Embed Size (px)
Citation preview

FEMS Microbiology Reviews 87 (1990) 103-112 103 Published by Els~ier
FEMSRE 06162
Uncommon pathways of metabolism among lactic acid bacteria
Jack London
Laboratory of Microbial Ecology, National Institute of Dental Research, National Institutes of Health, Bethesda, MD, U.S.A.
Key words: Metabolism; Energy Production; Gluconate; Malate; Xylitol; Ribitol
1. SUMMARY
A small number of lactic acid bacteria possess the ability to derive energy from organic mole- cules not utilized by the vast majority of repre- sentatives of this large group of microorganisms. Thus, strains of Lactobacillus casei and enteroc- occi readily grow at the expense of substrates such as gluconate, malate and pentitols. Transport of gluconate and pentitols is catalysed by phos- photransferase sytems unique to these bacteria. Similarly, ",he initial steps in peL, titol dissimilation are mediated by enzymes found only i~ Lb. casei and Streptococcus avium.
2. INTRODUCTION
less impressive than that round among the enteric bacteria [1,2]. Collectively, these bacteria have the ability to metabolize a large r~amber of mono- and oligosaccharides, polyalcohols, aliphatic com- pounds, mono-, di- and tricarboxylic acids and some amino acids. Synthetically, they possess the potential to manufacture a host of complex carbohydrates (expressed as cell wall antigens or loosely associated slime matrices) and, in at least one instance, some rare amino acids. Past contri- butions to this series of symposia have dealt with the general issues of energy metabolism, transport and metabolic regulation of the more familiar sugar substrates. This paper will emphasize re- search on some uncommon and ~musual metabolic pathways that have been observed in a restricted number of lactic acid bacteria.
Individually, most of the species that comprise the lactic acid bacteria utiDze a relatively re- stricted number of organic molecules as carbon and energy sources. However, taken in their en- tirety, this group of Gram-positive bacteria which includes the genera, Lactobacillus, Streptococcus, Pediococcus, Leuconostoc, Lactococcus and Enter- ococcus, possess a metabolic po:enfial only slightly
Correspondence to: J. London, Building 30, Room 314, Labora- tory of Microbial Ecology, National Institute of Dental Re- search, National Institutes of Health, Bethesda, MD, U.S.A.
3. GLUCONATE
It is a generally known and widely accepted observation that many, if not all, strains of Enter- ococcus (Streptococcus) faecalis readily grow at the expense of gluconate, althc.agh Bergey's Manual of Systematic Bacteriology [1] makes no mention of this property. Many strains of Lactobacillus casei possess the same trait (J. London, unpub- lished observations). Studies with E. faecalis [3] established that growth on gluconate induced a NAD+-dependent gluconate-6-P dehydrogenase
0168-6445/90/$03.50 © 1990 Federation of European Microbiological Societies
104
(6GPdh) 1 which coexisted with a constitutively synthesized NADP+-dependent 6GPdh 2 [4].
6GP + NAD + --* RuSP + CO 2 + NADH (1)
6GP + NADP +-* Ru5P + CO 2 + NADPH (2)
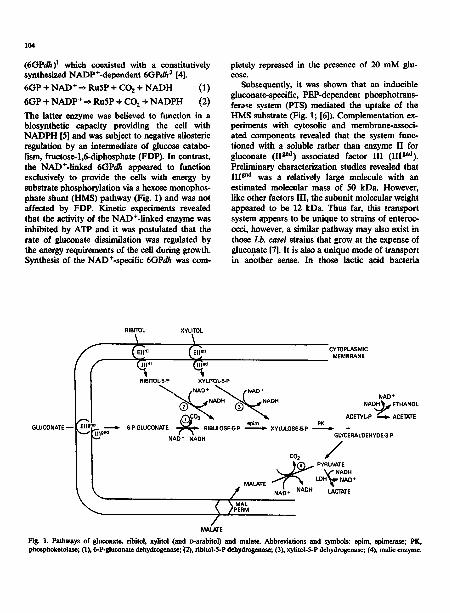
The latter enzyme was believed to function in a biosynthetic capacity providing the cell with NADPH [5] and was subject to negative allosteric regulation by an intermediate of glucose catabo- lism, fructose-l,6-diphosphate (FDP). In contrast, the NAD+-linked 6GPdh appeared to function exclusively to provide the cells with energy by substrate phosphorylation via a hexose monophos- phate shunt (HMS) pathway (Fig. 1) and was not affected by FDP. Kinetic experiments revealed that the activity of the NAD+-linked enzyme was inhibited by ATP and it was postulated that the rate of gluconate dissimilation was regulated by the energy requirements of the cell during growth. Synthesis of the NAD+-specific 6GPdh was com-
pletely repressed in the presence of 20 mM glu- cose.
Subsequently, it was shown that an inducible glucunate-specific, PEP-dependent phosphotrans- ferase system (PTS) mediated the uptake of the HMS substrate (Fig. 1; [6]). Complementation ex- periments with cytosolic and membrane-associ- ated components revealed that the system func- tioned with a soluble rather than enzyme II for gluconate (II gnd) associated factor III (IIIgnd). Preliminary characterization studies revealed that III gnd was a relatively large molecule with an estimated molecular mass of 50 kDa. However, like other factors Ill , the subunit molecular weight appeared to be 12 kDa. Thus far, this transport system appears to be unique to strains of enteroc- occi, however, a similar pathway may also exist in those Lb. casei strains that grow at the expense of gluconate [7]. It is also a unique mode of transport in another sense. In those lactic acid bacteria
RIBITOL XYLITOL
c==c
RIBtTOL-5-P XYLITOL-5-P
NADH NADH NADH~ ETHANOL
.(~.C0 2 ~'~ ~'~ ACETYbP ~ ACETATE . _ ~ epim PK
GUJCONATE~ ~ ~ ~ 6-P-GLUCONATE RIBULOSE-5-P ~ XYLULOSE-5-P I~ + "~ 1119~'~ GLYCERALDEHYDE-3-P NAD* NAOH
co, /" It ( ~ . P'~.UVATE
~ " ~ , '~f" NADH
\ MAI. ~ . ~ /PERM
MALATE
Fig. I. Pathways of gluconate, ribito], xylltol (and D-arabitol) and malatc. Abbreviations and symbols: epim, epimeras¢; PK, phosphoketolase; (1), 6-P-81uconate dehydrogenase; (2), ribitol-5-P dehydrogenase; (3), xylitol-5-P dehydrogenase; (4), medic enzyme.

capable of utilizing HMS pathway sugars, i.e. ribose, xylose and arabinose, the existence of specific pentose permeases is inferred from the presence of substrate-specific kinases detected in cell free extracts of these bacteria [8].
4. PENTITOLS
The utilization of five-carbon polyols, i.e. ribi- tol, xylitol and v-arabitol, is a relatively rare prop- erty occurring only in a handful of procaryotes and eucaryotes [9,10]. Strains of Streptococcus avium grow ~t the expense of the three polyal- cohols listed ,~bove while certain strains of Lb. casei utilize either ribitol or xylitol and ~arabitol. The enzymatic mechanisms by which these sub- strates are taken up and the initial steps in their catabolism are unique and have been observed only with two species of lactic acid bacteria. Sub- strate-specific phosphotransferase systems import ribitol or xyhtol and v-arabitol into the cell con- verting the substrates to their respective pentitol- 5-phosphates in the process (Fig. 1; [liD. In the cell, NAD+-specific dehydrogenases oxidize the pentitol-5-phosphates to their corresponding pentulose-5-phosphates [12]. At this point, the pathways merge with routes usually employed by the lactic acid bacteria to dissimilate pentoses and gluconate (Fig. 1).
Mechanistically, the pentitol PTS were very similar to the hexitol PTS used by a large variety of microorganisms to translocate mannitol [13-15]. However, the ribitol and xylitol transport systems were not only highly specific for their respective substrates, they also failed to import mannltol [11]. Because the xylitol PTS proved to be readily amenable to biochemical dissection, studies were concentrated on characterizing the I l l xt: compo- nent of this system. The purified III xtt was a slightly acidic protein that had a molecular mass of 12 kDa [16]. Monospecific rabbit antisera pre- pared against this purified soluble PTS component reacted with lII ~*l from other strains of lactoba- cilfi and S. avium in immunodiffusion experi- ments. This observation suggests that the anti- 8enic structure (and some portion of the amino
105
acid sequence) of this molecule have been struct- urally conserved. It has not yet been determined which of the five histidine residues found in Lb. casei III xtl catalyse transfer of the phosphate moiety form HPr to EII xtj [17,18].
The rtl-5-P and xtl-5-P NAD+-specific dehy- drogenases were purified from Lb. casei strains CI 16 and CI 83, respectively [12]. Substrate specific- ity was established with chemically or enzymati- eally synthesized ribitol-5-P (rtl-5-P), xylitol-5-P (xtl-5-P) and D-arabitol-5-P (atl-5-P). Ribitol-5-P dehydrogenase (Rtl-5-Pdh 3) reduced NAD + only in the presence of rtl-5-P while xylitol-5-P dehy- drogenase (xtl-5-Pdh 4) reduced NAD + with both atl-5-P and xtl-5-P as substrata.
rtl-5-P + NAD + --, ru-5-P + NADH (3)
xtl-5-P + NAD +--* xu-5-P + NADH (4)
In the native state, rtl-5-Pdh and xtl-5-Pdh exist as a dimer and tetramer, respectively, and migrate like proteins with estimated molecular masses of 115 kDa and 180 kDa. Like the III xt] of this group of bacteria, immunological cross reactivity be- tween the streptococcal and lactobacillus xtl-5oPdh was readily demonstrated. However, none was ob- served between the rtl-5-P and xtl-5-P dehydro- genases in either intra- and intergeneric cross matches.
Those strains of Lb. casei that grow at the expense of ribitol simultaneously induce a func- tional xylitol PTS, however, they have apparently lost the ability to synthesize a xtl-5-Pa~ and can- not further metabolize the phosphorylated xylitol intermediate [11,19]. The inability to metabolize intracellular xtl-5-P had serious consequences for growing cells. The addition of xyfitol to an ac- tively growing culture (with ribitiol as energy source) quickly leads to the PEP-dependent accu- mulation of high levels of non-metabolizable xtl-5- P. However, the microorganisms also possess a phosphohydrolase activity of broad specificity that acts on a number of phosphorylated intermediate products including xtl-5-P [20]. The net effect of these two activities, xyfitol transport and phos- phohydrolase, is the creation of a futile cycle that dissipates the cells' energy by continuously deplet- ing its PEP pool [21]. This effect is manifested by

106
a cessation of growth at the expense of ribitol; metabolism of this substrate merely primes the PEP dissipating pump. Ultimately, the cells over- came the xylitol-induced bacteriostasis by re- pressing synthesis of the EII xtl of the xylitol I r i s via a mechanism which is not yet understood. Repression of the Eli xu remained effective only so long as xylitol was present in the growth medium. Growth of repressed cells at the expense of ribitol, but in the absence of xylitol, returned the micro- organism to its xylitol-sensitive ground state. Xylitol-driven futile cycles reported in other streptococci have been reviewed elsewhere [10]. The action of the xylitol futile cycle is similar to the futile cycle driven by 2-deoxyglucose in strains of Streptococcus pyogenes and Lactococcus lactis [22,23].
Studies with chemically mutated isolates of Lb. casei strain C183 were used to ascertain how this microorganism regulates synthesis of the proteins that comprise the xylitol pathway, namely EII xtl, lI l xtl and xylitol-5-P dehydrogenate [10]. Pre- liminary experiments revealed that ribose and glu- conate, or their metabolic intermediate products, served as gratuitous inducers of the xylitol path- way. Two classes of mutations were observed; one group, which expressed the phenotype x t l - /a t l - , was unable to grow at the expense of either xylitol or D-arabitol. '12ds group consisted of mutants in which one of the three xylitol pathway compo- nents, EII xtl, III xtl or xti-5-Pdh, was not synthe- sized in an active form and these isolates were classified as structural gene mutants. The second major class of mutants exhibited one of the fol- lowing phenotypes: x t l - /a t l +, xtl+/atl - or x t l - /a t l - . The isolates possessed all three compo- nents of the pathway when cultivated on ribose or gluconate and would grow on either pentitol if sufficient gratuitous inducer was added to the medium. The multiplicity of inducing substrates, i.e. xylitol, D-arabitol, ribose and gluconate, and the L~olation of the three phenotypic expression groups described above was rationalized by pos- tulating that the cells' genome contained three regulatory genes whose products function in a positive fashion upon interacting with xtl-5-P, atl- 5-P or xu-5-P.
5. MALATE
With few exceptions, most lactic acid bacteria are capable of catalysing a malolactate fermenta- tion 5 in which malic acid (a dicarboxylic acid) is converted to lactate [24,25].
malate - , lactate + CO 2 (5)
Although the reaction is generally thought to be non-energy yielding process, the export of lactate in symport with proton(s) may establish an energy yielding proton motive gradient across the cells' membrane similar to that described with Lacw- coccus lactis ssp. cremoris [26,27]. Several malo- lactic enzymes have been either partially purified or purified to homogeneity and characterized. Studies with the partially purified preparation from Leuconostoc mesenteroides [28] and the pure preparation from Lb. plantarum [25] indicated that the malolactate fermentation is not catalysed by a heterogeneous enzyme complex performing a variety of individual steps, but rather that a single NAD+-dependent protein mediates the entire re- action. Because free NADH is not produced by the Lb. plantarum enzyme, the reaction appears to be fundamentally decarboxylative in nature [25,28]. Immunological studies with anti-Lb. plantarum malie enzyme suggests that the anti- genic structure of the protein is conserved within lactic acid bacteria. Recently, a L, iactis gene encoding a protein that acts as a positive regulator for inductic, i~ of the malolaedc enzyme was de- scribed [29]. Introduction of the cloned gene into mutants that were incapable of catalysing the malolactic reaction but that still possessed both the malate permease system and malolactic en- zyme, restored activity completely.
Strains of E. faecalis and Lb. casei differ from other lactic acid bacteria by virtue of their ability to utilize L-malate as a growth substrate. Studies established that this dicarboxylic TCA cycle inter- mediate product was dissimilated by an inducible system consisting of a specific permease which transported malate into the cell and a NAD +- specific 'malic enzyme' (eqn. 6) that oxidized the substrate to pyruvate and carbon dioxide [30,31].

malate + NAD +- , pyruvate + CO 2 + NADH
(6)
Energy was presumably derived from the conver- sion of pyruvate to acetate and carbon dioxide. Aerobically, acetate was the major end product while anaerobic growth resulted in the production of almost equimolar amounts of acetate and ethanol.
Synthesis of malate permease and mafic en- zyme were repressed by the addition of glucose at concentrations greater than 5 mM. However, the addition of 10 mM glucose to resting cells had no effect on the transport of the dicarboxylic acid suggesting that neither components of the glucose PTS nor intermediate products of the Embden- Meyerhof pathway affected the activity of this extant process, i.e. that glucose prevented the op- eration of the permease [32]. However, electro- phorefically homogeneous malic enzyme isolated from both E. faecalis and Lb. casei was inhibited by Embden-Meyerhof pathway intermediates FDP and 3-phosphoglycerate as well as ATP [33,34]. Inhibition by the two intermediate products was believed to provide a means of regulating carbon flow, energy production and NADH levels in the presence of two substrates like glucose and malate. Inhibition by ATP supposedly furnished the malate pathway with a means regulating energy production and carbon flow. Malic enzyme from both microorganisms exhibited very similar physi- cal and kinetic properties and shared a relatively high degree of immunological homology [35]. It would be of interest to learn whether a single permease imports malate for both the malolactic and mallc enzymes.
6. LACTATE
The lactic acid bacteria are defined, in part, by their ability to produce lactate from fermentable or oxidizable substrates. Yet, some members of this group are capable of using lactate as a source of energy; and enterococci exhibit a low, but measurable, amount of growth at the expense of
107
this metabolic product [36]. One of these organisms, a strain of E. faecalis, produces a lactate oxidase activity when cultivated on a variety of substrates under oxidative conditions. Expression of lactate oxidase activity was greatest when glycerol and pyruvate were used .as growth substrates. Glucose and gluconate permitted in- duction of only one-fifth of the enzyme activity observed with the three carbon compounds while growth on ribose resulted in the synthesis of a level of lactate oxidase between that observed with the three- and six-carbon substrates. For reasons that are still not clear, fructose completely re- pressed expression of the lactate oxidase activity. Although other species of streptococci appear to be capable of metabolizing lactate [37] or deriving energy from lactate symport [26,38], growth at the expense of lactate per se is limited to the one report cited above.
7. CARBOXYETHYL AMINO ACIDS
Recently, a most interesting series of papers have appeared in the literature which further at- tests to the metabolic diversity among the lactic acid bacteria. Strains of lactococci have been shown to produce unusual amino acid derivatives from ornithine and lysine. Analysis of the in- traceUular amino acid pool of Lactococc~.s lactls strain 133 during growth in 'spent' medium, re- vealed high levels of a neutral compound, tenta- tively identified as ' valine' [39]. This new amino acid, which was produced concomitant with the conversion of arginine to ornithine by cultures of L lactis, was subsequently isolated and identified as NS-(carboxyethyl)-ornithine [40]. Studies with L lactis strain K1 that were designed to elucidate the mechanism by which NS-(carboxyethyl)- ornithine was synthesized led to the isolation and identification of a second hitherto unknown amino acid, N6-(carboxyethyl)-lysine [41]. Fig. 2 shows the formulation for the synthesis and structure of N5-(carboxylethyl) ornithine (the lysine derivative has a similar structure). Naturally occurring carboxyethyl derivatives of amino acids are rare in nature (for review see [42]). Within the biosphere,

108
they appear, thus far, to be produced only by certain marine invertebrates [~3,44] and constitute the opines produced in crowri gall of plants in- fected by species of Agrobacterium [40,42].
Biosynthesis of the carboxyethyl derivatives of ornithlne and lysine occurs by a relatively simple NADPH-dependent reductive condensation be- tween the respective amino acid and pyruvate [45]. Both reactions are catalysed by a single enzyme, a NADP+-specific NS-(L-l-carboxyethyl)-L-orni thine oxidoreductase (EC. 1.5.1.24). The native enzyme is a tetramer consisting of four 38-kDa subunlts and is present in the cell at very low concentrations [45]. At present, the 'synthase' ap- pears to be restricted to strains of L. lactis and has not been observed in extracts of other species of lactic acid bacteria [46]. Although the enzyme can provide a means of reoxidizing NADPH, this may not be its primary function in the cell. An opine dehydrogenase of relatively broad specific- ity produced by a species of Arthrobacter con- denses the amino acids methionlne and isoleucine and with pyruvate to form the corresponding N- [1-R-(carboxy)ethyl] derivatives [47]. However, since this strain of Arthrobacter grows at the ex- pense of opines, presumably of plant origin, the function of the enzyme appears to be degradative rather than biosynthetic. The function of the lactococcal enzyme and the role(s) of the carbo- xethyl derivatives of ornithine and lysine in this organism's metabolism have yet to be established.
8. REGULATION OF MALATE AND PENTI- TOL METABOLISM BY LACTIC ACID BAC- TERIA: CONUNDRUMS AND RATIONALI- ZATIONS
The fine control systems observed in Staphylo- coccus aureus [48] and Gram-negative bacteria [49] which inhibit transport and metabolism of glycerol or glycerol and lactate metabolism, respectively, when a Errs sugar is present do not appear to operate so effectively in those lactic acid bacteria that metabolize pentitols and malate. Growing cultures of Lb. casei and E. faecalis have been observed to utilize pentitols and glucose simulta- neously, while resting cells of E. faecalis transport malate in the presence of bexose. The transport of pentitols and malate is mediated by a I r i s and permease, respectively, yet their uptake by grow- ing or resting cell suspensions was virtually unaf- fected by the addition of 10 mM glucose [19,20]. Uptake and, in the case of the pentitols, ntiliza- tion of these substrates, ceased only after several generations of growth in the presence of glucose. These data suggest that some glycolytic inter- mediate triggers the formation of a repressor of biosynthesis of both the transport enzymes and dehydrogenases. In time, the various activities were diluted to the point where they became non-func- tional. A similar phenomenon was observed dur- ing the operation of the xylitol futile cycle in Lb. casei cells in which the pentitol transport system
o H H
~ - o~ H÷ H~O ~ _ ~
Omithine ?ls.(L.l.carboxyethyl)- L-omithine
Fig. 2. Structure of NS-(carboxyethyl) omithine and equation describing the synthesis of the amino acid derivative (figure courtesy of Dr. J. Thompson).

drained away significant amounts of the cells' energy supply generated during growth on glucose [20]. Glucose (10 mM) had no apparent effect on xylitol uptake and the functioning futile cycle (which consists of the xylitol PTS and a phoso phohydrolase) reduced the growth rate markedly. Again, it appears that the primary source of con- trol of the futile cycle enzymes is exerted at the level of transcription because the inhibitory effects of xylitol were not overcome during glucose-sup- ported growth over the course of the experiment [20] and growth at the expense of ribitol was inhibited for as long as several days [19].
The xylitol-5-P dehydrogenase also remained detectable and functional in cells long after glu- cose had been added to the culture. However, malic enzyme was sensitive to glycolytic inter- mediates and conversion of malate to pyruvate and CO 2 could theoretically be inhibited by inter- mediate products of glucose catabolism [32]. The ribitol-5-P, xylitol-5-P and 6-phosphogluconate dehydrogenases differ from malic enzyme in that they are not subject to allosteric inhibition by glycolytic intermediates. The NAD +-linked malic enzyme clearly belongs to that of the group of catabolic and biosynthetic enzymes under the con- trol of specific glycolytic intermediates. It would appear that carbon channeling from gluconate, xylitol or ribitol into the Embden-Meyerhof path- way at the level of triose phosphate and acetyl phosphate is readily accommodated by streptococ- cal or lactobacillus cells metabolizing hexose simultaneously. This is probably due to the fact that only half of the carbon from ribitol and 25~ of the carbon from xylitol is processed via LDH. Moreover, excess reducing equivalents can be di- rected into the production of ethanol under anaerobic conditions. This does not appear to be the case for carbon derived from malate which enters the glycolytic pathway at the level of pyru- vate. In the absence of an external electron accep- tor like oxygen, the malate-derived pyruvate may compete with the pyruvate originating from glu- cose for a limited number of reducing equivalents and upset the ceils' metabolic balance. However, certain lactic acid bacteria apparently can coordi- nate the efficient utilization of two substrates simultaneously, at least for limited periods of time.
109
This property may endow these bacteria with a selective advantage over more rigorously regulated bacteria in an environment rich in organic nutri- ents.
9. COI~ICLUDING REMARKS
One of the primary purposes of this presenta- tion was to review some of the rarer metabolic traits and pathways of lactic acid bacteria that had been defined biochemically and enzymatically over the past 20 years. The genetic system responsible for the biosynthesis of the carbox-yethyl amino acid derivatives is currently under vigorous inves- tigation at the National Institute of Dental Re- search. This is not the case for the gluconate, pentitol or malate dissimilatory systems. Probes are either available or can readily be obtained for cloning the genes which encode proteins of the latter two systems. Such studies are vital to under- standing how these genes are arranged and how transcription is regulated in the lactic acid bacteria. It is hoped that this treatise will serve as a re- minder and impetus for future investigations of these pathways 'less travelled' in the lactic acid bacteria.
ACKNOWLEDGEMENTS
This paper is dedicated to my friend and col- league, Morrison Rogosa, who died March 28, 1989. His fife-long devotion to defming the lactic acid bacteria inspired his co-workers. I am also grateful to Drs. John Thompson, Paul Kolen- brander and Stan Robrish for their helpful criti- cism and comments.
REFERENCES
[1] Schleifer, K.H. (1986) Gram-positive cocci. In: lk-rgey's Manual of Systematic Bacteriology, Vol. 2 (Sheath, P.H.A, Mail', N.S., Shalpe, M.E. and Holt, J.G, Eds.), pp. 999- 1103. Williams and Wilkins, Baltimore.
[2] Kandler, O. and Weiss, N. (1986) Regular, non-spore- forming Gram-positive rods. In: Bergey's Manual of Sys- tematic Bacteriology, %ol. 2 (Sneath, P.H.A., Mair, N.S.,

110
Shari~ M.E. and Holt, J.G, Eds.), pp. 1208-1234. Wil- liams and Wilkins, Baltimore.
[3] Brown, A.T. and Wittanberger, C.L. (1972) Induction and regulation of a nientinamide adenine dinucleotide-specific dehydrogenase in Streptococcus faecalis. J. Bacteriol. 109, 106-115.
[4] Bridges, R.B., Palumbo, M.P. and Wittenberger, C.L. (1975) Purification and properties of an NADP-specific 6-phosphogluconate dehydrogenese from Streptococcus faecalis. J. Biol. Chem. 250, 6093-6100.
[5] Wittenberger, C.L. and Crow, V.L. (1976) Regulation of carbon flow within and between metahofic pathways in certain species of Streptococcus. In: Mi~obiulogical Aspects of Dental Caries (a supplement to Microbiologi- cal Abstracts), Vol. 2, pp. 617-629.
[6] Bernsmann, P., Alpert, C.A., Muss, P., Deutscher, J. and Hengstenberg, W. (1982) The bacterial PEP-dependent phosphotransferase system mechanism of gluennate phos- phorylation in Streptococcus foccalis. FEBS Left. 138, 101-103.
[7] London, J and Chace, N.M. (1979) Pentitol metabolism in Lactobacillus casei. J. Bacteriol. 140, 949-954.
[8] Heath, E.C., Hurwit~ J., Horecker, B.L. and Ginsburg, A. (1958) Pentose fermation by Lactobacillus plantarum. J. Biol. Chem. 231,1009-1031.
[9] Mortlock, R.P. (1976) Catabolism of unnatural carbo- • hydrates by micro-organisms. In: Andvances in Microbial Physiology, Vol 13, pp. 2-53 (Rose, H.A. and Tempest, D.W., Eds.), Academic Press, New York.
[10] London, J. (1987) Pentitol transport and metabolism in lactic a~'id bacteria. In: Sugar Transport and Metabolism in Gi'~m-Positive Bacteria, pp. 150-164 (Reizer, J. and Peterkofsky, P., Eds.), Ellis Horwood Ltd., Chichester.
[ll] London, J. and Chace, N.M. (1977) New pathways for the metabolism of pentitols. Prof. Natl. Acad. Sci. 74, 4296- 4300.
[12] Hausman, S.Z. and London, J. (1987) Purification and ch-~racterization of a ribitol-5-phosphate and xylitul-5- phosphate dehydrogenases from strains of Loctobacillus casei. J. Bacteriol. 169,1651-1655.
[13] Berkowitz, D. (1971) D-mannitol utilization in Salmonella typhimurium. J. Bacterioi. 105, 232-240.
[14] Patni, N.J. and Alexander, J.K. (1971) Catabolism of fructose and mannitol in Clostridum thermocellum: pres- ence of phosphoenol-pyruvate:fructose phos- photransferase, fructose-l-phosphate kinase, phos- phoenoipyruvate:mannitol phosphotransferase and man- nitoi-l-phosphate dehydrogenase in cell extracts. J. Bacteriol. 105, 226-231.
[15] Jacobson, G.R., Lee, C.A. and Saier, M.J. Jr. (1979) Purification of the mannitol-specific enzyme II of the £scherichia coli phosphoenolpyruvate:sugar phos- photransferase system. J. Biol. Chem. 254, 249-252.
[16] London, J and Hausman, S.Z. (1983) Purification and characterization of the III xti phospho-carrier protein of the phosphoenolpyruvate-dependent xylitol:phospho- transferase found in Lactobacillus casei C!83. J. Bacterioi. 156, 611-619.
[17] Hays, J.B., Simoni, R.D. and Roseman, S. (1973) Sugar transport. V. A trimeric lactose-specific phosph~arrier protein of the Staphylococcus aureus phosphotransferase system. J. Biol. Chem. 248, 941-956.
[18] Deutscher, J.K., Beyreuther, H.M., Sobek, K., Stuber, K. and Hengstenberg, W. (1982) Phosphoenolpyruvate-de- pendent phosphotransferase system of Siaphylococcas aureus: factor lII I~, a trimeric p h o s p h ~ e r protein that also acts as a phase transfer catalyst. Biochemistry 21, 4867-2617.
[19] London, J. and Hausman, S.Z. (1982) Xylitol-mediated transient inhibition of ribitol utilization by Lactobocillus casei. J. Bactariul. 150, 657-661.
[20] Hansman, S.Z., Thompson, J. and London, J. (1984) Futile xylitol cylcle in Lactobacillus casei. J. Bacterinl. 160, 211- 215.
[21] London, L, Hausman, S.Z. and Thompson, J. (1985) Characterization of a membrane-regulated sugar phos- phate phosphohydrolase from Lactobocillus casei. J. Bacterinl. 163, 951-956.
[22] Reizer, J. and Panos, C. (1980) Regulation of D-galacto- side phosphate accumulation in Streptococcus pyogenes by an expulsion mechanism. Prof. Natl. Acad. Sei. USA 77, 5497-5501.
[23] Thompson, J. and Chassy, B.M. (1982) Novel phos- phocnulpyruvate-dependent futile cycle in Streptococcus lactis: 2-D-deoxyglucose uncouples energy production from growth. J. Bacteriol. 156, 70-80.
[24] Konkec, R.E. (1974) Malolactic fermentation and wine- making, Adv. Chem. Res. 137,151-170.
[25] Caspritz, G. and Radler, F. (1983) Malolactic enzyme of Lactobacillus plantarum. J. Biol. Chem. 258, 4907-4910.
[26] Ten Brink, B., Otto, R., Hansen, U.P. and Konings, W.N. (1985) Energy recycling by lactate efflux in growing and non-growing cells of Streptococcus cremoris. J. Bacteriol. 162, 383-390.
[27] Renault, P., Galilardin, C. and Heslot, H. (1988) Role of the malolactic fermentation in lactic acid bacteria. Bin- chemie 70, 375-379.
[28] Lonvaod-Funel, A. and Stransser de Saad, A.M. (1982) Purification and properties of a malolactic enzyme from strains of l-~conostoc mesenteroides isolated from grapes. Appl. Environ. Microbiol. 43, 357-361.
[29] Renault, P., Galllardin, C. and Heslot, H. (1989) Product of Lactococcus Ioctis gene required for malolactic fermen- tation is homologous to a family of positive regulators. J. Bacteriol. 171, 3008-3114.
[30] London, J. and Meyer, E.Y. (1969) Malate utilization by a group D Streptococcus: physiological properties and puri- fication of an inducible medic enzyme. J. Bacteriol. 98, 705-711.
[31] London, J., Meyer, E.Y. and Kulczyk, S. (1971) Compara- tive biochemical and immunological study of malic en- zyme from two species of lactic acid bacteria: evolu- tionary implications. J. Bacteriol. 106,126-137.
[32] London, J. and Meyer, E.Y. (1970) Malate utilization by a group D Streptococcus; regulation of malic enzym,.-

synthesis by an inducible malate permease. J. Bacteriol. 102,130-137.
[33] London, J. and Meyer, E.Y. (1%9) Malate utilization by a group D Streptococcus. II. Evidence for allosteric inhibi- tion of an inducible malate dehydrogenase (decarboxylat- in8) by ATP and glycolytic intermediate products. Bin- chim. Biophys. Acta 178, 205-212.
[34] London, J., Meyer, E.Y. and Kulczyk, S. (1970) Allosteric control of a Lacwbocillus malate dehydsogenase (de- carboxylating) by two glycolytic intermediate products. Biochim. Biophys. Acta 212, 512-514.
[35] London, £, Meyer, E.Y. and Kulczyk, S.IL (1971) Detec- tion of relationships between Streptococcus faocalis and Lactobocillus casei by immunological studies with two forms of malic enzyme. J. Bactc:fiol. 108,1%-201.
[36] London, J. (1968) Regulation and function of lactate oxidation in Streptococcus faocium. J. Bacterini. 95,1380- 1387.
[37] Hu, G. and Sandham, H.J. (1972) Streptococcal utilization of lactic acid and its effect on pH. Arch. Oral Biol. 17, 729-743.
[38] Distler, W., Kagermcier, A., Hickal, R. and Kroancke, A. (1989) Lactate influx and efflux in the 'Streptococcus romans' group and Streptococcus sanguis. Caries Res. 23, 252--255.
[39] Thompson, J., Curtis, M.A. and Miller, S.F.P. (1986) NS-(1-ca,'boxyethyl)-ornlddne, a new amino acid from the intracellular pool of Streptococcur lactis. J. Bacteriol. 167, 522-529.
[40] Miller, S.P.F. and Thompson, J. (1987) Biosynthesis and s~reuchemical confi$uration of NS-(1-carboxyethyl)or - nithine. An unusual amino acid produced by Streptococcus /act/s. J. Biol. Chem. 262,16109-16115.
[41] Thompson, J. and Miller, S.P.F. (1988) Nt-(1-carboxy- ethyl) iysine formation by Streptococcus lactis. Purifica.
111
tion, synthesis and stereochemical structure. J. Biol. Chem. 263, 2064-2069.
[42] Thompson, J. and Miller, S.P.F. (1990) NS-(1-carbo- xyethyl) ornithine and related N-carboxylalkyl amino acids structure: biosynthesis and function. Adv. Enzymol. (in press).
[43] Fields, J.H.A., Eng, A.K., Ramsden, W.D., Hochacld~ P.W. and Weinstein, B. (1980) Alanopine and strombine are novel imino acids produced by a dchydrogenase found in the adductor muscle of the oyster, Cra.v.vostrea gigas. Arch. Biochem. Biophys. 201,110--114.
[44] Eberlee, J.C., Storey, £M., Storey, K.B. (1983) Anaerobio- sis, recovery from anoxia, and the role of strombine and alanopine in the oyster, Crassostrea virginica. Can. J. ZooL 21, 2682-2687.
[45] Thompson, J. (1989) NS..(L-l-carboxyethyl)-c-orni thine:NADP + oxidoreductase from Streptococcus lactis. Purification and partial characterization, j. Biol. Chem. 264, 9592-9601.
[46] Thompson, J., Harr, ILL. and Donkersloot, J.A. (1990) NS-(L-l-carboxyethyl) oraithine:NADP + oxidoreductase in Streptococcus lactis: distribution, constitutivity, and regulation. Curt. Microbiol. 20, 239-249.
[47] Asano, Y., Yamaguchi, K. and Kondo, K. (1989) A new NAD+-dependent opine dehydrosenas¢ from Arthro- bacter sp. strain 1C. J. Bacteriol. 171, 4466-4471.
[48] Saier Jr., M.H. and Simoni, R.D. (1976) Regulation of carbohydrate uptake in gram-positive bacteria. $. Biol. Chem. 251, 893-894.
[49i &tier Jr., M.H., Feucht, B.U. and Hofsladter, LJ. (1976) Regulation of carbohydrate uptake and adenylate cycAase activity mediated by th~ enzymes I1 of the phosphoenol- pyruvate:sugar phosphate system in Escherichia coil J. Biol. Chem. 251, 88~-892.














![[Product Monograph Template - Standard] · DRUG INTERACTIONS.....25 DOSAGE AND ADMINISTRATION ... on lactate metabolism (see Endocrine and Metabolism, Lactic Acidosis section below)](https://img.pdfslide.us/doc/110x75/5ea304d1f714896fc84299f2/product-monograph-template-standard-drug-interactions25-dosage-and-administration.jpg)














