Embed Size (px)
Citation preview

Typing of Rhizobium by phages
RYSZARD STANIEWSKI Department of General Microbiology, M. Curie-Sklodowska Crniversiry, Lrrblin, Poland
Received Decern ber 19, 1968
STANIEWSKI, R. 1970. Typing of Rhizobium by phages. Can. J. Microbiol. 16: 1003-1009. Two hundred and thirty strains of Rhizobi~tm trifolii, Rhizobi~rm legumirlosarum for pea, vetch, horse
bean, and Lathyr~rs spp., Rhizobium phaseoli and Rhizobi~lm meliloti were subjected to phage typing. On the basis of their sensitivity to phages these strains were divided into three groups: I, 11, and 111.
In group I, consisting of R. trifolii, R. leguminosarrrm for pea, vetch, and horse bean, and R. phaseoli, 18 phage types were established. Group II included some strains of R. trifolii and R. leg~rn~itzosarum for pea and vetch. Among them three phage types were distinguished. Group I11 included R. meliloti strains and one strain of Rhizobi~rtir I~rpitri for lupine. In that group 10 phage types were found.
Introduction R. phaseoli, while lucerne bacteria formed a
Phages were shown to have a wide application in epidemiological studies not only in the identi- fication of species of bacteria (Meynell 1964; Stocker 1955) but in the differentiation between pathogenic and saprophytic strains, e.g. staphy- lococci (Wentworth 1963), tubercle bacilli (Juhasz and Bijnicke 1965), and also in the de- termination of virulence of bacteria (Wentworth 1963). Craigie and Yen (1938) isolated phages acting on the Vi form of Salmonella typhi and worked out a scheme of phage typing of these bacteria.
Soil bacteria, including rhizobia which live in symbiosis with leguminous plants, have so far not been extensively studied as far as phage typing is concerned.
The taxonomy of Rhizobium was based on its ability to live symbiotically with some species of leguminous plants (Breed et al. 1957). Attempts were also made at Rhizobiutn classification ac- cording to its sensitivity to phages. On the basis of the results obtained by Gerretsen et al. (1923), Laird (1932), Dorosinskii (1941), Conn et al. (1945), Demolon (1951), Kleczkowska (1957), Staniewski et al. (1962), and Staniewski (1970) it may be concluded that the host range of phages propagated on the strains belonging to the same species of Rhizobium was often differ- ent. Therefore the degree of relationship between different species of Rhizobiurn was also defined diversely. The studies of Conn et al. (1945) and Staniewski (1970) on the susceptibility of dif- ferent species of Rkizobiunz to phages and studies of Graham (1964) on different physiological properties of these bacteria proved a relation- ship between R. trifolii, R . leguminosarum, and
separate group. However, there is lack of studies aimed at a
differentiation of particular strains within species of Rhizobium on the basis of their sensitivity to phages. Taking into account the results of Laird (1932), Conn et al. (1945), and the author (Staniewski 1970), attempts were made to find out whether strains belonging to various species of Rhizobium, which showed a similar sensitivity to the action of non-diluted phage suspensions, could be further differentiated by phage typing. It was found in earlier tests (Staniewski 1970), that all strains of Rhizobium used in the present studies were sensitive to non-diluted cultures of 28 phages tested.
In this paper results of detailed studies on the differentiation of strains of various species of Rhizobium with phage typing technique are presented.
Materials and Methods Rhizobillm Straitcs
Two hundred and thirty Rhizobium strains belonging to the following species were used for phage typing: R. trifblii 113 strains; R. leg~rminosar~m~ for pea 24 strains, for vetch 9 strains, for horse bean 1 strain, for Lathyrus spp. 1 strain; R. meliloti 70 strains; R. hrpi~i for lupine ! strain; R. pllaseoli 11 strains.
Most of the Rhizobiltrn strains were isolated from root nodules of different leguminous plants grown in various localities in Poland. Thirteen strains of Rhizobium (R. trifolii Nos. 21-26 and 28, R. leg~rmit~osarwn for pea Nos. 1 and 2 and for vetch No. 36, R. meliloti Nos. 12, 14, and 16b) isolated in Poland and 24 foreign ones were kindly supplied by the Department of Agricultural Microbiology, IUNG, Pulawy. Foreign strains of Rhizobiunc were of the following origin: R. rrifolii No. 28n and R. meliloti No. 16 from the Institute National de le Recher- c h e ~ Agronomiques, Versailles, France; R. meliloti
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
AL
GA
RY
on
10/0
1/12
For
pers
onal
use
onl
y.

1004 CANADIAN JOURNAL OF MICROBIOLOGY. VOL. 16, 1970
No. 11 from Rothamsted Experimental Station, Har- penden, Herts, England; R. legunzit~osarun~ for pea Nos. 3a, 36, P8, and PRE from Wageningen, Laboratory of Microbiology, Holland; R. frifolii Nos. 226, 237, and 243, R. le~~rrnd~osarunl for pea Nos. 402 and 404, R. meliloii No. L56 from B;l.ljvaxtlaboratoriet Lantsbruk- shogskola, Uppsala, Sweden; R. frifolii No. 211, R. legrrmitrosrrrrrtn for pea Nos. 3c, 301: 311, and 312, R. meliloii Nos. L60 and L61 from the University of Wisconsin, Ilepartment of Bacteriology, U.S.A.; R. frifolii No. 27, R. legurnit~osar~trn for pea No. 3, for horse bean No. 34, R. nlelilofi No. 13 from Vsesoyuznyi Nauchno Issledovatelskii Iilstitut Selskokhozyaistvennoi Mikro- biologii, Leningrad, U.S.S.R.
The other 183 strains were isolated in our laboratcry. Besides original strains of R. ~?lelilofi, 23 lysogenized
derivates of this species were also used. These strains were obtained by Kowalski (1965,1966) after lysogenization of two strains of li~cerne bacteria with temperate phages isolated from different lysogenic strains of these bacteria.
Strains Nos. 23c, 23e, and 23f of clover bacteria, No. 36f for vetch bacteria, No. LAa of lucerne bacteria and strain No. F l u of bean bacteria were semirough mutants of strains 23, 36, L4, and F1 respectively.
Bacf eriophoges Soil samples in which phages were sought were taken
from fields where leguminous plants were cultured. For phage isolation a modified method of Dernolon and Dunez (Vandecaveye and Katznelson 1936) or Klecz- kowska (1957), described in the early papers, was used (Staniewski et al. 1962; Staniewski 1970). The virulent phages were isolated from filtrates of roots of leguminous plants with their nodules and the adjacent soil and then propagated on sensitive Rliizobiuttl strains of respective species. About 150 phage strains were isolated. In pre- liminary studies the activity of phages was determined on a small number of RAizobiuni strains. For further study the 28 most active phages were selected. Multipli- cation, titration, and routine test dilution (RTD) deter- mination of the phages were carried out on the following host strains: R. frifolii(o)No. 21 for phages 2C, 3C,and5C, (b) No. 22 for phage lC, (c) No. 23f'for phages 6C and 8C, (d) No. 24 for phages 7C and 9C, (e) No. 26 for phage 4C; R. Iegio~rinosarrr~n (a) for pea No. 1 for phages 2P, 3P, and 4P, (b) for pea No. 2 for phage lP, (c) for vetch No. 36f for phages lV, 2V, and 3V, (d) for horse bean No. 34 for phages lH, 2H, and 3H; R.pliaseoli No. F4 for phages 1B and 2B; R. tnelilo~i(o) No. 11 for phage lA, (6)No. 13 for phages 2A and 3A, (c) No. L8 for phage 4A; R. Irrpitii for lupine No. 39 for phages lL, 2L, and 3L. Pure clones of phages were obtained by successive reisolations of phages from single plaques. The titer of phage stocks was determined by the double agar layer technique (Adarns 1959). For various phages those ranged from 2 X 107 to 3 X 1010 plaque-forming units.
For storing of Rllizobillm strains Thornton medium was used (Fred et al. 1932). Bacteria and phages were propagated on "mediilm 5" (Laird 1932). Titration of phages, determination of 1 X RTD and typing of Rhizo- biirnr strains were carried out on agar medium "Thl" (Kowalski efal. 1963). The expression 1 X RTD (routine test dilution) means the highest dilution of phages pro- ducing confluent lysis of host strains after a 24-h incuba-
tion at 28' (Craigie and Yen 1938). The 1 X RTD ranged from 3 X 103 to 2.4 X 106 plaque-forming units in a standard drop. For typing, concentrations equal to 4 X RTD of phages were applied.
Typing Tec~rlli~lre Liquid cultures of bacteria were grown in medium 5
with aeration at 28" for 12-14 h in fast-growing strains and for 24-36 h in slowly growing ones. The bacterial cultures were then poured on dried plates with medium Thl and dried again. Plates thus prepared were spotted with 4X RTD of the phage stocks. The results of phage activities were tested after 18, 24, and 48 h of incubation at 28'.
Results The phages used for typing showed different
morphology of plaques. The photographs 1 to 4 illustrate the plaque types observed among the 28 phages studied. One type of plaques (Fig. 1) produced the phages multiplied on clover, pea, and vetch bacteria. The second type of plaques (Fig. 2) corresponded to phages multiplied on horse bean bacteria; the third type (Fig. 3) the phages propagated on bean bacteria; and fourth type (Fig. 4) the phages of lucerne and lupine bacteria.
Lysis of Rhizobium strains by phages was completed mostly after 18 h of incubation. After the second readings of results at 24 h the plates were kept further at room temperature. Con- tinued incubation at this temperature gave a clearer picture of the lyse. Negative or doubtful results of lysis were controlled after 24 h and 48 h of incubation of the plates and sometimes after a longer period.
In phage-typing tests 31 phage types were distinguished in the three groups of Rhizobium strains.
Group I of' Rhizobiunz Strains Out of 146 strains of R. trifolii; R . legumino-
sarum for pea, vetch, horse bean, and Latlzyrus spp.; and R. phaseoli, 96 strains were sensitive to five phages. They formed 18 different phage types (Table Ia). Each of the five phages used for typing in this group of bacterial strains had a different host range. Phage No. 2H lysed the greatest number of strains, i.e. 81 strains from phage type 11 + I ~ + I ~ + I ~ + I ~ + I ~ + ~ ~ + I I I + I I ~ +I14+117.
Morphological mutants of Rhizobium, used by us in the typing, showed slight differences in phage sensitivity as compared to their original forms. Thus morphological mutant No. Fla of
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
AL
GA
RY
on
10/0
1/12
For
pers
onal
use
onl
y.
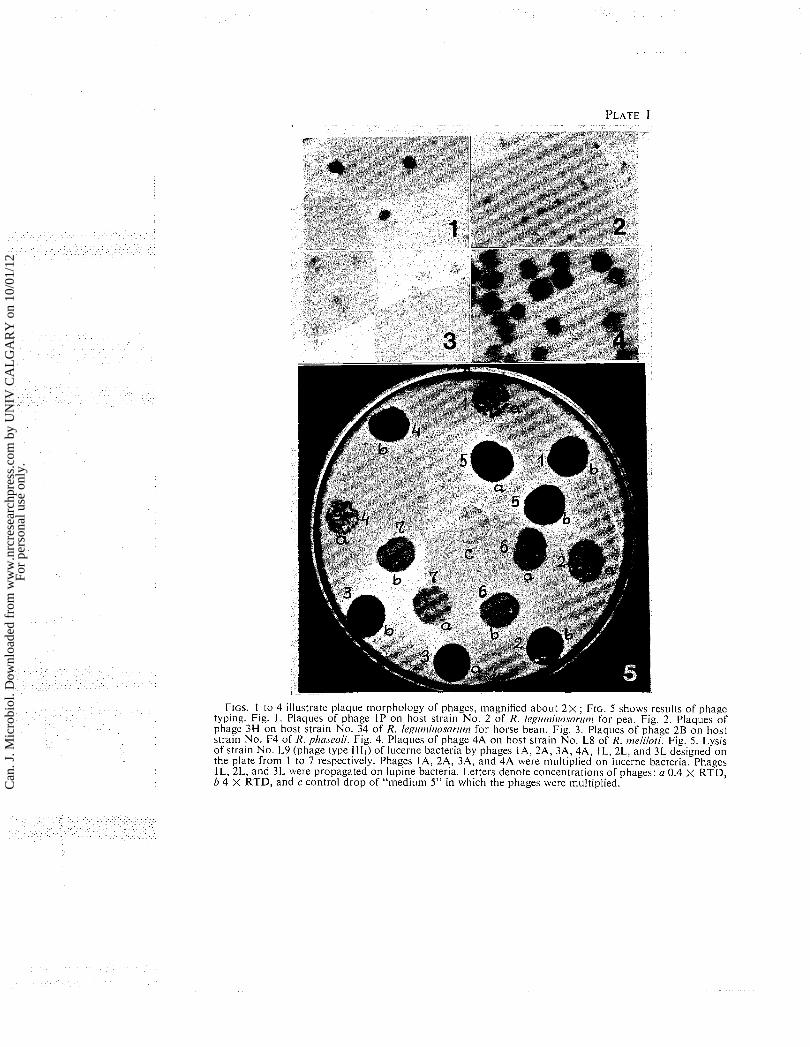
FIGS. 1 to 4 illustrate plaque morphology of phages, magnified about 2 X ; FIG. 5 shows resul(s of phage typing. Fig. 1. Plaques of phage I P on host strain No. 2 of R. Ic.~lit71i1losorlit~1 for pea. Fig. 2. P l a q ~ ~ e s of phage 3H on host strain No. 34 of R. l e g i ~ t i ~ i t r o ~ o r i t ~ for horse bean. Fig. 3. P l a q ~ ~ e s of phage 2B on host strain No. F4 of R. phoseoli. Fig. 4. P l a q ~ ~ e s of phage 4A on host strain No. L8 of R. t~ielilofi. Fig. 5 . Lysis of strain No. L9 (phage type 1111) of lucerne bacteria by phages IA, 2A, 3A, 4A, I L, 2L. and 3L designed on the plate from 1 to 7 respectively. Phages IA, 2A, 3A, and 4A were ni~lltiplied on 1ucel.ne bacteria. Phagcs l L , 2L, and 3L were propagated on lupine bacteria. Letters denote concentrations of phages: (1 0.4 X RTD, b 4 X RTD, and c control drop of "mediunl 5" in which the phages were multiplied. C
an. J
. Mic
robi
ol. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
CA
LG
AR
Y o
n 10
/01/
12Fo
r pe
rson
al u
se o
nly.
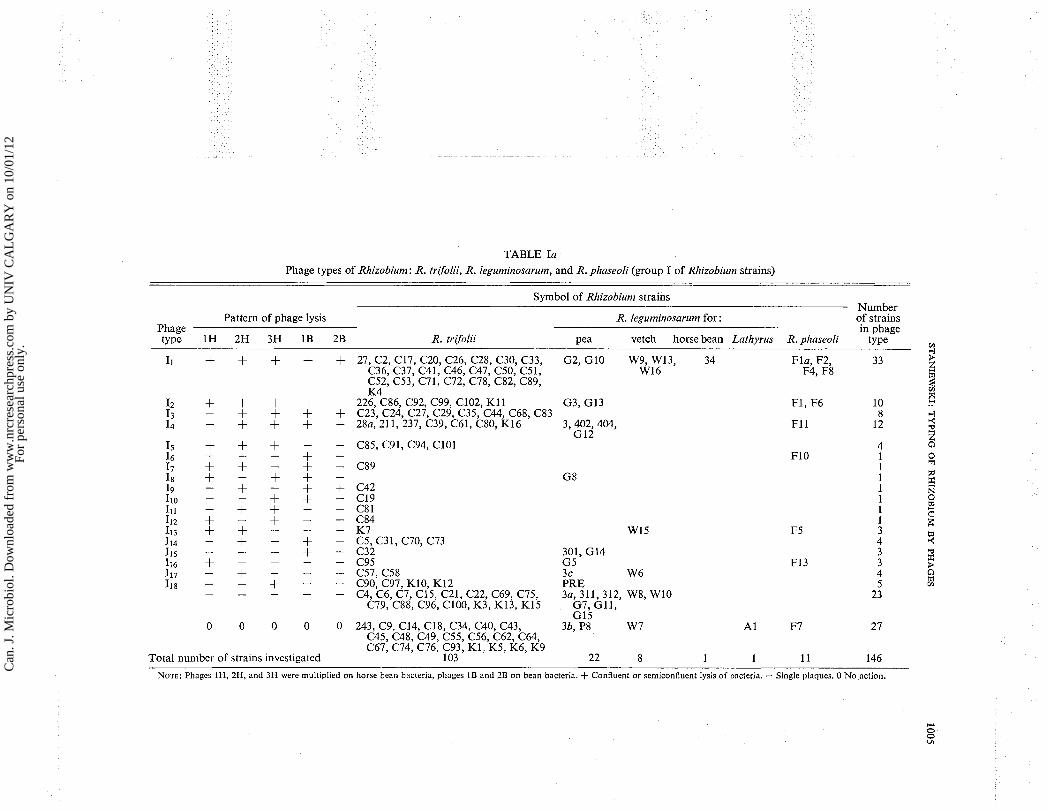
TABLE Ia Phage types of Rhizobiun~: R. trgolii, R. leguminosarum, and R. phaseoli (group I of Rhizobiurn strains)
Symbol of Rhizobium strains Numher . -. . . - . .
Pattern of phage lysis R. legumir~osarum for : of strains Phage in phage type 1H 2H 3H 1B 2B R. tr~yolii Pea vetch horse bean Latlvrus R. phaseoli type
11 + + + + + 27, C2, C17, C20, C26, C28, C30, C33, G2, G I 0 W 9 , W13, 34 Fla, F2, 33 C36, C37, C41, C46, C47, C50, C51, W16 F4, F8 C52, C53, C71, C72, C78, C82, C89, K 4
C85, C91, C94, ClOl
C57, C58 3 c W 6 C90, C97, K10, K12 PRE C4, C6, C7, C15, C21, C22, C69, C75, 3a, 311, 312, W 8 , W10
C79, C88, C96, C100, K3, K13, K15 G7, GI 1 , GI5
0 0 0 0 0 243, C9, C14, CIS, C34, C40, C43, 3b, P8 W 7 A 1 F7 27 C45, C48, C49, C55, C56, C62, C64, C67, C74, C76, C93, K1, K5, K6, K9
Total number of strains investigated 103 22 8 1 1 11 146
NOTE: Phages 11-1, 2H, and 3H were multiplied on horse bean bacteria, pliages 1B and 2B on bean bacteria. + Confluent or semiconfluent lysis of bacteria. - Single plaques. 0 No action.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
AL
GA
RY
on
10/0
1/12
For
pers
onal
use
onl
y.

1006 CANADIAN JOURNAL OF MICROBIOLOGY. VOL. 16, 1970
bean bacteria belonged to phage type 11, while its original strain No. F1 to type I2 (Table Ia).
Group 11 of Rhizobiurn Strains This group included R. trifolii (10 strains) and
R. legumi~zosarun~ for pea (2 strains) and vetch (1 strain), which were typed by 16 phages propa- gated on clover, pea, and vetch bacteria. Among these strains three phage types were distin- guished (Table Ib).
The original strain No. 23 of clover bacteria and its mutant No. 23e belonged to phage type E12; however, two other mutants (No. 23c and 23f) derived from the same culture of strain No. 23 belonged to phage type 111. Strain No. 36 of vetch bacteria was resistant to 16 phages while its mutant was sensitive to them (Table Ib).
Group 111 of Rhizobium Strains Various sensitivities of this group of Rhizobium
strains to seven phages constituted a basis for identification of 10 phage types (Table Ic, Fig. 5). The greatest number of bacterial strains was lysed by two phages: No. 3A (46 strains, i.e. strains from phage types 1111 to 1118) and No. 1L (47 strains, i.e. strains from phage types 1111 to IIIs + IIIlo). Strain No. L4 of lucerne bacteria was non-sensitive to seven phages, but its mor- phological mutant No. L4a (phage type 1118) showed sensitivity to two phages (Table Ic). Three phages (No. lL, 2L, and 3L) lysed most of the R. meliloti strains, but (except for the host strain) they did not lyse lupine bacteria.
Discussion The isolated rhizobiophages showed a wide
host range, They were not as specific as the phages for S. typhi or pathogenic staphylococci. However, the phages studied, like those obtained by Conn et al. (1945), did not act on Agrobac-
terium radiobacter; Arthrobacter globiforrne, or some other species of bacteria not belonging to Rhizobium (Staniewski 1970). On the other hand Roslycky et al. (1962) found phages lysing not only A. radiobacter but also R. meliloti and R. trifolii.
Both Laird (1932) and Conn et al. (1945) claimed that it was impossible to differentiate between Rhizobiunz species on the basis of bac- teriophage sensitivity, because phages used by them possessed an interspecies specificity.
A classification of the studied Rhizobiz1rr1 strains into phage-sensitive types proved to be possible because of the application of the "rou- tine test dilution" method and solid medium. Thus, a number of types both in the composed group of R. trifolii, R. leguminosarum, and R. phaseoli strains (Table Ia and Ib) as well as among strains of R. meliloti (Table Ic) were established.
The typing of rhizobia by diluted phages to 4 X RTD gave better results than the deter- mination of bacterial sensitivity to non-diluted phage preparations (Staniewski 1970). About 327, of the strains of Rhizobiunz studied were not sensitive to this concentration of phages.
Phages multiplied on horse bean, bean, lu- cerne, and lupine bacteria acted in a more dif- ferentiated way on the investigated strains of Rhizobium than phages propagated on clover, pea, and vetch bacteria. With the aid of phages of horse bean and bean bacteria (Table Ia) as well as with phages of lucerne and lupine bac- teria (Table Ib) the greatest number of phage types among the studied strains of Rhizobiurrz were distinguished.
Among the results obtained it is interesting to note the geographical distribution of strains belonging to the same phage type. Graham (1964)
TABLE I D R. !r.ifolii and R. Iegwt~itlosnrwn (group I1 of RlrizoDi~ml strains)
Symbol of RRizobi~m~ strains Number of
Pattern of phage lysis R. Ieg~~trrinosar~rttl for: strains Phage in phage type IC* 6C IP IV 2V R. trifolii Pea vetch type
NOTE: Phages 1C and 6C were multiplied on clover bacteria, phage 1P on pea bacteria, phages 1V and 2V on vetch bacteria. *Identical lytic patterns with those of 11 other phages: 2C, 3C, 4C, 5C, 7C, 8C, 9C multiplied on clover bacteria; 2P, 3P, 4P multiplied on pea
bacteria; 3V multiplied on vetch bacteria.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
AL
GA
RY
on
10/0
1/12
For
pers
onal
use
onl
y.
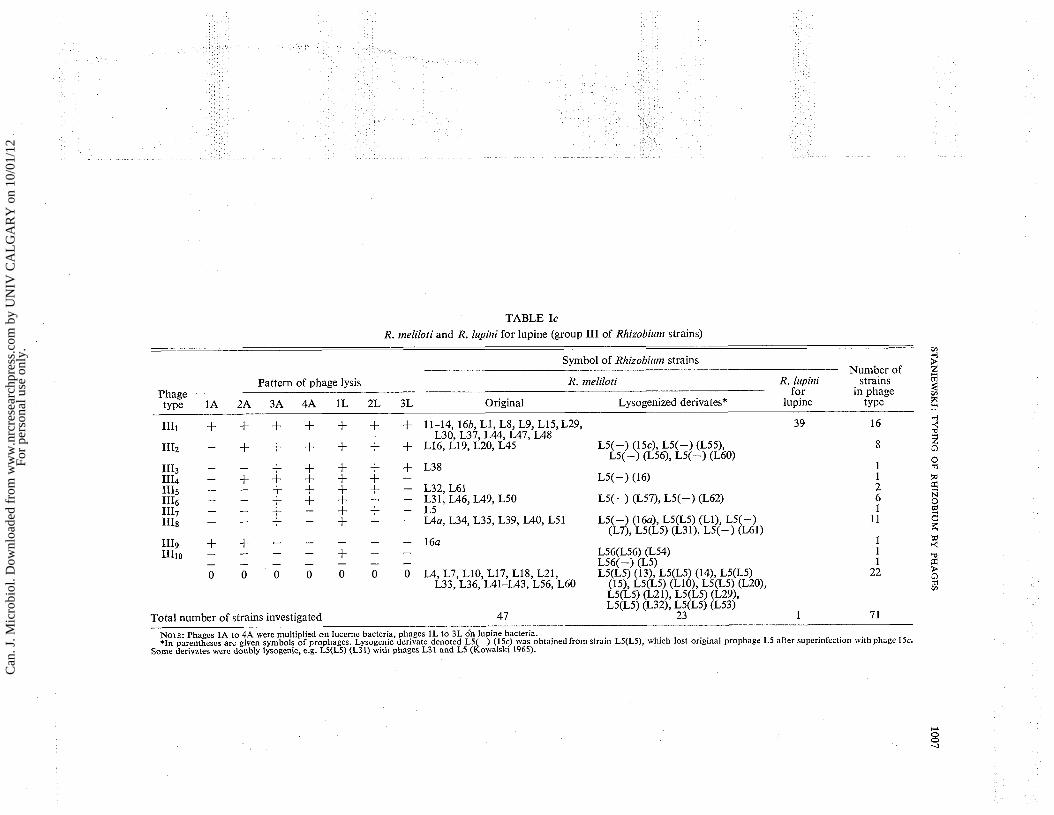
TABLE Ic R. 111eliloti and R. lupir~i for lupine (group I11 of Rhizobiurn strains)
- Symbol of Rllizobiurn strains
Number of Pattern of phage lysis R. meliloti R. lupini strains
Phage for in phage type 1A 2A 3A 4A 1L 2L 3L Original Lysogenized derivates* lupine type
. . . . .
Total number of strains investigated 47 23. . . 1 7 1
NOTE: Phages 1A to 4A were multiplied o n lucerne bacteria, phages 1L to 3L dn lupine bacteria. *In parentheses are given symbols of prophages. Lysogenic derivate denoted L5(-) ( 1 5 ~ ) was obtainedfrom strain L5(L5), which lost original prophage L5 after superinfection with phage 15c.
Some derivates were doubly lysogenic, e.g. L5(L5) (L31) with phages L31 and L5 (Kowalsk~ 1965).
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
AL
GA
RY
on
10/0
1/12
For
pers
onal
use
onl
y.

1008 CANADIAN JOURNAL OF MICROBIOLOGY. VOL. 16, 1970
established considerable similarities in terms of various properties shared by several R. trifolii strains from South Africa and some European strains of clover and pea bacteria. In our studies, out of 12 strains of R. trifolii isolated from var- ious sites of the same locality (Czechow Dolny, Poland), 6 strains (Nos. C46, C47, C50, C51, C52, and C53), belonged to the same phage type I1 (Table Ia). In a number of cases two to four strains isolated in the same locality repre- sented the same phage type. However, it hap- pened frequently that the same phage type was found to occur in native strains and in some strains isolated in foreign laboratories. Thus, for example, among five Rhizobium strains received from Sweden and sensitive to phages, three (Nos. 237, 402, and 404) belonged to phage I4 and one (No. 226) to phage type 12. Phage type 14, apart from the three Swedish strains, also included one strain (No. 211) from the United States, one (No. 280) from France, and one (No. 3) from the Soviet Union (Table Ia). Cases of considerable differences of phage types were found among strains isolated in the same local- ity. Thus, for example, each of the eight strains of R. trifolii (Nos. C78, C86, C83, C80, C85, C89, C84, and C90) isolated in the same locality (Wolka Abramowicka, Poland) belonged to a different phage type (11, 12, 13, 14, 15, 17, 112, and II8 respectively (Table Ia)).
Similar cases of relation between the belong- ing of Rlzizobiuilz strains to phage type and their geographical distribution were found also in the group of strains I1 and 111. Thus it seems that most strains originating from given areas belong to the same phage type.
The lysogeny of Rhizobium may influence its phage type. Some strains of lucerne bacteria were found (Staniewski and Kowalski 1965) to change its phage type after lysogenization. A change of phage type took place in cases when the lysogenizing phage replaced the original prophage in the lysogenic strain of lucerne bac- teria. Besides that, the lysogenized strain became either sensitive to more phages or lost sensitivity
serological relationship of Rlrizobium and their sensitivity to phages. Rhizobi~rnz strains classified to group I1 in Drozaliska's studies (1966) showed an antigenic relationship. However, in some cases, strains belonging to different phage types were agglutinated by the same antiserum. Kleczkowski and Thornton (1944) did not find any correlation between antigenic structure and phage sensitivity.
From studies of Lorkiewicz and Dusiliski (1963) and Lorkiewicz (personal communica- tion), who used a considerable number of mor- phological mutants of R. trifolii and R. meliloti, and from the author's results presented here it can be concluded that rough or semirough forms of Rhizobiu~~z which produce less slime were more sensitive to phage action.
Like the results reported by Kleczkowska (1957), the present study did not establish a correlation between the ability of strains of nodule bacteria to fix free nitrogen and their sensitivity to phages.
The author's results make him believe that the method of typing of bacteria by phages may be applied in attempts to determine relation- ships between different species of Rhizobium as well as in a more detailed differentiation of strains among Rhizobiuin species. Standard phage cultures against Rhizobirml, propagated on closely determined hosts, like phages for some species of salmonella and staphylococci, would provide results comparable with those of different investigators working on phage typing of Rhizobium.
Acknowledgments
This research was financed in part by a grant from the United States Department of Agricul- ture, Agricultural Research Service, for which the author expresses his thanks. Thanks are also due to the late Professor Jadwiga Ziemiecka and Professor Zbigniew Lorkiewicz for helpful discussion and advice.
to some phages. The phage type of lysogenized ADAMS, M. H. 1959. Bacteriophages. Interscience Publ. Inc. New York. nzeliloti strains On the properties BREED, R , S. , E. G. D. MURRAY, and N. R. SMITH, 1957.
of the bacterial strain and the nature of the in- Bergey's Manual of determinative bacteriology. 7th ed. troduced temperate phage. Baltimore.
studies of ~ ~ ~ ~ ~ - , ~ l l and villcent (1954) and CONN, H. J., E. J. B O ~ C H E R , and C. RANDALL. 1945. The value of bacteriophage in classifying certain soil
Vincent (1962) showed a correlation between bacteria. J. Bacteriol. 49: 359-373.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
AL
GA
RY
on
10/0
1/12
For
pers
onal
use
onl
y.

STANIEWSKI: TYPING OF RHIZOBIUM BY PHAGES 1009
CRAIGIE, J., and C. H. YEN. 1938. The demonstration of types of B. typhosus by means of preparations of type I1 Vi-phage. Can. J. Public Health, 29: 448484.
DEMOLON, A. 1951.Contribution a 1'6tude de la symbiose bactkrienne chez les legumineuses. Rev. Gen. Bot. 58: 562-572.
DOROSINSKII, L. M. 1941. Vllyanie bakteriofaga na razvitie klevera. Mlkrobiologiya, 10: 208-215.
DROZANSKA, D. 1966. Studies on the relationships of somatic antigens among different groups of Rhizobium. Acta Microbiol. Pol. 15: 323-334.
FRED, E. B., I. L. BALDWIN, and E. McCoy. 1932. Root nodule bacteria and leguminous plants. Unlv. WIS- consin Studies Sci. 5.
GERRETSEN, F. C., A. GRYNS, J. SACK, and N. L. SOHNGEN. 1923. Das Vorkommen eines Bakteriophagen in den Wurzelknollchen der Legum~nosen. Zentralbl. Bakteriol. Parasitenk. Infekionskr. 11, 60: 311-316.
GRAHAM, P.H. 1964. The application of computer techniques to the taxonomy of the root-nodule bacteria of legumes. J. Gen. Microbiol. 35: 511-517.
JUHASZ, S. E., and R. BONICKE. 1965. Possible classifica- tion of rapldly growlng mycobacteria on the basis of their phage susceptibility. Can. J. Microbiol. 11: 235-241.
KLECZKOWSKA. J. 1957. A study of the distribution and the effects of bacteriophage of root nodule bacteria in the soil. Can. J. Microbiol. 3: 171-180.
KLECZKOWSKI, A., and H. G. THORNTON. 1944. A sero- logical study of root nodule bacteria from pea and clover inoculation groups. J. Bacteriol. 48: 661-672.
KOWALSKI, M. 1965. The phenomenon of lysogeny of Rhizobium meliloti. Folia Soc. Sci. Lublinensis, Sec. B, 5(6L
KOWALSKI, M., R. STANIEWSKI, and Z. HALABIS. 1963. The influence of chemical agents on Rhizobiurn bacteriophages. Acta Microbiol. Pol. 12: 175-179.
LAIRD, D. G. 1932. Bacteriophage and the root nodule bacteria. Arch. Mikrobiol. 3: 159-193.
LORKIEWICZ, Z., and M. Dus~f i s~r . 1963. The antigenic structure of Rhizobium trifolii mutants. Acta Micro- biol. Pol. 12: 119-124.
MARSHALL, K. C., and J. M. VINCENT. 1954. Relation- ship between the somatic antigens of Rhizobium trifolii and susceptibility to bacteriophage. Aust. J. Sci. 17: 68-69.
MEYNELL, E. 1964. The significance of bacteriophage in bacterial classification. J. Gen. Microbiol. 36: 461469.
ROSLYCKY, E. B., 0. N. ALLEN, and E. McCoy. 1962. Phages for Agrobacterium radiobacter with reference to host range. Can. J. Microbiol. 8: 71-78.
STANIEWSKI, R. 1970. Relationship among different Rhizobium strains determined by phage lysis. Acta Microbiol. Pol. Ser. B, 2(19): 1: 3-12.
STANIEWSKI, R., and M. KOWALSKI. 1965. Effect of lysogenization on variability of phage type in Rhizo- bium meliloti. Acta Microbiol. Pol. 14: 231-236.
STANIEWSKI, R., M. KOWALSKI, E. GOGACZ, and F. SOKOLOWSKA. 1962. Susceptibility of Rhizobium strains to phages. Acta Microbiol. Pol. 11: 245-254.
STOCKER, B. A. D. 1955. Bacteriophage and bacterial classification. J. Gen. Microbiol. 12: 375-379.
VANDECAVEYE, S. C., and H. KATZNELSON. 1936. Bac- teriophage as related to the root nodule bacteria of alfalfa. J. Bacteriol. 31 : 465-477.
VINCENT, J. M. 1962. Australian studies of the root- nodule bacteria. A review. Proc. Linn. Soc. N.S.W. 87: 8-38.
\ ,
1966. Lysogeny in Rhizobium meliloti. Acta WENTWORTH, B. B. 1963. Bacteriophage typing of the Microbial. Pol. 15: 119-128. staphylococci. Bacteriol. Rev. 27: 253-272.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
AL
GA
RY
on
10/0
1/12
For
pers
onal
use
onl
y.