólico de
Calophyllum brasiliensis incubado simultaneamente à PHA foi semelhante ao
observado quando o ensaio realizado com esse extrato sem estímulo mitogênico no
cultivo celular (Figura 4A), apresentando inclusive efeito dose-dependente (R2=1,0)
(Figura 5A). A percentagem média de crescimento foi discretamente superior que na
incubação do extrato isoladamente no cultivo celular, de 15,3% a 96,3%, e
igualmente o extrato a 100 e 200 µg/mL mostrou maior indução da proliferação
celular, ambas significativamente superiores ao controle positivo PHA (12,2%)
(p=0,021). A concentração de 23,9 µg/mL foi estimada como necessária para a
indução de 50% do crescimento celular.
Algumas lectinas extraídas de plantas possuem a capacidade de se ligar
especificamente a certos resíduos de açúcares nas glicoproteínas da superfície do
linfócito T, no complexo TcR : CD3, sem interagir diretamente com o sítio de ligação
ao antígeno do receptor. Esta ligação suscita um sinal semelhante ao que é dado
pelo antígeno estranho ao sistema de defesa, que é interpretado pela célula como
iniciador de saída do estado de repouso e entrada no ciclo celular a fim de realizar
mitose (PAUL, 2003). A PHA, uma proteína vegetal polimérica, é a lectina mais
extensamente estudada e aplicada na ativação dos linfócitos T in vitro, ou seja, na
avaliação da resposta imune mediada por este tipo celular (PAUL, 2003; ROCHA;
GORESCU; BELTRAME, 2007).
Analisando os resultados do extrato metanólico de Calophyllum brasiliensis
descritos acima a partir desta informação, é possível sugerir que o mesmo estimula
a proliferação de linfócitos T, provavelmente potencializando a resposta imune
celular. Além disso, a presença de ácido gálico isolado a partir de espécies de
Calophyllum tem demonstrado evitar a oxidação do DNA , impedindo assim a morte
celular pelo acúmulo de reativos como óxido nítrico no meio de cultura no qual se
encontram as células (KUHLMAN; HORSCH; BURKAARDT, 2005).

46
O extrato metanólico de Matayba elaegnoides incubado simultaneamente à
PHA também apresenta perfil de ativação celular dose-dependente como observado
quando o extrato foi incubado isoladamente (R2=1,0) (Figura 5B). A percentagem
média de crescimento foi de -7,0% a 48,8%, sendo a primeira a 10 µg/mL e com
diferença significativa frente ao controle positivo (12,2%, p=0,021) (Figura 4B). A
concentração de 19,3 µg/mL foi estimada como necessária para a indução de 50%
do crescimento celular. Embora 10 µg/mL tenha sido a única concentração com
relevância estatística, e cuja percentagem média de crescimento foi negativa, os
resultados mostram uma tendência à ativação celular induzida pelo extrato junto à
PHA, e que não foi comprovada estatisticamente devido à variabilidade encontrada
nos experimentos.
Novamente a variabilidade de compostos presentes na planta, com atividades
diversas, pode explicar a variabilidade de resultados obtidos com o extrato de
Matayba elaegnoides. Cabe aqui ressaltar que o composto 3b-O-D-glicopiranosil–
sitosterol, identificado por Souza (2006) no extrato das cascas de Matayba
elaegnoides, apresentou potente atividade citotóxica com dose letal média em 67
µg/mL quando testado frente à Artemia salina (GALOTTA ; BOAVENTURA 2005).
A percentagem média de crescimento obtida com o extrato metanólico de
Maytenus robusta nas concentrações de 10, 50, 100 e 200 µg/mL incubados
simultaneamente com PHA foi de -10,2% a 0,6%, sendo os valores das
concentrações de 10 e 100 µg/mL significativamente inferiores à do controle positivo
(12,5%, p=0,878) (Figura 4C) e com correlação com a concentração do extrato
(R2=1,0) (Figura 5C). Os resultados obtidos com o extrato incubado isoladamente
comparado a estes sugerem inibição da proliferação celular, ou ainda lise celular
promovida pelo composto adicionado à cultura. Outra espécie deste gênero, a
Maytenus ilicifolia revelou propriedades biológicas, como atividade citotóxica,
antibaceriana e anti-ulcerogênica, devido à presença de triterpenos e compostos
fenólicos (OLIVEIRA et al., 1991; CARLINI, 1988; SOUZA-FORMIGONI et al., 1991;
ZHU, 1998).
O estudo realizado anteriormente (COELHO et al., 2007) avaliou o efeito do
extrato metanólico de Ipomoea pes-caprae e de Rubus imperialis simultaneamente à
PHA no cultivo de células esplênicas murinas, demonstrando que o primeiro induz à
proliferação celular a 200 µg/mL, enquanto que o segundo extrato apresentou efeito
inibitório na proliferação celular a 10 e 100 µg/mL. Este mesmo efeito inibitório foi

47
observado com o extrato metanólico de Vernonia scorpioides a 10 e 100 µg/mL em
outro estudo recente de proliferação de células esplênicas (MENDES et al., 2007). O
efeito citotóxico da Vernonia scorpioides tem sido descrito na literatura e relacionado
às lactonas sesquiterpênicas (BREHMER, 2005; PAGNO et al., 2006; BUSKÜHL,
2007), bem como a um recente composto isolado da planta, o poliacetilieno
(BUSKÜHL, 2007).
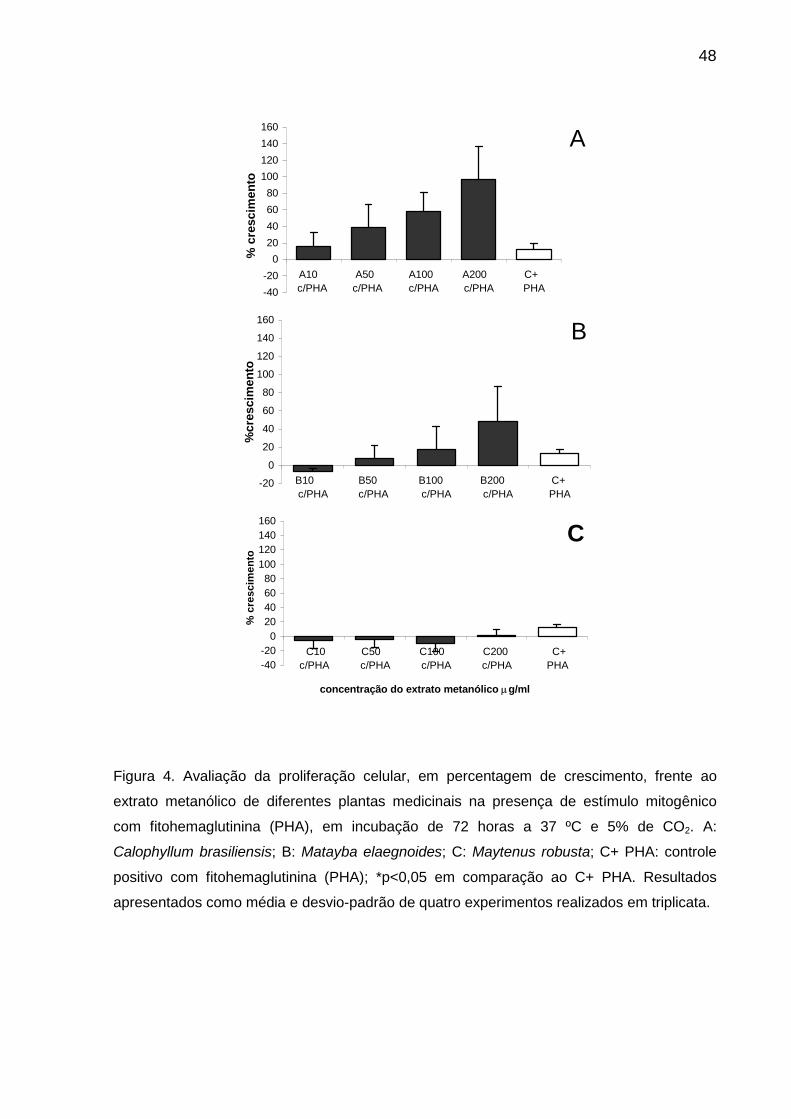
48
A
-40
-20
0
20
40
60
80
100
120
140
160
A10 c/PHA
A50 c/PHA
A100 c/PHA
A200 c/PHA
C+ PHA
% c
resc
imen
to
B
-20
0
20
40
60
80
100
120
140
160
B10 c/PHA
B50 c/PHA
B100 c/PHA
B200 c/PHA
C+ PHA
%cr
esci
men
to
Figura 4. Avaliação da proliferação celular, em percentagem de crescimento, frente ao
extrato metanólico de diferentes plantas medicinais na presença de estímulo mitogênico
com fitohemaglutinina (PHA), em incubação de 72 horas a 37 ºC e 5% de CO2. A:
Calophyllum brasiliensis; B: Matayba elaegnoides; C: Maytenus robusta; C+ PHA: controle
positivo com fitohemaglutinina (PHA); *p<0,05 em comparação ao C+ PHA. Resultados
apresentados como média e desvio-padrão de quatro experimentos realizados em triplicata.
C
-40-20
020406080
100120140160
C10 c/PHA
C50 c/PHA
C100 c/PHA
C200 c/PHA
C+ PHA
concentração do extrato metanólico mg/ml
% c
resc
imen
to
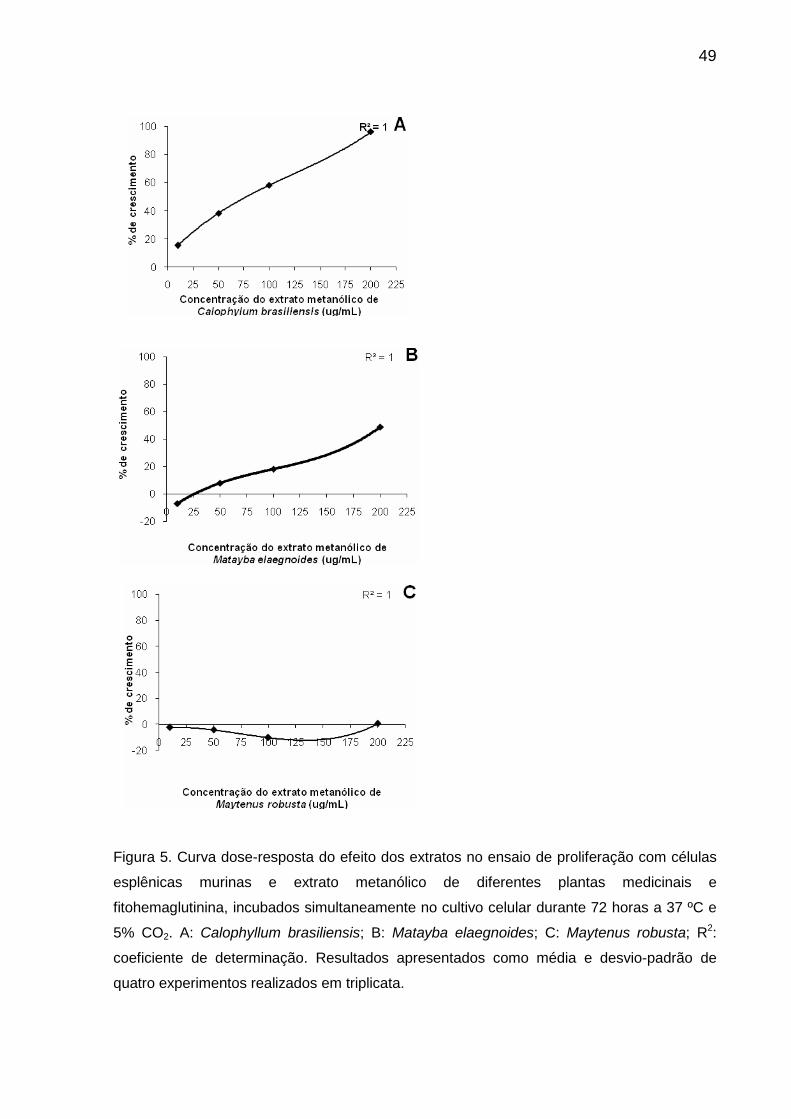
49
Figura 5. Curva dose-resposta do efeito dos extratos no ensaio de proliferação com células
esplênicas murinas e extrato metanólico de diferentes plantas medicinais e
fitohemaglutinina, incubados simultaneamente no cultivo celular durante 72 horas a 37 ºC e
5% CO2. A: Calophyllum brasiliensis; B: Matayba elaegnoides; C: Maytenus robusta; R2:
coeficiente de determinação. Resultados apresentados como média e desvio-padrão de
quatro experimentos realizados em triplicata.

50
Os resultados obtidos com os extratos metanólicos de Calophyllum
brasiliensis, Ipomoea pes-caprae, Matayba elaegnoides, Maytenus robusta, Rubus
imperialis e Vernonia scorpioides simultaneamente ao PWM no ensaio de
proliferação celular, tendo como controle positivo o próprio mitógeno incubado
isoladamente no cultivo celular, estão representados na Figura 6. Os resultados
foram apresentados como média e desvio padrões de quatro experimentos
realizados em triplicata.
Assim como observado nas Figuras 3 e 5, o extrato metanólico de
Calophyllum brasiliensis nas concentrações de 10, 50, 100 e 200 µg/mL incubado
simultaneamente com PWM mostrou indução do crescimento celular (Figura 6A)
com padrão dose-dependente (R2=1,0) (Figura 7A). A concentração de 73,5 µg/mL
foi estimada como necessária para a indução de 50% do crescimento celular. A
percentagem média de crescimento foi de 21,6% a 55,9%, sendo o menor e o maior
valor respectivamente para 10 e 200 µg/mL. Entretanto, as quatro concentrações
testadas não mostraram diferença significativa quando comparadas ao controle
positivo com PWM (10,1%) (p>0,149).
Este achado coincide com o indicativo de que o extrato induz a ativação e
proliferação de linfócitos T e que, mesmo não havendo diferença significativa nos
resultados em relação ao controle, pode-se notar um aumento na proliferação
celular. Este sutil estímulo à proliferação celular pode estar relacionado à presença
de composto da classe das xantonas, que mostrou atividade imunoestimulante tanto
in vivo quanto in vitro e um aumento significativo no título de anticorpos humorais
relacionados a ativação de linfócitos T e B em estudo realizado com extrato de
cascas da Mangífera indica (MAKARE; BODHANKAR; RANGARI, 2001).
O extrato metanólico de Ipomoea pes-caprae mostrou estímulo à proliferação
celular quando incubado junto ao PWM (Figura 6B), com correlação com a
concentração utilizada (R2=1,0) (Figura 7B). A proliferação celular foi mais intensa
nas concentrações de 100 e 200 µg/mL, diferindo significativamente do controle
positivo de PWM (10,1%) (p<0,043). A percentagem média de crescimento obtida
com o extrato variou de 20,7% a 78,2%, dependendo da concentração empregada.
A indução de 50% do crescimento celular foi estimada na concentração do extrato a
31,7 µg/mL. Estes dados, somados aos reportados no estudo com PHA (COELHO et
al., 2007), sugerem que tanto os linfócitos T quanto os linfócitos B são ativados pelo
extrato da planta. A literatura sugere que este efeito é encontrado em plantas que

51
apresentam alcalóides e flavonoides em sua composição, como no extrato aquoso
da Clausena excavata. Esta espécie apresenta atividade imumodulatória em
concentrações elevadas do extrato com estímulo dose-dependente na proliferação
de células esplênicas murinas estimuladas com PWM (MANOSROI;
SARAPHACHOTIWITTHAYA; MANOSROI, 2003).
O efeito promovido pelo extrato metanólico de Matayba elaegnoides, nas
concentrações de 10, 50, 100 e 200 µg/mL e incubado simultaneamente ao PWM na
proliferação celular (Figura 6C) foi semelhante ao observado quando o extrato foi
incubado isolado e simultaneamente à PHA no cultivo ou celular, ou seja, induziu à
proliferação celular de forma dose-dependente (R2=1,0) (Figura 7C). A percentagem
média de crescimento foi de -4,6% a 47,1%, esta última a 200 µg/mL e
significativamente superior ao controle positivo com PWM (7,5%) (p=0,021). A
concentração de 27,4 µg/mL foi estimada como necessária para a indução de 50%
do crescimento celular.
A proliferação celular significativa observada quando o extrato de Matayba
elaegnoides foi incubado isolado no cultivo celular, bem como simultâneo à PHA e
ao PWM, sugerem que o mesmo tem ação estimuladora sobre a ativação e
proliferação tanto de linfócitos T quanto de linfócitos B. Assim como a Clauseana
excavata, a Matayba elaegnoides apresenta na sua composição umbeliferona, que
tem sido relacionada com capacidade de estimular a proliferação celular quando
incubados simultaneamente ao PWM em cultura celular (MANOSROI:
SARAPHANCHOTIWITHAYA;MANOSROI, 2003).
O extrato metanólico de Maytenus robusta incubado simultaneamente ao
PWM, assim como o ensaio com o extrato isolado no cultivo celular, mostrou não
interferir no cultivo celular, seja com indução da proliferação, seja com inibição da
mesma (Figura 6D) (R2=1,0) (Figura 7D). A percentagem média de crescimento
obtida com o extrato nas concentrações de 10, 50, 100 e 200 µg/mL incubados
simultaneamente com PWM foi de 4,1% a 8,3%, tendo o controle positivo com PWM
percentagem média de crescimento de 7,5%. A concentração necessária para a
indução de 50% do crescimento celular com PWM foi estimada em 36,2 µg/mL. A
compilação dos resultados obtidos com o extrato isolado no cultivo celular, e
simultâneo à PHA e ao PWM, não permite uma conclusão acerca da atividade sobre
a proliferação de células esplênicas murinas.

52
Contudo, é possível sugerir que a presença de triterpenos e compostos com
atividade antioxidantes podem estar relacionados com os resultados encontrados co
o extrato de Maytenus robusta incubado como PHA. Nesta condição o extrato
mostrou que em doses menores ocorre inibição da proliferação celular, enquanto
que em doses maiores há diminuição deste efeito. A Clauseana excavata originária
da Índia apresenta compostos como terpenóides e flavonódes, tendo sido revelado
efeito estimulador dose-dependente sobre a proliferação celular (MANOSROI:
SARAPHANCHOTIWITHAYA;MANOSROI, 2003). Considerado que estas mesmas
classes de compostos estão presentes no extrato de Maytenus robusta, a ausência
de efeito inibidor na proliferação celular mesmo quando extrato foi incubado
simultaneamente com PWM parece ser justificada.
A inibição da proliferação celular induzida pelo extrato metanólico de Rubus
imperialis, novamente observado quando o mesmo foi incubado junto ao PWM no
cultivo celular (Figura 6E), com correlação dose-dependente (R2=1,0) (Figura 7E).A
percentagem média de crescimento para o extrato nesta condição e nas
concentrações de 10, 50, 100 e 200 µg/mL foi de -17,3% a 11,2%, sendo o efeito
observado nas concentrações de 10, 100 significantemente inferior ao controle
positivo de PWM (13,8%) (p=0,021), embora a concentração de 200 µg/mL tenha
sido superior ao mesmo controle (p=0,021). Esta variabilidade nos resultados não
permitiu a determinação da concentração necessária para o crescimento em 50% da
população celular. Esta inibição parece envolver os dois tipos de linfócitos e um
estudo mais aprofundado poderá avaliar a imunossupressão induzida pelas
substâncias presentes no extrato da planta.
O extrato metanólico de Vernonia scorpioides apresentou valores positivos na
percentagem média de proliferação para as concentrações de 10, 50, 100 e 200
µg/mL do extrato incubado simultaneamente com o mitógeno PWM (Figura 6F), com
correlação dose-dependente (R2=1,0) (Figura 7F). Contudo, os resultados não
mostraram significância estatística frente ao frente ao controle positivo realizado com
PWM (13,8%) (p>0,387). A percentagem média de crescimento foi de 3,5% a 20,8%.
A concentração de 59,5 µg/mL foi estimada como necessária para a indução de 50%
do crescimento celular. Como estes dados frente ao controle positivo de PWM não
apresentaram relevância estatística, é possível manter a indicação do efeito inibitório
observado quando o extrato foi colocado isoladamente junto às células na
concentração de 100 µg/mL, permanecendo em concordância com outros autores no

53
mesmo modelo de estudo (MENDES et al., 2007) e em estudos específicos de
citotoxicidade (BREHMER, 2005; PAGNO et al., 2006; BUSKÜHL, 2007).
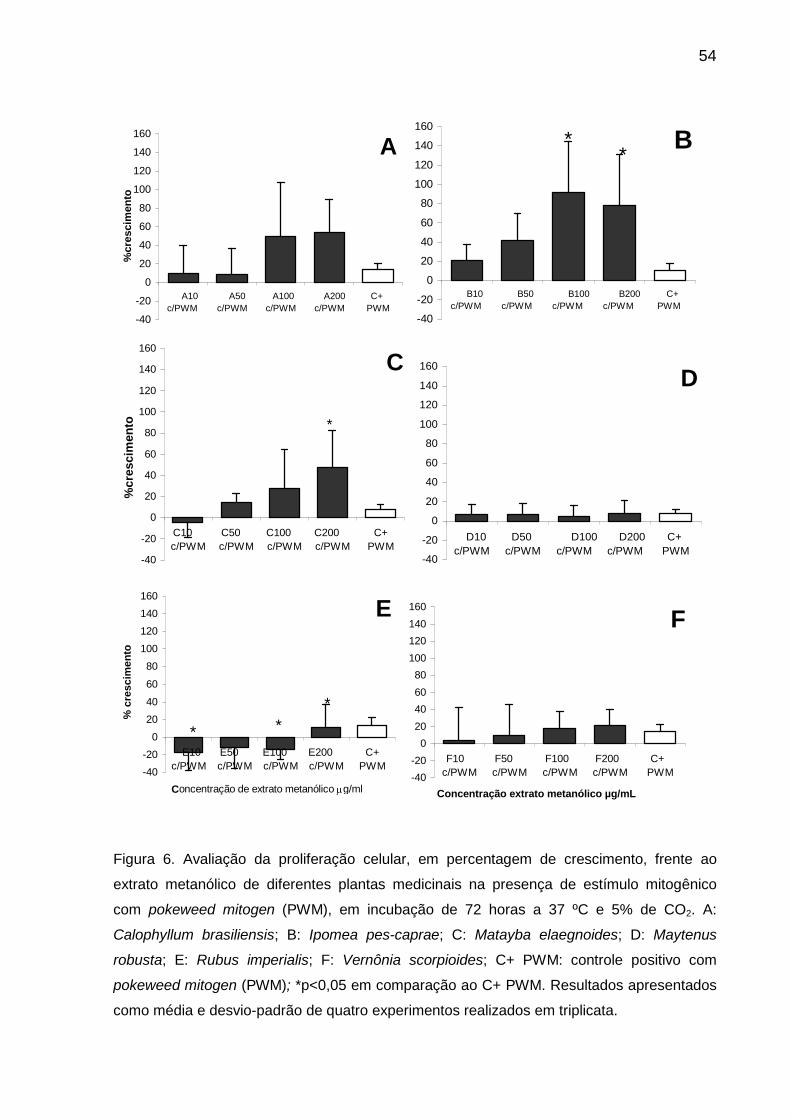
54
A
-40
-20
0
20
40
60
80
100
120
140
160
A10 c/PWM
A50 c/PWM
A100 c/PWM
A200 c/PWM
C+ PWM
%cr
esci
men
to
B
-40
-20
0
20
40
60
80
100
120
140
160
B10 c/PWM
B50 c/PWM
B100 c/PWM
B200 c/PWM
C+ PWM
**
C
-40
-20
0
20
40
60
80
100
120
140
160
C10 c/PWM
C50 c/PWM
C100 c/PWM
C200 c/PWM
C+ PWM
%cr
esci
men
to *
D
-40
-20
0
20
40
60
80
100
120
140
160
D10 c/PWM
D50 c/PWM
D100c/PWM
D200 c/PWM
C+ PWM
E
-40
-20
0
20
40
60
80
100
120
140
160
E10 c/PWM
E50 c/PWM
E100 c/PWM
E200 c/PWM
C+ PWM
Concentração de extrato metanólico mg/ml
% c
resc
imen
to
* **
F
-40
-20
0
20
40
60
80
100
120
140
160
F10 c/PWM
F50 c/PWM
F100 c/PWM
F200 c/PWM
C+ PWM
Concentração extrato metanólico µg/mL
Figura 6. Avaliação da proliferação celular, em percentagem de crescimento, frente ao
extrato metanólico de diferentes plantas medicinais na presença de estímulo mitogênico
com pokeweed mitogen (PWM), em incubação de 72 horas a 37 ºC e 5% de CO2. A:
Calophyllum brasiliensis; B: Ipomea pes-caprae; C: Matayba elaegnoides; D: Maytenus
robusta; E: Rubus imperialis; F: Vernônia scorpioides; C+ PWM: controle positivo com
pokeweed mitogen (PWM); *p<0,05 em comparação ao C+ PWM. Resultados apresentados
como média e desvio-padrão de quatro experimentos realizados em triplicata.
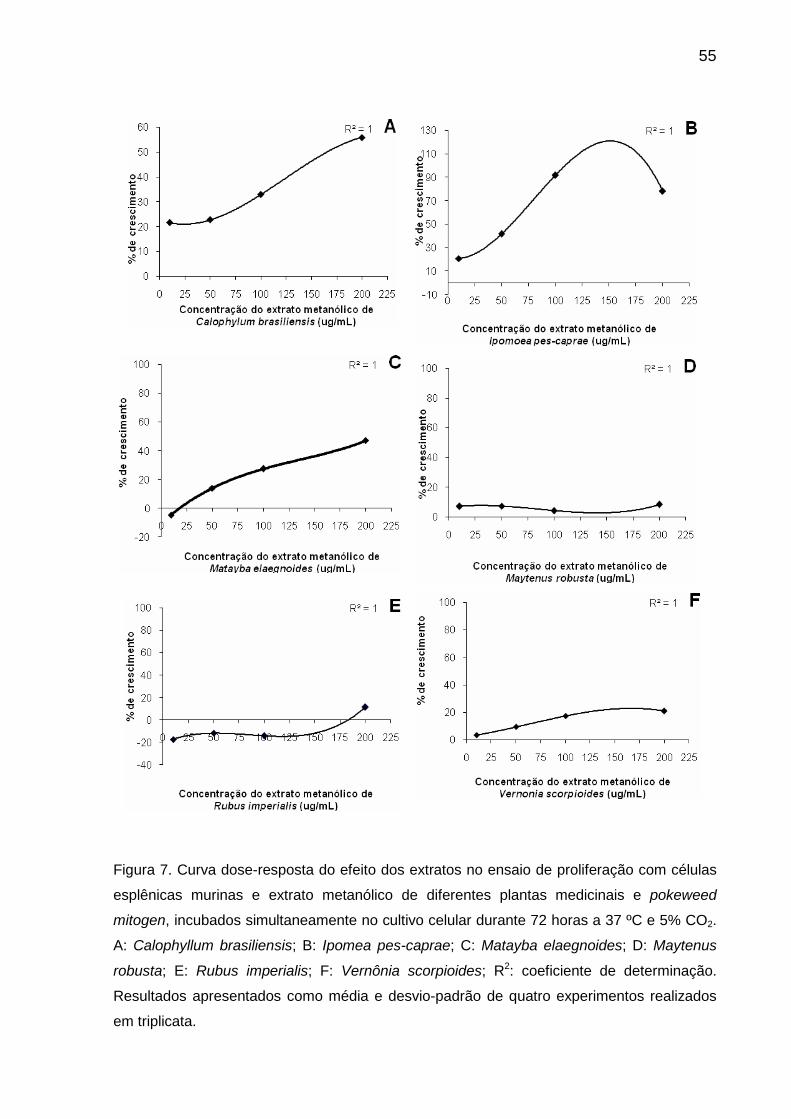
55
Figura 7. Curva dose-resposta do efeito dos extratos no ensaio de proliferação com células
esplênicas murinas e extrato metanólico de diferentes plantas medicinais e pokeweed
mitogen, incubados simultaneamente no cultivo celular durante 72 horas a 37 ºC e 5% CO2.
A: Calophyllum brasiliensis; B: Ipomea pes-caprae; C: Matayba elaegnoides; D: Maytenus
robusta; E: Rubus imperialis; F: Vernônia scorpioides; R2: coeficiente de determinação.
Resultados apresentados como média e desvio-padrão de quatro experimentos realizados
em triplicata.

56
6 CONCLUSÕES
1. O extrato metanólico de Calophyllum brasiliensis mostrou indução da
proliferação celular dose-dependente quando incubado isoladamente e
simultaneamente à PHA e ao PWM no cultivo celular. As concentrações de 23,2 a
73,5 µg/mL foram estimadas como necessárias para a indução de 50% do
crescimento celular. As concentrações mais elevadas do extrato, de 100 e 200
µg/mL, mostraram maior indução na proliferação celular em comparação com os
controles positivos PHA e PWM, tanto incubado isoladamente quanto em conjunto
com a PHA (p<0,029). Contudo, este efeito não foi observado quando o extrato foi
incubado simultaneamente ao PWM (p>0,149). Estes dados sugerem que o extrato
metanólico de Calophyllum brasiliensis estimula a proliferação de linfócitos T e
possibilita potencializar a resposta imune celular.
2. O extrato metanólico de Ipomoea pes-caprae incubado isolado no cultivo
celular induziu a proliferação celular de forma dose-dependente das células
esplênicas e significativamente superior aos controles positivos PHA e PWM na
concentração de 200 µg/mL (p=0,021). As concentrações de 31,7 a 40,7 µg/mL
foram estimadas como necessárias para a indução de 50% do crescimento celular.
Quando incubado simultaneamente ao PWM houve incremento da proliferação
celular com correlação com a concentração utilizada e significante. O estudo anterior
realizado pela equipe demonstrou que o extrato metanólico de Ipomoea pes-caprae
incubado simultaneamente à PHA no cultivo de células esplênicas murinas induz à
proliferação celular a 200 µg/mL (COELHO et al., 2007). Assim, a partir destes
dados é possível propor que tanto os linfócitos T quanto os linfócitos B são ativados
pelo extrato da planta.
3. O extrato metanólico de Matayba elaegnoides mostrou indução dose-
dependente da proliferação das células esplênicas quando incubado isolado e
simultâneo à PHA e ao PWM. As concentrações de 19,3 a 33,6 µg/mL foram
estimadas como necessárias para a indução de 50% do crescimento celular. O

57
extrato isolado mostrou inibição da proliferação celular a 10 µg/mL (p=0,030) e
indução do crescimento celular a 200 µg/mL frente ao controle positivo com PHA e
com PWM (p=0,021). Para o extrato incubado junto à PHA, a concentração de 10
µg/mL foi inferior ao controle positivo PHA (p=0,021), mas com tendência à indução
da proliferação celular em concentrações maiores. Quando incubado com PWM, o
extrato na concentração de 200 µg/mL mostrou indução do crescimento celular
superior ao controle positivo com PWM (p=,0,021). Desta forma, é sugestivo que o
extrato tem ação estimuladora sobre a ativação e proliferação tanto de linfócitos T
quanto de linfócitos B em concentrações maiores, enquanto que em baixas
concentrações o efeito é supressor.
4. O extrato metanólico de Maytenus robusta mostrou não interferir
significativamente no cultivo celular quando incubado isolado e simultâneo ao PWM
(p>0,083), embora o discreto efeito observado quando isolado no cultivo celular
tenha sido dose-dependente. Contudo, quando o extrato foi incubado nas
concentrações de 10 e 100 µg/mL junto à PHA houve inibição total do efeito
estimulador da mesma sobre os linfócitos T (p=0,021). Esta diversidade de
resultados não permite fazer inferências quanto à atividade do extrato sobre a
proliferação de células esplênicas murinas. Somado a estes achados, a
concentração de 36,2 foi estimada como necessária para a indução de 50% do
crescimento celular frente aos linfócitos B.
5. O extrato metanólico de Rubus imperialis apresentou inibição comprovada da
proliferação celular nas concentrações de 10 e 100 mg/mL frente aos controles
positivos com PHA e PWM (p<0,030), incubado isolado e simultaneamente ao PWM,
em concordância com estudos anteriores no mesmo modelo de avaliação
empregando PHA como mitógeno (COELHO et al., 2007). Ainda, o extrato
apresentou efeito dose-dependente quando incubado junto ao PWM no cultivo
ceular. A concentração necessária para a indução de 50% do crescimento celular
não foi possível de ser estimada devido à supressão promovida pelo extrato. A
inibição parece envolver linfócitos T e B, e um estudo mais aprofundado é indicado
para determinar se este efeito é citotóxico ou supressor, podendo então ser avaliada
a imunossupressão induzida pelas substâncias presentes no extrato da planta.

58
6. O extrato metanólico de Vernonia scorpioides teve comprovado efeito inibitório
apenas quando incubado a 100 µg/mL isoladamente no cultivo celular (p=0,043),
mas que é concordante com outros autores no mesmo modelo de estudo
empregando o mitógeno PHA (MENDES et al., 2007) e em estudos específicos de
citotoxicidade (BREHMER, 2005; PAGNO et al., 2006; BUSKÜHL, 2007). O efeito
inibidor na proliferação celular foi dose-dependente tanto quando o extrato foi
incubado isolado quanto quando incubado simultâneo ao PWM no ensaio de
proliferação celular. As concentrações de 11,3 a 59,5 µg/mL foram estimadas como
necessárias para a indução de 50% do crescimento celular com este extrato.

59
7. PERSPECTIVAS
Este estudo pioneiro sobre a atividade dos extratos de plantas medicinais
estudadas pelo grupo de pesquisadores do NIQFAR na proliferação de células
esplênicas de camundongos possibilitou identificar efeitos diversos dependendo da
planta estudada. Esta variação certamente é determinada pela composição química
apresentada pelos diferentes extratos utilizados, associação de moléculas esta que
beneficia a proliferação celular ou, contrariamente, tem efeito tóxico para as células.
A composição de substâncias presentes no extrato pode ainda atuar
potencializando ou apenas bloqueando os diferentes mecanismos envolvidos na
ativação das células do sistema imune. Estudos complementares de frações desses
extratos que apresentaram potencial, tanto no aumento da proliferação celular
quanto na atividade de inibir tal proliferação, devem ser ter seqüência, afim
identificar a(s) substância(s) resposável(is) pela atividade. O desenvolvimento de
estudos de Imunologia Celular e Molecular possibilitarão caracterizar o mecanismo
envolvido na ativação ou inibição celular, bem como no efeito citotóxico.
Por fim, a comprovação e caracterização de substância(s) capaz(es) de atuar
e modular as atividades do sistema imune possibilitarão, uma vez comprovada a
mesma atividade em modelo humano, ampliar as ferramentas utilizadas no combater
as desordens que acometem esse sistema. As plantas contêm centenas de
metabólitos secundários, mas geralmente, apenas os compostos presentes em
maior concentração são isolados e estudados pela fitoquímica clássica, já que a
obtenção destes é bastante complexa. Por isso, é indispensável análise biológica
das frações e das substâncias puras em relação à sua concentração, a fim de
predizer se o principal componente químico responsável pela atividade biológica foi
realmente determinado, ou se a ação biológica deve-se ao efeito sinérgico da vários
compostos.

60
REFERÊNCIAS
ABOSI, A. O.; RASEROKA, B. H. In vivo antimalarial activity of Vernonia amygdalina. Br. J. Biomed. Sci., v. 60, p. 89-91, 2003.
ALVES, M. H. A diversidade química das plantas como fonte de fitofarmacos. Cadr. Tem. Química Nova Esc., v. 3, 2001.
ANDRADE, F. S.; LEMOS, M.; COMUNELLO, E.; NOLDIN, F. V.; CHECHINEL FILHO, V.; NIERO, R. Evaluation of the antiulcerogenic activity of Maytenus robusta in different experimental ulcer models. J. Ethnopharmacol., v. 125, p. 247-265, 2007.
ARDENGHI, J.; KANEGUSUKU, M.; NIERO, R.; MONACHE, F. D.; CECHINEL FILHO, V.; SOUZA, M. M.; Analysis of the mechanism of antinoceptive action of niga-ichicoside obtained from Rubus imperialis. J. Pharm. Pharmacol, v. 58, p. 1669, 2006.
BALUNAS, M.J.; KINGHOM, A. D. Drug discovery from medicinal plants. Life Sci., v. 78, p. 31-41, 2005.
BOYUM, A. Separation of leukocytes from blood and bone marrow. Scand. J. Clin. Lab. Invest., v. 97, p. 7, 1968.
BREHMER, J. S. Estudo de extratos de plantas medicinais no desenvolvimento do tumor ascítico de Ehrlich. Dissertação (Mestrado em Ciências Farmacêuticas) - Universidade do Vale do Itajaí, UNIVALI, Itajaí, 2005.
BUSKÜHL, H. Avaliação in vitro do mecanismo de ação citotóxico de lactonas sesquiterpênicas e outras substâncias isoladas de Vernonia scorpioides (LAM) PERS. Dissertação (Mestrado em Ciênicas Farmacêuticas) - Universidade do Vale do Itajaí, Itajaí, 2007.
BUSS, A. D.;WAIGH, R. D. In Natural compounds as leads for new pharmaceuticals. Burger's medical chemistry and drug discovery, p. 983-1033, 1995.
CABRERA, A. L.; KLEIN, R. M. Compostas: 3. Tribo: Vernoniae. Fl. Ilustr. Catar., Itajaí, p. 354-355, 1980.
CAI, Q.; RANHN R. O.; ZHANG R. Dietary flavonoids, quercitin, luteolin and genistein reduce oxidative DNA damage and lipid peroxidation and quench free radicals. Cancer Letters, v. 210, p.99-107, 2004.
CALIXTO, J. B. Medicamentos Fitoterápicos. In: Yunes, R. A.; Calixto, J. B. Plantas Medicinais Sob a Ótica da Química Medicinal Moderna. Chapecó: Argos, p. 297-315, 2001.
CAMPOS, M.; OROPEZA, M.; PONCE, H.; FERNANDEZ, J.; JIMENEZ-ESTRADA, M.; TORRES, H.; REYES-CHILPA, R. Relaxation of uterine and aortic smooth muscle by glaucolides D and E from Vernonia liatroides. Biol. Pharm. Bull., v. 26, p. 112-115, 2003.

61
CARLINI, E.L.A. Estudo da ação antiúlcera gástrica de plantas brasileiras: Maytenus ilicifolia (Espinheira Santa) e outras. Brasília: CEME/AFIP, 1988.
CARVALHO, J. E. Fitoterápico: Alimento ou medicamento? In: MERCADANTEW, A. Z.; BOBBIO, F. O.; BOBBIO, P. A.; PEREIRA, J. L.; PASTORE, G. M. Ciências de alimentos: avanços e pespectivas. Campinas: UNICAMP, 196-202, 2001.
CECHINEL FILHO, V. Principais Avanços e perspectivas na Área de Produtos Naturais Ativos: Estudos Desenvolvidos no Niqfar/ Univali. Química Nova, v. 23, n. 5, p. 680-685, 2000.
CECHINEL FILHO, V.; YUNES, R. A. Algumas Considerações sobre a função e importância da fitoquímica de plantas medicinais para a obtenção de novos medicamentos. Química Nova, v. 21 n. 1, p. 99-105, 1998.
COELHO, F.; FERREIRA, J.; ZANDONAI, R. H.; NIERO, R.; CECHINEL FILHO, V.; BUENO, E. C. Proliferative activity of Ipomoea pes-caprae and Rubus imperialis methanolic extracts on lymphoblastogenesis assay using murine spleen cells stimulated with phytohemagglutinin. In: INTERNATIONAL CONGRESSO F PHARMACEUTICAL SCIENCES, 6., Ribeirão Preto, Anais. Ribeirão Preto: CIFARP, 2007.
COOPER, H. L. Studies on RNA metabolism during lymphocyte activation. Transplant. Rev., v. 11, p.3-38, 1972.
CORREA, M. P. Diccionario das Plantas úteis do Brasil e das exóticas cultivadas. Rio de Janeiro : Imprensa Nacional, 1978.
CRASTON, R.; KOH, M.; DERMOTT, A.; RAY, N.; PRENTICE, H. G.; LOWDELL, M. W. Temporal dynamics of CD69 expression on lymphoid cells. J. Immunol. Methods, v. 209, p. 37-45, 1997.
CRUZ-CHAMORRO, L.; PUERTOLLANO, M. A.; PUERTOLLANO, E.; CIENFUEGOS, G. A.; PABLO, M. A. In vitro biological activities of magainin alone or in combination with nisin. Peptides, v. 27, p. 1201-1209, 2006.
DENIZOT, F.; LANG, R. Rapid Colorimetric Assay for Cell Growth and Survival: Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods, v. 89, p. 271-277, 1986.
DHARMARATANE, H.R.W.; WANIGASEKERA, W.M.A.P. Xantones from root bark of Calophyllum thwateni. Phytochemestry, v. 42, p. 249-250, 1996.
FORMAN H, J.; TORRES, M. Reactive Oxygen Species and Cell Signaling Am. J. Respir. Crit. Care Med., v. 166, p. S4-S8, 2002.
FREIRE, M. F. I.; ABREU, H. S.; CRUZ, L. C. H.; FREIRE, R. B. Inhibition of fungal growth by extracts of Vernonia scorpioides (Lam.) Pers. Microbiology Reading, v. 27, p. 1-6, 1996.
GALOTTA, A.L..Q.A.; BOAVENTURA, M.A.D. Constituintes químicos da raiz e do talo da folha do açaí (Euterpe precatória, Mart Arecaceae) , Quimica Nova, v. 28, p. 610-613, 2005.
GASPAROTTO Jr., A .; MISLAINE, B. A.; IZABEL, C. P.; CORTEZ, A. G.; NAKAMURA, C. V.; PRADO, D. B.; RODRIGUES, E.; FERREIRA, A .G. Estudo fitoquímico e avaliação da atividade moluscicida do Calophyllum brasiliensis. Química Nova, v. 8, p. 575-578, 2005.

62
GREAVES, M.; JANOSSY, G. Elicitation of selective T and B lymphocyte responses by cell surface binding ligands. Transplant. Rev., v. 11, p.87-130, 1972.
GUPTA, M.; MAZUMDER, U.K.; MANIKANDAN, L.; BHATTACHARYA, S.; HALDAR, P.K.; ROY, S. Evaluation of antipyretic potential of Vernonia cinerea extract in rats. Phytomedicine, v.17, p. 804-806, 2003.
HENRY, J. B. Diagnóstico Clínico e Tratamento por Métodos de Laboratório: Aspectos Gerais do Sistema Imune, 19. ed., São Paulo: Manole, 2003, 1551 p.
HOSTETTMANN, K.; QUEIROZ, E. F.; VIEIRA, P. C. Princípios Ativos de Plantas Superiores. São Carlos: Edufscar, 2003. p. 9-43.
HOWARD M.; GARRA A.; ISHIDA H. Biological properties of interleukin 10. J. Clin. Immunol., v. 12, p. 239-247, 1992.
HOWARD, C. B.; STEVENS, J.; IZEVBIGIE, E. B.; WALKER, A.; MCDANIEL, O. Time and dose-dependent modulation of phase 1 and phase 2 gene expression in response to treatment of MCF-7 cells with a natural anti-cancer agent. Cell. Mol. Biol., v. 49, p. 1057-1065, 2003.
HUANG, Y.; DING, Z. H.; LIU, J. K. A new highly oxygenated flavone from Vernonia saligna. Z. Naturforsch., v. 58, p. 347-350, 2003.
ISAIAS, D. E. B.; NIERO, R.; NOLDIN, V. N.; BUZZI, F. C.; YUNES, R. A.; DELLEMONACHE, F.; CECHINEL FILHO, V.; Pharmacological and phytochemical investication of different parts of Calophyllum brasiliensis. Pharmazie, v.59, p. 879-881, 2004.
ITO, C.; ITOIGAWA, M.; MIYAMOTO, Y, RAO, K. S.; TAKAYASU, J.; OKUDA, Y.; MUKAINAKA, T.; TOZUDA, H.; NISHINO, H.; FURUKAWA, H. A new biflavonoide from Callophyllum panciflorum with antitumor-promoting activity. J. Nat. Prod., v. 62, p. 1668-16671, 1999.
ITOIGAWA, M.; ITO, C.; TAN, H. T. W.; KUCHIDE, M.; TOZUDA, H.; NISHINO, H.; FURKAWA, H. Câncer chemopreventive agentes, 4-phenylcoumarins from Calophyllum inophyllum. Câncer. Let., v.169, p. 15-19, 2001.
IWALEWA, E. O.; IWALEWA, O. J.; ADEBOYE, J. O. Analgesic, antipyretic, anti-inflammatory effects of methanol, chloroform and ether extracts of Vernonia cinerea less leaf. J. Ethnopharmacol., v. 86, p. 229-234, 2003.
IZEVBIGIE, E. B.; BRYANT, J. L.; WALKER, A. A novel natural inhibitor of extracellular signal-regulated kinases and human breast cancer cell growth. Exp. Biol. Med., v. 229, p.163-169, 2004.
KANEGUSUKU, M. Analise fitoquímica e biológica das raízes de Rubus imperialis. Itajaí 2001. Monografia . Curso de Farmácia - Universidade do Vale do Itajaí.
KAUSCH, I.; LINGNAU, A.; ENDL, E.; SELLMANN, K.; DEINERT, I.; RATLIFF, T. L.; JOCHAM, D.; SCZAKIEL, G.; GERDES, J.; BÖHLE, A. Anti-sense treatment against Ki-67 mRNA inhibits proliferation and tumor growth in vitro and in vivo. Int. J. Cancer, v. 105, p. 710-716, 2003.

63
KRAJEWSKA, M.; WANG, H. G.; KRAJEWSKI, S.; ZAPATA, J. M.; SHABAIK, A.; GASCOYNE, R.; REED, J. C. Immunohistochemical analaysis of in vivo patterns of expression of CPP32 (Caspase-3), a cell death protease. Cancer Research, v. 57, p. 1605-1613, 1997.
KRISHNA KUMARI, G. N.; MASILAMANI, S.; GANESH, M. R.; ARAVIND, S.; SRIDHAR, S. R. Zaluzanin D: a fungistatic sesquiterpene from Vernonia arborea. Fitoterapia, v. 74, p. 479-482, 2003
KROGH, R.; KROTH, R.; BERTI, C.; MADEIRA, A. O.; SOUZA, M. M.; CECHINEL FILHO, V.; DELLE-MONACHE, F.; YUNES, R. A. Isolation and identification of compounds with antinociceptive action from Ipomoea pes-caprae (L.) R. Br. Pharmazie, v. 54, n. 6, p. 464-466, 1999.
KRUEGER, C. L.; PETTRES, M. C.; SIMAS, R.; STEIL, A. A.; SATO, D.; BUENO, E. C. Padronização do teste de linfoproliferação com fitohemaglutinina para estudo da resposta imune celular. In: JORNADA FARMACÊUTICA, 6.; CICLO DE ATUALIZAÇÃO EM INVESTIGAÇÕES QUÍMICO-FARMACÊUTICAS – CAIQFAR, 6., 2005, Itajaí. Anais, Itajaí: UNIVALI, 2005. p. 13.
KUHLMAN M.K.; HORSCH E.; BURKAARDT G. Reduction of cisplatin toxicity in cultured renal tubular cells by the biflavonoides, quercitin. Arch Toxicol, v. 156, p. 536-540, 2005.
KUO, Y. H.; KUO, Y. J.; YU, A. S.; et al. Two novel sesquiterpene lactones, cytotoxic vernolide-A and -B, from Vernonia cinerea. Chem. Pharm. Bull., v. 51, p. 425-426, 2003.
LAMBERTINI, E.; PIVA, R.; KHAN, M. T.; LAMPRONTI, I.; BIANCHI, N.; BORGATTI, M.; GAMBARI, R. Effects of extracts from Bangladeshi medicinal plants on in vitro proliferation of human breast cancer cell lines and expression of estrogen receptor alpha gene. Int. J. Oncol., v.24, p. 419-423, 2004.
LEITE, S. N.; PALHANO, G.; ALMEIDA, S.; BIAVATTI, M. W. Wound healing activity and systemic effects of Vernonia scorpioides extract in guinea pig. Fitoterapia, v. 73, p. 496-500, 2002.
LIMA, H. C. Fatos e mitos sobre imunomoduladores. An. Bras. Dermatol., n. 82, p. 207-221, 2007.
LOCHER, C. P.; WITVROUW, M.; DE BÉTHUNE, M. P.; BURCH, M. T.; MOWER, H. F.; DAVIS, H.; LASURE, A.; PAUWELS, R.; DE CLERCQ, E.; VLIETINCK, A. J. Antiviral activity of Hawaiian medicinal plants against human immunodeficiency virus type-1 (HIV-1). Phytomedicine, v. 2, p. 259-264, 1996.
LORENZI, H. Árvores brasileiras. Nova Odessa: Editora Plantarum, 1998. 222p.
LORENZI, H. Plantas Daninhas do Brasil: terrestres, aquáticas parasitas e tóxicas. 3 ed. São Paulo: Instituto Plantarum, 2000. p. 208-209.
LUNA D.; RODRIGUES O.M.; RADA M.; MENDEZ J.; Actividad biológica de flavonoides. In: SIMPÓSIO INTERNACIONAL DE QUÍMICA DE PRODUCTOS NATURALES Y SUS APLICACIONES, 4. Anais, p. 134-135, 1996.
MAINO, V. C.; SUNI, M. A.; RUITENBERG, J. J. Rapid flow cytometric method for measuring lymphocyte subset activation. Cytometry, v. 20, p. 127-133, 1995.
MAKARE, N.; BODHANKAR, S.; RANGARI, V. Immunomodulatory activity of alcoholic extract of Mangigefa indica L. in mice. J. Ethnopharmacology, v. 78, p. 133-137, 2001.

64
MANOSROI, A.; SARAPHANCHOTIWITTHAYA, A.; MANOSROI, J. Immunomodulatory activities of Clausena excavate Burm. f. wood extracts. J. Ethnopharmacology, v. 89, p. 155-160, 2003.
MARGIS, M. M.; SANDRONI, M.; LUMERZHEIN, M.; OLIVEIRA, D. E. A defesa das plantas contra as doenças. Ciência Hoje, v. 25, p. 24-32, 1999.
MARTÍNEZ, M. L.; MORENO-CASASOLA. Effects of burial by sand on seedling growth and survival in six tropical sand dune species from the Gulf of Mexico. Journal of Coastal Research, v.12, p. 406-419, 1996.
MAZUMDER, U.K.; GUPTA, M.; MANIKANDAN, L.; BHATTACHARYA, S.; HALDAR, P.K; ROY, S. Evaluation of anti-inflammatory activity of Vernonia cinerea Less. extract in rats. Phytomedicine, v. 10, p. 185-188, 2003
MEHROTRA, S.; MISHRA, K. P.; MAURYA R.; SRIMAL R. C.; YADAV, V. S.; PANDEY, R.; SINGH V. K. Anticellular and immunosuppressive properties of ethanolic extract of Acorus calamus rhizome. Int. Immunopharmacol., v.3, p. 53-61, 2003.
MENDES, A. K. B.; COELHO, F.; FERREIRA, J.; SILVA, C. V.; DUARTE, B. M.; FELIPPE, M. E.; ZANDONAI, R. H.; BIAVATTI, M. W.; BUENO, E. C. Evaluation of the in vitro immunomodulatory activity of Vernonia scorpioides methanolic extract. In: INTERNATIONAL CONGRESSO F PHARMACEUTICAL SCIENCES, 6., Ribeirão Preto, Anais. Ribeirão Preto: CIFARP, 2007.
MONTEIRO, M.H.; GOMES-CARNEIRO, M.R.; FELZENSZWALB, I.; CHAHOUD, I.; PAUMGARTTEN, F.J. Toxicological evaluation of a tea from leaves of Vernonia condensata. J. Ethnopharmacol., v. 74, p. 149-157, 2001.
MOSMANN, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays . J. Immunol. Methods, Amsterdan, v. 65, p. 55-63, 1983.
MYERS, R. L. Immunology - a laboratory manual. Dubuque: WCB Publishers, 1995.
NERGARD, C. S. ; DIALLO, D. ; MICHAELSEN, T. E. et al. Isolation, partial characterisation and immunomodulating activities of polysaccharides from Vernonia kotschyana Sch. Bip. ex Walp. J. Ethnopharmacol., v. 91, p. 141-152, 2004.
NIERO, R.; KANEGUSUKU, M.; SOUZA, M. M.; YUNES, R. A.; CECHINEL FILHO, V. Antinocieptive action of extracts and fractions from Rubus imperialis (Rosaceae). Therapie, v. 57, p. 242-245, 2002.
NIERO, R.; MOSER, R.; BUSATO, A. C. B.; YUNES, R. A.; REIS, A.; CECHINEL FILHO, V. A comparative chemical study of Maytenus ilicifolia Mart. Reiss and Maytenus robusta Reiss (Celastraceae). Z. Naturforch., v. 56, p. 158-161, 2001.
NIERO, R.; CECHINEL FILHO, V.; SOUZA, M. M.; MONTANARI, J. L.; YUNES, R. A.; MONACHE F. D.; Antinonoceptive Activity of Niga – ichigoside F1 from Rubus imperialis. J. Nat. Prod., v. 62, p. 1145-1146, 1999.
OLIVEIRA, M.G.; MONTIERO,M.G.; MACAUBAS, C.; BARBOSA, V.P.; CARLINI, E.A. Pharmacology and toxicology effects of two Maytenus species in laboratory animals. J. Ethnopharmacol., v. 34, p. 29-41, 1991.

65
OTTO, R. G.; KAPLAN, C. A. M.; ORIN, R. M. Biodiversidade um enfoque químico. Rio de Janeiro: Edidtora UFRJ, p. 20-32, 1996.
PAGNO, T.; BLIND, L. Z. ; BIAVATTI, M. W. ; KREUGER, M. R. . Cytotoxic activity of the dichloromethane fraction from Vernonia scorpioides (Lam.) Pers. (Asteraceae) against Ehrlich's tumor cells in mice. Brazilian Journal of Medical and Biological Research, v. 39, p. 1483-1491, 2006.
PANDIMA DEVI, K.; SAI RAM, M.; SREEPRIYA, M.; ILAVAZHAGAN, G.; DEVAKI, T. Immunomodulatory effects of Premna tomentosa extract against Cr (VI) iducet toxicity in splenic lymphocytes – an in vitro study. Biomedicine & Pharmacotherapy, v. 57, p. 105-108, 2003.
PARK, J.; SHIM, K. J.; DO, W. J. ; KIM, Y. S. ; SEO, K. E.; KWON, L.; KIM, K. J.; CHOI, Y.D.; KIM, C.H. Immune-stimulating properties of polysaccharides from Phellodentri cortex. Glycoconjugate Journal, v. 16, p. 247-252, 1999.
PATEL A. V.; ROJAS-VERA J.; DACKE C. G. Therapeutic Constituents and Actions of Rubus Species. Current Medicinal Chemistry, v. 11, p. 1501-1512, 2004.
PATO KA J. Biologically active pentacyclic triterpenes and their current medicine signication. J. App. Biomed., v.1, p. 7 – 12, 2003.
PATWARDHAN, B.; KALBAG, D.; PATKI, P. S.; NAGSAMPAGI, B. A. Search of immunomodulatory agents: a review. Indian Drugs, v. 28, p. 348-358, 1990.
PAUL, W. E. Fundamental Immunology. 4 ed. Philadelphia: Lippincott Williams & Wilkins, 2003. 1502 p.
PETTRES, M. C.; KRUEGER, C. L.; SIMAS, R.; STEIL A. A.; SATO, D.; BUENO, E. C.; Padronização do teste de linfoproliferação com fitohemaglutinina para estudo da resposta imune celular. Cofa, v. 5, p. 14-17, 2006.
PHILLIPSON, J. D. 50 years of medicinal plant research – every progress in methodology is a progress in science. Planta Med., v. 69, p. 491-495, 2003.
PONGPRAYOON, U.; BOHLIN, L.; SANDBERG, F.; WASUWAT, S. Inhibitory effect of extract of Ipomoea pes-caprae on guinea-pig ileal smooth muscle. Acta Pharm. Nord., v. 1, p. 41-44, 1989.
PONGPRAYOON, U.; BOHLIN, L.; WASUWAT, S. Neutralization of toxic effects of different crude jellyfish venoms by an extract of Ipomoea pes-caprae (L.) R. Br. J. Ethnopharmacol., v. 35, n. 1, p. 65-69, 1991.
POSMANTUR, R.; WANG, K .K .W.; GILBERTSEN, R. B. Caspase-3 like activity is necessary for IL-2 release in activated Jurkt T cells. Exp. Cell Res., v. 244, p. 302-309, 1998.
RAO, Y. V.; CHAKRABARTI, R. Stimulation of immunity in Indian major carp Catla catla with herbal feed ingredients. Fish Shellfish Immunol., v.18, p. 327-334, 2005 .
REITZ, R. Flora Ilustrada Catarinense. Itajaí : Herbário Barbosa Rodrigues, 1980.

66
REYES, C.P.; NÚNEZ, M. J.; JIMÉNEZ, I. A..; BUSSEROLLES, J.; ALCARAZ, M. J.; BAZZOCCHI, I. L. Activity of lupane triterpenoids from Maytenus species as inhibitors of nitric oxide and prostaglandin. Bioorg. Méd. Chem. v. 14, p. 1573-1579, 2006.
RIBEIRO, A. Identificadas 124 plantas medicinais somente na região da Floresta Nacional. Caçador: Informe Empresarial, 2004. p. 6.
RISCO, E.; GHIA, F.; VILA, R.; IGLESIAS, J.; ALVAREZ, E.; CANIGUERAL, S. Immunomodulatory activity and chemical characterisation of sangre de drago (dragon's blood) from Croton lechleri. Planta Med., v. 69, n. 9, p. 785-794, 2003.
ROCHA, G. G. ; SIMÕES, M. ; LÚCIO, K. A. ; OLIVEIRA, R. R. ; KAPALN, M. A. C. ; GATTASS, C. R. . Natural triterpenoids from Cecropia lyratiloba are cytotoxic to both sensitive and multidrug resistant leukemia cell lines. Bioorganic & Medicinal Chemistry, v. 15, p. 7355-7360, 2007.
ROSE, B. A. Espinheira Santa. 1997. Jardim das Flores. Disponível no endereço eletrônico: <http:// www.jardimdeflores.com.br>. Acesso em: 15 dez. 2005.
ROSEGHINI, R.; MOREIRA ,P.; VALE, V.; PINHEIRO, A. M.; COSTA, J. F.; BITTENCOURT, T.; NASCIMENTO, I.; SCHAER, R.; VELOZO, E.; EL-BACHÁ, R.; MEYER, R.; FREIRE, S. Different effects of arborinine alkaloid obtained from Brazilian Erthela baihensis on spleen and thymus cells stimulated in vitro with different mitogens. Immunopharmacol Immunotoxicol., v.28, p. 361-376, 2006.
SANTOS, C. R.; ARRUDA, V. L. V. Floração, predação de flores e frutificação de Ipomoea pes-caprae e I. imperati (Convolvulceae). Insula, v. 24, p. 15-36, 1995.
SARTORI, N. T.; CANEPELLE, D.; SOUSA, Jr., P. T.; MARTINS, D. T. O. Gastroprotective effect from Calophyllum brasiliense Gamb.bark on experimental gastric lesionsin rats and mice. J. Ethnopharmacol. v.67, p. 149-156, 1999.
SCHOLZEN, T.; GERDES, J. The Ki-67 protein: from the known and the unknown. J. Cell Physiol., New York, v.182, p.311-322, 2000.
SHENG, Y., BRYNGELSSON, C.; PERO, R. W. Enhanced DNA repair, immune function and reduced toxicity of C-Med-100, a novel aqueous extract from Uncaria tomentosa. J. Ethnopharmacol., v. 69, p. 115-126, 2000.
SILVA, K. L; SANTOS, A. R. S.; CECHINEL FILHO, V.; Estudo fitoquímico e farmacológico. In: SEMINÁRIO INTEGRADO DE INICIAÇÃO CIENTÍFICA, 5., Joaçaba, Caderno de Resumos. Joaçaba, p. 203, 1999.
COELHO, F.; FERREIRA, J.; ZANDONAI, R. H.; NIERO, R.; CECHINEL FILHO, V.; BUENO, E. C. Proliferative activity of Ipomoea pes-caprae and Rubus imperialis methanolic extracts on lymphoblastogenesis assay using murine spleen cells stimulated with phytohemagglutinin. In: INTERNATIONAL CONGRESSO F PHARMACEUTICAL SCIENCES, 6., Ribeirão Preto, Anais. Ribeirão Preto: CIFARP, 2007.

67
SILVA, K. L; SANTOS, A. R. S.; MATTOS, P. E. O; YUNES, R. A; DELLE-MONACHE, F.; CECHINEL FILHO, V. Chemical composition and analgesic activity of Calophyllum brasiliensis. Therapie, v. 56, p. 431-434, 2001.
SILVA, M. G.; OHARA, M. T .; KATO, E. T. M.; MAIA, J. F. G.; ANDRADE, E. H. de; YOSHIDA, M. Estudo farmacognóstico de Myrciaria glomerata In: SIMPÓSIO DE PLANTAS MEDICINAIS DO BRASIL, 16., Recife. Anais. Recife, 2000. p. 172.
SMITH, K. A.; CANTRELLl, D. A. Interleukin 2 regulates its own receptors. Proc. Natl. Acad. Sci. USA., v. 82, p. 864-868, 1985.
SOUZA-FORMIGONI, M. L. O.; OLIVEIRA, M. G.; SILVEIRA FILHO, N. G.; BRAZ, S.; CARLINI, E. L. A. Antiulcerogenic effects of two Maytenus species in laboratory animals, J. Ethnophartmacol. v. 34, p.21-27, 1991.
SOUZA, M. M.; MADEIRA, A.; BERTI, C.; KROGH, R.; YUNES, R. A.; CECHINEL FILHO, V. Antinociceptive properties of the methanolic extract obtained from Ipomoea pes-caprae (L.) R. Br. J. Ethnopharmacol., v. 69, p. 85-90, 2000.
SOUZA, M. M.; SCHLEMPER, V.; JESUS, R.; CECHINEL FILHO, V. Analgesic profile of hidroalcoholic extract obtained from Marruium vulgare. Phytomedicine, v. 5, n. 2, p. 111-113, 1998.
SOUZA, M. T.; Estudo Fitoquímico e Avaliação de Atividade Biológica de Matayba elaeagnoides RADALK. Dissertação (Mestrado em Ciências Farmacêuticas)- Universidade do Vale do Itajaí, UNIVALI, Itajaí, 2006.
STARBORG, M.; GELL, K.; BRUNDELL, E.; HOOG, C. The murine Ki-67 cell proliferation antigen accumulates in the nucleolar and heterochromatic regions of intherphase cells and at the periphery of the mitotic chromosomes in a process essential for cell cycle progression. J. Cell Sci., v.109, p.143-153, 1996.
STITES, P.D.; TERR, A.I.;PARSOLW, T.C. Imunologia Médica.Rio de Janeiro: Editora Guanabara, 2000.
SWANSTON-FLATT, S. K.; DAY, C.; BAILEY, C. J.; FLATT, P. R. Traditional plant treatments for diabetes. Studies in normal and streptozotocin diabetic mice. Diabetologia, v. 33, p. 462-464, 1990.
TCHINDA, A. T.; TANE, P.; AYAFOR, J. F.; CONNOLLY, J. D. Stigmastane derivatives and isovaleryl sucrose esters from Vernonia guineensis (Asteraceae). Phytochemistry, v.63, p. 841-846, 2003.
VERROTTA, L.; APPENDINO, G.; JAKUPOVIC, J,; BOMBARDELLI, E. Hypeforine analogues from St. Jonh worth. Journal of Natural Products, v. 63, p. 412-415, 2000.
WAGNER, H. Pesquisa fitomédica no novo milênio: tendências e mudanças. In: Yunes, R. A.; CECHINEL FILHO, V. Química de produtos naturais, novos fármacos e a moderna farmacognosia. Itajaí: Editora da UNIVALI, 2007. p. 34-47.
WEISSMAN, I. Developmental switches in the imune system cell., 2.ed. New York: W. B. Saunders Company, 1994.

68
YASNI, S.; YOSHIIE K.; ODA H.; SUGANO M.; IAMIZUMI K. Dietary Curcuma xanthorrhiza Roxb. increases mitogenic responses of splenic lymphocytes in rats, and alters populations of the lymphocytes in mice. Journal of Nutricional Science and Vitaminology, v. 39, p. 345-354, 1993.
ZDZSI, B., REZESKI, W.; PADUCH, R.; SZUSTER-CIESIELSKA, A ; KACZOR, J.;WELKSZA, K.; KANDEFER-SZERSZE, M. Differential effect of betulin and betulinic acid on cytokine production in human whole blood cell cultures. Polish J. Pharmacol., v. 55, p. 235-238, 2003.
ZHU N. The glucosides from Maytenus ilicifolia. Phytochemestry, v. 47, p. 265-268, 1998.





























