Embed Size (px)
Citation preview

Ultrastructural evidence of stromal/epithelial interactions inthe human endometrial cycle
Daniel K. Roberts, MD, PhD, Nola ]. Walker, EMSA, and Lynn A. Lavia, PhD
Wichita, Kansas
We found ultrastructural evidence of interactions between glandular epithelium and superficial stromal cellsof the human endometrium during phases of the menstrual cycle. Four significant changes were observedin the transition from early proliferative (days 5 to 9) to early secretory (days 15 to 19) phases. Thesechanges included: (1) an increase in the number and size of lamina densa disruptions, (2) an increase inthe number and size of gap junctions, (3) an increase in the number and complexity of epithelial cellprojections that extended through the lamina densa, and (4) an increase in close contacts between stromaland epithelial cells. The complex epithelial cell projections that extended through the lamina densa were inclose proximity to stromal cells. These interactions were seen primarily in the early secretory phase. Afterthat time (days 20 to 28) the interactions were less frequent. These morphologic results reveal complexphysical interactions between epithelial and stromal cells of the adult endometrium . The interactions reachmaximal development during the preimplantation phase of the endometrial cycle. (AMJ OSSTET GVNECOL
1988;158:854-61.)
Key words: Menstrual cycle, stromal-epithelial interactions, morphology, transmissionelectron microscopy
Hormones have been shown to initiate growth anddifferentiation in tissues such as uterine glandular epithelial cells.1 Local substances such as growth factorsand other diffusible mediators influence these processes." Although the sources of these chemical mediators are largely unknown, it is known that stroma isrequired for normal epithelial cell development.' Furthermore, a close proximity of adjacent stromal andepithelial cells occurs in growth and differentiation ofembryonic tissues such as mouse tooth,' primary mesenchyme in mouse embryos," avian integument," andmurine lung.'. s
The adult human uterine glandular cells undergomonthly cycles ofgrowth and differentiation, yet littleis known of stromal-epithelial interactions that occurto facilitate this hormone-stimulated development.Therefore the studies reported in this article gatheredmorphologic evidence in support of: (1) the existenceof these interactions, (2) the specific chronological sequence, and (3) the nature of epithelial and stromalcell ultrastructural modifications that occur during the
From the Department of Obstetrics and Gynecology, University ofKansas School of Medicine,-Wichita.
Supported in partbya cancerresearch grantfrom theLadiesAuxiliaryto the Veterans of Foreign Wars, Department of Kansas, and inpart by the Wesley Medical Research Institutes.
Presented at theSixth Annual Meeting of theAmericanGynecologicaland Obstetrical Society, Phoenix,Arizona,September 10-12, 1987.
Reprint requests: Daniel K. Roberts, MD, PhD, Department of Obstetrics and Gynecology, University of KansasSchool ofM edicineWichita, HCA Wesley Medical Center, Wichita, KS 67214.
854
Fig. 1. An epithelial cell is seen with an extension protrudingthrough the lamina densa. The bod y of a stromal cell isseen below. Earl y proliferative phase. (Original magnificationx 3 1,933.)
Volume 158Number 4
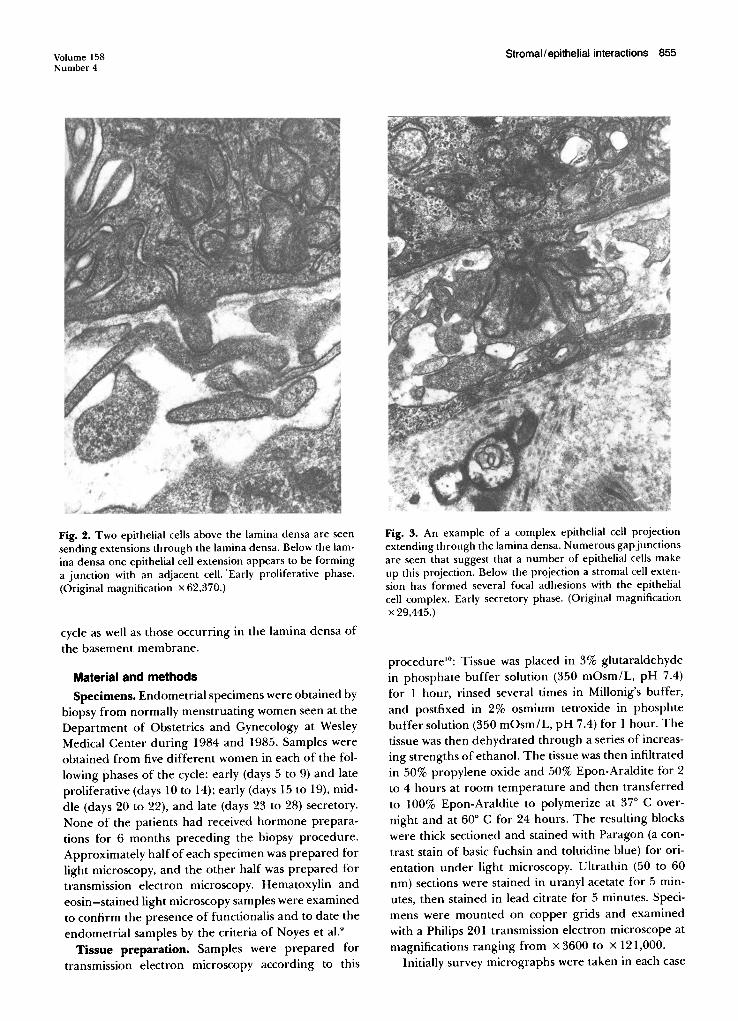
Fig. 2. Two epithelial cells above the lamina densa are seensending extensions through the lamina den sa. Below the lamina densa one epithelial cell extension appears to be forminga junction with an adjacent cell. 'Early proliferative phase.(Original magnification x 62,370.)
cycle as well as those occurring in the lamina densa ofthe basement membrane.
Material and methods
Specimens. Endometrial specimens were obtained bybiopsy from normally menstruating women seen at theDepartment of Obstetrics and Gynecology at WesleyMedical Center during 1984 and 1985. Samples wereobtained from five different women in each of the following phases of the cycle: early (days 5 to 9) and lateproliferative (da ys 10 to 14); early (days 15 to 19), middle (days 20 to 22), and late (days 23 to 28) secretory.None of the patients had received hormone preparations for 6 months preceding the biopsy procedure.Approximately half of each specimen was prepared forlight microscopy, and the other half was prepared fortransmission electron microscopy. Hematoxylin andeosin-stained light microscopy samples were examinedto confirm the presence of functionalis and to date theendometrial samples by the criteria of Noyes et al.?
Tissue preparation. Samples were prepared fortransmission electron microscopy accord ing to th is
Stromal/epithelial interactions 855
Fig. 3. An example of a complex epithelial cell projectionextending through the lamina den sa. Numerous gap j unctionsare seen that suggest that a number of epithelial cells makeup this projection. Below the projection a stromal cell extension has formed several focal adhesions with the epithelialcell complex. Earl y secretory pha se. (Original magnificationx 29 ,445.)
procedure": Tissue was placed in 3% glutaraldehydein phosphate buffer solution (350 mOsm/L, pH 7.4)for 1 hour, rinsed several times in Millonig 's buffer,and postfixed in 2% osmium tetroxide in phosphtebuffer solution (350 mOsm/L, pH 7.4) for 1 hour. Thetissue was then dehydrated through a series of increasing strengths of ethanol. The tissue was then infiltratedin 50% propylene oxide and 50% Epon-Araldite for 2to 4 hours at room temperature and then transferredto 100% Epon-Araldite to polymerize at 37° C overnight and at 60° C for 24 hours. The resulting blockswere thick sectioned and stained with Paragon (a contrast stain of basic fuchsin and toluidine blue) for orientation under light microscopy. Ultrathin (50 to 60nm) sections were stained in uranyl acetate for 5 minutes, then stained in lead citrate for 5 minutes. Specimens were mounted on copper grids and examinedwith a Philips 201 transmission electron microscope atmagnifications ranging from x 3600 to x 121,000.
Initially survey micrographs were taken in each case

856 Roberts, Walker, and Lavia April 1988Am J Obstet Gynecol
Fig. 4. Two epithelial cells are seen above a diffuse lamina densa. The complex cell projectionextending through the lamina densa appears to be composed of processes from at least two differentepithelial cells, separated by a developing gap junction. To the left of the complex epithelial cellprojection is a stromal cell with a focal adhesion between it and the epithelial cell projection. To theright of the complex epithelial cell projection is a stromal cell that is apparently fused with theprojection. Gap junctions as well as a desmosome are seen between the epithelial cells above thelamina densa. Late proliferative phase. (Original magnification X 29,445.)
and examined for interactions. Additional micrographswere taken, as required, to confirm the extent of theinteractions in a given endometrium. A minimum oftwo tissue blocks (50 sections per block) was examinedfor each endometrium. Additional blocks were used asnecessary to confirm and correlate the extent of interactions in a particular phase.
Results
Major structural changes occurred in the basal portion of the glandular epithelial cells, the superficialstromal cell layer, and the lamina densa. Although therewas regional variability in the degree to which the endometria participated in the changes described, consistent and discernible differences were noted betweenendometria obtained on different days of the cycle.
Development of epithelial projections through thelamina densa, The lamina densa of the glandular epithelium was largely intact during the early proliferativephase. Occasional focal interruptions were seen thatsometimes contained projections from single epithelialcells (Fig. 1). At other times two adjacent epithelial cellswere noted to send projections simultaneously throughthe interrupted lamina densa (Fig. 2).
In the early secretory phase after ovulation (days 15to 19) these epithelial projections became more complex (Fig. 3). Epithelial projections were often closely
apposed to an underlying stromal cell, where focaladhesions were identified (Fig. 3).
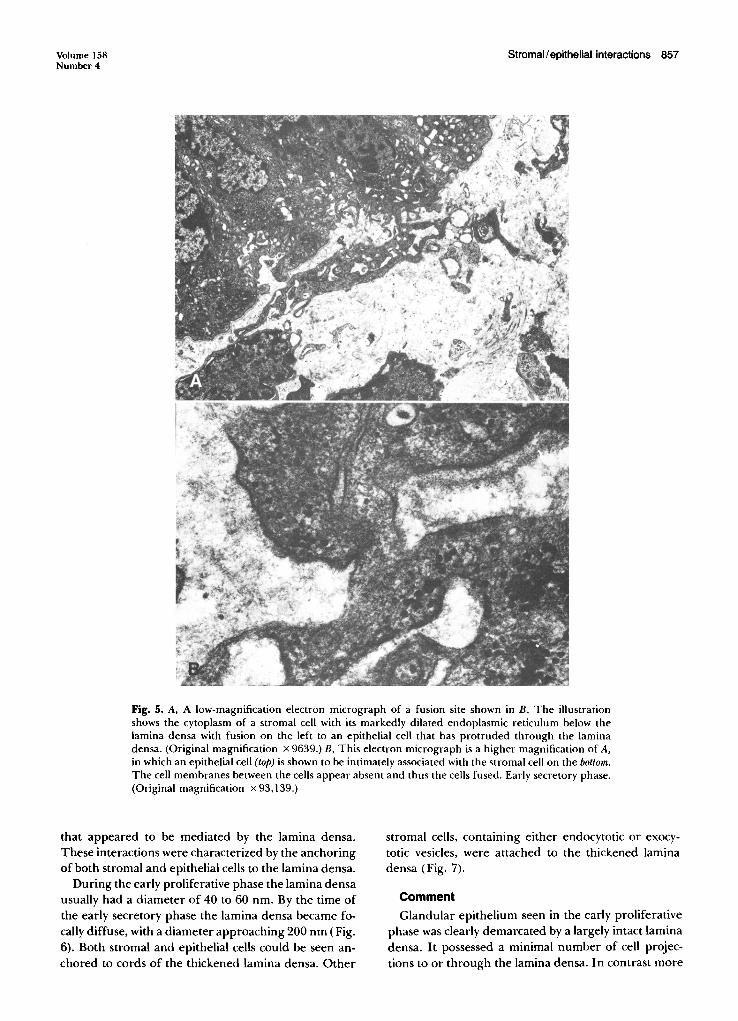
In addition to focal adhesion a second type ofstromal-epithelial cell interaction was noted in association with these complex cell projections (Fig. 4). Inthis type of interaction there were areas of extremelyclose stromal cell approximation to the epithelial cellprojection. No cell membrane could be identified ineither epithelial or stromal cells, which suggested thatthese cells were fused. At other times these apparentfusions between epithelial and stromal cells were moredirect and were not associated with complex cell projections (Fig. 5).
Some of the complex epithelial cell projections contained numerous large and convoluted gap junctions(Fig. 3). Elortgated gap junctions could sometimes. befound in basolateral areas of the adjacent epithelialcells. These gap junctions were also noted in epithelialcell membrane processes that did not project below thelamina densa. By the middle secretory (days 20 to 22)and late secretory (days 23 to 28) phases the frequencyand complexity of stromal-epithelial cell interactions,as well as gap junction development, had decreased.
Other lamina densa-related developments. In addition to the above-documented direct interactions between stromal and epithelial cells, other morphologicchanges were seen in both stromal and epithelial cells
Volume 158Number 4
Stromal/epithelial interactions 857
Fig. 5. A, A low-magnification electron micrograph of a fusion site shown in B. The illustrationshows the cytoplasm of a stromal cell with its markedly dilated endoplasmic reticulum below thelamina densa with fusion on the left to an epithelial cell that has protruded through the laminadensa. (Original magnification x 9639.) B, This electron micrograph is a higher magnification of A,in which an epithelial cell (top) is shown to be intimately associated with the stromal cell on the bottom.The cell membranes between the cells appear absent and thus the cells fused. Early secretory phase.(Original magnification x 93,139.)
that appeared to be mediated by the lamina densa.These interactions were characterized by the anchoringof both stromal and epithelial cells to the lamina densa.
During the early proliferative phase the lamina densausually had a diameter of 40 to 60 nm. By the time ofthe early secretory phase the lamina densa became focally diffuse, with a diameter approaching 200 nm (Fig.6). Both stromal and epithelial cells could be seen anchored to cords of the thickened lamina densa. Other
stromal cells, containing either endocytotic or exocytotic vesicles, were attached to the thickened laminadensa (Fig. 7).
CommentGlandular epithelium seen in the early proliferative
phase was clearly demarcated by a largely intact laminadensa. It possessed a minimal number of cell projections to or through the lamina densa. In contrast more

858 Roberts, Walker, and Lavia
Fig. 6. The stromal cell seen below is associated with theepithelial cell above through attachment of cords of thelamina densa. Late proliferative phase. (Original magnification x 71,010.)
complex stromal epithelial cell interactions were seenin the late proliferative and early secretory phases. Thedevelopment of these changes over the same timecourse, in which the whole endometrium is preparingfor implantation, suggests that the stromal-epithelialinteractions noted here are a part of the more generaldevelopment. Such a conclusion is supported by otherstudies on mouse tooth! embryonic' and postnatal murine lung," and avian integument" and lung," as wellas by interactions between the primary mesenchymeand the embryonic ectoderm in mouse embryos."
A major feature of the cyclic changes seen was thedevelopment of epithelial cell projections that began toappear in the early proliferative phase as small extensions, from either single or adjacent epithelial cells(Figs. I and 2). As these processes extended into thestroma, they appeared to develop gap junctions withadjacent epithelial cell processes (Fig. 2). By the timeof the late proliferative and early secretory phases theseextensions became complex epithelial cell projectionsthat appeared to continue to develop gapjunctions withadjacent epithelial cells (Figs. 3 and 4). These dramaticstructures with their gap junctions suggest that there
April 1988Am J Obstet Gynecol
must be extensive intercellular communication. Thiscommunication appears to be more extensive than hasbeen described for developing embryonic structures,such as murine primary mesenchyme' and lung."
The direct interactions noted here, between thestromal and epithelial cells, were of two types. The firsttype was that of the well-defined focal adhesion" witha 10 to 20 nm gap between the stromal and epithelialcell (Figs. 3 and 4). The second type was an even closerinteraction between epithelial cell projections and adjacent stromal cells (Figs. 4 and 5). Both the epithelialcell projection and the stromal cell appeared to lackcell membranes and thus appeared fused. Other artifactual reasons for this morphologic condition wereconsidered but were deemed unlikely because numerous areas of apparent cell fusion appeared nonrandomly. Heterotypic cell fusion has been known to occurfor some time." Close epithelial-stromal interactionsthat suggest such fusion have been reported in themouse integument." The exact nature of this interaction remains to be elucidated and eventually may require the reevaluation of cell interaction mechanisms.
The anchoring of epithelial and stromal cells to thethickened lamina densa (Fig. 6) appeared to be an additional process by which stromal and epithelial cellscommunicated. The meaning of these data, as well asthe finding of exocytotic or endocytotic vesicles (Fig.7) in the area of the diffuse lamina densa, is not known.It is suggested that these cells may be in the process ofremodeling the lamina densa or releasing intercellularmessages. 15
In menstrual endometrium the uptake of fluid andstromal substances through basal epithelial processesand phagocytosis has been reported. 16.17 It is suggestedthat these substances pass to the glandular lumen viavesicles within the epithelial cells as well as intercellularly.
Although the nature of the messages facilitated bythese contacts is unknown, the morphologic featuresare consistent with the idea that the preimplantationdevelopment of endometrial epithelium requires stromal interaction. Furthermore, the structures describedhere are 'consistent with those seen in a variety of developing embryonic tissues, in which stromal interaction is known to be important in epithelial cell differentiation. Therefore one could speculate that theproper differentiation and function of the endometrialepithelium in the preimplantation phase are directedby messages from the stroma and perhaps that theepithelium acts as a conduit for stromal products.
REFERENCES
I. Wynn RA, ed. Biology of the uterus. New York: PlenumPress, 1977.

Volume 158Number 4
Stromal/epithelial interactions 859
Fig. 7. The lamina densa is the anchor for the stromal cell on the bottom and the epithelial cell onthe top. Note the pinocytotic vesicles in the stromal cell membrane. (Original magnification x 85,882.)
2. Sirbasku DA, Leland FE, Benson RH. Properties of agrowth factor present in crude extracts of rat uterus.J Cell Physiol 1981;107:345-58.
3. Cunha GR, Bigsby RM, Cooke PS, Sugimura Y. Stromalepithelial interactions in adult organs. Cell Differ 1985;17:137-48.
4. Slavkin HC, Snead ML, Zeichner-David M, Jaskoll TF,Smith BT. Concepts of epithelial-mesenchymal interactions during development: tooth and lung organogenesis.J Cell Biochem 1984;26:117-25.
5. Franke WW, Grund C, Jackson BW, lllmensee K. Formation of cytoskeletal elements during mouse embryogenesis. IV. Ultrastructure of primary mesenchymal cellsand their cell-cell interactions. Differentiation 1983;25:121-41.
6. Mauger A, Demarchex M, Sengel P. Role of extracellularmatrix and of dermal-epidermal junction architecture inskin development. In: Kemp RB, Hinchliffe JR, eds. Matrices and cell differentiation. New York: Alan R Liss,1984: 115-28.
7. Bluemink JG, van Maurik P, Lawson KA. Intimate cellcontacts at the epithelial/mesenchymal interface in embryonic mouse lung. J Ultrastruct Res 1967;55:257-70.
8. Adamson IYR, King GM. Epithelial-mesenchymal interactions in postnatal rat lung growth. Exp Lung Res1985;8:261-74.
9. Noyes RW, Hertig AT, Rock J. Dating the endometrialbiopsy. Fertil Steril 1950; 1:3-26.
10. Lavia LA, Roberts DK, Walker NJ, Anderson K. Rat luminal cell nuclear area changes correlated with uterinegrowth responses induced by a low dose infusion or injection of estradiol-l Zp. Steroids 1985;45:519-37.
11. Gallagher BC. Branching morphogenesis in the avianlung: electron microscopic studies using cationic dyes.J Embryol Exp Morphol 1986;94:189-205.
12. Abercrombie M, Heaysman JEM, Pegrum SM. The locomotion offibroblasts in culture. IV. Electron microscopyof the leading lamella. Exp Cell Res 1971;67:359-67.
13. Harris H, Watkins JF. Hybrid cells derived from mouseand man: artificial heterokaryons of mammalian cellsfrom different species. Nature 1965;205:640-1.
14. Hardy MH, Goldberg EA. Morphological changes at the
basement membrane during some tissue interactions inthe integument. Can J Biochem Cell Bioi 1983;61:95765.
15. Stoker M, Perryman M. An epithelial scatter factor released by embryo fibroblasts. J Cell Sci 1985;77:209-23.
16. Flowers CE, Wilborn WHo New observations on the physiology of menstruation. Obstet Gynecol 1978;51: 16-24.
17. Wilborn WH, Flowers CEo Cellular mechanisms for endometrial conservation during menstrual bleeding. SeminReprod Endocrinol 1984;2:307-47.
Editors' note: This manuscript was revised after thesediscussions were presented.
DiscussionDR. HUGH M. SHINGLETON, Birmingham, Alabama.
Dr. Roberts and his colleagues are to be congratulated on an original contribution concerning stromalepithelial interactions during the endometrial cycle,which I believe is the first such study on human endometrium. The observations are in four categories:(1) lamina densa disruptions, (2) increase in numberand size of gap junctions, (3) increased numbers andcomplexity of epithelial cell projections through thelamina densa, and (4) an increase in close contacts between stromal and epithelial cells. Such evidence ofstromal-epithelial interactions supports the observations of Cunha et al. I that epithelial renewal and differentiation are tightly regulated to meet functionaldemands during reproductive cycles and that uncoordinated and altered interactions between stromal andepithelial cells are likely to be associated with the development of cancer and other disease states.
Recent work (Lawrence HD, Whitaker D, Sugimura Y, Cunha GR, Dickersin GR, Robboy SJ. An ultrastructural study of the developing urogenital tract















![Case Report A Case of Extrauterine Endometrial Stromal ...downloads.hindawi.com/journals/criog/2013/202458.pdfCase Reports in Obstetrics and Gynecology [ ]H.Y.Cho,M.K.Kim,S.J.Cho,J.W.Bae,andI.Kim,](https://img.pdfslide.us/doc/110x75/609e1f77ec2eb021e644bc5a/case-report-a-case-of-extrauterine-endometrial-stromal-case-reports-in-obstetrics.jpg)












