Embed Size (px)
Citation preview

ULTRAVIOLET LIGHT AND ITS EFFECT ON GERMINATION, GROWTH,
PHYSIOLOGY AND RESPONSES OF COOL SEASON GRASSES.
DISSERTATION
Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University
By
Edward John Nangle, M.S.
Graduate Program in Horticulture and Crop Science
The Ohio State University
2012
Dissertation Committee:
Dr. David Gardner, Adviser
Dr. T. Karl Danneberger
Dr. James D. Metzger
Dr. Luis E. Rodriguez-Saona

Copyrighted by
Edward John Nangle
2012

ii
ABSTRACT
The increase of ultraviolet (UV) light levels in the northern hemisphere raises the spectre
of possible problems for turfgrass plants due to long term exposure. Cool season
turfgrasses which are susceptible to photoinhibition may suffer from a loss of
productivity and growth, reducing their ability to sequester carbon. The effect on
germination, plant responses in relation to UV absorbing compounds and how turfgrasses
were evaluated for loss of quality with regards to linking new technology to visual ratings
have had no or limited research.
Enhancing the percentage of Kentucky bluegrass (Poa pratensis L.) seed
germination and speed could benefit establishment of the grass in a greater range of
environmental and geographical conditions. Potential for the use of ultraviolet light to
enhance Kentucky bluegrass seed germination exists through exposure to UV light. The
effect of ultraviolet light may be lost with seed age. In altering wavelength exposure,
there may be an opportunity to enhance the effect.
In measuring turf quality, traditionally human measurement has been the standard
method for both color and cover. Color, in particular, is thought to be controlled by
pigment changes. In evaluating a total of 51 cultivars of tall fescue (Schedenorus phoenix
Scop. Holub), perennial ryegrass (Lolium perenne L.) creeping bentgrass (Agrostis
stolonifera L.) cv. ‘Penncross’ and ‘L-93’ it was found that nitrogen content is

iii
most crucial in color measurements. Reflective measurements did not correlate with
nitrogen or chlorophyll content. Extract measurements had stronger correlation with
nitrogen content than pigmentation concentration. Current reflective measurement
equipment may not be closely linked to visual rating of turfgrass color possibly due to
variation in leaf surfaces.
There is a difference in response to UV light among grass species. Creeping
bentgrass ‘L-93’ produced increasing anthocyanin in response to UV light. The
characterization of Cyanidin – 3 – O – glucoside was the first reported in the literature in
creeping bentgrass. Carotenoids, zeaxanthin and β-carotene decrease in creeping
bentgrass after exposure to UV-B and turfgrass quality and vegetative production in
bentgrasses decreased to a greater extent than tall fescue or perennial ryegrass. All
grasses have the ability to initially accumulate phenolic compounds and flavonoids in the
tissue most exposed to light, although this still doesn’t mean a prevention in damage to
photosynthetic machinery. Turfgrass recovery and maintenance of optimal photosynthetic
rates will be crucial as breeders try to develop new cultivars that are adapted to higher
levels of UV light.

iv
DEDICATION
To Eddie and Catherine Nangle

v
ACKNOWLEDGEMENTS
I would like to thank Dr. Dave Gardner for giving me a chance at a PhD, it has been a
wonderful life changing experience here and I feel that I have achieved more than I could
ever have imagined. The friendship and comments, which were sharp, incredibly witty
and sometimes intelligent, were more than I could have asked for. I hope to carry on the
good name of the program wherever I may end up in a happy and successful career.
Thank you Dr. Metzger, for your invaluable advice and brevity. Dr. Danneberger, your
insight and always excellent stories kept me smiling and motivated. Thank you, Dr.
Rodriguez-Saona for your patience and willingness to teach and interact with me.
To the people in the lab, Dominic Petrella – I have learned as much from you as
you did from me; Aneta Studzinska, your determination to finish drove me on. To Pam
Sherratt, you took the first step and I am sure many more will faithfully follow your life
changing decision – I did! Dr. John Street, Emily, Matt, Dave Snodgrass, Jim Vent,
Andrew Muntz, Arly Drake and Phil Young, your help and input was greatly appreciated.
To Suemi, I couldn’t have gotten this far and further without you. To Mike O’Keeffe –
what you have done for me cannot be overestimated and I will be forever indebted.
Finally to my parents, it’s been tough and arduous, but this has been something
for which your support never wavered. For that, I will be eternally grateful and cannot
ever pay you back. I just hope you’re as proud as I am thankful.

vi
VITA
October 7, 1980 .............................................Born Enniscorthy, Wexford, Ireland
2008................................................................M.S. Horticulture and Crop Science,
The Ohio State University.
2008 to present ..............................................Graduate Research Associate, The Ohio
State University
PUBLICATIONS
Nangle, E.J., D.S. Gardner, J.D. Metzger, J. R. Street, and T.K. Danneberger. 2012. The effect of Nitrogen source and Trinexapac-ethyl on Creeping bentgrass (Agrostis stolonifera L.) physiology under neutral shade, deciduous tree shade and full sunlit conditions. Hortscience 47(7):1-7. Nangle, E., D. Gardner, A.S. Studzinska, T.K. Danneberger, and J.R. Street, 2010. Nitrogen absorption and movement in creeping bentgrass Agrostis stolonifera L. as affected by nitrogen source and shade. Proc. of 2nd Annual European Turfgrass Conference, Angers, France, 2010. Studzinska, A.K., D. S. Gardner, J. Yan, E. Nangle, and T. K. Danneberger. 2009. Development and Characterization of Transgenic Creeping Bentgrass Transformed with Arabidopsis BAS1 Gene. Int. Turf Soc. Res. J. 11:859-870.
FIELDS OF STUDY Major Field: Horticulture and Crop Science

vii
TABLE OF CONTENTS
ABSTRACT ........................................................................................................................ ii
DEDICATION ................................................................................................................... iv
ACKNOWLEDGEMENTS ................................................................................................ v
VITA .................................................................................................................................. vi
PUBLICATIONS ............................................................................................................... vi
FIELDS OF STUDY ......................................................................................................... vi
LIST OF TABLES ............................................................................................................. xi
LIST OF FIGURES ......................................................................................................... xiii
CHAPTER 1: LITERATURE REVIEW ............................................................................ 1
LIGHT .................................................................................................................... 1
ULTRAVIOLET LIGHT ........................................................................................ 2
CAROTENOIDS: LIGHT ABSORBING COMPOUNDS .................................... 6
FLAVONOIDS: UV ABSORBING COMPOUNDS WITH SPECIFIC
INTEREST IN ANTHOCYANINS ...................................................................... 11
PHENOLIC COMPOUNDS................................................................................. 18

viii
MEASUREMENT TECHNIQUES ...................................................................... 19
REFERENCE ........................................................................................................ 24
CHAPTER 2: SUBJECTIVE AND OBJECTIVE COLOR RATINGS OF
TURFGRASSES AND THEIR LINKS TO PIGMENTATION AND NITROGEN
CONCENTRATIONS. ..................................................................................................... 38
ABSTRACT .......................................................................................................... 38
INTRODUCTION ................................................................................................ 39
MATERIALS AND METHODS .......................................................................... 44
RESULTS ............................................................................................................. 49
DISCUSSION AND CONCLUSION .................................................................. 52
REFERENCE ........................................................................................................ 64
CHAPTER 3: INFLUENCE OF ULTRAVIOLET LIGHT ON GERMINATION RATE
AND SPEED OF KENTUCKY BLUEGRASS (Poa pratensis L.) ................................. 70
ABSTRACT .......................................................................................................... 70
INTRODUCTION ................................................................................................ 71
MATERIALS AND METHODS .......................................................................... 73
RESULTS AND DISCUSSION ........................................................................... 74
CONCLUSIONS .................................................................................................. 76
REFERENCE ........................................................................................................ 81

ix
CHAPTER 4: ULTRAVIOLET-B LIGHT INFLUENCE ON COOL SEASON
TURFGRASS GROWTH AND MORPHOLOGY .......................................................... 84
ABSTRACT .......................................................................................................... 84
INTRODUCTION ................................................................................................ 85
MATERIALS AND METHODS .......................................................................... 88
RESULTS ............................................................................................................. 91
DISCUSSION ....................................................................................................... 93
REFERENCE ...................................................................................................... 101
CHAPTER 5: PIGMENT CHANGES IN COOL SEASON TURFGRASSES IN
RESPONSE TO ULTRAVIOLET-B LIGHT IRRADIANCE ....................................... 106
ABSTRACT ........................................................................................................ 106
INTRODUCTION .............................................................................................. 107
MATERIALS AND METHODS ........................................................................ 113
RESULTS ........................................................................................................... 122
DISCUSSION ..................................................................................................... 125
CONCLUSION ................................................................................................... 129
REFERENCE ...................................................................................................... 141
CHAPTER 6: OVERALL CONCLUSIONS ON UV-B INFLUENCE ON TURFGRASS
PIGMENTS AND GROWTH ........................................................................................ 149

x
ABSTRACT ........................................................................................................ 149
DISCUSSION ..................................................................................................... 150
APPENDIX A: HANDHELD REFELCTANCE MEASUREMENTS OF TURFGRASS
TISSUE ........................................................................................................................... 154
APPENDIX B: COLOR OF LEAVES OF CREEPING BENTGRASS (Agrostis
stolonifera L.) TISUE AFTER EXPOSURE TO UV-B LIGHT .................................... 156
APPENDIX C: CHROMATOGRAM CAROTENOID STANDARDS ........................ 158
APPENDIX D: CHROMATOGRAM CHOKEBERRY ARONIA ................................ 160
APPENDIX E: CHLOROPHYLL A CONTENT AFTER D7 UV TREATMENT OF
CREEPING BENTGRASS ‘L-93’ ................................................................................. 162
APPENDIX F: CHROMATOGRAM BENTGRASS CAROTENOIDS ....................... 164
APPENDIX G: TALL FESCUE CAROTENOID CHROMATOGRAM ...................... 166
COMPLETE REFERENCE ........................................................................................... 168

xi
LIST OF TABLES
Table 2.1: Normality test results and data processing results for six cool season grasses
evaluated for color using transmission and reflection type measurements post
ROBUSTREG analysis. .................................................................................................... 56
Table 2.2: Correlation values for chlorophyll pigmentation and transmission
characteristics obtained using a Hunter XE colorquest colorimeter on extracts of six cool
season turfgrasses.............................................................................................................. 57
Table 2.3: Correlation values for chlorophyll pigmentation and tissue nitrogen levels with
visual ratings of six cool season turfgrasses. .................................................................... 58
Table 2.4: Correlation of visual ratings to transmission and reflection color characteristics
of six cool season turfgrasses. ........................................................................................... 59
Table 2.5: Correlation values for chlorophyll pigmentation and reflection characteristics
obtained using a Konica CR310 handheld colorimeter on surfaces of six cool season
turfgrasses. ........................................................................................................................ 60
Table 2.6: Correlations between reflectance measurements and transmission
measurements with tissue nitrogen for 6 cool season turfgrasses. .................................... 61

xii
Table 2.7: Human color rankings for cultivars of six cool grasses. (1= brown / 6 =
acceptable / 9 = optimum)................................................................................................. 62
Table 3.1: Germination percentages of Kentucky bluegrass (Poa pratensis L.) seed lots
treated with PAR (400 – 700 nm) and ultraviolet light (300 – 400 nm). .......................... 78
Table 4.1: Lighting conditions in control and ultraviolet treated light environments for
turfgrasses grown at The Ohio State University March – April 2011. ............................. 96
Table 5.1: Light intensity and light distribution for Conviron E15 growth chambers
illuminated using high output F12T12-CW.HO fluorescent tubes and QUV UV-B 313 nm
UV-B lights. .................................................................................................................... 131
Table 5.2: Anthocyanin pigmentation detected in creeping bentgrass ‘L-93’ after 7 days
of enhanced UV-B treatments. ........................................................................................ 132

xiii
LIST OF FIGURES
Figure 1.1: Carotenoid biosynthesis pathway to non-photochemical (NPQ) quenching
xantophyll cycle. ................................................................................................................. 8
Figure 1.2: Anthocyanin biosynthesis pathway. ............................................................... 14
Figure 2.1: Principal component analysis for turfgrass samples analyzed for color using
reflectance and transmission measurements. .................................................................... 63
Figure 3.1: Percentage germination of Kentucky bluegrass (Poa pratensis L.) seed treated
with photosynthetically active radiation (400 – 700 nm) and ultraviolet light (280 – 480
nm) in seed lot one. ........................................................................................................... 79
Figure 3.2: Percentage germination of Kentucky bluegrass (Poa pratensis L.) seed treated
with photosynthetically active radiation (400 – 700 nm) and ultraviolet light (280 – 480
nm) in seed lot two. ........................................................................................................... 80
Figure 4.1: Relative growth rate for four cool season grasses grown in control and
enhanced ultraviolet-B light conditions in repeated experiments. .................................... 97
Figure 4.2: Color ratings for four cool season grasses grown in control and enhanced
ultraviolet-B light conditions in repeated experiments. (1-9 scale, 1 = brown/dead, 9 =
dark green optimal color, 6 = acceptable). ........................................................................ 98

xiv
Figure 4.3: Number of tillers per plant for four cool season grasses grown in control and
enhanced ultraviolet-B light conditions in experiment one. ............................................. 99
Figure 4.4: Number of tillers per plant for four cool season grasses grown in control and
enhanced ultraviolet-B light conditions in experiment two. ........................................... 100
Figure 5.1: Fv / Fm values for Perennial Ryegrass and Creeping Bentgrass ‘Penncross’
treated with UV-B light (280 – 315 nm) and PAR light (400 – 700 nm) conditions over
seven days. ...................................................................................................................... 133
Figure 5.2: Total chlorophyll content for Tall Fescue and Creeping Bentgrass ‘L-93’
treated with UV-B light (280 – 315 nm) and PAR light (400 – 700 nm) conditions over
seven days. ...................................................................................................................... 134
Figure 5.3: Distribution of phenolics in creeping bentgrass ‘L-93’ after treatment with
both UV-B light (280 – 315 nm) and PAR light (400 – 700 nm). Data was pooled as all
grasses had the same response. ....................................................................................... 135
Figure 5.4: Distribution of total flavonoids in perennial ryegrass tissue after treatment
with both UV-B light (280 – 315 nm) and PAR light (400 – 700 nm). Data was pooled as
all grasses had the same response. .................................................................................. 136
Figure 5.5: Turfgrass flavonoid content in Creeping bentgrass, Tall Fescue and Perennial
Ryegrass grown in UV-B light (280 – 315 nm) and PAR light (400 – 700 nm) D0 to D7.
......................................................................................................................................... 137
Figure 5.6: Anthocyanin profile of creeping bentgrass ‘L-93’ exposed for 7 days to UV-B
light. Produced from HPLC analysis using C18 column and light absorbance measured at
520 nm using a PDA. ...................................................................................................... 138

xv
Figure 5.7: Chromatogram of Perennial ryegrass carotenoids with absorbance at 453 nm.
Data produce using C30 column and PDA connected to Chemstation software. ............ 139
Figure 5.8. β-carotene (top) and zeaxanthin (bottom) content in creeping bentgrass, tall
fescue and perennial ryegrass exposed to UV-B light D0 – D7. .................................... 140

1
CHAPTER 1
LITERATURE REVIEW
LIGHT
Plant growth and development depends mainly on energy derived from light
(Smith, 1982). The habitat and ecosystem of plants can vary greatly even when there are
minor changes in light (Schmitt and Wulff, 1993). There are two properties of light that
relate to plant growth and development: light quality and quantity. Light quality can refer
to proportions of visible light spread across the 400-700 nm spectra which influences
plant growth responses, e.g. changes in the ratio of red light to far red light can influence
biomass allocation (Smith, 1994; Maliakal et al., 1999) and light quantity is the actual
amount of energy associated with radiation hitting the earth’s surface (Smith, 1994).
There has been an increase in research concerning the effects of ultraviolet light (UV)
100-400 nm on plant growth and development (Caldwell et al., 1989; Mendez et al.,
1999; Rowland 2006; Sarkar et al., 2011). This includes UV-C (100-280nm), UV-B (315-
280nm) and UV-A (400-315nm). The ozone (O3) layer in the stratosphere between 15
km and 60 km above the earth filters UV wavelengths to hit the earth’s surface
selectively (Rind et al., 1990; Rowland, 2006). Ozone can absorb ultraviolet-B light,
while oxygen (O2) absorbs UV-C light (Caldwell, 1979; Björn et al., 1998). Exposure
UV-B is increasing due to the decline of ozone linked to chlorofluorocarbons and the
breakage of the O3 structure by chlorine (Rowland, 2006).

2
The levels of atmospheric ozone, however, have varied and declined since 1950 with
20% decreases in content in some locations (Björn et al., 1998; Frederick et al., 1989;
Kerr and McElroy, 1993; Rowland, 2006). Ozone is also created by UV striking O2 and
splitting it in two followed by the attachment to an unbroken O2 (Rozema et al., 1997).
The incidence of UV radiation varies by season with winter time increases as high 35%
per year while there is a 7% increase in summer due to greater ozone loss in colder
temperatures catalyzing chlorine depletion of the gas (Kerr and McElroy, 1993; Solomon,
1990). Cloud cover decreases UV radiation by as much as 20% compared to a clear day
due mainly to scattering (Lubin and Frederick, 1991).
ULTRAVIOLET LIGHT
The discovery of ultraviolet light as a wavelength occurred in 1800 with Ritter
(Berg, 2008). The issue of UV radiation hitting the earth’s atmosphere was first noted in
1881 by Hartley when he was able to measure ultraviolet energy hitting the earth’s
surface and found it varied depending on altitude (Hartley, 1881). The description of the
wavelengths occurred not long after with Hertz who developed a method for measuring
microwaves.
The difference in light energy associated with each photon in each UV light
portion has led to developments of action spectrums (Caldwell, 1971; Flint and Caldwell,
2003). The energy associated with each portion of the UV range elicits different
responses, creating the need for the spectrum. Currently, regions on earth between 40°N
and 40°S latitude receive 2-11 kJ m-2 d-1 of UV radiation and this has the potential to
increase due to ozone loss (Taalas et al., 2000; Kakani et al, 2003; Rowland, 2006).

3
Exposure to UV-B levels along different lines of latitudes vary due to ozone depletion,
notably at the poles. Increases in UV levels since 1970 are as great as 130% in the
Antarctic in the spring as compared to a 7% increase in northern hemisphere mid-
latitudes over the same time period (Madronich et al., 1998). Levels of ozone depleted by
40% in the Antarctic while negligible increases occurred in the mid-latitudes since the
1970s (McKenzie et al., 2003).
Ultraviolet-C and B, impacts DNA through the production of cyclobutane
pyrimidine dimers leading to mutations (McLennan, 1987) while UV-B has a greater
impact on plant physiology (Stapleton, 1992) and UV-A is considered to have little
impact (Flint and Caldwell, 2003). Exposure to UV-B also tends to promote dimerization
while UV-A exposure tends to promote repair enzymes such as photolyase which
photoreactivate DNA strands by dimer breakage (Strid et al., 1994; Britt, 1996). The
increased amount of UV-B is considered most important as it has physiological impact
(Caldwell, 1979; Stapleton, 1992). Ultraviolet-B levels are projected to increase by 50-
60% from current levels in the northern hemisphere with peak irradiance levels predicted
to occur between 2010 and 2019 in the spring time (Taalas et al., 2000). There is
variation in the possible increases with seasonal and geographical variations. Alpine
regions already have experienced an increase of UV radiation by 1.1% per anum
(Blumenthaler and Ambach, 1990) with higher elevations having higher levels of UV-B
due to a thinner atmosphere (Caldwell et al., 1980). UV light initially damages the
photosynthetic apparatus within the plant. The majority of research indicates that damage

4
is mainly in photosystem II of the light harvesting complex (Tevini et al., 1988;
Bornman, 1989).
The impact of ultraviolet-A light on plants had been noted in various ways
including phototrophic responses in the coleoptile tip in wheat (Triticum spp.) (Castle,
1935), and control of phytochrome via UV-A light - near blue was proposed by Galston
(1949). Pfeiffer (1928) reported that without UV irradiance, plants had longer shoots,
thinner leaves and poor root development. Popp and Brown found that there were only
negative attributes associated with UV light, (1936). They also disputed the results of
Pfeiffer based on lack of replication and poor lighting control. Other studies have
demonstrated a decrease in pollen production (Flint and Caldwell, 1984), declination in
competitive ability of plants against weeds (Barnes et al., 1990), and reductions in
biomass in a range of plants including pines (Pinaceae L.), (Tevini et al., 1981; Sullivan
and Teramura, 1988). Thinning of the leaf surface and increased levels of flavonoids and
phenolics are also found in plants (Tevini et al., 1981; Sarkar et al., 2011). The potential
for long term reduction in the ozone layer has driven interest in how long term adaptation
of plants will occur in response to increasing UV levels.
There are many causes for the variation of the impact of UV light on plants
observed in the literature. Plants with sensitivity to UV-B light produce increased
epicuticular wax on the adaxial surface while there is a reduction in leaf thickness in
cotton (Gossypium hirstum L.). Decreases in mesophyll and palisade tissue also occurs
(Kakani et al., 2003). Stomatal densities may also decrease but this is crop dependent and
other factors play a role. In saturated CO2 conditions, treatments of UV-B were more

5
effective in decreasing stomatal density (Ziska et al., 1992). Altitude changes increase the
UV effect on growth habits of plant material. Plant tissue found above 3000 m has
increased dark respiration, an earlier effort at reproduction and maintained photosynthetic
efficiency compared to plant material from 0 m above sea level when treated with
supplemental UV light due to adaptation to the conditions (Ziska et al., 1992). There can
be a greater than 40% increase in UV irradiance between the elevations (Caldwell et al.,
1980; Sullivan et al., 1992). Variations in levels of ozone with elevation change are
considered possible due to regional aerosol conditions solar elevation and also albedo
(Pfeifer et al., 2006).
Dilation of thylakoid membranes in (Pisium sativum L.) and damage to
chloroplast membranes occurred after UV-B irradiation (Brandle et al., 1977).
Herbaceous dicots cannot attenuate UV-B in the epidermis with 18-41% of UV-B light
reaching the mesophyll layer leaving increased chances for damage to photosynthetic
apparatus. Conifer needles, regardless of age, absorb essentially all UV-B light. Grasses
and woody dicots are considered intermediate in their ability to protect the mesophyll
layer from damage with attenuation in the epidermis ranging from 3-12% (Day et al.,
1992). In contrast to the negative effects, tomato ripening can be faster with treatments of
UV-B as colored pigments build up in the plant which can absorb the excess energy,
(Bacci et al., 1999). Flowering color can be enhanced with UV-B treatments in smoke
tree (Cotinus coggygria ‘Royal purple’). The increase in anthocyanin content in the leaf
in response to the light treatment resulted in more favorable performance ratings in color
(Oren-Shamir and Nissim, 1997).

6
CAROTENOIDS: LIGHT ABSORBING COMPOUNDS
The potential long term effects are considered to be both anatomical and physiological
and plants do possess response mechanisms for dealing with UV-B light. These are
mainly antioxidants such as photolyase and super oxide dismutase (Zhang et al., 2005;
Sarkar et al., 2011). These systems repair damage caused by excited radicals due to
excess energy from UV-B radiation. There are compounds however, that may absorb the
light directly and aid the reduction of UV-B light attenuation and energy dissipation in
the plant such as flavonoids and carotenoids (Gould, 2004; Rowland, 2006).
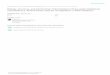
Carotenoids are lipid soluble isoprenoid compounds containing C40 chains
(tetraprenoids) which are ubiquitous in nature and have been investigated in relation to
human health benefits. Interest exists in looking at the capabilities they can have in plant
protection (Bartley and Scolnik, 1995; Müller et al., 2001). Biosynthesis of carotenoids
occurs in cellular plastids where they are associated with light harvesting complexes
(Kopsell and Kopsell, 2006). The basic C5 isoprenoid structures are converted to
isopentyl diphosphate (IPP) and into dimethylallyl diphosphate (DMAPP). These
compounds are substrates for the enzyme geranylgeranyl diphosphate synthase (GGPP)
(Cunningham and Gantt, 1998). The enzyme catalyzes GGPP formation which is paired
with another GGPP in a head to head condensation reaction creating phytoene (Gross,
1991). The pathway then branches after the conversion of phytoene to lycopene via
phytoene desaturase and ξ carotene desaturase; the following cyclization of the lycopene
rings can form carotenoids with two β rings (Fig. 1.1), (zeaxanthin, antheraxanthin,

7
violaxnathin) or one β ring and one ε ring (lutein, α carotene) on the C40 structures (Fig.
1.1), (Kopsell and Kopsell, 2006).
The reversibility of accumulation of lycopene with increased FR light sources
following red light exposure indicates that initial responses in carotenoid synthesis are
due in part to phytochrome (Alba et al., 2000). This hypothesis was first thought of early
in the 1950’s (Piringer and Heinze, 1954) when accumulations of certain pigments were
noted in response to the red light wavelengths. The inability to clarify this result further
inhibited work past the 1970’s. The general thought was that this physiological trait
occurred which was recently confirmed (Alba et al., 2000).

8
Geranylgeranyl diphosphate
Phytoene
ζ-carotene
Lycopene
β – carotene α - carotene
zeaxanthin lutein
NPQ antheraxanthin loroxanthin
violaxanthin
Figure 1.1: Carotenoid biosynthesis pathway to non-photochemical (NPQ) quenching xantophyll cycle.

9
The variations in the carotenoids impact various functions in the plant. Lutein is
believed to have a greater impact on light quenching than β carotene (Cuttriss and
Pogson, 2004). The xantophyll cycle has received a lot of attention due to its antioxidant
capacity and photoprotection element (Bartley and Scolnik, 1995; McElroy et al., 2006).
Carotenoids are essential in reducing light photooxidative stress to chlorophyll and the
light harvesting complex (Foyer et al., 1994, Moradas-Fereira et al., 1996). The
downward energy transfer of singlet oxygen (1O2) to zeaxanthin reduces the damage it
can cause to lipids and cell walls (Owens et al., 1992). In carotenoids with no rings at the
end of the structure, such as lycopene, the large amount of double bonds allows for a
similar effect (Kopsell and Kopsell, 2006). Light harvesting complexes have the -trans
configuration providing efficient singlet energy transfer to chlorophyll molecules. The -
cis forms tend to isomerize towards the -trans configuration upon photo excitation. The
carotenoids attached to reaction centers are also found to be in –cis conformation and are
unaffected by temperature change. This may make them preferred carotenoids for energy
dissipation (Lutz et al., 1978; Koyama and Fujii, 1999).
Creeping bentgrass xantophylls in high light conditions cumulatively increase as a
percentage of the total carotenoids in high irradiance but decrease in low irradiance.
Neoxanthin and β-carotene decrease in high irradiance but increase in low irradiance
(Bell and Danneberger, 1999; McElroy et al., 2006). Testing of creeping bentgrass
showed that after up to 24 hours of exposure there was an increase of carotenoids in
higher irradiance but a resultant decrease and probable degradation after 168hrs
(McElroy et al., 2006). Maximum buildup of lutein, β-carotene and chlorophyll B

10
occurred at 24 hours in Kale (Brassica oleracea L.) but an overall increase in
photoperiod also increased total pigment accumulations (Lesfrud et al., 2006). Seeds of
certain maize hybrids had lower carotenoid content grown at cooler temperatures (16 °C)
(Massacci et al., 1995).
The application of nutrient elements to plants increases carotenoid content.
Increasing nitrogen applications to spinach (Spinacia oleracea L.) almost doubled the
quantity of lutein on a fresh weight basis (Lesfrud et al. 2007). This is a benefit that may
already have been realized with current applications of nitrogen but also may allow for
improvements in areas which are prone to excess levels of sunlight. The addition of
nitrogen to plants increases production of many pigments (Greenwood et al., 1990) until
a level of nitrogen toxicity is reached (Carrow et al., 2001). In periods of reduced
nitrogen content rice plants may increase their content in an attempt to increase thermal
dissipation abilities in responses following decreases in chlorophyll (Huang et al., 2004).
The management of turfgrasses on sand based golf greens is dependent on sufficient
fertility in a growing medium that is classed as having poor nutrient retention capabilities.
Thus poor fertility management may lead to photoinhibition and loss of productivity.
Control or inhibition of carotenoid biosynthesis is a mode of action that has been
exploited in the recent development of herbicides. Mesotrione (2-[4-(methylsulfonyl)-2-
nitrobenzoyl]-1,3-cyclohexanedione) has proved useful in controlling common turfgrass
weeds (McCurdy et al., 2008; Beam et. al., 2006). Inhibition of certain carotenoids leads
to bleaching of the leaf and damage to the photosystems of weeds (McCurdy et al., 2008).
The damage to the photosystems then allows for improved turfgrass competition although

11
there is also potential for damage to turfgrass systems in certain situations (Beam et al.,
2006). This form of chemical control is relatively new to the turfgrass industry and
suggests the potential for more useful information related to the development and control
of secondary metabolites.
There have been efforts to enhance levels of carotenoids in plants. This has been
carried out for a variety of reasons including ecological fitness due to reduced
photoinhibition and increased human health benefits. Variations occur in tomato in
particular providing a wide range of varieties with different carotenoid profiles (Fraser
and Bramley, 2004; Ronen et al., 1999; Ronen et al., 2000). Genetic engineering of
carotenoids in plants has occurred at an increased rate as our understanding has
developed about the pathways. Control is offered with genetic manipulation and allows
for a greater understanding and possibility for greater change in compounds (Fraser and
Bramley, 2004). Research on the effects of UV-B impact on carotenoid profiles of
turfgrass species has not been carried out.
FLAVONOIDS: UV ABSORBING COMPOUNDS WITH SPECIFIC INTEREST IN
ANTHOCYANINS
Pigments such as flavonoids have specific absorption characteristics in the UV
region (Ziska et al., 1992). Flavonoids are compounds defined as having a 15 carbon
structure in a form of a C6 – C3 –C6 backbone (Anonymous, 1997). They can be broken
down into different pigments which have UV-B absorbing characteristics such as
anthocyanins (Gould, 2004). Anthocyanins are a pigment in plants which are associated
with various functions. They are derived from shikimic acid via flavonoid formation

12
(Bennett and Wallsgrove, 2006). There have been at least 4,000 flavonoids described and
many are found in higher plants (Seigler, 1998). Plants use them as a color source for
attraction of pollinators which can see in the UV wavelengths and also as a defense
mechanism for disease and herbivory (Briscoe and Chittka, 2001; Irani and Grotewold,
2005). They are primarily known for their bright red colors and are used in the food
industry as a colorant. In plants, anthocyanin response is related to environmental stresses
including drought stress, excessive UV-B conditions, heavy metal toxicity, cold
hardiness, insect and fungal attack (Chalker-Scott, 1999; Gould, 2004).
Anthocyanins are naturally found in higher levels in plants such as berries and red
fruits (Lohachoompol et. al, 2004). Anthocyanins are found in shoots of plants but in
general are predominantly found in the upper epidermis or just beneath it in the
mesophyll layer in leaves (Chalker-Scott, 1999). The areas in which they are most often
found tend to have greater levels of light exposure (McLure 1975, Chalker-Scott 1999). It
has been shown that the synthesis is autonomous to cells (Lancaster et al., 1994). The
localization of production would indicate that a specific screen for cell ultra-structure
may occur versus whole plant changes.
Anthocyanin synthesis involves approximately seven steps and the pigments are
assembled similarly to other flavonoids from two different streams (Bennett and
Wallsgrove, 2006). One stream involves the shikimate pathway to produce the amino
acid phenylalanine. The other stream produces malonyl-CoA, a C3 unit from a C2 unit
(acetyl-CoA). These streams meet and are coupled together by the enzyme chalcone
synthase (CHS), which forms an intermediate chalcone (Fig. 1.2). The chalcone synthase

13
is considered to be the first enzyme in all direct flavonoid biosynthesis. The chalcone is
subsequently isomerized by the enzyme chalcone isomerase (CHI) to the prototype
pigment naringenin (Fig 1.2), (Heller and Forkmann, 1988). Naringenin is oxidized and
then further reduced by the enzyme dihydroflavonol 4-reductase (DFR) to the
corresponding leucoanthocyanidins. The resulting, unstable anthocyanidins are further
coupled to sugar molecules by enzymes like UDP-3-O-glucosyl transferase to yield the
final relatively stable anthocyanins (Seigler, 1998). One of the final steps is light
controlled and in particular it seems that UV-B light induces increased synthesis
(Kubasek et al., 1992).
The initiation of synthesis is thought to be controlled by the light receptors
phytochrome or cryptochrome (Mancinelli et al., 1991; Sponga et al 1986).
Cryptochrome, a blue light receptor, is inferred to elicit anthocyanin synthesis in tomato
seedlings. Blue light applications significantly increased levels of anthocyanins in
tomato. The phytochrome response was more significant in cabbage, however, with about
7x efficiency in anthocyanin production with red light (650 – 800nm) applications
(Sponga et al., 1986). The initiation response of chalcone synthase to UV-A is mediated
by cryptochrome 1 (CRY1) but there is no response to UV-B light via either CRY1 or
CRY2 to enhance chalcone synthase, the key flavonoid biosynthesis enzyme (Wade et
al., 2001). The interaction between the two light receptors is also possibly important as
both phytochrome and cryptochrome converge in a single pool to initiate light responses
(Ang et al., 1998). It has been proposed that PHYB, in particular, equilibrates the flux
through CRY1 and UV-B signaling pathways (Wade et al., 2001).

14
Phenylalanine
4 – Coumaryl CoA
Malonyl CoA Chalcone synthase
Chalcone
Chalcone isomerase
Flavanone
Flavones, isoflavones Flavanone 3-hyrdoxylase (F3H) Flavan-4-ols (colorless)
Dihydroflavanol
Anthocyanins Flavanol glycosides (red pigmentation) (colorless)
Figure 1.2: Anthocyanin biosynthesis pathway.

15
Plant age affects response to changes in the light and induction of chalcone
synthase activity via phytochrome. In 6 days old mustard (Sinapis hirta L.) seedlings far
red, UV-B and UV-A / blue light induced the CHS transcripts (Kaiser el al., 1995). In
older mustard seedlings and mature Arabidopsis leaf tissue, however, there is no
phytochrome induction but there was a response due to UV-B and UV-A / blue light
treatments (Jackson et al., 1995).
There are 6 main anthocyanins and many others that occur less frequently. The six
aglycones (pelargonidin, cyanidin, delphinidin, petunidin, malvidin and peonidin) are
widely distributed among angiosperms and gymnosperms (Seigler, 1998). The
differences between the various anthocyanins are mainly due to the nature and number of
sugars attached to the ‘3 or ‘5 position on the C ring of the structure. The addition of
sugars and acids at either of these positions leads to a more stable structure and alters the
absorption abilities of the pigments (Garćia-Viguera and Bridle, 1999). Variations can be
caused by the nature and number of aliphatic or aromatic acids attached to the sugars
(Kong et al., 2003). The variations lead to color changes which can be seen across a
whole range of plants. The leaves of red cabbage contain eight acylated cyanidin
glucosides while red onion contains four major anthocyanins; cyanidin 3-glucoside,
cyanidin 3-laminaribioside, cyanidin 3-malonylglucoside, cyanidin 3-
malonyllaminaribioside and four minor ones (Brouillard, 1988; Donner et al., 1997).
There is some argument as to whether anthocyanins are directly related to general
stress tolerance or if they are produced as a direct result of the stress under which the
plant has been placed. The impact of cold hardiness, changes in light intensity and

16
wavelengths seems to enhance responses in particular. Photoprotection has been offered
as a specific anthocyanin function and hazelnut seeds lacking anthocyanins have died in
field conditions versus anthocyanin producing progeny (Mehlenbacher and Thompson,
1991). The anthocyanins also favor absorption of UV-B and green light over blue light
with only small quantities of red light being absorbed (McClure, 1975). The absorption of
the blue-green light means that there is only red light available to chloroplasts. This is
important as it reduces the potential for excessive energy build up (Pietrini and Massacci,
1998). An increased tolerance for photoinhibition in cold acclimated jack pine seed /
needles was a result of light attenuation by anthocyanins and increased utilization of light
energy (Krol et al., 1995).
Temperature changes elicit many anthocyanin responses. In fall, color changes in
leaves lead to increased quantities of carotenoids and anthocyanins giving a range of red,
orange and yellow colors. The increase in content has been related to masking of
chlorophylls as leaves are senescing thus decreasing potential for photo oxidation damage
reducing nutrient retrieval prior to winter-like conditions (Field et. al., 2001). The red
leaves are seen when there is de novo synthesis of anthocyanins unlike carotenoids which
are revealed with chlorophyll breakdown and there is no increase in levels (Matile et al.,
1992). Increased absorption in green yellow light by anthocyanins (Neill and Gould,
1999) may counteract the loss of chlorophyll and attenuate sufficient light to prevent
damage as leaves senesce (Field et al., 2001). The reflection of red light may be
independent of anthocyanin content and concentration of the pigments is more important
than their location (Neill and Gould, 1999).

17
Temperature also plays a role in the stability of anthocyanins, and they are
susceptible to degradation if exposed to prolonged warmer temperatures when used in
food colorants (Giusti, personal comm. 2008, Ahmed et al., 2004). In the natural
environment the anthocyanins are seen in varying temperatures depending on the plant
and the functionality of the anthocyanin. The alterations in temperature are seen as
leading to a response also. In cabbage (Brassica oleracea L.) colder temperatures
increase light dependent anthocyanin production; however, there is disagreement as to
whether the development is a result of temperature effects on the phytochrome system or
on chalcone synthase activity related to photoinhibition (Rabino and Mancinelli, 1986).
The pH plays a major role in coloration. Lower pH levels (<2) tend to produce a
red color while a pH greater than 4 results in predominantly colorless anthocyanins
(Seigler, 1998). The change in plant pH can be caused by a variety of environmental
effects including sunlight (Zu-Hua et al., 1990). The pH also affects anthocyanin bases
which are stabilized by forming very colorful complexes with metals. The complexes
with such minerals as aluminum, iron or magnesium which are extremely susceptible to
pH change and have narrow limitations (Seigler, 1998). The extraction and quantification
of anthocyanins tends to occur from plant material using a solvent containing a low
percentage of hydrochloric or formic acid. This allows samples to be kept at lower pH
and reducing the chances of degradation of non-acylated pigments (Kong et al., 2003).
Anthocyanins may also have potential to reduce aluminum toxicity. Formation of
complexes between anthocyanins and Al3+ allows for pigment stabilization and reduces

18
degradation due to oxygen reactions as well as internal Al3+ levels (Moncada et al.,
2003).
PHENOLIC COMPOUNDS The class of phenolic compounds is widespread and has been identified in large
numbers. Approximately 40% of the organic carbon is allocated to phenolic compounds
in the biosphere (Croteau et al., 2000). Their production is predominantly linked to the
shikimate and acetate pathways (Floss, 1997; Hare and Cress, 1997). They are largely
derived from phenylalanine and are formed usually in cells close to locations of stress.
The enzyme phenylalanine ammonia-lyase (PAL) is used to direct carbons from amino
acids into the synthesis of the metabolites. They can be constitutively present (e.g.
lignins) or they can be induced (Langebartels et al., 2002). Their production is highly
complex and variable as there are so many compounds with multi step reactions required
for their synthesis (Mustafa and Verpoorte, 2007). The structures have a minimum of 6
carbons and their structure has an aromatic phenyl ring. There are generally one or more
acidic hydroxyl groups attached to the aromatic ring (Croteau et al., 2000).
The action of UV within the photosynthetic apparatus is predominantly involved
in the formation of highly excited compounds such as singlet oxygen (Foyer et al., 1994).
A majority of phenolic compounds act as scavengers of singlet oxygen and have an
ability to donate hydrogen from hydroxyl groups around their aromatic ring to free
radicals. This then reduces the impact of oxidation of lipids and other structures within
the cell (Foti et al., 1994). This can be used to reduce protein denaturation in the
photosynthetic apparatus which undergoes continuous cycles of damage and repair and

19
can become overwhelmed in high light or enhanced UV-B conditions (Takashi and
Murata, 2008). The production of these compounds is stimulated by UV radiation as well
as flavonols (Tevini, 1993). The range of compounds means that they can be induced in
many situations. High concentrations of phenolic compounds are believed to reduce
herbivore attack (Dicke et al., 2003). In Brassicaceae, sinapate esters of phenolics
provide protection against UV irradiation due to specificity in UV light wavelength
absorption (Sheahan, 1996). Phenolics in epidermal cells have the ability to screen and
absorb UV irradiation (Bornman and Teramura, 1993).
Plants do also possess the ability to distribute protective compounds based on
where light exposure is highest. In Jersey cudweed (Gnaphalium luteo-album L.), surface
concentrations of flavonols increased in response to UV-B radiation (Cuadra et al., 1997).
Regular removal of tissue may reduce a plant’s ability to respond to these conditions and
enhance the impact of UV-B light stress. Phenolic compounds found in the epidermal
layers of leaves are believed to offer screening of elevated light levels to the
photosynthetic apparatus (Gould, 2004). The upper part of the plant may alter its
pigment composition in response to the light location also (Ju et al., 1999).
MEASUREMENT TECHNIQUES
The ability to separate and identify the compounds has increased as equipment
sensitivity and improved structural elucidation developed. This happens via use of
spectrophotometry, mass spectrometry, high performance liquid chromatography and
nuclear magnetic resonance spectrometry has created better separation techniques which
do not damage the compounds during analysis. This has allowed for the creation of a vast

20
amount of information on these materials and in food sources there is a lot of information
known about their structure and molecular weight (Wrolstad and Giusti, 2001). A range
of these systems are used to characterize and quantify pigmentation changes.
Spectrophotometry is dependent on variations in reflective or transmission properties in
compounds based on their reaction to excitation via light energy. The absorption of light
leads to a change in energy in bonding electrons (electronic, vibrational, and rotational).
The spectra arise when the molecules of interest absorb photons of a specific energy.
They remit the energy at a slightly longer wavelength which is then divided out by a
monochromator and absorbed by a light sensor. The values are then compared against a
set of standards (Hardy, 1938). Spectrophotometers can be used in a pH differential
method to quantify specific anthocyanin levels depending on the standards available.
There is a very high correlation with the standards (R >0.9), (Lee et al., 2005). Analysis
of all pigments in this study could be carried out with spectrophotometry (Wellburn,
1994).
More in-depth analysis can be carried out using high pressure liquid
chromatography (HPLC). The process is the separation of chemical substances by
partitioning them between two media. The most common form is via a stationary phase
and a mobile phase. The first may be a solid or liquid while the second maybe a liquid or
gas. The separation of substances depends on a range of situations – compound size,
polarity and volatility are some of the common methods. It provides a precise method
which can separate compounds which are very similar thus difficult to analyze using
other methods. Chromatography itself has many fields and methods such as gas

21
chromatography, thin layer chromatography, paper chromatography, and gel permeation
chromatography (Shackleton, 2010). Specific known wavelengths are again used with a
photodiode array along with retention time, order of elution, standards, solvents used in
phases and also the extraction methods to characterize and quantify the pigments.
Anthocyanins and carotenoids can be separated within the chemical classes using this
system and it has become consistent and reliable over time (Hong and Wrolstad, 1990;
McCurdy et al., 2008).
Enhancing the separation and developing greater precision requires mass analysis
of compounds. This can be carried out using mass spectrometry. It analyzes compounds
based on a mass to charge ratio of an analyte. This mass to charge is used in two ways;
the charge on the compound and its mass affect its flight through a charged field and
depending on this its structure can be determined using time of flight and distance flown.
The compounds are ionized and have a charge added and then analyzed. This
occurs simply through a five step process. The loading of samples is carried out usually
with an automatic injector or it can be done manually if single samples are being used.
The samples are then vaporized undergoing a phase transition from liquid to gas phase.
Ionization then occurs via a range of ways – including electrospray and lasers, and this
leaves a charged particle which is then accelerated through a larger column which is
surrounded by electromagnetic fields. The ions are then deflected by a magnetic field
according to their masses. This method has been used successfully to elucidate
anthocyanins (McEwen et al., 2005; Pascual-Teresa et al., 2002). These systems allow for
further information to be produced on turfgrass changes in response to both biotic and

22
abiotic stresses. These methods will allow for deeper insight into producing quality
turfgrasses with more specific topographical, geological and geographical variances
catered for. It will give turfgrass breeders new tools in selecting potential turfgrass
cultivars.
The changes that are currently ongoing in the earth’s atmosphere and potential
future issues with higher light intensities and alterations in spectral radiation mean that
understanding the protective pigment capacities of turfgrass in response to this stress is
important. In turfgrass there has been very little work on these pigments. There is a
known increase in total anthocyanin concentration in Kentucky bluegrass (Poa pratensis
L.) in response to UV-B light (Zhang et al., 2005), but there has been no characterization
of the pigments. Identification of changes within turfgrass plants based on possible
attainable levels of UV-B is important and could aid potential breeding efforts in trying to
combat the problem. Their influence on turfgrass color and productivity may not be
understood in this situation and this warrants further investigation. The physiological
aspect of the plant and the response location is also of importance. It can be expected that
tissue receiving more sunlight at the top of the canopy may have a dual purpose. It
creates shaded conditions for the understory, reducing potential for light damage while
also it could be expected that there is a concentration of protective light pigmentation
where the light intensity is greatest. This also may influence carbohydrate allocation and
force a change in plant growth habit. A reduction, for instance, in biomass accumulation
or tillering would lead to a reduction in recovery capability and thus limiting turfgrass use
in certain situations. These questions do pose a learning moment for physiologists and

23
breeders and the objective of this study is to give insight into what protective
pigmentation cool season turfgrasses possess and how plants may or may not use them in
relation to a stress response.

24
REFERENCE Ahmed, J., U.S. Shivhare, and G.S.V. Raghavan. 2004. Thermal degradation kinetics of
anthocyanin and visual colour of plum puree. Euro. Food Res. Tech. 218:525-528. Alba, R., M.M. Cordonnier-Pratt, and L.H. Pratt. 2000. Fruit-localized phytochromes
regulate lycopene accumulation independently of ethylene production in tomato. Plant Physiol. 123:363-370.
Ang, L.-H., S. Chattopadhyay, N. Wei, T. Oyama, K. Okada, A. Batschauer, and X.-W
Deng. 1998. Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol. Cell. 1:213–222.
Anonymous, 1997. Flavonoids (isoflavonoids and neoflavonoids). In McNaught A.D. and
A.Wilkinson (ed.) IUPAC Compendium of Chemical Terminology, 2nd ed. Available at http://old.iupac.org/goldbook/F02424.pdf (verified 28 Jun. 2012).
Bacci, L., D. Grifoni, F. Sabatini, and G. Zipoli. 1999. UV-B radiation causes early
ripening and reduction in size of fruits in two lines of tomato (Lycopersicon esculentum Mill.). Glob. Change Bio. 5:635-646.
Barnes, P.W., S.D. Flint, and M.M. Caldwell. 1990. Morphological responses of crop and weed species of different growth forms to ultraviolet-B radiation. Am. J. Bot. 77:1354-1360.
Bartley, G.E., and P.A. Scolnik. 1995. Plant carotenoids: Pigments for photoprotection, visual attraction and human health. Plant Cell. 7:1027-1038.
Beam, J.B., W.L. Barker, and S.D. Askew. 2006. Selective creeping bentgrass (Agrostis
stolonifera L.) control in cool season turfgrass. Weed Tech. 20:3430-344.

25
Bell, G.E., and T.K. Danneberger. 1999. Temporal shade on creeping bentgrass turf. Crop Sci. 39:1142-1146.
Bennett, R.N., and R.M. Wallsgrove. 2006. Secondary metabolites in plant defense
mechanisms. New Phytol. 127:617-633. Berg, H. 2008. Johann Wilhelm Ritter – The founder of scientific electrochemistry. Rev.
Polarography. 54:99-103.
Björn, L.O., T.V. Callaghan, C. Gehrke, U. Johanson, M. Sonesson, and D. Gwynn-
Jones. 1998. The problem of ozone depletion in northern Europe. Ambio 27:275-279.
Blumenthaler, M., and W. Ambach. 1990. Indication of increasing solar ultraviolet-B radiation flux in Alpine regions. Science 248:206-208.
Bornman, J.F. 1989. Target sites of UV-B radiation in photosynthesis of higher plants. J.
Photochem. Photobiol. 4:145-158.
Bornman, J.F., and A.H. Teramura. 1993. Effects of ultraviolet-B radiation on terrestrial plants. p. 427-471. In A.R. Young et al. (ed.) Environmental UV Photobiology. Plenum Press, New York.
Brandle, J.R., W.F. Campbell, W.B. Sisson, and M.M. Caldwell. 1977. Net photosynthesis, electron transport capacity and ultrastructure of Pisium sativum L. exposed to ultraviolet-B radiation. Plant Physiol. 60:165-169.
Briscoe, A.D., and L. Chittka. 2001. The evolution of colour vision in insects. Ann. Rev.
Entomol. 46:471-510 Britt, A.B. 1996. DNA damage and repair in plants. Annu. Rev. Plant Physiol. Plant Mol.
Biol. 47.75-100.

26
Brouillard, R. 1988. Flavonoids and flower color. p. 525-538. In J.B. Harborne (ed.) The flavonoids: Advances in research since 1980. Chapman Hall, London, United Kingdom.
Caldwell, M.M. 1971. Solar UV irradiation and the growth and development of higher
plants. Photophysiology 6:131-177.
Caldwell, M.M. 1979. Plant life and ultraviolet radiation: Some perspective in the history of the earth's UV climate. BioScience 29:520-525.
Caldwell, M.M., R. Robrecht, and W.D. Billings. 1980. A steep latitudinal gradient of
solar ultraviolet-B radiation in the arctic-alpine life zone. Ecology 6:600-611. Caldwell, M.M., A.H. Teramura, and M. Tevini. 1989. The changing solar ultraviolet
climate and the ecological consequences for higher plants. Trends Ecol. Evol. 4:363-367.
Carrow, R.N., D.V. Waddington, and P.E. Reike. 2001. Turfgrass soil fertility and
chemical problems. John Wiley & Sons, Inc., Hoboken, NJ. Castle, E.S. 1935. Photic excitation and phototropism in single plant cells. Cold Spring
Harbor Symp. Quant. Biol. 3:224-229. Chalker-Scott, L. 1999. Environmental significance of anthocyanins in plant stress
responses. Photochem. Photobiol. 70:1-9.
Croteau, R., T.M. Kutchan, and N.G. Lewis. 2000. Natural products (secondary
metabolites). p. 1250-1318. In B. Buchanan et al. (ed.) Biochem. Molec. Biol. Plts. Am. Soc. Plant. Phys.
Cuadra, P., J.B. Harborne, and P.G. Waterman. 1997. Increases in surface flavonols and
photosynthetic pigments in Gnaphalium luteo-album in response to UV-B radiation. Phytochemistry 45:1377-1383.

27
Cunningham, F.X., and E. Gantt. 2002. Genes and enzymes of carotenoid biosynthesis in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 49:577-583.
Cuttriss, A., and B. Pogson. 2004. Carotenoids. In: Plant pigments and their
manipulation. Ann. Plant Rev. 14:57-91.
Day, T.A., T.C. Vogelmann, and E.H. DeLucia. 1992. Are some plant life forms more
effective than others in screening out ultraviolet-B radiation? Oecologia 92:513-519.
Dicke, M., A.A. Agrawal, and J. Bruin. 2003. Plants talk, but are they deaf? Trends
Plant Sci. 8:403-405.
Donner, H., L. Gao, and G. Mazza. 1997. Separation and characterization of simple and
malonylated anthocyanins in red onions, Allium cepa L. Food Res. Int. 30:637-643.
Field, T.S., D.W. Lee, and N.M. Holbrook. 2001. Why leaves turn red in autumn. The
role of anthocyanins in senescing leaves of red osier dogwood. Plant Physiol. 127:566-574.
Flint, S.D., and M.M. Caldwell. 1984. Partial inhibition of in vitro pollen germination by
simulated solar ultraviolet‑B radiation. Ecology 65:792-795.
Flint, S.D., and M.M. Caldwell. 2003. A biological spectral weighting function for ozone
depletion research with higher plants. Physiol. Planta. 117:137-144. Floss, H.G. 1997. Natural products derived from unusual variants of the shikimate
pathway. Natural Prod. Rep. 14:433-452.
Foti, M., M. Piattelli, V. Amico, and G. Ruberto. 1994. Antioxidant activity of phenolic
meroditerpenoids from marine algae. Photochem. Photobiol. 26:159-164.

28
Foyer, C.H., M. Lelandais, and K.J. Kunert. 1994. Photooxidative stress in plants. Physiol. Planta. 92:696-717.
Foyer, C.H., P. Descourvieres, and K.J. Kunert. 1994. Protection against oxygen radicals:
an important defence mechanism studied in transgenic plants. Plant Cell Environ. 17:507-523.
Fraser, P.D., and P.M. Bramley. 2004. The biosynthesis and nutritional uses of
carotenoids. Prog. Lipid Res. 43:228-265.
Frederick, J.E., H.E. Snell, and E.K. Haywood. 1989 Solar ultraviolet radiation at the
earths surface. Photochem. Photobio. 50:443-450.
Galston, A.W., and R.S. Baker. 1949. Studies on the physiology of light action II. The
photodynamic action of riboflavin. Am. J. Bot. 36:773-780.
Garcı́a-Viguera, C., and P. Bridle. 1999. Influence of structure on colour stability of
anthocyanins and flavylium salts with ascorbic acid. Food Chem. 64:21-26.
Gould, K.S. 2004. Nature's swiss army knife: The diverse protective roles of
anthocyanins in leaves. J. Biomed. Biotech. 5:314-320.
Greenwood, D.J., G. LeMaire, G. Gosse, P. Cruz, A. Draycott, and J.J. Neeteson. 1990.
Decline in percentage N of C3 and C4 crops with increasing plant mass. Ann. Bot. 66:425-436.
Gross, J., 1991. Pigments in vegetables: chlorophylls and carotenoids. Van Nostrand
Reinhold, NY. Hardy, A.C. 1938. History of the design of the recording spectrophotometer. J. Opt.
Soc. Am. 28:360-363.
Hare, P.D., and W.A., Cress. 1997. Metabolic implications of stress-induced proline
accumulation in plants. Plant Growth Reg. 21:79-102.

29
Hartley, W.N. 1881. On the absorption of solar rays by atmospheric ozone. J. Chem. Soc. 39:111-128.
Heller, W., and G. Forkmann. 1988. Biosynthesis in the flavonoids: p. 319-425. In J.B.
Harborne (ed.) The flavonoids: Advances in research since 1980. Chapman Hall, London, United Kingdom.
Hong, V., and R.E. Wrolstad. 1990. Use of HPLC separation/photodiode array detection
for characterization of anthocyanins. J. Agric. Food Chem. 38:708-71. Huang, Z.-A., D.-A. Jiang, Y. Yang, J.-W. Sun, and S.-H. Jin. 2004. Effects of nitrogen
deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica. 42:357-364.
Irani, N.G., and E. Grotewold. 2005. Light-induced morphological alteration in
anthocyanin-accumulating vacuoles of maize cells. BMC Plant Bio. 5:7. Jackson, J.A., G. Fuglevand, B.A. Brown, M.J. Shaw, and G.I. Jenkins. 1995. Isolation of
Arabidopsis mutants altered in the light regulation of chalcone synthase gene expression using a transgenic screening approach. Plant J. 8:369-380.
Ju, Z., Y. Duan, and Z. Ju. 1999. Effects of covering the orchard floor with reflecting
films on pigment accumulation and fruit coloration in `Fuji' apples. Sci. Hortic. 82:47-56.
Kaiser, T., K. Emmler, B. Weisshaar, E. Schafer, and A. Batschauer. 1995. Promoter
elements of the mustard CHS1 gene are sufficient for light regulation in transgenic plants. Plant Mol. Biol. 28:219-229.
Kakani, V.G., K.R. Reddy, D. Zhao, and A.R. Mohammed. 2003. Effects of ultraviolet‐B
radiation on cotton (Gossypium hirsutum L.). Morph. Anat. Ann. Bot. 91:817-826.
Kerr, J.B., and C.T. McElroy. 1993. Evidence for large upward trends of ultraviolet-B
radiation linked to ozone depletion. Science 262:1032-1034.

30
Kong. J.M., C. Lian-Sai, G. Ngoh-Khang, C. Tet-Fatt, and R. Brouillard. 2003. Analysis and biological activities of anthocyanins. Phytochemistry 64:923-933.
Kopsell, D.A., and D.E. Kopsell. 2006. Accumulation and bioavailability of dietary
carotenoids in vegetable crops. Trends Plant Sci. 11:1360-1385. Koyama, Y., and R. Fujii, 1999. Cis-trans carotenoids in photosynthesis: configurations,
excited–state properties and physiological functions. p. 161-188. In H.A. Frank (ed.) The photochemistry of carotenoids. Kluwer Publishers, Dordrecht, Netherlands.
Krol, M., G.R. Gray, V.M. Hurry, G. Oquist, L. Malek, and N.P.A. Huner, 1995. Low
temperature stress and photoperiod effect on increased tolerance to photoinhibition in Pinus banksia seedlings. Canadian J. Bot. 73:1119-1127.
Kubasek, W.L., B.W. Shirley, A. McKillop, H.M. Goodman, W. Briggs, and F.M.
Ausubel. 1992. Regulation of flavonoid biosynthetic genes in germinating Arabidopsis seedlings. Plant Cell. 4:1229-1236.
Lancaster, J.E., J.E. Grant, C.E. Lister, and M.C. Taylor. 1994. Skin color in apples –
Influence of copigmentation and plastid pigments on shade and darkness of red color in five genotypes. J. Am. Soc. Hort. Sci. 119:63-69.
Langebartels, C., M. Schraudner, W. Heller, D. Ernst, and H. Sanderman Jr. 2002.
Oxidative stress and defense reactions in plants exposed to air pollutants and UV-B radiation. p. 105-137. In D. Inzé and M. Van Montagu (ed.) Oxidative Stress in Plants. Taylor and Francis New York, NY.
Lee, J., R.W. Durst, and R.E. Wrolstad. 2005. Determination of total monomeric
anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: collaborative study. J. of Ass. Analyt. Com. Int. 88:1269-1278.
Lesfrud, M.G., D.A. Kopsell, R.M. Auge, and A.J. Both. 2006. Biomass production and
pigment accumulation in kale grown under increasing photoperiods. Hortscience 41:603-606.

31
Lesfrud, M.G., D.A. Kopsell, and D.E. Kopsell. 2007. Nitrogen levels influence biomass, elemental accumulations, and pigment concentrations in spinach. J. Plant Nut. 30:171-185.
Lohachoompol, V., G. Srzednicki, and J. Craske. 2006. The change of total anthocyanins
in blueberries and their antioxidant effect after drying and freezing. J. Biomed. Biotechnol. 5:248-252.
Lubin, D., and J.E. Frederick. 1991. The ultraviolet radiation environment of the antarctic
peninsula: the roles of ozone and cloud cover. J. Appl. Meteorol. 30:478-493. Lutz, M., I. Agalidis, G. Hervo, R.J. Cogdell, and F. Reiss-Husson. 1978. On the state of
carotenoids bound to reaction centers of photosynthetic bacteria: A resonance raman study. Biochim. Biophys. Acta (BBA) – Bioenergetics. 503:287-303.
Madronich, S., R.L. McKenzie, L.O. Björn, M.M. Caldwell. 1998. Changes in
biologically active ultraviolet radiation reaching the earth's surface. J. Photochem. Photobiol. B: Biol. 46:5-19.
Maliakal, S.K, K. McDonnell, S.A. Dudley, and J. Schmitt. 1999. Effects of red to far-red
ratio and plant density on biomass allocation. Int. J. Plant Sci. 160:723-733. Mancinelli, A.L., F. Rossi, and A. Moroni. 1991. Cryptochrome, phytochrome and
anthocyanin production. Plant physiol. 96:1079-1085. Massacci, A., M. Adelaide-lannelli, F. Pietrini, and F. Loreto. 1995. The effect of growth
at low temperature on photosynthetic characteristics and mechanisms of photoprotection of maize leaves. J. Exp. Bot. 46:119-127.
Matile, P., B.M.P. Flach, and B.M. Eller. 1992. Autumn leaves of Ginkgo biloba L.:
optical properties, pigments, and optical brighteners. Bot. Acta. 105:13-17. McClure, J.W. 1975. Physiology and functions of flavonoids. p. 970-1055. In. J.
Harborne, and T. Mabry (ed.) The flavonoids. Chapman and Hall Ltd., United Kingdom.

32
McCurdy, J.D., J.S. McElroy, D.A. Kopsell, C.E. Sams, and J.C. Sorochan. 2008. Effects of mesotrione on perennial ryegrass (Lolium perenne L.) carotenoid concentrations under varying environmental conditions. J. Ag. Food Chem. 56:9133-9139.
McElroy, J.S., D.A. Kopsell, J.C. Sorochan, and C.E. Sams. 2006. Response of creeping
bentgrass carotenoid composition to high and low irradiance. Crop Sci. 46:2606-2612.
McEwen, C.N., R.G. McKay, and B.S. Larsen. 2005. Analysis of solids, liquids, and
biological tissues using solids probe introduction at atmospheric pressure on commercial LC/MS instruments. Anal. Chem. 77:7826-7831.
McKenzie, R.L., L.O. Björn , A. Bais, and M. Ilyas. 2003. Changes in biologically
active ultraviolet radiation reaching the earth's surface. Photochem. Photobiol. Sci. 2:5-1.
McLennan, A.G. 1987. DNA damage, repair, and mutagenesis. p. 135-186. In J.A.
Bryant and V.L. Dunham (ed.) DNA replication in Plants. CRC Press, Boca Raton, FL.
Mehlenbacher, S.A., and M.M. Thompson. 1991. Inheritance of a chlorophyll deficiency
in hazelnut. Hortscience 26:1414-1416. Mendez, M., D.G. Jones, and Y. Manetas. 1999. Enhanced UV-B radiation under field
conditions increases anthocyanin and reduces the risk of photoinhibition but does not affect growth in the carnivorous plant Pinguicula vulgaris. New Phytol. 144:275-282.
Moncada, M.C., S. Moura, M. João Melo, A. Roque, C. Lodeiro, and F. Pina. 2003.
Complexation of aluminum (III) by anthocyanins and synthetic flavylium salts: A source for blue and purple color. Inorganica Chimica Acta. 356:51-61.
Moradas-Fereira, P.V., P.P. Costa, and W. Mager. 1996. The molecular defenses against
reactive oxygen species in yeast. Mol. Microbiol. 19:651-658.

33
Müller P., L. Xiao-Ping, and K.K. Niyogi. 2001. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 125:1558-1566.
Mustafa, N.R., and R.Verpoorte. 2007. Phenolic compounds in Catharanthus
roseus. Phytochem. Rev. 6:243-258. Neill, S., and K.S. Gould. 1999. Optical properties of leaves in relation to anthocyanin
concentration and distribution. Can J. Bot. 77:1777-1782. Oren-Shamir, M., and A. Levi-Nissim. 1997. UV-light effect on the leaf pigmentation of
Cotinus coggygria ‘Royal Purple’. Sci. Hortic. 71:59-66. Owens, T.G., A.P. Shreve, and A.C. Atbrecht. 1992. Dynamics and mechanisms of
singlet energy transfer between carotenoids and chlorophylls: light harvesting and nonphotochemical fluorescence quenching. p. 170-186. In N. Murata (ed.) In Research in Photosynthesis 4. Kluwer, Dordrecht, Netherlands.
Pascual-Teresa, S. de., C. Santos-Buelga, and J.C. Rivas-Gonzalo. 2002. LC–MS analysis
of anthocyanins from purple corn cob. J. Sci. Food and Agric. 82:1003-1006. Pfeiffer, N.E. 1928. Anatomical study of plants grown under glasses transmitting light of
various ranges of wavelength. Bot. Gaz. 85:427-436. Pfeifer, M.T., P. Koepke, and J. Reuder. 2006. Effects of altitude and aerosol on UV
radiation. J. Geophys. Res. 111. D01203. doi:10.1029/2005JD006444. Pietrini, F., and A. Massacci. 1998. Leaf anthocyanin content changes in Zea mays grown
at low temperature: Significance for the relationship between quantum yield of PS II and the apparent quantum yield of CO2 assimilation. Photosynth. Res. 58:213-219.
Piringer, A.A., and P.H. Heinze. 1954. Effect of light on the formation of a pigment in
the tomato fruit cuticle. Plant Physiol. 29:467-472.

34
Popp, H.W., and F. Brown. 1936. The effect of ultra- violet radiation upon seed plants. p. 853-887. In B.M. Dugger (ed.) Biological effects of radiation. Vol. 2. McGraw-Hill Book Co., New York.
Rabino, I., and A.L. Mancinelli. 1986. Light, temperature and anthocyanin production.
Plant Physiol. 81:922-924. Rind, D., R. Suozzo, N.K. Balachandran, and M.J. Prather. 1990. Climate change and the
middle atmosphere. I: The doubled CO2 climate. J. Atmos. Sci. 47:475-494. Ronen, G., M. Cohen, D. Zamir, and J. Hirschberg. 1999. Regulation of carotenoid
biosynthesis during tomato fruit development: expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant delta. Plant J. 17:341-351.
Ronen, G., M. Cohen, D. Zamir, and J. Hirschberg. 2000. An alternative pathway to β-
carotene formation in plant chromoplasts discovered by map-based cloning of beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. U.S.A. 97:11102-11107.
Rowland, F.S. 2006. Stratospheric ozone depletion. Philos. Trans. R. Soc. London Ser. B.
361:769-790. Rozema, J., J. van de Staaij, L.O. Björn, and M. Caldwell. 1997. Review. UV-B as an
environmental factor in plant life: stress and regulation. Trends in Ecol. 12:22–28. Sarkar, D., P.C. Bhowmik, Y-I. Kwon, and K. Shetty. 2011. The role of proline-
associated pentose phosphate pathway in cool-season turfgrasses after UV-B exposure. Environ. Exp. Bot. 70:251-258.
Schmidt, R.E., and X. Zhang. 2001. Alleviation of photochemical activity decline of
turfgrasses exposed to soil moisture stress or UV radiation. Int. Turfgrass Soc. Res. J. 9:340-346.

35
Schmitt, J., and R.D. Wulff. 1993. Light spectral quality, phytochrome and plant competition. Trends in Ecol. Evol. 8:47-51.
Seigler, D.S. 1998. Plant secondary metabolism. Kluwer Publishers, Norwell, MA. Shackleton, C. 2010. Clinical steroid mass spectrometry: A 45-year history culminating
in HPLC–MS/MS becoming an essential tool for patient diagnosis. J. Ster. Biochem. Mol. Biol. 121:481-490.
Sheahan, J.J. 1996. Sinapate esters provide greater UV-B attenuation than flavonoids in
Arabidopsis thaliana (Brassicaceae). Am J. Bot. 83:679-686. Smith, H. 1982. Light quality, photo perception and plant strategy. Ann. Rev. Plant
Physiol. 33:481-518. Smith, H. 1994. The light environment. p. 377-416. In R.E. Kendrick and G.H.
Kronenberg. (ed.) Photomorphogenesis in plants. Kluwer Academic Publishers, Dordrecht, The Netherlands.
Solomon, S. 1990. Progress towards a quantitative understanding of Antarctic ozone
depletion. Nature 347:347-354. Sponga, F., G.F. Deitzer, and A.L. Mancinelli. 1986. Cryptochorme, Phytochrome and
the photoregualtion of anthocyanin production under blue light. Plant Physiol. 82:952-955.
Stapleton, A.E. 1992. Ultraviolet radiation and plants: Burning questions. Plant Cell. 4:1353-1358.
Strid, Å., and R.J. Porra. 1992. Alterations in pigment content in leaves of Pisium sativum after exposure to supplementary UV-B. Plant Cell Physiol. 33:1015-1023.
Strid, Å., W.S. Chow, and J.M. Anderson. 1994. UV-B damage and protection at the
molecular level in plants. Photosynth. Res. 39:475-489.

36
Sullivan, J.H., and A.H. Teramura. 1988. Effects of ultraviolet-B irradiation on seedling growth in the Pinaceae. Am. J. Bot. 75:225-230.
Taalas, P., J. Kaurola, and A. Kylling. 2000. The impact of greenhouse gases and
halogenated species on the future solar UV radiation doses. Geog. Res. Lett. 27:1127-1130.
Takashi, S., and N. Murata. 2008. How do environmental stresses accelerate
photoinhibition? Trends Plant. Sci. 13:178-182. Tevini, M., W. Iwanzik, and U. Thoma. 1981. Some effects of enhanced UV-B
irradiation on the growth and composition of plants. Planta 153:388-394. Tevini, M. 1993. Effects of enhanced UV-B radiation on terrestrial plants. p. 125-153. In
Tevini M. (ed.) UV-B Radiation and Ozone Depletion. Lewis Publ., Boca Raton, FL.
Wade, H.K., T.N. Bibikova, W.J. Valentine, and G.I. Jenkins. 2001. Interactions within a
network of phytochrome, cryptochrome and UV-B phototransduction pathways regulate chalcone synthase gene expression in Arabidopsis leaf tissue. Plant J. 25:675-685.
Wellburn, A.R. 1994. The spectral determination of chlorophylls a and b, as well as total
carotenoids using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 144:307-313.
Wrolstad, R.E., and M.M. Giusti. 2001. Characterization and measurement of
anthocyanins by UV-visible spectroscopy. Unit F1.2. p. F1.2.1-F1.2.13. In R.E. Wrolstad and S.J. Schwartz (ed.) Current Protocols in Food Analytical Chemistry. John Wiley & Sons, Inc. New York, NY.
Zhang, X., E.H. Ervin, and R.E. Schmidt. 2005. The role of leaf pigment and antioxidant
levels in UV-B resistance of dark - and light - green Kentucky bluegrass cultivars. J. Am Soc. Hort. Sci. 130:836-841.

37
Ziska, L.H, A.H. Teramura, and J.H. Sullivan. 1992. Physiological sensitivity to plants along an elevation gradient to UV-B radiation. Am. J. Bot. 79:863-871.
Zu-Hua, Y., K. Siebke, and U. Heber. 1990. Light dependent changes in leaves of C3
plants. III. Effect of inhibitors of photosynthesis and of developmental state of photosynthetic apparatus on cystolic and vacuolar pH changes. Planta 182:262-269.

38
CHAPTER 2
SUBJECTIVE AND OBJECTIVE COLOR RATINGS OF TURFGRASSES AND THEIR LINKS TO PIGMENTATION AND NITROGEN CONCENTRATIONS.
ABSTRACT
Turfgrass color is used to correlate turfgrass responses to management inputs such as
fertility treatments and response to environmental stresses. Visual perception of color is
considered a crucial measurement of turfgrass quality as it is the quickest method of
judging turfgrass performance. The measurement can be subjective; therefore the digital
analysis of color has been added as a means to add objectivity to standardize color
measurements. The purpose of this study was to test for a correlation between subjective
and objective turfgrass color rankings and chlorophyll pigment concentrations in several
cool season grasses on an area and volume basis. Fifty one cultivars of six different cool
season grasses were used; perennial ryegrass (Lolium perenne L.), Kentucky bluegrass
(Poa pratensis L.), creeping bentgrass (Agrostis stolonifera L.), tall fescue (Schedenorus
phoenix (Scop.) Holub, fine fescues (Festuca spp.), and colonial bentgrass (Agrostis
capillaris L.). Turfgrasses were germinated and maintained in fifteen cm pots for two
months. Subjective analysis was then carried out using a visual rating for greenness on a
1-9 scale.

39
Objective analysis of chlorophyll pigmentation on a mass and area basis was carried out
using a handheld colorimeter on the grass surface and a bench top colorimeter on pigment
extract. Both systems used the CIELab color measurement system. Tissue nitrogen and
chlorophyll content by area and mass were measured. The handheld system measuring
reflectance did not correlate to pigmentation concentration (P=0.05). Visual rating was
closely linked to nitrogen levels and the benchtop system was influenced by pigment
concentrations (P=0.05). Hand held systems using reflectance type measurements may
suffer from excess error due to characteristics such as surface texture and leaf thickness.
INTRODUCTION
Turfgrass color is traditionally ranked on a 1-9 scale with a ranking of one equal
to dead brown turf and nine equal to optimal green turf color in the eyes of a rater.
However, this is a subjective measurement. In trying to develop a rapid method of
measuring turfgrass responses to either stresses or management inputs, researchers have
found success with using handheld reflectance systems in field situations to measure
plant color. The measurement of plant disease damage and plant nutrient deficiencies
have been successfully tested in creeping bentgrass (Agrostis stolonifera L.), and corn
(Zea mays L.), (Nutter et al., 1993; Bausch and Duke, 1996). These systems provided
highly correlated data and were shown to have positively correlated (R) values to visual
ratings and even tended to be less biased (Nutter et al., 1993). There has been success
using systems which are wavelength dependent to indicate biomass status and
chlorophyll (680 – 780 nm) content (Filella and Penuelas, 1994). Wavelengths in the red

40
range 680-780nm are considered the best estimator of leaf area index in (Capsicum
annuum L.) and (Phaseolous vulgaris L.). Water stress can be measured using this
technology also, however this was seen only after the stress was well developed (Nutter
et al., 1993).
The digital color measurement systems in use have different functions. Digital
imaging has been found to be extremely successful (r2=0.99) in correlating Munsell color
chips to nitrogen treatments in creeping bentgrass (Karcher and Richardson, 2003). The
digital system used is also more precise when compared to traditional methods
(Richardson et al., 2001). The digital analysis uses photography and software to develop
ranking systems and has the ability to obtain large volumes of data from one picture, and
separate out colors with excellent reproducibility. Colorimeters, which depend on a
reflective light wavelength and intensity measurement have been used to separate
creeping bentgrass cultivars and different nitrogen treatments (Landschoot and Mancino,
2000; Mangiafico and Guillard, 2005). The method used in our trials is offered as an
alternative method for turfgrass color analysis which can use either handheld reflective
measurements or spectral transmission benchtop measurements and link them to nitrogen
measurement in turfgrass.
A majority of the color-based systems in use are dependent on a reflective
measurement of turfgrass leaf surfaces. However, leaf surfaces vary in many ways due to
environmental conditions and varietal differences. Tall fescue (Schedenorus phoenix
(Scop.) Holub.) cultivars have a leaf thickness of between 0.31 and 0.36mm under
drought stress and a drought free leaf width of 0.44 cm to 0.52 cm (Fu and Huang, 2004).

41
Nitrogen content causes variation in tall fescue leaf width from 0.31 cm to 0.75 cm
(Rademacher and Nelson, 2001). Colorimeter systems do not have the capability to
measure such narrow sizes creating the potential for inaccurate measurements due to
different leaf age and different nitrogen concentrations. Upper and lower leaf surfaces
also vary in thickness. The upper epidermal surface of Kentucky bluegrass can be 0.58
µm and 1.23 µm on the lower epidermal surface. This thickness variation in the leaf
surface plays a role in light absorption, reflection and transmission (Cameron, 1970;
Clark and Lister, 1975). Penetration and reflection of light varies by leaf thickness.
Treatment by UV-B of alfalfa (Brassica campestris L.) resulted in an increase of leaf
thickness by 45% - causing an increase in the amount of light back scattered (Bornman
and Vogelmann, 1991). Variations across and within turfgrass species leaf thickness
could influence reflective type measurements.
Human eyesight uses rods and cones in the retina. The cones are sensitive to
short, medium and long wavelengths and work to absorb opposing colors of red, green
and blue (Brown and Wald, 1964; Roorda and Williams, 1999). The methods of color
measurement by different handheld reflectance systems vary. The systems can use color
rankings from the Munsell system (Escadafal et al., 1989), Hunter color systems or CIE
Lab scales (Commission Internationale de l'éclairage), (Pattee et al., 1991). The use of the
Munsell color system (Munsell, 1915) was designed to rank color by Albert Munsell
(Nickerson, 1940). This is considered the initial color system, but it did not transfer
consistently into digital reproduction and so more recent systems were developed such as
CIELab and Hunter scales (Sharma and Trussell, 1997). These two systems attempt to

42
replicate how the rods and cones work in the retina and are based on a fixed scale so as
not to become device dependent and lacking reproducibility (Lemaire et al., 2005).
A handheld colorimeter system uses measurements in different color spaces and
L* is considered a measure of lightness while a* measures green (-) or red, (+) and b* is
a measure of blue (-) and yellow (+) colors on the wheel. Hue or the color attribute is also
measureable in a degree system to fit the color wheel while chromaticity is used to
measure the lightness (100 = white) or darkness (0 = black) of the color. This represents a
similar system to human eyesight and can be converted into a 3-D image for digital
analysis (American Society for Testing and Materials, 1989). The system emits a diffuse
uniform light source over a set measuring area that is reflected and collected by an optical
fiber cable which enables analysis of color (Minolta, Chroma Meter CR-300 Instruction
manual). The light source can be manipulated to equal daylight conditions D65 or C
which have similar light circumstances but D65 is preferred by CIE. There are other
illuminant types which can replicate indoor lighting. The handheld systems are fast,
simple and field usable (Shimada et al., 2004).
Previous work investigated correlating colorimeter values to creeping bentgrass
cultivars using the CIELab system and the Minolta CR-310 handheld colorimeter. The
results indicated that the hue angle – the component of the system that describes the true
color which is then further influenced by the lightness or chroma of the color – was the
most consistent part of the data and correlated favorably with data collected from visual
rankings (Landschoot and Mancino, 2000). The data, however, were not linked to any
plant characteristic thus offering no ability to replicate the study. CIELab system data can

43
be transferred to tristimulus based values such as XYZ values and so it offers a possible
system that is transferrable between multiple pieces of equipment. This then allows for
comparison of the same characteristics across grass types (Reinhard et al., 2001). Trials
on creeping bentgrass using the normalized difference vegetation indices (NDVI) system
compared to turfgrass pigmentation content indicated an influence on color ratings when
nitrogen, magnesium and iron fertilization rates were varied. The variation in the color
was only explained 36% of the time by chlorophyll pigmentation and that another factor
or combination of factors was responsible for the result (Stiegler et al., 2005).
Extract measurements of chlorophyllous pigments using benchtop colorimeters
can also give color measurements based on either transmittance or reflectance dimensions
(Ahmed et al., 2000). Transmitted or reflected light beams are collected through a lens
which is 8° from perpendicular to the sample surface. The beams are then integrated
using a sphere and flash beams. The flash reduces the noise and allows for higher
intensity light which normally reduces sensitivity of the pigments due to degradation
when exposed to light for an extended period (Joilot and Joilot, 1984).
Spectrophotometers have been used to test color differences between grasses extracts
(Kavanagh et al., 1985). The transmitted light is then passed through a diffraction grating
which separates and measures the wavelengths of light. These systems also possess the
ability to measure in L*a*b* color space (Yudin et al., 1987). The use of extracts requires
lab time however; this can be a downside to the use of benchtop color measurement due
to cost and time while field measurements are easily and quickly carried out.

44
These systems offer opportunities to standardize color system management and
take out subjectivity that can occur between trained and untrained raters of color
(Gooding and Gamble, 1990; Landschoot and Mancino, 1997). There, however, needs to
be a larger bank of color data compiled with these systems. There are limited data in
relation to pigmentation influence on color rating and whether or not surface reflectance
measurements or transmission type measurements are more appropriate for color
analysis. The objective of this study was to discover how chlorophyll pigments influence
color systems analysis and link pigment concentration to a particular trait within the color
system to create a baseline of data for breeders and other end users. The study aims to
determine what influences human color perception which could be related to stress
responses during periods of pigment decline such as exposure to UV-B. It further aims to
discover whether there is a correlation between visual rating and color system ratings.
MATERIALS AND METHODS Plant material
Two separate Turfgrass trials were run from March, 1st 2010 until June, 11th 2010,
and June 15th 2010 until September 30th 2010 at The Ohio State University, Howlett Hall
greenhouse, 680 Vernon Tharp St., Columbus OH, 43210. Six turfgrass varieties were
seeded in a sand based rootzone, of 80/20 volume/volume sand / peat mix. There were 9
cultivars of perennial ryegrass, Kentucky bluegrass, creeping bentgrass, tall fescue, fine
fescues, sheeps fescue (Festuca ovina L.), creeping red fescue (Festuca rubra L.), hard
fescue (Fesutca brevapila Tracey.). Six cultivars of colonial bentgrass were evaluated.

45
The grasses were chosen based on their color rankings from national turfgrass
evaluation program (NTEP) trial results for 2003 – 2007 for all grasses and based on
commercial availability. Cultivars were randomly separated by the rankings of high /
intermediate / low (Top 1/3, Middle 1/3 Lowest 1/3) of grasses on the NTEP evaluation
results.
Turfgrasses were watered with a mist system initially for 30 seconds every 10
minutes during germination (21 days). Once established, grasses were watered twice
daily for 3 minutes. All turfgrass samples were treated with 20-20-20 liquid fertilizer
(Scotts Co. Marysville, OH) at a rate of 4.06 kg ha-1 on a bi weekly basis. Each grass was
then grown out to a height of 10 cm and maintained at this height for two months in a
greenhouse with temperatures maintained at ~20° Celsius.
Data collection
Turfgrass samples were placed in a completely randomized design in full sun
conditions. A total of 44 untrained (not trained in evaluating turfgrass color) and trained
(turfgrass researchers with training and experience in ranking turfgrass color) evaluators
ranked grasses based on their color. Ranking was set with 1 = brown 6 = satisfactory and
9 = optimal dark green. Turfgrasses were irrigated twice on both days to prevent wilt and
plant material was unranked for 1 hour periods post irrigation to prevent any impact of
water remaining on leaf surfaces on rankings. On both occasions grasses were in full sun
for the total period of time during ranking. A total of forty two surveys were collected
between both experiments. Visual ratings by humans were carried out with permission of

46
institutional review board (IRB project #2010E0291) on June 12th and again on
September 22nd 2010.
Turfgrass samples were taken immediately following the visual rating and leaves
were collected by area (40 mm-2) and on a mass basis (0.05 g) with three subsamples
taken from each cultivar and placed in N,N-dimethylformamide (Sigma-Aldrich, St
Louis, MO) to extract chlorophyll, (Moran and Porath, 1980). Samples were stored in
dark conditions for a period of 48–72 hours after this. Extract was subjected to absorption
analysis using a UV-Vis spectrophotometer (Shimadzu Scientific Instruments Inc.,
Columbia, MD) at wavelengths of 664, 647, 625 and 603 nm. Data were calculated using
formulas from Moran, (1982) and chlorophyll totals, chlorophylls A and B were
quantified on an area and mass basis. The premise for this was that the hand-held system
measures directly on a set area on leaf tissue while benchtop systems require tissue
extract for analysis. Correlating color with chlorophyll content based on the area and
mass measurements were designed to fit this variation between color measurement
systems.
Freshly harvested samples were used for reflectance analysis. Turfgrass leaves
were layered four times to create an optically dense stack (Mangiafico and Guillard,
2005), then three light measurements were taken on the top layer (Appendix A), using a
Konica Minolta CRM 310 (Konica Minolta Sensing, Ramsey, NJ). The stacking and
multiple readings were done to reduce variability due to turfgrass surface irregularity.
The layers were then randomly re-ordered and measurements repeated. Samples were re-
configured three times for a total of nine color measurements per pot and thirty six

47
measurements per cultivar. Data were taken for chroma, hue lightness (L*) red/green
(±a*) yellow/blue (±b*). Prior to analysis the system was calibrated with a white plate
provided by the manufacturer.
Color analysis of extract was carried out using the Hunter Colorquest XE system
(Hunter Associates Laboratory Inc., Reston, VA). Turfgrasses were initially tested for
spectral transmission properties to assess which volume of turfgrass was suitable to
reduce high viscosity or high reflectance levels which could distort data. Turfgrass
samples weighing 0.5 g were determined to be most consistent across all grasses used.
Turfgrass samples were then placed in 15 mL glass tubes with 10mL of N,N-
dimethylformamide. N,N-dimethylformamaide was used to extract all color related to
chlorophyll pigments due to its high efficiency (Stiegler et al., 2004). Samples were
placed in reagent for 48hrs in darkness. Extract was then placed in a 20 mL cuvette with
10 mm path length. Each sample was then tested at illuminant settings of D65 and
transmittance settings with data acquired for chroma, hue lightness (L*) green/red (±a*)
yellow/blue (±b*). Prior to analysis, calibration was performed with a white plate
provided by manufacturer and dimethylformamide was used as a blank.
Tissue samples from each grass were taken and dried for 48 hrs at 60 °C prior to
tissue analysis for percentage nitrogen content at this time (A&L Eastern Laboratories,
Richmond, VA).

48
Data analysis and experimental design
Four replications of each cultivar were used in two repeated trials. The pots were
ranked by humans in a completely randomized design in an outdoor setting with full sun
on turfgrasses throughout the day. During the second experiment a precipitation event
forced the stoppage of ranking, this resumed once water was removed from leaf surface
and cloud cover was gone.
Data transformation of a*b* values is required to create the hue value which is in
a degree format to fit onto the 3-D color circle associated with the system. The
conversion formula is:
degrees ((Atan2(b*,a*))
Following this conversion data was tested using robust regression ROBUSTREG
analysis for data influence and outliers (Chen, 2002). Normality tests were carried out
using the Kolmogorov – Smirnov test while skew and kurtosis were tested also. Data was
then analyzed using multivariate principal components analysis with the R program (R
Development core team, 2011; Jaisingh et al., 2006) to gain an in initial understanding on
variable influence on color. The results of the multivariate analysis were then used to
develop regression and correlation data using PROCREG and PROCCORR with SAS
(SAS Institute, Cary, NC). Differences between cultivar color, nitrogen content and
chlorophyll content were analyzed using PROCGLM. All grasses were analyzed
separately and as a completely randomized design.

49
RESULTS
There was some influence from raw values in the data and outliers were discarded
accordingly (P=0.05). Normality tests (Kolmogorov-Smirnov) found the data to be non-
normal (P=0.05) and so data was tested for outliers using PROC ROBUSTREG. Data
with leverage due to errors in entry or large variability were removed. Kurtosis and skew
varied between grasses and also between transmission and reflectance measurements
(Table 2.1). Transmission data was distributed normally for fine fescue, colonial
bentgrass and Kentucky bluegrass. The data for other three grasses were also normally
distributed with tall fescue (0.04) (P=0.05), creeping bentgrass (0.03) and perennial
ryegrass (0.04) according to Kolmogorov-Smirnov tests. All grasses, with the exception
of fine fescue (0.03) and Kentucky bluegrass (0.03), were found to have normal data for
reflectance measurements (Table 2.1). Skew was similar for both transmission and
reflectance though no grass had a normal distribution based on kurtosis and skew analysis
(Table 2.1).
There was some correlation between hue and chlorophyll B in Kentucky
bluegrass (-0.6), colonial bentgrass (-0.6) and creeping bentgrass (-0.3) cultivars for
transmission analysis with the bench system (Table 2.2). The negative value obtained
correlation is due to the values obtained for the -A* measurements in the green region.
Perennial ryegrass, tall fescue and fine fescue had limited to poor correlation with hue,
chroma A* and B* values (Table 2.2). The strongest correlation across all pigments and
transmission values was -0.6 (Chl B – Hue : Colonial bentgrass / Kentucky bluegrass)

50
with all grasses except colonial bentgrass having no correlation between a certain
pigment and measurement value at least once (Table 2.2).
No, or limited, relationships existed between chlorophyll levels or variations in
chlorophyll pigments and visual rating (Table 2.3). Visual rating was closely linked in
particular with nitrogen levels in perennial ryegrass (0.7) but there was limited linkage
overall, either on an area basis or chlorophyll volume basis between visual rating and
chlorophyll content (Table 2.3). Visual analysis and correlation with the reflectance and
transmission data was also very limited with data having a correlation of no more than -
0.5 which occurred when correlating reflectance characteristics to the visual rating of fine
fescue (Table 2.4).
There was a correlation between nitrogen levels and visual rating of green color
(Figure 2.1). Lower levels of tissue nitrogen content led to lower rankings within species.
Neither pigment concentrations nor nitrogen content influenced CIElab measured
reflectance. The transmission analysis was influenced by all of the pigments. Visual
rating was independent of transmission and reflectance while N and grass variety had
positive correlations with human color rankings (Figure 2.1).
There was a very poor relationship found between reflectance measurements for
color and turfgrass pigmentation (Table 2.5). The strongest noted relationship existed
between the red/green (A*) component and blue yellow component (B*) of colonial
bentgrass with a value of 0.5 and -0.5 for both. Grasses such as tall fescue, perennial
ryegrass and fine fescue had an even more limited relationship between pigments and
color measurements (Table 2.5). Hue values were not seen to be linked closely to total

51
chlorophyll in all grasses while reflective analysis did have stronger correlation with Chl
b concentrations (Table 2.5).
The a* and b* values were influenced minimally on their own by chlorophyll
content in perennial ryegrass, creeping bentgrass and Kentucky bluegrass using both
transmission and reflectance methods (Tables 2.2, 2.5). Correlation between turfgrasses
fine fescue, tall fescue and colonial bentgrass and chlorophyll was comparatively stronger
but the strongest relationship was only found to be 0.5 between chlorophyll B content and
A* and B* reflectance measurement (Table 2.5). A correlation of 0.6 and -0.7 was found
between nitrogen levels and reflectance measurements for colonial bentgrass and A* and
B* (Table 2.6).
The reflectance measurements were linked closest to nitrogen content for A* and
B* with the strongest relationships occurring between nitrogen fine fescue, tall fescue
and colonial bentgrass (Table 2.6). Nitrogen levels had the strongest relationship in
reflectance measured hue for fine fescue, tall fescue and colonial bentgrass (-0.4) and a
higher correlation with chroma for the same three grasses (-0.6, -0.5, -0.6). Nitrogen did
correlate with colonial bentgrass in transmission measurements for hue (0.8) but
transmission analysis in general did not link as closely to nitrogen content in fine fescue
or tall fescue. Colonial bentgrass, creeping bentgrass and Kentucky blue grass had
stronger links between color characteristics and tissue nitrogen content when using
transmission analysis (Table 2.6).
Cultivar differences were noted within grasses. They did not follow visual rating
for reflection analysis and there was limited similarity in the rankings from the

52
transmission measurements (Table 2.4). The visual rating did separate cultivars though
not following the same ranking as NTEP evaluations to a large extent. Perennial ryegrass
had similar rankings but not values, while tall fescue had limited separation (P=0.05)
using human data (Table 2.7).
DISCUSSION AND CONCLUSION
The correlation between key components of turfgrass color and objective
measurements using refelectance and transmission systems in this study was limited.
Pigment concentrations in our study had a poor relationship with the handheld measuring
system in particular. This is consistent with Stiegler at al., (2005) who stated that pigment
concentration could only account for 36% of the variability in turfgrass color and that
other factors play a role when using reflective measuring devices. The studies that had
success with the reflectance system included variations in nitrogen rates as part of the
studies (Landschoot and Mancino, 1997; Mangiafico and Guillard, 2005). The variation
in nitrogen rates would presumably enhance differences in turfgrass colors making it
more likely that reflection analysis could distinguish differences similar to human
eyesight. Forcing of differences was not the aim of this trial so nitrogen rates were not
altered.
There was some what more correlation between pigment concentrations in our
study and transmission measurements. The use of the extract and use of the
spectrophotometer equipment may have been more beneficial for this relationship. In
using reflectance systems there also could have been an increase in light scattering from

53
the leaf surfaces, possibly reducing the precision of the results. The isolation of the
pigments through extraction would also have helped the transmission results and offered
a clear influence on the results. There may be a greater opportunity to more strongly
correlate the reflectance measurements if scattering from the leaf surfaces can be
reduced.
Other studies have concentrated on using smooth surface turfgrass leaves such as
Kentucky bluegrass and creeping bentgrass. Turfgrass surfaces which are known for
varied veination and narrower leaves such as the tall fescue, perennial ryegrass and fine
fescues used here offer a problem for attempting to obtain a consistent reflectance type
measurement. In tall fescue for instance with a change in ploidy from 4X to 8X, which
could have played a role in this study, there was a decrease in vein number on the leaf
surface of 14.2 to 10.2 in major veins. Minor veins on average increase from 4.2 to 6.6
with ploidy changes (Byrne et al., 1981). These surface differences and leaf angle play a
role in light reflectance properties; abaxial surfaces reflect greater levels of light in
grasses than adaxial surfaces. The structure of the epidermal layer (Stiff and Powell,
1974; Thomas and Barber, 1974) which varies in grasses plays a large role in their optical
properties (Sheehy, 1975). Glaucous leaves of eucalyptus (Eucalyptus obliqua L’Hér.)
absorbed only 37% of total radiation while green leaves absorbed 50% of total radiation
(Thomas and Barber 1974). These are all possible areas that would affect the use of a
handheld system in our study which could reduce its reproducibility.
For a more accurate assessment of the ability of these handheld systems other
pigments may need to be taken into account when working with reflectance

54
measurements. Flavonoids absorb light up to 430 nm (Saxby, 1964) and thus reach into
the visible light range; variations in their concentrations can occur with leaf age and also
location in the leaf (Morales et al., 2011). The carotenoids also absorb blue light in the
visible spectrum (Zscheile et al., 1942). The pools of carotenoids found in turfgrass
including lutein and the xantophylls can change based on irradiance levels (McElroy et
al., 2006). Anthocyanins also absorb light in the 500-530 nm region (Gould, 2004), and
may play a role in light wavelengths reflected in that region of PAR. We did not measure
these pigments and they may have played a role in creating the variability we found using
the handheld system in this study.
Leaf texture may also play a role in the lack of strong data correlation. Grasses
which possess leaf hairs and trichomes such as buffalograss (Buchloe dactyloides L.) also
offer a problem for this equipment. There may be increased scattering of radiation on
turfgrasses thus increasing diffuse reflectance (Gausman and Cardenas, 1969; Chien and
Sussex, 1996). Texture influences not only turfgrasses as it has been noted in the food
industry that these issues do exist when analyzing color (Giusti and Wrolstad, 2005.)
It could be expected that visual ratings by untrained evaluators in our study would
not follow NTEP rankings due to a lack of training and possible differences in light and
locations. There can also be variation between trained observers and their color rankings
(Landschoot and Mancino, 1997). The value of having trained observers may be lost in
this instance with greater argument for use of objective standardized rankings of color.
Variability in human eyesight and perspective of color allows for variation in comparison
to NTEP trials and also as compared to trained evaluators. Due to location the potential

55
for visual ratings varying is increased with disparities in light intensity and light angle
playing a role in color analysis (Giusti and Wrolstad, 2005). This issue is part of the
reason that there is interest in using a standardized system to analyze turfgrass color. The
transmission and reflectance methods which offer a standardized color system with
CIELab did not follow the visual ratings with NTEP or local visual ratings. This is
despite the fact the two systems are designed to replicate human eyesight in standard
daylight illumination. This is the first time such a broad scale measurement of turfgrass
species and turfgrass cultivars has been tested however, and previous studies did not aim
to link results to NTEP trial results.
Visual analysis is linked closest to nitrogen content, which has been noted before.
Higher nitrogen rates lead to higher color rankings (Guertal and Evans, 2006; Frank et
al., 2004). Transmission and reflection based analysis also linked most strongly with
nitrogen although in this research the correlation values of 0.4 are not considered strong.
Altering nitrogen rates could lead to a more favorable result (Landschoot and Mancino,
1997). The measurement of nitrogen rather than color may prove to be a better option for
future research as a broader database is created for turfgrasses and reflectance-based
measurements using the same field equipment or, at minimum, the same scale.

56
*Indicates significance in data different from normal at P=0.05. § NS, Non significant at P=0.05. † Data considered to have skew of 0 are considered normal. ‡ Data considered to have kurtosis of 0 are considered normal. Table 2.1: Normality test results and data processing results for six cool season grasses evaluated for color using transmission and reflection type measurements post ROBUSTREG analysis.
Transmission Reflectance Grass Skew† Kurtosis‡ Kolmogorov - Smirnov Skew Kurtosis Kolmogorov - Smirnov
Fine fescue 0.6 2.4 0.05 NS§ 0.6 1.5 0.03 * Tall fescue 0.7 1.5 0.04 * 0.5 0.6 0.05 NS
Colonial Bentgrass 0.3 0.8 0.05 NS 0.4 0.5 0.07 NS Creeping bentgrass 0.6 0.8 0.03 * 0.6 1.0 0.07 NS Perennial ryegrass 0.5 1.4 0.04 * 0.4 0.5 0.09 NS
Kentucky bluegrass 0.7 1.3 0.05 NS 0.6 0.7 0.03 *

57
† A measure of color using CIELab system – 3-d color system using lightness / red-green / blue-yellow components. ‡ A measure of intensity of color using CIElab. § The red-green component of the color system. ¶ The blue yellow component of the color system. Correlation values below 0.7 are considered to have limited relationship between components analyzed Table 2.2: Pearsons correlation values for chlorophyll pigmentation and transmission characteristics obtained using a Hunter XE colorquest colorimeter on extracts of six cool season turfgrasses.
Chl Total Chl a Chl b Grass Hue† Chroma‡ A*§ B*¶ Hue Chroma A* B* Hue Chroma A* B*
Fine fescue 0.1 0.2 -0.1 0.1 0.1 0.2 -0.1 0.1 0.1 0.0 0.0 0.1 Tall fescue 0.0 -0.4 -0.3 0.3 0.0 -0.3 -0.3 0.3 0.0 -0.3 -0.3 -0.3
Colonial bentgrass -0.5 0.2 0.3 -0.4 -0.4 0.2 0.2 -0.3 -0.6 0.1 0.4 -0.5 Creeping bentgrass -0.2 0.1 0.1 -0.1 -0.1 0.2 0.2 -0.2 -0.3 -0.2 -0.1 0.0 Perennial ryegrass 0.0 -0.4 -0.2 0.1 0.0 -0.4 -0.2 0.1 -0.0 -0.4 -0.2 0.1
Kentucky bluegrass -0.5 -0.1 0.1 -0.3 -0.4 -0.0 0.1 -0.2 -0.6 -0.4 0.1 -0.4

58
† Correlation values below 0.7 are considered to have limited relationship between components analyzed. Table 2.3: Pearsons correlation values for chlorophyll pigmentation and tissue nitrogen levels visual rating of six cool season turfgrasses.
Pigment / g-1 Pigment / m-2 Grass Chl total Chl a Chl b N Chl total Chl a Chl b N
Fine fescue -0.2† -0.2 -0.2 0.2 -0.4 -0.4 -0.2 0.2 Tall fescue -0.1 -0.0 -0.1 0.1 0.1 0.1 -0.0 0.0
Colonial bentgrass -0.2 -0.2 -0.1 0.2 -0.0 -0.0 0.0 0.1 Creeping bentgrass -0.2 -0.1 -0.2 -0.1 0.1 0.1 0.1 -0.1 Perennial ryegrass 0.1 0.1 0.1 0.7 0.1 0.1 0.1 0.7
Kentucky bluegrass -0.3 -0.3 -0.1 -0.3 -0.1 -0.1 -0.1 -0.3

59
† A measure of color using CIELab system – 3-d color system using lightness / red-green / blue-yellow components. ‡ A measure of intensity of color using CIElab. § The red-green component of the color system. ¶ The blue yellow component of the color system. Correlation values below 0.7 are considered to have limited relationship between components analyzed Table 2.4: Pearsons correlation of visual rating to transmission and reflection color characteristics of six cool season turfgrasses.
Transmission Reflectance Grass Hue† Chroma‡ A§ B¶ Hue Chroma A B
Fine fescue 0.1 0.0 -0.1 0.1 0.1 -0.5 0.3 -0.1 Tall fescue 0.0 0.2 0.0 0.0 0.1 -0.1 0.1 -0.1
Colonial bentgrass -0.1 -0.1 0.1 -0.1 -0.1 -0.0 -0.1 -0.0 Creeping bentgrass 0.1 -0.0 0.2 -0.1 0.1 -0.0 0.0 0.1 Perennial ryegrass -0.1 -0.0 -0.2 0.2 0.0 -0.1 -0.0 -0.0
Kentucky bluegrass -0.1 0.0 0.1 -0.1 0.0 0.1 -0.1 0.1

60
† A measure of color using CIELab system – 3-d color system using lightness / red-green / blue-yellow components. ‡ A measure of intensity of color using CIElab. § The red-green component of the color system. ¶ The blue yellow component of the color system. Correlation values below 0.7 are considered to have limited relationship between components analyzed Table 2.5: Pearsons correlation values for chlorophyll pigmentation and reflection characteristics obtained using a Konica CR310 handheld colorimeter on surfaces of six cool season turfgrasses.
Chl Total Chl a Chl b Grass Hue† Chroma‡ A§ B¶ Hue Chroma A B Hue Chroma A B
Fine fescue -0.4 0.2 0.0 -0.4 -0.4 0.2 0.0 -0.4 -0.4 0.1 0.1 -0.3 Tall fescue -0.1 -0.1 0.2 -0.1 -0.1 -0.1 0.2 -0.1 -0.0 -0.1 0.1 -0.1
Colonial bentgrass -0.2 -0.3 0.4 -0.4 -0.2 -0.3 0.4 -0.4 -0.3 -0.4 0.5 -0.5 Creeping bentgrass 0.0 -0.0 0.0 -0.0 0.0 -0.0 0.0 -0.0 0.0 -0.0 0.0 0.0 Perennial ryegrass 0.1 0.1 -0.1 0.1 0.1 0.1 -0.1 0.1 0.1 0.1 -0.1 0.1
Kentucky bluegrass 0.1 0.0 -0.0 0.1 0.1 0.0 -0.0 0.1 0.0 0.1 -0.1 0.0

61
† A measure of color using CIELab system – 3-d color system using lightness / red-green / blue-yellow components. ‡ A measure of intensity of color using CIElab. § The red-green component of the color system. ¶ The blue yellow component of the color system. Correlation values below 0.7 are considered to have limited relationship between components analyzed Table 2.6: Pearsons correlations between reflectance measurements and transmission measurements with tissue nitrogen for 6 cool season turfgrasses.
Reflectance Transmission Grass Hue† Chroma‡ A§ B¶ Hue Chroma A B
Fine fescue -0.4 -0.6 0.6 -0.7 0.1 0.3 -0.4 0.3 Tall fescue -0.4 -0.5 0.5 -0.5 -0.0 -0.4 -0.2 0.1
Colonial bentgrass -0.4 -0.6 0.6 -0.7 0.8 -0.2 -0.6 0.7 Creeping bentgrass 0.1 -0.0 -0.1 0.1 0.2 0.5 0.3 -0.2 Perennial ryegrass -0.1 -0.1 -0.1 0.1 -0.2 -0.0 -0.4 0.3
Kentucky bluegrass -0.1 -0.2 0.2 -0.2 0.4 0.5 -0.3 0.4

62
Grass Cultivar Colonial bentgrass Fine fescue Kentucky bluegrass Creeping bentgrass Perennial ryegrass Tall fescue
1 6.4 a† 7.7 ab 7.2 ab 6.6 a 6.9 a 8.2 a 2 5.4 c 6.4 c 6.2 def 4.9 b 7.1 a 7.4 b 3 6.1 ab 8.0 a 6.0 ef 5.3 b 6.5 ab 7.2 b 4 6.4 a 8.0 a 6.7 bcd 6.5 a 6.6 ab 8.1 a 5 5.2 c 6.1 c 6.0 ef 5.2 b 5.8 cd 7.0 b 6 5.6 ab 6.1 c 7.4 a 6.6 a 6.2 bcd 7.3 b 7 n/a‡ 5.5 d 7.0 abc 5.1 b 6.4 abc 8.0 a 8 n/a 7.3 b 5.9 f 5.0 b 5.1 e 7.0 b 9 n/a 7.3 b 6.5 cde 5.5 b 5.6 de 7.3 b
LSD§ (P=0.05) 0.5 0.6 0.6 0.7 0.7 0.5 † Data followed by a different letter in each column represent significant differences between cultivars (P=0.05). ‡ Only six cultivars could be found for colonial bentgrass. § Means separation carried out using Fishers least protected means (P=0.05). Table 2.7: Human color rankings for cultivars of six cool grasses. (1= brown / 6 = acceptable / 9 = optimum).

63
† Directional line indicates influence of component on either transmission or reflection analysis. PC = principal component. PC1 = Influence of chlorophyll total, chlorophyll a and b, hue, chroma l* a* b* on transmission color analysis. PC 2 = Influence of chlorophyll total, chlorophyll a and b, hue, chroma l* a* b* on reflection color analysis. Figure 2.1: Principal component analysis for turfgrass samples analyzed for color using reflectance and transmission measurements.

64
REFERENCE Ahmed, J., U.S. Shivhare, and G.D. Singh. 2000. Chlorophyll and color of green chilli
puree as affected by mesh size and temperature. Int. J. Food Propog. 3:305-315. American Society for Testing and Materials. 1989. Standard test method for specifying
color by the munsell system. ASTM standard D1535-89. ASTM, Philadelphia, PA.
Bausch, W.C., and H.R. Duke. 1996. Remote sensing of plant nitrogen status in corn.
Trans. ASAE. 39:1869-1875. Bornmann, J.F., and T.C. Vogelmann. 1991. Effect of UV-B radiation on leaf optical
properties measured with fiber optics. J. Exp. Bot. 42:547-554. Brown, P.K., and G. Wald. 1964. Visual pigments in single rods and cones of the human
retina. Science 1964. 144:45-52. Byrne, M.C., C.J. Nelson, and D.D. Randall. 1981. Ploidy effects on anatomy and gas
exchange of tall fescue leaves. Plant Physiol. 68:891-893. Cameron, R.J. 1970. Light intensity and the growth of Eucalyptus seedlings: II. The
effect of cuticular waxes on light absorption in leaves of Eucalyptus species. Aus. J. Bot. 18:275-284.
Chen, C. 2002. Robust regression and outlier detection with the ROBUSTREG
procedure. Paper 265–27. SAS Institute Inc., Cary, NC, Proceedings SUGI 27, Orlando FL Apr 14–17. Available at http://www2.sas.com/proceedings/sugi27/p265-27.pdf (verified, 28 June, 2012).

65
Chien, J.C., and I.M. Sussex. 1996. Differential regulation of trichome formation on the adaxial and abaxial leaf surfaces by gibberellins and photoperiod in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 111:1321-1328.
Clark, J.B, and G.R. Lister. 1975. Photosynthetic action spectra of trees: II. The
relationship of cuticle structure to the visible and ultraviolet spectral properties of needles from four coniferous species. Plant Physiol. 55:407-413.
Escadafal, R., M-C. Girard, and D. Courault. 1989. Munsell soil color and soil
reflectance in the visible spectral bands of landsat MSS and TM data. Remote Sens. Env. 27:37-46.
Filella, I., and J. Penuelas. 1994. The red edge and shape as indicators of plant
chlorophyll, biomass and hydric status. Int. J. Remote Sensing. 15:1459-1470.
Frank, K.W., R.E. Gaussoin, T.P. Riordan, R.C. Shearman, J.D. Fry, E.D. Miltner, and
P.G. Johnson. 2004. Nitrogen rate and mowing height effects on turf-type buffalograss. Crop Sci. 44:1615-1621.
Fu, J., and B. Huang. 2004. Leaf characteristics associated with drought resistance in tall
fescue cultivars. Acta Hortic. 661:233-239. Gausman, H.W., and R. Cardenas. 1969. Effect of leaf pubescence of Gynura aurantiaca
on light reflectance. Bot. Gaz. 130:158-162. Giusti, M.M., and R.E. Wrolstad. 2005. Characterization and measurement of
anthocyanins by UV-visible spectroscopy. p. 19–32. Unit F1.2. In Wrolstad R.E. (ed.) Handbook of food analytical chemistry—pigments, colorants, flavors, texture, and bioactive food components. John Wiley & Sons Inc., New York.
Gooding, M.J., and L.J. Gamble. 1990. Color evaluation of Poa pratensis cultivars. J.
Sports Turf Res. Inst. 66:134-140. Gould, K.S. 2004. Nature's swiss army knife: The diverse protective roles of
anthocyanins in leaves. J. Biomed. Biotech. 5:314-320.

66
Guertal, E.A., and D.L. Evans. 2006. Nitrogen rate and mowing height effects on TifEagle bermudagrass establishment. Crop Sci. 46:1772-1778.
Jaisingh, J., E.W.K. See-To, and K.Y. Tamin. 2006. The impact of open source software
on the strategic choices of firms developing proprietary software. J. Manage. Info. Sys. 25:241-275.
Joliot, P., and A. Joliot. 1984. Electron transfer between the two photosystems. I. flash excitation under oxidizing conditions. Biochim. Biophys. Acta. 765:210-218.
Karcher, D.E., and M.D. Richardson. 2003. Quantifying turfgrass color using digital
image analysis. Crop Sci. 43:943-951.
Kavanagh, T., D. O’Beirne, and T.P. Cormican. 1985. Use of the hunter colour difference
meter for colour evaluation in turfgrass experiments. p. 255-261. In: F. Lemaire (ed.) Proc. 5th Intl. Turfgrass Res. Conf., Avignon, France. 1-5 Jul.
Landschoot, P.J., and C.F. Mancino. 1997. Assessment of the minolta CR-310 chroma
meter for predicting nitrogen status of Agrostis stolonifera L. Intl. Turfgrass Soc. Res. J. 8:711-718.
Landschoot, P.J., and C.F. Mancino. 2000. A comparison of visual vs. instrumental
measurement of color differences in bentgrass turf. HortScience 35:914-916. Lemaire, G., G. Escadeillas, and E. Ringot. 2005. Evaluating concrete surfaces using an
image analysis process. Cons. Build. Mat. 19:604-611. Mangiafico, S.S., and K. Guillard. 2005. Turfgrass reflectance measurements,
chlorophyll, and soil nitrate desorbed from anion exchange membranes. Crop Sci. 45:259-265.
McElroy, J.S., D.A. Kopsell, J.C. Sorochan, and C.E. Sams. 2006. Response of creeping
bentgrass carotenoid composition to high and low irradiance. Crop Sci. 46:2606-2612.

67
Morales, L.O., R. Tegelberg, M. Brosché, A. Lindfors, S. Siipola, P.J. Aphalo. 2011. Temporal variation in epidermal flavonoids due to altered solar UV radiation is moderated by the leaf position in Betula pendula. Physiol. Planta. 143:1399-3054.
Moran, R. 1982. Formulae for determination of chlorophyllous pigments extracted with
N,N- dimethylformamide. Plant Physiol. 69:1376-1381. Moran, R., and D. Porath. 1980. Chlorophyll determination in intact tissues using N,N –
dimethylformamide. Plant Physiol. 65:478-479. Munsell, A.H. 1915. Atlas of the munsell color system. Wadsworth-Howland and
Company, Malden, MA. Nickerson, D. 1940. History of the munsell color system and its scientific application. J.
Optic. Soc. Am. 30:575-586. Nutter, F.W., Jr., M.L. Gleason, J.H. Jenco, and N.C. Christians. 1993. Assessing the
accuracy, intra-rater repeatability and inter-rate repeatability of disease assessment systems. Phytopathology 83:806-812.
Pattee, H.E., F.G. Giesbrecht, and C.T. Young. 1991. Comparison of peanut butter
color determination by CIELAB L*,a*,b* and Hunter color-difference methods and the relationship of roasted peanut color to roasted peanut flavor response. J. Agric. Food Chem. 39:519-523.
R Development Core Team. (2011). R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. Available at http://www.R-project.org/ (Verified 29 Jun. 2012).
Rademacher, I.F., and C.J. Nelson. 2001. Nitrogen effects on leaf anatomy within
intercalary meristems of tall fescue leaf blades. Ann. Bot. 88:893-903. Reinhard, E., M. Adhikkmin, B. Gooch, and P. Shirley. 2001. Color transfer between
images. Comp. Graphic. App. 21:34-41.

68
Richardson, M.D., D.E. Karcher, and L.C. Purcell. 2001. Quantifying turfgrass cover using digital image analysis. Crop Sci. 41:1884–1888.
Roorda, A., and D.R. Williams. 1999. The arrangement of the three cone classes in the
living human eye. Nature 397:520-522. SAS Institute. 2011. Release 9.2 SAS Inst. Cary, NC. Saxby, M.J. 1964. Characterization of flavonoids and coumarins by their absorption
spectra on paper chromatograms. Anal. Chem. 36:1145-1146. Sharma, G., and H.J. Trussell. 1997. Digital color imaging. Trans. Image Proc. 6:901-
932. Sheehy, J.E. 1975. Some optical properties of leaves of eight temperate forage grasses.
Ann. Bot. 39:377-386. Shimada. K., Y. Kakehashi, H. Matsumura and N. Tanoue. 2004. In vivo quantitative
evaluation of tooth color with hand-held colorimeter and custom template. J. Prosth. Dent. 91:389-391.
Stiegler, J.C., G. Bell and N. Maness. 2004. Comparison of acetone and N,N-
dimethylformamide for pigment extraction in turfgrass. Comm. Soil Sci. Plant Anal. 35:1801-1813.
Stiegler, J.C., G.E. Bell, N.O. Maness, and M.W. Smith. 2005. Spectral detection of
pigment concentrations in creeping bentgrass golf greens. Int. Turf. Res. J. 10:818-825.
Stiff, M.L., and J.B. Powell. 1974. Stem anatomy of turfgrass. Crop Sci. 14:181-186. Thomas, D.A., and H.N. Barber. 1974. Studies on leaf characteristics of a cline of
Eucalyptus urnigera from Mount Wellington, Tasmania. II. Reflection, transmission and absorption of radiation. Aust. J. Bot. 22:701-707.

69
Wang, Y., M. Zhang, X.Y. Zhang, and H.D. Dong. 2002. Comparative studies on elasticity and scrape tolerance characters of Zoysia japonica and Poa pratensis. Prataculture Sci. 19:56-59.
Yudin, L.S., C.W. Mitchell, and J.J Cho. 1987. Color preference of thrips (Thysanoptera: Thripidae) with reference to aphids (Homoptera: Aphididae) and leafminers in Hawaiian lettuce farms. J. Econo. Entomol. 80:51-55.
Zscheile, F.P., J.W. White, Jr., B.W. Beadle, and J.R. Roach. 1942. The preparation and
absorption spectra of five pure carotenoid pigments. Plant Physiol. 17:331-346.

70
CHAPTER 3
INFLUENCE OF ULTRAVIOLET LIGHT ON GERMINATION RATE AND SPEED OF KENTUCKY BLUEGRASS (Poa pratensis L.)
ABSTRACT
Kentucky bluegrass (Poa pratensis L.) is a commonly used turfgrass in the
temperate climates of the United States. The grass is predominantly established from sod,
because of its slow time to germinate (21 d). Ultraviolet light has been noted to enhance
germination speed of other crops and it may similarly enhance Kentucky Bluegrass
germination. Turfgrass seeds from two seed lots were placed in two separate germination
chambers and set on an 8hr / day 16hr / night schedule. Supplemental ultraviolet light
(11.2 kJ / m-2 / d-1) was added on a treatment of two 1 hr time periods split over a 3 hour
period versus no UV light. The treatments were applied for 21 days. Seed germination
was counted at 7, 14 and 21 days. Ultraviolet light increased the germination
completeness and speed of germination in the newer but not older seed lot. Seed
germination was greater (P=0.05) at day 7 and after 21 days there was a greater
germination percentage or completeness in ultraviolet light conditions than in control in
seed lot one but not lot two. These results suggest that treatment of bluegrass seed with a
UV light may enhance germination; however seed age may override the effect.

71
INTRODUCTION
Turfgrass seed germination is dependent on a number of environmental factors.
Manipulating the light environment may be a method to enhance germination speed and
completeness. Kentucky bluegrass (Poa pratensis L.) may reach only 60% germination
after 28 days and 90% germination after 4 months (Maguire and Steen, 1971). This is a
low rate of germination and leads to difficulties in establishment of the turfgrass.
Treatments to enhance this have included the use of hydrogen peroxide to increase fresh
weight of seed and germination by 20% and 30% respectively (Takabe, 2004). There are
limited methods to improve its germination completeness and hence the establishment of
Kentucky bluegrass lawns is limited by speed of seed germination. Kentucky bluegrass is
considered a favorable turfgrass in the northern United States variety due to its spreading
growth habit and desirable color especially when nitrogen rates are increased (Skogley
and Ledeboer, 1968).
Methods to enhance germination speed and vigor of grass seedlings include seed
placement in favorable moisture conditions prior to seeding. This resulted in a 40hr
decrease in germination time of crested wheatgrass (Agropyron cristatum L.) (Keller and
Bleak, 1968). The aim of this method is to have the seed active prior to seeding and thus
naturally increase the germination rate. Priming seed with initial hydration followed by
air drying has proven to increase germination speed and seedling fresh mass (Pill et al.,
1997) in Kentucky bluegrass. Osmoconditioning or pre-treating with water, of perennial
ryegrass (Lolium perenne L.) seeds increased germination rate and root growth even
under suboptimum temperatures (Danneberger et al., 1992). Sonication for 4 minutes

72
significantly increased germination in orchids (Calanthe discolor L.) by 50% (Miyoshi
and Mii, 1988). This was due to a perceived requirement for scarification of a thicker
seed coat. Different energy treatments using microwaves removes fungal propagules thus
reducing the need for fungicide applications and this had no effect on germination rates
(Cavalcante and Muchovej, 1993). Gamma radiation of tall fescue (Schedenorus phoenix
(Scop.) Holub) reduces endophyte levels by ~90% while not affecting germination
(Bagegni et al., 1990).
The potential for the use of ultraviolet-B (UV-B) light in promoting germination
has received no attention in turfgrass seedling studies. Light is crucial in controlling plant
germination in Kentucky bluegrass (Toole and Borthwick, 1971), as well as plant growth
and development (Frankhauser and Chory, 1997). Ultraviolet light, (200-400nm) has
higher energy compared to photosynthetically active radiation (400-700nm), (PAR). The
impact of ultraviolet-C (UV-C) has been found to be inhibitory in sunflower (Helianthus
annuus L.). This effect, however, can be reversed during dark storage of seeds after the
UV-C treatment (Torres et al., 1991). Kale, cabbage, (Brassica oleracea), radish
(Raphanus sativus L.) and agave (Agave L) treated with UV-B light germinated more
rapidly but subsequent growth was markedly inhibited (Noble, 2002). The number of
seeds and dry weight of seeds in Bromus catharticus receiving 90% UV-B almost
doubled compared to the control (Deckmyn and Impens, 1998). There was, however, not
an effect on germination. The objective of this study was to investigate if ultraviolet light
could be used as a treatment to speed germination of Kentucky bluegrass and to increase
average germination completeness.

73
MATERIALS AND METHODS
Plant material
Kentucky bluegrass seeds from two different seed lots based on age of the same
cultivar ‘Bariris’ were used in this trial (Barenbrug USA, Tangent, OR). Fifty turfgrass
seeds were placed in each of 9 petri-dishes (9 cm diam.) for a total 450 seeds per light
treatment. Two repeated studies for each lot were carried out in each of two growth
chambers with a day / night regime of 8 / 16 hours and temperatures of 25 / 15° C. The
visible light (400 nm – 700 nm) intensity was measured using cosine corrected light
meters (Spectrum Technologies, Plainfield, IL). Ultraviolet light was provided by four
Reptisun 10.0 UV overhead fluorescent tubes (Zoo Med Laboratories, San Luis Obispo,
CA) in one chamber. The ultraviolet light was measured using a portable IL1350
radiometer with UV-B sensor (International Light, Newburyport, MA). The UV light was
then reported as kJ / m-2 / d-1. Each study was conducted for twenty-one days. The lights
were switched between chambers for each experiment. The petri dishes were covered in
both chambers for 21 hours per day for moisture retention. Seeds were placed on standard
blue blotters (Anchor Paper Co., Plymouth, MN), to ensure seed stability and consistent
moisture levels. Prior to daily UV treatment initiation, covers were removed to prevent
any reflection of UV rays from seed surfaces.
Data collection and analysis
Initial treatment of seed with distilled H2O at 5 mL per day occurred and this then
decreased to 3 mL per day after 14 d as seed volume and water requirements decreased.

74
Radicle emergence was used to determine germination and seed count data were
collected at 7, 14 and 21 days. Germination rate based on radicle emergence against total
seeds used and germination vigor based on seed counts were analyzed. The petri dishes
were set up in a randomized complete block design to reduce the impact of lighting
variability. Data were analyzed using the general linear models (GLM) procedure of the
SAS program (SAS Institute, 2008). The data from both seed lots were pooled from each
repetition. Fisher’s protected least significant difference testing (LSD) was used to test
for differences in the average germination values and total germination percentages. Data
were combined to check for differences between seed lots.
RESULTS AND DISCUSSION
Treatments of UV totaled 11.2 kJ m-2 d-1 in the UV chamber, and daily PAR was
4.3 mol m-2 d-1. The control chamber had a total PAR of 4.2 mol m-2 d-1 but no detectable
level of UV light. There was no supply of UV light in the UV-A region (320 – 400 nm) in
either chamber. Due to significant differences (P=0.05) in overall percentage germination
between the two seed lots data were analyzed separately. The newer seed lot which was
one year old (Lot 1), had significantly higher (P=0.05) germination rates compared to the
older seed lot (>2 years old) in UV (Table 3.1). There was lower germination rates in
control light environment in Lot 1 compared to Lot 2 (Table 3.1). The age of the seed
may play a role in the germination percentage change in the UV chamber as has been
noted previously where older seeds had reduced germination percentages (Shaidaee et al.,
1969).

75
Overall total germination percentage was significantly higher (Figure 3.1) in lot 1
for UV treated seeds compared to control seeds. In tests on lot two UV germination
decreased from lot one. The control total germination percentage rates in lot two
increased from tests on lot one. This meant there was no overall separation between the
germination rates due to the light treatments (Figure 3.2).
Days to germination of lot one was affected by light treatments. Applications of
ultraviolet light significantly (P=0.05) enhanced seed germination on day 7 compared to
the control treatment. The cumulative germination percentage was still higher at day 14
for the UV chamber in lot one but not lot two (Figure 3.1 and 3.2). On day 21 both sets of
light treatments were significantly different in lot one. The observation of higher
germination on day 7 resulted in significantly faster germination times in the UV treated
grass chambers (Figure 3.1). The pattern of greater seed germination (P=0.05) on day 7
in the first experiment was also repeated in the second experiment for seed lot one
(Figure 1). Seed lot one almost reached germination completeness after 21 days in UV,
however data was not collected past this date.
The same results were not observed with the 2nd seed lot. UV treatment did not
result in increased germination by day 7 in seed lot 2 (Figure 3.2). The initial difference
at day 7 from seed lot one was not noticed in the second seed lot. The second seed lot
demonstrated the greatest percentage of germination on day 21 in both experiments and
there was no significant difference between light treatments (Figure 3.2). Overall
germination decreased in UV conditions in lot 2 compared to lot 1 while there was an
increase in germination from lot one to seed lot two in control conditions (Table 3.1).

76
CONCLUSIONS
The variation between the seed lots means that further work is required in order to
conclusively determine if a clear benefit due to UV irradiance exists. The initial results
were very promising and agree with data produced from lettuce and wheat seed (Noble,
2002, Wagné, 1966). The amount of published work done with ultraviolet light on
turfgrass germination is limited and variability in seed lots due to age and weight has
been noted previously regardless of light treatment (Larsen and Andreasen, 2004,
Shaidaee et al., 1969). This may be a large part of the reason for the difference in results
and a specific cultivar which has more readily available seed lots of similar age would be
desirable.
The use of specific wavelengths also requires investigation. The experimental
system provided elevated UV-B levels but it also had UV-A wavelengths. This was due
to a size restriction with UV-B specific lights. If the study focused solely around the main
UV-B wavelengths it may enhance the biologically activity spectrum (Caldwell, 1971). It
has been noted in that UV-C (100 nm - 290 nm) does not provide potential for
germination enhancement, with inhibition of sunflower germination (Helianthus annus
L.), (Torres et al., 1991). The wavelengths and energy associated with ultraviolet light
may have triggered response from the light receptor phytochrome – A, especially at low
fluence rates which irreversibly trigger germination (Shinomura et al., 1996).
Field testing is also a step that has to be considered as well as combing hydration
and temperature cycles with the UV irradiation in future studies. It does offer a potential
option however to produce enhanced Kentucky bluegrass germination if consistency can

77
be found with the seed lots. This would allow for greater seasonal use of the grass in
home lawn establishment.

78
Light Ultraviolet Control
Lot Df Germination # --------------------------- % --------------------------- 1 1 81.6 a† 60.3 b 2 1 71.6 b 70.7 a
LSD (0.05) 5.8 8.9 † Means followed by different letter were significantly different (P=0.05)
Table 3.1: Germination percentages of Kentucky bluegrass (Poa pratensis L.) seed lots treated with PAR (400 – 700 nm) and ultraviolet light (300 – 400 nm).

79
Figure 3.1: Percentage germination of Kentucky bluegrass (Poa pratensis L.) seed treated with photosynthetically active radiation (400 – 700 nm) and ultraviolet light (280 – 480 nm) in newer seed lot one.

80
Figure 3.2: Percentage germination of Kentucky bluegrass (Poa pratensis L.) seed treated with photosynthetically active radiation (400 – 700 nm) and ultraviolet light (280 – 480 nm) in older seed lot two.

81
REFERENCE Bagegni, A.M., D.A. Sleper, H.D. Kerr, and J.S. Morris. 1990. Viability of Acremonium
coenophialum in tall fescue seed after ionizing radiation treatments. Crop Sci. 30:1272-1275.
Caldwell, M.M. 1971. Solar ultraviolet radiation and the growth and development of
higher plants. p. 131-174. In: A.C. Giese (ed.) Photophysiology. Academic Press, New York.
Cavalcante, M.J.B., and J.J. Muchovej. 1993. Microwave irradiation of seeds and
selected fungal spores. Seed Sci. Technol. 21:247-253. Danneberger, T.K., M.B. McDonald Jr., C.A. Geron, and P. Kumari. 1992. Rate of
germination and seedling growth of perennial ryegrass seed following osmoconditioning. HortScicience 27:28-30.
Deckmyn, G., and I. Impens. 1998. Effects of solar UV-B irradiation on vegetative and
generative growth of Bromus catharticus. Environ. Exp. Bot. 40:179-185. Frankhauser, S., and J. Chory. 1997. Light control of plant development. Ann. Rev. Cell
Dev. Biol. 13:203-29. Keller, W., and A.T. Bleak. 1968. Preplanting treatment to hasten germination and
emergence of grass seed. J. Range Manage. 21:213-216. Larsen, S.U., and C. Andreasen. 2004. Light and heavy turfgrass seeds differ in
germination percentage and mean germination thermal time. Crop Sci. 44:1710-1720.

82
Larsen, S.U., and B.M. Bibby. 2004. Use of germination curves to describe variation in germination characteristics in three turfgrass species. Crop Sci. 44:891-899.
Maguire, J.D., and K.M. Steen. 1971. Effects of potassium nitrate on germination and
respiration of dormant and non dormant Kentucky bluegrass (Poa pratensis L.) seed. Crop Sci. 11:48-50.
Miyoshi, K., and M. Mii. 1985. Ultrasonic treatment for enhancing seed germination of
terrestrial orchid Calanthe discolor in a symbiotic culture. Sci. Hortic. 35:127-130.
Noble, R.E. 2002. Effects of UV-irradiation on seed germination. Sci. Total Environ.
299:173-176. Pill, W.G., J.J. Frett, and I.H. Williams. 1997. Matric priming of Kentucky bluegrass and
tall fescue seeds benefits seedling emergence. HortScience 32:1061-1063. Shaidaee, G, B.E. Dahl, and R.M. Hansen. 1969. Germination and emergence of different
age seeds of six grasses. J. of Range Manage. 22:240-243. Shinomura, T., A. Nagatani, H. Hanzawa, M. Kubota, M. Watanabe, and M. Furuya,
1996. Action spectra for phytochrome A and B-specific photoinduction of seed germination in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U.S.A. 15:8129-8133.
Skogley, C.R., and F.B. Ledeboer. 1968. Evaluation of several Kentucky bluegrass and
red fescue strains maintained as lawn turf under three levels of fertility. Agron. J. 60:47-49.
Takabe, T., A. Uchida, and T. Tetsuko. 2004. Effects of hydrogen peroxide on germination of Kentucky bluegrass. In Abstracts, Abs # 709. Plant Biology 2004: July 24 – July 28, 2004. Lake Buena Vista, FL.
Toole, V.K., and H.A. Borthwick. 1971. Effect of light, temperature and their interactions
on seeds of Kentucky bluegrass (Poa pratensis L.). J Am. Soc. Hort. Sci. 96:301-304.

83
Torres, M., G. Frutos, and J.M. Duran. 1991.Sunflower seed deterioration from exposure to UV-C radiation. Environ. Exp. Bot. 31:201-207.
Wagné, C. 1966. Effect of UV light on lettuce seed germination and on the unfolding of
grass leaves. Physiol. Planta. 19:128-133.

84
CHAPTER 4
ULTRAVIOLET-B LIGHT INFLUENCE ON COOL SEASON TURFGRASS GROWTH AND MORPHOLOGY
ABSTRACT
Ultraviolet light is a potential issue for plant growth and development due to its
effect on photosynthesis and plant productivity. Turfgrass plants may be affected by
ultraviolet light causing loss of quality and ability to recover from injury. The objective
of this study was to investigate the effects of ultraviolet light on turfgrass productivity
and morphology in three cool season grasses. Tall fescue, (Schedonorus phoenix (Scop.)
Holub.), creeping bentgrass (Agrostis stolonifera L.), cultivars ‘Penncross’, ‘L-93’ and
perennial ryegrass (Lolium perenne L.), were selected due to limited or no information on
their growth and development in elevated ultraviolet light conditions at lawn height of
10-12 cm. The impact of ultraviolet light treatment on color, root and shoot biomass and
tillering was measured over a four week period of exposure in repeated experiments.
Ultraviolet-B light was tested in particular and light levels were measured at 16 kJ m-2 d-1
biologically effective light in growth chambers programmed for a day/night regime of
14/10 hours. Ultraviolet-B light significantly (P=0.05) reduced shoot and root production
in all grasses and inhibited tiller production (P=0.05). Turfgrasses exposed to this level of
ultraviolet-B light at lawn height of cut showed visual damage and significantly lower

85
color rankings after 2 weeks. The experiment demonstrated that after extended exposure
to ultraviolet-B light there is a loss of productivity and color in cool season turfgrasses.
INTRODUCTION
Light plays a crucial role in turfgrass growth and development. It is normally
considered most important in the wavelengths of 400-700 nm; photosynthetically active
radiation (PAR), (Pons et al., 1993). Light wavelengths can often vary and spectral
distribution can fluctuate with environmental conditions (Bell et al., 2000). There is
interest in how the degradation of the ozone layer will influence spectral distribution on
the earth’s surface (Manney et al., 2011). The decrease in ozone is predicted to lead to a
significant increase in exposure to wavelengths in the ultraviolet (UV) range (Kerr and
McElroy, 1993). Ultraviolet light is found in the range of 10 – 400 nm and the energy
associated with these wavelengths is considered damaging when compared to PAR
(Fiscus and Booker, 1995). The UV-B wavelengths which hit the surface of the earth are
predicted to increase in springtime radiation by 50-60% from 2010-2020 (Taalas et al.,
2000) in the northern hemisphere. The reduction in ozone and a resulting increase in UV
irradiance has been linked to chlorofluorocarbons in the atmosphere (Rowland, 1990).
It has been shown that leaf area indices of some Poaceae species can change in
response to different levels of biologically effective UV-B irradiance (Caldwell, 1971;
Deckmyn and Impens, 1999). Increases in leaf and axillary shoot production in response
to UV-B exposure are noted in both monocot crops such as wheat (Triticum aestivum L.),
and dicots such as beans (Phaseolus vulgaris L.), (Barnes et al., 1990). Red fescue

86
(Festuca rubra L.) growth was positively affected by UV-B radiation in all seasons
(Deckmyn and Impens, 1999), showing potential for differences in response to exposure.
Epidermal thickness alters leaf absorbance properties so much that it is considered an
excellent predictor of the effectiveness of a plant’s ability to screen UV-B light (Day,
1993). Root length was inhibited at UV-B irradiances of 2.5 – 5 kJ m-2 d-1 in Antarctic
hairgrass (Deschampsia antartica L.), while there was an increase in tillering but no
overall impact in the relative growth rate due to the compensation in tiller production
compared to root loss (Rozema et al., 2001). Reductions in stem elongation and leaf
expansion in young plants of long spined thorn apple (Datura ferox L.) occur in UV-B
light (Ballare et al., 1996). Smaller leaf sizes in white clover (Trifolium repens L.) also
have been influential in inferring tolerance to UV-B exposure (Hoffman, 2000). In bean
plants, low levels of PAR, combined with UV-B treatments, led to a decrease in leaf area
of 47% and a decrease in leaf dry weight of ~25% (Cen and Bornman, 1990). Ultraviolet
light exposure is linked to reduced photosynthesis due to direct damage to photosystem II
(Brandle et al., 1977), which results in decreased leaf blade and internode lengths and
plant productivity.
Varietal differences in rice (Oryza sativa L.) following exposure to UV have been
observed. Some cultivars had smaller leaf areas, stunted plant height and a reduction in
relative growth rate whereas others showed little response. Plants with rapid growth rates
such as squash (Cucurbita pepo L.) tend to be more sensitive to damage while slower
growing plants such as peppers (Capsicum frutescens L.) are tolerant of the UV-B
irradiance (Smith et al., 2000). These effects were not noted until after 2 weeks of UV-B

87
treatment (Dai et al., 1992). Perennial ryegrass and orchardgrass (Dactylis glomerata L.)
biomass production was reduced in response to comparable spring and summer
treatments of elevated UV levels (Deckmyn and Impens, 1999). In poplars (Populus
cathayana Rehd.), however, there was no effect on biomass following exposure to
different levels of UV-B (Zhang et al., 2011). Variability among plant species in response
to UV-B has been noted in review papers and could be related to other issues such as
temperature, or concentrations of CO2 (Krupa and Nickert, 1989). Limited work has been
conducted to determine the impact of UV-B on vegetative productivity of turfgrasses that
would be used in homelawn situations covering potentially 163,800 km-2 ± 35,850 km-2
in the United States (Milesi et al., 2005).
Ultraviolet light has varying effects on quality ratings among the turfgrasses.
Chewings fescue (Festuca rubra L. ssp. commutata Gaud.), is considered the most UV-B
tolerant turfgrass with Kentucky bluegrass being considered the least tolerant (Zhang and
Ervin, 2009). Quality ratings were reduced by as much as 74% for Kentucky bluegrass
after 10 days of exposure. There was also cultivar variation in the tolerance of UV-B in
this trial. Darker green Kentucky bluegrass cultivars are thought to be also more tolerant
of UV-B conditions (Ervin et al., 2004c). In other plants such as apples (Malus
domestica) ‘Royal Gala’, increasing the red color post-harvest with the use of UV-B
treatments has been seen as a benefit to production (Dong et al., 1995). This response was
due to increased production of flavonoid-type pigments. Combined treatments of UV-B
and visible light were most effective in enhancing the color in other apple cultivars such

88
as (Malus domestica Borkh.), cv. ‘Aroma’, (Hagen et al., 2007). The resulting increase in
red color on the skin in both cases added value to the apple crop.
Limited research has been conducted to test the effect of UV-B exposure on
turfgrasses at higher heights of cut such as home lawns and the estimated area covered by
irrigated turfgrass crop covers potentially 163,800 km-2 (Milesi et al., 2005). The
objective of this study was to test four cool season grasses for their growth rate, tillering
and color responses to elevated UV-B conditions predicted to occur in this decade
(Taalas et al., 2000). It was hypothesized that thicker-leaved species, such as perennial
ryegrass and tall fescue would have greater tolerance to UV-B than thin leaved species
like Kentucky bluegrass and creeping bentgrass.
MATERIALS AND METHODS
Sample preparation
Experiments were initiated in March, 2011 and May, 2011 with no noticeable
differences in turfgrasses following establishment in Columbus, Ohio. Turfgrasses were
initially seeded into a mason sand rootzone containing >97% sand (Hummel & Co,
Trumansburg, NY) as a single seed with structured timing to match germination speeds.
The rootzone met United States Golf Association (USGA) classification and had a pH of
7.4 with 2.7% clay silt fraction. Creeping bentgrasses were seeded first into 3.8 cm
diameter ‘conetainers’ (Steuwe & Sons, Tangent, OR) and 4 days later perennial ryegrass
and tall fescue were seeded. Each conetainer contained a single plant. The plants were
placed in a mist house in a ten minute misting cycle of 15 seconds per water application

89
to maintain moisture. Turfgrasses were grown for 6 weeks and initially treated with
Peters 20-20-20 (Scotts-Miracle Gro, Marysville, OH) at a rate of 11.8 kg ha-1 every two
weeks. Six weeks after germination all plants were removed and placed in a greenhouse
maintained at ~20°C for four more weeks to allow for maturation. Imidacloprid N-[1-[(6-
Chloro-3-pyridyl) methyl]-4,5-dihydroimidazol-2-yl]nitramide (Bayer Environmental
Science, Research Triangle PK, NC) was applied to prevent insect damage. Mefenoxam
(R,S)-2-[(2,6-dimethylphenyl)-methoxyacetylamino]-propionic acid methyl ester
(Syngenta AG, Greensboro, NC) and thiophanate-methyl (dimethyl 4,4’-o-
phenylenebis[3-thioallophanate]) (Cleary Chemicals, Dayton, NJ) were applied for
preventative disease control. Water was applied for a 5 minute period daily to prevent
wilt, with automated heads using an ESP modular irrigation system (Rain Bird, Azusa,
CA). Ten weeks after turfgrass emergence, plants were placed in Conviron E15 growth
chambers (Controlled Environment Ltd., Winnipeg, Canada). Plants were left in complete
darkness at 20 °C for 24 hours.
Ultraviolet-B treatment was implemented using specific UV-B lamps that have
peak light emission at 313 nm, QUV UV-B 313 nm, (Q-Lab Corp., Westlake, OH). The
control treatment consisted of only light from fluorescent lamps. The UV-B lamps did not
produce any extra visible radiation. Treatment exposure was calculated based on the
predicted time of exposure and measured lamp irradiance. Radiation from ultraviolet
sources were measured using both UV-B and UV-A sensors attached to an IL1350
photometer (International Light, Newburyport, MA). The radiation is reported in kJ m-2
d-1. Photosynthetic active radiation was measured every 15 minutes for the period of the

90
study using a cosine corrected light sensor (Spectrum Technologies, Plainfield, Il) and
reported as mol m-2 d-1. The chambers were programmed for a day/night length of 14/10
hours. Photosynthetic active radiation (PAR) in the range of 400 – 700 nm was provided
in all chambers using 85 watt s-1 high output F12T12-CW.HO fluorescent tubes (General
Electric, Fairfield, CT). Water was applied twice daily to prevent wilting. Temperature in
each chamber was set to 20°C during light exposure and 17 °C in darkness. Fertility
treatments continued for the duration of each study. Turfgrass plants were placed 0.4 m
below the lighting in an even spacing.
Data collection and analysis
The experiment was designed as a randomized complete block with weeks being
randomized in each block. Under each light regime, there were three blocks with four
replications per block of each grass for a total of 12 samples of each grass per
measurement. Data were analyzed using the linear model (GLM) method in SAS Institute
9.2 (2008). Means separation was carried out using Fishers protected least significant
difference test (P=0.05) and data were pooled. At the end of each week samples were
harvested and separated into roots and shoots, then dried at 70 °C for two days. The
shoot: root ratio was then calculated based on dry weight (Gwynn-Jones and Johanson,
1996). Tissue was dried in a VWR 1350F (Univar USA, Redmond, WA) oven in 8.9 x
16.5 cm coin envelopes (Officemax, Naperville, IL). Relative growth rate was calculated
based on the formula of Gardner et al., (1985):

91
RGR = Relative growth weight; lnW2 = natural log weight 2; lnW1 = natural log weight
1; T1 = Time 1; T2 = Time 2.
RGR = (lnW2 – lnW1) / (T1 – T2)
Tillering was measured by counting daughter plants extending from the crown of
each seeded conetainer. Tillers were counted when samples were harvested for each week
of treatment at weeks 1, 2, 3 and 4. Tillers were defined as daughter plants arising
directly from the crown individual plants. Color rankings were carried out on a 1-9
ranking scale with 1 = dead/brown 9 = dark green 6 = acceptable. The color data were
collected prior to trial initiation and weekly during the trial under the same light
conditions.
RESULTS
Turfgrasses in the UV-B treated environment received a total of 24 kJ m-2 d-1.
Once this was adjusted for UV-B biologically effective radiation the total effective UV-B
light seen by turfgrasses (Caldwell, 1971) was decreased to 16 kJ m-2 d-1 (Table 4.1). The
daily photosynthetic integral totaled 18.5 mol m-2 d-1 (Table 4.1).
After 3 weeks, growth of all grasses in ultraviolet light was significantly inhibited
(P=0.05) compared to grasses grown in the control light environments. There was,
however, a decrease in growth rate in the control chambers after four weeks and so no
significant differences were observed. The growth rate in UV conditions, however, had

92
become negative as turfgrasses began to completely fail (Figure 4.1). There was
variability noted within the data, which occurred despite using twelve different samples
from each grass at each sampling date.
There was a significant decrease (P=0.05) in color quality ratings for all grasses
exposed to UV-B (Figure 4.2). There was a 22% decrease in the ‘L-93’ color and a 17%
decrease in ‘Penncross’ color. Tall fescue and perennial ryegrass had a loss of 18% and
15% in color quality respectively. After four weeks, both creeping bentgrass cultivars
were ranked below acceptable levels and visible damage was clearly seen with browning
and yellowing of the leaf surfaces (Figure 4.2), (Appendix B). Dark lesions similar to
burn spots were also noted on the UV-treated bentgrass plants in the UV-B treatment.
The loss of color was not observed until week two of the experiments and was consistent
in both trials (Appendix B). The color differences were consistent after week two until
the end of each experiment. There was some reddening of stems and tissue material but
this was not consistently observed.
Under UV-B conditions in experiment one there was a clear inhibition (P=0.05)
of plant tillering in three of the grasses (Figure 4.3). There was a 40% inhibition of
tillering in creeping bentgrass ‘L-93’ and 39% decrease in the creeping bentgrass
‘Penncross’ cultivar. Perennial ryegrass was not inhibited significantly in either
experiment. In experiment one, tall fescue tillering was significantly inhibited (P=0.05)
when it was treated with UV-B light. There was a 30% decrease of tall fescue tillering by
30% in experiment one but this did not occur in experiment two. In experiment two, there
was an overall decrease in tillering regardless of light environment. The bentgrasses

93
exposed to UV-B had lower tiller numbers in experiment one but they were not
significantly different from the control treatment in experiment two (Figure 4.4). This
was possibly due to greater light intensities in the maturation of plants in experiment 2
which occurred two months later in the year. Perennial ryegrass tiller number was not
significantly different in response to light regime, between the light treatments. The root :
shoot ratio did not change significantly in response to light regime, but the data were
highly variable.
DISCUSSION
The decrease in growth rate in the bentgrasses after exposure to UV light has been
reported previously (Schmidt and Zhang, 2001). Reducing the efficiency of the
photosynthetic machinery due to photolytic damage would effectively reduce plant
productivity and explain previous findings. Biological variation and seasonal light
changes affect plant maturation and may have affected the turfgrass plants in experiment
two. This had an effect on the control plants in experiment two, thus reducing the
potential for separation of treatment effects. The potential exists for differences in the leaf
surface characteristics of each of the turfgrasses. In particular, differences in leaf
thickness are known to alter light absorption and reflection (Cameron, 1970; Clark and
Lister, 1975). Exposure to UV-B light can also lead to thickening of the leaf surfaces
(Bornman and Vogelmann, 1991), thus affecting plant productivity though not
necessarily in a beneficial manner. The difference in productivity between the grasses
was also of interest. The two thicker-leafed grasses - tall fescue and perennial ryegrass -

94
tended to produce greater biomass levels than the thinner-leafed creeping bentgrass. The
issue of leaf thickness may be crucial in this situation as a plant tries to maintain normal
growth rates in high UV-B conditions.
The inhibition of tillering by UV-B could be considered problematic for turfgrass
wear-recovery from daily use as well as regular growth and development. This reduction
has been noted before in rice but there were also cultivar differences (Teramura at al.,
1991). In that study, 30% of the rice cultivars showed a significant sensitivity to UV-B
irradiance. This was also shown in other rice cultivars at ambient levels of UV-B of 8 kJ
m-2 d-1, where tillering was reduced occurred in cultivar ‘CL161’ but not in ‘Cocodrie’.
The results were not linked to hormonal or carbohydrate changes in the plants
(Mohammed et al., 2007). In Antarctic hairgrass (Deschampsia antarctica), tillering was
increased by UV-B; however, the levels of UV-B were not as high as in this study
(Rozema et al., 2001). The lack of separation in the root and shoot ratio was somewhat
surprising; however, the light levels may have suppressed the growth and allocation of
carbohydrate equally. This does not agree with previous results (Gwynn-Jones, 2001), but
the light levels were higher in this study and it was conducted over a shorter period of 35
days compared to 45 days.
Light intensity variation plays a role in the overall responses of plants to both UV-
B exposure and standard PAR irradiance. Its effect varies regarding the amount of
exposure that is received by plants. In lapland reedgrass, (Calamagrostis lapponica) and
(Calamgrostis pupurea), levels of UV-B simulating 15% loss of ozone reduced tillering.
In light levels simulating 25% ozone depletion, there was an increase in tillering (Gwynn-

95
Jones and Johanson, 1996). The exposure value equated to 6.5 kJ m-2 d-1, which is lower
than turfgrasses were subjected to in this study. Visible light intensities in this study were
also lower than normal daily integrals. The effect of UV-B could have been increased due
to lower levels of light protecting pigments in the lower visible light conditions
(Teramura et al., 1980). In an attempt to gain a consistent turfgrass response, UV-B levels
in our study were elevated above normal and this allowed for a clearer insight into the
turfgrass response. Another goal of our approach was to subject turfgrasses to equivalents
of extended exposures to UV-B irradiation which could simulate a season-long exposure.
The visual evaluation of color and differences between ratings agrees with
previous studies (Millington and King, 2010). Turfgrasses may have suffered from
probable chlorophyll degradation and this has been connected to a decrease in
photochemical efficiency (Ervin et al., 2004c). The loss of color is a principal factor for
evaluating turfgrass quality and while the thicker-leaved plants had significant color loss,
they still maintained acceptable color levels compared to the two creeping bentgrass
cultivars. This measurement can be construed as subjective and future potential use of
objective color measurements could offer more accurate and consistent visual
representations of the changes. This may be an important factor to consider regarding
tolerance of cool season grasses to UV light stress. If the predicted changes in UV light
levels occur over time, further research into leaf cuticle thickness and changes in resins
and waxes in the cuticular layers will be required. The use of synthetic, reflective type
pigments as plant coatings may be an alternative to breeding for increased UV-B
tolerance in turfgrasses.

96
Light Chamber UV-B (280-320nm) UV-A (320-400nm) PAR (400-700nm)
______________ kJ m-2 d-1 ______________ mol / m-2 / d-1 Control 0.0 0.1 18.6
UV- Chamber 24.0 0.4 18.4
Table 4.1: Lighting conditions in control and ultraviolet treated light environments for turfgrasses grown at The Ohio State University March – April 2011.

97
† Significant difference between grasses grown in ultraviolet and control conditions on this date (P=0.05). Figure 4.1: Relative growth rate for four cool season grasses grown in control and enhanced ultraviolet-B light conditions in repeated experiments

98
† Significant difference between grasses grown in ultraviolet and control conditions on this date (P=0.05).
Figure 4.2: Color ratings for four cool season grasses grown in control and enhanced ultraviolet-B light conditions in repeated experiments. (1-9 scale, 1 = brown/dead, 9 = dark green optimal color, 6 = acceptable).

99
† Bars signify least significant difference between treatments (P=0.05). ‡ Histograms with different letters for each grass signify differences at (P=0.05).
Figure 4.3: Number of tillers per plant for four cool season grasses grown in control and enhanced ultraviolet-B light conditions in experiment one.

100
† Bars signify least significant difference between treatments (P=0.05). ‡ Histograms with different letters for each grass signify differences at (P=0.05).
Figure 4.4: Number of tillers per plant for four cool season grasses grown in control and enhanced ultraviolet-B light conditions in experiment two.

101
REFERENCE
Ballare, C.L., A.L. Scopel, A.E. Stapleton, and M.J. Yanovsky. 1996. Solar ultraviolet-B radiation affects seedling emergence, DNA integrity, plant morphology, growth rate, and attractiveness to herbivore insects in Datura ferox. Plant Physiol. 112:161-170. Barnes, P.W., S.D. Flint, and M.M. Caldwell. 1990. Morphological responses of crop and weed species of different growth forms to ultraviolet-B radiation. Am. J. Bot. 77:1354-1360. Bell, G.E., T.K. Danneberger, and M.J. McMahon. 2000. Spectral irradiance available for turfgrass growth in sun and shade. Crop Sci. 40:189-195. Bornman, J.F., and T.C. Vogelmann. 1991. Effect of UV-B radiation on leaf optical properties measured with fiber optics. J. Exp. Bot. 42:547-554. Brandle, J.R, W.F. Campbell, W.B Sisson, and M.M. Caldwell. 1977. Net photosynthesis, electron transport capacity, and ultrastructure of Pisium sativum
L. exposed to ultraviolet-B radiation. Plant Physiol. 60:165-169. Caldwell, M.M. 1971. Solar ultraviolet radiation and the growth and development of higher plants. p. 131-177. In A.C. Giese (ed.) Photophysiology: Academic Press, NY. Cameron, R.J. 1970. Light intensity and the growth of Eucalyptus seedlings: II. The effect of cuticular waxes on light absorption in leaves of Eucalyptus species. Aus. J. Bot. 18:275-284.

102
Clark, J.B., and G.R. Lister. 1975. Photosynthetic action spectra of trees: II. The relationship of cuticle structure to the visible and ultraviolet spectral properties of needles from four coniferous species. Plant Physiol. 55:407-413. Cen, Y.-P., and J.F. Bornman. 1990. The response of bean plants to UV-B radiation
under different irradiances of background visible light. J. Exp. Bot.41:1489-1495. Dai, Q., V.P. Coronel, B.S. Vergara, P.W. Barnes, and A.T. Quintos. 1992. Ultraviolet-B radiation effects on growth and physiology of four rice cultivars. Crop Sci. 32:1269-1274. Day, T.A. 1993. Relating UV-B radiation screening effectiveness of foliage to absorbing- compound concentration and anatomical characteristics in a diverse group of plants. Oecologia 95:542-550. Deckmyn, G., and I. Impens. 1999. Seasonal responses of six Poaceae to differential levels of solar UV-B radiation. Environ. Exp. Bot. 41:177-184. Dong, Y. hu, D. Mitra, A. Kootstra, C. Lister, and J. Lancaster. 1995. Postharvest stimulation of skin color in royal gala apple. J. Am. Soc. Hortic. Sci. 120:95-100. Ervin, E.H., X. Zhang, and J.H. Fike. 2004a. Ultraviolet-B radiation damage on Kentucky bluegrass I: antioxidant and colorant effects. HortScience 39:1465-1470. Ervin, E.H., X. Zhang, and J.H. Fike. 2004c. Ultraviolet-B radiation damage on Kentucky bluegrass. III. Cultivar effects. HortScience 39:1475-1477. Fiscus, E.L., and F.L. Booker 1995. Is increased UV-B a threat to crop photosynthesis
and productivity? Photosynth. Res. 43:81-92. Gardner, F.P., R.B. Pearce, and R.L. Mitchell (ed.) 1985. Physiology of crop plants. Iowa State Univ. Press, Ames IA.

103
Gwynn-Jones, D., and U. Johanson. 1996. Growth and pigment production in two subarctic grass species under four different UV-B irradiation levels. Physiol. Planta. 97:701-707. Gwynn-Jones, D. 2001. Short-term impacts of enhanced UV-B radiation on photo- allocation and metabolism: a possible interpretation for time-dependent inhibition of growth. Plant Ecol. 154:67-73. Hagen, S.F., G.I.A. Borge, G.B. Bengtsson, W. Bilger, A. Berge, K. Haffner, and K.A. Solhaug. 2007. Phenolic contents and other health and sensory related properties of apple fruit (Malus domestica Borkh., cv. Aroma): Effect of postharvest UV-B irradiation. Postharvest Bio. Technol. 45:1-10. Hofmann, R.W. 2000. Population studies of ultraviolet-B radiation responses in white clover (Trifolium repens L.). Ph.D. Thesis, Massey University, N.Z. http://muir.massey.ac.nz/bitstream/handle/10179/2248/02_whole.pdf?sequence=1
(Verified 30 Jun. 2012). Kerr, J.B., and C.T. McElroy. 1993. Evidence for large upward trends of ultraviolet-B radiation linked to ozone depletion. Science 262:1032-1034. Krupa, S.V., and R.N. Kickert. 1989. The Greenhouse effect: Impacts of ultraviolet-B (UV-B) radiation, carbon dioxide (CO2), and Ozone (O3) on vegetation. Environ. Pollut. 61:263-393. Manney, G.L., M.L. Santee, M. Rex, N.J. Livesey, M.C. Pitts, P. Veefkind, E.R. Nash, I. Wohltmann, R. Lehmann, L. Froidevaux, L.R. Poole, M.R. Schoeberl, D.P. Haffner, J. Davies, V. Dorokhov, H. Gernandt, B. Johnson, R. Kivi, E. Kyrö, N. Larsen, P.F. Levelt, A. Makshtas, C.T. McElroy, H. Nakajima, M.C. Parrondo, D.W. Tarasick, P. von der Gathen, K.A. Walker, and N.S. Zinoviev. 2011. Unprecedented Arctic ozone loss in 2011. Nature 478:469-475. Milesi, C., S.W. Running, C.D. Elvidge, J.B. Dietz, and B.T. Tuttle. 2005. Mapping and modeling the biogeochemical cycling of turf grasses in the United States. Environ. Manage. 36:426-438.

104
Millington, K.R., and A.L. King. 2010. Measuring colour and photostability of small fleece wool samples. Anim. Prod. Sci. 50:589-592. Mohammed, A.R., E.W. Rounds, and L. Tarpley. 2007. Response of rice (Oryza sativa L.) tillering to sub-ambient levels of ultraviolet-B radiation. J. Agron. Crop Sci. 193:324-335. Pons, T.L., H. van Rijnberk, I. Scheurwater, and A. van der Werf. 1993. Importance of the gradient in photosynthetically active radiation in a vegetation stand for leaf nitrogen allocation in two monocotyledons. Oecologia 95:416-424. Rowland, F.S. 1990. Stratospheric ozone depletion by chlorofluorocarbons. Ambio 19:281-292. Rozema, J., R. Broekman, D. Lud, H.J. Huiskes, T. Moerdijk, N. de Bakker, B. Meijkamp, and A. van Beem. 2001. Consequences of depletion of stratospheric ozone for terrestrial antarctic ecosystems: the response of Deschampsia antarctica to enhanced UV-B radiation in a controlled environment. Plant Ecol. 154:103- 115. Smith, J.L., D.J. Burritt, and P. Bannister. 2000. Shoot dry weight, chlorophyll and UV- B-absorbing compounds as indicators of a plant's sensitivity to UV-B radiation. Annal. Bot. 86:1057-1063. SAS Version 9.2. SAS Copyright, 2002-2008 by SAS Institute Inc., Cary, NC, USA. Schmidt, R.E., and X. Zhang. 2001. Alleviation of photochemical activity decline of turfgrasses exposed to soil moisture stress or UV radiation. Int. Turfgrass Res. J. 9:340-346. Teramura, A.H., R. Hilton Biggs, and S. Kossuth. 1980. Effects of ultraviolet-B
irradiances on Soybean II. Interaction between ultraviolet-B and photosynthetically active radiation on net photosynthesis, dark respiration, and transpiration. Plant Physiol. 65:483-488.

105
Teramura, A.H., L.H. Ziska, and A.E. Sztein. 1991. Changes in growth and photosynthetic capacity of rice with increased UV-B radiation. Physiol. Planta. 83:373-380. Zhang, S., H. Jiang, S. Peng, H. Korpelainen, and C. Li. 2011. Sex-related differences in morphological, physiological, and ultrastructural responses of Populus cathayana to chilling. J. Exp. Bot. 62:675-686. Zhang, X., and E.H. Ervin. 2009. Physiological assessment of cool-season turfgrasses under ultraviolet-B stress. HortScience 44:1785-1789.

106
CHAPTER 5
PIGMENT CHANGES IN COOL SEASON TURFGRASSES IN RESPONSE TO ULTRAVIOLET-B LIGHT IRRADIANCE
ABSTRACT
Pigments involved in protection of photosynthetic apparatus in high light
conditions have potential to aid turfgrasses in periods of enhanced ultraviolet-B (UV-B)
radiation. These pigments have light absorbing capacities in the UV-B range (280-320
nm) and also may aid in energy dissipation as an initial protective mechanism from
photosystem damage. These pigments and their function in elevated UV-B conditions
have received little attention. This study aimed to characterize protective pigment
responses to enhanced UV-B conditions in cool-season turfgrass. Three cool season
grasses were used - tall fescue (Schedenorus phoenix (scop.) Holub), perennial ryegrass
(Lolium perenne L.) and creeping bentgrass (Agrostis stolonifera L.) cultivars ‘L93’ and
‘Penncross’. Turfgrasses were treated for pigment responses over a one week period and
subjected to 16 kJ m-2 d-1 of UV-B in growth chambers. The chambers were run on a day
/ night regime of 14 / 10 hours and plants were subjected to 18.5 mol m-2 d-1
photosynthetically active radiation (PAR) and 23 / 20 °C. Samples were collected at day

107
0, 1, 3, 5 and 7, and chlorophyll fluorescence was measured and chlorophyll
pigmentation, flavonoids, phenolics, anthocyanins and carotenoids content were
determined. Fluorescence measurements and chlorophyll levels decreased significantly
(P=0.05) in UV-B conditions compared to the control environment. All species possessed
significantly (P=0.05) higher quantities of total phenolics and flavonoids in the top of the
tissue canopy compared to roots or tissue at the soil surface. After day one all of these
compounds decreased but there were more significant decreases in UV-B conditions.
Anthocyanins were only characterized in creeping bentgrass ‘L93’ with Cyanidin – 3 – O
– glucoside making up 60% of the anthocyanins identified. The carotenoids, zeaxanthin
and β-carotene declined in total content in UV-B treatments in creeping bentgrass ‘L-93’
and ‘Penncross’ after seven days, but not in perennial ryegrass or tall fescue.
INTRODUCTION
Light drives many photosynthetic responses in plants. High irradiance can lead to
photoinhibition and chlorophyll degradation in plants. This may cause a decrease in
productivity (Curran et al., 1990; Anderson and Aro, 1994; Gitelson et al., 2006) and
leaves plants susceptible to poor recovery from wear and biotic stress. The irradiance that
plants receive on a daily basis varies in both quantity and spectral distribution. The light
distribution refers to the photosynthetically active radiation (PAR) that occurs in the 400-
700 nm wavelengths. It can be measured instantaneously in μmoles m-2 s-1 (Smith, 1994).
The direct effect of wavelength variations differs however from high levels of irradiance.
Light low in red light leads to alterations in photoreceptors signaling whole plant and

108
biochemical changes (Wherley et al., 2005). In Italian ryegrass (Lolium multiflorum
Lam.), there was reduced tillering while increasing length of leaf blades and reproductive
shoots due to increased red light (655 nm – 665 nm) (Casal et al., 1985). On the other end
of the spectrum, concentrations of blue light enhanced activity of cryptochrome leading
to chalcone synthase induction in Arabaidposis thaliana, (Lin et al., 1996). Outside of
PAR, other wavelengths such as ultraviolet light (UV) impact plant growth and
physiology. Ultraviolet light wavelengths are found between 100 nm and above 400 nm
(Rosario et al., 1979).
The effects of these wavelengths vary on plant physiology and growth.
Ultraviolet-A increases biomass by 22% in cucumber compared to UV-B wavelengths
but standard PAR additionally increased biomass by 14% above UV-A (Krizek et al.,
1997). The influence of UV-B in particular is considered to be negative due to
development of reactive oxygen species, inhibition of photosynthesis and effects on
genetic expression (Mackerness, 1999; Teramura and Sullivan, 1994). Lower
regeneration of ribulose bisphosphate, decreases in fluorescence, increased D1
polypeptide turnover within photosystem II have also been linked to UV-B exposure
(Greenburg et al., 1989; Strid et al., 1990; Ziska et al., 1993). Pea plants (Pisium sativum
L.) and cotton (Gossypium hirsutum L.) suffer decreases in chlorophyll content when
exposed to UV-B (Ziska et al., 1993; Gao et al., 2003). These losses occur within plants
particularly from UV-B radiation (Caldwell, 1971). The potential for further damage
could increase from 2010-2020. Peak irradiance of UV-B wavelengths are predicted to

109
occur in this time period prior to degradation of chlorine and bromine containing species
(Taalas et al., 2000).
The protective mechanisms that plants possess vary in their action and also in
location of response. Plant pigments such as anthocyanins, a class of UV-B absorbing
flavonoids are produced in Kentucky bluegrass (Poa pratensis L.), in response to
elevated UV-B conditions (Zhang et al., 2005). Anthocyanins are flavonoids which
absorb in the UV-B region (Gould, 2004). Polyacylated anthocyanins completely reduced
DNA lesions in blue morning glory (Ipomoea tricolor cv. ‘Heavenly Blue’), (Mori et al.,
2005). The anthocyanins themselves can vary in their absorption spectrum due to
changes in methylation or acylation (Gitz et al., 1998; Do and Cormier, 1991). These
changes are affected predominantly by pH alterations within the plant which are light
driven (Yin et al., 1990). The actual location of the pigments in the adaxial epidermis of
leaf surfaces is also considered crucial for UV protection in coleus (Solenostemon L.)
(Burger and Edwards, 1996). There are also other pigments which are used by plants with
the aim of reducing light damage to photosynthetic apparatus. The light absorption
characteristics are different from anthocyanins but offer light protection.
Carotenoids are another set of compounds which have another role in light
protection in high intensity light. A pool of the 40 carbon compounds known as
xantophylls alters its composition in creeping bentgrass (Agrostis stolonifera L.) in
response to high and low light conditions (Bell and Danneberger, 1999; McElroy et al.,
2006). Zeaxanthin is found in larger proportion to violaxanthin in high light conditions
but reverts to equal or lower levels in low light conditions. Lutein has also been shown to

110
be the carotenoid of greatest concentration regardless of light conditions (McElroy et al.,
2006). In Kentucky bluegrass treated with UV-B, total carotenoid content decreased over
10 days with recovery occurring after 15 days of treatment completion. There was also a
positive correlation (>0.7) between these compounds and turfgrass quality in enhanced
UV-B conditions (Zhang et al., 2005). The ability of carotenoids to quench singlet
oxygen and other excited free radicals after UV-B treatment (Mathews-Roth, 1996), may
be responsible for reduction in damage to LHC in plants and limit the loss of plant
quality. The damage to photosynthetic apparatus is used to infer possible reductions in
photosynthetic efficiency and this damage has also a positive correlation to total
carotenoid content (Zhang et al., 2005).
The repair and defense mechanisms can also include more than pigments.
Enzymes which act as catalysts for energy-quenching reactions such as super oxide
dismutase build up in response to UV-B exposure in Kentucky bluegrass (Zhang et al.,
2005). Glucose-6-phosphate dehydrogenase activity and higher levels of proline occur in
creeping bentgrass, Kentucky bluegrass, perennial ryegrass and tall fescue treated with
UV-B. A possible repair mechanism allocating resources from glycolysis to the pentose
phosphate pathway leads to a buildup in enzymatic activity in turfgrasses exposed to UV-
B light (Sarkar et al., 2011). These repair systems stabilize the D1 protein within PSII and
offer a method of scavenging excess levels of energy. The systems also help to split up
cyclobutane dimers in DNA between two adjacent pyrimidines which are probably
formed during UV exposure. Photolyase uses excision repair or recombination repair to
complete restoration of DNA damaged from UV-B exposure (Sancar and Sancar, 1988).

111
The damage to photosynthetic apparatus linked to high energy light within PAR is
known as photoinhibition and UV-B light is known to cause similar problems for plants
(Friso et al., 1994). The enhanced rate of degradation of both the D1 and D2 proteins
inhibits the rate of photosynthesis. The damage to the D2 protein is increased in high UV-
B conditions which are not considered normal. In general the D1 protein degrades at a
more rapid rate than D2 with the exception where UV-B treatments are imposed in
combination with visible light (Jansen et al., 1993). This is due to the UV absorption by
the manganese cluster which lies in the oxygen evolving complex. This may act as a
component magnifying the protein degradation (Babu et al., 1999; Tyystjärvi, 2008). The
extent of the damage may initially be expressed in PSII and then damage will begin to
occur in the more robust PSI. The damage in PSII may be construed as an enhanced rate
of protein degradation which can be repaired. This damage, however, then leads to
damage of PSI which requires complete breakdown and reconstruction and long term
damage to PSI (Takahashi and Murata, 2008). The degradation of PSII leads to a buildup
of reactive oxygen species and peroxyl radicals with increased lipid peroxidation (Hideg
and Vass, 1996; He and Häder, 2002).
In measuring damage to photosynthetic apparatus within plants, damage to
photosystem II is most prevalent in relation to UV-B damage (Vass et al., 1996). This
damage can be measured indirectly via fluorescence of reaction centers of chlorophyll A
(Maxwell and Johnson, 2000). The degradation of the centers leads to a decrease in
fluorescence. Chlorophyll content within tissue, which also degrades due to
photoinhibition, can also be measured (Strid and Porra, 1992). This degradation in UV-B

112
conditions is at a higher rate than normal PAR and leads to a decrease in productivity
from plants and can occur on a time scale of minutes to hours (Aro et al., 1993).
Turfgrass photoinhibition occurs under elevated UV-B light conditions with Kentucky
bluegrass suffering significant decreases in the ratio of variable fluorescence to maximal
fluorescence (Fv/Fm) (Zhang et al., 2005). This results in a decrease in photosynthetic
activity due to increased levels of fluorescence within the photosynthetic apparatus.
Under elevated UV-B light conditions it has been shown that photoinhibition also affects
the PSII apparatus in tall fescue and perennial ryegrass but not as badly as Kentucky
bluegrass (Zhang and Ervin, 2009).
There is a need to elucidate the variations that occur within the plant canopy while
also characterizing pigment changes across a range of cool season turfgrasses exposed to
UV-B irradiation. The influence of UV-B may create different responses within cool
season turfgrasses and how they protect their photosynthetic apparatus in an attempt to
retain productivity. The objective of this study was to characterize pigmentation
alteration in UV-B conditions and investigate if these changes offer potential for
protection from UV-B damage to the photosynthetic apparatus in cool season turfgrasses
via energy dissipation and direct absorption of excess light. We hypothesized that
turfgrasses would produce higher quantities of protective pigments in response to the
UV-B stress. These higher pigment quantities would then allow turfgrasses to retain their
photosynthetic capacity during the period of stress. This will aid in reducing summer
stress and allow turfgrasses to maintain photosynthetic activity close to normal levels.

113
MATERIALS AND METHODS
Sample preparation
The studies were conducted in October, 2010 and March, 2011 in Columbus,
Ohio. Turfgrasses were initially seeded into a mason sand rootzone >97% sand (Hummel
& Co, Trumansburg, NY). The rootzone met USGA requirements and had a pH of 7.4
with a clay silt fraction of less than 5%. The seeding was timed to match germination
speeds between the grasses. Creeping bentgrasses were seeded first, and 4 days later
perennial ryegrass and tall fescue were seeded into 3.8 cm diameter ‘conetainers’
(Steuwe & Sons, Tangent, OR). Moisture was maintained by placing turfgrasses on a 30
second irrigation cycle every 10 minutes in a mist house. Turfgrasses were allowed to
establish for six weeks in the mist house.
Grasses were treated with Peters 20-20-20 (Scotts-Miracle Gro, Marysville, OH)
at a rate of 11.8 kg ha-1 every two weeks during this period. After germination, plants
were removed and placed in standard greenhouse conditions for a further four weeks to
allow for maturation. Turfgrasses were maintained at a height of ~10cm with clippings
removed 3 times weekly. Fertility treatments were continued and imidacloprid N-[1-[(6-
chloro-3-pyridyl) methyl]-4,5-dihydroimidazol-2-yl]nitramide [Bayer Environmental
Science, Research Triangle PK, NC] was applied to prevent insect damage. Mefenoxam
(R,S)-2-[(2,6-dimethylphenyl)-methoxyacetylamino]-propionic acid methyl ester
[Syngenta AG, Greensboro, NC] and thiophanate-methyl (dimethyl 4,4’-o-
phenylenebis[3-thioallophanate]) [Cleary Chemicals, Dayton, NJ] were applied for
preventative disease control. Water was applied to prevent wilt, with automated heads

114
using an ESP modular irrigation system (Rain Bird, Azusa, CA) once daily. Ten weeks
after initial turfgrass emergence, plants were placed in Conviron E15 growth chambers
(Controlled Environment Ltd., Winnipeg, Canada). To equalize plants they were placed
in dark growth chambers at 20 °C. This happened for 24 hours for all plants in both
experiments.
Ultraviolet-B treatment commenced using UV-B lights having peak emission at
313 nm, QUV UV-B 313 nm, (Q-Lab Corp., Westlake, OH). Treatment exposure was
calculated using time of exposure multiplied by measured irradiance energy per second.
Wavelengths within UV-B and UV-A were measured using an IL1350 photometer
(International Light, Newburyport, MA) with specific sensors for each portion of the UV
light spectrum. Biologically effective radiation was calculated from spectrum provided
by light producers. Ultraviolet radiation is reported in kJ m-2 d-1. Photosynthetically
active radiation was instantaneously measured every 15 minutes during the study and
reported as mol m-2 d-1 using cosine corrected light sensors (Spectrum Technologies,
Plainfield, Il). Light treatments of 14 hours daytime and night time periods of 10 hours
were controlled by the chamber. The lamps were pre-lit for 48 hours to prevent any
fluctuation in energy output during trials. Photosynthetic active radiation (PAR) which
lies in the range of 400 – 700 nm was provided using fluorescent lamps. The same lamps
were used in the control and UV chambers, 85 w s-1 high output F12T12-CW.HO
fluorescent tubes (General Electric, Fairfield, CT). The turfgrasses were irrigated twice
daily to prevent wilt in the chambers. Each chamber was fixed at 20°C during light

115
periods and 17 °C in darkness. In each chamber, plants were placed 0.4 m below the
lighting in an even spacing.
Chlorophyll content and fluorescence
The experiment was set up in a randomized complete block design with four
blocks and three replications per block for each grass. Analysis was carried out at Day0
(D0), Day1 (D1), Day3 (D3), Day5 (D5) and Day7 (D7) for all data. After each treatment
turfgrasses were placed in darkness for one hour. Fluorescence measurements were then
taken with 12 total replications for each grass per block to reduce variability. The Fv/Fm
values were obtained using a handheld chlorophyll fluorometer OS-30p (Opti-Sciences,
Hudson, NH). Leaf tissue from the top 5 cm was used and measured in-vivo with initial
660 nm pulses from an integral probe fluoresced to a photodiode with a 700 – 750 nm
bandpass filter. Values were averaged for each block and analyzed separately due to the
fact that different turfgrass plants were used for each sample date.
Three turfgrass tissue samples from the top of each grass of 40 mm-2 were taken
and placed into 5 mL of N,N-dimethylformamide (Sigma-Aldrich, St. Louis, MO) for 48
hours in the dark at 4°C. The extract was then placed in 4 mL cuvettes and absorbances
were measured at 664, 647, 625, 603 nm. This was carried out using a UV1800
spectrophotometer (Shimadzu Corp., Colombia, MD). Total chlorophyll, chlorophyll A
and chlorophyll B were calculated using pre-determined formulae (Moran, 1982).
Chlorophyll content was reported in µg m-2. The rest of the tissue material was separated
into the top 5 cm (T5) of tissue, all tissue between the root / shoot interface and the top

116
cm (T) and roots (R) (Appendix C), and frozen in liquid nitrogen at -210 °C. All samples
were then freeze dried using a Virtis ultra 35 LE lyophilizer (The Virtis Co., Gardiner,
NY) under a vacuum of 125 mTorr. This ran on an air temperature of -40°C to 10°C over
a 20 hour period and maintained at 10°C for a further four hours while tray temperatures
were maintained at -20°C. Samples were then stored at -20°C while analysis was
completed.
Total flavonoids
Soluble flavonoids were measured using the method described by Pourmorad et
al., (2006). Samples of 0.035 g were added to 10 mL of methanol and vortexed in 15 mL
conical centrifuge tubes and allowed to stand for 10 minutes at room temperature. The
samples were centrifuged at 3,400 rpm for 5 minutes. The material stood with an
occasional vortex for at least 10 minutes before centrifuging. A standard curve was
prepared using quercitin solutions at concentrations of 0, 10, 20, 40 and 50 μg mL in
methanol. The extract was then placed into a tube with 1.5 mL CH3OH, 0.1 mL of 10%
AlCl3, 0.1 mL potassium acetate and 2.8 mL distilled H2O. The material was vortexed
and incubated at room temperature for 30 minutes. The supernatant absorbance was
measured at 415 nm with a UV1800U spectrophotometer. Linear regression analysis was
used to determine concentrations of flavonoids in each sample. This was carried out for
R, T and T5 of all samples to determine concentrations of pigments based on location.
This test was performed for each day in both experiments.

117
Total phenolics
Total phenolics were measured in accordance McDonald et al., (2001). Gallic acid
standard curves were made using concentrations of 0, 5, 10, 20, 40 and 80 mg L in
MeOH:H2O, 50:50 v/v. Plant tissue (0.035 g) was placed in 15 mL of methanol and
vortexed and let sit for 10 minutes. Tissue sample was then centrifuged for 3 minutes at
3600 rpm. Supernatant (0.5 mL) was collected and placed in a 15 mL tube with 5 mL of
10% folin ciocalteu reagent and 4 mL of 1M Na2CO3 and let sit at room temperature for
15 minutes. Total phenol concentration was then determined at absorbance of 765 nm
using a UV1800U spectrophotometer. Linear regression analysis was then used to
estimate phenol concentrations in each sample on a µg g-1 basis. This was carried out for
R, T and T5 of all samples to determine concentrations of pigments based on location.
This was carried out for each day in both experiments.
Carotenoid extraction and analysis
Carotenoid extraction was carried out with slight modifications from a method
described by Ferruzzi et al., (1998). All experiments were carried out under caution of
low light to prevent carotenoid degradation. Tissue sample (0.5 g) was placed in 10 mL
of MeOH in a 15 mL centrifuge tube and vortexed for 30 seconds. Samples were then
homogenized using a polytron PT10 (Brinkman Institute, Westbury, NY), and then
centrifuged for 7 minutes with an Accuspin 3R (Fisher Scientific, Pittsburgh, PA).
Supernatant was removed and the pellet was extracted 3 times in 10 mL acetone/hexane
1:1 (v/v). All extracts were pooled in a 50 mL tube, a small volume of NaCl crystals were

118
added, and the extract was brought up to 50 mL with water and vortexed. The upper
hexane layer was brought to 10 mL in a volumetric flask with hexane and transferred to a
25 mL conical flask.
Samples were run through glass wool and Na2SO4 crystals and then saponified to
remove the phytol tail from the chlorophyll pigments to allow them to partition into the
aqueous phase for removal. Samples (10 mL) were placed in 50 mL conical flasks with
10% KOH (Sigma Aldrich), in a 1:2 v/v with the extract. Flasks were then placed on a
benchtop shaker for 2 hours covered in foil. The saponified mixture was then transferred
to a 125 mL separatory funnel. The green chlorophyll separated into the lower phase
which was removed. The remaining upper phase was washed with 40% Na2SO4 three
times. The remaining sample was then dried under nitrogen at less than 2 PSI using a
reactive-vap 18780 (Pierce, Rockford, IL). Samples were then resuspended in 2 mL of
methyl tert-butyl ether: MeOH 1:1 (v/v) and passed through a 0.22 µm nylon syringe
filter (Fisher Scientific, Pittsburgh PA) into 2 mL amber HPLC vials (Waters, Milford,
MA).
Sample volumes of 20 µL were injected into an HP1050 series HPLC, with a
diode array detector (DAD), were used to quantitate the carotenoids. Samples (20 µL)
were injected onto a YMC C30 carotenoid column 4.6 x 250 mm, 3 µm particle size,
(Waters, Milford, MA). A gradient system was set up and a tertiary solvent system was
used. The gradient was used based on a modified method of Moros et al., (2002), with a
flow rate of mL/min. Solvent A consisted of methanol/MTBE/water (81:15:4), solvent B
consisted of methanol/MTBE (9:91) and solvent C consisted of MTBE alone. The initial

119
conditions were 100% A, with a linear increase to 45% A and 55% B over 21 min,
changing to 100% C and holding for 2 minutes and returning to 100% A and holding for
5 minutes. Data was collected using HP Chemstation 3D and 453 nm wavelength was
used for quantitation of the compounds. External calibration curves of zeaxanthin
(CaroteNature, Lupsingen, Switzerland), lutein, β-carotene (Sigma Aldrich),
antheraxanthin and violaxanthin (Chromadex, Irving, CA) were used to quantitate the
respective carotenoids. Turfgrass samples were used from D0 and D7 of the trials to look
at overall changes in concentrations of carotenoids. Samples were taken from the T5
portion of turfgrasses
Anthocyanins
Anthocyanins were extracted using a method modified from Giusti et al., (1999).
Freeze dried tissue (0.5g) was crushed into a fine powder using a mortar and pestle while
suspended in liquid nitrogen. Samples were then placed in 10 mL of 100% acetone in 15
mL tubes, vortexed and left to sit for 5 minutes. Samples were then centrifuged at 3600
rpm for 5 minutes. Supernatant was collected and samples were re-extracted with 0.1%
HCL acidified 70:30% acetone:H2O (V:V). Samples were re-extracted two more times
until supernatant was clear. Extract was combined in a 50 mL centrifuge tube and
brought to 30 mL. Supernatant and tissue were then combined and passed through a No.1
filter (Whatman, Kent, United Kingdom) to maximize extract collection in a Büchner
funnel. This was carried out under vacuum using a Rocker 300 pump (Rocker,
Kaohsiung, Taiwan), and samples were collected in a conical flask. To separate water

120
soluble from lipid soluble compounds, 2:1 volume chloroform was added and samples
were sat overnight in dark cool conditions in 125 ml separating funnels. Chloroform and
acetone (now mixed together) were removed and the top aqueous portion was collected
and retained. To purify the anthocyanins, C-18 cartridges (Biotage, Charlotte, NC) were
used. To prepare the cartridges 100% MeOH x2 volumes of cartridge were flushed
through under an Alltech 210351 12-port vacuum manifold (Alltech, Deerfield, IL) and
washed with 3 volumes of acidified water (0.1% v/v HCL). Samples were then loaded
into the cartridge and anthocyanins, sugars and other phenolics were adsorbed onto the C-
18 column under vacuum. To remove water soluble compounds (0.01%) acidified water
was then passed through the cartridge followed by ethyl acetate to elute non-anthocyanin
phenolics. Anthocyanins were finally recovered using v/v 0.01% HCL acidified MeOH in
concentrated format. In all stages, the pH of extract was sustained at 2 or lower to retain
stability. Samples were collected in 2.5 mL microfuge tubes and spun at 5000 rpm x 5
minutes to remove any debris. Samples were then concentrated using a Büchi R114
rotoevaporator and re suspended in 1 mL distilled deionized water which was acidified
with 0.01% HCL.
Samples were passed through a .22 µ filter using 3 mL luer lock syringes and
injected into 2 mL amber LCMS vials with 300 µL inserts with springs. Samples were
then injected into a LCMS apparatus set up as follows. A CTO-20A (Shimadzu Scientific
Inst., Columbia, MD) oven was used to maintain temperatures at 35°C, flow of 0.8 mL
min was controlled with LC-20AD – LC pumps, while 50 µL sample injection was
managed with a SIL-20AC autosampler. Samples were injected onto a Varian C18-A,

121
150 x 4.6 mm i.d. column (Varian Inc., Lake Forest CA). Samples were eluted through a
SPD-M20A PDA (Shimadzu) with wavelengths 280, 320 and 520 nm simultaneously
selected. Spectral analysis of wavelengths between 250 and 700 nm were recorded for all
peaks. Solvents for mobile phase were first filtered through .22 µm filters. A binary flow
mobile phase consisting of (A) 4.5% formic acid:H2O v/v and (B) acetonitrile with a flow
rate of 0.8 mL min was used. Anthocyanins were separated using a solvent gradient from
5 % B to 15 % in 5 minutes followed by an increase in B to 30 % at 20 minutes. From 20
minutes to 23 minutes solvent B increased to 100 % and was maintained there until 25
minutes. Concentration of B then decreased to 5 % at 28 minutes and sample injection
completed after 35 minutes. Samples were then injected into an electron spray mass
spectrometer and fragmentation analysis carried out. Due to limited samples only
qualitative data was obtained. Known anthocyanin pigments (Chandra et al., 2001) from
(Aronia melanocarpa) were injected under the same conditions and used as a reference
material (Appendix D). Compound mass, elution order, spectral absorbance and retention
time were used to characterize anthocyanins found in turfgrass samples.
Data analysis
Data analysis for chlorophyll, fluorescence, total phenolics and flavonoids was
carried out using the general linear model from the SAS program (SAS Inst., 2008).
Analysis was carried on block averages for comparison of data from each light
environment collected on each specific date. Comparisons between light environments
were carried out for each variable on each date because different samples were used for

122
each date. Carotenoid content for each light environment was calculated as a difference
between D0 and D7 to compare overall changes during treatments. The lack of samples
due to growth chamber size constrictions for anthocyanin analysis meant that only
characterization of these pigments could occur at this time.
RESULTS
Light intensities for the two chambers were very similar within the PAR region of
light (400-700 nm). The main difference between chambers was measured in the UV-B
region with turfgrasses subjected to 16.9 KJ m-2 d-1 of UV-Bbe light in the UV chamber
compared to the 0 in the control chamber (Table 5.1).
Fluorescence measurements in all grasses followed the same pattern in both
experiments and data was pooled after testing for differences between experiments. In
both chambers there was a relative decrease in photosynthetic activity from D0 in all
grasses. However, significantly lower fluorescence measurements occurred in chambers
that had elevated UV-B conditions (P=0.05) after D3 and this continued through D7. All
grasses suffered greatest decline in Fv / Fm up to D5. Perennial ryegrass and “Penncross’
creeping bentgrasses had declines of Fv / Fm greater than 10% (Figure 5.1). No grass
exhibited complete loss of photosynthetic activity.
Chlorophyll pigmentation exhibited a similar pattern in all grasses in both
experiments. Treatment of UV-B resulted in significant declines in total chlorophyll
content (P=0.05). The decreases were noted after D1 in both perennial ryegrass and tall
fescue. This separation between control and UV treated grasses did not occur until D3 in

123
the creeping bentgrass cultivars. There were also declines in the control chamber in all
grasses; however, the decline in chlorophyll content in the control chambers was faster in
the bentgrass cultivars (Figure 5.2). Overall, chlorophyll levels were higher on an area
basis in perennial ryegrass and tall fescue compared to the creeping bentgrass cultivars.
The change in chlorophyll A and B also followed the same pattern. Significant declines
(P=0.05) occurred in both of these chlorophyll pigments in UV-B conditions (Appendix
E).
The distribution of both phenolics and flavonoids were similar across all grasses
regardless of light environment. This occurred in both experiments. Turfgrasses exhibited
the ability to accumulate significantly higher (P=0.05) quantities of both phenolics and
flavonoids in the T5 portion of the tissue sample. There was almost a 50% difference in
total phenolics between the T5 and T in creeping bentgrass ‘L-93’ in control conditions at
the end of the trial. On that same date there was an 85% difference between T5 and R
tissues in ‘L-93’ (Figure 5.3). There was an initial increase in phenolic content in both
portions of tissue on D1 and this was then followed by a decline. Flavonoids also
exhibited similar significant separation with T5 tissue containing significantly greater
concentrations. In perennial ryegrass, concentrations of total flavonoids varied between
T5 and T tissue was as much as 50% after D7 (Figure 5.4). There was for the most part a
spike in response after day 1 but this was followed by a decline in concentration in both
parts of the plant tissue.
The overall concentration of flavonoids increased on D1 in both light
environments. This response was then followed by a decline in all grasses until D5 when

124
quantities then leveled off. There was significant decreases in all grasses in UV-B
irradiated conditions at D7. There was on average a 27.5% decrease in all grasses at D7
(Figure 5.5). A similar decrease was observed in both light environments in total phenolic
concentrations (Data not shown). The difference at D7 between control and UV-B
chambers was only significantly different in the two creeping bentgrass cultivars (Data
not shown).
Anthocyanin responses to UV-B exposure were very limited. Extracts of tissue
from D1, D3 and D5 yielded no data when passed through HPLC conditions at 520 – 530
nm. This occurred in all grasses for both sets of light environments and in both
experiments. In D7 tissue from creeping bentgrass ‘L-93’, HPLC analysis indicated that
the presence of anthocyanins sufficiently strong for detection at 520 nm (Figure 5.6) and
320 nm. Electrospray ionization of compounds resulted in masses for four main pigments
in the ‘L-93’ tissue. The elution order and approximate retention time was similar to
Aronia samples which were also injected and checked in previous literature (Chandra et
al., 2001). This information was then used to characterizing the anthocyanins found in
creeping bentgrass ‘L-93’ (Table 5.2). First eluted compound at 8.4 minutes was cyanidin
3 – O – glucoside which was followed by peonidin 3 – O – galactoside at 9.95 minutes.
At 11.05 minutes cyanidin 3 – O – malonyl glycoside was detected and this was followed
by cyanidin 3 – O – succinyl glucoside at 13.25 minutes (Figure 5.6). No other grasses
displayed detectable anthocyanin increases at D7 in either light environment.
Using internal spikes to quantify potential pigment degradation resulted in noted
loss of antheraxanthin, lutein and violaxanthin due to saponification. Zeaxanthin and β-

125
carotene were identified based on absorbance, peak shape, elution order and retention
time linked to standards (Figure 5.7). Zeaxanthin eluted at 11.1 minutes while β-carotene
eluted at 20.1 minutes. Peaks that did fit any of standards available eluted at 16, 19 and
24 minutes (Figure 5.7). Degradation of β-carotene was ~10% through saponification and
zeaxanthin had ~15% degradation. Saponification was required to separate chlorophyll
pigments from carotenoids prior to HPLC analysis. There was no difference in carotenoid
content between light environments at D0 or D7. There was however greater degradation
in the UV environment from D0 to D7 with significant decreases in β-carotene content in
both of the bentgrasses (Appendix F) in the UV chamber. Perennial ryegrass and tall
fescue (Appendix G) did not have such a significant loss of β-carotene in UV conditions
(Figure 5.8a). Creeping bentgrass ‘Penncross’ also had significant β-carotene loss
between samples from D0 to D7 in control light conditions (Data not shown). Creeping
bentgrass ‘L-93’ had significant losses (P=0.05) of zeaxanthin in both UV and control
chambers. No other grass exhibited this loss of zeaxanthin and this did not happen in the
control chamber (Figure 5.8b). There was no difference in zeaxanthin content between
UV and control treated turfgrasses at D7.
DISCUSSION
The decrease in photosynthetic activity in our study occurred regardless of grass
and agreed with previous data in cool season turfgrass (Zhang et al., 2005; Sarkar et al.,
2011). This loss of activity in our study could be attributed to the decrease in phenolics or
also loss of enzymes used for repair of the photosynthetic apparatus. The decrease in

126
carotenoid pigments may also have played a role in the decrease in photosynthetic
activity in this study. The loss after D1 of UV the absorbing compounds would indicate
that the photosynthetic apparatus would have been suffering damage at the same time.
This was born out in the results show using the flouresence measurements.
The loss of chlorophyll pigmentation in this study was despite taller turfgrasses
having a dense canopy. The loss of chlorophyll coincided with the decrease in
fluorescence also and this would then lead to a loss of biomass productivity as has been
noted in wheat (Triticum aestivum L.) which had a decline of 18% (Correia et al., 1999)
in response to elevated UV-B levels. Degradation of chlorophyll is a common response to
UV-B radiation in plants. Kentucky bluegrass has exhibited this response before as have
sunflower cotyledons and other plants (Zhang et al., 2005; Costa et al., 2002; Tevini et
al;, 1981). The energy associated with the photons in the UV-B wavelengths (3.94 – 4.43
electronvolts) is considered strong enough to destroy many chemical bonds and it may
have contributed directly to the chlorophyll degredation (Kovács and Keresztes, 2002).
The variation in distribution of phenolics and flavonoids in our study could be
attributed to two causes. Turfgrass plants in such a dense canopy may have led to self-
shading thus the elevated levels in the upper canopy of these UV absorbing compounds is
a response to the light conditions in the grasses tested. This argument is further
strengthened by the lack of response and much lower levels of each set of compounds in
the root tissue (Buer and Muday, 2004). Similar distributions of flavonols have been
noted (Cortell and Kennedy, 2006) and also in phenolic compounds in roots and leaf
tissue of cow lily (Nuphar advena) and large leaf pondweed (Potamogeton amplifolius

127
L.) (Cronin and Lodge, 2003). The second cause could relate to age of the leaf tissue
which can be more susceptible to damage from UV-B exposure when it is younger (Lois,
1994). We feel this distribution of compounds in our study is due to a light response as
material was clipped 3 days before trial initiation to reduce the influence of leaf age.
The actual quantities of phenolics and flavonoids had an initial increase from D0
to D1 in both light environments in our study. This type of response is considered a plant
protective response in an attempt by the plant to reduce the impact of the UV-B
wavelengths on the photosynthetic apparatus. Phenolics have antioxidant capacity and
offer a method to reduce oxidized radicals and reduce damage to the photosynthetic
apparatus (Duval et al., 1999). Mutant Arabidopsis plants unable to accumulate
flavonoids are highly susceptible to UV damage (Landry et al., 1995). There was then a
decline in flavonoid and phenolic levels in all grasses, with greater declines for both in
UV-B. This loss may be related to an inability of turfgrass plants to allocate enough
energy to retain initial response levels due to lower levels of visible light in the chambers
(Teramura et al., 1980). The UV-B exposure may have overwhelemed the plants
capability to repair itself and recover from free radical damage in this study.
The potential benefit of anthocyanins was only seen after seven days of exposure
only in the ‘L-93’ cultivar of the creeping bentgrass in our study. This delay in
production was after a decline in photosynthetic activity and so the benefit was both
delayed and limited. Increased anthocyanins have been found in Kentucky bluegrass in
response to UV-B exposure (Zhang et al., 2005). The UV-B absorption capabilities of
anthocyanins are argued as a benefit and that plants possessing this response are

128
acclimated to UV-B stress (Gould, 2004). There is some argument to the photoprotective
capabilities of these compounds as only marginally higher photosynthetic efficiency was
found in red Kermes oak (Quercus coccifera L.) leaves compared to greener ones
(Karageorgou and Manetas, 2006). Also limited or no photoprotective value was found in
myrtle plants with anthocyanins in the epidermis (Woodall and Stewart, 1998).
In our study the creeping bentgrass cultivar ‘L-93’ only exhibited such a response
while ‘Penncross’ did not. The variety differences in anthocyanin response noted in our
study have also been noted before though not in turfgrass. In maize (Zea mays L.), of
seven varieties tested for UV-B response, only two varieties responded with anthocyanin
production while one other showed a strong anthocyanin development response to far red
light (Beggs and Wellmann, 1985). The other reason that both the perennial ryegrass and
tall fescue may have had limited response in this study was that epidermal thickness plays
a large role in determining the transmission of light into the epidermis. Tall fescue has
large variations in its thickness and it can significantly alter its leaf thickness from 0.31
cm to 0.36 cm in response to drought (Fu and Huang, 2004). This may require much
larger consideration for UV-B screening effectiveness (Day, 1993) combined with
pigment concentrations.
The concentration of β-carotene decreased in both light environmentsin our study
but the decrease was greatest in UV conditions and also only in the bentgrass species.
The loss in β-carotene content has also been linked to a loss of photosynthetic activity in
Dunlaliella cells exposed to UV-B (White and Jahnke, 2002). This loss of an important
pigment may relate to the loss of quality and also the greater loss of photosynthetic

129
activity in the UV chamber-treated bentgrasses. Zeaxanthin decreases have occurred over
extended periods of UV- B exposure in maize (Carletti et al., 2003). There have been
initial increases in content also but possible changes in energy allocation may reduce
their levels after prolonged exposure. This degradation of both carotenoids reduces the
turfgrass capabilities to moderate damage from reactive oxygen species. The impact of
carotenoids may be crucial in high UV conditions due to their capabilities of energy
dissipation and is particularly important for the thinner leaf bentgrasses. Further work
completing the xantophyll cycle response and also quantifying and characterizing these
compounds is required to gain a fuller understanding of their value.
The loss of xantophylls due to saponification has been observed in prior studies
(Kimura et al., 1990). It is the first time that the problem has been reported in turfgrass
and may reduce the capability to cleanly separate carotenoid and chlorophyllous
pigments when using HPLC analysis. The alkalinity of the methanolic KOH leads to
hydrolysis of the compounds; however, it was thought that the lower percentage of KOH
would reduce the problem (de Sá and Rodriguez-Amaya, 2003).
CONCLUSION The loss of photosynthetic activity in turfgrasses observed in this experiment due
to exposure to UV-B was expected. It was interesting to note a decrease in quantity of
UV-B absorbing pigments over extended exposure. However, the combination of high
UV and low visible light levels may have played a role in this. The loss of carotenoids in
the creeping bentgrasses is of interest as is the production of anthocyanins in ‘L-93’

130
creeping bentgrass due to their UV absorbing abilities. The reduction in carotenoids
would lead to an increase in highly energized products thus requiring some form of
antioxidant response and also a compound that absorbs the excess light. Immediate
breeding responses for a crop that covering potentially 163,800 km-2 (Milesi et al., 2005),
of cultivated land are not possible. In an attempt to address the problem rapidly, the use
of synthetic coatings may offer a practical response and tool for turfgrass managers. This
first characterization and extraction of turfgrass anthocyanins also offers some potential
for further research in turfgrass stress physiology. Further work on carotenoid
characterization and chromatogram publication is also required due to their importance in
light stress and new herbicide research.

131
Light _______________ Chambers _______________ Control Ultraviolet-B
PAR (400-700 nm) 26.2 mol m-2 d-1 25.9 mol m-2 d-1
UV-A (400 – 320 nm) 1.0 KJ m-2 d-1 0.8 KJ m-2 d-1 UV-B be† ( 320 – 280 nm) 0.0 KJ m-2 d-1 16.9 KJ m-2 d-1
†Biologically effective radiation within UV-B wavelengths based on Caldwell, (1971).
Table 5.1: Light intensity and light distribution for Conviron E15 growth chambers illuminated using high output F12T12-CW.HO fluorescent tubes and QUV UV-B 313 nm UV-B lights.

132
Compound Tr (min) † [M] +m / z‡ Fragments m / z§ UV – vis nm¶ Area % (520 nm) # Identity 1 8.42 449 287 280, 516 60.5 Cyanidin 3 – O – glucoside 2 9.95 463 301 324, 516 4.3 Peonidin 3 – O – galactoside 3 11.05 535 287 281, 519 27.5 Cyanidin 3 – O – malonyl glycoside 4 13.25 549 287 329, 516 7.4 Cyanidin 3 – O – succinyl glucoside
† Retention time. ‡ Mass of whole compound. § Breakdown of compound sizes after passage through mass spectrometer. ¶ Peak absorption of compounds in PAR and UV regions. # Percentage of total compounds identified. Table 5.2: Anthocyanin pigmentation detected in creeping bentgrass ‘L-93’ after 7 days of enhanced UV-B treatments.

133
*† Significantly different at (P=0.05)
Figure 5.1: Fv / Fm values for Perennial Ryegrass and Creeping Bentgrass ‘Penncross’ treated with UV-B light (280 – 315 nm) and PAR light (400 – 700 nm) conditions over seven days.

134
*† Significantly different at (P=0.05)
Figure 5.2: Total chlorophyll content for Tall Fescue and Creeping Bentgrass ‘L-93’ treated with UV-B light (280 – 315 nm) and PAR light (400 – 700 nm) conditions over seven days.

135
*† Significantly different at (P=0.05) between Roots (r), Top (T) and Top 5 cm (T5) tissue.
Figure 5.3: Distribution of phenolics in creeping bentgrass ‘L-93’ after treatment with both UV-B light (280 – 315 nm) and PAR light (400 – 700 nm). Data was pooled as all grasses had the same response.

136
*† Significantly different at (P=0.05) between Top (T) and Top 5 cm (T5) tissue Figure 5.4: Distribution of total flavonoids in perennial ryegrass tissue after treatment with both UV-B light (280 – 315 nm) and PAR light (400 – 700 nm). Data was pooled as all grasses had the same response.
.

137
()† Significant difference occurred in flavonoid content for each grass on each day where letters are different. Significance at (P=0.05). Figure 5.5: Turfgrass flavonoid content in Creeping bentgrass, Tall Fescue and Perennial Ryegrass grown in UV-B light (280 – 315 nm) and PAR light (400 – 700 nm) D0 to D7.

138
† Retention times, mass, spectral information and order of elution provided in Table 5.2.
Figure 5.6: Anthocyanin profile of creeping bentgrass ‘L-93’ exposed for 7 days to UV-B light. Produced from HPLC analysis using C18 column and light absorbance measured at 520 nm using a PDA.

139
† 1 – Zeaxanthin; 2 – β – carotene. Figure 5.7: Chromatogram of Perennial ryegrass carotenoids with absorbance at 453 nm. Data produce using C30 column and PDA connected to Chemstation software.

140
†
Means followed by different letters significantly different (P=0.0).
Figure 5.8. β-carotene (top) and zeaxanthin (bottom) content in creeping bentgrass, tall fescue and perennial ryegrass exposed to UV-B light D0 – D7.

141
REFERENCE
Anderson, J.M., and E-M. Aro. 1994. Grana stacking and protection of Photosystem II in
thylakoid membranes of higher plant leaves under sustained high irradiance: Ann. Hypoth. Photosynth. Res. 41:315-326.
Aro, E-M., I. Virgin, and B. Andersson. 1993. Photoinhibition of photosystem II.
Inactivation, protein damage and turnover. Biochim. Biophys. Acta. 1143:113-34. Babu, T.S., M.A.K. Jansen, B.M. Greenberg, V. Gaba, S. Malkin, A.K. Mattoo, and M.
Edelman. 1999. Amplified degradation of photosystem II D1 and D2 proteins under a mixture of photosynthetically active radiation and UV-B radiation: Dependence on redox status of photosystem II. Photochem. Photobiol. 69:553-559.
Beggs, C.J., and E. Wellmann. 1985. Analysis of light-controlled anthocyanin formation
in coleoptiles of Zea mays L.: The role of UV-B, blue, red and far-red light. Photochem. Photobiol. 41:481-486.
Bell, G.E., and T.K. Danneberger, 1999.Temporal shade on creeping bentgrass turf. Crop
Sci. 39:1142-1146. Britton, G., S. Liaaen-Jensen, and H. Pfander. 2004. Carotenoids handbook. Birkhäuser
Verlag, Boston, MA.
Buer, C.S., and G.K. Muday. 2004. The transparent testa4 mutation prevents flavonoid synthesis and alters auxin transport and the response of Arabidopsis roots to gravity and light. Plant Cell. 16:1191-1205.

142
Burger, J., and G. E. Edwards. 1996. Photosynthetic efficiency, and photodamage by UV and visible radiation, in red versus green leaf coleus varieties. Plant Cell Physiol. 37:395-399.
Caldwell, M.M. 1971. Solar UV irradiation and the growth and development of higher
plants. p. 131-177. In A.C. Giese (ed.) Photophys. Academic Press, NY. Caldwell, M.M., J.H. Richards, D.A. Johnson, R.S. Nowak, and R. S. Dzurec. 1981.
Coping with herbivory: Photosynthetic capacity and resource allocation in two semiarid Agropyron bunchgrasses. Oecologia 50:14-24.
Carletti, P., A. Masi, A. Wonisch, D. Grill, M. Tausz, and M. Ferretti. 2003. Changes in
antioxidant and pigment pool dimensions in UV-B irradiation maize seedlings. Environ. Exp. Bot. 50:149-157.
Casal, J.J., V.A. Deregibus, and R.A. Sánchez. 1985. Variations in tiller dynamics and
morphology in Lolium multiflorum Lam. vegetative and reproductive plants as affected by differences in red/far-red irradiation. Ann. Bot. 56:553-559.
Chandra, A., J. Rana, and Y. Li. 2001. Separation, identification, quantification, and
method validation of anthocyanins in botanical supplement raw materials by HPLC and HPLC-MS. J. Agric. Food Chem. 49:3515-3521.
Correia, C.M., M.S. Torres-Pereira, and J.M.G. Torres-Pereira. 1999. Growth,
photosynthesis and UV-B absorbing compounds of Portuguese Barbela wheat exposed to ultraviolet-B radiation. Environ. Pollut. 104:383-388.
Cortell, J.M., and J.A. Kennedy. 2006. Effect of shading on accumulation of flavonoid
compounds in (Vitis vinifera L.) pinot noir fruit and extraction in a model system. J. Agric. Food Chem. 49:3515-3521.
Costa, H., S.M. Gallego, and M.L. Tomaro. 2002. Effect of UV-B radiation on
antioxidant defense system in sunflower cotyledons. Plant Sci. 162:939-945.

143
Cronin, G., and D.M. Lodge. 2003. Effects of light and nutrient availability on the growth allocation, carbon/nitrogen balance, phenolic chemistry and resistance to herbivory of two freshwater macrophytes. Oecologia 137:31-41.
Curran, P.J., J.L. Dungan, and H.L. Gholz. 1990. Exploring the relationship between
reflectance red edge and chlorophyll content in slash pine. Tree Physiol. 7:33-48. Day, T.A. 1993. Relating UV-B radiation screening effectiveness of foliage to absorbing-
compound concentration and anatomical characteristics in a diverse group of plants. Oecologia 95:542-550.
de Sá, M.C. and D.B. Rodriguez-Amaya. 2003. Carotenoid composition of cooked green
vegetables from restaurants. Food Chem. 83:595-600. Do, C.B., and F. Cormier. 1991. Accumulation of peonidin 3-glucoside enhanced by
osmotic stress in grape (Vitis vinifera L.) cell suspension. Plant Cell Tiss. Organ Cult. 24:49-54.
Duval, B., K. Shetty, and W.H. Thomas. 1999. Phenolic compounds and antioxidant
properties in the snow alga Chlamydomonas nivalis after exposure to UV light. J. App. Phycol. 11:559-566.
Ferruzzi, M.G., L.C. Sander, C.L. Rock, and S.J. Schwartz. 1998. Carotenoid
determination in biological microsamples using liquid chromatography with a coulometric electrochemical array detector. Anal. Biochem. 256:74-81.
Friso, G., R. Barbato, G.M. Giacometti, and J. Barber. 1994. Degradation of D2 protein
due to UV-B irradiation of the reaction center of photosystem II. FEES Lett. 339: 217-221.
Fu, J., and B. Huang. 2004. Leaf characteristics associated with drought resistance in tall
fescue cultivars. Acta Hortic. 661:233-239.

144
Gao, W., Y. Zheng, J.R. Slusser, and G.M. Heisler. 2003. Impact of enhanced ultraviolet- B irradiance on cotton growth, development, yield, and qualities under field conditions. Agric. For. Meteorol. 120:241-248.
Gitelson, A.A., A. Vina, S.B. Verma, D.C. Rundquist, T.J. Arkebauer, G. Keydan, B.
Leavitt, V. Ciganda, G.G. Burba, and A.E. Suyker. 2006. Relationship between gross primary production and chlorophyll content in crops: Implications for the synoptic monitoring of vegetation productivity. J. Geophys. Res. 111: D08S11 Available at http://dx.doi.org/10.1029/2005JD006017 (Verified 30 Jun. 2012).
Gitz, D., D.L. Liu, and J.W. McClure. 1998. Phenolic metabolism, growth, and UV-B
tolerance in phenylalanine ammonia-lyase inhibited red cabbage seedlings. Photochemistry 49:377-386.
Giusti, M.M., L.E. Rodríguez-Saona, D. Griffin, and R.E. Wrolstad. 1999. Electrospray
and tandem mass spectroscopy as tools for anthocyanin characterization. J. Agric. Food Chem. 47:4657-4664.
Gould, K.S.2004. Nature's swiss army knife: The diverse protective roles of anthocyanins
in leaves. J Biomed. Biotechnol. 5:314-320. Greenburg, B.M., V. Gaba, A.K. Mattoo, and M. Edelman. 1989. Degradation of the 32-
KDa Photosystem II reaction center protein in UV, visible and far red light occurs through a common 23.5 KDa intermediate. Z Naturforsch. C:Biosci. 44:450-452.
He, Y-Y. D-P. Häder. 2002. UV-B-induced formation of reactive oxygen species and
oxidative damage of the cyanobacterium Anabaena sp.: protective effects of ascorbic acid and N-acetyl-L-cysteine. J. Photochem. Photobiol. B. 66: 115-124.
Hideg, É. and I. Vas. 1996. UV-B induced free radical production in plant leaves and
isolated thylakoid membranes. Plant Sci. 115:251-260. Jansen, M.A.K, B. Depka, A. Trebst, and M. Edelman. 1993. Engagement of specific
sites in the plastoquinone niche regulates degradation of the D1 protein in Photosystem II. J. Biol. Chem. 268:21246-21252.

145
Karageorgou, P. and Y. Manetas. 2006. The importance of being red when young: anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light. Tree Physiol. 26:613-621.
Kimura, M., D.B. Rodriguez-Amaya, and H.T. Godoy. 1990. Assessment of the
saponification step in the quantitative determination of carotenoids and provitamins Ann. Food Chem. 35:187-195.
Kovács, E., and Á. Keresztes. 2002. Effect of gamma and UV-B/C radiation on plant
cells. Micron 33:199-210. Krizek, D.T., R.M. Mirecki, and S.J. Britz. 1997. Inhibitory effects of ambient levels of
solar UV-A and UV-B radiation on growth of cucumber. Physiol. Plant. 100:886-893.
Landry, L.G., C.C. Chapple, and R.L. Last. 1995. Arabidopsis mutants lacking phenolic
sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant Physiol. 109:1159-1166.
Lin, C., M. Ahmad, and A.R. Cashmore. 1996. Arabidopsis cryptochrome 1 is a soluble
protein mediating blue light-dependent regulation of plant growth and development. Plant J. 5:893-902.
Lois, R. 1994. Accumulation of UV-absorbing flavonoids induced by in Arabidopsis
thaliana L. I. Mechanisms of UV - resistance in Arabidopsis. Planta 194:498- 503.
Mackerness, S.A-H. 2000. Plant responses to ultraviolet-B (UV-B: 280–320 nm) stress:
What are the key regulators? Plant Growth. Reg. 32:27-39. Mathews-Roth, M.M. 1986. Carotenoids quench evolution of excited species in
epidermis exposed to UV-B (290-320 nm) light. Photochem. Photobiol. 43:91–93. Maxwell, K., and G.N. Johnson. 2000. Chlorophyll fluorescence – a practical guide. J.
Exp. Bot. 51:659-668.

146
McDonald, S., P.D. Prenzler, M. Autolovich, and K. Robards. 2001. Phenolic content and antioxidant activity of olive extracts. Food Chem. 73:73-84.
McElroy, J.S., D.A. Kopsell, J.C. Sorochan, and C.E. Sams. 2006. Response of creeping
bentgrass carotenoid composition to high and low irradiance. Crop Sci. 46:2606-2612.
Milesi, C., S.W. Running, C.D. Elvidge, J.B. Dietz, and B.T. Tuttle. 2005. Mapping and
modeling the biogeochemical cycling of turf grasses in the United States. Environ. Manage. 36:426-438.
Moran, R. 1982. Formulae for determination of chlorophyllous pigments extracted with
N,N-dimethylformamide. Plant Physiol. 69:1376-381. Mori, M., K. Yoshida, Y. Ishigaki, T. Matsunaga, O. Nikaido, K. Kameda, and T. Kondo.
2005. UV-B protective effect of a polyacylated anthocyanin, HBA, in flower petals of the blue morning glory, Ipomoea tricolor cv. Heavenly Blue. Bioorg. Med. Chem. 13:2015-2020.
Moros, E.E., D. Dsrnoko, M. Cheryan, E.G. Perkins, and J. Jerrell. 2002. Analysis of
xantophylls in corn by HPLC. J. Agric. Food Chem. 50:5787-5790. Pourmorad, F., S.J. Hosseinimehr, and N. Shahabimajd. 2006. Antioxidant activity,
phenol and flavonoid contents of some selected Iranian medicinal plants. Afric. J. Biotechnol. 5:1142-1145.
Rosario, R., G.J. Mark, J.A. Parrish, and M.C. Mihm Jr. 1979. Histological changes
produced in skin by equally erythemogenic doses of UV-A, UV-B, UV-C and UV-A with psoralens. Brit. J. Derm. 101:299-308.
Sancar, A. and G.B. Sancar. 1988. DNA repair enzymes. Ann. Rev. Biochem. 57:29-67. Sarkar, D., P.C. Bhowmik, Y.I. Kwon, and K. Shetty. 2011. The role of proline-
associated pentose phosphate pathway in cool-season turfgrasses after UV-B exposure. Environ. Exp. Bot. 70:251-258.

147
Smith, H. 1994. Sensing the light environment: the functions of the phytochrome family. p. 377-416. In J.R.E. Kendrick and G.H Kronenberg (ed.) Photomorphogenesis in Plants. Kluwer Academic Publishers, Dordrecht, The Netherlands.
Strid, A., W.S. Chow, and J.M. Anderson. 1990. Effects of supplementary ultraviolet – B
radiation on photosynthesis in Pisium sativum. Biophys. Biochim. Acta. 1020:260-268.
Taalas, P., J. Kaurola, A. Kylling, D. Shindell, R. Sausen, M. Dameris, V. Grewe, J.
Herman, J. Damski, and B. Steil. 2000. The impact of greenhouse gases and halogenated species on future solar UV radiation doses. Geophys. Res. Lett. 27:1127-1130.
Takahashi, S. and N. Murata. 2008. How do environmental stresses accelerate
photoinhibition? Trends Plant Sci. 13:178-182. Teramura, A.H., R. Hilton Biggs, and S. Kossuth. 1980. Effects of ultraviolet-B
irradiances on Soybean II. Interaction between ultraviolet-B and photosynthetically active radiation on net photosynthesis , dark respiration, and transpiration. Plant Physiol. 65:483-488
Teramura, A.H., and J.H. Sullivan. 1994. Effects of UV-B radiation on photosynthesis
and growth of terrestrial plants. Photosynth. Res. 39:463-473. Tevini, M., M.W. Iwanzik, and U. Thoma. 1981. Some effects of enhanced UV-B
irradiation on the growth and composition of plants. Planta 153:388-394. Tyystjärvi, E. 2008. Photoinhibition of photosystem II and photodamage of the oxygen
evolving manganese cluster. Coordination Chemistry Reviews. The Role of Manganese in Photosystem II. 252:361-376.
Vass, I., L. Sass, C. Spetea, A. Bakou, D.F. Ghanotakis, and V. Petroulea. 1996. UV-B-
induced inhibition of photosystem II electron transport studied by EPR and chlorophyll fluorescence. Impairment of donor and acceptor side components. Biochem. J. 35:8964-8973.

148
Wherley, B.G., D.S. Gardner, and J.D. Metzger. 2005. Tall fescue photomorphogenesis as influenced by changes in the spectral composition and light intensity. Crop Sci. 45:562-568.
White, A.L., and L.S. Jahnke. 2002. Contrasting effects of UV-A and UV-B on
photosynthesis and photoprotection of β-carotene in two Dunaliella spp. Plant Cell Physiol. 43:877-884.
Woodall, G.S., and G.R. Stewart. 1998. Do anthocyanins play a role in UV protection of
the red juvenile leaves of Syzygium? J. Exp. Bot. 49:1447-1450. Yin, Z-H., S. Neimanis, U. Wagner, and U. Hebe. 1990. Light-dependent pH changes in
leaves of C3 plants I. Recording pH changes in various cellular compartments by fluorescent probes. Planta 182:244-252.
Zhang, X., E.H. Ervin, and R.E. Schmidt. 2005. The role of leaf pigment and antioxidant
levels in UV-B resistance of dark- and light-green Kentucky bluegrass cultivars. J. Am Soc. Hortic. Sci. 130:836-841.
Zhang, X., and E.H. Ervin. 2009. Physiological assessment of cool-season turfgrasses
under ultraviolet-B stress. HortScience 44:1785-1789. Ziska, L.H., A.H. Teramura, J.H. Sullivan, and A. McCoy. 1993. Influence of ultraviolet-
B (UV-B) radiation on photo-synthetic and growth characteristics in field-grown cassava (Manihot esculentum Crantz). Plt. Cell Environ. 16:73-79.

149
CHAPTER 6
OVERALL CONCLUSIONS ON UV-B INFLUENCE ON TURFGRASS PIGMENTS AND GROWTH
ABSTRACT
The studies conducted for this project have yielded a range of insights as to the
effects of UV-B on different cool season turfgrasses. In looking at growth and
germination there is an applied element to the turfgrass knowledge which adds to the
expanding base of information in place. Investigating the changes in pigmentation and
developing new information and areas of research has created potential for new turfgrass
cultivar breeding and also offers insight into physiological changes that previously were
undocumented in turfgrass. Overall conclusions and ideas for future work are offered.

150
DISCUSSION
The influence of UV-B light from these studies on turfgrass is varied. From the
initial growth and germination stage to maturity and canopy effects, there were some
benefits and some negative impacts. The use of UV to enhance Kentucky bluegrass seed
germination offers the potential to increase chances of successful establishment in a
broader range of environmental conditions. Further studies, including field research,
should be conducted in this area as this was the first trial using UV light and Kentucky
bluegrass seed. There were also some issues with both the chambers and the lighting.
Seed age seemed to reduce the effectiveness of UV light in seed germination overriding
any benefit. The exposure of Kentucky bluegrass seedlings to UV did not seem to
damage seed coats prior to germination and further work on long term exposure of
Kentucky bluegrass seedlings to UV is warranted. Enzymatic responses and breakdown
of starch reserves undoubtedly will paint a clearer picture for the method’s potential use.
Investigating what affects turfgrass color raised some interesting questions.
Previous research suggests that adding nitrogen darkens turfgrass. The pigment
correlation or lack ofit in cultivars which were not forced by different rates of nitrogen
applications was somewhat surprising as plants were not chlorotic and seemed dark green
in the trial. Nitrogen was supplied at a single rate and thus this would explain the green
color on its own. The data analysis indicated that the hand-held system which depended
on light reflection failed to be influenced by pigment content and did not correlate to
human visual quality rankings. This is somewhat of a concern as handheld systems are
becoming prevalent and may mean that a whole new ranking database will need to be

151
built up if they are going to become standard. There was a relatively strong relationship
found between both systems and leaf tissue nitrogen content, thus handheld systems
measuring nitrogen might be more appropriate. The extract analysis undoubtedly offered
more information and links to pigment changes; however its cost and time delay is
problematic. Other pigments could also play a role in color which we did not account for
of course. Pigments such as carotenoids absorb light and may completely reduce
reflection of light in the blue region. This would account for the fact that the systems
could not separate out green colors. Trained evaluators, using the long standard NTEP +9
scale, may still be the most consistent and precise method for color rankings.
Increasing UV-B light in this situation on mature stands of cool season turfgrasses
produced some interesting and exciting responses. Characterizing anthocyanins for the
first time in turfgrass is a mile stone and offers a whole new area of research in turfgrass.
Potential lies for pigments in a range of areas and if turfgrasses can be controlled in a
way that naturally enhances pigment production, then a potential for improved stress
tolerance exists. The result here was not overly consistent; however, by varying light
intensity and possibly temperature then opportunity exists for quantification and greater
characterization of the pigments. The perennial habit of turf also offers a possibility to be
harvested with the aim of using the pigments possibly as natural food colorants or light
absorbing dyes. The characterization of the impact of UV-B on specific carotenoid
content is also a first in turfgrass management and shows how important these pigments
are in grass. Reductions in carotenoid content and the publication of chromatograms are a
first within the turfgrass research area on carotenoids. As herbicide modes of action

152
continue to diversify into bleaching and photoinhibition due to damage to photosynthetic
apparatus, knowledge of these pigments is required within grasses to offer potential for
selective control.
The turfgrass decline in both the UV-B and control chambers is of interest. In
summer periods PAR light intensity is far higher than was found in these studies and so
turfgrass undergoes a daily stress of photo-inhibition. This low light however is a
problem which occurs when using growth chambers, depending on the grass, turfgrasses
may still be susceptible to photoinhibition in chambers. The photoinhibition can vary
between rapidly repaired dynamic photoinhibition to chronic photoinhibition leaving
turfgrass with a reduced capacity for optimal productivity. This is particularly true for C3
turfgrasses, and during the period of the year when opportunities for carbon sequestration
are greatest due to higher light levels. This undoubtedly plays into a loss of quality during
stressful periods of the year. It also may reduce turfgrass plant’s competitive ability with
weedy grasses and further work is currently ongoing looking into this acclimation during
seasonal changes in light.
The difference between the quality of grasses that have a shiny leaf surface and
thicker leaves compared to the thin-leaved bentgrasses is also of interest. Perennial
ryegrass and tall fescue which are considered to have shiny leaf surfaces were not as
affected as the bentgrasses and so reflective characteristics of the leaf surface may be
important. Limited attention has been paid to turfgrass leaf epidermal content changes in
high light conditions. This may become more of an interest going forward as new
herbicides aim for selectivity with greater potential for damage to target species due to an

153
ability to penetrate leaf surfaces. It is also of concern for herbicide use in summer periods
where potential damage to non-target species can occur due to changes in epidermal
surfaces. Reducing the impact of higher UV-B levels becomes more important from a
plant health standpoint with the likely increase in levels penetrating the ozone in the next
ten years (2010-2020).

154
APPENDIX A: HANDHELD REFELCTANCE MEASUREMENTS OF TURFGRASS TISSUE

155

156
APPENDIX B: COLOR OF LEAVES OF CREEPING BENTGRASS (Agrostis stolonifera L.) TISUE AFTER EXPOSURE TO UV-B LIGHT

157

158
APPENDIX C: CHROMATOGRAM CAROTENOID STANDARDS

159
# Standard Time (min) Area Area % 1 Violaxanthin 6.5 78.9 2.6 2 Antheraxanthin 8.3 1482.6 50.4 3 Lutein 10.0 510.6 17.3 4 Zeaxanthin 11.2 267.2 9.1 5 Β-carotene 23.0 603.6 20.5
Time (min)

160
APPENDIX D: CHROMATOGRAM CHOKEBERRY ARONIA

161

162
APPENDIX E: CHLOROPHYLL A CONTENT AFTER D7 UV TREATMENT CREEPING BENTGRASS ‘L-93’

163
0
100
200
300
400
0 2 4 6 8
Chl
orop
hyll
A c
onte
nt (µ
g m
-2)
Day
Chlorophyll A Creeping bentgrass 'Penncross'
UVCTRL

164
APPENDIX F: CHROMATOGRAM BENTGRASS CAROTENOIDS

165
# Time (min) Area Area% Zeaxanthin 10.4 43.1 3.2 2 17.5 72.4 5.4 3 19.5 31.1 2.3 4 20.2 50.3 3.8 β-carotene 22.5 891.4 66.7 6 23.6 168.6 12.6
Time (min)

166
APPENDIX G: TALL FESCUE CAROTENOID CHROMATOGRAM

167
# Time (min) Area Area% Zeaxanthin 10.5 54.6 1.6 2 17.5 32.9 0.8 3 19.4 66.9 2.0 4 20.0 143.3 4.2 β-carotene 22.53 2686.3 80.4 6 23.3 339.3 10.16
Time (min)

168
COMPLETE REFERENCE Ahmed, J., U.S. Shivhare, and G.D. Singh. 2000. Chlorophyll and color of green chili
puree as affected by mesh size and temperature. Int. J. Food Prop. 3:305-315. Ahmed, J., U.S. Shivhare, and G.S.V. Raghavan. 2004. Thermal degradation kinetics of
anthocyanin and visual colour of plum puree. Euro. Food Res. Tech. 218:525-528 Alba, R., M.M. Cordonnier-Pratt, and L.H. Pratt. 2000. Fruit-localized phytochromes regulate lycopene accumulation independently of ethylene production in tomato.
Plant Physiol. 123:363-370. American Society for Testing and Materials. 1989. Standard test method for specifying color by the munsell system. ASTM standard D1535–89. ASTM, Philadelphia,
PA. Anderson, J.M., and E.-M. Aro. 1994. Grana stacking and protection of Photosystem II in thylakoid membranes of higher plant leaves under sustained high irradiance: Ann.
Hypoth. Photosynth. Res. 41:315-326. Ang, L.-H., S. Chattopadhyay, N. Wei, T. Oyama, K. Okada, A. Batschauer, and X.-W
Deng. 1998. Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol. Cell. 1:213–222.
Anonymous, 1997. Flavonoids (isoflavonoids and neoflavonoids). McNaught A.D. and
A.Wilkinson (ed.) IUPAC Compendium of Chemical Terminology, 2nd ed. Available at http://old.iupac.org/goldbook/F02424.pdf (verified 28 Jun. 2012).
Aro, E-M., I. Virgin, and B. Andersson. 1993. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta. 1143:113-34.

169
Babu, T.S., M.A.K. Jansen, B.M. Greenberg, V. Gaba, S. Malkin, A.K. Mattoo, and M. Edelman. 1999. Amplified degradation of photosystem II D1 and D2 proteins
under a mixture of photosynthetically active radiation and UV-B radiation: Dependence on redox status of photosystem II. Photochem. Photobio. 69:553-559.
Bacci, L., D. Grifoni, F. Sabatini, and G. Zipoli. 1999. UV-B radiation causes early ripening and reduction in size of fruits in two lines of tomato (Lycopersicon
esculentum Mill.). Glob. Change Bio. 5:635-646. Bagegni, A.M., D.A. Sleper, H.D. Kerr, and J.S. Morris. 1990. Viability of Acremonium coenophialum in tall fescue seed after ionizing radiation treatments. Crop Sci.
30:1272-1275. Ballare, C.L., A.L. Scopel, A.E. Stapleton, and M.J. Yanovsky. 1996. Solar ultraviolet-B radiation affects seedling emergence, DNA integrity, plant morphology, growth
rate, and attractiveness to herbivore insects in Datura ferox. Plant Physiol. 112:161-170.
Barnes, P.W., S.D. Flint, and M.M. Caldwell. 1990. Morphological responses of crop and weed species of different growth forms to ultraviolet-B radiation. American J. of
Bot. 77:1354-1360. Bartley, G.E., and P.A. Scolnik. 1995. Plant carotenoids: Pigments for photoprotection, visual attraction and human health. Plant Cell. 7:1027-1038. Bausch, W.C., and H.R. Duke. 1996. Remote sensing of plant nitrogen status in corn. Trans. ASAE. 39: 1869-1875. Beam, J.B., W.L. Barker, and S.D. Askew. 2006. Selective creeping bentgrass (Agrostis stolonifera L.) control in cool season turfgrass. Weed Tech. 20:3430-344. Beggs, C.J., and E. Wellmann. 1985. Analysis of light-controlled anthocyanin formation in coleoptiles of Zea mays L.: The role of UV-B, blue, red and far-red light.
Photochem. Photobio. 41:481-486.

170
Bell, G.E., and T.K. Danneberger. 1999. Temporal shade on creeping bentgrass turf. Crop Sci. 39:1142-1146. Bell, G.E., T.K. Danneberger, and M.J. McMahon. 2000. Spectral irradiance available for growth in sun and shade. Crop Sci. 40:189-195. Bennett, R.N., and R.M. Wallsgrove. 2006. Secondary metabolites in plant defense mechanisms. New Phytol. 127:617-633. Berg, H. 2008. Johann Wilhelm Ritter – The founder of scientific electrochemistry. Rev. Polarography. 54:99-103. Bilger, W., M. Veit, L. Schreiber, and U. Schreiber. 1997. Measurement of leaf epidermal transmittance of UV radiation by chlorophyll fluorescence. Physiolog.
Planta. 101:754-763. Björn, L.O., T.V. Callaghan, C. Gehrke, U. Johanson, M. Sonesson, and D. Gwynn- Jones. 1998. The problem of ozone depletion in northern Europe. Ambio 27:275-
279. Blumenthaler, M., and W. Ambach. 1990. Indication of increasing solar ultraviolet-B radiation flux in Alpine regions. Science 248:206-208. Bornman, J.F., and T.C. Vogelmann. 1988. Penetration of blue and UV radiation measured by fiber optics in spruce and fir needles. Physiolog. Planta. 72:699-705. Bornman, J.F. 1989. Target sites of UV-B radiation in photosynthesis of higher plants. J. Photochem. Photobiol. 4:145-158. Bornman, J.F., and T.C. Vogelmann. 1991. Effect of UV-B radiation on leaf optical properties measured with fiber optics. J. Exp. Bot. 42:547-554.

171
Bornman, J.F., and A.H. Teramura. 1993. Effects of ultraviolet-B radiation on terrestrial plants. p. 427-471. In A.R. Young et al. (ed.) Environmental UV Photobiology.
Plenum Press, New York. Brandle, J.R., W.F. Campbell, W.B. Sisson, and M.M. Caldwell. 1977. Net photosynthesis, electron transport capacity and ultrastructure of Pisium sativum L.
exposed to ultraviolet-B radiation. Plant Physiol. 60:165-169. Briscoe, A.D., and L. Chittka. 2001. The evolution of colour vision in insects. Ann. Rev.
Entomol. 46:471-510 Britt, A.B. 1996. DNA damage and repair in plants. Annu. Rev. Plant Physiol. Plant Mol.
Biol. 47.75-100. Britton, G., S. Liaaen-Jensen, and H. Pfander. 2004. Carotenoids handbook. Birkhäuser Verlag, Boston, MA. Brouillard, R. 1988. Flavonoids and flower color. P. 525–538. In J.B. Harborne (ed.) The flavonoids: Advances in research since 1980. Chapman Hall, London, United
Kingdom. Brown, P.K., and G. Wald. 1964. Visual pigments in single rods and cones of the human retina. Science 1964. 144:45-52. Buer, C.S., and G.K. Muday. 2004. The transparent testa4 mutation prevents flavonoid synthesis and alters auxin transport and the response of Arabidopsis roots to
gravity and light. Plant Cell. 16:1191-1205. Burger, J., and G. E. Edwards. 1996. Photosynthetic efficiency, and photodamage by UV and visible radiation, in red versus green leaf coleus varieties. Plant Cell Physiol.
37:395-399. Byrne, M.C., C.J. Nelson, and D.D. Randall. 1981. Ploidy effects on anatomy and gas exchange of tall fescue leaves. Plant Physiol. 68:891-893.

172
Caldwell, M.M. 1971. Solar ultraviolet radiation and the growth and development of higher plants. p. 131-174. In: A.C. Giese (ed.) Photophysiology. Academic Press,
New York. Caldwell, M.M. 1979. Plant life and ultraviolet radiation: Some perspective in the history of the earth's UV climate. BioScience 29:520-525. Caldwell, M.M., R. Robrecht, and W.D. Billings. 1980. A steep latitudinal gradient of solar ultraviolet-B radiation in the arctic-alpine life zone. Ecology 6:600-611. Caldwell, M.M., J.H. Richards, D.A. Johnson, R.S. Nowak, and R. S. Dzurec. 1981. Coping with herbivory: Photosynthetic capacity and resource allocation in two
semiarid Agropyron bunchgrasses. Oecologia 50:14-24. Caldwell, M.M., A.H. Teramura, and M. Tevini. 1989. The changing solar ultraviolet climate and the ecological consequences for higher plants. Trends Ecol. Evol.
4:363-367. Cameron, R.J. 1970. Light intensity and the growth of Eucalyptus seedlings: II. The effect of cuticular waxes on light absorption in leaves of Eucalyptus species. Aus.
J. Bot. 18:275-284. Carletti, P., A. Masi, A. Wonisch, D. Grill, M. Tausz, and M. Ferretti. 2003. Changes in antioxidant and pigment pool dimensions in UV-B irradiation maize seedlings.
Environ. Exp. Bot. 50:149-157. Carrow, R.N., D.V. Waddington, and P.E. Reike. 2001. Turfgrass soil fertility and
chemical problems. John Wiley & Sons, Inc., Hoboken, NJ. Casal, J.J., V.A. Deregibus, and R.A. Sánchez. 1985. Variations in tiller dynamics and morphology in Lolium multiflorum Lam. vegetative and reproductive plants as
affected by differences in red/far-red irradiation. Ann. Bot. 56:553-559. Castle, E.S. 1935. Photic excitation and phototropism in single plant cells. Cold Spring Harbor Symp. Quant. Biol. 3:224-229.

173
Cavalcante, M.J.B., and J.J. Muchovej. 1993. Microwave irradiation of seeds and selected fungal spores. Seed Sci. Technol. 21:247-253. Cen, Y.-P., and J.F. Bornman. 1990. The response of bean plants to UV-B radiation
under different irradiances of background visible light. J. Exp. Bot. 41:1489-1495. Chalker-Scott, L. 1999. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol. 70:1-9. Chandra, A., J. Rana, and Y. Li. 2001. Separation, identification, quantification, and method validation of anthocyanins in botanical supplement raw materials by
HPLC and HPLC-MS. J. Agric. Food Chem. 49:3515-3521. Chen, C. 2002. Robust regression and outlier detection with the ROBUSTREG procedure. Paper 265-27. SAS Institute Inc., Cary, NC, Proceedings SUGI 27,
Orlando FL Apr 14–17. Available at http://www2.sas.com/proceedings/sugi27/p265-27.pdf (verified, 28 June, 2012). Chien, J.C., and I.M. Sussex. 1996. Differential regulation of trichome formation on the adaxial and abaxial leaf surfaces by gibberellins and photoperiod in Arabidopsis
thaliana (L.) Heynh. Plant Physiol. 111:1321-1328. Clark, J.B, and G.R. Lister. 1975. Photosynthetic action spectra of trees: II. The relationship of cuticle structure to the visible and ultraviolet spectral properties of
needles from four coniferous species. Plant Physiol. 55:407-413. Correia, C.M., M.S. Torres-Pereira, and J.M.G. Torres-Pereira. 1999. Growth, photosynthesis and UV-B absorbing compounds of Portuguese Barbela wheat
exposed to ultraviolet-B radiation. Environ. Pollut. 104:383-388. Cortell, J.M., and J.A. Kennedy. 2006. Effect of shading on accumulation of flavonoid compounds in (Vitis vinifera L.) pinot noir fruit and extraction in a model system.
J. Agric. Food Chem. 49:3515-3521.

174
Costa, H., S.M. Gallego, and M.L. Tomaro. 2002. Effect of UV-B radiation on antioxidant defense system in sunflower cotyledons. Plant Sci. 162:939-945. Cronin, G., and D.M. Lodge. 2003. Effects of light and nutrient availability on the growth allocation, carbon/nitrogen balance, phenolic chemistry and resistance to
herbivory of two freshwater macrophytes. Oecologia 137:31-41. Croteau, R., T.M. Kutchan, and N.G. Lewis. 2000. Natural products (secondary metabolites). p. 1250-1318. In B. Buchanan et al. (ed.) Biochem. Molec. Biol.
Plants Am. Soc. Plant Physiol. Cuadra, P., J.B. Harborne, and P.G. Waterman. 1997. Increases in surface flavonols and photosynthetic pigments in Gnaphalium luteo-album in response to UV-B
radiation. Phytochemistry 45:1377-1383. Cunningham, F.X., and E. Gantt. 2002. Genes and enzymes of carotenoid biosynthesis in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 49:577-583. Curran, P.J., J.L. Dungan, and H.L. Gholz. 1990. Exploring the relationship between reflectance red edge and chlorophyll content in slash pine. Tree Physiol. 7:33-48. Cuttriss, A., and B. Pogson. 2004. Carotenoids. In: Plant pigments and their manipulation. Ann. Plant Rev. 14:57-91. Dai, Q., V.P. Coronel, B.S. Vergara, P.W. Barnes, and A.T. Quintos. 1992. Ultraviolet-B radiation effects on growth and physiology of four rice cultivars. Crop Sci.
32:1269-1274. Danneberger, T.K., M.B. McDonald Jr., C.A. Geron, and P. Kumari. 1992. Rate of germination and seedling growth of perennial ryegrass seed following
osmoconditioning. HortScience 27:28-30. Day, T.A., T.C. Vogelmann, and E.H. DeLucia. 1992. Are some plant life forms more effective than others in screening out ultraviolet-B radiation? Oecologia 92:513-
519.

175
Day, T.A. 1993. Relating UV-B radiation screening effectiveness of foliage to absorbing- compound concentration and anatomical characteristics in a diverse group of
plants. Oecologia 95:542-550. Deckmyn, G., and I. Impens. 1999. Seasonal responses of six Poaceae to differential levels of solar UV-B radiation. Environ. Exp. Bot. 41:177-184. Deckmyn, G., and I. Impens. 1998. UV-B and PAR in a grass (Lolium perenne L.) canopy. Plant Ecol. 137:13-19. Deckmyn, G., and I. Impens. 1998. Effects of solar UV-B irradiation on vegetative and generative growth of Bromus catharticus. Env. Exp. Bot. 40:179-185. de Sá, M.C. and D.B. Rodriguez-Amaya. 2003. Carotenoid composition of cooked green vegetables from restaurants. Food Chem. 83:595-600. Dicke, M., A.A. Agrawal, and J. Bruin. 2003. Plants talk, but are they deaf? Trends Plant Sci. 8:403-405. Do, C.B., and F. Cormier. 1991. Accumulation of peonidin 3-glucoside enhanced by osmotic stress in grape (Vitis vinifera L.) cell suspension. Plant Cell Tiss. Organ
Cult. 24:49-54. Dong, Y. hu, D. Mitra, A. Kootstra, C. Lister, and J. Lancaster. 1995. Postharvest stimulation of skin color in royal gala apple. J. Am. Soc. Hortic Sci. 120:95-100. Donner, H., L. Gao, and G. Mazza. 1997. Separation and characterization of simple and malonylated anthocyanins in red onions, Allium cepa L. Food Res. Int. 30:637-
643. Duval, B., K. Shetty, and W.H. Thomas. 1999. Phenolic compounds and antioxidant properties in the snow alga Chlamydomonas nivalis after exposure to UV light. J.
Appl. Phycol. 11:559-566.

176
Ervin, E.H., X. Zhang, and J.H. Fike. 2004a. Ultravolet-B radiation damage on Kentucky bluegrass I: antioxidant and colorant effects. HortScience 39:1465-1470. Ervin, E.H., X. Zhang, and J.H. Fike. 2004c. Ultraviolet-B radiation damage on Kentucky bluegrass. III. Cultivar effects. HortScience 39:1475-1477. Escadafal, R., M-C. Girard, and D. Courault. 1989. Munsell soil color and soil reflectance in the visible spectral bands of landsat MSS and TM data. Remote
Sens. Environ. 27:37-46. Ferruzzi, M.G., L.C. Sander, C.L. Rock, and S.J. Schwartz. 1998. Carotenoid determination in biological microsamples using liquid chromatography with a
coulometric electrochemical array detector. Anal. Biochem. 256:74-81. Field, T.S., D.W. Lee, and N.M. Holbrook. 2001. Why leaves turn red in autumn. The role of anthocyanins in senescing leaves of red osier dogwood. Plant Physiol.
127:566-574. Filella, I., and J. Penuelas. 1994. The red edge and shape as indicators of plant chlorophyll, biomass and hydric status. Int. J. Remote Sensing. 15:1459-1470. Fiscus E.L., and F.L. Booker. 1995. Is increased UV-B a threat to crop photosynthesis and productivity? Photosynth. Res. 43:81-92. Flint, S.D., and M.M. Caldwell. 1984. Partial inhibition of in vitro pollen germination by simulated solar ultraviolet‑B radiation. Ecology. 65:792-795. Flint, S.D., and M.M. Caldwell. 2003. A biological spectral weighting function for ozone depletion research with higher plants. Physiol. Planta. 117:137-144. Floss, H.G. 1997. Natural products derived from unusual variants of the shikimate pathway. Natural Prod. Rep. 14:433-452.

177
Foti, M., M. Piattelli, V. Amico, and G. Ruberto. 1994. Antioxidant activity of phenolic meroditerpenoids from marine algae. J. Photochem. Photobiol. 26:159-164. Foyer, C.H., M. Lelandais, and K.J. Kunert. 1994. Photooxidative stress in plants. Physiol. planta. 92:696-717. Foyer, C.H., P. Descourvieres, and K.J. Kunert. 1994. Protection against oxygen radicals: an important defence mechanism studied in transgenic plants. Plant Cell and Env.
17:507-523. Frank, K.W., R.E. Gaussoin, T.P. Riordan, R.C. Shearman, J.D. Fry, E.D. Miltner, and P.G. Johnson. 2004. Nitrogen rate and mowing height effects on turf-type
buffalograss. Crop Sci. 44:1615–1621. Frankhauser, S., and J. Chory. 1997. Light control of plant development. Ann. Rev. Cell Dev. Biol. 13:203-29. Fraser, P.D., and P.M. Bramley. 2004. The biosynthesis and nutritional uses of carotenoids. Prog. Lipid Res. 43:228-265. Frederick, J.E., H.E. Snell, and E.K. Haywood. Solar ultraviolet radiation at the earths surface. Photochem. Photobio. 50:443-450. Friso, G., R. Barbato, G.M. Giacometti, and J. Barber. 1994. Degradation of D2 protein due to UV-B irradiation of the reaction center of photosystem II. FEES Lett. 339:
217-221. Fu, J., and B. Huang. 2004. Leaf characteristics associated with drought resistance in tall fescue cultivars. Acta Hortic. 661:233-239. Galston, A.W., and R.S. Baker. 1949. Studies on the physiology of light action II. The photodynamic action of riboflavin. Am. J. Bot. 36:773-780.

178
Gao, W., Y. Zheng, J.R. Slusser, and G.M. Heisler. 2003. Impact of enhanced ultraviolet- B irradiance on cotton growth, development, yield, and qualities under field
conditions. Agric. For. Meteorol. 120:241-248. Garcı́a-Viguera, C., and P. Bridle. 1999. Influence of structure on colour stability of anthocyanins and flavylium salts with ascorbic acid. Food Chem. 64:21-26. Gardner, F.P., R.B. Pearce, and R.L. Mitchell (ed.) 1985. Physiology of crop plants. Iowa State Univ. Press, Ames IA. Gausman, H.W., and R. Cardenas. 1969. Effect of leaf pubescence of Gynura aurantiaca on light reflectance. Bot. Gaz. 130:158-162. Gitelson, A.A., A. Vina, S.B. Verma, D.C. Rundquist, T.J. Arkebauer, G. Keydan, B. Leavitt, V. Ciganda, G.G. Burba, and A.E. Suyker. 2006. Relationship between
gross primary production and chlorophyll content in crops: Implications for the synoptic monitoring of vegetation productivity. J. Geophys. Res. 111: D08S11 Available at http://dx.doi.org/10.1029/2005JD006017 (Verified 30 Jun. 2012).
Gitz, D., D.L. Liu, and J.W. McClure. 1998. Phenolic metabolism, growth, and UV-B
Tolerance in phenylalanine ammonia-lyase inhibited red cabbage seedlings. Photochemistry 49:377-386.
Giusti, M.M., L.E. Rodríguez-Saona, D. Griffin, and R.E. Wrolstad. 1999. Electrospray and tandem mass spectroscopy as tools for anthocyanin characterization. J. Agric.
Food Chem. 47:4657-4664. Giusti, M.M., and R.E. Wrolstad. 2005. Characterization and measurement of anthocyanins by UV-visible spectroscopy. p. 19-32. Unit F1.2. In Wrolstad R.E.
(ed.) Handbook of food analytical chemistry—pigments, colorants, flavors, texture, and bioactive food components. John Wiley & Sons Inc., New York.
Gooding, M.J., and L.J. Gamble. 1990. Color evaluation of Poa pratensis cultivars. J. Sports Turf Res. Inst. 66:134-140.

179
Gould, K.S. 2004. Nature's swiss army knife: The diverse protective roles of anthocyanins in leaves. J. Biomed. Biotechnol. 5:314-320. Greenburg, B.M., V. Gaba, A.K. Mattoo, and M. Edelman. 1989. Degradation of the 32- KDa Photosystem II reaction center protein in UV, visible and far red light occurs
through a common 23.5 KDa intermediate. Z Naturforsch. C:Biosci. 44:450-452. Greenwood, D.J., G. LeMaire, G. Gosse, P. Cruz, A. Draycott, and J.J. Neeteson. 1990. Decline in percentage N of C3 and C4 crops with increasing plant mass. Ann. Bot.
66:425-436. Gross, J., 1991. Pigments in vegetables: chlorophylls and carotenoids. Van Nostrand Reinhold, NY. Guertal, E.A., and D.L. Evans. 2006. Nitrogen rate and mowing height effects on TifEagle bermudagrass establishment. Crop Sci. 46:1772-1778. Gwynn-Jones, D., and U. Johanson. 1996. Growth and pigment production in two subarctic grass species under four different UV-B irradiation levels. Physiol.
Planta. 97:701-707. Gwynn-Jones, D. 2001. Short-term impacts of enhanced UV-B radiation on photo- assimilate, allocation and metabolism: a possible interpretation for time-
dependent inhibition of growth. Plant Ecol. 154:67-73. Hagen, S.F., G.I.A. Borge, G.B. Bengtsson, W. Bilger, A. Berge, K. Haffner, and K.A. Solhaug. 2007. Phenolic contents and other health and sensory related properties
of apple fruit (Malus domestica Borkh., cv. Aroma): Effect of postharvest UV-B irradiation. Postharvest Bio. Technol. 45:1-10.
Hardy, A.C. 1938. History of the design of the recording spectrophotometer. J. Optic. Soc. Am. 28:360-363. Hare, P.D., and W.A., Cress. 1997. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Reg. 21:79-102.

180
Hartley, W.N. 1881. On the absorption of solar rays by atmospheric ozone. J. Chem. Soc. 39:111-128. He, Y-Y. D-P. Häder. 2002. UV-B-induced formation of reactive oxygen species and oxidative damage of the cyanobacterium Anabaena sp.: protective effects of
ascorbic acid and N-acetyl-L-cysteine. J. Photochem. Photobio. B: Biology. 66: 115-124.
Heller, W., and G. Forkmann. 1988. Biosynthesis in the flavonoids: p. 319-425. In J.B. Harborne (ed.) The flavonoids: Advances in research since 1980. Chapman Hall,
London, United Kingdom. Hideg, É. and I. Vas. 1996. UV-B induced free radical production in plant leaves and isolated thylakoid membranes. Plant Sci. 115:251-260. Hofmann, R.W. 2000. Population studies of ultraviolet-B radiation responses in white clover (Trifolium repens L.). Ph.D. Thesis Massey University, N.Z.
http://muir.massey.ac.nz/bitstream/handle/10179/2248/02_whole.pdf?sequence=1 (Verified 30 Jun. 2012).
Hong, V., and R.E. Wrolstad. 1990. Use of HPLC separation/photodiode array detection for characterization of anthocyanins. J. Agric. Food Chem. 38:708-71 Huang, Z.-A., D.-A. Jiang, Y. Yang, J.-W. Sun, and S.-H. Jin. 2004. Effects of nitrogen
deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica. 42:357-364.
Irani, N.G., and E. Grotewold. 2005. Light-induced morphological alteration in anthocyanin-accumulating vacuoles of maize cells. BMC Plant Bio. 5:7. Jackson, J.A., G. Fuglevand, B.A. Brown, M.J. Shaw, and G.I. Jenkins. 1995. Isolation of Arabidopsis mutants altered in the light regulation of chalcone synthase gene
expression using a transgenic screening approach. Plant J. 8:369-380.

181
Jaisingh, J., E.W.K. See-To, and K.Y. Tamin. 2006. The impact of open source software on the strategic choices of firms developing proprietary software. J. of Manage.
Info. Sys. 25:241-275. Jansen, M.A.K, B. Depka, A. Trebst, and M. Edelman. 1993. Engagement of specific
sites in the plastoquinone niche regulates degradation of the D1 protein in Photosystem II. J. Biol. Chem. 268:21246-21252.
Joliot, P., and A. Joliot. 1984. Electron transfer between the two photosystems. I. flash excitation under oxidizing conditions. Biochim. Biophys. Acta. 765:210-218. Ju, Z., Y. Duan, and Z Ju. 1999. Effects of covering the orchard floor with reflecting films on pigment accumulation and fruit coloration in `Fuji' apples. Sci. Hortic.
82:47-56. Kaiser, T., K. Emmler, B. Weisshaar, E. Schafer, and A. Batschauer. 1995. Promoter elements of the mustard CHS1 gene are sufficient for light regulation in
transgenic plants. Plant Mol. Biol. 28:219-229. Kakani, V.G., K.R. Reddy, D. Zhao, and A.R. Mohammed. 2003. Effects of ultraviolet‐B radiation on cotton (Gossypium hirsutum L.). Morph. Anat. Ann. Bot. 91:817-
826. Karageorgou, P., and Y. Manetas. 2006. The importance of being red when young: anthocyanins and the protection of young leaves of Quercus coccifera from insect
herbivory and excess light. Tree Physiol. 26:613-621. Karcher, D.E., and M.D. Richardson. 2003. Quantifying turfgrass color using digital image analysis. Crop Sci. 43:943-951. Kavanagh, T., D. O’Beirne, and T.P. Cormican. 1985. Use of the hunter colour difference meter for colour evaluation in turfgrass experiments. p. 255-261. In: F. Lemaire
(ed.) Proc. 5th Intl. Turfgrass Res. Conf., Avignon, France. 1-5 Jul.

182
Keller, W., and A.T. Bleak. 1968. Preplanting treatment to hasten germination and emergence of grass seed. J. Range Manage. 21:213-216. Kerr, J.B., and C.T. McElroy. 1993. Evidence for large upward trends of ultraviolet-B radiation linked to ozone depletion. Science 262:1032-1034. Kimura, M., D.B. Rodriguez-Amaya, and H.T. Godoy. 1990. Assessment of the saponification step in the quantitative determination of carotenoids and
provitamins A. Food Chem. 35:187-195. Kong, J.M., C. Lian-Sai, G. Ngoh-Khang, C. Tet-Fatt, and R. Brouillard. 2003. Analysis and biological activities of anthocyanins. Phytochemistry 64:923-933. Kopsell, D.A., and D.E. Kopsell. 2006. Accumulation and bioavailability of dietary carotenoids in vegetable crops. Trends Plant Sci. 11:1360-1385. Kovács, E., and Á. Keresztes. 2002. Effect of gamma and UV-B/C radiation on plant cells. Micron 33:199-210. Koyama, Y., and R. Fujii, 1999. Cis-trans carotenoids in photosynthesis: configurations, Excited–state properties and physiological functions. p.161-188. In H.A. Frank (ed) The photochemistry of carotenoids. Kluwer Publishers, Dordrecht, Netherlands. Krizek, D.T., R.M. Mirecki, and S.J. Britz. 1997. Inhibitory effects of ambient levels of solar UV-A and UV-B radiation on growth of cucumber. Phys. Planta. 100:886-
893. Krol, M., G.R. Gray, V.M. Hurry, G. Oquist, L. Malek, and N.P.A. Huner, 1995. Low Temperature stress and photoperiod effect on increased tolerance to
photoinhibition in Pinus banksia seedlings. Canadian J. Bot. 73:1119-1127. Krupa, S.V., and R.N Kickert. 1989. The Greenhouse effect: Impacts of ultraviolet-B (UV-B) radiation, carbon dioxide (CO2), and Ozone (O3) on vegetation. Environ.
Pollut. 61:263-393.

183
Kubasek, W.L., B.W. Shirley, A. McKillop, H.M. Goodman, W. Briggs, and F.M. Ausubel. 1992. Regulation of flavonoid biosynthetic genes in germinating
Arabidopsis seedlings. Plant Cell. 4:1229-1236. Lancaster, J.E., J.E. Grant, C.E. Lister, and M.C. Taylor. 1994. Skin color in apples – Influence of copigmentation and plastid pigments on shade and darkness of red
color in five genotypes. J. Am. Soc. Hortic. Sci. 119:63-69. Landry, L.G., C.C. Chapple, and R.L. Last. 1995. Arabidopsis mutants lacking phenolic sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant
Physiol. 109:1159-1166. Landschoot, P.J., and C.F. Mancino. 1997. Assessment of the minolta CR-310 chroma meter for predicting nitrogen status of Agrostis stolonifera L. Intl. Turfgrass Soc.
Res. J. 8:711-718. Landschoot, P.J., and C.F. Mancino. 2000. A comparison of visual vs. instrumental measurement of color differences in bentgrass turf. HortScience 35:914-916. Langebartels, C., M. Schraudner, W. Heller, D. Ernst, and H. Sanderman Jr. 2002. Oxidative stress and defense reactions in plants exposed to air pollutants and UV-
B radiation. p.105-137. In D. Inzé and M. Van Montagu (ed.) Oxidative Stress in Plants. Taylor and Francis New York, NY.
Larsen, S.U., and C. Andreasen. 2004. Light and heavy turfgrass seeds differ in germination percentage and mean germination thermal time. Crop Sci. 44:1710-
1720. Larsen, S.U., and B.M. Bibby. 2004. Use of germination curves to describe variation in germination characteristics in three turfgrass species. Crop Sci. 44:891-899. Lee, J., R.W. Durst, and R.E. Wrolstad. 2005. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and
wines by the pH differential method: collaborative study. J. Ass. Analyt. Com. Int. 88:1269-1278.

184
Lemaire, G., G. Escadeillas, and E. Ringot. 2005. Evaluating concrete surfaces using an image analysis process. Cons. Build. Mat. 19:604-611. Lesfrud, M.G., D.A. Kopsell, R.M. Auge, and A.J. Both. 2006. Biomass production and pigment accumulation in kale grown under increasing photoperiods. Hortscience
41:603-606. Lesfrud, M.G., D.A. Kopsell, and D.E. Kopsell. 2007. Nitrogen levels influence biomass, elemental accumulations, and pigment concentrations in spinach. J. Plant Nut.
30:171-185. Lin, C., M. Ahmad, and A.R. Cashmore. 1996. Arabidopsis cryptochrome 1 is a soluble protein mediating blue light-dependent regulation of plant growth and
development. Plant J. 5:893-902. Lohachoompol, V., G. Srzednicki, and J. Craske. 2006. The change of total anthocyanins in blueberries and their antioxidant effect after drying and freezing. J. Biomed.
Biotechnol. 5:248-252. Lois, R. 1994. Accumulation of UV-absorbing flavonoids induced by in Arabidopsis thaliana L. I. Mechanisms of UV - resistance in Arabidopsis. Planta 194:498- 503. Lubin, D., and J.E. Frederick. 1991. The ultraviolet radiation environment of the antarctic peninsula: the roles of ozone and cloud cover. J. Appl. Meteorol. 30:478-493. Lutz, M., I. Agalidis, G. Hervo, R.J. Cogdell, and F. Reiss-Husson. 1978. On the state of
carotenoids bound to reaction centers of photosynthetic bacteria: A resonance raman study. Biochim. Biophys. Acta (BBA) – Bioenergetics. 503:287-303.
Mackerness S.A-H. 2000. Plant responses to ultraviolet-B (UV-B: 280–320 nm) stress: What are the key regulators? Plant Growth Reg. 32:27-39.

185
Madronich, S., R.L. McKenzie, L.O. Björn, M.M. Caldwell. 1998. Changes in biologically active ultraviolet radiation reaching the earth's surface. J. Photochem. Photobiol. B: Biol. 46:5-19.
Maguire, J.D., and K.M. Steen. 1971. Effects of potassium nitrate on germination and respiration of dormant and non dormant Kentucky bluegrass (Poa pratensis L.)
seed. Crop Sci. 11:48-50. Maliakal, S.K, K. McDonnell, S.A. Dudley, and J. Schmitt. 1999. Effects of red to far-red ratio and plant density on biomass allocation. Int. J. Plant Sci. 160:723-733. Mancinelli, A.L., F. Rossi, and A. Moroni. 1991. Cryptochrome, phytochrome and anthocyanin production. Plant Physiol. 96:1079-1085. Mangiafico, S.S., and K. Guillard. 2005. Turfgrass reflectance measurements, chlorophyll, and soil nitrate desorbed from anion exchange membranes. Crop Sci.
45:259-265. Manney, G.L., M.L. Santee, M. Rex, N.J. Livesey, M.C. Pitts, P. Veefkind, E.R. Nash, I. Wohltmann, R. Lehmann, L. Froidevaux, L.R. Poole, M.R. Schoeberl, D.P. , J.
Davies, V. Dorokhov, H. Gernandt, B. Johnson, R. Kivi, E. Kyrö, N. Larsen, P.F. Levelt, A. Makshtas, C.T. McElroy, H. Nakajima, M.C. Parrondo, D.W. Tarasick, P. von der Gathen, K.A. Walker, and N.S. Zinoviev. 2011. Unprecedented Arctic ozone loss in 2011. Nature 478:469-475.
Massacci, A., M. Adelaide-lannelli, F. Pietrini, and F. Loreto. 1995. The effect of growth at low temperature on photosynthetic characteristics and mechanisms of
photoprotection of maize leaves. J. Expt. Bot. 46:119-127. Mathews-Roth, M.M. 1986. Carotenoids quench evolution of excited species in epidermis exposed to UV-B (290-320 nm) light. Photochem. Photobiol. 43:91-93 Matile, P., B.M.P. Flach and B.M. Eller. 1992. Autumn leaves of Ginkgo biloba L.: optical properties, pigments, and optical brighteners. Bot. Acta. 105:13-17.

186
Maxwell, K., and G.N. Johnson. 2000. Chlorophyll fluorescence – a practical guide. J. Exp. Bot. 51:659-668. McClure, J.W. 1975. Physiology and functions of flavonoids. p. 970-1055. In. J. Harborne, and T. Mabry (ed.) The flavonoids. Chapman and Hall Ltd., United
Kingdom. McCurdy, J.D., J.S. McElroy, D.A. Kopsell, C.E. Sams, and J.C. Sorochan. 2008. Effects of mesotrione on perennial ryegrass (Lolium perenne L.) carotenoid
concentrations under varying environmental conditions. J. Ag. Food Chem. 56:9133-9139.
McDonald, S., P.D. Prenzler, M. Autolovich, and K. Robards. 2001. Phenolic content and antioxidant activity of olive extracts. Food Chem. 73:73-84. McElroy, J.S., D.A. Kopsell, J.C. Sorochan, and C.E. Sams. 2006. Response of creeping bentgrass carotenoid composition to high and low irradiance. Crop Sci. 46:2606-
2612. McEwen, C.N., R.G. McKay, and B.S. Larsen. 2005. Analysis of solids, liquids, and biological tissues using solids probe introduction at atmospheric pressure on
commercial LC/MS instruments. Anal. Chem. 77:7826-7831. McKenzie, R.L. L.O. Björn, A. Bais, and M. Ilyas. 2003. Changes in biologically
active ultraviolet radiation reaching the earth's surface. Photochem. Photobiol. Sci. 2:5-1.
McLennan, A.G. 1987. DNA damage, repair, and mutagenesis. p. 135-186. In J.A. Bryant and V.L. Dunham (ed.) DNA replication in Plants. CRC Press, Boca
Raton, FL. Mehlenbacher, S.A., and M.M. Thompson. 1991. Inheritance of a chlorophyll deficiency in hazelnut. Hortscience 26:1414-1416.

187
Mendez, M., D.G. Jones, and Y. Manetas. 1999. Enhanced UV-B radiation under field conditions increases anthocyanin and reduces the risk of photoinhibition but does
not affect growth in the carnivorous plant Pinguicula vulgaris. New Phytol. 144:275-282.
Milesi, C., S.W. Running, C.D. Elvidge, J.B. Dietz, and B.T. Tuttle. 2005. Mapping and modeling the biogeochemical cycling of turf grasses in the United States.
Environ. Manage. 36:426-438. Millington, K.R., and A.L. King. 2010. Measuring colour and photostability of small fleece wool samples. Anim. Prod. Sci. 50:589-592. Miyoshi, K., and M. Mii. 1985. Ultrasonic treatment for enhancing seed germination of terrestrial orchid Calanthe discolor in a symbiotic culture. Sci. Hortic. 35:127-
130. Mohammed, A.R., E.W. Rounds, and L. Tarpley. 2007. Response of rice (Oryza sativa L.) tillering to sub-ambient levels of ultraviolet-B radiation. J. Agron. Crop
Sci.193:324-335. Moncada, M.C., S. Moura, M. João Melo, A. Roque, C. Lodeiro, and F. Pina. 2003.
Complexation of aluminum (III) by anthocyanins and synthetic flavylium salts: A source for blue and purple color. Inorganica Chimica Acta. 356:51-61.
Moradas-Fereira, P.V., P.P. Costa, and W. Mager. 1996. The molecular defenses against reactive oxygen species in yeast. Mol. Microbiol. 19:651-658. Morales, L.O., R. Tegelberg, M. Brosché, A. Lindfors, S. Siipola, and P.J. Aphalo. 2011. Temporal variation in epidermal flavonoids due to altered solar UV radiation is
moderated by the leaf position in Betula pendula. Phys. Planta. 143:1399-3054. Moran, R. 1982. Formulae for determination of chlorophyllous pigments extracted with N,N- dimethylformamide. Plant Physiol. 69:1376-1381.

188
Moran, R., and D. Porath. 1980. Chlorophyll determination in intact tissues using N,N – dimethylformamide. Plant Physiol. 65:478-479. Mori, M., K. Yoshida, Y. Ishigaki, T. Matsunaga, O. Nikaido, K. Kameda, and T. Kondo. 2005. UV-B protective effect of a polyacylated anthocyanin, HBA, in flower
petals of the blue morning glory, Ipomoea tricolor cv. Heavenly Blue. Bioorg. Med. Chem. 13:2015-2020.
Moros, E.E., D. Dsrnoko, M. Cheryan, E.G. Perkins, and J. Jerrell. 2002. Analysis of xantophylls in corn by HPLC. J. Agric. Food Chem. 50:5787-5790. Müller, P., L. Xiao-Ping, and K.K. Niyogi. 2001. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 125:1558-1566. Munsell, A.H. 1915. Atlas of the munsell color system. Wadsworth-Howland and Company, Malden, MA. Mustafa, N.R., and R.Verpoorte. 2007. Phenolic compounds in Catharanthus roseus. Phytochem. Rev. 6:243-258. Neill, S., and K.S. Gould. 1999. Optical properties of leaves in relation to anthocyanin concentration and distribution. Can J. Bot. 77:1777-1782. Nickerson, D. 1940. History of the munsell color system and its scientific application. J. Optic. Soc. Am. 30:575-586. Noble, R.E. 2002. Effects of UV-irradiation on seed germination. Sci. Total Environ. 299:173-176. Nutter, Jr., F.W., M.L. Gleason, J.H. Jenco, and N.C. Christians. 1993. Assessing the accuracy, intra-rater repeatability and inter-rate repeatability of disease
assessment systems. Phytopatholgy 83:806-812.

189
Oren-Shamir, M., and A. Levi-Nissim. 1997. UV-light effect on the leaf pigmentation of Cotinus coggygria ‘Royal Purple’. Sci. Hortic. 71:59-66. Owens, T.G., A.P. Shreve, and A.C. Atbrecht. 1992. Dynamics and mechanisms of
singlet energy transfer between carotenoids and chlorophylls: light harvesting and nonphotochemical fluorescence quenching. p. 170-186. In N. Murata (ed.) In Research in Photosynthesis 4. Kluwer, Dordrecht, Netherlands.
Pascual-Teresa, S. de., C. Santos-Buelga, and J.C. Rivas-Gonzalo. 2002. LC–MS analysis of anthocyanins from purple corn cob. J. Sci. Food and Agric. 82:1003-1006. Pattee, H.E., F.G. Giesbrecht, and C.T. Young. 1991. Comparison of peanut butter color determination by CIELAB L*, a*, b* and Hunter color-difference methods
and the relationship of roasted peanut color to roasted peanut flavor response. J. Agric. Food Chem. 39:519-523.
Pfeiffer, N.E. 1928. Anatomical study of plants grown under glasses transmitting light of various ranges of wavelength. Bot. Gaz. 85:427-436. Pfeifer, M.T. P. Koepke, and J. Reuder. 2006. Effects of altitude and aerosol on UV
radiation. J. Geophys. Res. 111. D01203. doi:10.1029/2005JD006444. Pietrini, F., and A. Massacci. 1998. Leaf anthocyanin content changes in Zea mays grown at low temperature: Significance for the relationship between quantum yield of PS
II and the apparent quantum yield of CO2 assimilation. Photosynth. Res. 58:213-219.
Pill, W.G., J.J. Frett, and I.H. Williams. 1997. Matric priming of Kentucky bluegrass and tall fescue seeds benefits seedling emergence. HortScience 32:1061-1063. Piringer, A.A., and P.H. Heinze. 1954. Effect of light on the formation of a pigment in the tomato fruit cuticle. Plant Physiol. 29:467-472.

190
Pons, T.L., H. van Rijnberk, I. Scheurwater, and A. van der Werf. 1993. Importance of the gradient in photosynthetically active radiation in a vegetation stand for leaf
nitrogen allocation in two monocotyledons. Oecologia 95:416-424. Popp, H.W., and F. Brown. 1936. The effect of ultra- violet radiation upon seed plants. p. 853-887. In B.M. Dugger (ed.) Biological effects of radiation. Vol. 2. McGraw-
Hill Book Co., New York. Pourmorad, F., S.J. Hosseinimehr, and N. Shahabimajd. 2006. Antioxidant activity, phenol and flavonoid contents of some selected Iranian medicinal plants. Afric. J.
Biotechnol. 5:1142-1145. R Development Core Team. (2011). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-
900051-07-0. Available at http://www.R-project.org/ (Verified 29 Jun. 2012). Rabino, I., and A.L. Mancinelli. 1986. Light, temperature and anthocyanin production. Plant Physiol. 81:922-924. Rademacher, I.F., and C.J. Nelson. 2001. Nitrogen effects on leaf anatomy within intercalary meristems of tall fescue leaf blades. Ann. Bot. 88:893-903. Reinhard, E., M. Adhikkmin, B. Gooch, and P. Shirley. 2001. Color transfer between images. Comp. Graphic App. 21:34-41. Richardson, M.D., D.E. Karcher, and L.C. Purcell. 2001. Quantifying turfgrass cover using digital image analysis. Crop Sci. 41:1884-1888. Rind, D., R. Suozzo, N.K. Balachandran, and M.J. Prather. 1990. Climate change and the middle atmosphere. I: The doubled CO2 climate. J. of Atmos. Sci. 47:475-494. Ronen, G., M. Cohen, D. Zamir, and J. Hirschberg. 1999. Regulation of carotenoid biosynthesis during tomato fruit development: expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant
delta. Plant J. 17:341-351.

191
Ronen, G., M. Cohen, D. Zamir, and J. Hirschberg. 2000. An alternative pathway to β- carotene formation in plant chromoplasts discovered by map-based cloning of beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. U.S.A.
97:11102-11107. Roorda, A., and D.R. Williams. 1999. The arrangement of the three cone classes in the living human eye. Nature 397:520-522. Rosario, R., G.J. Mark, J.A. Parrish, and M.C. Mihm Jr. 1979. Histological changes produced in skin by equally erythemogenic doses of UV-A, UV-B, UV-C and
UV-A with psoralens. Brit. J. Derm. 101:299-308. Rowland, F.S. 1990. Stratospheric ozone depletion by chlorofluorocarbons. Ambio 19:281-292. Rowland, F.S. 2006. Stratospheric ozone depletion. Philos. Trans. R. Soc. London, Ser. B. 361:769-790. Rozema, J., J. van de Staaij, L.O. Björn, and M. Caldwell. 1997. Review. UV-B as an
environmental factor in plant life: stress and regulation. Trends in Ecol. 12:22–28. Rozema, J., R. Broekman, D. Lud, H.J. Huiskes, T. Moerdijk, N. de Bakker, B. Meijkamp, and A. van Beem. 2001. Consequences of depletion of stratospheric
ozone for terrestrial Antarctic ecosystems: the response of Deschampsia antarctica to enhanced UV-B radiation in a controlled environment. Plant Ecol. 154:103-115.
Sancar, A., and G.B. Sancar. 1988. DNA repair enzymes. Ann. Rev. Biochem. 57:29-67. Sarkar, D., P.C. Bhowmik, Y-I. Kwon, and K. Shetty. 2011. The role of proline- associated pentose phosphate pathway in cool-season turfgrasses after UV-B
exposure. Environ. Exp. Bot. 70:251-258. SAS Institute. 2011. Version 9.2 SAS Inst. Cary, NC.

192
Saxby, M.J. 1964. Characterization of flavonoids and coumarins by their absorption spectra on paper chromatograms. Anal. Chem. 36:1145-1146. Schmidt, R.E., and X. Zhang. 2001. Alleviation of photochemical activity decline of turfgrasses exposed to soil moisture stress or UV radiation. Int. Turfgrass Soc. Res. J. 9:340-346. Schmitt, J., and R.D. Wulff. 1993. Light spectral quality, phytochrome and plant competition. Trends in Ecol. Evol. 8:47-51. Seigler, D.S. 1998. Plant secondary metabolism. Kluwer Publishers, Norwell, MA. Shackleton, C. 2010. Clinical steroid mass spectrometry: A 45-year history culminating in HPLC–MS/MS becoming an essential tool for patient diagnosis. J. Ster.
Biochem. Mol. Biol. 121:481-490. Shaidaee, G, B.E. Dahl, and R.M. Hansen. 1969. Germination and emergence of different age seeds of six grasses. J. of Range Manage. 22:240-243. Sharma, G., and H.J. Trussell. 1997. Digital color imaging. Trans. Image Proc. 6:901- 932. Sheahan, J.J. 1996. Sinapate esters provide greater UV-B attenuation than flavonoids in Arabidopsis thaliana (Brassicaceae). Am. J. Bot. 83:679-686. Sheehy, J.E. 1975. Some optical properties of leaves of eight temperate forage grasses. Ann. Bot. 39:377-386. Shimada, K., Y. Kakehashi, H. Matsumura, and N. Tanoue. 2004. In vivo quantitative evaluation of tooth color with hand-held colorimeter and custom template. J.
Prosth. Dent. 91:389-391. Shinomura, T., A. Nagatani, H. Hanzawa, M. Kubota, M. Watanabe, and M. Furuya. 1996. Action spectra for phytochrome A and B-specific photoinduction of seed
germination in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U.S.A. 15:8129-8133.

193
Skogley, C.R., and F.B. Ledeboer. 1968. Evaluation of several Kentucky bluegrass and red fescue strains maintained as lawn turf under three levels of fertility. Agron. J.
60:47-49. Smith, H. 1982. Light quality, photo perception and plant strategy. Ann. Rev. Plant Physiol. 33:481-518. Smith, H. 1994. Sensing the light environment: the functions of the phytochrome family. p. 377-416. In J.R.E. Kendrick and G.H Kronenberg (ed.) Photomorphogenesis in
Plants. Kluwer Academic Publishers, Dordrecht, The Netherlands. Smith, J.L., D.J. Burritt, and P. Bannister. 2000. Shoot dry weight, chlorophyll and UV-
B-absorbing compounds as indicators of a plant's sensitivity to UV-B radiation. Annal. Bot. 86:1057-1063.
Solomon, S. 1990. Progress towards a quantitative understanding of Antarctic ozone
depletion. Nature 347:347-354 Sponga, F., G.F. Deitzer, and A.L. Mancinelli. 1986. Cryptochorme, phytochrome and the photoregualtion of anthocyanin production under blue light. Plant Physiol.
82:952-955. Stapleton, A.E. 1992. Ultraviolet radiation and plants: Burning questions. Plant Cell. 4:1353-1358. Stiegler, J.C., G. Bell and N. Maness. 2004. Comparison of acetone and N,N- dimethylformamide for pigment extraction in turfgrass. Comm. Soil Sci. Plant
Anal. 35:1801-1813. Stiegler, J.C., G.E. Bell, N.O. Maness, and M.W. Smith. 2005. Spectral detection of pigment concentrations in creeping bentgrass golf greens. Int. Turf. Res. J.
10:818-825.

194
Stiff, M.L., and J.B. Powell. 1974. Stem anatomy of turfgrass. Crop Sci. 14:181-186. Strid, Å., W.S. Chow, and J.M. Anderson. 1990. Effects of supplementary ultraviolet – B radiation on photosynthesis in Pisium sativum. Biophys. Biochim. Acta.
1020:260-268. Strid, Å., and R.J. Porra. 1992. Alterations in pigment content in leaves of Pisium
sativum after exposure to supplementary UV-B. Plant Cell Physiol. 33:1015-1023. Strid, Å., W.S. Chow, and J.M Anderson. 1994. UV-B damage and protection at the
molecular level in plants. Photosynth. Res. 39:475-489. Sullivan, J.H., and A.H. Teramura. 1988. Effects of ultraviolet-B irradiation on seedling growth in the Pinaceae. Am. J. Bot. 75:225-230. Taalas, P., J. Kaurola, A. Kylling, D. Shindell, R. Sausen, M. Dameris, V. Grewe, J. Herman, J. Damski, and B. Steil. 2000. The impact of greenhouse gases and
halogenated species on future solar UV radiation doses. Geophys. Res. Lett. 27:1127-1130.
Takabe, T., A. Uchida, and T. Tetsuko. 2004. Effects of hydrogen peroxide on germination of Kentucky bluegrass. In Abstracts, Abs # 709. Plant Biology 2004:
July 24 – July 28, 2004. Lake Buena Vista, FL. Takashi, S., and N. Murata. 2008. How do environmental stresses accelerate photoinhibition? Trends Plant. Sci. 13:178-182. Teramura, A.H., R. Hilton Biggs, and S. Kossuth. 1980. Effects of ultraviolet-B
irradiances on Soybean II. Interaction between ultraviolet-B and photosynthetically active radiation on net photosynthesis, dark respiration, and transpiration. Plant Physiol. 65:483-488.
Teramura, A.H., L.H. Ziska, and A.E. Sztein. 1991. Changes in growth and photosynthetic capacity of rice with increased UV-B radiation. Physiol. Planta.
83:373-380.

195
Teramura, A.H., and J.H. Sullivan. 1994. Effects of UV-B radiation on photosynthesis and growth of terrestrial plants. Photosynth. Res. 39:463-473. Tevini, M., W. Iwanzik, and U. Thoma. 1981. Some effects of enhanced UV-B irradiation on the growth and composition of plants. Planta 153:388-394. Tevini, M. 1993. Effects of enhanced UV-B radiation on terrestrial plants. p. 125-153. In Tevini M. (ed.) UV-B Radiation and Ozone Depletion. Lewis Publ., Boca Raton,
FL. Thomas, D.A., and H.N. Barber. 1974. Studies on leaf characteristics of a cline of Eucalyptus urnigera from Mount Wellington, Tasmania. II. Reflection,
transmission and absorption of radiation. Aust. J. Bot. 22:701-707. Toole, V.K., and H.A. Borthwick. 1971. Effect of light, temperature and their interactions on seeds of Kentucky bluegrass (Poa pratensis L.). J Am. Soc. Hortic. Sci.
96:301-304. Torres, M., G. Frutos, and J.M. Duran. 1991.Sunflower seed deterioration from exposure to UV-C radiation. Environ. Exp. Bot. 31:201-207. Tyystjärvi, E. 2008. Photoinhibition of photosystem II and photodamage of the oxygen evolving manganese cluster. Coordination Chemistry Reviews. The Role of
Manganese in Photosystem II. 252:361-376. Vass, I., L. Sass, C. Spetea, A. Bakou, D.F. Ghanotakis, and V. Petroulea. 1996. UV-B- induced inhibition of photosystem II electron transport studied by EPR and
chlorophyll fluorescence. Impairment of donor and acceptor side components. Biochemistry 35:8964-8973.
Wade, H.K., T.N. Bibikova, W.J. Valentine, and G.I. Jenkins. 2001. Interactions within a network of phytochrome, cryptochrome and UV-B phototransduction pathways
regulate chalcone synthase gene expression in Arabidopsis leaf tissue. Plant J. 25:675-685.

196
Wagné, C. 1966. Effect of UV light on lettuce seed germination and on the unfolding of grass leaves. Physiol. Planta. 19:128-133. Wang, Y., M. Zhang, X.Y. Zhang, and H.D. Dong. 2002. Comparative studies on elasticity and scrape tolerance characters of Zoysia japonica and Poa pratensis.
Prataculture Sci. 19:56-59. Wellburn, A.R. 1994. The spectral determination of chlorophylls a and b, as well as total carotenoids using various solvents with spectrophotometers of different
resolution. J. Plant Physiol. 144:307-313. Wherley, B.G., D.S. Gardner, and J.D. Metzger. 2005. Tall fescue photomorphogenesis as influenced by changes in the spectral composition and light intensity. Crop
Sci. 45:562-568. White, A.L., and L.S. Jahnke. 2002. Contrasting effects of UV-A and UV-B on photosynthesis and photoprotection of β-carotene in two Dunaliella spp. Plant
Cell Physiol. 43:877-884. Woodall, G.S., and G.R. Stewart. 1998. Do anthocyanins play a role in UV protection of the red juvenile leaves of Syzygium? J. Exp. Bot. 49:1447-1450. Wrolstad, R.E., and M.M. Giusti. 2001. Characterization and measurement of anthocyanins by UV-visible spectroscopy. Unit F1.2. p. F1.2.1-F1.2.13. In R.E.
Wrolstad and S.J. Schwartz (ed.) Current Protocols in Food Analytical Chemistry. John Wiley & Sons, Inc. New York, NY.
Yin, Z-H., S. Neimanis, U. Wagner, and U. Hebe. 1990. Light-dependent pH changes in leaves of C3 plants I. Recording pH changes in various cellular compartments by
fluorescent probes. Planta 182:244-252. Yudin, L.S., C.W. Mitchell, and J.J Cho. 1987. Color preference of thrips (Thysanoptera: Thripidae) with reference to aphids (Homoptera: Aphididae) and leafminers in
Hawaiian lettuce farms. J. Econo. Entomol. 80:51-55.

197
Zhang, S., H. Jiang, S. Peng, H. Korpelainen, and C. Li. 2011. Sex-related differences in morphological, physiological, and ultrastructural responses of Populus cathayana
to chilling. J. Exp. Bot. 62:675-686. Zhang, X., E.H. Ervin, and R.E. Schmidt. 2005. The role of leaf pigment and antioxidant levels in UV-B resistance of dark- and light-green Kentucky bluegrass cultivars.
J. Am Soc. HortScience 130:836-841. Zhang, X., and E.H. Ervin. 2005. Exogenous salicylic acid and jasmonic acid effects on Kentucky bluegrass UV-B tolerance. Int. Turfgrass Soc. Res. J. 10:910-915. Zhang, X., and E.H. Ervin. 2009. Physiological assessment of cool-season turfgrasses under ultraviolet-B stress. HortScience. 44:1785-1789. Ziska, L.H, A.H. Teramura, and J.H. Sullivan. 1992. Physiological sensitivity to plants along an elevation gradient to UV-B radiation. Am. J. Bot. 79:863-871. Ziska, L.H., A.H. Teramura, J.H. Sullivan, and A. McCoy. 1993. Influence of ultraviolet- B (UV-B) radiation on photo-synthetic and growth characteristics in field-grown
cassava (Manihot esculentum Crantz). Plant Cell Environ. 16:73-79. Zscheile, F.P., J.W. White, Jr., B.W. Beadle, and J.R. Roach. 1942. The preparation and
absorption spectra of five pure carotenoid pigments. Plant Physiol. 17:331-346. Zu-Hua, Y., K. Siebke, and U. Heber. 1990. Light dependent changes in leaves of C3 plants. III. Effect of inhibitors of photosynthesis and of developmental state of
photosynthetic apparatus on cystolic and vacuolar pH changes. Planta 182:262-26.