Embed Size (px)
Citation preview

Two-timing zinc finger transcriptionfactors liaising with RNAJon Burdach1,2, Mitchell R. O’Connell2, Joel P. Mackay2 and Merlin Crossley1
1 School of Biotechnology and Biomolecular Sciences, University of New South Wales, NSW 2052, Australia2 School of Molecular Biosciences, University of Sydney, Sydney, NSW 2006, Australia
Review
Classical zinc fingers (ZFs) are one of the most commonprotein domains in higher eukaryotes and have beenknown for almost 30 years to act as sequence-specificDNA-binding domains. This knowledge has come, how-ever, from the study of a small number of archetypalproteins, and a larger picture is beginning to emerge thatZF functions are far more diverse than originally sus-pected. Here, we review the evidence that a subset of ZFproteins live double lives, binding to both DNA and RNAtargets and frequenting both the cytoplasm and thenucleus. This duality can create an important additionallevel of gene regulation that serves to connect transcrip-tional and post-transcriptional control.
ZF proteins as multifunctional regulatorsGene expression comprises various layers (such as tran-scription, splicing, mRNA export, mRNA stability andtranslation) that come together to determine the expres-sion of any gene. These layers are not independent but areinextricably linked, and this crosstalk helps to generatethe level of regulatory complexity required for sophisti-cated biological systems. Proteins that act on multiplelayers of gene regulation are well suited to facilitate theconnections required for an effective regulatory system. Inthis review, we focus on one remarkable group of multi-functional regulators: namely, transcription factors thatcontact both DNA and RNA through their ZF domains(Table 1). These proteins can interact not only with theirconventional DNA targets but also with RNA transcripts,and can thereby connect transcriptional and post-tran-scriptional regulatory networks, often utilizing feedbackmechanisms.
Given the multifunctional nature of proteins, it is notunexpected that some proteins possess domains dedicatedto DNA or RNA binding. Indeed, there are several exam-ples of proteins involved in transcription, RNA processing,RNA sequestration, translational regulation and otherprocesses that have both DNA and RNA binding activitythrough separate domains [1–9]. Here we examine aunique group of multifunctional regulators that can usea single classical ZF domain (Box 1) for both DNA and RNAbinding. The dual function of the ZF domains containedwithin these transcription factors seems to allow for cross-regulation of DNA- and RNA-binding events. It is notablethat, in several cases discussed here, RNA binding resultsin the inhibition of the ‘normal’ transcription factor activity
Corresponding author: Crossley, M. ([email protected]).
0968-0004/$ – see front matter � 2012 Elsevier Ltd. All rights reserved. doi:10.1016/j.tibs.2012.
of the protein. As the same domain is used for mediatingboth DNA and RNA interactions, binding to one ligandphysically precludes binding to the other. In addition, theresultant protein–RNA complexes are often observed to belocalized to the cytoplasm, further reducing the potentialfor DNA-directed transcriptional activity. Sequestration ofthe transcription factor in this manner represents a usefulstrategy for effecting short-term adjustments to transcrip-tional activity without the need to alter protein levels. Inthe case of transcription factor for polymerase III (TFIIIA)and transformer-1 (TRA-1), this capability has allowedfeedback loops to be established, thereby connecting tran-scriptional and post-transcriptional gene regulatory pro-cesses. RNA interactions by these multifunctionalregulators can also lead to the coupling of transcriptionand translation through, for example, direct regulation ofthe translation of the bound mRNA.
In the following sections, we describe the known ZFtranscription factors that bind both DNA and RNA, discussthe functional implications of dual binding in the broadercontext of gene regulation and examine the structuralbasis for these DNA and RNA interactions.
In the beginning there was TFIIIATFIIIA is perhaps the best-studied example of a multifunc-tional classical ZF transcription factor. It was initially iso-lated from a 7S RNP complex as an unknown 40-kDa proteinbound to 5S rRNA in Xenopus laevis oocyte cell extracts [10].Remarkably, the protein was soon found to have a dual roleas both a transcription factor capable of activating 5S rRNAgene expression and as an RNA-binding protein, associatingwith the 5S rRNA transcript itself [11,12]. TFIIIA is an RNApol III transcription factor that drives expression of 5SrRNA by binding to the internal 5S rRNA promoter. Fol-lowing transcription, TFIIIA then binds to the resultanttranscript to form a 7S RNP complex, facilitating exportfrom the nucleus (Figure 1) [13]. In contrast to most RNA polII transcription factors, RNA pol III transcription factorssuch as TFIIIA often bind to an internal promoter. Thisdifference could be significant because, during transcrip-tion, the proximity of TFIIIA to the nascent transcript mightfacilitate the interaction of TFIIIA and the 5S rRNA tran-script. Following nuclear export, TFIIIA then remains in thecytoplasm until the 5S rRNA is released for ribosome as-sembly. Release of the 5S rRNA allows TFIIIA to return tothe nucleus, where it can activate the expression of addi-tional 5S rRNA. The binding of TFIIIA to the 5S rRNAtranscript establishes an autoregulatory feedback loop.
02.001 Trends in Biochemical Sciences, May 2012, Vol. 37, No. 5 199
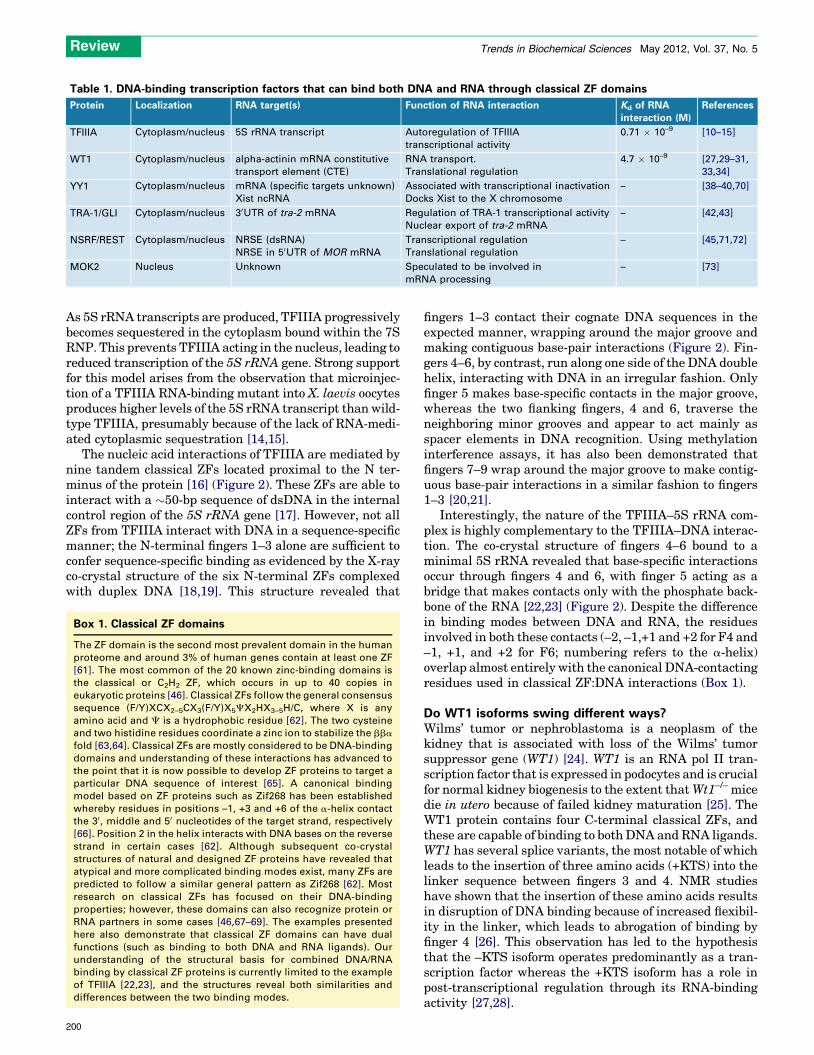
Table 1. DNA-binding transcription factors that can bind both DNA and RNA through classical ZF domains
Protein Localization RNA target(s) Function of RNA interaction Kd of RNA
interaction (M)
References
TFIIIA Cytoplasm/nucleus 5S rRNA transcript Autoregulation of TFIIIA
transcriptional activity
0.71 � 10–9 [10–15]
WT1 Cytoplasm/nucleus alpha-actinin mRNA constitutive
transport element (CTE)
RNA transport.
Translational regulation
4.7 � 10–9 [27,29–31,
33,34]
YY1 Cytoplasm/nucleus mRNA (specific targets unknown)
Xist ncRNA
Associated with transcriptional inactivation
Docks Xist to the X chromosome
– [38–40,70]
TRA-1/GLI Cytoplasm/nucleus 30UTR of tra-2 mRNA Regulation of TRA-1 transcriptional activity
Nuclear export of tra-2 mRNA
– [42,43]
NSRF/REST Cytoplasm/nucleus NRSE (dsRNA)
NRSE in 50UTR of MOR mRNA
Transcriptional regulation
Translational regulation
– [45,71,72]
MOK2 Nucleus Unknown Speculated to be involved in
mRNA processing
– [73]
Review Trends in Biochemical Sciences May 2012, Vol. 37, No. 5
As 5S rRNA transcripts are produced, TFIIIA progressivelybecomes sequestered in the cytoplasm bound within the 7SRNP. This prevents TFIIIA acting in the nucleus, leading toreduced transcription of the 5S rRNA gene. Strong supportfor this model arises from the observation that microinjec-tion of a TFIIIA RNA-binding mutant into X. laevis oocytesproduces higher levels of the 5S rRNA transcript than wild-type TFIIIA, presumably because of the lack of RNA-medi-ated cytoplasmic sequestration [14,15].
The nucleic acid interactions of TFIIIA are mediated bynine tandem classical ZFs located proximal to the N ter-minus of the protein [16] (Figure 2). These ZFs are able tointeract with a �50-bp sequence of dsDNA in the internalcontrol region of the 5S rRNA gene [17]. However, not allZFs from TFIIIA interact with DNA in a sequence-specificmanner; the N-terminal fingers 1–3 alone are sufficient toconfer sequence-specific binding as evidenced by the X-rayco-crystal structure of the six N-terminal ZFs complexedwith duplex DNA [18,19]. This structure revealed that
Box 1. Classical ZF domains
The ZF domain is the second most prevalent domain in the human
proteome and around 3% of human genes contain at least one ZF
[61]. The most common of the 20 known zinc-binding domains is
the classical or C2H2 ZF, which occurs in up to 40 copies in
eukaryotic proteins [46]. Classical ZFs follow the general consensus
sequence (F/Y)XCX2–5CX3(F/Y)X5CX2HX3–5H/C, where X is any
amino acid and C is a hydrophobic residue [62]. The two cysteine
and two histidine residues coordinate a zinc ion to stabilize the bba
fold [63,64]. Classical ZFs are mostly considered to be DNA-binding
domains and understanding of these interactions has advanced to
the point that it is now possible to develop ZF proteins to target a
particular DNA sequence of interest [65]. A canonical binding
model based on ZF proteins such as Zif268 has been established
whereby residues in positions –1, +3 and +6 of the a-helix contact
the 30, middle and 50 nucleotides of the target strand, respectively
[66]. Position 2 in the helix interacts with DNA bases on the reverse
strand in certain cases [62]. Although subsequent co-crystal
structures of natural and designed ZF proteins have revealed that
atypical and more complicated binding modes exist, many ZFs are
predicted to follow a similar general pattern as Zif268 [62]. Most
research on classical ZFs has focused on their DNA-binding
properties; however, these domains can also recognize protein or
RNA partners in some cases [46,67–69]. The examples presented
here also demonstrate that classical ZF domains can have dual
functions (such as binding to both DNA and RNA ligands). Our
understanding of the structural basis for combined DNA/RNA
binding by classical ZF proteins is currently limited to the example
of TFIIIA [22,23], and the structures reveal both similarities and
differences between the two binding modes.
200
fingers 1–3 contact their cognate DNA sequences in theexpected manner, wrapping around the major groove andmaking contiguous base-pair interactions (Figure 2). Fin-gers 4–6, by contrast, run along one side of the DNA doublehelix, interacting with DNA in an irregular fashion. Onlyfinger 5 makes base-specific contacts in the major groove,whereas the two flanking fingers, 4 and 6, traverse theneighboring minor grooves and appear to act mainly asspacer elements in DNA recognition. Using methylationinterference assays, it has also been demonstrated thatfingers 7–9 wrap around the major groove to make contig-uous base-pair interactions in a similar fashion to fingers1–3 [20,21].
Interestingly, the nature of the TFIIIA–5S rRNA com-plex is highly complementary to the TFIIIA–DNA interac-tion. The co-crystal structure of fingers 4–6 bound to aminimal 5S rRNA revealed that base-specific interactionsoccur through fingers 4 and 6, with finger 5 acting as abridge that makes contacts only with the phosphate back-bone of the RNA [22,23] (Figure 2). Despite the differencein binding modes between DNA and RNA, the residuesinvolved in both these contacts (–2, –1,+1 and +2 for F4 and–1, +1, and +2 for F6; numbering refers to the a-helix)overlap almost entirely with the canonical DNA-contactingresidues used in classical ZF:DNA interactions (Box 1).
Do WT1 isoforms swing different ways?Wilms’ tumor or nephroblastoma is a neoplasm of thekidney that is associated with loss of the Wilms’ tumorsuppressor gene (WT1) [24]. WT1 is an RNA pol II tran-scription factor that is expressed in podocytes and is crucialfor normal kidney biogenesis to the extent that Wt1–/– micedie in utero because of failed kidney maturation [25]. TheWT1 protein contains four C-terminal classical ZFs, andthese are capable of binding to both DNA and RNA ligands.WT1 has several splice variants, the most notable of whichleads to the insertion of three amino acids (+KTS) into thelinker sequence between fingers 3 and 4. NMR studieshave shown that the insertion of these amino acids resultsin disruption of DNA binding because of increased flexibil-ity in the linker, which leads to abrogation of binding byfinger 4 [26]. This observation has led to the hypothesisthat the –KTS isoform operates predominantly as a tran-scription factor whereas the +KTS isoform has a role inpost-transcriptional regulation through its RNA-bindingactivity [27,28].

5S rRNA
(i) Gene activation
(ii) Export
(iii) Sequestration
Ribosome assembly
(iv) Release
Transcriptionally active Transcriptionally inactive
TFIIIA
TFIIIATFIIIA
TFIIIA
TiBS
Figure 1. Transcription factor for polymerase III A (TFIIIA) binds to the 5S rRNA transcript to facilitate nuclear export and to autoregulate its own transcriptional activity.
Following activation of 5S rRNA gene, TFIIIA binds to the 5S rRNA transcript and facilitates export of the transcript from the nucleus. While bound to the 5S rRNA, TFIIIA is
transcriptionally inactive and becomes localized to the cytoplasm. Once the bound 5S rRNA is released for ribosome assembly, TFIIIA is able to return to the nucleus to
again activate the 5S rRNA gene.
Review Trends in Biochemical Sciences May 2012, Vol. 37, No. 5
Data have begun to emerge that point towards the invivo RNA targets of WT1. In 2006, it was shown that the+KTS but not the –KTS isoform was able to facilitateexport of unspliced mRNA from the nucleus and enhancetranslation of the bound transcript. This activity wasdependent on the binding of WT1 to a constitutive trans-port element (CTE) within the RNA sequence [29]. Inter-estingly, the CTE that was shown to interact with WT1contained a 208-bp motif with remarkable homology to anRNA that had been identified by another group from RNAco-immunoprecipitated with WT1 (+KTS) [29]. In additionto the CTE, a second in vivo RNA target has also beenidentified. Using RNA co-immunoprecipitated with WT1 ina yeast three-hybrid system, WT1 was found to bind to the50 end of the alpha-actinin 1 mRNA (ACTN1) [30,31].Although the physiological role of this interaction is asyet unknown, it could be significant that mutations in theactinin genes can lead to defects in the podocyte cytoskele-ton, an effect also associated with reduced levels of WT1[30]. Overall, what is clear from these studies is that WT1prefers RNA sequences that form stable stem–loop struc-tures. It is apparent that both the size and composition ofthe stem, the length of the loop and the composition of theflanking sequences are all crucial to maximal WT1 binding,suggesting that each finger probably plays a separate rolein the recognition of discrete elements within these RNAspecies.
The structure of WT1 bound to DNA shows that all fourclassical ZFs are involved in DNA binding, with fingers 2–4conferring all of the sequence specificity, again using resi-dues located in the canonical DNA-binding positions on thea-helix of each ZF [32]. Finger 1, on the contrary, isoriented differently with respect to the DNA and onlyprovides non-base-specific contacts to the DNA.
By contrast (and perhaps in accord with the situationdescribed above for TFIIIA), it has been demonstrated thatfinger 1 is absolutely required for interaction between WT1and RNA [33], and further studies have demonstrated that
whereas finger 1 is required, fingers 2 and 3 make majorcontributions to the binding affinity [34]. At the amino acidlevel, it appears that a largely overlapping subset of resi-dues located in the canonical binding positions of a-helicesof ZFs 2 and 3 is also required for its interaction with RNA;the individual amino acid contributions of ZF 1 were not,however, described [34]. We eagerly await a structure ofWT1 bound to an RNA target.
The Yin and Yang of nucleic acid contactsYin-Yang 1 (YY1) is a ubiquitously expressed vertebrateRNA pol II transcription factor that is involved in manycellular processes including differentiation, developmentand proliferation (for reviews see [35,36]). YY1 has four C-terminal classical ZF domains (as is the case with WT1),and can act as an activator and repressor of transcriptiondepending on the gene context [36]. Although YY1 mRNAand protein are abundant throughout oogenesis and em-bryonic development in X. laevis, DNA binding and tran-scriptional activity are only observed at the initiation ofoogenesis and the terminal stages of embryogenesis [37].During the intervening stages, when YY1 is transcription-ally inactive, it is localized to the cytoplasm and is boundwithin mRNP complexes in an RNase-dependent manner[37,38]. In vitro, YY1 binds strongly to polyU20 andpolyA:U duplexes [39], and the protein appears to preferinternal U-rich/A:U-rich regions in maternal mRNA whileshowing little binding to polyA or cap structures [40].Importantly, injection of polyU20 into oocytes disruptsthe formation of YY1:mRNP complexes, indicating thatYY1 RNA-binding activity is responsible for the formationof the YY1 mRNP particles [38]. This suggests that, in asimilar manner to TFIIIA, YY1 might be regulated bysequestration in the cytoplasm through its ability to bindRNA. However, at this stage, neither the detailed RNA-binding specificity of YY1 nor its in vivo target RNA(s) haveyet been defined .Although no mRNA targets for YY1 haveyet been defined, YY1 has recently been shown to bind to
201

N
C
F1
F2F3
F4
F5
F6
F4
F5
F6
N
C
(c)
(a)
(b)
N
Specific DNA binding
Specific RNA binding
F1 F2 F3 F4 F5 F6 F7 F8 F9
TiBS
Figure 2. Transcription factor for polymerase III A (TFIIIA) is able to interact specifically with DNA or RNA through distinct subsets of its zinc finger (ZF) motifs. (a) TFIIIA
recognizes the 5S rRNA gene through specific interactions mediated by ZFs F1–F3, F5 and F7–F9, whereas the interaction with 5S rRNA occurs specifically through ZFs F4 and F6.
(b) X-ray crystal structure of ZF1–ZF6 of TFIIIA in complex with DNA (PDB ID: 1TF6). TFIIIA uses base-specific contacts between F1–3 and 5 and DNA (indicated by red dashes in
boxes) to specifically recognize the 5S rRNA gene promoter. Residues are numbered according to amino acid position and a-helical position (in parentheses). DNA bases are
numbered as in the 5S rRNA gene. Hydrogen bonding and van der Waals interactions are indicated according to [19] (red dashes). (c) X-ray crystal structure of ZFs F4–F6 in
complex with minimal 5S rRNA (PDB ID: 1UN6). F4 uses the N-terminal tip of the a-helix (positions +1 and +2) to make base-specific contacts with 5S rRNA [23]. Residues in the –
1, +1 and +2 positions of the a-helix of finger 6 also make base-specific contacts [22,23]. Interactions indicated by red dashes are as reported previously [22].
Review Trends in Biochemical Sciences May 2012, Vol. 37, No. 5
the ncRNA Xist, facilitating docking of Xist to the X chro-mosome [70].
The crystal structure of YY1 bound to DNA (the adeno-associated virus P5 initiator element) has revealed that, inline with its similarity to WT1, YY1 uses predominatelyfingers 2–4 to make sequence-specific contacts with duplexDNA, whereas finger 1 interacts with the major groove ofthe DNA in a non-canonical manner, making only one base-specific interaction with the DNA by way of a single lysineside chain [41].
TRA-1 worms its way into the cytoplasmTRA-1 is a classical ZF RNA pol II transcription factorcrucial for sexual determination in the nematode Caenor-habditis elegans and is the orthologue of the human GLItranscription factors (GLI1 to 3). Both TRA-1 and GLIproteins contain five classical ZFs in their N-terminalregions. TRA-1 is an important regulator of the sex deter-mination pathway in C. elegans, where it cooperates withTRA-2 to promote female sexual fate. TRA-1 was firstshown to bind RNA through its five classical ZF domainsfollowing the observation that tra-1–/– worms showed dif-ferential expression and subcellular localization of tra-2transcripts. This led to the discovery that TRA-1 binds tothe 30UTR of the tra-2 mRNA [42].
202
Further work revealed that the subcellular localizationof the TRA-1 protein is regulated through interaction witha nuclear retention element located in the 30UTR of the tra-2 mRNA [43]. TRA-1 binding to the 30UTR blocks thenuclear retention element and facilitates the export ofTRA-1 to the cytoplasm (Figure 3). The export of theTRA-1/tra-2 mRNA complex to the cytoplasm is importantfor regulating nuclear TRA-1 levels and forms a switch forselection of sexual fate [43]. When nuclear export of TRA-1was blocked using leptomycin B, TRA-1 accumulated in thenucleus and increased the activation of a green fluorescentprotein (GFP) reporter gene linked to a tra-1 promoter,showing that nuclear export of TRA-1 is important indetermining its transcriptional activity [43], in a mannerreminiscent of TFIIIA. Reduced export of TRA-1 was alsoobserved in a C. elegans strain expressing a tra-2 deletionmutant that does not include the TRA-1 binding site in the30UTR. This strain showed a propensity for female sexselection that could be rescued by the forced expression ofthe tra-2 30UTR [43].
Intriguingly, the crystal structure of the GLI1:DNAcomplex revealed that, whereas most fingers make se-quence-specific contacts with the DNA, finger 1 is againnot involved in any specific interaction with the DNA [44].Furthermore, in vitro RNA binding assays show that the

tra-2 mRNA
TRA-1 transcriptionally active TRA-1 transcriptionally inactive
TRA-1
TRA-1 TRA-1
TRA-2TRA-2
TRA-1
tra-2
tra-2 translation
TiBS
Figure 3. The subcellular localization and transcriptional activity of transformer-1 (TRA-1) is regulated by the tra-2 mRNA, facilitating a switch in sexual fate. Nuclear TRA-1 binds
to a nuclear retention element in the 30UTR of the tra-2 mRNA before being exported from the nucleus in an mRNP complex. Export of the TRA-1/tra-2 complex results in a
decrease in nuclear TRA-1 and a concomitant decrease in TRA-1 transcriptional activity. Export of tra-2 mRNA also results in translation of the tra-2 transcript and, in turn, the
increased levels of TRA-2 protein inhibit the expression of TRA-1. These multiple mechanisms reduce TRA-1 transcriptional activity and drive female sexual fate selection.
Review Trends in Biochemical Sciences May 2012, Vol. 37, No. 5
first two fingers are essential for RNA binding [42]. Duringthe work that established TRA-1 as an RNA-binding pro-tein, the authors also found that the ZFs of GLI1 were ableto bind the 30UTR of the tra-2 mRNA in vitro [42]. Despitethe publication of this finding over 10 years ago, we are notaware of any advances on the roles of human GLI proteinsin RNA binding.
State of the ZF RNAtionEvolution has favored the emergence of highly complex andinterconnected systems to control gene expression. TheDNA/RNA-binding ZF proteins discussed here are impor-tant in connecting transcriptional and post-transcriptionalregulatory networks. The important feature that enablesthese proteins to play this two-faced game is the presenceof a domain that can bind to either DNA or RNA but not toboth simultaneously. As discussed, this characteristicenables the presence of RNA to negatively regulate DNAbinding and can facilitate the establishment of feedbackloops that cross transcriptional and post-transcriptionallayers of the gene regulatory system.
The advantages of this system are readily apparent. Denovo synthesis of new protein is slow, and degradation offunctional protein is metabolically costly for the cell. Thus,sequestration of transcription factors in RNP complexesrepresents a useful strategy to enable short-term adjust-ments to be made to transcriptional activity without need-ing to alter protein levels. A second advantage conferred bythis approach is that the transcription factor can use thecopy number of a particular transcript or group of tran-scripts as an input for a regulatory circuit. Both TFIIIA andTRA-1 are able to directly sense the level of 5S rRNA and tra-2 mRNA, respectively. The level of these transcripts thencontrols the concentration of active transcription factor inthe nucleus. This mechanism of regulation is fairly sophis-ticated (as it uses transcript levels as input information) andis an interesting alternative to post-translational modifica-tion for regulation of transcription factor activity.
Alternative functions for DNA-binding proteins beyondautoregulation through mRNA binding might also exist.Given the abundance of non-coding RNAs (ncRNAs) andthe increasing understanding of their role in gene regula-tion, it would be reasonable to consider these as potentialbinding partners for dual-binding ZF transcription factors.Indeed, the DNA-binding ZF protein, neuron restrictivesilencing factor (NRSF/REST), has been shown to bind toa short ncRNA in an interaction that can modulate theexpression of NSRF/REST target genes [45] and YY1 hasalso recently been reported to bind the ncRNA Xist [70].
Thus far, only a handful of classical ZF proteins havebeen demonstrated to exhibit dual DNA/RNA bindingcapacity. Even so, several common themes emerge betweenthese proteins and the mechanism by which they interactwith both DNA and RNA. A combination of DNA/RNA-binding studies and structural biology has clearly demon-strated that not all ZFs contained within these proteins areequal; that is, different ZFs possess varying degrees ofDNA- and/or RNA-binding affinity and specificity. InTFIIIA, this property is manifested as a discrete partition-ing of the interactions that confer specificity to DNA orRNA binding into different ZF domains. Although less isknown about the dual-binding specificity determinants ofWT1, YY1 and TRA-1/GLI1, it is clear that in all casesfinger 1 appears to be dispensable for specific DNA binding.In the case of WT1, finger 1 is crucial for RNA binding butour current lack of structural and biochemical data meansthat the RNA-binding role of finger 1 in YY1 and TRA-1/GLI1 remains unknown.
It is also clear that, in each case for which we have data,ZFs bind primarily through a similar set of residues at theN-terminal tip of their respective a-helices (namely acombination of residues at positions –1, +1, +2, +3 and+6), irrespective of whether DNA or RNA is the target.The only observable difference lies in their orientation withrespect to their target sequence, such that when interact-ing specifically with DNA, the a-helix generally sits snugly
203

Review Trends in Biochemical Sciences May 2012, Vol. 37, No. 5
along the major groove, whereas when interacting withRNA, the very N-terminal tip of the a-helix points directlytowards the RNA. This situation contrasts with what isknown about protein–protein interactions mediated byZFs, where the ZFs use a far greater diversity of theirsurface to contact protein partners [46]. It is tempting tospeculate that, whereas it is unlikely that DNA and RNAbinding can occur simultaneously, ZF-mediated protein–protein interactions might be able to occur in concert withDNA/RNA binding, if separate binding interfaces can beutilized. In fact, one example of this latter situation hasbeen described: the N-terminal ZF of GATA1 can bindsimultaneously to both DNA and a ZF from the co-regula-tor FOG1 ZF [47], and there are also reports that the ZFs ofboth YY1 and GLI1 can mediate protein–protein interac-tions [47–51].
In light of the data presented in this review, it is reason-able to question whether the dual DNA/RNA-binding phe-nomenon is more common than currently appreciated. Theclassical ZF is widely distributed among the proteomes ofhigher organisms, but the tendency in the past has been tolabel these domains as DNA-binding modules. However, alarge majority of these ZFs occur in tandem repeats within asingle protein containing anywhere from 3 up to 40 ZFmotifs. It has been demonstrated in a range of studies thatproteins containing more than three ZFs use only a subset ofthese to contact DNA specifically [26]. It could well be thatproteins containing more than three ZFs are best suited forusing their additional domains for new functions such asRNA or protein–protein interactions.
Moreover, transcription factors other than ZF proteinscan also possess dual-binding domains. The protein bicoidhas been shown to bind both DNA and RNA through itshomeodomain motif [52], and there are several examples ofother transcription factors binding to RNA, albeit throughdifferent domains from those responsible for DNA binding[1,4,8,9]. It is also logical to consider that multifunctionalDNA/RNA-binding domains could have many and variedroles beyond transcription. Proteins involved in RNA bind-ing, splicing, RNA editing/processing, DNA repair andother nucleic acid binding events could also potentiallyhave dual DNA/RNA-binding functions. The activity ofthese proteins might be similarly regulated by the pres-ence of alternate DNA or RNA ligands.
Elucidating the mechanisms of action of these multi-functional proteins is difficult and could explain the slowprogress in the field to date. However, with the advent ofnew technologies, uncovering the RNA targets and thecomplex functions of these proteins is becoming easier.RNA immunoprecipitation followed by microarray analy-sis (RIP-chip) or high-throughput sequencing (RIP-seq,CLIP-seq) have recently been used to characterize thein vivo targets of several RNA-binding proteins includingHuR, Argonaute proteins, polycomb proteins and Msi1[53–58]. Despite considerable progress with other RNA-binding proteins, no reports exist describing RIP-chip orRIP-seq experiments on an RNA-binding classical ZFtranscription factors. This could be a focus of future re-search to enable a better understanding of how thesehighly abundant proteins fit into the broader theme ofgene regulation. We consider it is also worthwhile to
204
further explore the structure of the RNAs involved inthese interactions. It is now apparent that RNA secondarystructure is widely conserved across the transcriptomeand sophisticated methodologies using deep-sequencingtechnology have recently enabled the high-throughputelucidation of RNA secondary structure (for a review,see [59]). Indeed, transcriptome-wide secondary structureanalysis in yeast has indicated that RNAs that possesssimilar quantities of secondary structure tend to sharebiological function and/or subcellular localization [60]. Itis possible that multi-ZF proteins might recognize partic-ular subsets of structurally conserved cis-regulatory ele-ments in a variety of RNAs, enabling crosstalk betweenfunction/pathway-related RNAs. With the array of newtechnologies now at our disposal, we are hoping for rapidadvances that will reveal the true extent of RNA-bindingactivity by classical ZF proteins and a greater understand-ing of the networks that they regulate.
AcknowledgmentsM.C. and J.M. are supported by research grants from the AustralianResearch Council and the National Health and Medical Research Council.J.B. and M.O’C. are supported by Australian Postgraduate Awards andM.O’C. also holds a Research Scholar Award, from the Cancer InstituteNSW.
References1 Cassiday, L.A. and Maher, L.J. (2002) Having it both ways:
transcription factors that bind DNA and RNA. Nucleic Acids Res.30, 4118–4126
2 Ladomery, M. (1997) Multifunctional proteins suggest connectionsbetween transcriptional and post-transcriptional processes. Bioessays19, 903–909
3 Ladomery, M. and Dellaire, G. (2002) Multifunctional zinc fingerproteins in development and disease. Ann. Hum. Genet. 66, 331–342
4 Wilkinson, M.F. and Shyu, A.B. (2001) Multifunctional regulatoryproteins that control gene expression in both the nucleus and thecytoplasm. Bioessays 23, 775–787
5 Yang, J. et al. (2006) Deletion of the DNA/RNA-binding protein MSY2leads to post-meiotic arrest. Mol. Cell. Endocrinol. 250, 20–24
6 Warraich, S.T. et al. (2010) TDP-43: a DNA and RNA binding proteinwith roles in neurodegenerative diseases. Int. J. Biochem. Cell Biol. 42,1606–1609
7 Bandiera, A. et al. (2005) In-vitro dual binding activity of anevolutionarily related subgroup of hnRNP proteins. Mol. Cell.Biochem. 268, 121–127
8 Challen, C. et al. (2012) Recombinant human MDM2 oncoprotein showssequence composition selectivity. Int. J. Oncol. 40, 851–859
9 Riley, K.J. and Maher, L.J., III (2007) p53–RNA interactions: new cluesin an old mystery. RNA 13, 1825–1833
10 Picard, B. and Wegnez, M. (1979) Isolation of a 7S particle fromXenopus laevis oocytes: a 5S RNA-protein complex. Proc. Natl. Acad.Sci. U.S.A. 76, 241–245
11 Pelham, H.R. and Brown, D.D. (1980) A specific transcription factorthat can bind either the 5S RNA gene or 5S RNA. Proc. Natl. Acad. Sci.U.S.A. 77, 4170–4174
12 Engelke, D.R. et al. (1980) Specific interaction of a purifiedtranscription factor with an internal control region of 5S RNAgenes. Cell 19, 717–728
13 Guddat, U. et al. (1990) Protein-mediated nuclear export of RNA: 5SrRNA containing small RNPs in xenopus oocytes. Cell 60, 619–628
14 Setzer, D.R. et al. (1996) Functional interactions between the zincfingers of Xenopus transcription factor IIIA during 5S rRNA binding.RNA 2, 1254–1269
15 Rollins, M.B. et al. (1993) Role of TFIIIA zinc fingers in vivo: analysis ofsingle-finger function in developing Xenopus embryos. Mol. Cell. Biol.13, 4776–4783
16 Miller, J. et al. (1985) Repetitive zinc-binding domains in the proteintranscription factor IIIA from Xenopus oocytes. EMBO J. 4, 1609–1614

Review Trends in Biochemical Sciences May 2012, Vol. 37, No. 5
17 Churchill, M.E.A. et al. (1990) Mode of interaction of the zinc fingerprotein tfiiia with a 5S RNA gene of Xenopus. Proc. Natl. Acad. Sci.U.S.A. 87, 5528–5532
18 Liao, X.B. et al. (1992) Specific interaction of the first three zinc fingersof TFIIIA with the internal control region of the Xenopus 5S RNA gene.J. Mol. Biol. 223, 857–871
19 Nolte, R.T. et al. (1998) Differing roles for zinc fingers in DNArecognition: structure of a six-finger transcription factor IIIAcomplex. Proc. Natl. Acad. Sci. U.S.A. 95, 2938–2943
20 Clemens, K.R. et al. (1992) Definition of the binding sites of individualzinc fingers in the transcription factor IIIA-5S RNA gene complex.Proc. Natl. Acad. Sci. U.S.A. 89, 10822–10826
21 Hayes, J.J. and Clemens, K.R. (1992) Locations of contacts betweenindividual zinc fingers of Xenopus laevis transcription factor IIIAand the internal control region of a 5S RNA gene. Biochemistry 31,11600–11605
22 Lu, D. et al. (2003) Crystal structure of a zinc-finger–RNA complexreveals two modes of molecular recognition. Nature 426, 96–100
23 Lee, B.M. et al. (2006) Induced fit and ‘lock and key’ recognition of5S RNA by zinc fingers of transcription factor IIIA. J. Mol. Biol. 357,275–291
24 Haber, D.A. et al. (1990) An internal deletion within an 11p13 zincfinger gene contributes to the development of Wilms’ tumor. Cell 61,1257–1269
25 Kreidberg, J.A. et al. (1993) WT-1 is required for early kidneydevelopment. Cell 74, 679–691
26 Laity, J.H. et al. (2000) Molecular basis for modulation of biologicalfunction by alternate splicing of the Wilms’ tumor suppressor protein.Proc. Natl. Acad. Sci. U.S.A. 97, 11932–11935
27 Morrison, A.A. et al. (2008) The post-transcriptional roles of WT1, amultifunctional zinc-finger protein. Biochim. Biophys. Acta 1785, 55–62
28 Hohenstein, P. and Hastie, N.D. (2006) The many facets of the Wilms’tumour gene, WT1. Hum. Mol. Genet. 15, R196–R201
29 Bor, Y.C. et al. (2006) The Wilms’ tumor 1 (WT1) gene (+KTS isoform)functions with a CTE to enhance translation from an unspliced RNAwith a retained intron. Genes Dev. 20, 1597–1608
30 Morrison, A.A. et al. (2006) The Wilms tumour suppressor protein WT1(+KTS isoform) binds alpha-actinin 1 mRNA via its zinc-finger domain.Biochem. Cell Biol. 84, 789–798
31 Nurmemmedov, E. et al. (2010) Kinetic behaviour of WT 1’s zinc fingerdomain in binding to the alpha-actinin-1 mRNA. Arch. Biochem.Biophys. 497, 21–27
32 Stoll, R. et al. (2007) Structure of the Wilms tumor suppressor proteinzinc finger domain bound to DNA. J. Mol. Biol. 372, 1227–1245
33 Caricasole, A. et al. (1996) RNA binding by the Wilms tumor suppressorzinc finger proteins. Proc. Natl. Acad. Sci. U.S.A. 93, 7562–7566
34 Weiss, T.C. and Romaniuk, P.J. (2009) Contribution of individualamino acids to the RNA binding activity of the Wilms’ tumorsuppressor protein WT1. Biochemistry 48, 148–155
35 Castellano, G. et al. (2009) The involvement of the transcriptionfactor Yin Yang 1 in cancer development and progression. Cell Cycle8, 1367–1372
36 Gordon, S. et al. (2006) Transcription factor YY1: structure, function, andtherapeutic implications in cancer biology. Oncogene 25, 1125–1142
37 Ficzycz, A. et al. (2001) Expression, activity, and subcellularlocalization of the Yin Yang 1 transcription factor in Xenopusoocytes and embryos. J. Biol. Chem. 276, 22819–22825
38 Ficzycz, A. and Ovsenek, N. (2002) The Yin Yang 1 transcription factorassociates with ribonucleoprotein (mRNP) complexes in the cytoplasmof Xenopus oocytes. J. Biol. Chem. 277, 8382–8387
39 Belak, Z.R. et al. (2008) Biochemical characterization of Yin Yang 1–RNA complexes. Biochem. Cell Biol. 86, 31–36
40 Belak, Z.R. and Ovsenek, N. (2007) Assembly of the Yin Yang 1transcription factor into messenger ribonucleoprotein particlesrequires direct RNA binding activity. J. Biol. Chem. 282, 37913–37920
41 Houbaviy, H.B. et al. (1996) Cocrystal structure of YY1 bound to theadeno-associated virus P5 initiator. Proc. Natl. Acad. Sci. U.S.A. 93,13577–13582
42 Graves, L.E. et al. (1999) TRA-1 regulates the cellular distribution ofthe tra-2 mRNA in C. elegans. Nature 399, 802–805
43 Segal, S.P. et al. (2001) RNA-regulated TRA-1 nuclear export controlssexual fate. Dev. Cell 1, 539–551
44 Samad, A. and Carroll, R.B. (1991) The tumor suppressor p53 is boundto RNA by a stable covalent linkage. Mol. Cell. Biol. 11, 1598–1606
45 Kuwabara, T. et al. (2004) A small modulatory dsRNA specifies the fateof adult neural stem cells. Cell 116, 779–793
46 Brayer, K.J. and Segal, D.J. (2008) Keep your fingers off my DNA:protein-protein interactions mediated by C2H2 zinc finger domains.Cell Biochem. Biophys. 50, 111–131
47 Koyabu, Y. et al. (2001) Physical and functional interactions betweenZic and Gli proteins. J. Biol. Chem. 276, 6889–6892
48 Lee, J.S. et al. (1993) Evidence for physical interaction between thezinc-finger transcription factors YY1 and Sp1. Proc. Natl. Acad. Sci.U.S.A. 90, 6145–6149
49 Seto, E. et al. (1993) Interaction between transcription factors Sp1 andYY1. Nature 365, 462–464
50 Zhou, Q. et al. (1995) Transcriptional repression of the c-fos gene byYY1 is mediated by a direct interaction with ATF/CREB. J. Virol. 69,4323–4330
51 Kurisaki, K. et al. (2003) Nuclear factor YY1 inhibits transforminggrowth factor beta- and bone morphogenetic protein-induced celldifferentiation. Mol. Cell. Biol. 23, 4494–4510
52 Dubnau, J. and Struhl, G. (1996) RNA recognition and translationalregulation by a homeodomain protein. Nature 379, 694–699
53 Calaluce, R. et al. (2010) The RNA binding protein HuR differentiallyregulates unique subsets of mRNAs in estrogen receptor negative andestrogen receptor positive breast cancer. BMC Cancer 10, 126
54 de Sousa Abreu, R. et al. (2009) Genomic analyses of musashi1downstream targets show a strong association with cancer-relatedprocesses. J. Biol. Chem. 284, 12125–12135
55 Jayaseelan, S. et al. (2011) RIP: an mRNA localization technique.Methods Mol. Biol. 714, 407–422
56 Wang, W.X. et al. (2010) Anti-argonaute RIP-chip shows that miRNAtransfections alter global patterns of mRNA recruitment tomicroribonucleoprotein complexes. RNA 16, 394–404
57 Wen, J. et al. (2011) MicroRNA transfection and AGO-bound CLIP-seqdata sets reveal distinct determinants of miRNA action. RNA 17,820–834
58 Zhao, J. et al. (2010) Genome-wide identification of polycomb-associated RNAs by RIP-seq. Mol. Cell 40, 939–953
59 Wan, Y. et al. (2011) Understanding the transcriptome through RNAstructure. Nat. Rev. Genet. 12, 641–655
60 Kertesz, M. et al. (2010) Genome-wide measurement of RNA secondarystructure in yeast. Nature 467, 103–107
61 Tadepally, H.D. et al. (2008) Evolution of C2H2-zinc finger genes andsubfamilies in mammals: species-specific duplication and loss ofclusters, genes and effector domains. BMC Evol. Biol. 8, 176
62 Wolfe, S.A. et al. (2000) DNA recognition by Cys2His2 zinc fingerproteins. Annu. Rev. Biophys. Biomol. Struct. 29, 183–212
63 Lee, M.S. et al. (1989) Three-dimensional solution structure of a singlezinc finger DNA-binding domain. Science 245, 635–637
64 Rhodes, D. and Klug, A. (1993) Zinc fingers. Sci. Am. 268, 56–59 62–5565 Mandell, J.G. and Barbas, C.F. (2006) Zinc finger tools: custom DNA-
binding domains for transcription factors and nucleases. Nucleic AcidsRes. 34, W516–W523
66 Pavletich, N.P. and Pabo, C.O. (1991) Zinc finger-DNA recognition:crystal structure of a Zif268-DNA complex at 2.1 A. Science 252, 809–817
67 Brown, R.S. (2005) Zinc finger proteins: getting a grip on RNA. Curr.Opin. Struct. Biol. 15, 94–98
68 Hall, T.M.T. (2005) Multiple modes of RNA recognition by zinc fingerproteins. Curr. Opin. Struct. Biol. 15, 367–373
69 Kelly, S.M. et al. (2007) Recognition of polyadenosine RNA by zincfinger proteins. Proc. Natl. Acad. Sci. U.S.A. 104, 12306–12311
70 Jeon, Y. and Lee, J.T. (2011) YY1 tethers Xist RNA to the inactive Xnucleation center. Cell 146, 119–133
71 Kuwabara, T. et al. (2005) The NRSE smRNA specifies the fate of adulthippocampal neural stem cells. Nucleic Acids Symp. Ser. (Oxf.) 87–88
72 Kim, C.S. et al. (2008) Novel function of neuron-restrictive silencerfactor (NRSF) for posttranscriptional regulation. Biochim. Biophys.Acta 1783, 1835–1846
73 Arranz, V. et al. (1997) Human and mouse MOK2 proteins areassociated with nuclear ribonucleoprotein components and bindspecifically to RNA and DNA through their zinc finger domains.Mol. Cell. Biol. 17, 2116–2126
205





























