Embed Size (px)
Citation preview

Unique Regulation of Long Term Potentiationin the Rat Ventral Hippocampus
Nicola Maggio and Menahem Segal*
ABSTRACT: Morphological and electrophysiological evidence has beenaccumulated in recent years to indicate that a functional gradient existsalong the septo-temporal axis of the hippocampus such that spatial mem-ory is associated primarily with the dorsal (septal) sector while the func-tions of the ventral sector are not yet clearly defined. Also, the ventralhippocampus (VH) is reported to express a much smaller long termpotentiation of responses to afferent stimulation than the dorsal sector.In the present study, we first confirmed that CA1 region of VH slicesexpresses significantly smaller LTP than the dorsal hippocampus. Strik-ingly, much larger LTP was obtained in VH slices following low fre-quency priming stimulation applied prior to the tetanic stimulation.DHPG ((S)-3,5-Dihydroxyphenylglycine hydrate) a metabotropic gluta-mate receptor agonist, produced a similar potentiating effect on LTP asthat produced by the priming stimulation. In both cases, the spectralanalysis of spontaneous electrical activity recorded from the same loca-tion in the slice revealed an increase in peak amplitude around 30 Hz.MCPG, a metabotropic glutamate receptor antagonist, and both thapsi-gargin and cyclopiazonic acid, inhibitors of Ca2+ release from stores,blocked the potentiating action of both DHPG and the priming stimula-tion. These results indicate that the ventral hippocampus possesses differ-ent network properties compared to the dorsal hippocampus and that itsability to undergo plastic changes is controlled by a metabotropic gluta-mate receptor. VVC 2006 Wiley-Liss, Inc.
KEY WORDS: ventral hippocampus; LTP; LTD; metabotropic glu-tamate receptors; thapsigargin
INTRODUCTION
The hippocampus has been assumed to play a key role in memoryprocesses (Squire, 2004). However, recent evidence (Witter and Amaral,2004) suggests that a functional differentiation exists along the septo-temporal axis of the hippocampus (Colombo et al., 1998; Moser andMoser, 1998; Strange et al., 1999; Pothuizen et al., 2004). Specifically, itwas proposed that while the dorsal hippocampus is involved in spatialmemory, the temporal pole is devoted to the processing of informationrelated to the motivational and homeostatic state of the animal (Ban-nerman et al., 1999, 2004; Moser and Moser, 1998). Later studies alsodemonstrated that the ventral hippocampus, more than the dorsal one, isinvolved in the development of affective disorders (Kjelstrup et al., 2002;
Degroot and Treit, 2004). In addition, it was shownboth in vivo (Maruki et al., 2001) and in vitro (Papa-theodoropoulos and Kostopoulos, 2000a; Colgin et al.,2004) that the ability to evoke short and long termplasticity differs between dorsal and ventral hippocam-pus (DH and VH), respectively. Particularly, the mag-nitude of LTP, a cellular model of learning and mem-ory (Bliss and Collingridge, 1993; Dudek and Bear,1993), is significantly smaller in VH compared to DH.Paired pulse responses also reveal asimilarities betweenthe two regions (Papatheodoropoulos and Kostopoulos,2000b; Papatheodoropoulos et al., 2002).
The objectives of the present study are to investigatethe differences in plastic properties between DH andVH. In addition, the spontaneous activity was charac-terized in the VH slice in relation to the evoked neuro-nal network activity.
MATERIALS AND METHODS
Chemicals
(S)-3,5-Dihydroxyphenylglycine hydrate (DHPG)was purchased from Sigma (St. Louis, MO), (RS)-methyl-4-carboxyphenylglycine (MCPG) was pur-chased from Tocris (Ellisville, MO), Thapsigargin wasa generous gift from Alomone Labs (Jerusalem, Israel).Pharmaceuticals were added to the perfusion mediumwith special care to prevent changes in temperature,pH, flow rate, or degree of oxygenation of the artificialCSF (aCSF).
Slice Preparation
Hippocampal slices from 10 to 12 week-old maleWistar rats were prepared as follows: After rapid decap-itation, the hippocampus was removed, and 350 lmslices were prepared using a McIlwain tissue slicer. Spe-cifically, transverse slices were cut from the regionextending more than 1 mm and less than 3.5 mm fromthe septal or temporal pole of the hippocampus(Fig. 1A). Experiments involving comparisons betweendorsal and ventral hippocampus were performed on sli-ces derived from the same hippocampus at the same ex-perimental conditions. To cut both regions perpendicu-larly to the longitudinal axis of the hippocampus theplate supporting the structure was rotated so that sliceswere cut from the respective ends of the hippocampus.
Department of Neurobiology, The Weizmann Institute, Rehovot, Israel*Correspondence to: Menahem Segal, Department of Neurobiology, TheWeizmann Institute of Science, 76100 Rehovot, Israel.E-mail: [email protected] for publication 7 September 2006DOI 10.1002/hipo.20237Published online 8 November 2006 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 17:10–25 (2007)
VVC 2006 WILEY-LISS, INC.
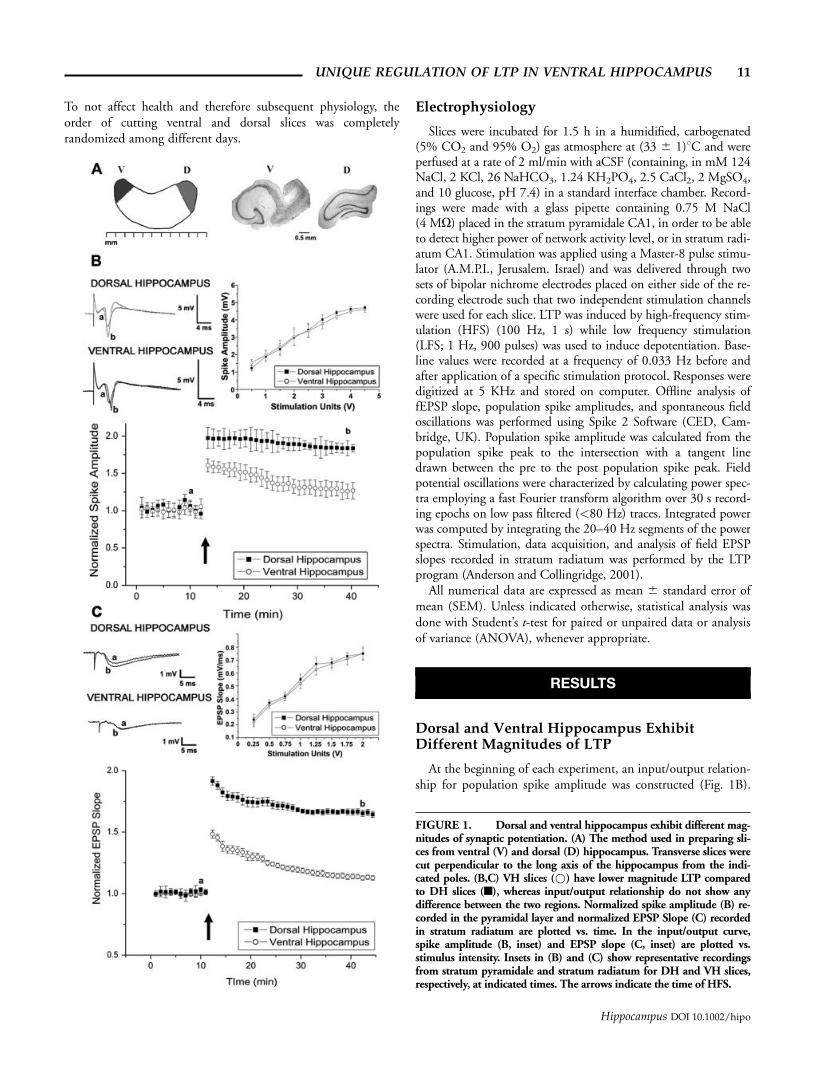
To not affect health and therefore subsequent physiology, theorder of cutting ventral and dorsal slices was completelyrandomized among different days.
Electrophysiology
Slices were incubated for 1.5 h in a humidified, carbogenated(5% CO2 and 95% O2) gas atmosphere at (33 6 1)8C and wereperfused at a rate of 2 ml/min with aCSF (containing, in mM 124NaCl, 2 KCl, 26 NaHCO3, 1.24 KH2PO4, 2.5 CaCl2, 2 MgSO4,and 10 glucose, pH 7.4) in a standard interface chamber. Record-ings were made with a glass pipette containing 0.75 M NaCl(4 MO) placed in the stratum pyramidale CA1, in order to be ableto detect higher power of network activity level, or in stratum radi-atum CA1. Stimulation was applied using a Master-8 pulse stimu-lator (A.M.P.I., Jerusalem. Israel) and was delivered through twosets of bipolar nichrome electrodes placed on either side of the re-cording electrode such that two independent stimulation channelswere used for each slice. LTP was induced by high-frequency stim-ulation (HFS) (100 Hz, 1 s) while low frequency stimulation(LFS; 1 Hz, 900 pulses) was used to induce depotentiation. Base-line values were recorded at a frequency of 0.033 Hz before andafter application of a specific stimulation protocol. Responses weredigitized at 5 KHz and stored on computer. Offline analysis offEPSP slope, population spike amplitudes, and spontaneous fieldoscillations was performed using Spike 2 Software (CED, Cam-bridge, UK). Population spike amplitude was calculated from thepopulation spike peak to the intersection with a tangent linedrawn between the pre to the post population spike peak. Fieldpotential oscillations were characterized by calculating power spec-tra employing a fast Fourier transform algorithm over 30 s record-ing epochs on low pass filtered (<80 Hz) traces. Integrated powerwas computed by integrating the 20–40 Hz segments of the powerspectra. Stimulation, data acquisition, and analysis of field EPSPslopes recorded in stratum radiatum was performed by the LTPprogram (Anderson and Collingridge, 2001).
All numerical data are expressed as mean 6 standard error ofmean (SEM). Unless indicated otherwise, statistical analysis wasdone with Student’s t-test for paired or unpaired data or analysisof variance (ANOVA), whenever appropriate.
RESULTS
Dorsal and Ventral Hippocampus ExhibitDifferent Magnitudes of LTP
At the beginning of each experiment, an input/output relation-ship for population spike amplitude was constructed (Fig. 1B).
FIGURE 1. Dorsal and ventral hippocampus exhibit different mag-nitudes of synaptic potentiation. (A) The method used in preparing sli-ces from ventral (V) and dorsal (D) hippocampus. Transverse slices werecut perpendicular to the long axis of the hippocampus from the indi-cated poles. (B,C) VH slices (*) have lower magnitude LTP comparedto DH slices (n), whereas input/output relationship do not show anydifference between the two regions. Normalized spike amplitude (B) re-corded in the pyramidal layer and normalized EPSP Slope (C) recordedin stratum radiatum are plotted vs. time. In the input/output curve,spike amplitude (B, inset) and EPSP slope (C, inset) are plotted vs.stimulus intensity. Insets in (B) and (C) show representative recordingsfrom stratum pyramidale and stratum radiatum for DH and VH slices,respectively, at indicated times. The arrows indicate the time of HFS.
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 11
Hippocampus DOI 10.1002/hipo

The relationship between stimulation intensity and magnitude ofpopulation spike was similar in ventral and dorsal hippocampus(n ¼ 9 slices in each group). At stimulus intensities that yielded50% of maximal responses the population spikes were 2.44 60.38 mV for VH and 2.31 6 0.35 mV for DH. Before deliveringthe HFS, a 15 min baseline was recorded for both VH and DHslices. LTP was induced by a 100 Hz stimulation, which led topotentiation in both groups of slices. However, the level of shortterm potentiation was lower in the VH compared to the DH(1.60 6 0.08 in VH and 1.97 6 0.14 in DH, P < 0.05). Thedecay rate of the potentiation was also faster in VH compared toDH; 25 min after the tetanic stimulation the responses reached1.26 6 0.096 in VH and 1.84 6 0.065 in the DH, P < 0.05,Fig. 1B).
Since population spikes are generated at higher stimulationintensities, it is possible that the different magnitude of LTP inVH vs. DH could be due to different levels of saturation of theresponse in the two regions. To investigate this possibility, thesame experiment described earlier was also conducted by stimu-lation of lower amplitude and recording EPSP in stratum radia-tum (Fig. 1C). In this case, the input/output curve also did notshow any differences between DH and VH (n ¼ 9 slices in eachgroup). At stimulus intensities that yielded 50% of maximalresponses, the EPSP slopes were 0.52 6 0.042 mV/ms for VHand 0.55 6 0.041 mV/ms for DH. After the delivery of HFS(100 Hz), LTP was induced in slices from both hippocampalregions. However, the VH showed a lower initial response to thetetanic stimulation compared to DH (1.48 6 0.032 in VH and1.91 6 0.033 in DH, P < 0.001). In addition, a faster decayrate of the potentiation was found in VH as seen earlier:Twenty-five minutes after the tetanic stimulation, the responsesreached 1.20 6 0.03 in VH and 1.70 6 0.027 in DH (P <0.001). Taken together, these data confirm previous experimen-tal findings (Papatheodoropoulos and Kostopoulos, 2000a,b)and support the assumption that different mechanisms mayunderlie synaptic plasticity in DH and VH.
One possible cause for the lower level of potentiation in theVH slices is that the potentiated pathways in the VH slice are al-ready saturated. A common procedure for depotentiation of asaturated pathway is to apply a low frequency stimulation trainwhich causes a suppression of the synaptic transmission and sub-sequently enables tetanic-induced potentiation of the pathway. Alow frequency priming stimulation (LFS) protocol producedsimilar decreases in reactivity to the stimulated pathway in bothDH and VH, yet produced remarkably different effects on thesubsequent HFS-induced LTP. Results of these experiments areshown in Figure 2, which compares the effects of LFS in DH(n ¼ 9) and VH (n ¼ 9). In both cases, low frequency stimula-tion was delivered to a single pathway after a 15 min baselineand reliably induced short term depression in slices from bothregions. The magnitude of the depotentiation was 0.71 6 0.047for fEPSP slope and 0.73 6 0.06 for population spike in DH(Fig. 2A) while in VH (Fig. 2B) it was 0.66 6 0.065 and0.79 6 0.08, respectively. No significant difference was found inthe magnitude of depression between the two regions. A HFSwas then delivered to the depotentiated pathway 20 min after
the LFS. Surprisingly, a significantly larger magnitude LTP wasnow found in VH slices. In both slice types, the degree of poten-tiation was calculated relative to the baseline prior to priming,but the priming did not make a big difference in the calculationof the evoked LTP. The fEPSP slope change at the onset of LTPover baseline was 1.75 6 0.095 while for the population spike itwas 2.39 6 0.14. 30 min after the HFS, the responses stabilizedat a plateau level 1.54 6 0.083 for fEPSP slope and 2.12 60.076 for population spikes. LTP was produced also in depoten-tiated DH slices. The magnitude of the potentiation was 1.56 60.064 for the fEPSP slope and 1.72 6 0.05 for the populationspike. Thirty minutes following the first HFS to the \naı̈ve"pathway, the same tetanic stimulation, described earlier, wasdelivered to the \unprimed" pathway both in DH and VH.Interestingly, in VH, this channel showed a decreased level ofLTP as described in the previous experiments (1.29 6 0.09 forthe EPSP and 1.52 6 0.073 for population spike). In the DH afull magnitude LTP was observed (1.77 6 0.05 for fEPSP slopeand 1.85 6 0.056 for population spike). This LTP was signifi-cantly higher (for EPSP slope and population spike, P < 0.05)than the potentiation elicited by the HFS on the depotentiatedchannel.
To get a better estimate of the LTP induced by HFS followingthe priming protocol, some experiments were also conductedwith the recording electrode placed in stratum radiatum of CA1,both in DH and VH. EPSP slopes recorded in stratum radiatumallow a closer comparison between pre synaptic and post synap-tic components of the field responses following the stimulation.
Figures 3A,B show the input/output relationship for EPSPslopes plotted against fiber volley amplitudes in DH and VHperformed at the beginning of each experiment (n ¼ 6 slices ineach group). As it was shown earlier, no significant differenceswere found between DH and VH slices. In both dorsal and ven-tral slices, higher stimulation intensities produced higher ampli-tudes of fiber volleys, which were not significantly different inthe two stimulated pathways. After a 15 min baseline recording,one of the pathways was stimulated by the LFS protocol. Thisresulted in a selective depression of the responses to the stimu-lated pathway (at 45 min of recordings: 0.789 6 0.06 in DHand 0.736 6 0.05 in VH). After 50 min of recording, HFS wasdelivered to the depotentiated pathway, producing LTP in bothdorsal and ventral slices. In dorsal hippocampus, the LTP in-duced was not different from the one expressed in the pathway thatdid not get the LFS protocol (after 15 min from HFS: 1.70 60.05 for pathway 1 and 1.67 6 0.07 for pathway 2, P ¼ 0.58).In VH, the unprimed pathway, after a HFS, expressed a shortterm potentiation with a much faster decay compared to theone induced in the depotentiated pathway (after 15 min fromHFS, 1.69 6 0.05 for pathway 1 and 1.17 6 0.07 for pathway2, P < 0.001). However, in both VH and DH slices, LFS andHFS did not produce any change in the fiber volleys amplitude(Figs. 3A,B), indicating that these stimulation protocols affectonly the post synaptic responses. These results, together withthe findings reported earlier, confirm that in VH, unlike theDH, LTP can be markedly facilitated by a priming depotentiat-ing stimulation.
12 MAGGIO AND SEGAL
Hippocampus DOI 10.1002/hipo

Finally, to test the hypothesis that a HFS protocol followingthe LFS is necessary for the induction of LTP in ventral hippo-campal slices, an additional experiment was carried out in stra-tum radiatum CA1. Figure 3C shows that, in these slices, thepriming protocol alone produced a depotentiation (at 70 min
from the beginning of the recordings: 0.75 6 0.02), which didnot result in an enhancement of synaptic responses about 1 h afterthe LFS protocol. This indicates that the priming stimulation doesnot produce a delayed increase of the transmission per se, but itprimes the synapses to respond at the subsequent HFS.
FIGURE 2. Low frequency priming stimulation (LFS) increasesLTP level in ventral but not in dorsal hippocampus recorded inCA1 stratum pyramidale. In DH slices (A), a decreased level of LTPcould be evoked by HFS in a previous primed pathway (three bars,in this and following figures, *) compared to its control (n);whereas in VH slices (B), a larger LTP level was obtained in the
primed (*) but not in the control (n) pathway. Averaged normal-ized EPSP slope (left column in A and B) and spike amplitude(right column) are plotted vs. time. Representative traces for DHand VH slices at indicated times are shown above A and B. Thearrows indicate the time of HFS delivery while the three bars denoteLFS.
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 13
Hippocampus DOI 10.1002/hipo
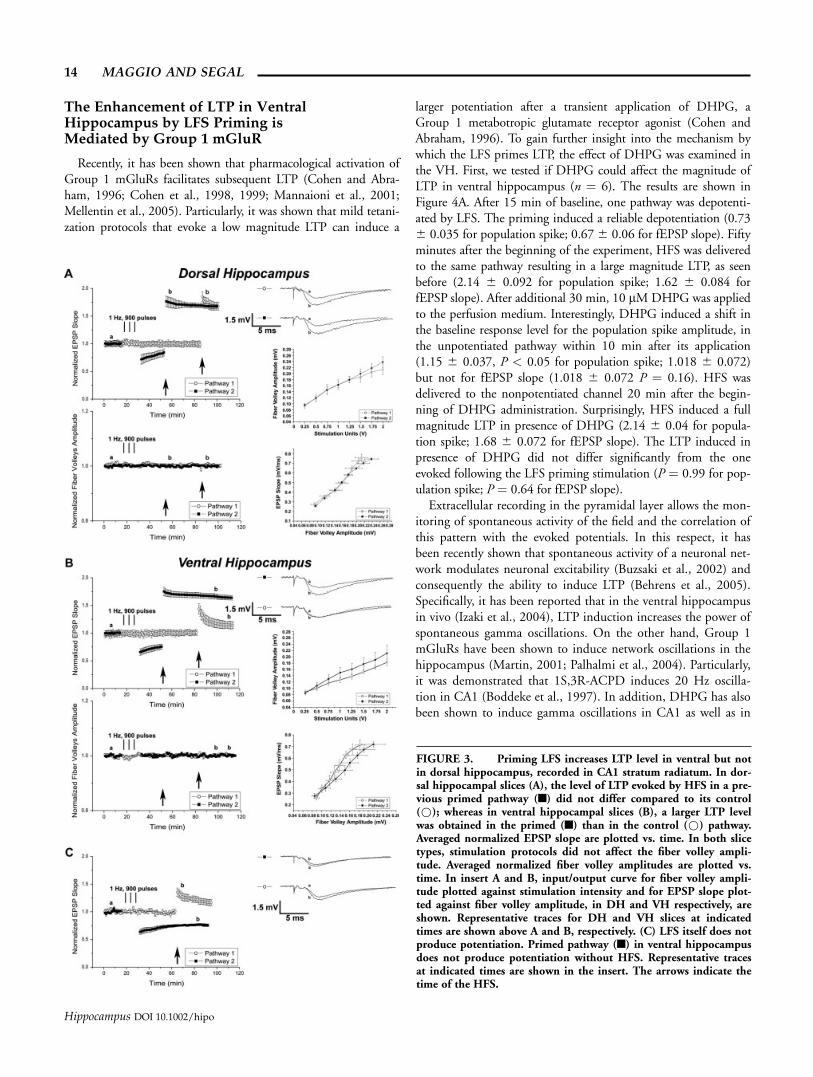
The Enhancement of LTP in VentralHippocampus by LFS Priming isMediated by Group 1 mGluR
Recently, it has been shown that pharmacological activation ofGroup 1 mGluRs facilitates subsequent LTP (Cohen and Abra-ham, 1996; Cohen et al., 1998, 1999; Mannaioni et al., 2001;Mellentin et al., 2005). Particularly, it was shown that mild tetani-zation protocols that evoke a low magnitude LTP can induce a
larger potentiation after a transient application of DHPG, aGroup 1 metabotropic glutamate receptor agonist (Cohen andAbraham, 1996). To gain further insight into the mechanism bywhich the LFS primes LTP, the effect of DHPG was examined inthe VH. First, we tested if DHPG could affect the magnitude ofLTP in ventral hippocampus (n ¼ 6). The results are shown inFigure 4A. After 15 min of baseline, one pathway was depotenti-ated by LFS. The priming induced a reliable depotentiation (0.736 0.035 for population spike; 0.676 0.06 for fEPSP slope). Fiftyminutes after the beginning of the experiment, HFS was deliveredto the same pathway resulting in a large magnitude LTP, as seenbefore (2.14 6 0.092 for population spike; 1.62 6 0.084 forfEPSP slope). After additional 30 min, 10 lMDHPG was appliedto the perfusion medium. Interestingly, DHPG induced a shift inthe baseline response level for the population spike amplitude, inthe unpotentiated pathway within 10 min after its application(1.15 6 0.037, P < 0.05 for population spike; 1.018 6 0.072)but not for fEPSP slope (1.018 6 0.072 P ¼ 0.16). HFS wasdelivered to the nonpotentiated channel 20 min after the begin-ning of DHPG administration. Surprisingly, HFS induced a fullmagnitude LTP in presence of DHPG (2.14 6 0.04 for popula-tion spike; 1.68 6 0.072 for fEPSP slope). The LTP induced inpresence of DHPG did not differ significantly from the oneevoked following the LFS priming stimulation (P ¼ 0.99 for pop-ulation spike; P ¼ 0.64 for fEPSP slope).
Extracellular recording in the pyramidal layer allows the mon-itoring of spontaneous activity of the field and the correlation ofthis pattern with the evoked potentials. In this respect, it hasbeen recently shown that spontaneous activity of a neuronal net-work modulates neuronal excitability (Buzsaki et al., 2002) andconsequently the ability to induce LTP (Behrens et al., 2005).Specifically, it has been reported that in the ventral hippocampusin vivo (Izaki et al., 2004), LTP induction increases the power ofspontaneous gamma oscillations. On the other hand, Group 1mGluRs have been shown to induce network oscillations in thehippocampus (Martin, 2001; Palhalmi et al., 2004). Particularly,it was demonstrated that 1S,3R-ACPD induces 20 Hz oscilla-tion in CA1 (Boddeke et al., 1997). In addition, DHPG has alsobeen shown to induce gamma oscillations in CA1 as well as in
FIGURE 3. Priming LFS increases LTP level in ventral but notin dorsal hippocampus, recorded in CA1 stratum radiatum. In dor-sal hippocampal slices (A), the level of LTP evoked by HFS in a pre-vious primed pathway (n) did not differ compared to its control(*); whereas in ventral hippocampal slices (B), a larger LTP levelwas obtained in the primed (n) than in the control (*) pathway.Averaged normalized EPSP slope are plotted vs. time. In both slicetypes, stimulation protocols did not affect the fiber volley ampli-tude. Averaged normalized fiber volley amplitudes are plotted vs.time. In insert A and B, input/output curve for fiber volley ampli-tude plotted against stimulation intensity and for EPSP slope plot-ted against fiber volley amplitude, in DH and VH respectively, areshown. Representative traces for DH and VH slices at indicatedtimes are shown above A and B, respectively. (C) LFS itself does notproduce potentiation. Primed pathway (n) in ventral hippocampusdoes not produce potentiation without HFS. Representative tracesat indicated times are shown in the insert. The arrows indicate thetime of the HFS.
14 MAGGIO AND SEGAL
Hippocampus DOI 10.1002/hipo
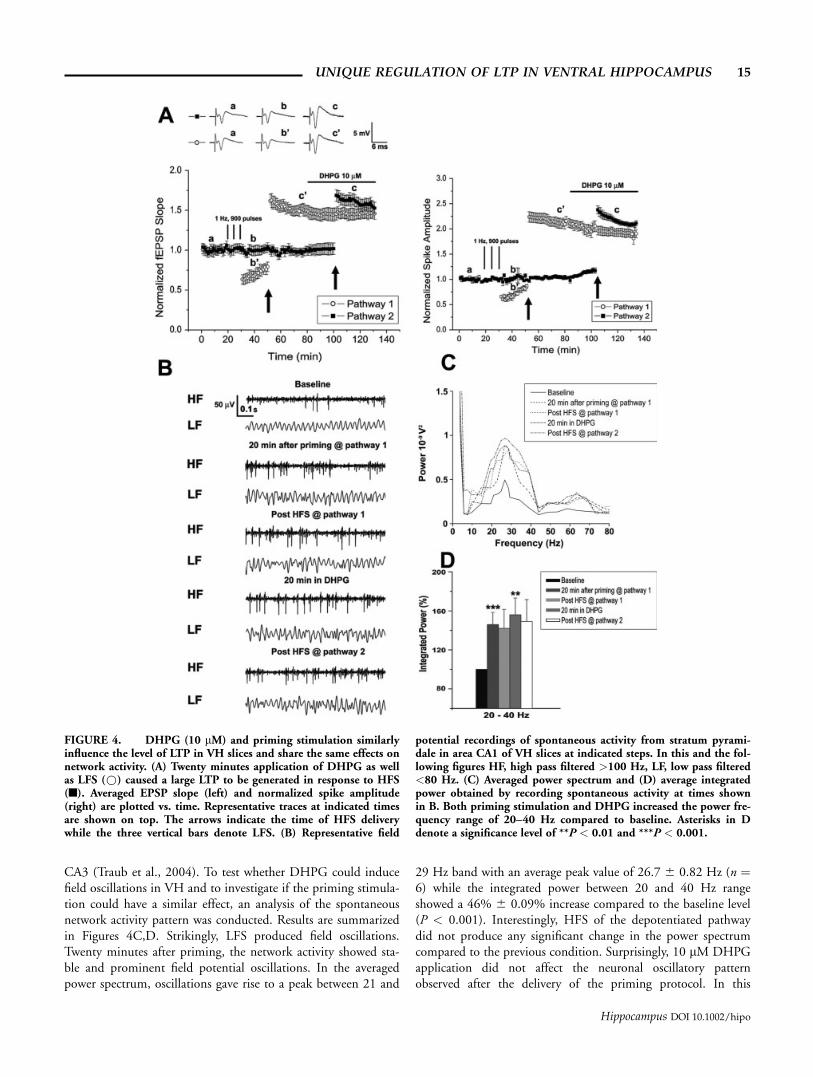
CA3 (Traub et al., 2004). To test whether DHPG could inducefield oscillations in VH and to investigate if the priming stimula-tion could have a similar effect, an analysis of the spontaneousnetwork activity pattern was conducted. Results are summarizedin Figures 4C,D. Strikingly, LFS produced field oscillations.Twenty minutes after priming, the network activity showed sta-ble and prominent field potential oscillations. In the averagedpower spectrum, oscillations gave rise to a peak between 21 and
29 Hz band with an average peak value of 26.7 6 0.82 Hz (n ¼6) while the integrated power between 20 and 40 Hz rangeshowed a 46% 6 0.09% increase compared to the baseline level(P < 0.001). Interestingly, HFS of the depotentiated pathwaydid not produce any significant change in the power spectrumcompared to the previous condition. Surprisingly, 10 lM DHPGapplication did not affect the neuronal oscillatory patternobserved after the delivery of the priming protocol. In this
FIGURE 4. DHPG (10 lM) and priming stimulation similarlyinfluence the level of LTP in VH slices and share the same effects onnetwork activity. (A) Twenty minutes application of DHPG as wellas LFS (*) caused a large LTP to be generated in response to HFS(n). Averaged EPSP slope (left) and normalized spike amplitude(right) are plotted vs. time. Representative traces at indicated timesare shown on top. The arrows indicate the time of HFS deliverywhile the three vertical bars denote LFS. (B) Representative field
potential recordings of spontaneous activity from stratum pyrami-dale in area CA1 of VH slices at indicated steps. In this and the fol-lowing figures HF, high pass filtered >100 Hz, LF, low pass filtered<80 Hz. (C) Averaged power spectrum and (D) average integratedpower obtained by recording spontaneous activity at times shownin B. Both priming stimulation and DHPG increased the power fre-quency range of 20–40 Hz compared to baseline. Asterisks in Ddenote a significance level of **P < 0.01 and ***P < 0.001.
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 15
Hippocampus DOI 10.1002/hipo

respect, after 20 min of drug treatment, the averaged power spec-trum showed a peak between 20 and 34 Hz with an average peakvalue at 26.9 6 1.46 Hz. Integrated power indicated a 56% 60.175% increase of field activity over baseline (P < 0.01).
Altogether, these results indicate that LFS and DHPG affect theability to produce LTP in VH and evoke similar field oscillations.These results also suggest that both protocols may activate thesame receptor and if so, they should eventually be blocked by the
same receptor antagonist. To this end, MCPG (200 lM), a GroupI/II metabotropic glutamate receptors antagonist, was used.
In the first experiment (VH, n ¼ 6, Fig. 5A), 10 lM DHPGwas applied to the perfusion bath after 15 min of baseline. HFSwas delivered to one of the pathways 20 min following the drugapplication. This reliably reproduced the facilitation of LTP drivenby DHPG (2.07 6 0.055 for population spike; 1.86 6 0.07 forfEPSP). Nevertheless, MCPG produced a decay of the response in
FIGURE 5. MCPG (200 lM) blocks the LTP facilitation inducedby DHPG (10 mM). (A) In ventral hippocampal slices 20 min appli-cation of MCPG (n) impairs LTP facilitation induced by DHPGapplication (*). Averaged EPSP slope and average spike amplitude areplotted vs. time. Representative traces at indicated times are showed ontop. The arrows indicate the time of HFS. (B) Representative field
potential recordings of spontaneous activity from stratum pyramidalein area CA1 at indicated steps. (C) Average power spectrum and (D) av-erage integrated power obtained by recording spontaneous activity atthe time intervals shown in B. MCPG decreased the level of power fre-quency range around 20–40 Hz induced by application of DHPG.Asterisks in D denote a significance level of ***P < 0.001.
16 MAGGIO AND SEGAL
Hippocampus DOI 10.1002/hipo

the potentiated pathway. However, after 25 min of MCPG treat-ment, the potentiation level stabilized at 1.74 6 0.08 for popula-tion spike and at 1.53 6 0.08 for fEPSP. On the other hand, inpresence of its antagonist, DHPG was unable to facilitate LTP inVH: a smaller magnitude LTP was evoked in the second pathwayunder these conditions (1.696 0.046 for population spike; 1.38 60.074). Interestingly, the level of potentiation reached by path-way 2 differed statistically from the one observed in the controlpathway (P < 0.05). Besides, MCPG was unable to affect theincrease in the baseline of the population spike response medi-ated by DHPG (1.17 6 0.066).
The effect of MCPG on the field oscillation induced byDHPG is summarized in Figures 5C,D. The Group 1 mGluRsagonist induced field oscillations. Twenty minutes after its bathapplication, the averaged power spectrum showed activity thatpeaked in a band of 19–39 Hz with an average peak value of27.53 6 1.64 Hz. The integrated power at 20–40 Hz band in-terval showed a 56% 6 0.12% (P < 0.001) increase over base-line of network activity. However, in the presence of MCPG,DHPG did not have the same effect on the spontaneous activity.The power of the oscillations decreased significantly, reaching8% 6 0.11% (P ¼ 0.42) over baseline level in the integratedpower. Interestingly, the frequency of oscillations was not af-fected: the band peak was between 19 and 34 Hz with an aver-age peak value of 27.8 6 1.57. These results confirm thatMCPG blocks the field activity induced by DHPG.
The effect of MCPG on the induction of LTP driven by thepriming protocol in VH (n ¼ 6, Fig. 6A) was then examined.When delivered in the control pathway, LFS induced depotentia-tion (0.72 6 0.036 for population spike; 0.72 6 0.05 for fEPSP)and facilitated subsequent LTP (2.00 6 0.039 for populationspike; 1.85 6 0.08 for EPSP). MCPG markedly reduced theresponse of the primed pathway, reaching a plateau (1.60 60.054 for population spike; 1.46 6 0.08 for fEPSP, P < 0.001)after 30 min of drug application. In presence of MCPG, the prim-ing protocol produced a depotentiation (0.79 6 0.01 for popu-lation spike; 0.79 6 0.05 for fEPSP) that was not significantlydifferent from the one evoked in the control pathway (P ¼0.095). However, in presence of MCPG, the priming protocoldid not facilitate the induction of LTP (1.59 6 0.021 for popu-lation spike; 1.41 6 0.07 for fEPSP, P < 0.001), suggesting thatGroup1 mGluRs are involved in the effect mediated by thepriming protocol.
In addition, MCPG blocked the field oscillations induced byLFS (Figs. 6C,D). LFS at pathway 1 produced field oscillationsthat peaked between 20 and 38 Hz with an average peak at 26.760.95 Hz. The percentage of change in the power of the field oscil-latory activity compared to baseline was 45% 6 0.094% (P <0.001). Application of MCPG impaired this effect. Under theseconditions, the integrated power analysis in the 20–40 Hz bandshowed a mere 6% 6 0.06% increase of oscillation power com-pared to baseline values (P ¼ 0.26). In addition, the frequency ofoscillations did not show any change from baseline conditions(band peak 22–34 Hz and average peak 28.61 6 1.23 Hz). Theseresults suggest that the LFS affects the field oscillations by activat-ing Group1 mGluRs.
To investigate whether LTP in dorsal hippocampus could alsobe modulated by Group1 mGluRs and to get further insights intothe effects of those receptors on LTP in ventral hippocampus,some experiments were performed in stratum radiatum CA1.
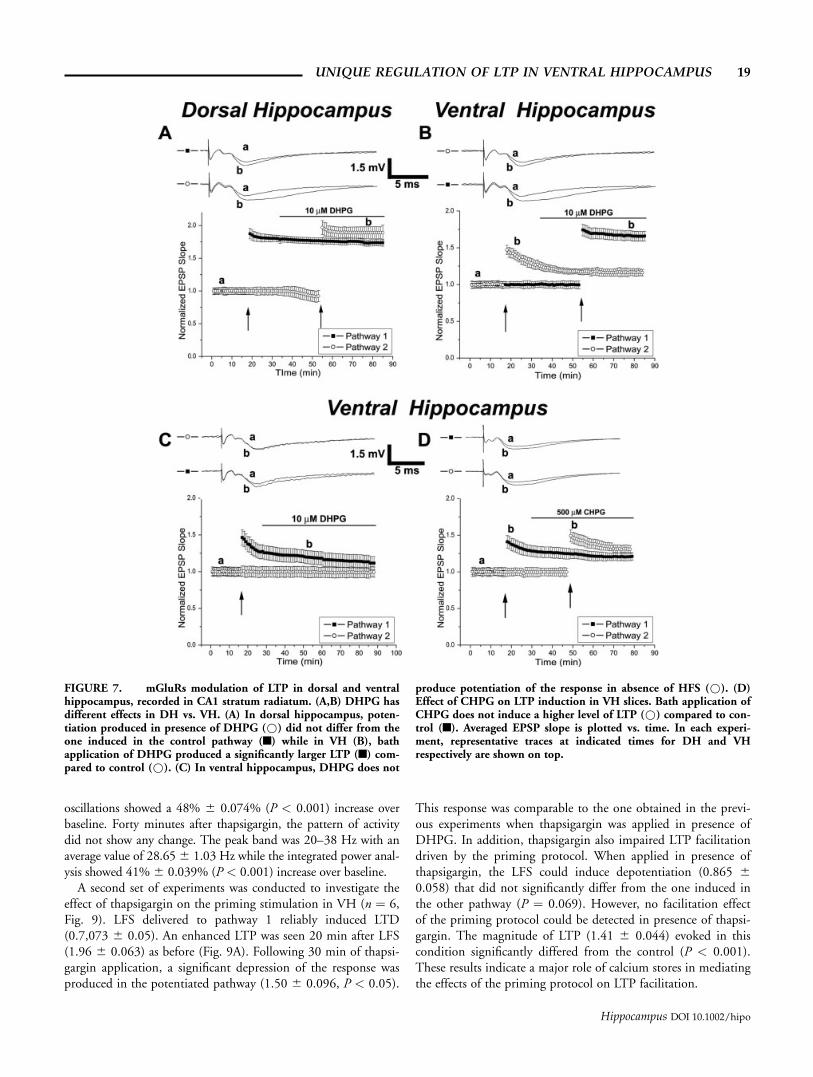
Figures 7A,B show the different effect of 10 lM DHPG onLTP in DH vs. VH slices (n ¼ 6 in each group). In both slices,a HFS was delivered after 15 min of baseline. This stimulationled to LTP production in DH (10 min after HFS: 1.80 60.054), while VH slices expressed the characteristic short termpotentiation after a HFS of same intensity (10 min after HFS:1.30 6 0.066). Interestingly, as previously reported (Cohenet al., 1998), DHPG induced a depression of the transmissiononly in dorsal hippocampus (0.91 6 0.085). However, in ourcase, this effect was only slightly significant (P ¼ 0.043). Deliv-ery of HFS after DHPG application produced LTP in bothslices. In dorsal hippocampus, this potentiation reached a levelof 1.89 6 0.08 10 min after HFS, which was not statisticallysignificant from the level achieved by the previously tetanizedpathway (P ¼ 0.38). In ventral hippocampus, the level of LTPreached a value of 1.69 6 0.07 10 min after HFS and differedsignificantly from the potentiation produced in absence ofDHPG (P < 0.01). These experiments confirm that DHPG hasa unique effect in the modulation of LTP in ventral hippocampalslices. To test the possibility that DHPG alone could affect thesynaptic transmission in VH and possibly lead to potentiation,even without HFS delivery, an additional experiment was carriedout. The results (Fig. 7C) clearly show that DHPG in stratumradiatum of CA1 was not able to affect basal EPSP responses if aHFS was not delivered.
Group 1 mGluRs is a family consisting of two receptors:mGluR 1 and mGluR 5. To better understand which receptorcould mediate the effects of DHPG, an experiment with 500 lMCHPG, a selective mGluR 5 agonist, was performed in stratumradiatum CA1. Figure 7D shows that bath application of CHPGalso produced a potentiation following HFS. This potentiationshowed a higher, not statistically significant, onset compared tothe response obtained in absence of CHPG (1.49 6 0.07 and1.42 6 0.08, respectively, P ¼ 0.36). Furthermore, the decay ofthe potentiation obtained in presence of CHPG also did notshow any significant difference with respect to the one obtainedwithout CHPG (1.37 6 0.06 and 1.27 6 0.07 respectively, P ¼0.12). This indicates that mGluR5 are not likely to be involvedin the effects mediated by DHPG.
Intracellular Calcium Stores Mediate the Effect ofthe Priming Protocol in Ventral Hippocampus
Group 1 mGluR agonists regulate intracellular calcium homeo-stasis (Rae and Irving, 2004) by releasing calcium from intracellu-lar stores through IP3R signaling (Rose and Konnerth, 2001; Rosset al., 2005). In this way they play a major role in induction ofLTP (Wilsch et al., 1998). The possibility that intracellular cal-cium stores mediate the effect of DHPG and LFS was thereforeexamined. Thapsigargin (2 lM), an inhibitor of endoplasmic re-ticulum Ca2+ ATPases, was used in two series of experiments:
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 17
Hippocampus DOI 10.1002/hipo
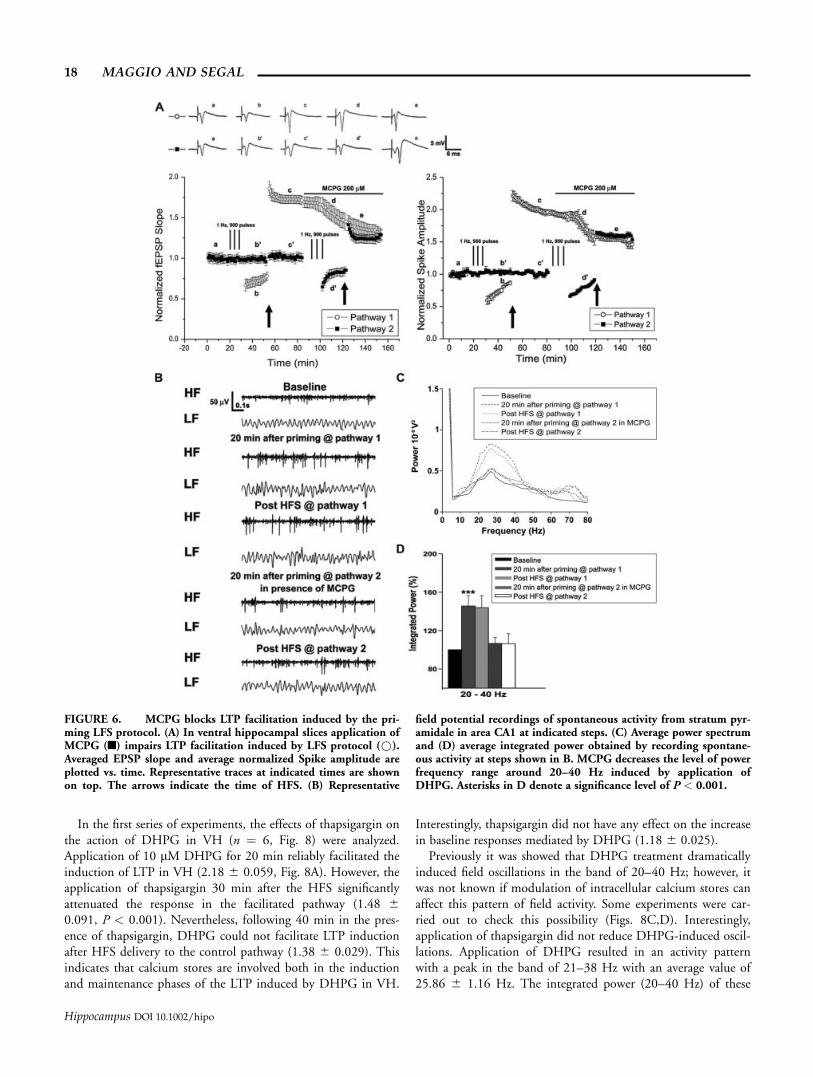
In the first series of experiments, the effects of thapsigargin onthe action of DHPG in VH (n ¼ 6, Fig. 8) were analyzed.Application of 10 lM DHPG for 20 min reliably facilitated theinduction of LTP in VH (2.18 6 0.059, Fig. 8A). However, theapplication of thapsigargin 30 min after the HFS significantlyattenuated the response in the facilitated pathway (1.48 60.091, P < 0.001). Nevertheless, following 40 min in the pres-ence of thapsigargin, DHPG could not facilitate LTP inductionafter HFS delivery to the control pathway (1.38 6 0.029). Thisindicates that calcium stores are involved both in the inductionand maintenance phases of the LTP induced by DHPG in VH.
Interestingly, thapsigargin did not have any effect on the increasein baseline responses mediated by DHPG (1.18 6 0.025).
Previously it was showed that DHPG treatment dramaticallyinduced field oscillations in the band of 20–40 Hz; however, itwas not known if modulation of intracellular calcium stores canaffect this pattern of field activity. Some experiments were car-ried out to check this possibility (Figs. 8C,D). Interestingly,application of thapsigargin did not reduce DHPG-induced oscil-lations. Application of DHPG resulted in an activity patternwith a peak in the band of 21–38 Hz with an average value of25.86 6 1.16 Hz. The integrated power (20–40 Hz) of these
FIGURE 6. MCPG blocks LTP facilitation induced by the pri-ming LFS protocol. (A) In ventral hippocampal slices application ofMCPG (n) impairs LTP facilitation induced by LFS protocol (*).Averaged EPSP slope and average normalized Spike amplitude areplotted vs. time. Representative traces at indicated times are shownon top. The arrows indicate the time of HFS. (B) Representative
field potential recordings of spontaneous activity from stratum pyr-amidale in area CA1 at indicated steps. (C) Average power spectrumand (D) average integrated power obtained by recording spontane-ous activity at steps shown in B. MCPG decreases the level of powerfrequency range around 20–40 Hz induced by application ofDHPG. Asterisks in D denote a significance level of P < 0.001.
18 MAGGIO AND SEGAL
Hippocampus DOI 10.1002/hipo
oscillations showed a 48% 6 0.074% (P < 0.001) increase overbaseline. Forty minutes after thapsigargin, the pattern of activitydid not show any change. The peak band was 20–38 Hz with anaverage value of 28.656 1.03 Hz while the integrated power anal-ysis showed 41% 6 0.039% (P < 0.001) increase over baseline.
A second set of experiments was conducted to investigate theeffect of thapsigargin on the priming stimulation in VH (n ¼ 6,Fig. 9). LFS delivered to pathway 1 reliably induced LTD(0.7,073 6 0.05). An enhanced LTP was seen 20 min after LFS(1.96 6 0.063) as before (Fig. 9A). Following 30 min of thapsi-gargin application, a significant depression of the response wasproduced in the potentiated pathway (1.50 6 0.096, P < 0.05).
This response was comparable to the one obtained in the previ-ous experiments when thapsigargin was applied in presence ofDHPG. In addition, thapsigargin also impaired LTP facilitationdriven by the priming protocol. When applied in presence ofthapsigargin, the LFS could induce depotentiation (0.865 60.058) that did not significantly differ from the one induced inthe other pathway (P ¼ 0.069). However, no facilitation effectof the priming protocol could be detected in presence of thapsi-gargin. The magnitude of LTP (1.41 6 0.044) evoked in thiscondition significantly differed from the control (P < 0.001).These results indicate a major role of calcium stores in mediatingthe effects of the priming protocol on LTP facilitation.
FIGURE 7. mGluRs modulation of LTP in dorsal and ventralhippocampus, recorded in CA1 stratum radiatum. (A,B) DHPG hasdifferent effects in DH vs. VH. (A) In dorsal hippocampus, poten-tiation produced in presence of DHPG (*) did not differ from theone induced in the control pathway (n) while in VH (B), bathapplication of DHPG produced a significantly larger LTP (n) com-pared to control (*). (C) In ventral hippocampus, DHPG does not
produce potentiation of the response in absence of HFS (*). (D)Effect of CHPG on LTP induction in VH slices. Bath application ofCHPG does not induce a higher level of LTP (*) compared to con-trol (n). Averaged EPSP slope is plotted vs. time. In each experi-ment, representative traces at indicated times for DH and VHrespectively are shown on top.
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 19
Hippocampus DOI 10.1002/hipo

Application of thapsigargin did not produce any effect on theoscillatory pattern evoked by the priming protocol (Figs. 9C,D).LFS at one pathway induced prominent activity that peakedbetween 22 and 36 Hz with an average value of 24.66 6 1.4 Hzwhile the integrated power showed an increase of 46% 60.042% (P < 0.001) compared to baseline level. Interestingly, asin the case of DHPG, thapsigargin did not change this patternof oscillations (band peak between 21 and 35 Hz, average peakvalue of 24.42 6 1.3 Hz, 36% 6 0.02% in the integratedpower analysis, P < 0.01). These results, together with the effect
of thapsigargin on the oscillatory activity evoked by DHPG,suggest that intracellular calcium release does not play a signifi-cant role in sustaining LFS and DHPG induced field activity. Inaddition, they indicate that the development of such oscillatorypattern is not correlated with the ability to facilitate LTP.
The observation that thapsigargin is able to block LTP was al-ready reported (Harvey and Collingridge, 1992). Nevertheless, itwas never reported that a block of intracellular calcium storescould impair pre-established LTP. We explored further this obser-vation in ventral hippocampal slices using cyclopiazonic acid
FIGURE 8. Thapsigargin blocks the LTP facilitation inducedby DHPG. (A) In ventral hippocampal slices 40 min application ofthapsigargin (n) impairs LTP facilitation induced by DHPG appli-cation (*). Averaged EPSP slope and averaged normalized spikeamplitude are plotted vs. time. Representative traces at indicatedtimes are shown on top. The arrows indicate the time of HFS. (B)Representative field potential recordings of spontaneous activity
from stratum pyramidale in area CA1 at indicated steps. (C,D)Averaged power spectrum (C) and averaged integrated power (D)obtained by recording spontaneous activity at steps shown in B.Thapsigargin does not affect the level of power frequency rangearound 20–40 Hz induced by application of DHPG. Asterisks in Ddenote a significance level of P < 0.001 over control.
20 MAGGIO AND SEGAL
Hippocampus DOI 10.1002/hipo

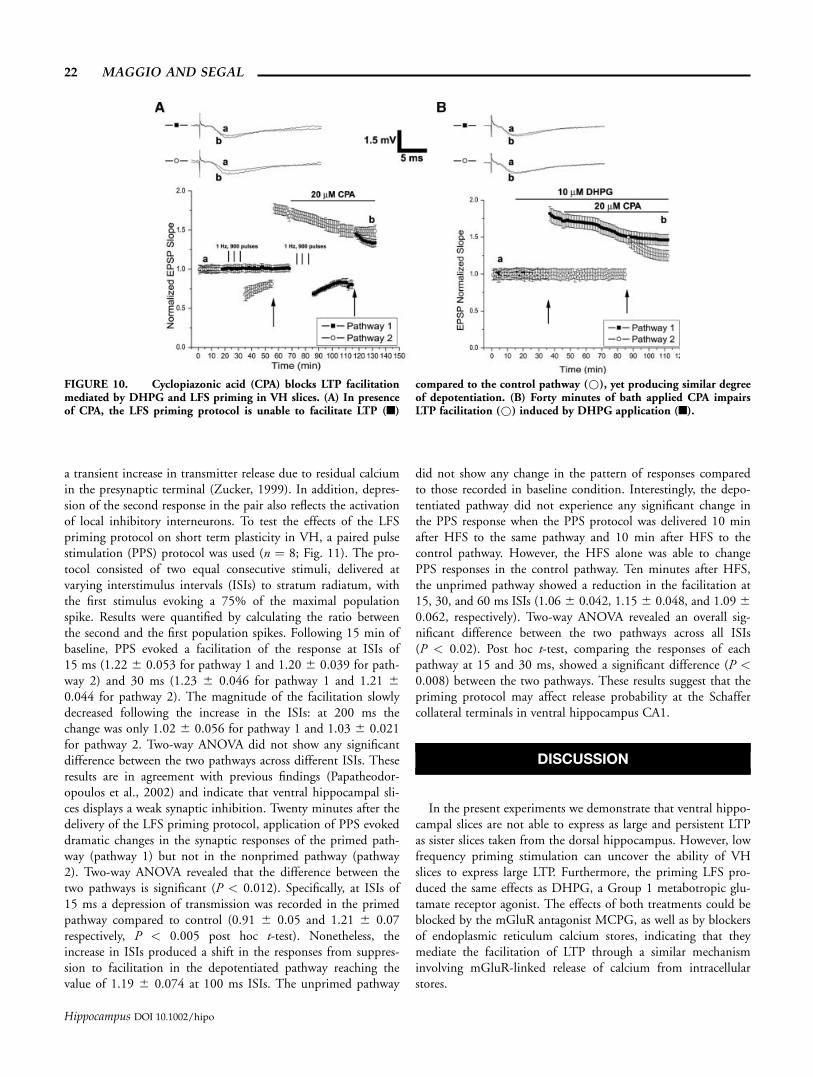
(CPA), another compound that blocks sarco-endoplasmic reticu-lum Ca2+ ATPases. Results are shown in Figures 10A,B. Bathapplication of 20 lM CPA was able to block the potentiation (10min after HFS, 1.74 6 0.09) facilitated by DHPG (Fig. 10B),which reached a value of 1.50 6 0.07 (P < 0.05), after 30 minfrom the application of CPA. In addition, like thapsigargin, CPAalso impaired the LTP induced in the other pathway in presenceof DHPG (10 min after HFS: 1.31 6 0.07, P < 0.01). Similarresults were obtained when CPA was bath applied after the HFSinduction of the LTP in the LFS primed pathway (Fig. 10A). Inthis case, the prior established LTP (10 min after HFS: 1.71 60.06) reached a plateau level of 1.55 6 0.04 (P < 0.05) 30 minafter CPA application. The HFS induction of LTP in a secondprimed pathway was also blocked following CPA wash (10 min
after HFS, 1.34 6 0.05, P < 0.01). Interestingly, CPA did notaffect the level of the depotentiation produced by the LFS(10 min after each LFS, 0.76 6 0.05 for the pathway 2 and0.776 0.03 for the pathway 1, P ¼ 0.72).
These results indicate that intracellular calcium stores are im-portant for the induction as well as maintenance of LTP facili-tated by DHPH or priming protocol stimulation in ventralhippocampus.
Priming Stimulation Affects Paired PulseResponses in Ventral Hippocampus
The change in response to the second of a pair of stimuli isconsidered to constitute a form of short term plasticity. It reflects
FIGURE 9. Thapsigargin blocks LTP facilitation induced byLFS priming protocol (A). (C,D) In ventral hippocampal slices 40min application of thapsigargin does not influence the level of
power frequency range around 20–40 Hz induced by LFS protocoldelivery. **P < 0.01 and ***P < 0.001.
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 21
Hippocampus DOI 10.1002/hipo
a transient increase in transmitter release due to residual calciumin the presynaptic terminal (Zucker, 1999). In addition, depres-sion of the second response in the pair also reflects the activationof local inhibitory interneurons. To test the effects of the LFSpriming protocol on short term plasticity in VH, a paired pulsestimulation (PPS) protocol was used (n ¼ 8; Fig. 11). The pro-tocol consisted of two equal consecutive stimuli, delivered atvarying interstimulus intervals (ISIs) to stratum radiatum, withthe first stimulus evoking a 75% of the maximal populationspike. Results were quantified by calculating the ratio betweenthe second and the first population spikes. Following 15 min ofbaseline, PPS evoked a facilitation of the response at ISIs of15 ms (1.22 6 0.053 for pathway 1 and 1.20 6 0.039 for path-way 2) and 30 ms (1.23 6 0.046 for pathway 1 and 1.21 60.044 for pathway 2). The magnitude of the facilitation slowlydecreased following the increase in the ISIs: at 200 ms thechange was only 1.02 6 0.056 for pathway 1 and 1.03 6 0.021for pathway 2. Two-way ANOVA did not show any significantdifference between the two pathways across different ISIs. Theseresults are in agreement with previous findings (Papatheodor-opoulos et al., 2002) and indicate that ventral hippocampal sli-ces displays a weak synaptic inhibition. Twenty minutes after thedelivery of the LFS priming protocol, application of PPS evokeddramatic changes in the synaptic responses of the primed path-way (pathway 1) but not in the nonprimed pathway (pathway2). Two-way ANOVA revealed that the difference between thetwo pathways is significant (P < 0.012). Specifically, at ISIs of15 ms a depression of transmission was recorded in the primedpathway compared to control (0.91 6 0.05 and 1.21 6 0.07respectively, P < 0.005 post hoc t-test). Nonetheless, theincrease in ISIs produced a shift in the responses from suppres-sion to facilitation in the depotentiated pathway reaching thevalue of 1.19 6 0.074 at 100 ms ISIs. The unprimed pathway
did not show any change in the pattern of responses comparedto those recorded in baseline condition. Interestingly, the depo-tentiated pathway did not experience any significant change inthe PPS response when the PPS protocol was delivered 10 minafter HFS to the same pathway and 10 min after HFS to thecontrol pathway. However, the HFS alone was able to changePPS responses in the control pathway. Ten minutes after HFS,the unprimed pathway showed a reduction in the facilitation at15, 30, and 60 ms ISIs (1.06 6 0.042, 1.15 6 0.048, and 1.09 60.062, respectively). Two-way ANOVA revealed an overall sig-nificant difference between the two pathways across all ISIs(P < 0.02). Post hoc t-test, comparing the responses of eachpathway at 15 and 30 ms, showed a significant difference (P <0.008) between the two pathways. These results suggest that thepriming protocol may affect release probability at the Schaffercollateral terminals in ventral hippocampus CA1.
DISCUSSION
In the present experiments we demonstrate that ventral hippo-campal slices are not able to express as large and persistent LTPas sister slices taken from the dorsal hippocampus. However, lowfrequency priming stimulation can uncover the ability of VHslices to express large LTP. Furthermore, the priming LFS pro-duced the same effects as DHPG, a Group 1 metabotropic glu-tamate receptor agonist. The effects of both treatments could beblocked by the mGluR antagonist MCPG, as well as by blockersof endoplasmic reticulum calcium stores, indicating that theymediate the facilitation of LTP through a similar mechanisminvolving mGluR-linked release of calcium from intracellularstores.
FIGURE 10. Cyclopiazonic acid (CPA) blocks LTP facilitationmediated by DHPG and LFS priming in VH slices. (A) In presenceof CPA, the LFS priming protocol is unable to facilitate LTP (n)
compared to the control pathway (*), yet producing similar degreeof depotentiation. (B) Forty minutes of bath applied CPA impairsLTP facilitation (*) induced by DHPG application (n).
22 MAGGIO AND SEGAL
Hippocampus DOI 10.1002/hipo

Previous studies (Papatheodoropoulos and Kostopoulos,2000a,b; Colgin et al., 2004) have demonstrated that ventralhippocampus CA1 (VH) has a decreased ability to express LTP.This deficit was also shown in vivo (Maruki et al., 2001), verify-ing that the reduced LTP is a genuine property of this region,and is not an artifact of the angle of slicing the VH, which maysevere afferent pathways to the recorded cells.
A transient exposure to higher DHPG concentrations (30–100 lM) has been reported to induce a presynaptic Ca2+ inde-pendent LTD in hippocampal slices (Fitzjohn et al., 2001;Rammes et al., 2003). In addition, Cohen et al., 1998 reportedthat a 10 min application of 20 lMDHPG significantly enhancesthe ability to induce LTP with a weak HFS. Aside from primingLTP, DHPG produced a transient depression of the synapticresponses during the drug application. On the other hand, DHPGhas been shown to induce a slow onset potentiation in the dentategyrus (DG) in vivo (Manahan-Vaughan and Reymann, 1996).Interestingly, in our experiments, 10 lM DHPG facilitated LTPbut also produced an increase in the baseline responses. Thisresponse was neither blocked by the receptor antagonist MCPG,
nor by thapsigargin, a blocker of calcium release. However, thiseffect was seen only with the population spike amplitude whilethe EPSP slope was not affected, suggesting that a different mech-anism, e.g., a reduction in GABAergic transmission, could medi-ate this effect. On the other hand, 10 lM DHPG induced adepression of the EPSP slope responses in stratum radiatum CA1of DH, indicating that Group 1 mGluRs differently modulatesynaptic transmission in DH and VH.
MCPG is known to block field oscillations mediated bymGluR agonists both in CA3 (Taylor et al., 1995) and in CA1(Boddeke et al., 1997). On the other hand, considerable contro-versy exists over the involvement of mGluRs in LTP induction/maintenance and whether MCPG can block LTP (Anwyl, 1999).Here we demonstrate that MCPG reliably blocked field oscilla-tions in the range of 20–40 Hz induced by DHPG or primingstimulation. In addition, MCPG was quite effective in blockingboth the induction and the maintenance of LTP in each protocolused to facilitate LTP in VH. In this respect our results supportthe hypothesis that stimulation of Group 1 mGluRs leads, viaphospholipase C, to the activation of PKC and CaMKII that alsorequire release of Ca2+ from intracellular stores (Bortolotto et al.,2005). PKC activation has been shown to be particularly impor-tant in LTP maintenance, which can be blocked by inhibitors ofPKC, and MCPG seems to prevent normal PKC modulation(Riedel et al., 1995). Our findings also suggest that mGluR1rather than mGluR5 is mediating the effect of DHPG since themGluR5 specific agonist, CHPG, failed to facilitate LTP in VH.
In our experiments, thapsigargin and CPA also blocked LTPinduction/maintenance. Besides the established role of calciumstores in LTP induction (Harvey and Collingridge, 1992), we de-monstrate here that calcium stores are probably involved also in theinitial phases of LTP maintenance. In our experiments, drug appli-cation started 30 min after HFS to the control pathway. A possiblerole of calcium stores in LTP maintenance could involve the activa-tion of protein kinases, such as PKC, that is modulated by intracel-lularly released Ca2+. Even though further experiments are requiredto investigate the action of thapsigargin and CPA, our findings areconsistent with the hypothesis that a permanent activation of PKCis required for the persistence of LTP (Malinow et al., 1989).
Previous studies suggest that the smaller amplitude LTP, evokedin VH CA1 is due to the presence of spontaneous sharp wave rip-ples (SPW-R) (Colgin et al., 2004). In our experiment, no SPW-R event was recorded. This could probably be due to the fact thatthe mechanical trauma due to the slice procedure severely impairsthe viability of granule cells and part of CA3 pyramidal cells, tworegions considered to be important for the generation of SPW-R.On the other hand, several studies on LTP in CA1 were per-formed using this preparation and have demonstrated that theconnectivity of CA3–CA1 synapses is very well preserved. Theseresults, together with our own, suggest that, regardless of theinfluence of the SPW-R, the lower magnitude LTP is a uniqueproperty of VH CA1 region of the hippocampus.
In several experiments we have demonstrated that LFS primingprotocol induced field oscillations in the range of 20–40 Hz.These oscillations resembled the ones evoked by DHPG in VH.Activation of type 1mGluR-induced field oscillations in CA3 and
FIGURE 11. LFS priming protocol affects paired pulse responses.Twenty minutes after delivery of priming protocol a depression oftransmission was recorded in the primed pathway compared to theunpotentiated pathway at 15 ms interstimulus interval (ISI). In addi-tion, the increase in ISI’s shifted the responses from suppression tofacilitation in the depotentiated pathway. Data are represented asmean 6 SEM. Representative traces at indicated ISI’s are shown ontop. Asterisks denote a significance level of P < 0.05.
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 23
Hippocampus DOI 10.1002/hipo

CA1 has been proposed (Palhalmi et al., 2004; Traub et al., 2004).It was thought that these oscillations in the gamma frequency (20–80 Hz) range are involved in LTP induction (Traub et al., 1998).In the VH slice preparation used to carry out the experimentsdescribed before, both DHPG treatment and LFS could mediatethe appearance of such oscillations together with the ability to facil-itate LTP. This effect could prove a possible role of this pattern ofactivity in LTP generation. However, thapsigargin could not blockthe field oscillation while reliably affecting LTP stability. This sug-gests that the relations between network activity and synaptic plas-ticity are more complicated than originally proposed.
In conclusion, we have demonstrated that the induction ofLTP is differently regulated in DH vs. VH. In addition, LFS pro-tocol as well as exposure to DHPG can facilitate LTP in VHthrough a mechanism that requires the release of calcium from in-tracellular stores.
Acknowledgments
We thank Dr. A. Avital for statistical advices and S. Sapoznikfor help with the preparation of the figures.
REFERENCES
Anderson WW, Collingridge GL. 2001. A data acquisition program foron-line analysis of long-term potentiation and other synaptic events.J Neurosci Methods 108:71–83.
Anwyl R 1999. Metabotropic glutamate receptors: Electrophysiologicalproperties and role in plasticity. Brain Res Brain Res Rev 29:83–120.
Bannerman DM, Yee BK, Good MA, Heupel MJ, Iversen SD, Rawlins JN.1999. Double dissociation of function within the hippocampus: Acomparison of dorsal, ventral, and complete hippocampal cytotoxiclesions. Behav Neurosci 113:1170–1188.
Bannerman DM, Rawlins JN, McHugh SB, Deacon RM, Yee BK, Bast T,Zhang WN, Pothuizen HH, Feldon J. 2004. Regional dissociationswithin the hippocampus–memory and anxiety. Neurosci BiobehavRev 28:273–283.
Behrens CJ, van den Boom LP, de Hoz L, Friedman A, Heinemann U.2005. Induction of sharp wave-ripple complexes in vitro and reor-ganization of hippocampal networks. Nat Neurosci 8:1560–1567.
Bliss TV, Collingridge GL. 1993. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 361:31–39.
Boddeke HW, Best R, Boeijinga PH. 1997. Synchronous 20 Hz rhythmicactivity in hippocampal networks induced by activation of metabo-tropic glutamate receptors in vitro. Neuroscience 76:653–658.
Bortolotto ZA, Collett VJ, Conquet F, Jia Z, vanderPutten H, CollingridgeGL. 2005. The regulation of hippocampal LTP by the molecular switch,a form of metaplasticity, requires mGlu5 receptors. Neuropharmacology49(Suppl 1):13–25.
Buzsaki G, Csicsvari J, Dragoi G, Harris K, Henze D, Hirase H. 2002.Homeostatic maintenance of neuronal excitability by burst dischargesin vivo. Cereb Cortex 12:893–899.
Cohen AS, Abraham WC. 1996. Facilitation of long-term potentiationby prior activation of metabotropic glutamate receptors. J Neurophy-siol 76:953–962.
Cohen AS, Raymond CR, Abraham WC. 1998. Priming of long-termpotentiation induced by activation of metabotropic glutamate recep-tors coupled to phospholipase C. Hippocampus 8:160–170.
Cohen AS, Coussens CM, Raymond RC, Abraham WC. 1999. Long-lasting increase in cellular excitability associated with the priming ofLTP induction in rat hippocampus. J Neurophysiol 82:3139–3148.
Colgin LL, Kubota D, Jia Y, Rex CS, Lynch, G. 2004. Long-termpotentiation is impaired in rat hippocampal slices that producespontaneous sharp waves. J Physiol 558(Part 3):953–961.
Colombo M, Fernandez T, Nakamura K, Gross CG. 1998. Functionaldifferentiation along the anterior-posterior axis of the hippocampusin monkeys. J Neurophysiol 80:1002–1005.
Degroot A, Treit D. 2004. Anxiety is functionally segregated within thesepto-hippocampal system. Brain Res 1001:60–71.
Dudek SM, Bear MF. 1993. Bidirectional long-term modification of syn-aptic effectiveness in the adult and immature hippocampus. J Neuro-sci 13:2910–2918.
Fitzjohn SM, Palmer MJ, May JE, Neeson A, Morris SA, Collingridge GL.2001. A characterisation of long-term depression induced by metabo-tropic glutamate receptor activation in the rat hippocampus in vitro.J Physiol 537(Part 2):421–430.
Harvey J, Collingridge GL. 1992. Thapsigargin blocks the induction oflong-term potentiation in rat hippocampal slices. Neurosci Lett 139:197–200.
Izaki Y, Takita M, Nomura M, Akema T. 2004. Effects of ventralhippocampal long-term potentiation and depression on the gamma-band local field potential in anesthetized rats. Exp Brain Res157:147–151.
Kjelstrup KG, Tuvnes FA, Steffenach HA, Murison R, Moser EI, MoserMB. 2002. Reduced fear expression after lesions of the ventral hip-pocampus. Proc Natl Acad Sci USA 99:10825–10830.
Malinow R, Schulman H, Tsien RW. 1989. Inhibition of postsynapticPKC or CaMKII blocks induction but not expression of LTP. Sci-ence 245:862–866.
Manahan-Vaughan D, Reymann KG. 1996. Metabotropic glutamate re-ceptor subtype agonists facilitate long-term potentiation within a dis-tinct time window in the dentate gyrus in vivo. Neuroscience 74:723–731.
Mannaioni G, Marino MJ, Valenti O, Traynelis SF, Conn PJ. 2001.Metabotropic glutamate receptors 1 and 5 differentially regulate CA1pyramidal cell function. J Neurosci 21:5925–5934.
Martin SJ 2001. Activation of metabotropic glutamate receptors indu-ces gamma frequency oscillations in the rat dentate gyrus in vivo.Neuropharmacology 40:634–637.
Maruki K, Izaki Y, Nomura M, Yamauchi T. 2001. Differences inpaired-pulse facilitation and long-term potentiation between dorsaland ventral CA1 regions in anesthetized rats. Hippocampus 11:655–661.
Mellentin C, Moller M, Jahnsen, H. 2006. Properties of long-term syn-aptic plasticity and metaplasticity in organotypic slice cultures of rathippocampus. Exp Brain Res 170:522–531.
Moser MB, Moser EI. 1998. Functional differentiation in the hippo-campus. Hippocampus 8:608–619.
Palhalmi J, Paulsen O, Freund TF, Hajos N. 2004. Distinct propertiesof carbachol- and DHPG-induced network oscillations in hippo-campal slices. Neuropharmacology 47:381–389.
Papatheodoropoulos C, Kostopoulos G. 2000a. Decreased ability of rattemporal hippocampal CA1 region to produce long-term potentia-tion. Neurosci Lett 279:177–180.
Papatheodoropoulos C, Kostopoulos G. 2000b. Dorsal-ventral differen-tiation of short-term synaptic plasticity in rat CA1 hippocampalregion. Neurosci Lett 286:57–60.
Papatheodoropoulos C, Asprodini E, Nikita I, Koutsona C, Kostopoulos G.2002. Weaker synaptic inhibition in CA1 region of ventral comparedto dorsal rat hippocampal slices. Brain Res 948:117–121.
Pothuizen HH, Zhang WN, Jongen-Relo AL, Feldon J, Yee BK. 2004.Dissociation of function between the dorsal and the ventral hippo-campus in spatial learning abilities of the rat: A within-subject,within-task comparison of reference and working spatial memory.Eur J Neurosci 19:705–712.
Rae MG, Irving AJ. 2004. Both mGluR1 and mGluR5 mediate Ca2+
release and inward currents in hippocampal CA1 pyramidal neu-rons. Neuropharmacology 46:1057–1069.
24 MAGGIO AND SEGAL
Hippocampus DOI 10.1002/hipo

Rammes G, Palmer M, Eder M, Dodt HU, Zieglgansberger W,Collingridge GL. 2003. Activation of mGlu receptors induces LTDwithout affecting postsynaptic sensitivity of CA1 neurons in rat hip-pocampal slices. J Physiol 546(Part 2):455–60.
Riedel G, Casabona G, Reymann KG. 1995. Inhibition of long-termpotentiation in the dentate gyrus of freely moving rats by themetabotropic glutamate receptor antagonist MCPG. J Neurosci 15:87–98.
Rose CR, Konnerth A. 2001. Stores not just for storage.intracellular cal-cium release and synaptic plasticity. Neuron 31:519–522.
Ross WN, Nakamura T, Watanabe S, Larkum M, Lasser-Ross N. 2005.Synaptically activated Ca2+ release from internal stores in CNS neu-rons. Cell Mol Neurobiol 25:283–295.
Squire LR, Stark CE, Clark RE. 2004. The medial temporal lobe. AnnuRev Neurosci 27:279–306.
Strange BA, Fletcher PC, Henson RN, Friston KJ, Dolan RJ. 1999.Segregating the functions of human hippocampus. Proc Natl AcadSci USA 96:4034–4039.
Taylor GW, Merlin LR, Wong RK. 1995. Synchronized oscillations inhippocampal CA3 neurons induced by metabotropic glutamate re-ceptor activation. J Neurosci 15:8039–8052.
Traub RD, Spruston N, Soltesz I, Konnerth A, Whittington MA,Jefferys GR. 1998. g-Frequency oscillations: A neuronal populationphenomenon, regulated by synaptic and intrinsic cellular processes,and inducing synaptic plasticity. Prog Neurobiol 55:563–575.
Traub RD, Bibbig A, LeBeau FE, Buhl EH, Whittington MA. 2004.Cellular mechanisms of neuronal population oscillations in the hip-pocampus in vitro. Annu Rev Neurosci 27:247–278.
Wilsch VW, Behnisch T, Jager T, Reymann KG, Balschun D. 1998.When are class I metabotropic glutamate receptors necessary forlong-term potentiation? J Neurosci 18:6071–6080.
Witter MP, Amaral DG. 2004. Hippocampal formation. In: Paxinos G,editor. The Rat Nervous System. Amsterdam: Elsevier. pp 637–687.
Zucker RS 1999. Calcium- and activity-dependent synaptic plasticity.Curr Opin Neurobiol 9:305–313.
UNIQUE REGULATION OF LTP IN VENTRAL HIPPOCAMPUS 25
Hippocampus DOI 10.1002/hipo





























