Embed Size (px)
Citation preview

Ultrastructure and Lectin Cytochemistry of the Cloacal VentralGlands in the Male Newt Triturus marmoratus marmoratusE. ROMO, R. PANIAGUA,* B. FRAILE, AND M.P. DE MIGUELDepartment of Cell Biology and Genetics, University of Alcala, E-28871 Alcala de Henares, Madrid, Spain
ABSTRACT Ventral glands are found in the cloacal walls of male urodele amphibians except forsirenids. These glands are mucous, and secrete substances that will form part of the spermatophoreused in transfer of sperm during fertilization. Ventral glands are formed by secretory and ductalportions; both possess epithelial and myoepithelial cells with different characteristics. Urodeleshave cyclic reproduction, and cloacal ventral glands show seasonal differences with electronmicroscopy. The glycoproteins secreted by these glands have been studied by means of lectinhistochemistry. The labeling was detected mainly in the nuclei, rough endoplasmic reticulum, Golgicomplex, and cytosol. Secretory granules in these glands are composed by mucous glycoproteins thatbind PNA lectin (which binds galactose) and SBA and HPA lectins (N-acetylgalactosamine), UEA-I(fucose), and LcA (glucose and/or mannose). These findings suggest that the mucins secreted byventral glands contain both N- and O-linked oligosaccharides. Ventral glands secrete higherquantity and more diverse mucous substances in the reproductive period, as confirmed bylectin-histochemical reactions. Based on these results, the major similarity between ventral cloacalglands and accessory mammalian glands, can be established with bulbourethral glands. Microsc.Res. Tech. 45:122–129, 1999. r 1999 Wiley-Liss, Inc.
INTRODUCTIONThe cloacal glands of male urodele amphibians play
an important role in reproduction. These glands areresponsible for the secretion of several substances thatsurround the sperm and form the spermatophore forfemale fertilization (Russell et al., 1981; Sever andHouck, 1985; Zalisko et al., 1984), and act as phero-mones for mating (Malacarne et al., 1984; Malacarneand Giacoma, 1986; Sever et al., 1990).
There have been several light microscopical studiesconcerning the cloacal glands of Salamandridae (Sever,1981, 1991, 1992), including those of the genus Triturus(Romo et al., 1996; Sever et al., 1990). In these studies,four main types of cloacal glands have been distin-guished: pelvic, dorsal, ventral, and Kingsbury’s glands(Romo et al., 1996; Sever, 1992).
Ventral glands are a large group of glands that occuralong the entire length of the inferior border of thecloaca. These glands are formed by a basophilic flat-tened epithelium surrounded by thin myoepithelialcells. The glands secrete into the cloacal chamberthrough tubular ducts, which are composed of thinepithelial cells and thick myoepithelial cells.
In urodeles reproduction is cyclic, and usually takesplace in spring (Geniez and Grillet, 1990). The reproduc-tive cycle displays testicular changes (Saez et al., 1989;Tanaka and Iwasawa, 1979), which are associated withchanges in the morphology and secretion of the cloacalglands (Romo et al. 1996; Sever et al., 1990). In theseglands, seasonal differences have been observed. In thereproductive period, the total glandular volume and thediameter of the tubular lumen increase significantly,although the epithelial height does not show differencesduring the reproductive cycle (Romo et al., 1996).
Glycoproteins generally form part of secretions andalso can act as structural proteins in membranes
(Rademacher et al., 1988). Glycoproteins are also pre-sent in the cytosol and nucleoplasm, but their functionin these areas is not yet completely elucidated (Halti-wanger et al., 1990). The oligosaccharides incorporatedinto glycoproteins can be divided in two groups: N-oligosaccharides and O-oligosaccharides, depending onthe amino acid that binds the sugar residue (Kornfeldand Kornfeld, 1985; Strouss and Dekker, 1992).
Ultrastructural studies on the cloacal glands of malesalamandrids have been done in the species Salamand-rina terdigitata (Brizzi et al., 1986, 1988, 1990) butthere were not ultramicroscopic works in Triturusmarmoratus marmoratus. A preliminary lectin histo-chemistry study (at light microscopy level) demon-strated very poor affinity for any lectin examined in theventral glands of T. marmoratus (Romo et al., 1996). Inthis study, we attempted the ultrastructural character-ization of the ventral glands of Triturus marmoratusmarmoratus by conventional methods and by lectincytochemistry.
MATERIALS AND METHODSMaterials
Sixteen male marbled newts (Triturus marmoratusmarmoratus Latreille) were collected from ponds of theProvince of Leon, in Spain (42o 30’N, 5o 45’W; altitude800 m). Eight animals were captured in February, atthe beginning of the reproductive period, and the othereight newts in August, when the reproductive periodhad ended (non-reproductive period). The animals wereanaesthetized with 0.1% methanesulphonate (MS-222;
*Correspondence to: Dr. R. Paniagua, Department of Cell Biology and Genetics,University of Alcala, E-28871 Alcala de Henares, Madrid, Spain.
Received 8 September 1998 ; accepted in revised form 22 December 1998
MICROSCOPY RESEARCH AND TECHNIQUE 45:122–129 (1999)
r 1999 WILEY-LISS, INC.

Sandoz Barcelona, Spain), killed, and their cloacaewere removed.
Conventional Electron MicroscopyThe cloacae were fixed for 6 hours in a 0.1 M
phosphate-buffered mixture of 3% paraformaldehydeand 1% glutaraldehyde at pH 7.4. Afterwards, thematerial was washed, postfixed with osmium tetroxide,dehydrated, and embedded in either epoxy resin. Sec-tions (1-µm-thick) were stained with toluidine blue.Ultrathin sections were double stained with uranylacetate and lead citrate.
Lectin HistochemistryThe cloacae were fixed for 6 hours in a mixture of
2.5% paraformaldehyde and 0.5% glutaraldehyde inequal proportions, at pH 7.4. Afterwards, tissues wererinsed in phosphate buffer, dehydrated, and embeddedin Lowicryl K4M. Semithin sections were stained withtoluidine blue. Ultrathin sections were placed on nickelgrids and processed by means of cytochemical lectingold method (Roth, 1983). In brief, grids were placed ondrops of 0.2 M Tris buffer (pH 7.6) containing 0.1%glycine and 2% BSA for 30 minutes. Afterwards, theywere incubated with different lectin gold complexes atconcentration of 10–20µg/ml in Tris buffer containingBSA (2%) for 1 hour at room temperature. The lectingold complexes used were (Table 1): wheat germ agglu-tinin (Triticum vulgaris, WGA); concanavalin A (Cana-valia ensiformis, ConA), Helix pomatia agglutinin(HPA), Ulex europaeus agglutinin I (UEA-I), peanutagglutinin (Arachis hypogaea, PNA), soybean aggluti-nin (Glycine max, SBA) and Lens culinaris agglutinin(LcA). All lectin gold complexes tested were supplied bySigma (Barcelona, Spain). The size of gold particles was10 nm in all cases except for ConA and HPA (20 nm).
For negative controls, the following competing sugarswere added at a final concentration of 2 mg/ml to therespective lectin solutions 1 hour prior to incubation:N-acetyl-D-glucosamine for WGA, D-mannose for ConA,N-acetyl-D-galactosamine for HPA, L-fucose for UEA-I,D-galactose for PNA, N-acetyl-D-galactosamine for SBA,and D-glucose for LcA. Grids were then incubated aspreviously described. All grids were stained with uranylacetate and examined in a Zeiss (Thornwood, NY) EM10C transmission electron microscope.
To evaluate the staining intensity of the differentlectins, the average number of colloidal gold particlesper cell surface area and per extracellular space (back-ground reaction) was calculated for each subject in 8–16electron-micrographs (at a final magnification of330,000) of 5–12 non-consecutive ultrathin sections, togive the density expressed as gold particles per µm2 ofcytoplasmic surface. The number of sections and micro-graphs of each sections that should be used for calcula-tions were determined for each subject by successiveapproaches to obtain the minimum numbers requiredto reach the lowest standard deviations (SD) (Lucocq,1994).
RESULTSUltrastructural Study
Reproductive period. In the marbled newt, theventral glands were composed of tubulo-alveolar glandsshowing two well-differentiated portions: a distal secre-
tory region and a proximal ductal portion (Fig. 1). Theepithelial cells of the secretory portion formed a lowcolumnar epithelium. These cells possessed a hetero-chromatic nucleus, located near the basement mem-brane, and a very electron-dense cytoplasm that wasalmost filled by clear secretory granules containing afibrous material (Fig. 2). The granules varied widely insize and frequently fused one to another.
Cytoplasmic organelles occupied a small part of thecell. The most conspicuous organelles were the roughendoplasmic reticulum, which displayed parallely ar-ranged cisternae occupying the basal and lateral cyto-plasm (Fig. 3), elongated mitochondria with a electron-dense matrix, a well-developed Golgi apparatus (Fig.4), scanty smooth endoplasmic reticulum tubules, andsome lysosomes, which appeared only occasionally andwere more electron-dense than the secretory granules.
Lateral plasma membranes of adjacent cells werejoined by junctional complexes and interdigitations.The apical cell surface presented microvilli, mainlynear the cell borders (Fig. 2). The glandular lumencontained the same type of fibrous material as thesecretory granules.
The epithelium was surrounded by a discontinuouslayer of elongated, flattened myoepithelial cells. Thesecontained a large amount of filaments, small mitochon-dria, a few rough endoplasmic reticulum cisterna, andsome smooth endoplasmic reticulum tubules.
The epithelial cells lining the ductal portion wereflattened and their cytoplasm possessed small emptyvesicles and scanty organelles, mainly mitochondria,and both rough and smooth endoplasmic reticulum(Fig. 5).
The ductal epithelium was completely surrounded bymyoepithelial cells that were much thicker than thoseof the glandular portion. The ductal myoepithelial cellsshowed a crescent-shaped nucleus and a clear cyto-plasm that was nearly filled by filaments and mitochon-dria (Fig. 5). Numerous electron-dense bodies could beseen in the cytoplasm of myoepithelial cells, mainlybeneath the plasma membrane (Fig. 6). Adjacent cellswere joined by desmosomes and interdigitations (Figs.7 and 8).
Non-reproductive period. The ultrastructuralcharacteristics of the glandular cells changed in thenon-reproductive period. The most striking differenceswere that the glandular lumen appeared nearly oc-cluded and that the cytoplasm of epithelial cells waslacking in secretory granules (Fig. 9). The most abun-dant organelles were lysosomes, including multivesicu-lar bodies (Figs. 10 and 11). Myoepithelial cells in thesecretory portion are more concentrated than in thereproductive period but their ultrastructural character-istics have not changed.
The ductal portion showed a completely occludedlumen. The ductal epithelial cells presented a big andirregular nucleus and scanty cytoplasm. Myoepithelialcells were irregular in shape and showed the sameultrastructural features as in the reproductive period.
Lectin CytochemistryReproductive period. The results of lectin cyto-
chemistry in the reproductive period are summarizedin Table 1. The labeling pattern of the glandularepithelial cells was as follows: The nuclei labeled to
123CLOACAL VENTRAL GLANDS IN THE MALE NEWT

WGA, ConA, LcA, PNA, and SBA lectins (Fig. 12).Cytosolic reaction was observed for PNA, SBA, andHPA (Fig. 13). The rough endoplasmic reticulum la-beled to WGA, ConA, and LcA lectins (Fig. 14), and theGolgi complex and secretory granules labeled to LcA,UEA-I, PNA, SBA, and HPA (Figs. 15 and 16). Therewere marked differences among granules in the label-ing to PNA, SBA, and HPA lectin: the number of goldparticles was higher in the granules located in thecenter of the cell and basal cytoplasm than in thoselocated in the apical cell portion (Table 1).
The epithelial cells of the ducts and the myoepithelialcells of both glandular and the ductal portions wereonly labeled in their nucleus. This labeling was for thesame lectins as the epithelial glandular cells (except forLcA), although the density of gold particles was lowerthan that of these cells (Fig. 17).
Non-reproductive period. The results obtained inthe non-reproductive period are shown in Table 2. Theglandular epithelial cells were only labeled in theirnucleus and cytosol. Nuclear labeling was to the samelectins as in the reproductive period, except for PNA,and weaker than that recorded in the reproductiveperiod. Cytosolic staining was only observed for SBA.The labeling pattern of the ductal epithelial cells in thenon-reproductive period was similar to that presentedin the reproductive period, except for the absence ofnuclear ConA labeling, and the presence of cytosolicSBA labeling. The labeling pattern of myoepithelialcells in both glandular portion and ducts only differedfrom that of these cells in the reproductive period in theappearance of cytosolic labeling to both PNA and SBAlectins.
DISCUSSIONThe ultrastructural characteristics of the glandular
epithelial cells of the ventral glands of Triturus marmo-ratus marmoratus in the reproductive period are notdifferent from those of a typical secretory cell; this is,the cytoplasm of these cells exhibited abundant roughendoplasmic reticulum, a well-developed Golgi com-plex, and numerous secretory vesicles. These vesicleswill transport the material to the spermatophore, be-cause the secretion of these glands forms the base of thespermatophore (Sever and Houck, 1985).
The ductal portion of ventral glands has thin epithe-lial cells and thick myoepithelial cells. This observationsuggests that ducts are structures that do not contrib-ute to ventral glands secretion but to fluid motilitythroughout myoid cell contraction.
Several authors have reported that the cloaca is anandrogen-dependent organ (Norris and Moore, 1975;Norris et al., 1989). Both cloacal size (Sever et al., 1990)and blood androgen levels (Norris et al., 1989; Romo etal., 1996) are maximal in the period of the year previousto reproduction, and reach their minimal volume afterthe breeding season. After the breeding season, theepithelial cells of the secretory portion lose their secre-tory granules, and the cytoplasm develops multiple,large lysosomes. These organelles have also been ob-served in mammalian epididymal cells and have beenrelated to the resorptive ability of these cells (Friend,1969; Moniem and Glover, 1972). The lysosomes pres-ent in the ventral glands in this period of the year might
TA
BL
E1.
Lec
tin
cyto
chem
istr
yof
the
ven
tral
glan
ds
inth
ere
prod
uct
ive
peri
od*
Lec
tin
Car
boh
ydra
tebi
ndi
ng
Bac
kgro
un
dre
acti
on
Gla
nds
Du
cts
Epi
thel
ialc
ells
Myo
epit
hel
ialc
ells
Epi
thel
ialc
ells
Myo
epit
hel
ial
cell
sS
G
NC
RE
RG
Su
pran
uc
Infr
anu
cN
CN
CN
C
WG
A(G
lcN
Ac)
n.
Neu
Ac
166
449
86
184
—49
612
——
—28
86
125
—72
69
—28
46
81—
Con
Aa
-D-M
an.
a-D
-Glc
326
790
56
265
—63
67
——
—36
86
86—
243
620
—33
86
67—
LcA
a-D
-Man
.a
-D-G
lc12
63
426
5—
296
1119
68
356
1339
615
——
——
——
UE
A-I
a-L
-Fu
c11
63
——
—25
69
102
650
986
41—
——
——
—P
NA
D-G
al-(
b1=
3),D
Gal
NA
c12
63
100
640
516
46—
216
952
612
986
29—
—33
69
—31
611
—S
BA
a,b
-D-G
alN
Ac
156
440
36
8310
06
44—
246
1154
610
114
632
131
624
—80
620
—10
36
28—
HPA
a-D
-Gal
NA
c10
63
—35
620
—19
67
236
873
636
——
——
——
*Dat
aar
eex
pres
sed
asn
um
ber
ofgo
ldpa
rtic
les
per
10µ
m2
(mea
n6
SD
)in
the
cell
ula
rlo
cati
onm
enti
oned
.Gal
:Gal
acto
se.G
lcN
Ac:
N-A
cety
lglu
cosa
min
e.M
an:m
ann
ose.
Glc
:glu
cose
.Gal
NA
c:N
-ace
tylg
alac
tosa
min
e.F
uc:
fuco
se.N
:nu
cleu
s.C
:cyt
osol
.RE
R:r
ough
endo
plas
mic
reti
culu
m.G
:Gol
gico
mpl
ex.S
G:s
ecre
tory
gran
ule
s.S
upr
anu
c:su
pran
ucl
ear
cyto
plas
m(i
ncl
udi
ng
the
hig
her
hal
fof
the
nu
cleu
s).I
nfr
anu
c.:i
nfr
anu
clea
rcy
topl
asm
(in
clu
din
gth
elo
wes
th
alfo
fth
en
ucl
eus)
.Val
ues
that
did
not
diff
erfr
omba
ckgr
oun
dva
lues
are
omit
ted
(—).
124 E. ROMO ET AL.
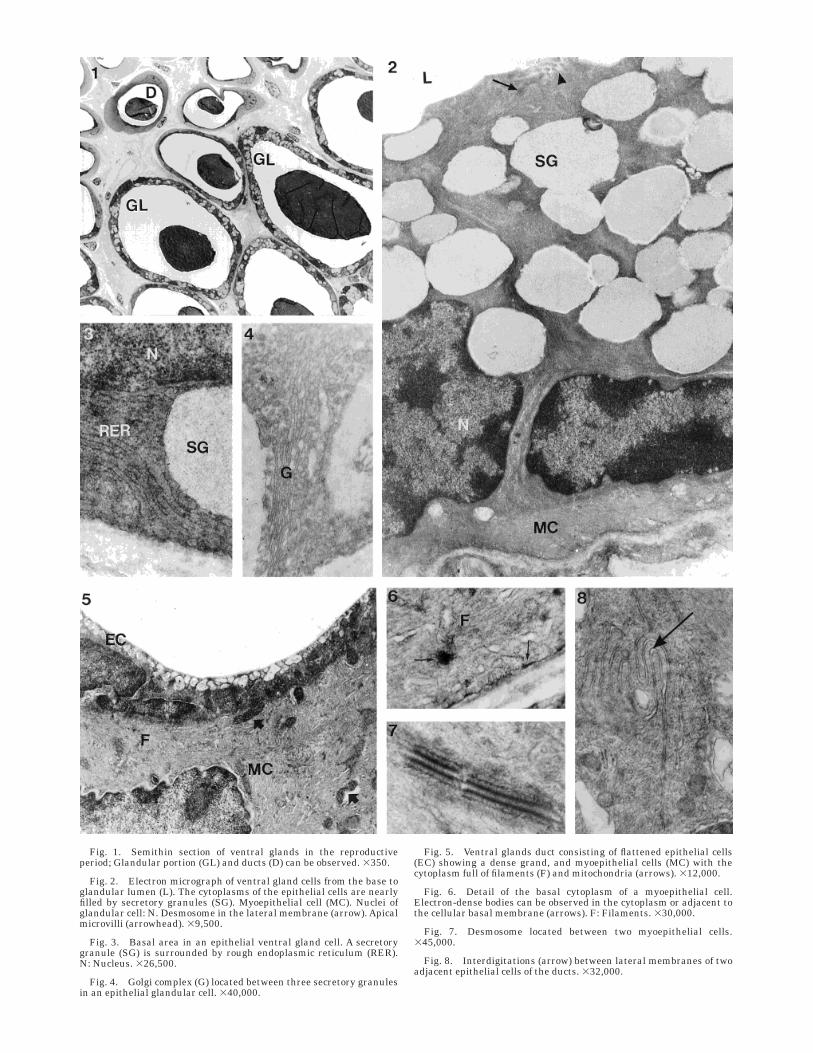
Fig. 1. Semithin section of ventral glands in the reproductiveperiod; Glandular portion (GL) and ducts (D) can be observed. 3350.
Fig. 2. Electron micrograph of ventral gland cells from the base toglandular lumen (L). The cytoplasms of the epithelial cells are nearlyfilled by secretory granules (SG). Myoepithelial cell (MC). Nuclei ofglandular cell: N. Desmosome in the lateral membrane (arrow). Apicalmicrovilli (arrowhead). 39,500.
Fig. 3. Basal area in an epithelial ventral gland cell. A secretorygranule (SG) is surrounded by rough endoplasmic reticulum (RER).N: Nucleus. 326,500.
Fig. 4. Golgi complex (G) located between three secretory granulesin an epithelial glandular cell. 340,000.
Fig. 5. Ventral glands duct consisting of flattened epithelial cells(EC) showing a dense grand, and myoepithelial cells (MC) with thecytoplasm full of filaments (F) and mitochondria (arrows). 312,000.
Fig. 6. Detail of the basal cytoplasm of a myoepithelial cell.Electron-dense bodies can be observed in the cytoplasm or adjacent tothe cellular basal membrane (arrows). F: Filaments. 330,000.
Fig. 7. Desmosome located between two myoepithelial cells.345,000.
Fig. 8. Interdigitations (arrow) between lateral membranes of twoadjacent epithelial cells of the ducts. 332,000.
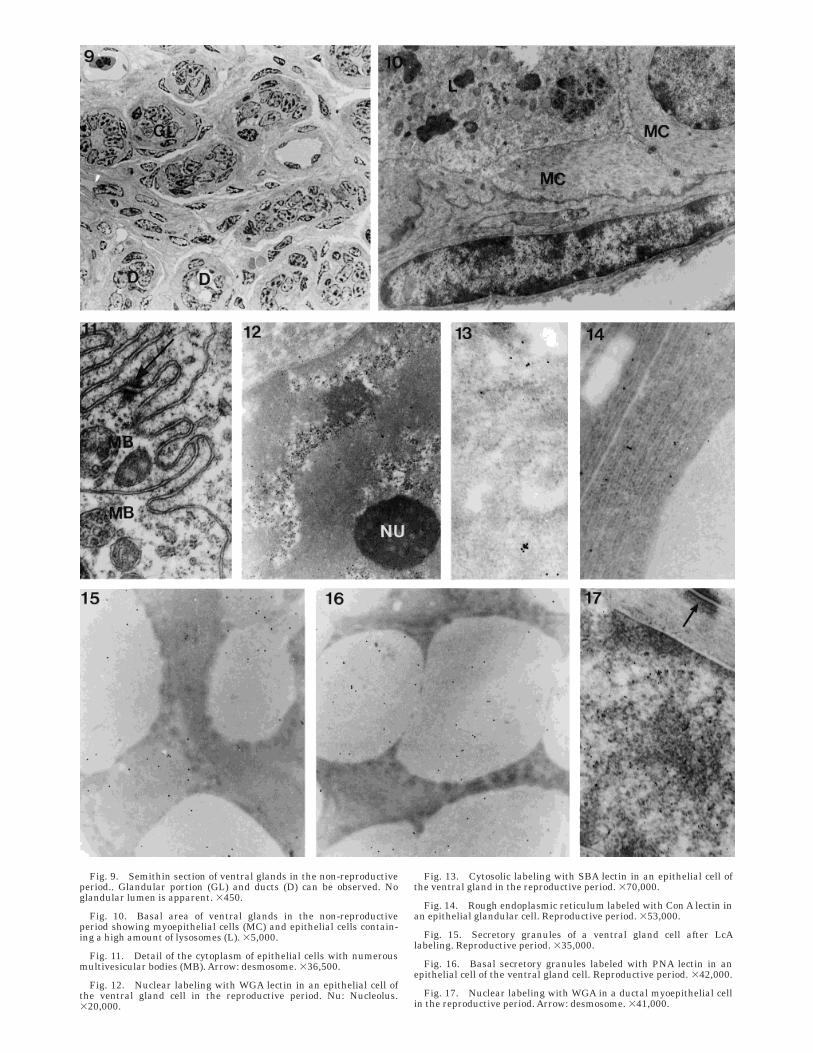
Fig. 9. Semithin section of ventral glands in the non-reproductiveperiod.. Glandular portion (GL) and ducts (D) can be observed. Noglandular lumen is apparent. 3450.
Fig. 10. Basal area of ventral glands in the non-reproductiveperiod showing myoepithelial cells (MC) and epithelial cells contain-ing a high amount of lysosomes (L). 35,000.
Fig. 11. Detail of the cytoplasm of epithelial cells with numerousmultivesicular bodies (MB). Arrow: desmosome. 336,500.
Fig. 12. Nuclear labeling with WGA lectin in an epithelial cell ofthe ventral gland cell in the reproductive period. Nu: Nucleolus.320,000.
Fig. 13. Cytosolic labeling with SBA lectin in an epithelial cell ofthe ventral gland in the reproductive period. 370,000.
Fig. 14. Rough endoplasmic reticulum labeled with Con A lectin inan epithelial glandular cell. Reproductive period. 353,000.
Fig. 15. Secretory granules of a ventral gland cell after LcAlabeling. Reproductive period. 335,000.
Fig. 16. Basal secretory granules labeled with PNA lectin in anepithelial cell of the ventral gland cell. Reproductive period. 342,000.
Fig. 17. Nuclear labeling with WGA in a ductal myoepithelial cellin the reproductive period. Arrow: desmosome. 341,000.

be involved in reabsorption of the excess of secretionmaterial synthesized in the previous period.
Norris (1987) suggested that cloacal glands in amphib-ians, reptiles, and birds may be homologous to struc-tures derived from the urogenital sinus in mammals. Aclear functional similarity cannot be established be-tween the ventral glands of urodeles and the glands ofthe mammalian urogenital tract, such as the seminalvesicles (Aumuller and Riva, 1992), prostate (Aumuller,1979), urethral glands (Testa-Riva et al., 1994), andbulbourethral glands (Testa-Riva et al., 1994) becausethe main function of ventral glands is to contribute tothe formation of a structure (the spermatophore) that isabsent in mammals. However, the ventral glands areultrastructurally very similar to the mammalian bulbo-urethral glands, which exhibit mucous secretory gran-ules like those of the ventral glands and are alsosurrounded by myoid cells (Testa-Riva et al., 1994).
In a previous light microscopy study, we demon-strated weak labeling with the lectins used in theventral glands of T. marmoratus located principally inbasal positions of the cells (Romo et al., 1996). In thepresent study, at the electron microscopic level, differ-ent lectins were localized at different sites in the cells ofthese glands, mainly in the basal cytoplasm, where thenucleus and cellular organelles are localized.
In the ventral glands of Triturus marmoratus, all celltypes have glycoconjugates in their nucleus, mainlyN-acetylglucosamine and/or sialic acid residues (WGA),mannose and/or glucose (ConA and LcA), galactose(PNA), and N-acetylgalactosamine (SBA and HPA).Nuclear glycoproteins are generally assumed to play aregulatory function in the nuclei (Londono and Ben-dayan, 1987) . It is known that Con A (which bindsmannose residues) also binds to non-histone glycopro-teins in the nuclear chromatin of rat liver cells (Rizzoand Bustin, 1977) and goblet cells of the chickenduodenum (Roth, 1984). In the ventral glands of themarbled newt, nuclear labeling to those residues ismore pronounced during the reproductive period in allcell types, mainly in the secretory cells. The diminutionof sugar residue content in all cell types during thenon-reproductive period might be related to a decreasein cell activity in this period.
The cytosol of ventral gland secretory cells during thereproductive period showed a positive reaction to thoselectins that bind N-acetylgalactosamine (SBA and HPA)or galactose (PNA), both residues typical of O-linkedoligosaccharides. The presence of sugar residues thatare free in the cytosol has been reported in several
mammalian and non-mammalian cells including theparasite Leishmania braziliensis (Nagakura et al.,1986), rabbit leucocytes (Feigenson et al., 1975), bovineadrenal glands (Meyer and Burger, 1976), and mousebrain (Hart et al., 1989). These authors have suggestedthat these cytosolic glycoproteins might form part ofcytoskeletal proteins and transcription factors or berelated to the glycogen synthesis process (Haltiwangeret al., 1990). In the ventral glands, the positive stainingto these three lectins only in the secretory cells suggeststhat these cytosolic lectins are indirectly related tosecretion.
The rough endoplasmic reticulum of the ventralgland epithelial secretory cells showed positive reactionto N-acetylglucosamine and/or sialic acid (WGA), andmannose and/or glucose residues (ConA and LcA).Similar labeling has also been observed in other glandu-lar cells such as the guinea-pig prostate (Chang andWong, 1992), and the rat exocrine pancreas (Pinto daSilva et al., 1981). In the rough endoplasmic reticulum,the N-glycosilation process starts with the incorpora-tion of the above-mentioned sugar residues (exceptsialic acid; Hirschberg and Snider, 1987). Since theseoligosaccharides were not detected in the ventral glandcytosol, it may be deduced that they are limited to theGolgi complex where N-glycosilation finishes and O-glycosilation occurs. In addition, these residues couldform part of the endoplasmic reticulum membraneglycoproteins (Kreibich et al., 1982).
The Golgi complex and secretory granules of ventralgland epithelial cells labeled those lectins that bind tomannose and/or glucose (LcA), fucose (UEA-I), galac-tose (PNA), and N-acetylgalactosamine (SBA and HPA).This latter residue is not typical of N-glycosilation butof O-glycosilation GalNAc (Strous and Dekker, 1992)and their presence suggests that the ventral glandsecretion contains both N- and O-linked oligosacchari-des. The Golgi complex negative labeling to somelectins that were detected in the rough endoplasmicreticulum suggests that some sugar residues that wereexposed in this organelle have been masked by newresidues added in the Golgi complex.
The secretions of the ventral glands of T. marmoratus(Romo et al., 1996) and other urodeles (Sever et al.,1990; Sever, 1992) have been previously described asacidic glycoproteins by means of conventional histo-chemistry. Mucous drops secreted by bulbourethralglands of mammals such as the goat (Tsukise andYamada, 1987) and man (Riva et al., 1989; Testa-Riva etal., 1994) are composed of acidic mucins that are rich in
TABLE 2. Lectin cytochemistry of the ventral glands in the non-reproductive period*
Lectin Carbohydrate bindingBackground
reaction
Glands Ducts
Epithelial cells Myoepithelial cells Epithelial cells Myoepithelial cells
N C N C N C N C
WGA (GlcNAc)n . NeuAc 16 6 4 44 6 11 — 63 6 5 — — — 113 6 12 —Con A a-D-Man . a-D-Glc 32 6 7 121 6 25 — 161 6 60 — 182 6 30 — 192 6 33 —LcA a-D-Man . a-D-Glc 12 6 3 31 6 7 — — — — — — —UEA-I a-L-Fuc 11 6 3 — — — — — — — —PNA D-Gal-(b1 = 3), DGalNAc 12 6 3 — — 19 6 9 25 6 10 29 6 7 — — 25 6 10SBA a,b-D-GalNAc 15 6 4 209 6 41 71 6 44 136 6 42 20 6 9 176 6 37 76 6 5 105 6 71 30 6 8HPA a-D-GalNAc 10 6 3 — — — — — — — —
*Data are expressed as number of gold particles per 10 µm2 (mean 6 SD) in the cellular location mentioned. Gal: Galactose. GlcNAc: N-acetylglucosamine. Man:mannose. Glc: glucose. GalNAc: N-acetylgalactosamine. Fuc: fucose. N: nucleus. C: cytosol. Values that did not differ from background values are omitted (—).
127CLOACAL VENTRAL GLANDS IN THE MALE NEWT

sialic acid. Although ventral glands also secrete acidicmucins and it is possible that sialic acid is present intheir secretion, we have not used specific lectins todetect sialic acid in the present study.
The secretory granules that appeared intensely la-beled with PNA, SBA, and HPA lectins seem to be thosemost recently formed, because they were located besidethe perinuclear zone where the rough endoplasmicreticulum and Golgi complex were placed. This sug-gests that during the maturation process that accompa-nies the migration of the granules to the glandularlumen, glycoproteins agglutinate, masking the bindingsites of these lectins, such as has been reported in ratPaneth cells (Leis et al., 1997).
Comparing the lectin labeling obtained in the differ-ent periods of the reproductive cycle, the most remark-able difference is the higher degree of labeling in thereproductive period, probably because the functionalactivity of the gland is increased in this period. In thistime, there is an active formation of glycoproteins in thenuclei, rough endoplasmic reticulum, and Golgi com-plex, and secretion of the glandular products is associ-ated with spermatophore formation, whereas in thenon-reproductive period a general glandular regressionoccurs, lacking even secretory granules.
Since, as has been mentioned above, the cloaca is anandrogen-dependent organ, the effects of castration inmammals could be compared to those of the non-reproductive period in marbled newt, because in thisperiod blood androgen concentration is lower (Norrisand Moore, 1975). A strong lectin labeling pattern in ratepididymis after castration has been reported (Aryaand Vanha-Perttula, 1984, 1985). These authors hypoth-esize that this fact was due to an increased activity ofglycosidases in the epithelial cells to lysate the largeamount of macromolecules, including glycoproteins,that are reabsorbed by the epithelial cells from theepididymal fluid. The decreased lectin labeling of theventral glands of the marbled newt in the non-reproductive period suggests that glycosidase activitystimulation during the non-reproductive period doesnot occur, at least in these glands, although this findingmay not be assumed for other cloacal glands.
REFERENCESArya M, Vanha-Perttula T. 1984. Distribution of lectin binding in rat
testis and epididymis. Andrologia 16:495–508.Arya M, Vanha-Perttula T. 1985. Effect of castration on lectin staining
in rat epididymis. Andrologia 17:327–337.Aumuller G. 1979. Prostate gland and seminal vesicles. Berlin:
Springer-Verlag.Aumuller G, Riva A. 1992. Morphology and functions of the human
seminal vesicle. Andrologia 24:183–196.Brizzi R, Delfino G, Calloni C. 1986. Accessory structures in the
genital apparatus of Salamandrina terdigitata (Amphibia: Salaman-dridae). I. Ultrastructural patterns of the male abdominal gland. ZMikrosk Anat Forsch, Leipzig 100:397–409.
Brizzi R, Delfino G, Calloni C. 1988. Accessory structures in thegenital apparatus of Salamandrina terdigitata (Amphibia: Salaman-dridae). II. Structural and ultrastructural specializations in themale cloacal region. Z Mikrosk Anat Forsch, Leipzig 102:833–857.
Brizzi R, Calloni C, Delfino G. 1990. Accessory structures in thegenital apparatus of Salamandrina terdigitata (Amphibia: Salaman-dridae). IV. Male cloacal glands: A study under light and scanningelectron microscopes. Z Mikrosk Anat Forsch, Leipzig 104:871–897.
Chang L, Wong YC. 1992. Localization of prostatic glycoconjugates bylectin gold method. Acta Anat 143:27–40.
Feigenson ME, Schnebli HP, Baggiolini M. 1975. Demonstration ofricin-binding sites on the outer face of azurophil and specific
granules of rabbit polymorphonuclear leukocytes. J Cell Biol 66:183–188.
Friend DS. 1969. Cytochemical staining of multivesicular body andGolgi vesicles. J Cell Biol 41:269–279.
Geniez P, Grillet P. 1990. Les salamandres et les tritons. EditionsPayot Lausanne. Suisse.
Haltiwanger RS, Holt GD, Hart GW. 1990. Enzymatic addition ofO-GlcNAc to nuclear and cytoplasmic proteins. Identification of auridine diphospho-N-acetylglucosamine peptide b-N-acetylglu-cosaminyltransferase. J Biol Chem 265: 2563–2568.
Hart GW, Haltiwanger RS, Holt GD, Kelly WG. 1989. Glycosilation inthe nucleus and cytoplasm. Ann Rev Biochem 58:841–874.
Hirschberg CB, Snider MD. 1987. Topography of glycosilation in therough endoplasmic reticulum and Golgi apparatus. Ann Rev Bio-chem 56:63–87.
Kornfeld R, Kornfeld S. 1985. Assembly of asparagine-linked oligosac-charides. Ann Rev Biochem 54:631–664.
Kreibich G, Ojakian G, Rodriguez-Boulan E, Sabatini DD. 1982.Recovery of ribophorins and ribosomes in ‘‘inverted-rough’’ vesiclesderived from rat liver rough microsomes. J Cell Biol 93:111–121.
Leis O, Madrid JF, Ballesta J, Hernandez F. 1997. N- and O-linkedoligosaccharides in the secretory granules of rat Paneth cells: anultrastructural cytochemical study. J Histochem Cytochem 45:285–293.
Londono I, Bendayan M. 1987. Ultrastructural localization of manno-side residues on tissue sections: comparative evaluation of theenzyme-gold and the lectin-gold approaches. Eur J Cell Biol 45:88–96.
Lucocq J. 1994. Quantitation of gold labeling and antigens in immuno-labelled ultrathin sections. J Anat 184:1–13.
Malacarne G, Giacoma C. 1986. Chemical signals in European newtcourtship. Boll Zool 53:79–83.
Malacarne G, Bottoni L, Massa R, Vellano C. 1984. The abdominalgland of the crested newt: a possible source of courtship phero-mones. Preliminary ethological and biochemical data. Monitore ZoolItal 18:33–39.
Meyer DI, Burger MM. 1976. The chromaffin granule surface. Localiza-tion of carbohydrate on the cytoplasmic surface of an intracellularorganelle. Biochim Biophys Acta 443:428–436.
Moniem KA, Glover TD. 1972. Comparative histochemical localizationof lysosomal enzymes in mammalian epididymides. J Anat 111:437–452.
Nagakura K, Tachibana H, Kaneda Y, Nakae T. 1986. Leishmaniabraziliensis: localization of glycoproteins in promastigotes. ExpParasitol 61:335–342.
Norris DO. 1987. Regulation of males gonoducts and sex accessorystructures. In: DO Norris DO, Jones RE, editors. Hormones andreproduction in fishes, amphibians and reptiles. New York: Plenum.p 327–354.
Norris DO, Moore FL. 1975. Antagonism of testosterone-inducedcloacal development by estradiol-17 in immature larval tiger sala-manders (Ambystoma tigrinum). Herpetologica 31:255–263.
Norris DO, Norman MF, Pancak MK, Duvall D. 1985. Seasonalvariations in spermatogenesis, testicular weights, vasa deferentisand androgen levels in neotenic male tiger salamanders, Ambys-toma tigrinum. Gen Comp Endocrinol 60:51–57.
Pinto da Silva P, Torrisi MR, Kachar B. 1981. Freeze-fracture cytochem-istry. Localization of wheat germ agglutinin and concanavalin Alectin binding sites on freeze-fractured pancreatic cells. J Cell Biol91:361–372.
Rademacher TW, Parekh RB, Dwek RA. 1988. Glycobiology. Ann RevBiochem 57:785–838.
Riva A, Usa IE, Scarpa R, Cossu M, Lantini MS. 1989. Fine structureof the accessory glands of the human male genital tract. In: MottaPM, editor. Developments in ultrastructure of reproduction. Progressin clinical and biological research. Vol. 296. New York: Alan R. Liss.
Rizzo WB, Bustin M. 1977. Lectin as probes of chromatin structure.Binding of concanavalin A to purified rat liver chromatin. J BiolChem 252:7062–7067.
Romo E, De Miguel MP, Arenas MI, Frago L, Fraile B, Paniagua R.1996. Histochemical and quantitative study of the cloacal glands ofTriturus marmoratus marmoratus (Amphibia: Salamandridae). JZool Lond 239:177–186.
Roth J. 1983. The colloidal gold marker system for light and electronmicroscopic cytochemistry. In: Bullock GR, Petrusz P, editors.Techniques in Immunocytochemistry. Vol. 2. New York: AcademicPress. p 217–284.
Roth J. 1984. Cytochemical localization of terminal N-Acetyl-D-Galactosamine residues in cellular compartments of intestinalgoblet cells: implication for the topology of O-glycosilation. J CellBiol 98:399–406.
128 E. ROMO ET AL.

Russell LD, Brandon RA, Zalisko EJ, Martan J. 1981. Spermatophoresof the salamander Ambystoma texanum. Tissue Cell 13:609–621.
Saez FJ, Fraile B, Paniagua, R. 1989. Histological and quantitativechanges in the annual testicular cycle of Triturus marmoratusmarmoratus. Can J Zool 68:63–72.
Sever DM. 1981. Cloacal anatomy of male salamanders in the familiesAmbystomatidae, Salamandridae and Plethodontidae. Herpeto-logica 37:142–155.
Sever DM. 1991. Comparative anatomy and phylogeny of the cloacaeof salamanders (Amphibia: Caudata). I. Evolution at the familylevel. Herpetologica 47:165–193.
Sever DM. 1992. Comparative anatomy and phylogeny of the cloacaeof salamanders (Amphibia: Caudata). IV. Salamandridae. Anat Rec232:229–244.
Sever DM, Houck LD. 1985. Spermatophore formation in Desmogna-thus ochrophaeus (Amphibia: Plethodontidae). Copeia 1985:394–402.
Sever DM, Verrell PA, Halliday TR, Griffiths M, Waights V. 1990. Thecloaca and the cloacal glands of the male smooth newt, Triturus
vulgaris vulgaris (Linnaeus), with especial emphasis on the dorsalgland. Herpetologica 46:160–168.
Strouss GJ, Dekker J. 1992. Mucine type proteins. Crit Rev BiochemMol Biol 27:57–92.
Tanaka S, Iwasawa, H. 1979. Annual changes in testicular structureand sexual character of the Japanese red-bellied newt Cynopspyrrogaster pyrrogaster. Zool Mag 88:295–305.
Testa-Riva F, Riva A, Congiu T, De Lisa A, Motta PM. 1994. Humanbulbourethral and urethral glands. In: Riva A, Testa-Riva F, MottaPM, editors. Ultrastructure of the male urogenital glands: prostate,seminal vesicles, urethral and bulbourethral glands. Boston: Klu-wer Academic Publishers. p 163–176.
Tsukise A, Yamada K. 1987. Histochemistry of glycoconjugates in thesecretory epithelium of the goat bulbourethral gland. Acta Anat129:344–352.
Zalisko EJ, BrandonA, Martan J. 1984. Microstructure and histochem-istry of salamander spermatophores (Ambystomatidae, Salamandri-dae and Plethodontidae). Copeia 1984:741–749.
129CLOACAL VENTRAL GLANDS IN THE MALE NEWT


















![Cloacal exstrophy associated with gastroschisis: Case ...gastroschisis, omphalocele, bladder exstrophy, and cloacal exs-trophy [1,2]. Gastroschisis is a defect of the anterior abdominal](https://img.pdfslide.us/doc/110x75/5f82b6822991d932fc2027c1/cloacal-exstrophy-associated-with-gastroschisis-case-gastroschisis-omphalocele.jpg)










