Embed Size (px)
Citation preview

1
UNIVERSIDADE DE CAXIAS DO SUL CENTRO DE CIÊNCIAS AGRÁRIAS E BIOLÓGICAS
CURSO DE BACHARELADO EM CIÊNCIAS BIOLÓGICAS
FLEXIBILIDADE ALIMENTAR E COMPORTAMENTAL DO BUGIO-RUIVO Alouatta guariba clamitans CABRERA, 1940
(PRIMATES, ATELIDAE) EM RELAÇÃO AO USO DE ESPÉCIES VEGETAIS ALÓCTONES EM CAXIAS DO SUL, RS.
Juliana Nascimento Martins
Caxias do Sul
2008

2
Juliana Nascimento Martins
FLEXIBILIDADE ALIMENTAR E COMPORTAMENTAL DO BUGIO-RUIVO Alouatta guariba clamitans CABRERA, 1940 (PRIMATES, ATELIDAE) EM RELAÇÃO AO USO
DE ESPÉCIES VEGETAIS ALÓCTONES EM CAXIAS DO SUL, RS.
Monografia apresentada ao Curso de Ciências Biológicas da Universidade de Caxias do Sul, como requisito para a obtenção do grau de bacharel.
Orientador Dr. Alois Eduard Schäfer
Co-orientador Dr. Rodrigo Cambará Printes.
Caxias do Sul
2008

iii
Dedico este trabalho aos meus pais, Ubirajara e
Mara, com os quais aprendi desde cedo a não
desistir dos meus sonhos e a empenhar todas as
minhas forças na realização dos meus objetivos.

iv
Agradecimentos
Ao meu orientador Alois Shäfer, que apesar de atuar em outra linha de pesquisa,
ajudou no que estava ao seu alcance e acreditou na realização desse trabalho.
Ao meu co-orientador Rodrigo Cambará Printes, que foi meu guia nessa caminhada,
por toda paciência com meus trilhões de e-mails e pelo apoio.
Ao Valmor Barni, diretor da Fepagro Agroindústria, pela autorização e apoio para
desenvolver esta pesquisa na área, mas, também, pelo seu entusiasmo e carisma. Aos
funcionários da Fepagro Agroindústria, que de uma forma e outra me ajudaram em muitas
ocasiões.
Ao biólogo Dr. Marcos Fialho de Souza, pelas sugestões, críticas construtivas e
revisão do projeto. Ao biólogo Martin Grings e ao colega Eduardo Pasini pelo auxílio nas
identificações botânicas.
Aos meus queridos amigos, que estiveram comigo a campo, muito obrigada pela
ajuda, sugestões, discussões, incentivo e, acima de tudo, pela amizade e companheirismo:
Camila Trentin, Cassiano Marchett, Cleber Saraiva, Liziane Crippa, Maína Marchioro,
Márcio Bonesso, Otávio Ruivo, Shana Ferrarini. Aos meus outros amigos, não menos
especiais, que estiveram torcendo pelo sucesso deste trabalho: Alan Bolzan, Flaviane Magrini,
Márcia Toigo e Thais Michel.
Ao meu pai Ubirajara, que foi fundamental para este trabalho ocorrer, pois, além do
apoio e incentivo, estava sempre disposto a levar-me a campo e discutir sobre as minhas mais
recentes observações.
À minha mãe Mara, minha irmã Luciana e meu padrasto Marcelo, que sempre
apoiaram minhas decisões e vibraram comigo nas conquistas, mesmo que à distância.
Ao meu namorado, Leonardo Brito, pelo carinho e compreensão e por aceitar, mesmo
que enciumado, meu grande amor pelos bugios.

v
ANEXOS
Anexo 1: Fotos dos indivíduos de Alouatta guariba clamitans do grupo de estudo 100
Anexo 2. Fotos de algumas espécies alimentares (alóctones e autóctones)
consumidas 101
Anexo 3. Detalhes da área de estudo e acontecimentos 108

vi
ÍNDICE
ANEXOS v
LISTA DE TABELAS vii
LISTA DE FIGURAS viii
LISTA DE FIGURAS viii
Resumo x
Astract xi
1. INTRODUÇÃO 12
2. REVISÃO DE LITERATURA 14
2.1. Gênero Alouatta Lacépède, 1799 (Primates, Atelidae) 14
2.2. O bugio-ruivo Alouatta guariba clamitans Cabrera, 1940 17
2.3. Dieta 19
2.4. Alouatta em ambientes fragmentados 20
3. MATERIAIS E MÉTODOS 24
3.1. Área de estudo 24
3.2. Coleta de dados 30
3.2.1. Definição do grupo focal 30
3.2.2. Análise comportamental 31
3.2.3. Análise da dieta 33
3.2.4. Determinação da área de vida e uso do espaço 34
4. RESULTADOS E DISCUSSÃO 36
4.1. Orçamento de atividades diárias 37
4.2. Dieta 48
4.2.4. Análise qualitativa 48
4.2.5. Análise quantitativa 55
4.3. Área de vida e uso do espaço 69
5. CONCLUSÕES 86
6. REFERÊNCIAS BIBLIOGRÁFICAS 88

vii
LISTA DE TABELAS
Tabela 1. Descrição dos comportamentos presentes no etograma. ....................................................... 33
Tabela 2. Porcentagem de tempo gasto nas atividades diárias. Média mensal e média total para os meses de estudo..................................................................................................................................... 39
Tabela 3. Lista de espécies vegetais e itens alimentares utilizados na alimentação de Alouatta guariba clamitans no período de janeiro a setembro de 2008, em um remanescente de Floresta Ombrófila Mista em Caxias do Sul, RS. Itens: f = folhas, fl = flores, fr = frutos, s = sementes. Metodologia: obs. direta = observação direta dos animais, fezes = análise fecal. .............................................................. 49
Tabela 4. Meses de consumo dos itens e espécies vegetais por A. g. clamitans, em um remanescente de Floresta Ombrófila Mista em Caxias do Sul, RS no ano de 2008. verde = folhas, vermelho = flores, amarelo = frutos, marrom = sementes. .................................................................................................. 53
Tabela 5. Freqüência de consumo de espécies vegetais por Alouatta guariba clamitans no período de fevereiro a setembro de 2008, em um remanescente de Floresta Ombrófila Mista em Caxias do Sul, RS. Em ordem decrescente de freqüência dos registros de alimentação. FA = freqüência absoluta, FR =freqüência relativa, N = Numero de meses de uso (máximo 8 meses). .............................................. 56
Tabela 6. Parâmetros ecológicos (diversidade, riqueza e equidade) das espécies alimentares, analisadas por estação do ano (verão, outono e inverno de 2008).......................................................................... 62
Tabela 7. Encontros intergrupais observados durante o período de estudo. ......................................... 75
Tabela 8. Área de vida e coeficiente de variação do uso da área durante as estações de verão, outono e inverno de 2008..................................................................................................................................... 80
Tabela 9. Estimativas de tamanho de área de vida e tamanho de grupo em diversos estudos realizados com a espécie Alouatta guariba, utilizando o método do esquadrinhamento (25 x 25m). ................... 83

viii
LISTA DE FIGURAS
Figura 1: Localização da área de estudo no município de Caxias do Sul, no estado do Rio Grande do Sul e no Brasil. ...................................................................................................................................... 24
Figura 2. Média das temperaturas médias e da precipitação de Caxias do Sul dos últimos 10 anos. ... 25
Figura 3. Temperatura média máxima e mínima de Caxias do Sul de janeiro a setembro de 2008...... 25
Figura 4. Precipitação de Caxias no Sul de janeiro a setembro de 2008............................................... 25
Figura 5. Imagem da área de estudo...................................................................................................... 28
Figura 6. Vista geral da área de estudo, mostrando a mata secundária (mata 1) no centro da imagem e à direita a linha com espécies vegetais alóctones..................................................................................... 29
Figura 7. Vista geral da linha de espécies vegetais alóctones. .............................................................. 29
Figura 8. Interior da mata secundária (mata 1). .................................................................................... 29
Figura 9. Vista geral das espécies vegetais alóctones. .......................................................................... 30
Figura 10. Grupo de A. g. clamitans observado neste estudo................................................................ 31
Figura 11: Total de horas mensais de observação do grupo de estudo. ................................................ 36
Figura 12. Média das porcentagens de tempo em que o grupo de estudo alocou às diferentes atividades, durante as observações de fevereiro a setembro de 2008.................................................... 37
Figura 13. Proporção de tempo gasto nas diferentes categorias comportamentais durante as estações de verão, outono e inverno de 2008. .......................................................................................................... 40
Figura 14. Variação temporal média das principais atividades realizadas pelo grupo de bugios durante as estações de verão (4 dias), outono (7 dias) e inverno (7 dias) do ano de 2008................................. 42
Figura 15. Variação temporal da atividade descanso, durante as estações de verão (4 dias), outono (7 dias) e inverno (7 dias) do ano de 2008. ............................................................................................... 43
Figura 16. Variação temporal da atividade alimentação, durante as estações de verão, outono e inverno de 2008. ................................................................................................................................................. 45
Figura 17. Freqüência de consumo dos itens alimentares nas estações de verão, outono e inverno de 2008....................................................................................................................................................... 58
Figura 18. Número de espécies consumidas pelo grupo de A. g. clamitans em dias completos (mín. 8 horas), nos meses de estudo em 2008. .................................................................................................. 59
Figura 19. Porcentagem de consumo das diferentes espécies vegetais pelo grupo de estudo nas estações de verão (a), outono (b) e inverno (c) de 2008........................................................................ 61
Figura 20. Contribuição das espécies vegetais autóctones e alóctones na dieta do grupo de estudo, nas estações de verão, outono e inverno de 2008. ....................................................................................... 63

ix
Figura 21. Contribuição das espécies vegetais alóctones na dieta do grupo de estudo, nas estações de verão (a), outono (b) e inverno (c) de 2008........................................................................................... 65
Figura 22. Uso cumulativo de quadrados pelo grupo de bugios, durante os dias amostrados por varredura instantânea, em Caxias do Sul, RS........................................................................................ 69
Figura 23. Imagem da área de estudo com a grade de quadrículas (25x25m). ..................................... 70
Figura 24. Uso do espaço pelo grupo de bugios antes do corte dos Pinus (a) e depois do corte (b), de janeiro a setembro de 2008.................................................................................................................... 71
Figura 25. Áreas nucleares de utilização (a) e sítios de pernoite (b) utilizados pelo grupo de bugios, de fevereiro a setembro de 2008. ............................................................................................................... 73
Figura 26. Uso de espécies vegetais autóctones e alóctones como sítio de pernoite pelo grupo focal nos meses de observação do ano de 2008.................................................................................................... 74
Figura 27. Uso do espaço no verão, outono e inverno de 2008............................................................. 78

x
RESUMO
A fragmentação dos hábitats e os efeitos de borda decorrentes, bem como a introdução de espécies alóctones alteram os ambientes florestais, exigindo da fauna adaptações e estratégias de sobrevivência. Desta forma, a flexibilidade alimentar e comportamental de um grupo de três bugios (Alouatta guariba clamitans Cabrera, 1940) em relação às espécies vegetais alóctones foi estudada em um fragmento de mata perturbada em Caxias do Sul, Rio Grande do Sul, Brasil. O fragmento de estudo é uma mata secundária inserida no Bioma Floresta Ombrófila Mista, com alta regeneração de espécies alóctones. A matriz da paisagem é atividade agrícola. Este fragmento é conectado a uma linha de divisa de propriedades plantada com as espécies arbóreas alóctones Eucalyptus sp., Pinus spp., Hovenia dulcis, Quercus robur e Carya illinoensis. Aspectos do comportamento, dieta, área de vida e uso do espaço foram analisados durante oito meses, pelo método de varredura instantânea (quatro unidades amostrais/h) e esquadrinhamento com o auxílio de bússola e trena (25 x 25m). O grupo dedicou 73% do tempo ao descanso, 11% à locomoção, 12% à alimentação e 3% à atividades sociais. Os bugios consumiram 34 espécies vegetais, sendo nove alóctones. Três espécies foram identificadas exclusivamente pela análise de fezes. As folhas de lianas contribuíram com 24% da dieta, sendo 12,3% desta composta por folhas e frutos de H. dulcis, 8,6% por folhas e sementes de Araucaria angustifolia, 6,64% por flores de Pyrostegia venusta. Frutos e folhas de Eucalyptus sp. e Hovenia dulcis foram consumidos durante o período de observação, exceto em junho e janeiro, respectivamente. Outras espécies alóctones utilizadas foram: folhas de Pinus spp., frutos de Eryobotrya japonica, folhas e frutos de Q. robur, folhas e fruto de Ligustrum lucidum, frutos de Vitis americana e frutos de Diospyrus kaki. Dentre os itens alimentares consumidos, 64% foram folhas, 25% frutos, 8,7% flores e 1,7% sementes. Ao todo, 63% da dieta do grupo foi composta por espécies autóctones e 37% alóctones. A área de vida total do grupo foi 3,25 ha, sendo usados 0,73 ha no verão, 0,92 ha no outono e 1,04 ha no inverno. Quando uma parte da área de vida do grupo (a plantação de Pinus sp.) foi suprimida, os animais passaram a utilizar mais a área de mata, aumentando sua área de uso de 1,8 ha para 2,5 ha. O grupo de estudo demonstrou alta flexibilidade alimentar e comportamental através da incorporação de espécies alóctones à dieta, aumento o tempo dedicado ao descanso e ajuste da área de vida ao território do grupo vizinho. Além disso, o grupo focal foi capaz de se ajustar à perda de parte de sua área de vida, causada pelo ser humano.
Palavras-chave: Alouatta guariba clamitans; comportamento alimentar; flexibilidade; espécies alóctones; área de vida.

xi
ABSTRACT
The habitat fragmentation and the decurrent edge effects in addition to exotic species introduction, modify the forest habitat demanding adaptations and strategies to wildlife survival. Thus, the alimentary and behavioral flexibility of a three howler monkey group (Alouatta guariba clamitans Cabrera, 1940) in relation to exotic tree species was studied in a disturbed fragment forest at Caxias do Sul, Rio Grande do Sul, Brazil. The fragment is a secondary formation in the atlantic rain forest, with a high level of exotic species occupation. The landscape matrix is agriculture. The fragment is connected to a narrow line of exotic tree Eucalyptus sp., Pinus spp., Hovenia dulcis, Quercus robur and Carya illinoensis that divides two properties. Aspects of behavior, diet, home range and space use were investigated during eight months, through method of “scan sampling” (four units sampling/hour) and grid cell method using a compass and a tape measure (25 x 25m). The group dedicated 73% of time to resting, 11% to moving, 12% to feeding and 3% in social activities. The howlers consumed 34 plant species, nine exotic species and three species exclusively registered in feces content analysis. The leaves of lianas were 24% of the howler’s diet, H. dulcis leaves and fruits were 12.3%; Araucaria angustifolia leaves and seed were 8.6% and Pyrostegia venusta flowers werw 6.64%. Fruits and leaves of Eucalyptus sp. and H. dulcis were consumed in all months, except in June and January. Others exotic species consumed were: Pinus spp (leaves)., Eryobotrya japonica (fruits), Q. robur (leaves and fruits), Ligustrum lucidum (leaves and fruits), Vitis americana (fruits) and Diospyrus kaki (fruits). In relation to the diet compounds, 64% was leaves, 25% fruits, 8.7% flowers and 1.7% seed. The native species were 63% of diet against 37% exotic. The home range was 3.25 ha, with 0.73 ha in the summer, 0.92 ha in the autumn and 1.04 ha in the winter. When a part o home range with Pinus spp. was cutted away the animals change its use patterns. The area was from 1.8 to 2.5 ha and the upgrade was due to native forest incorporated. The howler’s group presents a dietary and ethological flexibility, using alien species to feed, enlargement the resting time, fitting its home range to the other group and supporting the cut of its home range.
Key-words: Alouatta guariba clamitans; alimentary behaviour; flexibility; exotic species; home range.

12
1. INTRODUÇÃO
A silvicultura com as espécies alóctones dos gêneros Pinus e Eucalyptus é uma
atividade econômica comum e crescente no estado do Rio Grande do Sul, e é muito
questionada quanto aos impactos que traz aos ecossistemas. Com a utilização das terras para
silvicultura e agricultura, as atividades humanas têm causado diversas mudanças na paisagem,
diminuindo a quantidade e qualidade de hábitats disponíveis para muitas espécies.
Além disso, fragmentação de hábitat e o efeito de borda decorrente, a perda da
paisagem original e a introdução de espécies alóctones alteram os ambientes florestais,
exigindo da fauna adaptações e estratégias de sobrevivência. Tais alterações podem ter efeitos
sobre o padrão de movimento e comportamento dos animais dentro do seu ambiente natural,
efeitos estes ainda não bem compreendidos.
Várias espécies de primatas do gênero Alouatta (Primates, Atelidae) conhecidos como
bugio, guariba, barbado, macaco-uivador, têm sido amplamente estudadas no Brasil,
Argentina e América Central em seu hábitat natural. Entretanto, diante de tantas mudanças
nos ambientes destes primatas, se torna necessário conhecer as estratégias comportamentais e
alimentares que estes animais desenvolveram para persistir em ambientes alterados. Tais
estudos são importantes, frente à necessidade de conservação e manejo das populações,
principalmente fora de unidades de conservação.
Pesquisas vêm reportando que Alouatta é tolerante à fragmentação do hábitat e se sabe
que o bugio-ruivo Alouatta guariba clamitans Cabrera, 1940 pode viver em pequenos
fragmentos de mata e em locais com alta pressão antrópica, como ocorre com os bugios que
habitam a zona rural de Porto Alegre. Entretanto, ainda há poucos estudos destes animais nos
demais municípios do Estado e, principalmente, em áreas alteradas, nas quais foram
introduzidas espécies arbóreas alóctones ou exóticas.

13
Dados sobre a influência que as espécies alóctones exercem sobre a dieta, os
orçamentos diários de atividades e área de vida destes animais são importantes para a
compreensão do quão tolerante estes primatas são às alterações do hábitat e podem ter
implicações diretas para o manejo da espécie.
Além da carência de estudos sobre as adaptações do A. g. clamitans às crescentes
alterações ambientais, o bugio-ruivo está citado na lista de espécies vulneráveis à extinção do
Estado do Rio Grande do Sul, havendo poucas pesquisas da espécie em Floresta Ombrófila
Mista.
Desta forma, o objetivo deste trabalho é avaliar a flexibilidade alimentar e
comportamental de um grupo de bugio-ruivo Alouatta guariba clamitans, em relação à
utilização de espécies vegetais alóctones para alimentação e descanso, num fragmento de
mata perturbada no município de Caxias do Sul, Rio Grande do Sul, Brasil.

14
2. REVISÃO DE LITERATURA
2.1. Gênero Alouatta Lacépède, 1799 (Primates, Atelidae)
Entre as espécies de primatas ameaçadas de extinção, encontram-se os animais do
gênero Alouatta Lacépède, 1799, conhecidos vulgarmente como bugios, roncadores, guaribas
ou barbados. Pertencente à família Atelidae, o gênero atualmente é composto por nove
espécies, das quais seis ocorrem no Brasil (Rylands et al., 2000): A. seniculus (Linnaeus,
1766), A. belzebul (Linnaeus, 1766), A. guariba (Humboldt, 1812), A. caraya (Humboldt,
1812), Alouatta nigerrima Lonnberg, 1941; Alouatta sara Elliot, 1910.
A grande amplitude de distribuição geográfica do gênero, a maior dentre os primatas
neotropicais, desde o Estado de Vera Cruz no México até o Estado do Rio Grande do Sul no
Brasil, passando pela Província de Corrientes na Argentina é decorrente da ocupação de
hábitats de características tão distintas quanto florestas tropicais de terra firme, florestas
inundadas, florestas decíduas, florestas de galeria, cerrado, floresta com araucárias, florestas
de altitude, além de florestas secundárias, fragmentadas e alteradas (Neville et al., 1988;
Printes et al., 2001; Bicca-Marques, 2003).
O Alouatta guariba (Humboldt, 1812) é endêmico da Mata Atlântica e pode ser
encontrado em florestas costeiras do sudeste do Brasil, desde o Estado da Bahia até o Rio
Grande do Sul e em Missiones na Argentina. Esse território é subdividido pela ocupação de
duas subespécies: encontrando-se o A. guariba guariba (Humboldt, 1812) na Bahia até o
Espírito Santo e norte do Rio de Janeiro, além de algumas áreas de Minas Gerais. A outra
subespécie, Alouatta guariba clamitans Cabrera, 1940, ocorre desde o Espírito Santo até a
bacia do rio Camaquã no Rio Grande do Sul e no nordeste da Argentina (Hirsch et al., 1991;
Rylands et al., 2000; Printes et al., 2001).

15
No Estado do Rio Grande do Sul, ocorrem A. guariba clamitans (bugio-ruivo) e A.
caraya (bugio-preto), além de Cebus nigritus nigritus (Marques, 2003). A. caraya ocorre nos
enclaves florestais do bioma pampa, A. g. clamitans ocupa a floresta ombrófila densa, a
floresta ombrófila mista e a floresta estacional semi-decídua, enquanto Cebus nigritus tende a
acompanhar a distribuição geográfica da floresta ombrófila mista densa e as florestas do Alto
Uruguai.
Alouatta possui várias adaptações diferentes dos outros gêneros da família, como
corpo maciço com longa pelagem (7 cm) que se avoluma na mandíbula e lados da face,
formando grande barba. Sob a barba se oculta o osso hióide que se transformou em caixa de
ressonância para sua potente voz, sendo maior o osso nos machos, chegando a possuir um
volume interno de 50 cm2. Devido ao aparato hióidico, a vocalização é muito desenvolvida e
característica, sendo o “rugido” ouvido a quilômetros de distância e emitido em encontros
intergrupais (Carpenter, 1934; Chivers, 1969 apud Auricchio, 1995) e na manutenção do
espaçamento entre os grupos (Carpenter, 1934; Altmann, 1959).
As espécies do gênero Alouatta são de hábito diurno e geralmente vivem em grupos
coesos multi-machos e fêmeas, com 3 a 13 animais (Mendes, 1989, Marques, 2001), onde
normalmente contém de 1 a 4 machos adultos com uma proporção sexual de 1 macho: 2-4
fêmeas (Sussman, 2000). São animais arborícolas, preferindo os estratos médio e superior da
floresta, onde permanecem em repouso a maior parte do dia (Carpenter, 1934, Milton, 1980,
Mendes, 1985).
Alouatta consome uma alta proporção de folhas na dieta (Milton, 1980; Chapman,
1987; Neville et al., 1988; Bonvicino, 1989; Mendes, 1989; Chiarello, 1994; Bicca-Marques
& Calegaro-Marques, 1994; Valle et al., 2001; Fuentes et al., 2003; Aguiar et al., 2003;
Miranda & Passos, 2004) sendo este considerado o mais folívoro entre os primatas

16
neotropicais (Carpenter, 1934; Eisenberg et al., 1972; Jolly, 1985 apud Jardim 2005; Queiroz,
1995).
No entanto, foi observado um consumo maior de frutos em períodos ou locais de
maior disponibilidade deste recurso (Altmann, 1959; Hladik & Hladik, 1969; Milton, 1980;
Bonvicino, 1989; Queiroz, 1995; Jardim, 1997; Estrada et al., 1999; Fialho, 2000), sendo
considerada uma espécie bastante oportunista (Chiarello, 1992), consumindo também flores,
pecíolos, brotos, sementes, caules e ramos. Em função disto, os bugios são melhor
caracterizados como folívoro-frugívoros (Crockett & Eisenberg, 1987 apud Jardim, 2005).
As folhas não constituem boas fontes energéticas (Milton, 1980), além de serem ricas
em compostos de difícil digestão, alcalóides, celulose, lignina, taninos, dentre outros (Garber,
1987). Insetos são eventualmente consumidos por Alouatta (Milton, 1980; Chitolina &
Sander, 1981; Mendes, 1989).
O gênero possui algumas adaptações morfológicas associadas à folivoria, como o
alargamento do ceco e cólon e fermentação pós-gástrica (Queiroz, 1995). Por estes animais
não apresentarem adaptações morfo-fisiológicas tão acentuadas quanto os colobíneos, são
mais apropriadamente denominados folívoros comportamentais (Milton, 1978), sendo a pouca
energia obtida pelo alimento compensada por estratégias comportamentais de conservação de
energia (Milton, 1979).
Algumas estratégias usadas são a alta seletividade na alimentação, preferência por
folhas novas, limitação no percurso diário (Milton, 1978) e grande proporção do tempo do dia
utilizado em descanso (mais de 50% durante o dia) (Glander, 1975; Smith, 1977; Milton,
1978; Nagy & Milton, 1979; Milton et al., 1980; Chiarello, 1994). Glander (1975) acredita
que o maior tempo dedicado ao descanso é necessário para a desintoxicação dos compostos

17
secundários resultantes de uma dieta mais seletiva, capacitando-os a sobreviver com uma
dieta relativamente de baixa qualidade (Milton, 1978).
Os bugios respondem à dispersão espacial e temporal de fontes de alimentos
(principalmente frutos) através da movimentação de distâncias variadas, para encontrar o
indivíduo de uma espécie freqüentemente utilizada durante o ano (Estrada e Coates-Estrada,
1986; Limeira, 2000; Lunardelli, 2000). Entre as espécies de Alouatta, diferentes padrões são
observados em resposta a períodos de menor disponibilidade de recursos, desde reduções no
tamanho da área de uso e no comprimento dos deslocamentos para minimizar gastos
energéticos (Milton, 1980; Mendes, 1989; Queiroz, 1995; Marques, 1996) a expansões da
área e dos percursos em busca dos recursos alimentares importantes (Bonvicino, 1989;
Limeira, 1996).
O tamanho da área de vida de primatas é correlacionado positivamente com a massa
corporal, tamanho do grupo e tamanho do fragmento florestal e de forma negativa à proporção
de folhas na dieta e à densidade populacional (Milton & May, 1976; Clutton-Brock & Harvey,
1977; Crockett & Eisenberg, 1987; Steinmetz, 2001; Bicca-Marques, 2003).
É sugerido que em casos de alta densidade os grupos tenham duas opções: ou
restringem o tamanho da área de vida ou aumentam a taxa de sobreposição de área com
grupos adjacentes (Jardim, 2005). O tamanho das áreas de uso ocupadas pelos grupos é
bastante variável entre as espécies, desde 0,3 ha observado para A. caraya (Bicca-Marques,
1990) a 182 ha para A. seniculus (Palacios & Rodriguez, 2001).
2.2. O bugio-ruivo Alouatta guariba clamitans Cabrera, 1940
Alouatta guariba clamitans Cabrera, 1940, são primatas neotropicais endêmicos da
Floresta Atlântica lato sensu ocupando várias fisionomias florestais como a Floresta Atlântica

18
sensu strictu, a Floresta com Araucária e a Floresta Estacional Semidecidual (Ihering, 1914;
Cabrera, 1957; Hirsch et al., 1991; Chiarello & Galetti, 1994; Auricchio, 1995).
A despeito de sua ampla distribuição geográfica a espécie é enquadrada na categoria
vulnerável nas listas oficiais de fauna ameaçada dos estados de Minas Gerais (Machado et al.,
1998), São Paulo (São Paulo, 1998), Paraná (Tossulino et al., 1995) e Rio Grande do Sul
(Marques, 2003). Em escala global aparece listada na categoria "quase ameaçada" (Near
threatened-NT) (IUCN, 2007). As principais ameaças são a acelerada destruição da Mata
Atlântica, a caça e o comércio ilegal.
Gregorin (2006) realizou uma revisão taxonômica para o gênero Alouatta, variação
geográfica e não-geográfica dos diferentes táxons de bugios que ocorrem no Brasil, propondo
a denominação Alouatta clamitans. Entretanto, no presente trabalho optou-se por se referir à
subspécie, visto que ainda há contradições por parte dos especialistas.
Em A. g. clamitans há um acentuado dicromatismo sexual na coloração da pelagem,
onde as fêmeas adultas e os indivíduos imaturos são geralmente castanhos escuros e os
machos adultos apresentam-se ruivos, vermelhos ou alaranjados em devido a liberação de um
pigmento por glândulas apócrinas (Ihering, 1914; Hirano et al., 2003). As fêmeas pesam cerca
de 4,1 a 5Kg e os machos em torno de 5,3 a 7,2Kg (Rowe, 1996).
São encontrados em florestas em regeneração e áreas próximas a centros urbanos,
sendo em alguns locais a única espécie de primata presente. Esta ampla variedade de hábitats
reflete a sua capacidade de adaptação a diferentes condições ecológicas. Esta flexibilização
comportamental é associada à base de alimentação folívora, item menos sazonal, à habilidade
de mudanças nas estratégias de forrageamento, ao uso diferencial da área de vida e à
capacidade de viver com grupos de tamanho reduzido (Schwarzkopf & Rylands, 1989; Gilbert
& Setz, 2001; Bicca-Marques, 2003).

19
Estudos constataram que os bugios-ruivos são seletivos em sua dieta, apresentam
baixa freqüência de interações sociais e orçamentos temporais de atividades diárias que se
mantém relativamente estáveis independente das variações entre os hábitats (Chitolina &
Sander, 1981; Prates et al., 1990 a,b; Jardim, 1992; Cunha, 1994; Marques, 1996, 2001;
Fortes, 1999; Oliveira, 2000; Sammarco, 2000; Fialho, 2000; Buss, 2001; Jerusalinsky, 2001;
Silveira & Codennotti, 2001; Liesenfeld, 2003; Koch, 2004; Jardim, 2005). Os principais
ajustes se referem às adaptações ao clima, como posturas corporais e adequação dos horários
das atividades em relação ao fotoperíodo e temperaturas extremas (Jardim & Oliveira, 2000;
Marques, 2001) e às características do ambiente como abundância de recursos alimentares,
densidade populacional e tamanho do fragmento (Bicca-Marques & Calegaro-Marques,
1994).
As estratégias de forrageamento e a composição da dieta parecem ser flexibilizadas de
acordo com a disponibilidade e disposição espacial dos itens alimentares, variando o grau de
folivoria e a forma de utilização do espaço (Chiarello & Galetti, 1994; Juan et al., 2000;
Zunino et al., 2001; Bicca-Marques, 2003; Bravo & Sallenave, 2003). Os grupos possuem de
dois a 13 indivíduos, com áreas de uso que variam de 2,4 ha (Jardim, 2005) a 41 ha
(Steimmetz, 2000). Normalmente forrageiam e descansam juntos, mas ocasionalmente
formam subgrupos para estas atividades.
2.3. Dieta
A dieta dos bugios é baseada em itens fáceis de encontrar e consumir, porém dificeis
de ingerir (Chiarello, 1994). Geralmente poucas espécies vegetais constituem a base da dieta
dos bugios, enquanto que um grande número de espécies é consumido em proporções muito
reduzidas (Hladik & Hladik, 1969; Smith, 1977; Glander, 1978; Milton, 1980; Chapman,
1988), e esta diversidade está associada a uma forma de balanceamento para satisfazerem suas
necessidades metabólicas (Milton, 1979, Milton et al., 1981; Nagy & Milton, 1979).

20
Pesquisas realizadas sobre o teor nutritivo e tóxico da dieta mostraram que folhas são
um ítem rico em proteína, principalmente as folhas jovens, e os frutos contêm alto teor de
energia prontamente assimilável (Smith, 1977; Milton, 1980 apud Martins, 1997). Desta
forma, a seletividade também ocorre em termos de itens. Há evidências claras de que os
bugios preferem os itens sazonais (folhas jovens, flores e frutos) aos perenes, como por
exemplo, as folhas maduras (Smith, 1977; Glander, 1978; Milton, 1980). Esta preferência se
justifica pelo melhor valor nutritivo dos itens sazonais (Milton, 1979; Glander, 1978).
Em estudos realizados em Mata de Araucária no Paraná e no Rio Grande do Sul, A. g.
clamitans utilizou cerca de 30 espécies para alimentação. As famílias mais consumidas foram:
Lauraceae, Flacourtiaceae, Myrtaceae, Sapindaceae e Compositae. As espécies que mostraram
maior importância na dieta do bugio-ruivo foram: Araucaria angustifolia, Gochnatia
polymorpha, Matayba elaeagnoides, Mimosa scabrella, Ocotea porosa, O. odorifera, O.
puberulla, O. pulchella e Zanthoxylum rhoifolium (Jardim & Oliveira, 2000; Miranda &
Passos, 2004).
Os bugios são conhecidos por apresentarem uma dieta altamente eclética e adaptável
às condiçoes ambientais (Bicca-Marques & Calegaro-Marques, 1994; Crockett, 1988 apud
Bicca-Marques et al., 2006), podendo incluir espécies exóticas na dieta (Bicca-Marques &
Calegaro-Marques, 1994; Miranda, 2004; Prates, 2007).
2.4. Alouatta em ambientes fragmentados
A fragmentação de hábitats é uma das maiores ameaças à diversidade biológica. Um
dos maiores desafios da biologia da conservação atualmente é o estudo detalhado da
fragmentação e dos efeitos da mesma sobre a biodiversidade (Soulé & Kohn, 1989 apud
Martins, 1997).

21
Extensas áreas de florestas tropicais vêm sofrendo ações antrópicas em um acelerado
processo de destruição. Em muitos casos são formados fragmentos imersos em áreas agora
ocupadas por agricultura, pastagem ou outras atividades de cunho econômico (Chiarello,
2000; Seoane et al., 2000 apud Carvalho, 2007).
A principal causa do declínio das populações de bugios é a fragmentação de hábitats
(Mittermeier et al., 1982; Mittermeier et al., 1989 apud Miranda, 2004.) Por apresentar um
hábito arborícola, os bugios encontram uma certa dificuldade para atravessar, pelo solo,
médias e grandes distâncias que separam fragmentos florestais. Este fato pode desencadear
dois problemas principais para a conservação da espécie: impossibilidade de recolonizar
fragmentos e ausência de fluxo gênico entre populações (Chiarello & Galetti, 1994;
Steinmetz, 2001)
Desta forma, os primatas são espécies indicadoras importantes, devido a seus hábitat e
dieta especializados e, por isso, podem ser usados como ferramentas para análise da extensão
dos distúrbios ecológicos, principalmente da fragmentação ambiental (Strier, 2000).
Entretanto, na ausência de uma forte pressão de caça, as espécies de Alouatta podem existir
em fragmentos de mata, hábitats perturbados e próximos a populações humanas (Crockett,
1998).
Segundo Bicca-Marques (2003) a fragmentação resulta na diminuição da área de vida,
especialmente em pequenos fragmentos onde os animais são incapazes de aumentar sua área
de atividade devido aos limites da floresta. Spironello (2001) considera a área de vida um
fator determinante no efeito da fragmentação em primatas, que possibilita as análises de
requerimentos de áreas para futuras iniciativas de conservação em fragmentos florestais.
Bicca-Marques (op. cit.), a partir de uma revisão do gênero em ambientes
fragmentados, apresentou dados referentes à grande adaptabilidade destes animais em viver

22
nesses ambientes, sendo estratégicos para sua adaptação a dieta eclética, que pode incluir
espécies exóticas, e a folivoria, as quais permitem que grupos sociais de bugios sobrevivam
em fragmentos com dimensões muito reduzidas.
Miranda & Passos (2004) observaram o consumo de frutas e plantas exóticas ao
ambiente de Floresta Ombrófila Mista por A. g. clamitans, tais como Eryobothria japonica
(Thunb.) Lindl. (ameixa-amarela) e Diospyrus kaki L.F. (caqui), mostrando uma boa
capacidade deste primata em se adaptar a ambientes moderadamente alterados.
Um grupo de bugios-preto (A. caraya) estudado em Alegrete, Rio Grande do Sul,
utilizou como fonte de alimento nove espécies vegetais nativas e cinco espécies exóticas,
sendo Citrus cinensis a principal fonte de frutos (25,6% da dieta) (Prates, 2007).
Algumas populações do gênero Cebus (macaco-prego) conseguem utilizar-se de fontes
alimentares provenientes do meio antrópico, tal como pomares e monoculturas, dentre estas:
milho, cana-de-açúcar, mandioca e Pinus (Galetti & Pedroni, 1994; Koehler & Firkowski,
1996; Rocha, 2000; Siemers, 2000; Defler, 2004; Rocha-Mendes et al., 2005; Ludwig et al.,
2005; Ludwig et al., 2006; Carvalho, 2007). Estudos com grupos de Callithrix penicillata no
Cerrado, também relatam a inclusão de frutos de espécies exóticas na dieta dos animais
(Miranda, 1997; Vilela & Faria, 2002).
Todavia, a despeito de sua capacidade de sobreviver em fragmentos bastante
reduzidos, os bugios são mais vulneráveis nestas áreas à caça, doenças, predação, escassez de
recursos alimentares e depressão por endocruzamento. Em hábitats fragmentados as
populações de A. pigra existem em densidades particularmente altas e o número de machos
adultos nos grupos é significativamente mais baixo que em populações em hábitat contínuo
(Van Belle & Estrada, 2005).

23
A sobrevivência de primatas em florestas é determinada por variáveis complexas
como composição florística, níveis de conservação ou grau de perturbação, incluindo o
tamanho dos fragmentos, isolamento e tamanho das áreas de vida das espécies, entre outras.
Desta forma, informações precisas sobre a qualidade, composição florística e a estrutura
destas florestas são importantes para compreender a biologia dos primatas que ali vivem,
assim como subsidiar estratégias para a sua conservação, tais como programas de translocação
e reintrodução de espécies (Lemos de Sá & Strier, 1992). Recentemente, foram realizadas
translocações de A. caraya com sucesso no Brasil, devido ao melhor entendimento da biologia
da espécie (Printes & Malta, 2007; Perin, 2008).
Estudos realizados em paisagens heterogêneas contendo agroecossistemas arbóreos e
fragmentos de vegetação nativa no México, Guatemala e Costa Rica demonstraram que cinco
espécies de primatas podem residir temporária ou permanentemente em um conjunto de 15
agroecossistemas (Estrada, 2007). A presença e as atividades de alimentação dos primatas
podem beneficiar estes ambientes através da aceleração da produtividade primária, da
facilitação da regeneração natural de suas fontes e frutos e da adição de importantes
quantidades de nutrientes no solo pela dispersão de suas fezes (Estrada, op. cit.).
Na Costa Rica e Nicarágua, A. palliata tem sido encontrado em plantações de café
(McCann et al., 2003; Somarriba et al., 2004), e em Los Tuxtlas, México, A. palliata e Ateles
geoffroyi exploram plantações de cacau e café (Estrada & Coates-Estrada, 1996).
Desta forma, a realização de estudos em diferentes hábitats é a única forma de se
compreender e delimitar a amplitude da variabilidade adaptativa de uma espécie (Estrada,
1984). Apesar de A. g. clamitans ser relativamente bem estudado (Bicca-Marques, 2003),
dados sobre sua ecologia em ambiente de Floresta Ombrófila Mista (Miranda, 2004) e em
ambientes alterados ainda são escassos.

24
3. MATERIAIS E MÉTODOS
3.1. Área de estudo
O presente estudo foi desenvolvido na área da FEPAGRO AGROINDÚSTRIA -
Centro de Pesquisa em Agropecuária da Fundação Estadual de Pesquisa Agropecuária, no
distrito de Fazenda Souza, à 25 km do centro de Caxias do Sul, localizada na Encosta
Superior do Nordeste do Estado do Rio Grande do Sul, nas coordenadas UTM SAD69 (22 J)
501100/ 6778400 (Figuras 1 e 2).
O clima da região, segundo a classificação de Köppen-Geiger, é do tipo "Cfb"
(mesotérmico sempre úmido com verões brandos) e apresenta inverno com geadas freqüentes.
A temperatura do mês mais quente inferior a 22°C e a do mês mais frio superior a 3°C.
Apresenta quatro estações bem definidas, com chuvas durante todos os meses do ano, tendo
uma precipitação anual média de 1.915mm. As precipitações são regularmente distribuídas
durante o ano. A altitude da área de estudo é de aproximadamente 760 m.
Brasil
RS
Caxias do Sul
FEPAGRO
Figura 1: Localização da área de estudo no município de Caxias do Sul, no estado do Rio
Grande do Sul e no Brasil.

25
Caxias do Sul (RS)
-102030405060708090
100110120
jan fev mar abr mai jun jul ago set out nov dez
T (°C)
020406080100120140160180200220240
P (mm)
Precipitação Temperatura média
Figura 2. Média das temperaturas médias e da precipitação de Caxias do Sul dos últimos 10 anos.
0
5
10
15
20
25
30
Jan Fev Mar Abr Mai Jun Jul Ago Set
Meses
Tem
pera
tura
(C)
temperatura média máxima temperatura média mínima
Figura 3. Temperatura média máxima e mínima de Caxias do Sul de janeiro a setembro de 2008.
020406080
100120140160180
Jan Fev Mar Abr Mai Jun Jul Ago Set
Meses
Prec
ipita
ção
(mm
)
Figura 4. Precipitação de Caxias no Sul de janeiro a setembro de 2008.

26
A vegetação é Floresta Ombrófila Mista e o fragmento de estudo (mata 1) é uma mata
secundária em estágio avançado de regeneração, com 10,21 ha (Figuras 5 e 8), com
regeneração no sub-bosque e grande presença de lianas.
Algumas espécies florestais presentes na área são: Allophylus edulis, Allophilus
guaraniticus, Araucaria angustifolia, Casearia decandra, Cupania vernalis, Dasyphyllum
tomentosum, Ilex paraguaensis, Lithraea brasiliensis, Matayba elaeagnoides, Mimosa
scabrela, Myrciaria delicatula, Nectandra sp., Ocotea spp., Parapiptadenia rigida,
Piptocarpha angustifolia, Rapanea ferruginea, Roupala brasiliensis, Syagrus romanzoffiana,
Vitex megapotamica, Zanthoxylum rhoinfolium, além da presença de espécies alóctones como
Eriobotrya japonica, Ligustrum lucidum e Hovenia dulcis. A matriz da paisagem se
caracteriza pela presença de solo modificado pela atividade agrícola, com plantios de maçã,
pêra, ameixa, uva, caqui, kiwi, entre outros.
No interior da mata é freqüente o ruído originado pelos habitantes do entorno e de seus
animais domésticos, como cães e galinhas, bem como tratores e veículos que passam nas
estradas dentro e no entorno da propriedade.
O fragmento de mata é conectado à nordeste a um fragmento maior de 16,7 ha (mata
2). Conectado ao fragmento menor há uma linha de divisa de propriedades (Figuras 5 e 7) de
1,4 ha plantada com espécies alóctones, tais como Eucalyptus sp., Pinus taeda, Pinus elliottii,
Hovenia dulcis, Quercus robur e Carya illinoensis.
Na mata 1, é freqüente a observação de indivíduos de Dasyprocta azarae e de Sciurus
ingrami. Foram observados também no local Coendou prehensilis, Lepus europaeus, rastros
de Procyon cancrivorus e Cerdocyon thous. Na avifauna, destacam-se algumas espécies mais
comumente avistadas: Colaptes campestris, Cyanocorax caeruleus, Leptotila verreauxi,

27
Penelope obscura, Piaya cayana, Picumnus spp., Ramphastos dicollorus, Rupornis
magnirostris, Theristicus caudatus, Trogon sp. e dendrocolaptídeos.
Foram avistados bugios em ambas as áreas de mata, possuindo três grupos na mata 1.
O grupo estudado neste trabalho foi observado utilizando a mata secundária (mata 1) e a área
plantada com as espécies vegetais alóctones, sendo mais comumente avistado no segundo
local (Figura 5), durante o início do estudo.

28
Linha de espécies alóctones: 1,4 ha
Mata 1: 10,21ha Mata 2: 16,7 ha
Pomares
Sede da FEPAGRO
Figura 5. Imagem da área de estudo.

29
Figura 6. Vista geral da área de estudo, mostrando a mata secundária (mata 1) no centro da
imagem e à direita a linha com espécies vegetais alóctones.
Figura 7. Vista geral da linha de espécies vegetais alóctones.
Figura 8. Interior da mata secundária (mata 1).

30
Figura 9. Vista geral das espécies vegetais alóctones.
3.2. Coleta de dados
3.2.1. Definição do grupo focal
O reconhecimento e a seleção do grupo (Figura 10) foram realizados no período de
janeiro a fevereiro de 2008, considerado como etapa piloto.
O grupo foi definido e identificado conforme a composição sexo-etária dos indivíduos
(Mendes, 1989) e sua localização, sendo este composto por três indivíduos: juvenil II macho
(indivíduo completamente independente e de tamanho maior que infantes e menor que fêmeas
adultas), fêmea adulta (coloração escura, variando de preto ao castanho avermelhado,
genitália descoberta de forma triangular ou levemente arredondada quando vista de um ângulo
ventro-posterior) e um macho adulto (nitidamente maior que fêmeas adultas, barba longa,
pelagem bem densa, cuja coloração que pode variar do vermelho acastanhado, passando pelo
ruivo até o alaranjado, e com o saco escrotal bem desenvolvido e à mostra).

31
Figura 10. Grupo de A. g. clamitans observado neste estudo.
As saídas para coleta de dados foram realizadas mensalmente, exceto no mês de junho,
devido à chuva. O grupo foi acompanhado durante quatro dias na estação de verão (fevereiro
e março), sete dias no outono (abril, maio, junho) e sete dias no inverno (agosto e setembro),
durante o ano de 2008. As observações do grupo ocorreram desde o inicio da atividade dos
animais - em torno das 7:00 - até o anoitecer – após as 18:00 - somando em média 11h/dia.
3.2.2. Análise comportamental
A habituação do grupo foi realizada durante os meses de janeiro e fevereiro de 2008
(Williamson & Feistner, 2003), registrando-se os principais comportamentos através da
amostragem ad libitum.
O método de registro adotado para a análise comportamental foi o de varredura
instantânea (“scan sampling method”) (Altmann, 1959; Altmann, 1974; Setz & De Hoyos,
1985; Cullen Jr. & Valladares-Padua, 1997; Fortes & Bicca-Marques, 2005), com cinco
minutos de período amostral (PA) e dez minutos de período de intervalo (PI) (= quatro

32
unidades amostrais/hora). As observações foram realizadas com o auxílio de um binóculo
10x50mm. Em dias chuvosos não foram realizadas observações devido a dificuldade de
acompanhamento dos animais e visualização.
A forma de anotação dos registros foi pontual, com amostragem não instantânea (Setz,
1991), que consiste no registro da primeira atividade de cada individuo, com duração de pelo
menos 5 s. Em cada registro foi anotado o horário, atividade desempenhada pelos animais e a
espécie utilizada.
Os dados sobre comportamento foram utilizados para obtenção do orçamento de
atividades (Clutton-Brock, 1977). Os dados obtidos foram agrupados por quantidade de
indivíduos do grupo que estavam fazendo determinada atividade em determinado tempo.
A equação abaixo foi utilizada para o cálculo da porcentagem de determinada
atividade (Cullen Jr. & Valladares-Padua, 1997).
Proporção de tempo gasto da atividade Y
% Y = nº total de observações da atividade Y x 100
nº total de observações de todas as atividades
A seguir, os dados de comportamento foram separados por horas do dia, para
construção dos gráficos de orçamento diário.
Do etograma da espécie, já consagrado na literatura, foram escolhidos alguns
comportamentos para observação, de acordo com os objetivos do projeto (Damerose &
Hopkins, 2002; Albuquerque & Codenotti, 2006). Tais comportamentos são apresentados na
Tabela 1.

33
Tabela 1. Descrição dos comportamentos presentes no etograma.
COMPORTAMENTO DESCRIÇÃO
Descanso Quando o animal está parado por mais de 5 segundos, em postura neutra,
dormindo ou não, mas sem estar envolvido em atividades sociais ou em
alimentação.
Locomoção Qualquer deslocamento do animal, na horizontal ou na vertical, por curtas ou
longas distâncias.
Social Envolve os comportamentos de catação, brincadeira, comportamentos
sexuais e vocalização.
Alimentação Quando o animal está selecionando, mastigando, segurando e/ou ingerindo
itens alimentares.
3.2.3. Análise da dieta
Para avaliação da freqüência com que diferentes alimentos são ingeridos (Struhsaker,
1975; Oates, 1977), durante a coleta de dados pelo método de varredura instantânea, nas
atividades referentes à alimentação, foi anotado o item alimentar (folha, fruto, flor, semente)
consumido pelo indivíduo e as espécies foram identificadas. Quando as espécies não foram
reconhecidas no campo, as árvores utilizadas foram marcadas com fichas plásticas para
posterior identificação da espécie, com o auxílio de floras, monografias, artigos, exsicatas do
herbário da Universidade de Caxias do Sul e especialistas. Outro procedimento para estudo
qualitativo da dieta foi a análise do conteúdo fecal, que consistiu na identificação de sementes
presentes nas fezes coletadas.
Foi utilizada a equação abaixo, para cálculo da porcentagem de utilização de uma
espécie ou item alimentar na dieta (Cullen Jr. & Valladares-Padua, 1997).

34
Composição alimentar
% espécie ou item A = nº total da espécie ou item A x 100
nº total de todas as espécies
utilizadas ou dos itens consumidos
No mês de janeiro foram coletados dados através da análise fecal e por observações ad
libitum (Altmann, 1974). Uma lista com as espécies vegetais consumidas pelo grupo foi
elaborada a partir dos dados coletados (análise qualitativa). Dados obtidos dentro da
metodologia de varredura fizeram parte das análises de freqüência de consumo de cada
espécie. Para cálculo do índice de diversidade da dieta foi usado o índice de Shannon-Wiener
(Shannon, 1948), introduzido na ecologia por Pielou (1975).
Para cada estação foram analisados as porcentagens e os índices de diversidade da
dieta para os itens consumidos e para as espécies utilizadas, bem como da contribuição das
espécies vegetais autóctones e alóctones na dieta (análises quantitativas).
3.2.4. Determinação da área de vida e uso do espaço
A área de vida (“home range”) do grupo de estudo foi determinada pelo método de
esquadrinhamento da área. Este método foi escolhido devido a melhor acurácia nos resultados
(Jardim, 2005; Ludwig, 2006). Para isso, a área foi percorrida com bússola e trena, em
transecções de sul a norte, marcando-se pontos a intervalos de 25 m com estacas de madeira.
Esses pontos marcados foram unidos por transecções de leste a oeste, formando uma retícula
de quadrantes de 25 m de lado.
Um mapa da área de estudo foi confeccionado a partir das imagens obtidas pelo
Google Earth, utilizando os softwares ArcView 3.2 e ENVI® 4.5. As imagens foram geo-
referenciadas e foi plotada sobre elas uma grade de quadrados de 25 x 25m. Os fragmentos
foram vetorizados em Arc View para obtenção das áreas de mata.

35
Todos os quadrados onde passou pelo menos uma rota foram considerados parte da
área de vida dos bugios. Para as análises de área de vida por estação do ano foram
considerados somente os registros obtidos pelo método de varredura instantânea.
O uso do espaço é a forma pelo qual os animais se utilizam do ambiente, tanto em
termos horizontais quanto verticais em sua área de vida (Miranda, 2004). Para análise do uso
do espaço horizontal, durante a varredura instantânea foi anotada a quadrícula em que o grupo
se encontrava, e, então, calculada a frequência de uso das quadrículas em cada estação.
A área nuclear é definida como uma área muito mais utilizada dentro da área de vida ,
seja porque reúnem árvores de alimentação, abrigo ou outros recursos importantes (Samuel et
al., 1985 apud Miranda 1997), sendo neste estudo definida como a área que o grupo usou
acima de 3% do tempo do periodo de estudo.
O uso diferencial dos quadrados foi quantificado através do Coeficiente de Variação
(C.V.) (Rasmussen, 1980), expresso pela equação:
____________ C.V.= √ ∑ ( ti – t )2 / N , sendo ¯ t ¯
t i = quantidade de dias que o grupo usou o quadrado “i”
t = quantidade média de dias em que cada quadrado foi usado
N = número de quadrados usados. ¯
Este índice é igual a zero se todos os quadrados tiverem a mesma densidade
ocupacional, ou seja, se todos foram usados por um número igual de dias. Os valores se
tornam maiores do que zero à medida que aumenta a variabilidade no uso dos quadrados
(Rasmussen, 1980). Os coeficientes de variação foram analisados pelo teste de Mann-
Whitney-U.
36
4. RESULTADOS E DISCUSSÃO
A etapa piloto, realizada entre janeiro e fevereiro de 2008, totalizou 99 horas de
esforço de campo, distribuídos em 13 dias, com 46 horas de observação direta do grupo de
estudo.
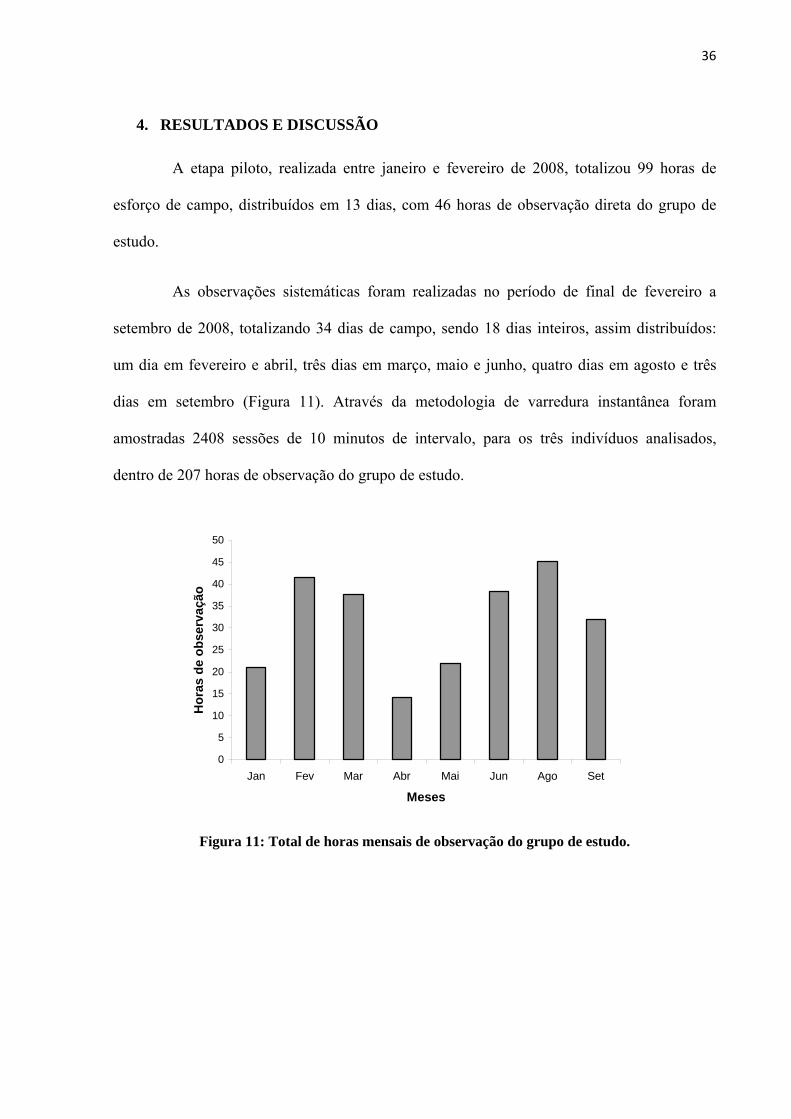
As observações sistemáticas foram realizadas no período de final de fevereiro a
setembro de 2008, totalizando 34 dias de campo, sendo 18 dias inteiros, assim distribuídos:
um dia em fevereiro e abril, três dias em março, maio e junho, quatro dias em agosto e três
dias em setembro (Figura 11). Através da metodologia de varredura instantânea foram
amostradas 2408 sessões de 10 minutos de intervalo, para os três indivíduos analisados,
dentro de 207 horas de observação do grupo de estudo.
0
5
10
15
20
25
30
35
40
45
50
Jan Fev Mar Abr Mai Jun Ago Set
Meses
Hor
as d
e ob
serv
ação
Figura 11: Total de horas mensais de observação do grupo de estudo.

37
4.1. Orçamento de atividades diárias
O modo pelo qual um organismo distribui seu tempo entre as diferentes atividades tem
conseqüências importantes da sua sobrevivência e reprodução (Watts, 1988 apud Martins,
1997), e indica a forma pela qual ele utiliza o ambiente e se amolda a mudanças (Ricklefs,
1990 apud Martins, 1997).
A Figura 12 mostra as porcentagens médias anuais do tempo diário, que o grupo focal
alocou para as diferentes atividades. O comportamento predominante foi o descanso,
ocupando 72,2% do dia. A alimentação e a locomoção foram a segunda (12,3%) e terceira
(11,4%) atividades mais observadas, respectivamente. As interações sociais (3,5%) e a
excreção (0,6%) foram reduzidas, visto que ocorrem em curtos períodos de tempo.
Alimentação12,3%
Locomoção11,4%
Social3,5%
Excreção0,6%
Descanso72,2%
Figura 12. Média das porcentagens de tempo em que o grupo de estudo alocou às diferentes
atividades, durante as observações de fevereiro a setembro de 2008.
Em comparação aos dados apresentados na literatura em Floresta Ombrófila Mista, o
grupo focal teve a maior parte do tempo gasto em descanso e o menor tempo dedicado à
alimentação. Os presentes dados ficam mais próximos dos valores registrados por Martins
(1997) com A. g. clamitans (em São Paulo) com 77% do tempo dedicado ao descanso, 10%
em alimentação e 12% em locomoção. Em comparação aos demais trabalhos, a freqüência de

38
descanso fica próxima ao encontrado em Caratinga (72%) e superior aos dados registrados em
Alegrete (68,5%), Santa Genebra (64%), Cantareira (59,5%), Aracuri (20,22%; 59,3%) e
Ribeirão Cachoeira (60%) (Calegaro-Marques,1992; Mendes, 1989; Chirello, 1993; Oliveira,
1993; Marques, 1996; Jardim & Oliveira, 2000; Gaspar, 1997).
Em contrapartida, a atividade social foi superior ao registrado na literatura, sendo
normalmente relacionada à catação entre mãe-filhote e/ou fêmea-macho, bem como algumas
atividades de vocalização para reunião do grupo ou em encontros intergrupais. Jardim &
Oliveira (2000), em Floresta Ombrófila Mista, registraram 0,9% do tempo em
comportamentos sociais. Os autores relatam ainda um gasto de 23,3% do tempo em
alimentação e 15,9% em locomoção, durante os seis meses de estudo em Esmeralda, Rio
Grande do Sul.
O alto tempo dedicado ao descanso pela espécie é decorrente do hábito folívoro. A
digestão de folhas é um processo demorado, que envolve fermentação bacteriana na quebra da
celulose, e as folhas não provém energia rapidamente (MacNab, 1978 apud Martins, 1997). O
alto tempo de descanso também é necessário para a desintoxicação do organismo devido ao
consumo de folhas com muitos compostos secundários (Glander, 1975) ou é devido à
escassez de recursos alimentares de alto valor energético.
A Tabela 2 mostra as porcentagens médias do tempo diário, que o grupo alocou para
as diferentes atividades durante os meses de estudo. Os dados estão apresentados por tipo de
atividade, não estando estas associadas em classes como nas demais análises. O mês de
fevereiro obteve a menor proporção de tempo utilizada para o descanso (51%) e um maior
tempo gasto em deslocamento (22,1%), em comparação aos outros meses de estudo. Isto pode
ser decorrente de os animais estarem recém habituados à presença dos observadores. As
atividades de alimentação foram maiores nos meses de fevereiro (15,5%) e setembro (17,1%)

39
e em menor proporção no mês de abril (6,7%). Deve-se levar em consideração que no mês de
fevereiro e abril só foi possível observar os animais durante um dia completo.
Brincadeiras foram mais observadas no início do estudo (fevereiro-abril), época em
que os animais freqüentavam diariamente a área com Eucalyptus, e ao fim do dia, entre os
períodos de alimentação, havia bastante interação entre os membros do grupo. O
comportamento catação foi mais registrado até o mês de junho. As vocalizações foram mais
freqüentes após o mês de maio, e ocorriam, normalmente, para reunião do grupo, visto que em
algumas ocasiões o macho se dispersou do grupo durante períodos de alimentação.
Comportamentos sexuais não constam na tabela, pois só foi verificada uma cópula
(fevereiro), com duração de cerca de 40 segundos, em um Pinus. Entretanto, durante o tempo
da pesquisa, não foi observado o nascimento de filhotes.
Tabela 2. Porcentagem de tempo gasto nas atividades diárias. Média mensal e média
total para os meses de estudo.
Atividade Fevereiro Março Abril Maio Junho Agosto Setembro Média
Descanso 51,0 72,1 64,2 70,9 73,1 75,0 72,6 68,4
Parado 7,4 5,7 4,2 5,9 0,3 1,7 1,4 3,8
Locomoção 22,1 6,6 16,7 11,9 7,3 8,9 6,4 11,4
Brincadeira 1,3 0,5 5,0 - 0,5 - 0,5 1,1
Catação 2,7 2,8 3,3 0,6 2,1 1,0 1,0 1,9
Vocalização - 0,5 - 1,2 0,8 0,5 - 0,4
Excreção - 1,9 - 0,6 0,5 0,5 1,0 0,6
Alimentação 15,4 9,9 6,7 8,9 15,5 12,2 17,1 12,3
Jardim & Oliveira (2000) não verificaram diferenças significativas das proporções de
tempo gasto nas três principais atividades (descanso, locomoção e alimentação) entre os
meses estudados.
40
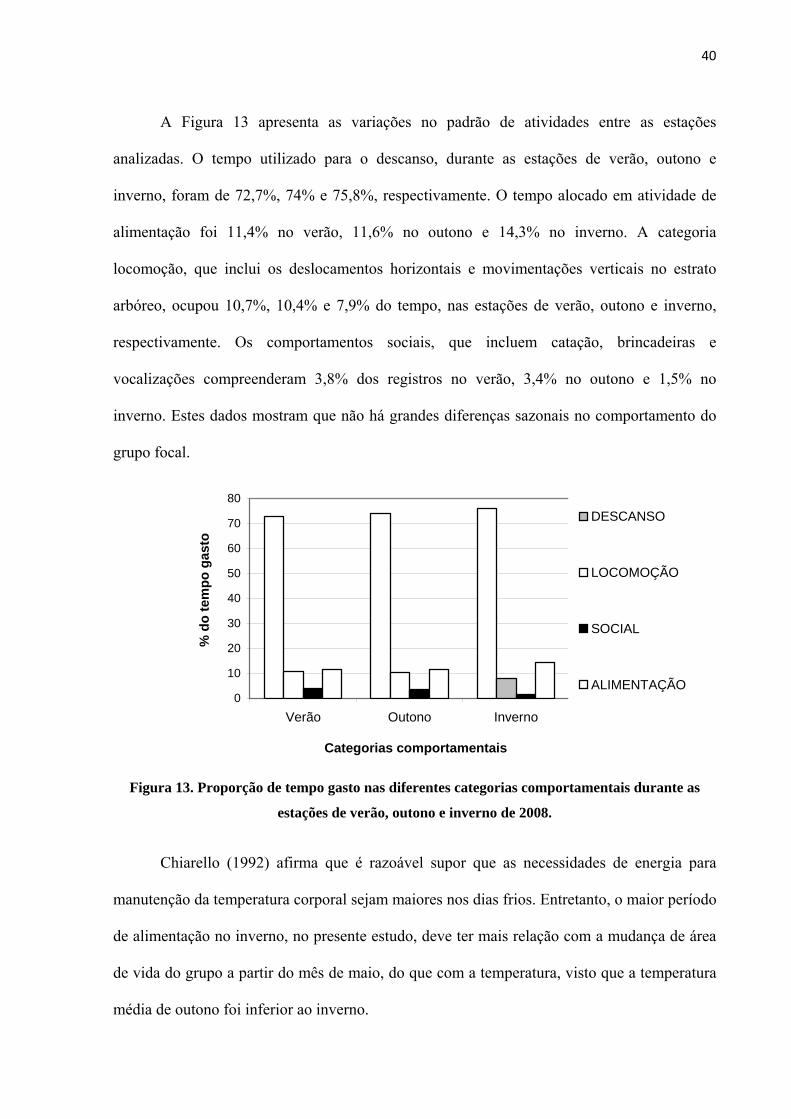
A Figura 13 apresenta as variações no padrão de atividades entre as estações
analizadas. O tempo utilizado para o descanso, durante as estações de verão, outono e
inverno, foram de 72,7%, 74% e 75,8%, respectivamente. O tempo alocado em atividade de
alimentação foi 11,4% no verão, 11,6% no outono e 14,3% no inverno. A categoria
locomoção, que inclui os deslocamentos horizontais e movimentações verticais no estrato
arbóreo, ocupou 10,7%, 10,4% e 7,9% do tempo, nas estações de verão, outono e inverno,
respectivamente. Os comportamentos sociais, que incluem catação, brincadeiras e
vocalizações compreenderam 3,8% dos registros no verão, 3,4% no outono e 1,5% no
inverno. Estes dados mostram que não há grandes diferenças sazonais no comportamento do
grupo focal.
0
10
20
30
40
50
60
70
80
Verão Outono Inverno
Categorias comportamentais
% d
o te
mpo
gas
to
DESCANSO
LOCOMOÇÃO
SOCIAL
ALIMENTAÇÃO
Figura 13. Proporção de tempo gasto nas diferentes categorias comportamentais durante as
estações de verão, outono e inverno de 2008.
Chiarello (1992) afirma que é razoável supor que as necessidades de energia para
manutenção da temperatura corporal sejam maiores nos dias frios. Entretanto, o maior período
de alimentação no inverno, no presente estudo, deve ter mais relação com a mudança de área
de vida do grupo a partir do mês de maio, do que com a temperatura, visto que a temperatura
média de outono foi inferior ao inverno.

41
O grupo focal utilizou a estratégia de aumento no período de alimentação e diminuição
de deslocamentos para minimizar gastos energéticos, durante os meses mais frios. Além
disso, nos dias mais frios, permanecerem agrupados durante o descanso e procuraram galhos
que estivessem expostos ao sol, corroborando observações de outros autores (Prates at al.,
1990b; Cunha, 1994).
A Figura 14 apresenta a variação temporal dos comportamentos descanso, locomoção
e alimentação do grupo de estudo, durante as diferentes estações do ano. Pode-se observar que
no verão os animais iniciaram suas atividades antes das 7 horas da manhã, possuindo um pico
de atividade após as 9 horas e voltando a descansar após as 11 horas, período em que a
temperatura do dia está mais elevada. Após as 14 horas, reiniciaram suas atividades,
encerrando-as após as 19 horas. Cunha (1994) obteve resultados diferentes, estudando os
bugios em um remanescente de vegetação primária no Parque Estadual de Itapuã
(Viamão/RS), onde observou picos de atividade no verão entre 11 e 12 horas.
No outono, os animais iniciaram suas atividades após as nove horas da manhã,
normalmente após a incidência do sol na mata, onde eles abrigavam-se para manter os corpos
aquecidos. Uma maior atividade ocorreu no período compreendido entre 11hs30min às
12hs30min, devido ao aumento da temperatura ambiente, bem como após as 16 horas,
variando conforme a temperatura do dia, considerando que em alguns dias que os animais
foram observados estava muito frio e, em outros, a temperatura estava mais amena.
No inverno os animais tiveram uma baixa atividade durante o dia, intensificando-a a
partir das 16 horas e 18 horas. As atividades de alimentação ficaram mais restritas ao fim da
tarde (16 horas), bem como a locomoção.

42
Verão
0
20
40
60
80
100
07:00 08:00 09:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Horas do dia
% d
as a
tivid
ades
Outono
0
20
40
60
80
100
07:00 08:00 09:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Horas do dia
% d
as a
tivid
ades
Inverno
0
20
40
60
80
100
07:00 08:00 09:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Horas do dia
% d
as a
tivid
ades
Descanso Locomoção Alimentação
Figura 14. Variação temporal média das principais atividades realizadas pelo grupo de bugios
durante as estações de verão (4 dias), outono (7 dias) e inverno (7 dias) do ano de 2008.

43
Jardim & Oliveira (2000), estudando os bugios em Floresta Ombrófila Mista (RS),
observaram uma correlação positiva entre o número de horas possíveis de brilho de sol e o
tempo médio de duração das atividades em cada mês. Assim, durante o período de inverno as
atividades estiveram restritas a um número menor de horas. Os dados aqui apresentados
corroboram Cunha (1994), que percebeu a relação entre a diminuição do nível de atividades
dos animais com horários de temperatura extrema, tanto alta como baixa.
A Figura 15 apresenta a variação temporal do comportamento descanso, durante as
diferentes estações. Pode-se observar que conforme há uma diminuição da temperatura em
cada estação, há um aumento no descanso pela parte da manhã, acarretando um atraso no
início das atividades. Tanto no verão quanto no inverno houve um incremento de atividades às
9 horas da manhã. A partir das 11 horas da manhã há uma diminuição de descanso no outono
e um aumento no verão, fato este que deve estar novamente relacionado com o aumento de
temperatura, corroborando os dados da literatura (Chitolina & Sander, 1981; Mendes, 1989;
Chiarello, 1993; Oliveira, 1993). No inverno os animais se recolheram para o descanso
noturno mais cedo, em comparação às demais estações, corroborando Miranda (1997).
0
20
40
60
80
100
07:00 08:00 09:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Horas do dia
% d
esca
nso
Verão Outono Inverno
Figura 15. Variação temporal da atividade descanso, durante as estações de verão (4 dias),
outono (7 dias) e inverno (7 dias) do ano de 2008.

44
Seria esperado que em uma situação em que a busca de alimento se mostrasse pouco
lucrativa do ponto de vista energético, houvesse uma diminuição de atividade (Cunha, 1994),
podendo ser esse o motivo pelo qual o animais do grupo focal apresentaram baixa atividade
em todos meses de estudo.
A Figura 16 apresenta a variação temporal do comportamento alimentação, durante
as diferentes estações. No verão, os animais começaram a alimentar-se a partir das 7h30min,
após acordarem e realizarem a atividade de excreção em grupo, seguido de deslocamento até
árvores próximas ao local dormitório. Um pico de atividade ocorreu às nove horas da manhã e
finalizou-se após as 10h30min. O segundo período de alimentação ocorre a partir das
13h30min, com um aumento as 14h30min e aumentando novamente até atingir um pico de
atividade após as 18 horas até os animais se recolherem para o descanso noturno. Cunha
(1994) observou no verão um pico de atividade de alimentação às 16 horas.
Durante o outono, a atividade de alimentação durante as primeiras horas do dia teve
baixa freqüência, apresentando um aumento após as 10 horas, com pico às 12h30min e
decréscimo após as 14 horas. Outro período de alimentação ocorre após as 15 horas, com um
pico às 16 horas e decréscimo após as 17 horas, com posterior aumento, terminando somente
nas ultimas horas do dia.
No inverno, os animais se alimentaram em diferentes momentos da manhã, com um
aumento considerável a partir das 14 horas, com pico às 16 horas e decréscimo da atividade
até as 18 horas, quando os animais se recolhiam para o descanso noturno, sendo este mais
cedo nesta estação, quando comparada às demais. No trabalho de Cunha (1994), o pico de
atividade de alimentação no inverno ocorre às 12 horas.

45
0
10
20
30
40
50
60
70
07:00 08:00 09:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Horas do dia
% a
limen
taçã
o
Verão Outono Inverno
Figura 16. Variação temporal da atividade alimentação, durante as estações de verão, outono e
inverno de 2008.
Chitolina & Sander (1981) e Oliveira (1993) relatam dois picos de alimentação,
ocorrendo um pela manhã e um ao final da tarde. Isso também foi evidenciado a campo,
porém em alguns dias os animais tiveram um período de alimentação a mais entre esses dois
picos.
Em duas ocasiões foram observados animais descendo ao solo para beber água em um
córrego na entrada da mata. Na primeira ocasião, em junho, às 13 horas, a fêmea desceu de
um Ligustrum lucidum, ficando agarrada neste pela cauda, por cerca de um minuto, observada
pelo seu filhote que vocalizava, vigilante. Na segunda ocasião, no mês de agosto, perto das 16
horas, observou-se o juvenil descendo para beber água no mesmo local que sua mãe havia
descido em junho, durando o comportamento cerca de meio minuto.
Em outros estudos também foi observado bugios descerem ao solo para beber água
acumulada da chuva ou de arroios, água acumuladas em ocos dos troncos, em bromélias e em
briófitas (Jardim & Oliveira, 2000; Fialho, 2000; Moro-Rios et al., 2008). Miranda et al.
(2005) também registraram a descida de uma fêmea com um infante ao solo para consumo de

46
água, nas estações de outono e inverno. Fialho (2000) observou um maior consumo de água
durante a estação de inverno.
Conforme estudo de Almeida-Silva et al. (2005), as observações de consumo de água
em um pequeno córrego por A. g. clamitans foram aos meses de julho (N=4), agosto (N=4) e
início de setembro (N=2), que coincidem com os períodos secos na região de Caratinga, MG.
Os registros dos animais bebendo água são mais comuns em períodos nos qual a
proporção de folhas na dieta é maior, pois esse item proporciona a obtenção de pequenas
quantidades de água e contém compostos secundários que necessitam de água para serem
eliminados (Bonvicino, 1989; Serio-Silva & Ricco-Gray, 2000; Miranda et al., 2005 apud
Moro-Rios et al., 2008). No presente estudo, o consumo de folhas em junho foi de 83%,
porém em agosto a ingestão foi de 32%, o que não explicaria o comportamento de descida ao
solo para beber água neste mês.
Moro-Rios et al. (2008) observaram um maior consumo de água em Floresta
Ombrófila Mista nos meses de julho (32% dos registros), maio (27%) e junho (19%). Nos
meses nos quais houve o maior consumo de água, foi detectado grande consumo de folhas e
de sementes de Araucaria angustifolia (21% em maio, 53% em junho e 10% em julho), sendo
confirmada uma correlação positiva entre o consumo das sementes e a ingestão de água.
Existe a possibilidade que os bugios aumentem a taxa de consumo de água de forma a
otimizar a digestão e excreção dos compostos secundários presentes nas sementes de A.
angustifolia. Moro-Rios et al. (op.cit.) não registraram uma correlação com o baixo consumo
de frutos e a necessidade de consumo de água.
No presente estudo, no mês de junho o consumo de pinhões fez parte de 8,2% da dieta.
Já em agosto, não foi registrado consumo da semente. O consumo de frutos nos meses de

47
junho e agosto foi 8,2% e 17,2%, respectivamente, oriundos de Hovenia dulcis, Eucalyptus
sp. e Eryobotrya japonica.
Durante a pesquisa, no mês de maio, iniciaram o corte dos Pinus presente na linha de
árvores que divide uma propriedade privada da FEGAPRO. Nesta época, os animais estavam
usando continuadamente esta área para pernoitar e, muitas vezes, passavam o dia nela. Nos
períodos de alimentação, quando não se deslocavam para o mato, faziam um alto consumo de
folhas de lianas que se desenvolveram sobre os Pinus, como também de acículas de Pinus,
folhas e frutos de Eucalyptus, folhas de Zanthoxylum rhoinfolium, folhas de Dasyphyllum
tomentosum, folhas e frutos de Quercus robur e frutos e folhas de Hovenia dulcis.
Dois dias antes de iniciarem o corte Pinus, o macho alfa sumiu durante o meio da
tarde, provavelmente tendo se deslocado pelo chão. A fêmea e o juvenil passaram o resto do
dia e o dia seguinte sem comer, vocalizando do tipo “choro”. No dia anterior ao corte,
perderam-se os dois animais de vista. No dia do corte das árvores, avistou-se um macho
solitário no inicio da tarde na entrada na mata e, no fim da tarde do mesmo dia, este animal
estava nos Pinus, sozinho.
Três dias após o inicio do corte das árvores avistou-se o grupo completo na entrada
da mata. Somente após um mês e meio observaram-se novamente os animais usando a linha
de árvores, inicialmente indo até a quadrícula B7 e, após, ficando restritos até a quadrícula
B9.

48
4.2. Dieta
4.2.4. Análise qualitativa
Os bugios utilizaram como fonte de recursos alimentares 34 espécies, distribuídas em
21 famílias botânicas, utilizando 42 itens alimentares diferentes. As espécies e os itens
alimentares utilizados pelo grupo de estudo estão listados na Tabela 3. As famílias mais
consumidas pelos bugios foram: Myrtaceae, Fabacea, Rosaceae e Sapindaceae.
As análises de fezes revelaram nove espécies consumidas, sendo três espécies
exclusivamente amostradas por este método. Foram identificadas sementes das espécies
autóctones Rapanea ferruginea, Casearia decandra, Nectandra megapotamica,
Campomanesia xanthocarpa, Lithraea molleoides, Vitex megapotamica e vestígios de flores
de Pyrostegia venusta. As espécies alóctones presentes na dieta foram Hovenia dulcis, Vitis
americana, Eucalyptus sp.

49
Tabela 3. Lista de espécies vegetais e itens alimentares utilizados na alimentação de
Alouatta guariba clamitans no período de janeiro a setembro de 2008, em um remanescente de
Floresta Ombrófila Mista em Caxias do Sul, RS. Itens: f = folhas, fl = flores, fr = frutos, s =
sementes. Metodologia: obs. direta = observação direta dos animais, fezes = análise fecal.
Família/ Espécie Nome popular Item Mês de registro Método
Araucariaceae
Araucaria angustifolia (Bertol.) O. Kuntze
pinheiro-brasileiro f, s mai, jun, ago, set obs. direta
Sapindaceae
Allophylus edulis (A. St.-Hil) Radlk. chal-chal f fev, ago obs. direta
Matayba elaeagnoides Radlk. camboatá-branco f mai, jun, ago, set obs. direta
Cupania vernalis Cambess. camboatá-vermelho
f jun obs. direta
Myrsinaceae
Myrsine ferruginea Spreng. capororoca f, fr jan, jun, set obs. direta + fezes
Flacourtiaceae
Casearia decandra Jacq. guassatonga fr jan, fev obs. direta + fezes
Fagaceae
Quercus robur L. * carvalho f, fr fev, mar, set obs. direta
Rhamnaceae
Hovenia dulcis Thunb. * uva-do-japão f, fr fev, mar, abr, mai, jun, ago, set
obs. direta + fezes
Lauraceae
Ocotea puberulla (Rich.) Ness canela-sebo f ago,set obs. direta
Nectandra megapotamica (Spreng.) Mez.
canela-preta fr jan, fev fezes
Vitaceae
Vitis americana Bartram* uva fr jan, fev obs.direta + fezes

50
Família/ Espécie Nome popular Item Mês de registro Método
Rutaceae
Zanthoxylum rhoinfolium Lam. mamica-de-cadela f fev, jun, ago, set obs. direta
Ebenaceae
Diospyrus kaki Linnaeus F. * caqui fr mai obs. direta
Myrtaceae
Blepharocalix salicifolius (Kunth) O.Berg
murta f mai, ago obs. direta
Campomanesia xanthocarpa O. Berg. guabiroba fr jan fezes
Eugenia involucrata DC. cerejeira f set obs. direta
Eucalyptus sp. L'Hér * eucalipto f, fr jan, fev, mar, abr, mai, ago, set
obs. direta + fezes
Pinaceae
Pinus taeda L.* pinus f fev, mar, abr, mai, jun
obs. direta
Pinus elliottii Engelm. * pinus f fev, mar, abr, mai, jun
obs. direta
Fabaceae
Parapiptadenia rigida (Benth.) Brenan angico f jun obs. direta
Inga virescens Benth. ingá f set obs. direta
Mimosa scabrella Benth. bracatinga f, fl ago, set obs. direta
Verbenaceae
Vitex megapotamica (Spreng.) Moldenke tarumã f, fr jan, fev, mar, mai obs. direta + fezes
Anacardiaceae
Lithraea molleoides (Vell.) Engl. aroeira-branca fr jan fezes
Lithraea brasiliensis March. bugreiro f jun, ago, set obs. direta
Bignoniaceae
Pyrostegia venusta (Ker-Gawler) Miers. cipó-de-são-joão fl ago, set obs. direta + fezes

51
Família/ Espécie Nome popular Item Mês de registro Metodologia
Asteraceae
Piptocarpha angustifolia Dusen vassourão-branco f set obs. direta
Dasyphyllum tomentosum (Spreng.) Cabrera
sucará f mar, jun, ago,set obs. direta
Oleaceae
Ligustrum lucidum W.T. Aiton * ligustro f, fr mai, jun obs. direta
Meliaceae
Cedrela fissilis Vell. cedro f set obs. direta
Rosaceea
Eryobotrya japonica (Thumb.) Lindl. * ameixa-amarela fr ago, set obs. direta
Quillaja brasiliensis (A.St.-Hil. & Tul.) Mart.
sabão-de-soldado f jun obs. direta
Prunus myrtifolia (L.) Urb. pessegueiro-bravo f set obs. direta
Euphorbiaceae
Sebastiania commersoniana (Baill.) L.B. Sm. & Downs.
branquilho f mar, jun obs. direta
* espécies vegetais alóctones
Em estudos realizados em Mata de Araucária no PR e RS, A. g. clamitans, as famílias
mais consumidas foram: Lauraceae, Flacourtiaceae, Myrtaceae, Sapindaceae e Compositae
(Jardim & Oliveira, 2000; Miranda & Passos, 2004).
Miranda & Passos (2004) reportam o consumo de 43 itens diferentes dentre as 34
espécies consumidas, em Floresta Ombrófila Mista, no Paraná. A partir de análises de fezes,
os autores relatam o consumo de 13 espécies, com sete espécies exclusivamente amostradas
dessa forma, sendo superior aos dados encontrados neste trabalho. Jardim & Oliveira (2000)
observaram o consumo 32 espécies vegetais em Floresta Ombrófila Mista, RS, estando mais
próximo dos valores encontrados neste trabalho.

52
O número de espécies consumidas pelos bugios está intimamente relacionado com a
disponibilidade de alimentos que cada ambiente oferece. Chiarello (1994) relata o consumo de
68 espécies, em Floresta Estacional Semidecidual, em São Paulo. Aguiar et al. (2003)
registraram 49 espécies em Floresta Estacional Semidecidual, no norte do Paraná. Bicca-
Marques & Calegaro-Marques (1994) observaram o consumo de 27 espécies como fonte de
alimento para A. caraya em uma mata seminatural, em Alegrete, RS.
Na Tabela 4 podem-se visualizar os meses de consumo das espécies alóctones e
autóctones pelo grupo focal. As lianas foram consumidas em todas as estações do ano
analisadas. Frutos e folhas de Eucalyptus sp. foram consumidos em todos os meses
analisados, exceto em junho. Frutos e folhas de Hovenia dulcis foram utilizadas na
alimentação do grupo durante todo estudo, não sendo observado o consumo somente em
janeiro nas observações ad libitum.
Outras espécies alóctones consumidas, além das já citadas acima, foram: frutos de
Eryobotrya japonica, folhas de Pinus, folhas e fruto de Ligustrum lucidum, frutos de Vitis
americana obtidos de uma parreira abandonada na entrada da mata, fruto de Diospyrus kaki
oriundos do pomar próximo aos Pinus e frutos e folhas de Quercus robur (Tabela 4).
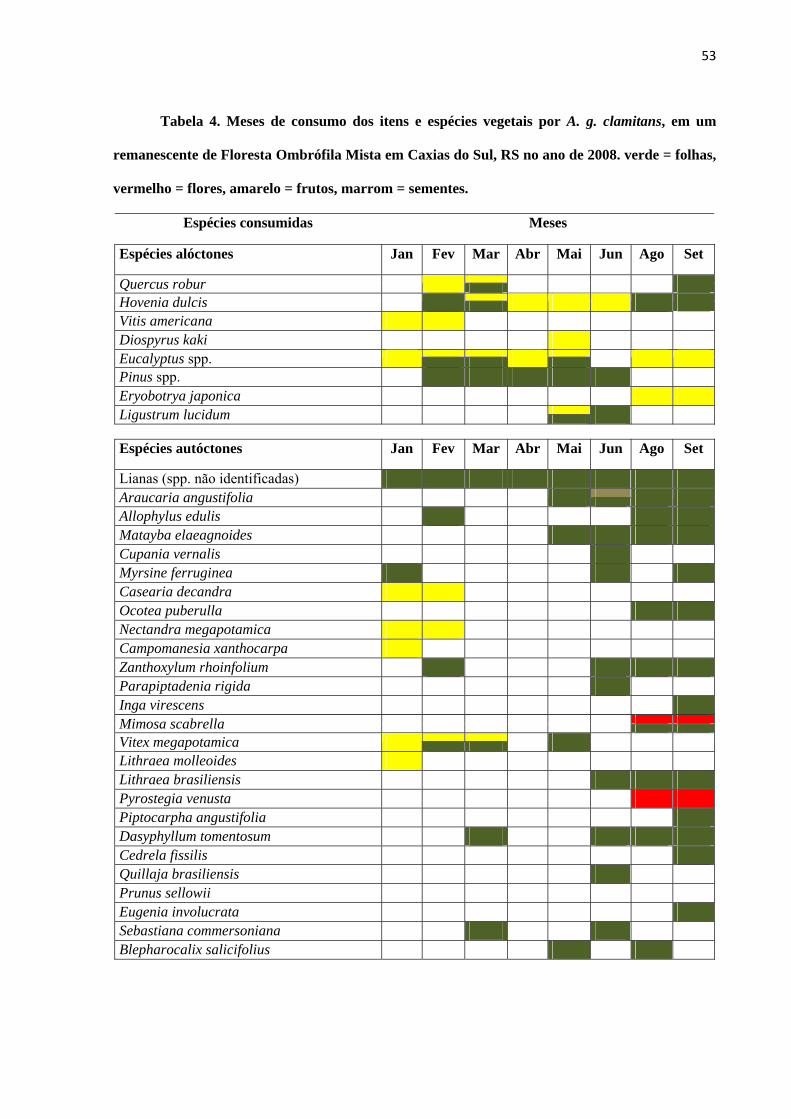
53
Tabela 4. Meses de consumo dos itens e espécies vegetais por A. g. clamitans, em um
remanescente de Floresta Ombrófila Mista em Caxias do Sul, RS no ano de 2008. verde = folhas,
vermelho = flores, amarelo = frutos, marrom = sementes.
Espécies consumidas Meses
Espécies alóctones Jan Fev Mar Abr Mai Jun Ago Set
Quercus robur Hovenia dulcis Vitis americana Diospyrus kaki Eucalyptus spp. Pinus spp. Eryobotrya japonica Ligustrum lucidum Espécies autóctones Jan Fev Mar Abr Mai Jun Ago Set
Lianas (spp. não identificadas) Araucaria angustifolia Allophylus edulis Matayba elaeagnoides Cupania vernalis Myrsine ferruginea Casearia decandra Ocotea puberulla Nectandra megapotamica Campomanesia xanthocarpa Zanthoxylum rhoinfolium Parapiptadenia rigida Inga virescens Mimosa scabrella Vitex megapotamica Lithraea molleoides Lithraea brasiliensis Pyrostegia venusta Piptocarpha angustifolia Dasyphyllum tomentosum Cedrela fissilis Quillaja brasiliensis Prunus sellowii Eugenia involucrata Sebastiana commersoniana Blepharocalix salicifolius

54
Alves & Zaú (2005) observaram um alto consumo de folhas de lianas na maioria dos
meses de estudo, não sendo consumida somente no mês de junho. Freitas et al. (2008) relatam
o consumo de Eucalyptus sp. por Cebus libidinosus nos meses de maio, novembro e
dezembro.
Jardim & Oliveira (2000) relatam um alto consumo de sementes de A. angustifolia nos
meses de maio e junho. Vários autores sugerem que a A. angustifolia seja o recurso-chave
para o bugio (Crespo, 1954; Silva, 1981; Jardim, 1992). Jardim & Oliveira (2000) consideram
que a espécie realmente tenha estreita relação com o bugio-ruivo, tanto do ponto de vista do
aporte nutricional do pinhão, como pelo aspecto estrutural das árvores que favorecem o
descanso.
Entretanto, não foi observado um grande consumo de pinhões pelo grupo de estudo.
Era esperado inclusive que, durante o mês de maio, os animais abandonassem a área de
espécies alóctones e se direcionassem para as regiões da mata com a presença da araucária.
No entanto, o que se pode observar foi a utilização mais intensa da área de mato somente após
o corte dos Pinus. Além disso, os animais consumiram mais a folha da araucária do que a
semente.

55
4.2.5. Análise quantitativa
Um total de 305 varreduras instantâneas de comportamento alimentar foram analisadas
e 33 espécies vegetais consumidas. Dentre as espécies consumidas (Tabela 5), as folhas de
lianas (espécies não identificadas) contribuíram com 24% da dieta do grupo. A espécie
alóctone Hovenia dulcis foi consumida em 12,3% das observações realizadas, contribuindo
com folhas e frutos na alimentação dos animais.
Araucaria angustifolia também teve um papel importante na alimentação do grupo,
fazendo parte de 8,6% dos registros de alimentação, porém o consumo de folhas desta espécie
foi maior do que o de sementes, sendo este poucas vezes observado. Em observações ad
libitum, foi possível obter mais registros de outros grupos de bugios consumindo pinhões, do
que do grupo em estudo. As flores da liana Pyrostegia venusta contribuíram com 6,64% da
dieta do grupo. As acículas de Pinus spp. fizeram parte de 5,32% da alimentação do grupo.
As demais espécies que contribuíram com cerca de 5 % da dieta foram: frutos de
Eryobotrya japonica, flores e folhas de Mimosa scabrella. Outra espécie alóctone utilizada
pelo grupo foi Quercus robur (4,65%), sendo utilizada as folhas e frutos desta árvore.

56
Tabela 5. Freqüência de consumo de espécies vegetais por Alouatta guariba clamitans no
período de fevereiro a setembro de 2008, em um remanescente de Floresta Ombrófila Mista em
Caxias do Sul, RS. Em ordem decrescente de freqüência dos registros de alimentação. FA =
freqüência absoluta, FR =freqüência relativa, N = Numero de meses de uso (máximo 8 meses).
Espécie Família FA FR% N
Lianas (spp. não identificadas) - 73 24,25 8
Hovenia dulcis Rhamnaceae 37 12,29 7
Araucaria angustifolia Araucariaceae 26 8,64 4
Pyrostegia venusta Bignoniaceae 20 6,64 2
Pinus spp. Pinaceae 16 5,32 5
Eryobotrya japonica Rosaceae 15 4,98 2
Mimosa scabrella Fabaceae 15 4,98 2
Quercus robur Fagaceae 14 4,65 3
Zanthoxylum rhoifolium Rutaceae 12 3,99 4
Dasyphyllum tomentosum Asteraceae 12 3,99 4
Ligustrum lucidum Oleaceae 12 3,99 2
Matayba elaeagnoides Sapindaceae 12 3,88 4
Eucalyptus sp. Myrtaceae 10 3,32 7
Lithraea brasiliensis Anacardiaceae 8 2,91 3
Vitis americana Vitaceae 5 1,66 2
Parapiptadenia rigida Leguminosae 4 1,33 2
Vitex megapotamica Verbenaceae 3 0,97 4
Casearia decandra Flacourtiaceae 2 0,66 3
Diospyrus kaki Ebenaceae 2 0,66 1
Piptocarpha angustifolia Asteraceae 2 0,66 1
Cedrela fissilis Meliaceae 2 0,66 1
Allophylus edulis Sapindaceae 1 0,33 2
Quillaja brasiliensis Rosaceae 1 0,33 1
Myrsine ferruginea Myrsinaceae 1 0,33 3
Ocotea puberula Lauraceae 1 0,33 2
Inga virescens Fabaceae 1 0,33 1
Blepharocalix salicifolius Myrtaceae 1 0,33 2

57
Em estudos realizados em Mata de Araucária no PR e RS, as espécies que mostraram
maior importância na dieta do bugio-ruivo foram: Araucaria angustifolia, Gochnatia
polymorpha, Matayba elaeagnoides, Mimosa scabrella, Ocotea porosa, O. odorifera, O.
puberulla, O. pulchella e Zanthoxylum rhoifolium (Jardim & Oliveira, 2000; Miranda &
Passos, 2004). Apesar das diferenças encontradas na proporção de consumo destas espécies
entre o presente trabalho e os trabalhos citados, a maioria delas foi utilizada na alimentação
do grupo focal.
Freitas et al. (2008) registrou o consumo de espécies alóctones por Cebus libidinosus
em SP. 22% da dieta foi composta por Zea mays, 9,4% por Saccharum officinarum, 0,3% por
frutos, sementes e néctar de Eucalyptus sp., 0,1% por polpa de Citrus sp. e frutos de Melia
azedarach.
No presente trabalho, os itens alimentares foram divididos em categorias: folha, fruto,
flor e semente (Figura 17). Em todas as estações, o consumo de folhas (64%) foi superior ao
consumo dos outros itens alimentares, sendo esta diferença mais evidenciada no outono. Os
frutos foram consumidos em 25% do tempo, as flores em 8,7% e sementes 1,7%.
Durante o verão, a dieta dos bugios foi composta basicamente por folhas e frutos. No
outono a ingestão de folhas atingiu 80% e, além do consumo de frutos, os animais ingeriram
também sementes de Araucaria angustifolia. No inverno, além das folhas e frutos, houve um
consumo de flores das espécies Pyrostegia venusta e Mimosa scabrella. O consumo de folhas foi
maior no outono do que nos demais meses e frutos foram mais consumidos no verão.

58
0
1020
3040
5060
7080
90
folhas frutos sementes flores
Itens alimentares
Freq
üênc
ia (%
)
verão
outono
inverno
Figura 17. Freqüência de consumo dos itens alimentares nas estações de verão, outono e inverno
de 2008.
Dentre os primatas neotropicais, os primatas do gênero Alouatta são os mais dependes
de folhas (Franceschinelli et al., 2005). Em Floresta Ombrófila Mista, Miranda & Passos
(2004) observaram uma maior freqüência do período de alimentação relacionada às folhas
(57,3%), seguidos de frutos (41%) e flores (1,7%). Jardim & Oliveira (2000) estudando
bugios em ambiente florestal semelhante registraram um consumo de folhas superior, sendo
82% do tempo de alimentação dedicado a esse item, seguido de sementes (11,1%) e flores
(6,9%). Alves & Zaú (2007) obtiveram dados semelhantes no que se refere ao consumo de
folhas (81% da dieta). Brotos fizeram parte de 10% da alimentação dos bugios, seguidos de
7% de frutos e 2% de flores, em Floresta Estacional Semidecidual Submontana, no RJ.
Conforme trabalho de Mendes (1989), na Estação Biológica de Caratinga (MG),
houve um consumo de 77% de folhas, 14% frutos e 9% flores. Lunardelli (2000) reporta o
consumo de 55% do tempo em folhas, 15% frutas e 29% flores. Aguiar et al. (2003)
registraram o consumo de 50,3% folhas, 47,9% frutos, 1,4% flores e 0,3% caules, em Floresta
Estacional Semidecidual, no PR.

59
Em estudo com A. caraya, 60,9% dos registros de alimentação estavam relacionados
ao consumo de folhas, 28,9% de frutos e sementes, 6,4% de caules, 2,7% de flores, 1,0% de
cascas e 0,1% de liquens (Bicca-Marques & Calegaro-Marques, 1994).
No presente estudo, observou-se a alta preferência dos bugios pelos itens sazonais,
principalmente frutos e flores. Este comportamento alimentar também foi observado por
outros autores (Miranda & Passos, 2004; Milton, 1980; Mendes, 1989; Bicca-Marques &
Calegaro-Marques, 1995). O consumo de flores somente foi observado no inverno,
corroborando os dados de Fialho (2000), estudando bugios em mata de encosta (Porto Alegre,
RS). Em contrapartida, Aguiar et al. (2003) observou um maior consumo deste item no
outono (Floresta Estacional Semidecidual, PR).
Com o passar dos meses, os bugios aumentaram o número de espécies consumidas por
dia de observação (Figura 18). No mês de fevereiro consumiram cinco espécies, em março a
média foi quatro, em abril quatro, em maio 5,5, em junho e agosto seis e em setembro a média
foi de 13 espécies. É possível visualizar que o número de espécies aumentou após o mês de
maio, devido a mudança da área de vida e incremento na alimentação do grupo.
0
2
4
6
8
10
12
14
16
Fev Mar Abr Mai Jun Ago Set
Meses
Nº d
e es
péci
es
Figura 18. Número de espécies consumidas pelo grupo de A. g. clamitans em dias completos
(mín. 8 horas), nos meses de estudo em 2008.

60
A Figura 19 mostra a porcentagem do consumo das espécies vegetais em cada estação
do ano. No verão (N=11), houve um alto consumo de folhas de lianas (43%), seguido de
frutos de uva-do-japão (H. dulcis), perfazendo 20% da dieta. No outono (N=14) o consumo de
lianas também foi maior que das demais espécies (27%). A espécie A. angustifolia foi
bastante representativa durante esta estação (15%), devido ao consumo de suas folhas e
sementes. Pinus spp. contribuiu com 14% da dieta do grupo de bugios durante o outono.
Na estação do inverno houve o consumo de um maior número de espécies (N=18),
com um alto consumo de flores de P. venusta (17%), seguidos de flores de M. scabrella e
frutos de E. japonica (12%).

61
Verão
Lianas
Hovenia dulcis
Quercus robur
Vitis americana
Eucalyptus sp.
Dasyphyllum tomentosum
Casearia decandra
Pinus spp.
Allophylus edulis
Vitex megapotamica
Zanthoxylum rhoifolium
(a)
Outono
LianasAraucaria angustifoliaPinus spp.Ligustrum sp.Matayba elaeagnoidesHovenia dulcisParapiptadenia rigidaVitex megapotamicaDiospyrus kaki Lithraea brasiliensisBlepharocalix salicifoliusZanthoxylum rhoifoliumEucalyptus sp.Quillaja brasiliensis
(b)
Inverno Pyrostegia venustaMimosa scabrellaEriobotrya japonicaHovenia dulcisAraucaria angustifoliaZanthoxylum rhoifoliumLianasDasyphyllum tomentosumLithraea brasiliensisEucalyptus sp.Matayba elaeagnoidesPiptocarpha augustifoliaCedrela fissilis Rapanea ferrugineaQuercus roburInga virescens Ocotea puberula (c)
Figura 19. Porcentagem de consumo das diferentes espécies vegetais pelo grupo de estudo nas
estações de verão (a), outono (b) e inverno (c) de 2008.

62
A espécie Pyrostegia venusta, amplamente utilizada pelo grupo focal na alimentação
durante a estação de inverno, também foi um recurso muito utilizado no trabalho de Martins
(1997). Segundo o autor, Pyrostegia venusta (cipó-de-são-joão) é uma espécie de cipó que
ocorre em áreas perturbadas, geralmente em bordas e após a ação do fogo.
A diversidade das espécies utilizadas na alimentação do bugio-ruivo durante as
estações de verão, outono e inverno calculada através do índice de Shannon (1948) pode ser
verificada na Tabela 6. A estação de verão teve o menor índice de diversidade e equidade,
devido ao alto consumo de lianas durante este período. Já o inverno, apresentou o maior
índice de diversidade e equidade, visto que a alimentação do grupo foi composta por um
grande número de espécies que contribuíram de forma mais homogênea na dieta dos animais.
Tabela 6. Parâmetros ecológicos (diversidade, riqueza e equidade) das espécies
alimentares, analisadas por estação do ano (verão, outono e inverno de 2008).
Estação Hs S Hmax Equidade
Verão 1,72 11 2,40 0,71
Outono 2,16 14 2,64 0,82
Inverno 2,62 18 2,89 0,91
Hs = índice de diversidade
S = número de espécies
Hmax = índice do Shannon-Wiener maximizado
Equidade = igual a 1 é homogênea.
O grupo focal utilizou menos espécies no verão, possivelmente por que na área que
eles estavam utilizando durante este período há menor disponibilidade de alimento. Martins
(1997) também obteve uma diversidade de espécies menor no verão, devido a uma maior
seletividade dos animais e por escolherem itens e espécies mais nutritivos.
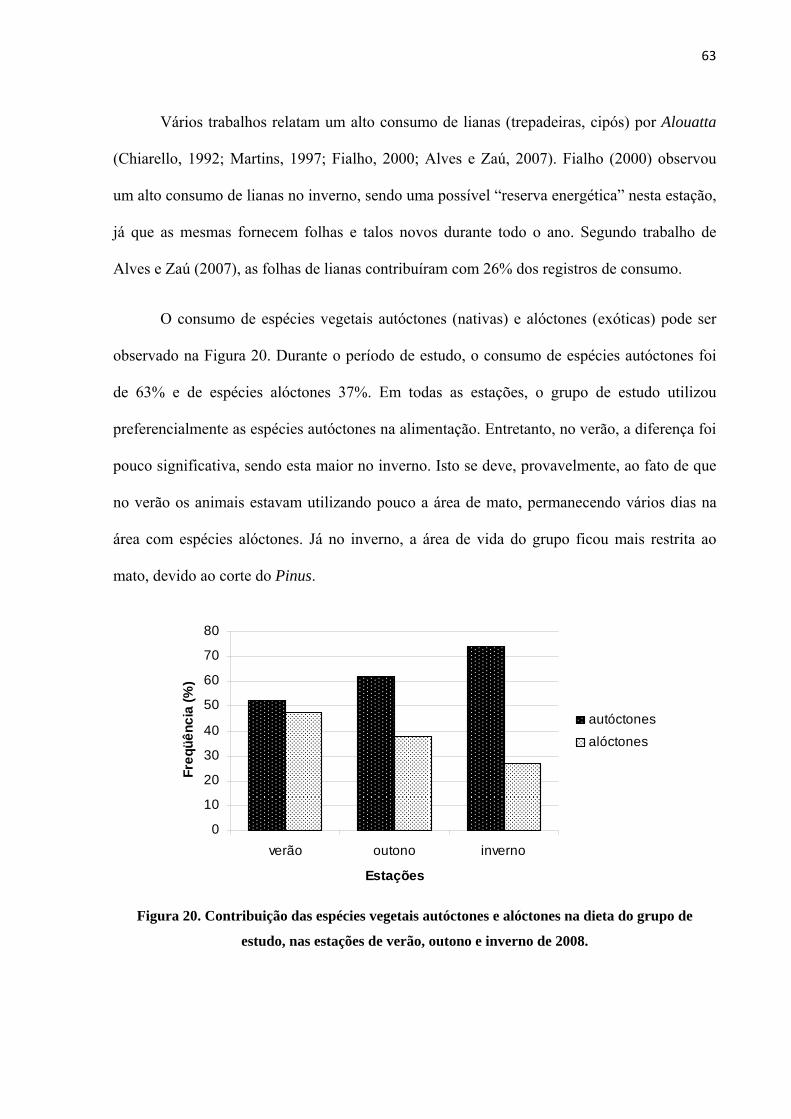
63
Vários trabalhos relatam um alto consumo de lianas (trepadeiras, cipós) por Alouatta
(Chiarello, 1992; Martins, 1997; Fialho, 2000; Alves e Zaú, 2007). Fialho (2000) observou
um alto consumo de lianas no inverno, sendo uma possível “reserva energética” nesta estação,
já que as mesmas fornecem folhas e talos novos durante todo o ano. Segundo trabalho de
Alves e Zaú (2007), as folhas de lianas contribuíram com 26% dos registros de consumo.
O consumo de espécies vegetais autóctones (nativas) e alóctones (exóticas) pode ser
observado na Figura 20. Durante o período de estudo, o consumo de espécies autóctones foi
de 63% e de espécies alóctones 37%. Em todas as estações, o grupo de estudo utilizou
preferencialmente as espécies autóctones na alimentação. Entretanto, no verão, a diferença foi
pouco significativa, sendo esta maior no inverno. Isto se deve, provavelmente, ao fato de que
no verão os animais estavam utilizando pouco a área de mato, permanecendo vários dias na
área com espécies alóctones. Já no inverno, a área de vida do grupo ficou mais restrita ao
mato, devido ao corte do Pinus.
0
10
20
30
40
50
60
70
80
verão outono inverno
Estações
Freq
üênc
ia (%
)
autóctones
alóctones
Figura 20. Contribuição das espécies vegetais autóctones e alóctones na dieta do grupo de
estudo, nas estações de verão, outono e inverno de 2008.

64
A contribuição das espécies vegetais alóctones e seus itens alimentares na dieta do
grupo, para cada estação do ano, pode-se ser visualizada na Figura 21. No verão, os frutos de
Hovenia dulcis foram o item mais consumido pelos bugios, em relação aos demais itens
derivados de espécies alóctones. Houve um grande consumo de frutos de Quercus robur,
seguidos de frutos de Vitis americana. As folhas das espécies alóctones foram pouco
consumidas.
No outono, houve um alto consumo de folhas de Pinus spp. e Ligustrum lucidum.
Frutos de Hovenia dulcis foram novamente um importante recurso alimentar dos bugios nesta
estação, sendo a dieta complementada com frutos de Diospyrus kaki e Ligustrum lucidum. No
inverno, os bugios utilizaram como parte da dieta frutos de Eriobotrya japonica, folhas de
Hovenia dulcis e frutos de Eucalyptus sp. Os animais ingeriram também, em baixa
porcentagem, folhas de Quercus robur e frutos de Hovenia dulcis.
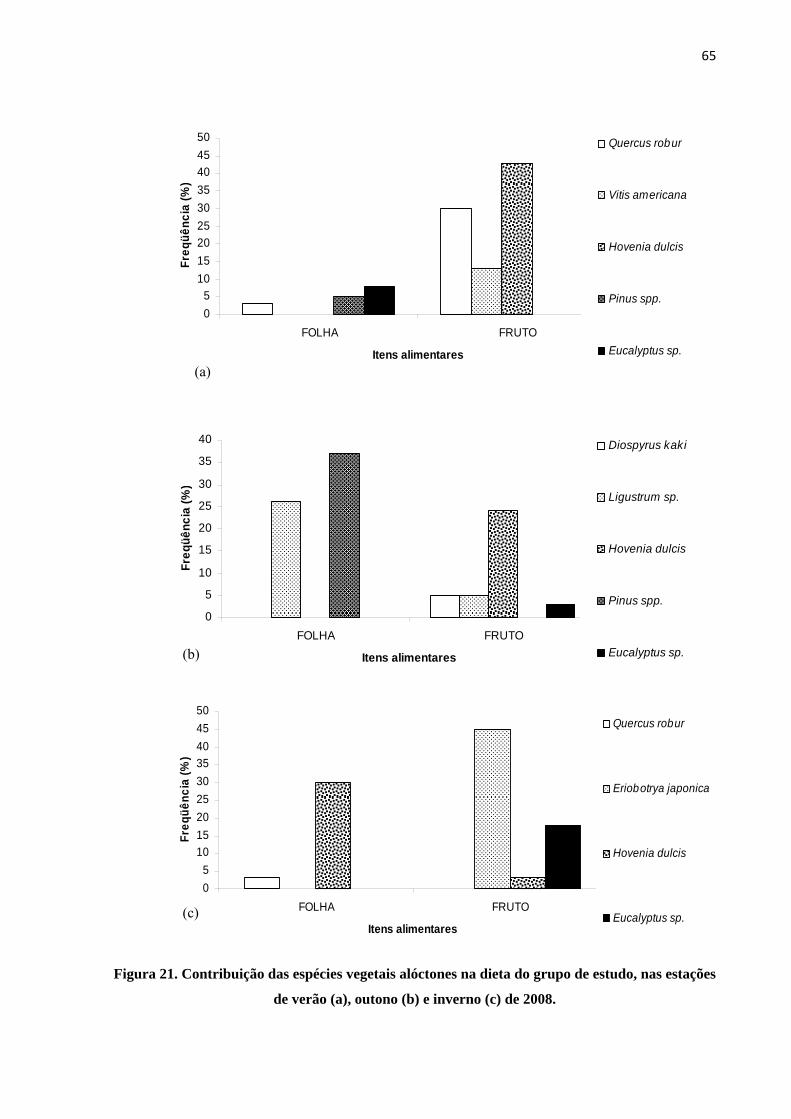
65
05
101520253035404550
FOLHA FRUTO
Itens alimentares
Freq
üênc
ia (%
)
Quercus robur
Vitis americana
Hovenia dulcis
Pinus spp.
Eucalyptus sp.
(a)
0
5
10
15
20
25
30
35
40
FOLHA FRUTO
Itens alimentares
Freq
üênc
ia (%
)
Diospyrus kaki
Ligustrum sp.
Hovenia dulcis
Pinus spp.
Eucalyptus sp.
(b)
05
101520253035404550
FOLHA FRUTO
Itens alimentares
Freq
üênc
ia (%
)
Quercus robur
Eriobotrya japonica
Hovenia dulcis
Eucalyptus sp.
(c)
Figura 21. Contribuição das espécies vegetais alóctones na dieta do grupo de estudo, nas estações
de verão (a), outono (b) e inverno (c) de 2008.

66
Bicca-Marques & Calegaro-Marques (1994) relatam o consumo de espécies
autóctones durante 62,4% dos registros e alóctones em 37,7% dos mesmos, estudando A.
caraya em Alegrete, RS. Dentre as espécies alóctones consumidas, incluem-se sementes e
cascas de Eucalyptus sp. Estes valores ficam muito próximo aos observados no presente
estudo (63% e 37%).
Miranda & Passos (2004) observaram um grande consumo de alimentos alóctones ao
ambiente de Floresta Ombrófila Mista, no período entre maio e agosto, tais como Eryobotrya
japonica (Thunb.) Lindl. e Diospyrus kaki L.F. Silveira & Codenotti (2001) também reportam
o consumo de Diospyrus kaki para A. g. clamitans no Rio Grande do Sul. Aguiar et al. (2003)
observou o consumo de Hovenia dulcis, no PR.
Existem trabalhos que relatam a utilização de Pinus spp. na dieta de algumas espécies
animais, como roedores e os macacos dos gêneros Cebus e Cercopithecus (Bussche & Zee,
1985; Koehler & Firkowski, 1996; Rocha, 2000; Miranda, 2005; Andreiv & Firkowski, 2006;
Gonçalves et al., 2007; Carvalho, 2007). O consumo desta espécie ainda não havia sido
documentada para A. g. clamitans.
De um total de 73 espécies consumidas por Cebus nigritus, no PR, seis eram exóticas
(Zea mays, duas espécies de Citrus spp., Hovenia dulcis, Caryota urens e Musa paradisiaca)
(Ludwig et al., 2005). Freitas et al. (2008) observou um alto consumo de cultivares por Cebus
libidinosus em épocas que a produção de frutos zoocóricos foi baixa nos fragmentos de
estudo.
Prates (2007) estudou um grupo de bugios-pretos (A. caraya) em Alegrete, RS, com
12 a 14 indivíduos vivendo em um pequeno pomar de 0,7 ha, com dossel descontínuo e aberto
e com baixa riqueza de espécies vegetais, confirmando a adaptabilidade da espécie em
sobreviver em condições ambientais extremas. A autora também observou que não houve

67
alterações significativas nos padrões comportamentais destes animais, reforçando a hipótese
de que Alouatta spp. são adaptados para sobreviver em ambientes com diferentes graus de
alteração.
O conhecimento de quais espécies são importantes para a manutenção de Alouatta se
faz importante para projetos de desenvolvimento de corredores ecológicos e recomposição de
áreas que possam, de alguma maneira, aumentar a probabilidade de sobrevivência da
população em questão. A compreensão dos limites de elasticidade alimentar e das bases que
os explicam permite estimar o quanto uma mata é potencialmente adequada ao
estabelecimento de uma população (Lindeberg & Santini, 1984 apud Fialho, 2000).
Os macacos defecam as sementes dos frutos ingeridos em grupos ao se deslocarem
pela floresta ou sob seus dormitórios (Julliot, 1997; Izar, 1999). Os bugios são
reconhecidamente importantes dispersores de sementes, colaborando para a manutenção e
sustentabilidade em longo prazo do ecossistema florestal (Chapman, 1989; Figueiredo, 1993;
Julliot, 1996; Chapman & Onderdonk, 1998; Liesenfeld, 1999; Liesenfeld, 2003; Andresen,
2006). Algumas espécies, inclusive, aumentam sua taxa de germinação após a passagem no
trato digestivo destes animais (Figueiredo, 1993; Bravo et al., 1995).
Em análises de germinação de sementes, em 95% das espécies encontradas nas fezes
dos bugios, a taxa de sementes intactas foi maior que as predadas, resultado este que parece
atribuir os bugios a um status de bons dispersores (Ribeiro et al., 2007).
Em restauração de ambientes degradados, há tentativas de gerar uma restauração
induzida, utilizando os vertebrados frugívoros como importantes agentes no fluxo de
sementes (Jordano et al., 2006). Agindo como dispersores de sementes, os bugios poderiam
favorecer a recuperação de áreas degradadas e alteradas.

68
Entretanto, pelo que foi observado neste estudo é provável que os bugios estejam
agindo como dispersores de Hovenia dulcis (espécie alóctone), devido ao grande número de
sementes intactas encontradas nas fezes dos animais. Este fato é preocupante, visto que pode
desencadear uma “contaminação ambiental” pela dispersão de espécies alócontes ao invés de
uma restauração do hábitat.
Alguns trabalhos têm demonstrado que reflorestamentos com eucalipto, que
apresentem sub-bosque, constituem um habitat freqüentado por diversas espécies de aves e
mamíferos potencialmente dispersores de sementes (Motta Jr., 1990; Stallings, 1990;
Machado & Lamas, 1996; Willis, 2002 apud Jordano et al., 2006). Neste estudo, foi
observado o uso da área com Eucalyptus pelos mamíferos: A.g.clamitans, Lepus europaeus
(lebre), Dasyprocta azarae (cutia), Coendou prehensilis (ouriço). Na área com Pinus
observaram-se A.g.clamitans e D. azarae e as aves Penelope obscura (jacu), Cyanocorax
caeruleus (gralha-azul), Rupornis magnirostris (gavião carijó). Foi registrado um bando de
Pyrrhura frontalis (tiriba-da-testa-vermelha) ingerindo sementes de Pinus. Estas aves
ingeriram também sementes de Carya illionensis e C. prehensilis consumiu folhas de Quercus
robur.
Outro fato interessante é que apesar da área de mata possuir exemplares de Syagrus
romanzoffiana, relatado na literatura como fonte de frutos para os bugios (Prates et al., 1990;
Cunha, 1994; Fialho, 2000; Miranda & Passos, 2004), não foi observado o consumo desta
espécie no presente trabalho.
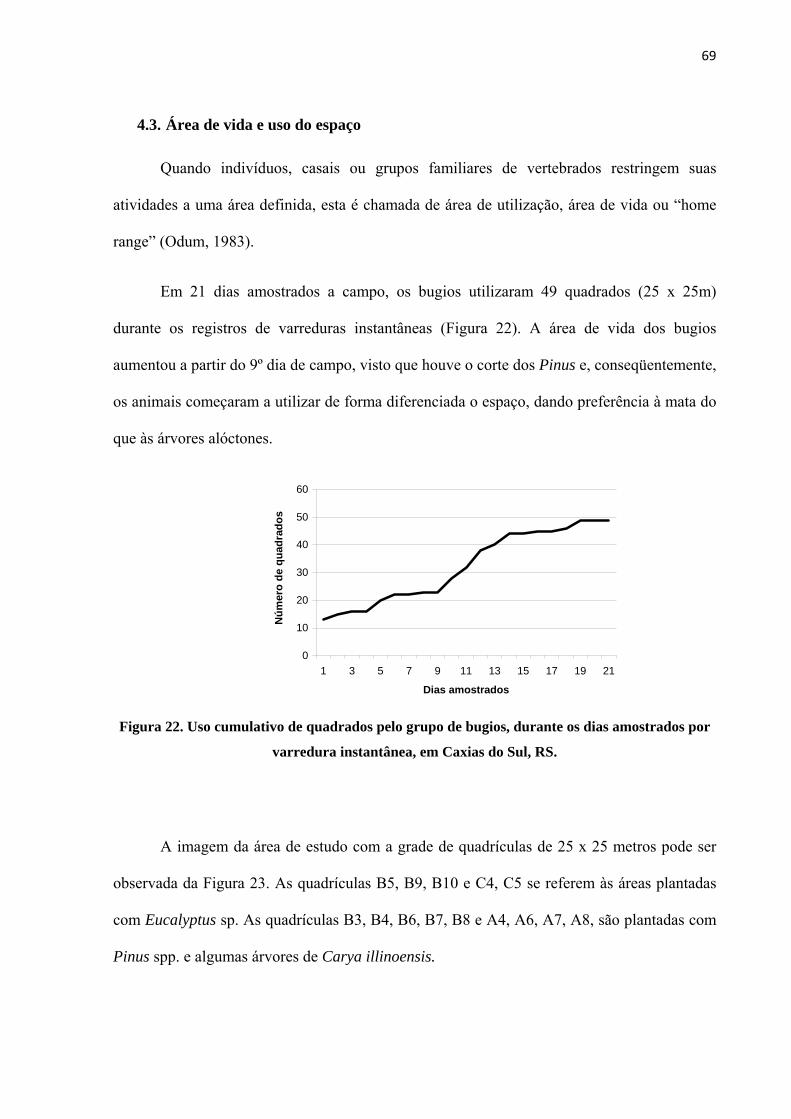
69
4.3. Área de vida e uso do espaço
Quando indivíduos, casais ou grupos familiares de vertebrados restringem suas
atividades a uma área definida, esta é chamada de área de utilização, área de vida ou “home
range” (Odum, 1983).
Em 21 dias amostrados a campo, os bugios utilizaram 49 quadrados (25 x 25m)
durante os registros de varreduras instantâneas (Figura 22). A área de vida dos bugios
aumentou a partir do 9º dia de campo, visto que houve o corte dos Pinus e, conseqüentemente,
os animais começaram a utilizar de forma diferenciada o espaço, dando preferência à mata do
que às árvores alóctones.
0
10
20
30
40
50
60
1 3 5 7 9 11 13 15 17 19 21
Dias amostrados
Núm
ero
de q
uadr
ados
Figura 22. Uso cumulativo de quadrados pelo grupo de bugios, durante os dias amostrados por
varredura instantânea, em Caxias do Sul, RS.
A imagem da área de estudo com a grade de quadrículas de 25 x 25 metros pode ser
observada da Figura 23. As quadrículas B5, B9, B10 e C4, C5 se referem às áreas plantadas
com Eucalyptus sp. As quadrículas B3, B4, B6, B7, B8 e A4, A6, A7, A8, são plantadas com
Pinus spp. e algumas árvores de Carya illinoensis.

70
Figura 23. Imagem da área de estudo com a grade de quadrículas (25x25m).
Em decorrência da mudança na paisagem após a supressão do Pinus, houve uma
mudança na área de vida e uso do espaço pelo grupo focal e, também, da sua dieta,
evidenciando a flexibilidade da espécie.
A área de uso é geralmente definida como a área usada para alimentação e descanso
(Jolly, 1972). Diferenças no tamanho da área de vida e no padrão de uso do espaço em
primatas são decorrentes das variáveis ambientais e sociais tais como distribuição e
abundância das fontes de alimento, disponibilidade de água, disposição espacial de áreas de
dormida, disponibilidade de parceiros sexuais, presença ou ausência de competidores e/ou

71
predadores, manutenção do território, tamanho do corpo e necessidades metabólicas (Macnab,
1963; Struhsaker, 1974; Gautier-Hion et al., 1981; Terborgh, 1983).
O uso do espaço do grupo antes da supressão dos Pinus e depois desta pode ser
observada na Figura 24. As áreas com coloração mais escura se referem a maior porcentagem
de uso e com cores mais claras, uma menor porcentagem de utilização. Fica evidente a busca
de uma nova área de vida pelo grupo, após o corte das árvores, aumentando a área de uso do
grupo de 1,8 para 2,5 ha. Entretanto, não houve diferenças significativas segundo o Teste de
Mann Whitney-U quanto ao coeficente de variação do uso do espaço entre as duas situações
(C.V.: 0,82 antes do corte; C.V.: 1,18 após o corte).
Figura 24. Uso do espaço pelo grupo de bugios antes do corte dos Pinus (a) e depois do corte (b),
de janeiro a setembro de 2008. Freqüência de uso analisada a partir de fevereiro. Linhas mais
espessas se referem ao uso das quadrículas fora dos períodos de varredura instantânea.

72
Partes da área de uso utilizadas mais intensamente são geralmente denominadas áreas
nucleares (Jolly, 1985). A Figura 25 apresenta as áreas nucleares de utilização pelo grupo e os
sítios de pernoite. As áreas que tiveram uma maior percentagem de uso, em sua maioria, se
referem às áreas que foram utilizadas como dormitório, visto que estas eram utilizadas por
boa parte da manhã e nas ultimas horas do dia. Nas quadrículas F18 e F19 há alguns
exemplares de araucária e na quadrícula C15 há ligustros e grande presença de lianas.
As árvores utilizadas como sítio de pernoite foram repetidas ao longo das observações,
às vezes, em dias consecutivos. Exemplares de Pinus e Eucalyptus foram utilizados como
sítios de pernoite na maioria dos registros (Figura 25). Este fato deve-se possivelmente à
maior proteção que estas árvores oferecem contra predadores devido ao seu grande porte e
também pela espessura e disposição dos galhos, considerando que a mata da área de estudo é
uma mata secundária, apresentando poucas árvores emergentes além da Araucaria
angustifolia. Piptocarpha angustifolia e Parapiptadenia rigida são árvores de grande porte
que poderiam servir como árvore dormitório, todavia estas árvores estão concentradas na
região oeste da mata, onde os animais foram observados somente durante os períodos de
alimentação.
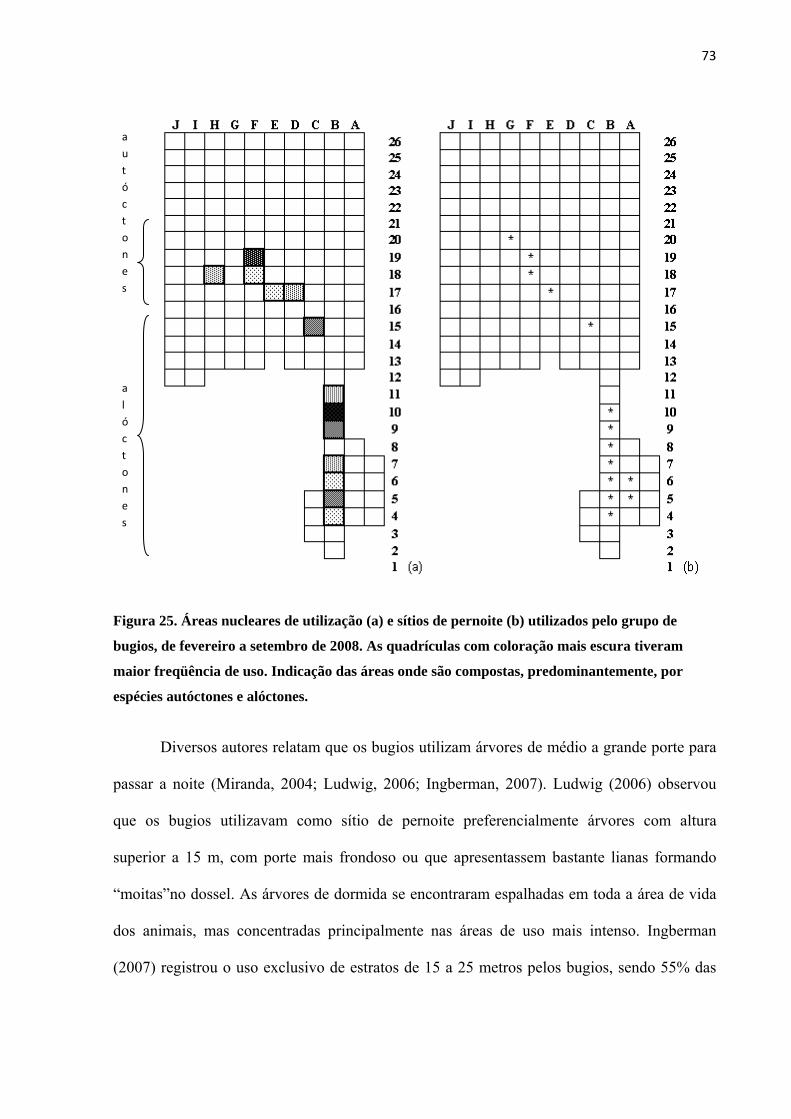
73
alóctones
autóctones
Figura 25. Áreas nucleares de utilização (a) e sítios de pernoite (b) utilizados pelo grupo de
bugios, de fevereiro a setembro de 2008. As quadrículas com coloração mais escura tiveram
maior freqüência de uso. Indicação das áreas onde são compostas, predominantemente, por
espécies autóctones e alóctones.
Diversos autores relatam que os bugios utilizam árvores de médio a grande porte para
passar a noite (Miranda, 2004; Ludwig, 2006; Ingberman, 2007). Ludwig (2006) observou
que os bugios utilizavam como sítio de pernoite preferencialmente árvores com altura
superior a 15 m, com porte mais frondoso ou que apresentassem bastante lianas formando
“moitas”no dossel. As árvores de dormida se encontraram espalhadas em toda a área de vida
dos animais, mas concentradas principalmente nas áreas de uso mais intenso. Ingberman
(2007) registrou o uso exclusivo de estratos de 15 a 25 metros pelos bugios, sendo 55% das

74
observações a 15 metros, 36% a 20m e somente 9% nas árvores emergentes de altura de 25
metros.
Entretanto, segundo Jardim & Oliveira (2000) existe a necessidade dos animais
descerem a níveis mais baixos de altura para uma maior exploração de recursos alimentares e
retornarem ao estrato superior para realizar as outras atividades com maior segurança. No
presente estudo verificou-se a utilização de árvores de baixo e médio porte durante os
períodos de alimentação.
Em 59% dos registros de árvores dormitório do grupo de estudo foram em espécies
alóctones (Pinus e Eucalyptus) e 41% de espécies autóctones (Araucaria angustifolia e, em
uma ocasião, outra espécie nativa) (Figura 26). Até a supressão dos Pinus, estes eram
utilizados como sítio de pernoite pelo grupo.
0
1
2
3
4
5
Fevereiro Março Abril Maio Junho Agosto Setembro
Meses
Reg
istr
os
Espécies autóctones Espécies alóctones
Figura 26. Uso de espécies vegetais autóctones e alóctones como sítio de pernoite pelo grupo focal
nos meses de observação do ano de 2008.
Jardim & Oliveira (2000) relatam uma forte preferência no uso de Araucaria
angustifolia para o descanso noturno, onde 74% dos registros foram nesta espécie. Durante o
período de disponibilidade de pinhão, os bugios utilizaram as araucárias em 100% dos
registros.

75
Em seis ocasiões durante os meses de junho, agosto e setembro observaram-se a
aproximação de um grupo de três indivíduos, de forma agonística e agressiva, sendo o macho
alfa maior que o macho do grupo focal. Estes encontros (Tabela 7), ocorridos nas quadrículas
G18, G19 e E17 causaram o deslocamento do grupo focal para as quadrículas B15 e C15, e,
posteriormente, para a área com Eucalyptus.
Nos encontros observados, o macho do grupo vizinho vocalizou para o outro grupo e,
em duas ocasiões, foi até a quadricula D17, que é a ultima quadricula ao sul da mata,
vocalizou e defecou, possivelmente numa atitude de defesa de território. Em contrapartida, o
grupo focal demonstrou submissão, retirando-se da área sem enfrentar e/ou vocalizar para o
outro grupo. Em um dos encontros o macho do grupo vizinho tentou atacar a fêmea e o
juvenil do grupo focal, podendo-se ouvir “gritos” dos mesmos. Chiarello (1995) registrou 15
agressões físicas em encontros intergrupais (35% dos casos), estudando A. g. clamitans na
Reserva Santa Genebra.
Tabela 7. Encontros intergrupais observados durante o período de estudo.
Data Local do encontro (quadrícula)
23/06/08 E17
30/06/08 G18 até D17
30/06/08 G19 até D16
23/08/08 G18 até D16
14/09/08 G18
15/09/08 G19 até E17
Os encontros intergrupais podem ser o motivo pelo qual o grupo usa a área com
espécies alóctones, visto que após todas ocorrências os animais abandonaram a área de mato,
retornando após algumas horas ou somente no dia posterior. Chiarello (1995) afirma que os
bugios são animais territoriais, que defendem ativamente sua área de uso através de

76
vocalizações, principalmente pelos machos adultos. Ludwig (2006), estudando A. caraya
observou 13 encontros intergrupais durante 12 meses de monitoramento.
Neville et al. (1988) citaram que os primatas do gênero Alouatta possuem grupos
que defendem com rigor suas áreas, principalmente através de vocalizações. Acredita-se
também que a ocorrência de coro matinal sirva para manter a distância entre os grupos,
evitando confrontos (Cunha, 1994). Este comportamento não foi observado pelo grupo focal
e, raramente, pelos grupos que habitam a mata de estudo. O macho alfa do grupo de estudo
vocalizou predominantemente em situações que necessitassem reunir o grupo ou inicialmente,
na presença dos observadores, sendo nos encontros intergrupais a vocalização quase
imperceptível.
Segundo Jardim (2005), um aspecto relevante que deve influenciar os tamanhos de
área de vida são as relações de dominância/subordinação entre grupos adjacentes. É razoável
supor que grupos dominantes de primatas ocupem as melhores porções da floresta,
restringindo grupos subordinados a áreas menos favoráveis. Entretanto, a pressão exercida
pelos grupos dominantes não significa que estes vão ocupar áreas maiores, pois a distribuição
dos recursos importantes é variável e grupos em áreas com menor disponibilidade de recursos
podem ser obrigados a percorrer áreas maiores para encontrar alimentos, aumentando
sobreposição de área com outros grupos ou expandindo para áreas onde não tenham grupos de
bugios estabelecidos. De forma semelhante a história de vida dos grupos deve influenciar,
pois grupos com formação mais recente devem ocupar áreas marginais e possivelmente
menos favoráveis.
Isso vem ao encontro das observações realizadas no presente estudo, que sugerem que
o grupo focal seja um grupo novo, pressionados pelo grupo dominante a utilizar uma área de
baixa qualidade, necessitando, inclusive, adaptar sua dieta ao espaço que lhe é disponível. Os
animais utilizaram diariamente, durante seu deslocamento entre a área com espécies alóctone

77
a mata, árvores de baixo porte e marginais, expondo os animais à proximidade com os cães da
propriedade e a presença humana.
Na Figura 27 pode-se visualizar o uso do espaço nas diferentes estações do ano.
Durante o verão os animais passaram boa parte do tempo na linha de espécies alóctones,
utilizando uma parte da área de mata durante os períodos de alimentação, quando se
deslocavam até a quadrícula D18 para consumir frutos de Casearia decandra e frutos de Vitis
americana na quadrícula D16.
Deve-se levar em consideração que o uso da mata foi maior do que o apresentado
neste trabalho, já que houve menos tempo de observações no período do início do verão. A
quadrícula B5 foi muito utilizada como sítio de pernoite. A quadrícula B11 foi muito utilizada
nos períodos de alimentação, visto que nela há uma grande árvore de Hovenia dulcis.
No outono, a área de vida foi ampliada após o corte dos Pinus, passando a utilizar com
mais freqüência a área de mato, inclusive como sítio de pernoite. Nesta época as árvores
utilizadas como dormitório eram as araucárias, as quais estavam na época de maturação das
pinhas, utilizadas para alimentação. Tanto, no outono quanto no verão, os animais iam para a
mata em busca de árvores de Hovenia dulcis, nas quais consumiam grande quantidade de
frutos. Segundo Chapman (1988), uma única árvore em frutificação pode fortemente
influenciar o modo de uso do espaço por primatas.
No inverno, a área foi novamente incrementada, devido a mudanças na dieta. Algumas
quadrículas passaram a ser utilizadas devido ao consumo de frutos de Eryobotrya japonica
(ex. quadricula B17). É importante ressaltar que nesta estação houve a maior diversidade de
consumo de espécies, estando esta interligada a um uso diferenciado da mata, propiciando
uma melhor diversidade na oferta de alimentos. Nesta estação os animais utilizaram a área
com Eucalyptus, inclusive como dormitório, mas não puderam mais utilizar a área com Pinus,
devido à dificuldade de locomoção entre as poucas árvores que restaram após o corte.
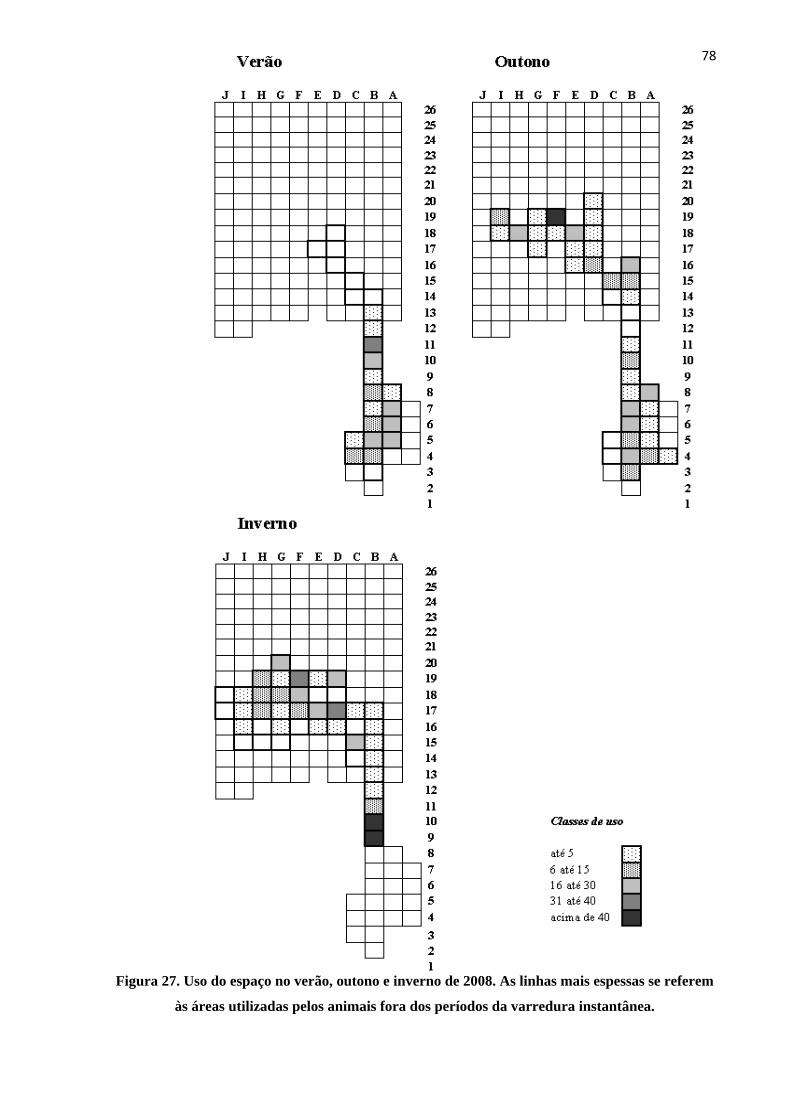
78
Figura 27. Uso do espaço no verão, outono e inverno de 2008. As linhas mais espessas se referem
às áreas utilizadas pelos animais fora dos períodos da varredura instantânea.

79
Segundo Spironello (2001) o tamanho da área de vida e modo de uso do espaço entre
os primatas pode depender de aspectos sociais e de estratégias alimentares comportamentais.
Estas podem mudar de acordo com a disponibilidade de recursos alimentares em diferentes
áreas de floresta ao longo do tempo (Cullen Jr. & Valladares-Padua, 1997; Jardim, 2005;
Ludwig, 2006), de lugares adequados para o descanso e, ainda, de fontes de água (Altmann,
1974).
A área de vida também é relacionada com o tamanho do grupo e inversamente à
porcentagem de folhas na dieta dos primatas, ou seja, animais frugívoros possuem maiores
áreas de vida do que os que dependem de folhas, recurso este, geralmente mais abundante e
de distribuição mais homogênea (Milton & May, 1976; Chiarello, 1992, 1993).
O Coeficiente de Variação (C.V.) da área de vida pode ser visualizado na Tabela 8.
Apesar do C.V. no inverno ser maior que os das demais estações, não houve diferenças
significativas pelo teste de Mann-Whitney-U. Entretanto, verificando os mapas com as
distribuições das observações dos bugios, podemos perceber um padrão diferenciado de uso
entre as estações.
Durante o período de estudo observou-se que o grupo utilizou 52 quadrados, o que
corresponde a 3,25 ha (área de vida total). A área de vida média foi de 0,9 ha, equivalendo a
0,3 ind/ha. A área de vida média do grupo foi de 0,73 ha no verão, 0,92 ha no outono e 1,04
ha no inverno (Tabela 8). Estes dados discordam do trabalho de Chiarello (1992), que obteve
uma maior área de vida no verão e a menor área no inverno. Ludwig (2006) também observou
uma área de vida maior nas estações de primavera e verão, do que outono e inverno.
Entretanto, em Floresta Ombrófila Mista, Miranda (2004) registrou um aumento nas
áreas de vida no outono e depois no inverno, em relação ao verão. O autor acredita que este

80
aumento na área esteja relacionado à escassez e agrupamento dos itens sazonais durante estas
estações. O aumento na área não acarretou o aumento nos percursos diários.
Estas diferenças na área de vida do grupo em estudo em relação a outros grupos
estudados possivelmente estão relacionadas a uma adaptação do grupo a uma nova área de
vida, após o corte dos Pinus. A intervenção de manejo florestal pode ter causado o incremento
do tamanho da área de uso no inverno em relação ao verão.
Tabela 8. Área de vida e coeficiente de variação do uso da área durante as estações de
verão, outono e inverno de 2008.
Estações Área de vida (ha) C. V. da área de vida
Verão 0,73 0,76
Outono 0,92 1,07
Inverno 1,04 1,22
Média anual 0,9 1,02
Total anual 3,25
Em comparação aos dados de área de vida de outras pesquisas já realizadas com
Alouatta guariba, o presente trabalho obteve o segundo menor valor de área de vida já
registrado (Tabela 9), sendo o menor valor de área de vida (2,4 ha) documentado por Jardim
(2005). Entretanto, deve-se levar em consideração que o tamanho do grupo é um fator
determinante no tamanho de área de vida (Milton & May, 1976), tendo o grupo focal o menor
tamanho em relação às demais pesquisas, além do fragmento ser o menor entre os demais.
Pequenas áreas de vida aumentam as chances das espécies sobreviverem em florestas
fragmentadas (Crockett, 1998). Segundo Bravo & Sallenave (2003), pequenas áreas de vida
podem estar relacionadas com uma dieta composta por recursos distribuídos de forma
abundante e uniforme no tempo e no espaço, com alto retorno energético. Como no presente

81
estudo os animais utilizaram como fonte alimentar plantas alóctones que disponibilizaram
frutos durante boa parte do ano (Vitis americana, Quercus robur, Hovenia dulcis, Diospyrus
kaki, Eryobotrya japonica, Eucalyptus sp.) é possível que haja uma relação entre a pequena
área de uso e o aporte energético fornecido por estas plantas, próximas aos locais de dormida.
É sugerido que os bugios precisem de áreas maiores em ambientes com uma menor
riqueza de espécies. Na Floresta Ombrófila Mista, um ambiente com menor riqueza quando
comparado a outros biomas ocupados pelos A.g.clamitans, os bugios apresentem áreas de vida
maiores que em ambientes fragmentados de tamanho aproximado (Perez, 1997; Miranda,
2004). Trabalhos realizados em Mata de Araucária documentam valores superiores de área de
vida do que encontrado nesse trabalho, com valores entre 9,2 ha a 16,75 ha (Marques, 1996;
Perez, 1997; Miranda, 2004).
Outro fator determinante para área de vida é a qualidade do ambiente. Em teoria, a
floresta primária alterada seria melhor para os bugios que a floresta secundária, sendo que os
grupos viventes na segunda deveriam ter áreas de vida maiores para compensar uma menor
qualidade ambiental (Dunbar, 1988 apud Miranda, 2004). Na prática, não foi corroborada essa
hipótese, haja visto que os animais não apresentaram uma área de vida maior mesmo em
ambiente de baixa qualidade.
Menores áreas de vida estariam relacionadas com um maior consumo de folhas.
Apesar da área de vida reduzida no presente estudo (3,25 ha), o consumo de folhas (64%)
esteve próximo aos valores encontrados para a espécie (entre 50% a 80%) (Mendes, 1989;
Lunardelli, 2000, Jardim & Oliveira, 2000; Aguiar et al., 2003; Miranda & Passos, 2004;
Alves & Zaú, 2007).

82
Em contrapartida, a pequena área de uso poderia estar relacionada a uma minimização
de gastos energéticos, com a diminuição das distâncias percorridas nos deslocamentos. Milton
(1980) classifica os bugios como “minimizadores de viagem”. Miranda (2004) propõe que
numa floresta em regeneração os bugios não tenham necessidade de se deslocar mais para
obtenção de recursos. No que se refere ao percurso diário, existiria uma equivalência em
qualidade de habitat para a espécie. Bicca-Marques (2003) mostra que os percursos diários
não se alteram entre diferentes tamanhos de fragmento.
O tamanho da área de vida possui correlação negativa com a densidade populacional.
Desta forma, análises de densidade seriam importantes para um melhor entendimento da
situação. Porém, durante o estudo foi observado em varias ocasiões dois grupos além do
grupo focal, um com três indivíduos e outro com quatro. Estes grupos habitam a área com
araucárias, um na região norte da mata e outro à noroeste. Em duas ocasiões foram avistados
dois outros grupos, com um maior numero de indivíduos, mas é possível que estes animais
estivessem somente se deslocando pela área já que não foram mais avistados.
Supondo que a área abrigue os três grupos mais avistados, resultaria em uma
densidade de 0,83 ind/ha. Este valor está dentro da variação registrada para o gênero e para a
espécie (Cunha, 1994), mas superior ao encontrado em Floresta Ombrófila Mista (0,38 ind/ha)
por Miranda (2004). Entretanto, a densidade normalmente é maior em fragmentos florestais
menores (Steinmetz, 2001).
No presente estudo é mais provável que a pequena área de vida seja decorrente do
forte territorialismo do grupo vizinho, que força o grupo focal a utilizar uma área menor para
evitar confrontos e pelo pequeno tamanho do fragmento. Caso a linha de árvores alóctones
fosse conectada a uma outra área de mato, é plausível supor que os animais utilizariam os
Pinus e Eucalyptus como uma espécie de corredor ecológico e ampliariam a sua área de vida.
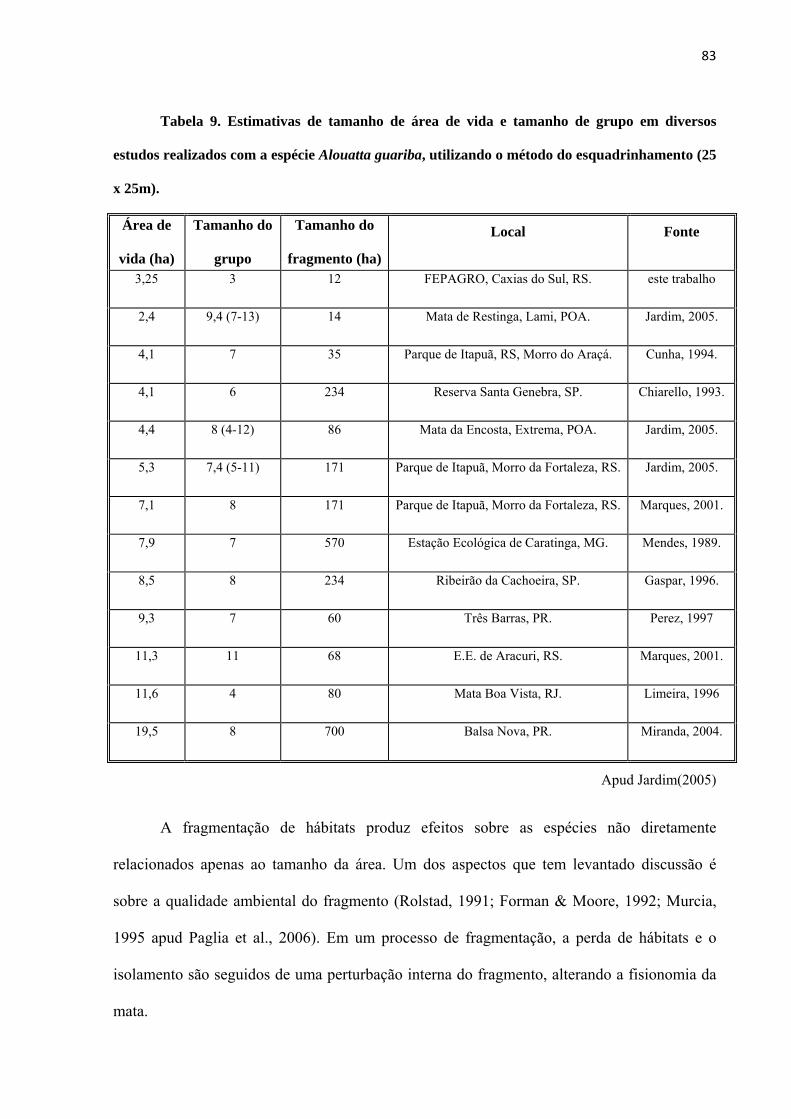
83
Tabela 9. Estimativas de tamanho de área de vida e tamanho de grupo em diversos
estudos realizados com a espécie Alouatta guariba, utilizando o método do esquadrinhamento (25
x 25m).
Área de
vida (ha)
Tamanho do
grupo
Tamanho do
fragmento (ha)
Local Fonte
3,25 3 12 FEPAGRO, Caxias do Sul, RS. este trabalho
2,4 9,4 (7-13) 14 Mata de Restinga, Lami, POA. Jardim, 2005.
4,1 7 35 Parque de Itapuã, RS, Morro do Araçá. Cunha, 1994.
4,1 6 234 Reserva Santa Genebra, SP. Chiarello, 1993.
4,4 8 (4-12) 86 Mata da Encosta, Extrema, POA. Jardim, 2005.
5,3 7,4 (5-11) 171 Parque de Itapuã, Morro da Fortaleza, RS. Jardim, 2005.
7,1 8 171 Parque de Itapuã, Morro da Fortaleza, RS. Marques, 2001.
7,9 7 570 Estação Ecológica de Caratinga, MG. Mendes, 1989.
8,5 8 234 Ribeirão da Cachoeira, SP. Gaspar, 1996.
9,3 7 60 Três Barras, PR. Perez, 1997
11,3 11 68 E.E. de Aracuri, RS. Marques, 2001.
11,6 4 80 Mata Boa Vista, RJ. Limeira, 1996
19,5 8 700 Balsa Nova, PR. Miranda, 2004.
Apud Jardim(2005)
A fragmentação de hábitats produz efeitos sobre as espécies não diretamente
relacionados apenas ao tamanho da área. Um dos aspectos que tem levantado discussão é
sobre a qualidade ambiental do fragmento (Rolstad, 1991; Forman & Moore, 1992; Murcia,
1995 apud Paglia et al., 2006). Em um processo de fragmentação, a perda de hábitats e o
isolamento são seguidos de uma perturbação interna do fragmento, alterando a fisionomia da
mata.

84
Segundo Limeira (1996), a fragmentação de habitat cauda perda de espécies preferidas
por A. g. clamitans, além de poder tornar a dieta mais fibras e/ou tóxica, com altas
concentrações de compostos secundários.
Outro efeito da fragmentação é o chamado efeito de borda, que pode resultar em um
aumento do fluxo de energia e a entrada de componentes químicos pela borda, afetando a
composição e a distribuição das espécies. O aumento no fluxo de radiação na borda implica
rápido crescimento de lianas, trepadeiras e outras espécies ao redor da borda da floresta
remanescente (Paglia et al., 2006).
Na área de estudo observa-se o efeito de borda agindo favorável à expansão das
espécies alóctones na orla do fragmento, principalmente o Ligustrum lucidum. As lianas
também estão bastante presentes no fragmento e compõem diariamente a dieta dos bugios.
Segundo Bicca-Marques (2003), a fragmentação de habitat resulta num decréscimo da
área de vida, especialmente em pequenos fragmentos. O tamanho do fragmento prediz a área
de vida, o numero de plantas usadas como recurso alimentar e a diversidade de folhas e frutas
na dieta. Em contrapartida, não pode predizer a área de vida diária, a contribuição de
diferentes itens alimentares da dieta, e o orçamento de atividades. Além disso, o tamanho do
grupo não pode predizer o tamanho da área de vida, exceto para A. palliata.
Bravo & Sallenave (2003) demonstram em seus estudos que áreas de vida muito
pequenas somente são possíveis em locais onde haja a disponibilidade de recursos de alto
valor energético. No presente estudo, o consumo de frutos de Hovenia dulcis e Eryobotrya
japonica devem fornecer um alto aporte energético para os animais.
Juan et al. (2000), estudando grupos de A. palliata vivendo em fragmentos de 3, 35 e
250 ha em Los Tuxtlas, México, observaram que em fragmentos maiores os animais

85
utilizaram um maior numero de espécies como fonte de alimento, consumiram mais frutas do
que folhas e tiveram um tempo maior dedicado ao deslocamento do que nos fragmentos
menores.
Este trabalho vai ao encontro com Bicca-Marques (2003), que conclui que os primatas
do gênero Alouatta lidam com a fragmentação sem mostrar mudanças direcionais na maioria
dos aspectos da sua ecologia alimentar e comportamental. Sua habilidade para conviver com
uma variável composição florística se deve a uma dieta flexível, composta por uma ampla
variedade de plantas, incluindo espécies exóticas, sendo isto especialmente relevante para o
seu sucesso em florestas fragmentadas.

86
5. CONCLUSÕES
A partir das observações realizadas neste trabalho foi possível verificar que o grupo de
bugio-ruivo A. g. clamitans estudado demonstrou alta flexibilidade alimentar e
comportamental em um ambiente fragmentado e alterado ao longo dos anos, com a introdução
e dispersão de espécies alóctones.
O grupo utilizou como alimento muitas destas espécies, tais como Pinus spp.
Eucalyptus spp., Hovenia dulcis, Eryobotria japonica, Quercus robur, Diospyrus kaki,
Ligustrum lucidum e Vitis americana. A inclusão de frutos de espécies alóctones à dieta
sugere que o gênero Alouatta seja tão frugívoro quanto possível em termos de ambiente.
Além disso, aumentou o tempo de descanso, possivelmente para metabolização de
compostos secundários, minimização de gastos energéticos ou para evitar confrontos com os
grupos vizinhos e diminuiu a área de vida, em comparação aos demais trabalhos realizados
com a espécie.
O grupo focal foi capaz de ajustar a área de vida passando a utilizar a área com
espécies alóctones, possivelmente devido ao territorialismo do grupo que habita a mata
secundária, e que pressiona o grupo focal a sair dessa área.
A flexibilidade do grupo foi fortemente demonstrada após a supressão de parte da sua
área de uso (área plantada com Pinus spp.), já que o grupo passou a utilizar uma nova área
dentro da mata secundária, retornado a área que restou após o corte das árvores (área com
Eucalyptus sp.), quando havia confrontos com o grupo vizinho.
Entretanto, apesar do bugio-ruivo poder incluir espécies alóctones à sua dieta, é
importante que se protejam as matas nativas e, que, se possível, haja um enriquecimento
destas matas com espécies autóctones, importantes para o bugio-ruivo. Possivelmente os

87
animais do grupo de estudo estejam utilizando novas fontes de alimentos por haver um déficit
de itens alimentares com alto valor energético no interior da mata.
Ainda, mesmo não havendo pressão de caça na localidade e os animais estarem
habituados à presença humana, isso não garante a conservação da população local à longo
prazo, já que área é fragmentada e a proximidade com animais domésticos (cães e galinhas)
geram riscos à saúde e a integridade dos animais.

88
6. REFERÊNCIAS BIBLIOGRÁFICAS
Aguiar, L.M.; Reis, N.R.; Ludwig, G. & Rocha, V.J. (2003). Dieta, área de vida, vocalizações e estimativas populacionais de Alouatta guariba em um remanescente florestal no Norte do estado do Paraná. Neotropical Primates. 11: 78-86.
Albuquerque, V.J. de & Codenotti, T.L. (2006). Etograma de um grupo de bugios-pretos, Alouatta caraya (Humboldt, 1812) (Primates, Atelidae) em um habitat fragmentado. Revista de Etologia. 8: 97-107.
Almeida, S.B; Guedes, P.G.; Boubli, J.P. & Strier, K.B. (2005). Deslocamento terrestre e o comportamento de beber em um grupo de barbados (Alouatta guariba clamitans Cabrera, 1940) em Minas Gerais, Brasil. Neotropical Primates. 13: 1–3.
Almeida-Silva, B.; Guedes, P.G.; Boubli, J.P. & Strier, K.B. (2005). Deslocamento terrestre e o comportamento de beber em um grupo de barbados (Alouatta guariba clamitans Cabrera, 1940) em Minas Gerais, Brasil. Neotrop Primates. 13: 1–3.
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour. 49: 227-267.
Altmann, S.A. (1959). Field observations on howling monkey society. J. Mamm. 40: 317-330.
Alves, S.L. & Zaú, A.S. (2005). A importância da Área de Relevante Interesse Ecológico Floresta da Cicuta (RJ) na conservação do bugio-ruivo (Alouatta guariba clamitans Cabrera, 1940). Rev. Univ. Rural. 25: 41-48.
Alves, S.L. & Zaú, A.S. (2007). Aspectos ecológicos de Alouatta guariba clamitans Cabrera, 1940 na Área de Relevante Interesse Ecológico Floresta da Cicuta, Rio de Janeiro, Brasil. Neotropical Primates. 14: 127-130.
Andreiv, J. & Firkowski, C. (2006). Técnicas de redução de danos causados por roedores em povoamentos de Pinus. Floresta. 36: 305-310.
Andresen, E. (2006). Primary seed dispersal by red howler monkeys and the effect of defecation patterns on the fate of dispersed seeds. Biotropica. 34: 261-272.
Auricchio, P. (1995). Primatas do Brasil. São Paulo: Terra Brasilis, 168p.
Bicca-Marques, J.C & Calegaro-Marques, C. (1994). Feeding behaviour of the black howler monkey (Alouatta caraya) in a seminatural forest. Acta Biologica Leopoldensia. 1: 64-84.
Bicca-Marques, J.C. (1990). A new southern limit for the distribution of Alouatta caraya in Rio Grande do Sul State, Brazil. Primates. 31: 449-451.
Bicca-Marques, J.C. (2003). How do howler monkeys cope with habitat fragmentation? In: Marsh, L.K. (Ed.). Primates in Fragments: Ecology and Conservation. New York: Kluwer Academic/Plenum Publishers, p283-303.

89
Bicca-Marques, J.C.; Silva, V.M. da; Gomes, D.F. (2006). Ordem Primates. In: Mamíferos do Brasil, cap. 5. p101-148.
Bonvicino, C.R. (1989). Ecologia e comportamento de Alouatta belzebul (Primates: Cebidae) na mata atlântica. Revista Nordestina de Biologia. 6: 149-179.
Bravo, S.P. & Sallenave, A. (2003). Foraging behavior and activity patterns of Alouatta caraya in the Northeastern Argentinean Flooded Forest. International Journal of Primatology. 24: 825-846.
Bravo, S.P.; Kowalewski, M.M. & Zunino, G.E. (1995). Dispersion y germinacion de semillas de Ficus monckii por el mono aullador negro (Alouatta caraya). Bol. Primatol Lat. 5: 25–27.
Buss, G. (2001). Densidade populacional do bugio-ruivo nas formações florestais do morro do Campista, Parque Estadual de Itapuã. Dissertação de mestrado. Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil, 76p.
Bussche, G.H. & Zee, D. (1985). Damage by samango monkeys, Cercopithecus (mitis) albogularis to Pine trees in the Northern Transvaal. South African Forestry Journal. p43-48.
Cabrera, A. (1940). Los nombres científicos de algunos monos americanos. Ciencia. 9: 402-405.
Cabrera, A. (1957). Catalogo de los mamiferos de America del Sur. Museo Argentino de Ciencias Naturales Bernardino Rivadavia e Instituto Nacional de Investigación de las Ciencias Naturales. 4: 1-307.
Calegaro-Marques, C. (1992). Comportamento social de um grupo de Alouatta caraya (Primates, Cebidae) em Alegrete, RS, Brasil. Dissertação de mestrado. Universidade de Brasília, DF, Brasil.
Carpenter, C.R. (1934). A field study of behavior and social relations of howling monkeys. Comp. Psychol. Monogr. 48: 1-168.
Carvalho, D.R.J. de (2007). Predação em Pinus spp. por Cebus nigritus (Goldfuss, 1809) (Primates:Cebidae) na região nordeste do Paraná - Brasil. Dissertação de mestrado. Universidade Federal do Paraná, Curitiba, PR, Brasil. 74 p.
Chapman, C.A. (1987). Flexibility in diets oh three species of Costa Rican Primates. Folia Primatologica. 49: 90-105.
Chapman, C.A. (1988). Patterns of foraging and range use by three species of Neotropical primates. Primates. 29: 177-194.
Chapman, C.A. & Balcomb, S.R. (1998). Population characteristics of howlers: ecological conditions or group history. Int. J. Primatol. 19: 385-403.
Chapman, C.A. (1989). Primate seed dispersal: the fate of dispersed seeds. Biotropica. 21: 148-154.

90
Chiarello, A.G. & Galetti, M. (1994). Conservation of the brown howler monkey in southeast Brazil. Oryx. 28: 37-42.
Chiarello, A.G. (1992). Dieta, padrão de atividades e área de vida de um grupo de bugios (Alouatta fusca), na reserva de Santa Genebra, Campinas-SP. Dissertação de mestrado, Universidade Estadual de Campinas, Campinas, SP, Brasil. 80p.
Chiarello, A.G. (1994). Diet of the brown howler monkey Alouatta fusca in a semi-deciduous forest fragment of southeastern Brazil. Primates. 35: 25-34.
Chiarello, A.G. (1995). Role of loud calls in brown howlers, Alouatta fusca. American Journal of Primatology. 36: 213-222.
Chitolina, O.P. & Sander, M. (1981). Contribuição ao conhecimento da alimentação de Alouatta guariba clamitans (Cabrera, 1940) em habitat natural no Rio Grande do Sul (Cebidae, Alouattinae). lheringia, Sér. Zoologia, 59: 37-44.
Clutton-Block, T.H. & Harvey, P.H. (1977). Species differences in feeding and ranging behavior in primates. In: Clutton-Brock, T.H. (Ed). Primate ecology: studies of feeding and ranging behaviour in lemurs, monkeys and apes. London: Academic Press. p557-584.
Clutton-Brock, T.H. (1977). Some aspects of intraspecific variation in feeding and ranging behaviour in primates. In: Clutton-Brock, T.H. (Ed). Primate ecology: studies of feeding and ranging behaviour in lemurs, monkeys and apes. London: Academic Press. 539-556p.
Crespo, J.A. (1954). Presence of the reddish howling monkey (Alouatta guariba clamitans Cabrera) in Argentina. Journal of Mammalogy. 35: 117–118.
Crockett, C.M. & Eisenberg, J.F. (1987). Howlers: Variations in group size and demography. In: Smuts, B.B.; Cheney, D. L.; R. M. Seyfarth, R.W. Wrangham & Struhsaker, T.T. (Eds.) Primates Societies. The University of Chicago Press, Chicago and London. 54-68p.
Crockett, C.M. (1998). Conservation Biology of howler. Int. J. Primatol. 19: 549-578.
Cullen Jr., L. & Valladares-Padua, C.B. (1977). Métodos para estudos de ecologia, manejo e conservação de primatas na natureza. Pp. 239-269. In: Padua, C.V.; Bodmer, R. & Cullen, L. (eds.). Manejo e Conservação de Vida Silvestre no Brasil. Brasília, Sociedade Civil Mamirauá – Cnpq. 296p.
Cunha, A.S. (1994). Aspectos sócio-ecológicos de um grupo de bugios (Alouatta fusca clamitans) do Parque Estadual de Itapuã, RS. Dissertação de mestrado, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil. 75p.
Damerose, E. & Hopkins, W.D. (2002). Scan and focal sampling: Reliability in the laterality for maternal cradling and infant nipple preferences in olive baboons, Papio anubis. Animal Behaviour. 63: 511-518.
Defler, T.R. (2004). Primates of Colombia. Colombia: Conservation International, 550p.

91
Eisenberg, J.F. & Muckenhirn, N.A. & Rudran, R. (1972). The relation between ecology and social structure in primates. Science. 176: 863-874.
Estrada, A. & Coates-Estrada, R. (1986). Frugivory by howling monkeys (Alouatta palliata) at Los Tuxtlas, Mexico: dispersal and fate of seeds. In: Estrada, A. & Fleming, T. H. (Eds.). Frugivores and seed dispersal. Dr. W. Junk Publishers. Dordrecht/Boston. p93-104.
Estrada, A. (1984). Resource use by howler monkey (Alouatta palliata) in the rain Forest of Los Tuxtlas, Vera Cruz, México. International Journal of Primatology. 5: 105-131.
Estrada, A. (2007). Primate conservation in fragmented neotropical landscapes: human dimension of the problem and conservation value of agroecosystems. In: Bicca-Marques, J.C. (Ed). A Primatologia no Brasil. 10: 37-69.
Estrada, A.; Juan-Solano, S.; Martínez, T.O. & Coates-Estrada, R. (1999). Feeding and general activity patterns of a howler monkey (Alouatta palliata) troop living in a Forest fragment at Los Tuxtlas, México. Am. J. Primatol. 48: 167-183.
Fialho, M.S. (2000). Ecologia do Alouatta fusca em Floresta de Encosta e de Restinga no Sul do Brasil. Dissertação de mestrado. Universidade Estadual de Campinas, Campinas, SP, Brasil. 171p.
Fortes, V. B. & Bicca-Marques, J.C. (2005). Ecologia e comportamento de primatas: métodos de estudo de campo. In: Métodos de estudo em biologia. Caderno La Salle XI. 2: 207-218.
Fortes, V. B. (1999). Dieta, atividades e uso do espaço por Alouatta fusca (Geoffroy, 1812, Primates: Cebidae), na Depressão Central do Rio Grande do Sul. Dissertação de mestrado. Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil. 122p.
Franceschinelli, E.D.; Almeida, E.A.B.; Antonini, Y., Cabral, B.C.; Carmo R.M. do; Damasceno, A.; Fontenelle, J.C.R.; Garcia, V.L.A.; Guilherme, M.S.; Laps, R.R.; Leitão, G.G.; Mikich, S.B.; Moreira, D.L.; Nascimento, M.T.; Nemésio, A.; Ribon, R.; Silveira, F.A. & Vidigal, T.H.D.A. (2005). Interações entre animais e plantas. 276-295p. In: Fragmentação de Ecossistemas: causas, efeitos sobre a diversidade e recomendações de políticas públicas. Ministério do Meio Ambiente. Brasília, DF. 508p.
Freitas, H.F. de; Setz, E.Z.F.; Araújo, A.R.B. & Gobbi, N. (2008). Agricultural crops in the diet of bearded capuchin monkeys, Cebus libidinosus Spix (Primates: Cebidae), in forest fragments in southeast Brazil. Revista Brasileira de Zoologia. 25: 32-39.
Fuentes, E.; Estrada, A.; Franco, B.; Magãna, M.; Decena, Y.; Muñoz, D. & García, Y. (2003). Reporte preliminar sobre el uso de recursos alimenticios por una tropa e mono aulladores, Alouatta palliata, en el Parque La Venta, Tabasco, México. Neotropical Primate. 11: 24-29.
Galetti, M. & Pedroni, F. (1994). Seasonal diet of capuchin monkeys (Cebus apella) in a semideciduous Forest in south-east Brazil. Journal of Tropical Ecology. 10: 27-39.
Garber, P.A. (1987). Foraging strategies among living primates. Annu. Rev. Anthrop. 16: 339-364.

92
Gautier-Hion, A.; J.P. Gautier & R. Quris. (1981). Forest structure and fruit availability as complementary factors influencing habitat use by a troop of monkeys (Cercopithecus cephus). Terre Vie. 35: 511-536.
Gilbert, K.A. & Setz E.A. (2001). Primates in a fragmented landscape: six species in Central Amazonia. In: Bierregaard Jr, R. O.; Gascon, C.; Lovejoy, T.E.; & Mesquita, R. (Ed). Lessons from Amazonia: the Ecology and Conservation of a Fragmented Forest. Yale University Press: New Haven. p. 262-270.
Glander, K.E. (1975). Habitat description and resource utilization: a preliminary report on mantled howling monkey ecology. In: Tuttle, R.H. (Ed.). Socioecology and Psychology of Primates. The HaHague, Mouton. p. 37-57.
Gonçalves, G.L.; Faria-Correa, M.A.; Cunha, A.S. & Freitas, T.R.O. (2007). Bark consumption by the spiny rat Euryzygomatomys spinosus (G. Fischer) (Echimyidae) on a Pinus taeda Linnaeus (Pinaceae) plantation in South Brazil. Revista Brasileira de Zoologia. 24: 260–263.
Gregorin, R. (2006). Taxonomia e variação geográfica das espéciesdo gênero Alouatta Lacépède (Primates, Atelidae) no Brasil. Revista Brasileira de Zoologia. 23: 64-144.
Hirsch, A.; Landau; E.C; Tedeschi, A.C. & Menegueti, J.O. (1991). Estudo comparativo das espécies do gênero Alouatta Lacèpéde, 1799 (Platyrrhini, Atelidae) e sua distribuição geográfica na América do Sul. In: Rylands, A.B. & Bernardes, A.T. (Eds). A Primatologia no Brasil. 3: 239-262.
Hladik, A.; Hladik, C.M. (1969). Rapports trophiques entre végétation et primates dans la forêt de Barro Colorado (Panamá). Terre et Vie. 1: 25-117.
Ihering, H.V. (1914). Os bugios do gênero Alouatta. Revista do Museu Paulista. 9: 231-280.
Ingberman, B. (2007). Análise populacional de Alouatta clamitans Cabrera, 1940 no Parque Estadual Ilha do Cardoso, litoral sul do estado de São Paulo, Brasil. Dissertação de mestrado. Universidade Federal do Paraná, Curitiba, PR, Brasil. 51p.
IUCN (2007). 2007 IUCN Red List of Threatened Species. <www.iucnredlist.org>. (Acessado em: 01 de abril de 2008).
Izar, P. (1999). Aspectos de ecologia e comportamento de um grupo de macacos-prego (Cebus apella) em área de Mata Atlântica, São Paulo. Tese de doutoramento. Universidade de São Paulo, São Paulo, São Paulo, Brasil. 144p.
Jardim, M.M.A. (1997). Estratégias de forrageamento e uso do espaço por Alouatta belzebul (PRIMATES, CEBIDAE) na Estação Científica Ferreira Penna, Melgaço, Pará. Dissertação de mestrado, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil. 121p.
Jardim, M.M.A. (2005). Ecologia populacional do bugio-ruivo nos municípios de Porto Alegre e Viamão, Rio Grande do Sul, Brasil. Tese de doutorado. Universidade Estadual de Campinas, Campinas, São Paulo, Brasil. 114p.

93
Jardim, M.M.A. & Oliveira, L.F.B. (2000). Aspectos ecológicos e do comportamento de Alouatta fusca (Geoffroy, 1812) na Estação Ecológica de Aracuri, RS, Brasil. In: Alonso, C & Langguth, A. (Eds.). A Primatologia no Brasil. 7: 151-169.
Jardim, M.M.A. (1992). Aspectos ecológicos e comportamentais de Alouatta fusca clamitans (Cabrera, 1940) na Estação Ecológica de Aracuri, RS, Brasil (Primates, Cebidae). Monografia de bacharelado. Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil. 70p.
Jerusalinsky, L. (2001). Diversidade em seqüências mitocondriais do bugio-ruivo (Alouatta guariba): implicações para a história evolutiva e a conservação da espécie. Dissertação de mestrado. Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil. 80p.
Jolly, A. (1972). The Evolution of Primate Behavior. Second edition. New York: Macmillan Publishing Company.
Jolly, A. (1985). The evolution of primate behavior. 2nd edition, New York: Macmillan Publishing Company.
Jordano, P.P.; Galetti, M.; Pizo, M.A. & Silva, W.R. (2006). Ligando frugivoria e dispersão de sementes à biologia da conservação. In: Duarte, C.F.F.; Bergallo, H.G.; Santos, M. A. dos (Eds.). Biologia da conservação: essências. São Paulo: Editorial Rima. p411-436.
Juan, S., Estrada, A. & Coates-Estrada, R. (2000). Contrastes y similitudes en el uso de recursos y patrón general de actividades en tropas de monos aulladores (Alouatta palliata) en fragmentos de selva en Los Tuxtlas, México. Neotropical Primates. 8: 131-135.
Julliot, C. (1996). Seed dispersal by red howling monkeys (Alouatta seniculus) in the tropical rain forest of French Guiana. Inter. J. of Primatol. 17: 239-258.
Julliot, C. (1997). Impact of seed dispersal by howler monkeys Alouatta seniculus on the seedling population in the understorey of a tropical rain forest. Journal of Ecology. 85: 431-440.
Koch, F. (2004). Padrão de atividades diárias, comportamento alimentar e área de uso de um grupo de bugios-ruivos (A. g. clamitans) no sul do Brasil. Monografia de bacharelado. Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil.
Koehler, A. & Firkowski, C. (1996). Descascamento de Pinus por macaco-prego (Cebus apella). Floresta. 24: 61-75.
Lemos de Sá, R.M.L. & Strier, K.B. (1992). A preliminary comparison of forest structure and use by two isolated groups of woolly spider monkeys, Brachyteles arachnoides Biotropica. 24: 455–459.
Liesenfeld, M.V.A. (2003). Ecologia da dispersão de sementes de maria-preta (Diospyros inconstans) pelo bugio-ruivo (Alouatta guariba) em uma mata subtropical de Porto Alegre, RS, Brasil. Dissertação de mestrado. Universidade Estadual de Campinas. Campinas, SP, Brasil. 80p.

94
Liesenfeld, M.V.A. (1999). Regeneração natural em fragmentos florestais de Porto Alegre e sua relação com o bugio-ruivo. Monografia de bacharelado. Departamento de Botânica, UFRGS, Porto Alegre, RS, Brasil.
Limeira, V.L.A.G. (1996). Comportamento alimentar, padrão de atividades e uso de espaço por Alouatta fusca (Primates, Platyrrhini) em um fragmento degradado de Floresta Atlântica no estado do Rio de Janeiro. Dissertação de Mestrado. Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brasil.
Ludwig, G.; Aguiar, L.M. & Rocha, V.J. (2005). Uma avaliação da dieta, da área de vida e das estimativas populacionais de Cebus nigritus (Goldfuss, 1809) em um fragmento florestal no norte do estado do Paraná. Neotropical Primates. 13: 12-18.
Ludwig, G.; Aguiar, L.M. & Rocha, V.J. (2006). Comportamento de obtenção de Manihot esculenta Crantz (Euphorbiaceae), mandioca, por Cebus nigritus (Goldfuss) (Primates, Cebidae) com uma adaptação alimentar em períodos de escassez. Revista Brasileira de Zoologia. 23: 888-890.
Lunardelli, M.C. (2000). Padrões de atividade e efeitos de compostos fenólicos na ecologia alimentar de um grupo de bugios-ruivos (Alouatta fusca) no Sudeste Brasileiro. Dissertação de mestrado. Universidade de São Paulo, SP, Brasil. 70p.
Machado, A.B.M.; Fonseca, G.A.B., Machado, R.B.; Aguiar, L.M.S. & Lins, L.V. (1998). Livro vermelho das espécies ameaçadas de extinção da fauna de Minas Gerais. Belo Horizonte, Fundação Biodiversitas. 605p.
Macnnab, B. K. (1963). Bioenergetics and the determination of home range size. The American Naturalist. 97: 133-140.
Marques, A. A. B. (1996). O bugio ruivo Alouatta fusca clamitans (Cabrera, 1940) (Primates, Cebidae) na Estação Ecológica de Aracuri, RS: Variações sazonais de forrageamento. Dissertação de mestrado, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, RS, Brasil.
Marques, A. A. B. (2001). Estratégias de uso do espaço por Alouatta guariba clamitans Cabrera, 1940 em habitats temperados e subtropicais no sul do Brasil. Tese de doutoramento. Universidade Federal de Minas Gerais. Belo Horizonte, MG, Brasil.
Marques, A.A.B. (2003). Primatas, p. 499-506. In: Fontana, C.S.; Bencke, G.A. & Reis, R.E. (Eds). Livro vermelho da fauna ameaçada de extinção no Rio Grande do Sul. Porto Alegre, EDIPUCRS, 632p.
Martins, C.S. (1997). Uso do habitat pelo bugio-ruivo, Alouatta fusca clamitans, em um fragmento florestal em Lençóis Paulista-SP. Dissertação de mestrado. Universidade Estadual de Campinas, Campinas, São Paulo, Brasil. 87p.
Mccann, C.; Williams-Guillen, K.; Koontz, F.; Roque-Espinoza, A.A.; Martinez-Sanchuez, J.C. & Koontz, C. (2003). Shade coffee plantations as wildlife refuge for mantled howler monkeys (Alouatta palliata) in Nicaragua. In: Marsh, L.K. (Ed.). Primates in Fragments: Ecology and Conservation New York: Kluwer Academic, p321-342.

95
Mendes, S.L. (1989). Estudo ecológico de Alouatta fusca (Primates: Cebidae) na Estação Biológica de Caratinga, MG. Revista Nordestina de Biologia. 6: 71-104.
Mendes, S.L. (1985). Uso do espaço, padrões de atividades diárias e organização social de Alouatta fusca (Primates, Cebidae) em Caratinga, MG. Dissertação de mestrado. Universidade de Brasília, Brasília. 70 p.
Milton, K. & May, M.L. (1976). Body weight, diet and home range area in primates. Nature. 259: 459-462.
Milton, K. (1978). Relation entre las estrategias empleadas en la busqueda de alimentos y la distribuicion y seleccion de estos en el caso del mono aullador (Alouatta palliata). Actas del IV Simposium International de Ecologia Tropical, Panamá: 3-16.
Milton, K. (1979). Factors influencing leaf choice by howler monkeys: A test of some hypotheses of food selection by generalist herbivores. Am. Nat. 114: 362-379
Milton, K. (1980). The foranging strategy of howler monkeys. New York: Columbia University Press, 165p.
Milton, K.; Van Soest, P. J. & Robertson, J. B. (1980). Digestive efficiencies of wild howler monkeys. Physiol. Zool. 53: 402-409.
Miranda, J. M. D. (2005). Dieta de Sciurus ingrami Thomas em Floresta com Araucária do Sul do Brasil. Revista Brasileira de Zoologia. 22: 1141-1145.
Miranda, J.M.D. & Passos, F.C. (2004). Hábito alimentar de Alouatta guariba (Humboldt) (Primates, Atelidae) em Floresta de Araucária, Paraná, Brasil. Revista Brasileira de Zoologia. 21: 821-826.
Miranda, J.M.D. (2004). Ecologia, densidade e conservação de Alouatta guariba clamitans Cabrera, 1940 em Floresta Ombrófila Mista no estado do Paraná, Brasil. Dissertação de mestrado. Universidade Federal do Paraná, Curitiba, Paraná, Brasil. 82p.
Miranda, J.M.D.; Bernardi, I.P., Abreu, K.C. & Passos, F.C. (2005). Predation on Alouatta guariba clamitans Cabrera (Primates, Atelidae) by Leopardus pardalis (Linnaeus) (Carnivora, Felidae). Revista Brasileira de Zoologia. 22: 793-795.
Moro-Rios, R.F.; Serur-Santos, C.S.; Miranda, J.M.D. & Passos, F.C. (2008). Obtenção de água por um grupo de Alouatta clamitans (Primates: Atelidae) em Floresta com Araucária: variações sazonais, sexo-etárias e circadianas. Revista Brasileira de Zoologia. 25: 558-562.
Nagy, K. & Milton, K. (1979). Energy metabolism and food consumption by wild howler monkeys (Alouatta palliata). Ecology. 60: 475-480.
Neville, M.K.; Glander, K.E.; Braza, F. & Rylands, A.B. (1988). The Howling Monkeys, Genus Alouatta, p. 349-453. In: Mittermeier, R.A.; Rylands, A.B.; Coimbra-Filho, A. & Fonseca, G.A.B. (Eds). Ecology and behavior of Neotropical Primates. Washington, 2, 610p.

96
Oates, J.F. (1977). The ecology and behaviour of the black and white colobus monkey (Colobus guereza ruppell) in East Africa. Tese de doutorado. Universidade de Londres.
Odum, E.P. (1983). Ecologia. Rio de Janeiro: Interamericana, 434p.
Oliveira, E.G.R. (2000). Tamanho de árvore e uso de recurso alimentar em um bando de bugios-ruivos (Alouatta fusca, Geoffroy, 1812) no Parque Estadual de Itapuã, Viamão, RS, Brasil. Monografia de bacharelado. Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil.
Paglia, A.P.; Fernandez, F.A.S. & Marco Jr, E.P. (2006). Efeitos da fragmentação de habitats: quantas espécies, quantas populações, quantos indivíduos, e serão eles suficientes? In: Rocha, C.F.D.; Bergallo, H.G.; Sluys, M.V. & Alves, EM.A.S. (Eds). Biologia da conservação: essências. Rima, São Carlos.
Palacios, E. & Rodriguez, A. (2001). Ranging pattern and use of space in a group of red howler monkey (Alouatta seniculus) in the Southeastern Colombian rainforest. Am J. Primatol. 55: 233-251.
Perez, D.M. (1997). Estudo ecológico do bugio-ruivo em uma floresta com araucária do sul do Brasil (Alouatta fusca, Ihering 1914-PRIMATES, ATELIDAE). Dissertação de mestrado. Universidade de São Paulo, SP, Brasil. 90p.
Perin, C. (2008). Projeto de translocação e revigoramento populacional de bugios, Alouatta caraya (Primates, Atelidae) no campus da Universidade de São Paulo, em Ribeirão Preto. Dissertação de mestrado.Universidade de São Paulo, Ribeirão Preto, SP, Brasil. 128p.
Pielou, E.C. (1975). Ecological diversity. New York.
Prates, H.M. (2007). Ecologia e comportamento de um grupo de bugios-pretos (Alouatta caraya) habitante de um pomar em Alegrete, RS, Brasil. Dissertação de mestrado. Pontifícia Universidade do Rio Grande do Sul, Porto Alegre, RS, Brasil.
Prates, J.C.; Gayer, S.M.P.; Kunz Jr., L.F. & Buss, G. (1990a). Feeding habitats of the brown howler monkey Alouatta guariba clamitans (Cabrera, 1940) (Cebidae, Alouatinae) in the ltapuã State Park- a preliminary report. Acta Biol. Leopoldensia. 12: 175-188.
Prates, J.C.; Kunz Jr., L.F. & Buss, G. (1990b). Comportamento postural e locomotor de Alouatta guariba clamitans (Cabrera, 1940) em floresta subtropical. Acta Biol. Leopoldensia. 12: 189-200.
Printes, R. C. & Malta, M.C.C. (2007). Translocação de duas fêmeas de bugio-preto (Alouatta caraya Humboldt, 1812) por ocasião da formação do lago da Hidrelétrica de Queimado, Minas Gerais, Brasil. A Primatologia no Brasil.10: 207-223.
Printes, R. C.; Liesenfeld, M. V. A. & Jerusalinsky, L. (2001). Alouatta guariba clamitans Cabrera, 1940: a new southern limit for the species and for neotropical primates. Neotropical Primates. 9: 118-121.
Queiroz, H. L. (1995). Preguiças e guaribas – os mamíferos folívoros arborícolas do Mamirauá. Sociedade Civil Mamirauá. Ed. Marigo. 161p.

97
Rasmussen, D.R. (1980). Clumping and consistency in primate’s patterns of range use: definitions, sampling, assessment and applications. Folia Primatologica. 34: 111-139.
Ribeiro, A. P.; Port-Cavalho, M.; Aguiar, O. T.de & Sarzi, I. (2007). Avaliação do potencial de bugios (Alouatta clamitans, Cabrera, 1940) (Primates: Atelidae) como dispersores de sementes no Parque Estadual da Cantareira. IF Sér. Reg. 31: 147-151.
Rocha, V.J. (2000). Macaco-prego, como controlar esta nova praga florestal? Floresta. 30: 95-99.
Rocha-Mendes, F.; Mikich, S.B.; Bianconi, G.V. & Pedro, W.A. (2005). Mamíferos do município de Fênix, Paraná, Brasil: etnozoologia e conservação. Revista Brasileira de Zoologia. 22: 991-1002.
Rowe, N. (1996). The pictorial guide to the living primates. East Hampton, New York: Pogonias Press.
Rylands, A.B.; Schneider, H.; Langguth, A.; Mittermeier, R.A.;Groves, C.P. & Rodriguez-Luna, E. (2000). An assessment of the diversity of new world monkeys. Neotropical Primates. 8: 61 – 93.
Sammarco, Y. M. (2000). Caracterização de três loci de DNA microssatélite em bugio-ruivo: uma ferramenta para estudos populacionais, filogeografia e conservação. Monografia de Bacharelado. Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil.
SÃO PAULO. Secretaria do Meio Ambiente. (1998). Fauna ameaçada no estado de São Paulo. Série documentos ambientais PROBIO, São Paulo: SMA/CED, 60p.
Schwarzkopf, L. & Rylands, A.B. (1989). Primate species richness in relation to habitat structure in amazoniam rainforest fragments. Biol. Conserv. 48: 1 -12.
Setz, E.Z.F. & De Hoyos, A. (1985). Participação do tempo: o problema da dependência entre observações comportamentais sucessivas. A Primatologia no Brasil. 2: 191-201.
Setz, E.Z.F. (1991). Métodos de quantificação de comportamento de primatas em estudos de campo. In: A primatologia no Brasil. Belo Horizonte, MG. 3: 411-435
Shannon, C.E. (1948). A mathematical theory of communications. Bull System Techn. J. 27:379-423, 623-655.
Siemers, B.M. (2000). Seasonal variation in food resource and forest strata use by brown capuchin monkeys (Cebus apella) in a disturbed forest fragment. Folia Primatologica. 71: 181-184.
Silva, E.C.J. (1981). A preliminary survey of brown howler monkeys (Alouatta fusca) at the Cantareira Reserve (São Paulo, Brazil). Revista Brasileira de Biologia. 41: 897–909.
Silveira, R.M.; Codenotti, T.L. (2001). Interações sociais e dieta do bugio-ruivo, Alouatta guariba clamitans no Parque Estadual de Itapuã, Rio Grande do Sul, Brasil. Neotropical Primates. 9: 15-19.

98
Smith, C.C. (1977). Feeding behaviour and social organization in hower monkeys. ln: Clutton-Brock, T. H. (Ed). Primate Ecology: Studies of Feeding and Ranging Behaviour in Lemurs, Monkeys and Apes. London, Academic Press, p97-126.
Somarriba, E.; Harvey, C.A.; Samper, M.; Anthony, F.; Gonzales, J.; Slaver, C.H. & Rice, R.A. (2004). Biodiversity conservation in Neotropical coffee (Coffea arabica) plantations. In: Schroth, G.; Fonseca, G.; Gascon; C.; Vasconcelos, H.; Izac, A.M. & Harvey, C.A. (Eds). Agroforestry and conservation of biodiversity in tropical landscapes. Island Press, New York, p198-226.
Spironello, W.R. (2001). The brown capuchin monkey (Cebus apella): ecology and home range requirements in central Amazonia. In: Bierregaard, R.O.; Gascon, C; Lovejoy, T.E. & Mesquita, C.G. (Eds). Lessons from Amazonia: the ecology and conservation of a fragmented forest. New Haven:Yale U Pr. p271-83.
Steinmetz, S. (2000). Ecologia e comportamento do bugio (Alouatta guariba clamitans, Atelidae-Primates) no Parque Estadual de Intervales, SP. Dissertação de mestrado, Universidade de São Paulo, São Paulo, Brasil.
Steinmetz, S. (2001). Densidade e conservação do bugio (Alouatta fusca) no Parque Estadual de Intervales. Neotrop. Primates. 9: 69-73.
Strier, K.B. (2000). Primate behavioral ecology. Needham Heights: Allyn & Bacon.
Struhsaker, T.T. (1974). Correlates of ranging behaviour in a group of red colobus monkeys, Colobus badius tephrosceles. The American Zoologist. 14: 177-184.
Struhsaker, T.T. (1975). The red colobus monkey. The University of Chicago Press, Chicago.
Sussman, R.W. (2000). Primate Ecology and Social Structure: New World Monkeys. v.2. Washington: Pearson Custom Publishing, 207p.
Terborgh, J. (1983). Five new world primates: a study in comparative ecology. Princeton University Press, New Jersey. 260p.
Tossulino, M.G.P.; Margarido, T.C.C.; Straube, F.C.; Moura-Leite, J. C.; Morato, S.A.A.; Bérnils, R.S.; Casagrande, M.M. & Mielke, H.H. (1995). Lista vermelha de animais ameaçados de extinção do Estado do Paraná. Curitiba: SEMA/ GTZ, 176p.
Valle, Y.G.; Muñoz, D.; Magaña-Alejandro, M.; Estrada, A. & Franco, B. (2001). Uso de plantas como alimento por monos aulladores, Alouatta palliata, en el Parque Yumká, Tabasco, México. Neotropical Primates. 9: 112-118.
Van Belle, S. & Estrada, A. (2005). Cambios demográficos en poblaciones del mono aullador negro (Alouatta pigra) como consecuencia de la fragmentación del hábitat. Universidad Y Ciencia. II: 1-9.
Vilela, S.L. & D.S. Faria. (2002). Dieta do Callithrix penicillata (Primates, Callithrichidae) em Áreas de Cerrado no Distrito Federal, Brasil. Neotropical Primates. 10: 17-20.

99
Williamson, E.A. & Feistner, A.T.C. (2003). Habituating primates: processes, techniques, variables and ethics. In: Setchell, J.M. & Curtis, D.J. (Eds). Field and Laboratory Methods in Primatology: A Practical Guide. Cambridge University Press, p. 25-39.
Zunino, G.E.; González, V.; Kowalewski, M.M. & Bravo, S.P. (2001). Alouatta caraya: Relations among habitat, density and social organization. Primate report. 61: 37-46.

100
Anexo 1: Fotos dos indivíduos de Alouatta guariba clamitans do grupo de estudo
`
Fêmea adulta. Juvenil II macho em um Eucalyptus sp.
Macho alfa.

101
Anexo 2. Fotos de algumas espécies alimentares (alóctones e autóctones) consumidas.
Consumo do fruto de Quercus robur (carvalho-europeu).
Consumo de frutos de Diospyrus kaki (caqui).

102
Consumo de frutos de Hovenia dulcis (uva-do-japão).
Consumo de frutos de Eryobotrya japonica (ameixa-amarela)

103
Consumo de acículas de Pinus spp. (pinus)
Consumo de folhas de Araucaria angustifolia (pinheiro-brasileiro)

104
Consumo de folhas de Zanthoxylum rhoifolium (mamica-de-cadela).
Consumo de flores de Pyrostegia venusta (cipó-de-são-joão)

105
Consumo de folhas e flores de Mimosa scabrella (bracatinga)
Fezes com sementes de Casearia decandra e sementes e casca do fruto de Vitis americana (jan/2008).

106
1 cm
Fezes com sementes de C. decandra, N. megapotamica e frutos de Eucalyptus sp. (fev/2008).
1 cm
Detalhe das fezes com sementes de C. decandra e frutos de Eucalyptus sp. (fev/2008).

107
Anexo 3. Detalhes da área de estudo e acontecimentos.
Vitis americana (uva) inserida da mata de estudo (quadrícula D16).
Detalhes da Vitis americana inserida da mata de estudo (quadrícula D16).

108
Fêmea do grupo focal aguardando o retorno no macho alfa, sumido no dia anterior, um dia antes do corte dos Pinus (maio).
Área com Pinus spp., no dia que iniciaram os cortes.

109
Área de espécies alóctones após a supressão dos Pinus spp.
Descida da fêmea ao solo para beber água em um córrego na entrada da mata.





























