Embed Size (px)
Citation preview

4600/208 N. Matsuda et al.: EGF receptor and osteoblastic differentiationJ Hum Genet (2002) 47:208–212 © Jpn Soc Hum Genet and Springer-Verlag 2002
SHORT COMMUNICATION
Hiroyasu Iwasa · Masahiko Kurabayashi · Ryozo NagaiYusuke Nakamura · Toshihiro Tanaka
Twenty single-nucleotide polymorphisms in four genes encoding cardiacion channels
Received: January 7, 2002 / Accepted: January 29, 2002
Since mutations in any of these five genes can be detectedin nearly 40% of LQTS families, this syndrome in the re-maining families is likely to be caused by other unknowngenes.
Voltage-gated ion channels play important roles in theexcitement of cardiomyocytes, and all the genes responsiblefor LQTS identified so far belong to this group. Althoughmany channel proteins have been identified at present, onlya few have been reported to be involved in cardiac excite-ment. For example, in mice, decreased expression of Kv1.5,encoded by KCNA5, prolongs QT-intervals of ECGs(London et al. 1998), and hKv�1.3, encoded by alternativesplicing of KCNAB1, is expressed in human ventricle andalters the functional properties of hKv1.5 (England et al.1995). A novel class of auxiliary proteins for Kv4α subunits,K-channel interacting proteins (KChIPs), modulates thetransient inward potassium current (Ito) (An et al. 2000).KChIP2, encoded by KCNIP2, is expressed with a steepgradient across the human ventricle and might be involvedin the transmural gradient of Ito in the free ventricular wall(Rosati et al. 2001), and KChIP2�/� mice are susceptible tocardiac arrhythmias (Kuo et al. 2001). An inward calciumion current generated by an L-type voltage-dependent cal-cium ion channel, encoded by CACNA1C, is implicated inearly afterdepolarizations (January and Riddle 1989), oscil-lations in cell membrane voltage thought to initiatearrhythmias caused by long-QT syndrome.
Here we report single-nucleotide polymorphisms (SNPs)and other genetic variations found in four genes encodingcardiac ion channels along with their allelic frequenciesamong normal and LQTS-affected Japanese subjects.
Subjects and methods
In the course of a search for mutations in these candidategenes in 73 unrelated Japanese LQTS patients, we identi-fied genetic variations. Our screening method was describedpreviously (Itoh et al. 1998). In brief, we prepared genomicDNA from blood samples according to standard protocols,
Abstract We here report 20 novel single-nucleotide poly-morphisms in four genes that are potentially involved in theexcitement of cardiomyocytes: 1 in KCNA5 (encodingKv1.5), 5 in KCNAB1 (encoding Kv�1.3), 5 in KCNIP2(encoding KChIP2), and 9 in CACNA1C (encoding acardiac L-type voltage-dependent calcium ion channel,dihydropyridine receptor). We also examined their allelicfrequencies in Japanese individuals. These data will be use-ful for genetic association studies designed to investigatesecondary long QT syndrome or other circulatory disorders.
Key words Long QT syndrome · Single-nucleotide poly-morphism · Japanese population · Voltage-gated potassiumion channel · KChIP · L-type voltage-dependent calcium ionchannel
Introduction
Long QT syndrome (LQTS), an arrhythmogenic disordercharacterized by prolongation of the QT interval on electro-cardiograms (ECGs), often causes syncope or sudden car-diac death as a result of recurrent and lethal arrhythmia.Five genes in which inherited mutations are responsible forthis syndrome have been identified to date: KCNA9(KVLQT1, KCNQ1), KCNH2 (HERG), KCNE1, KCNE2,and SCN5A (Bennett et al. 1995; Curran et al. 1995; Wanget al. 1996a, b; Splawski et al. 1997; Abbott et al. 1999).
H. Iwasa · Y. Nakamura · T. Tanaka (*)Laboratory of Molecular Medicine, Human Genome Center,Institute of Medical Science, University of Tokyo, 4-6-1Shirokanedai, Minato-ku, Tokyo 108-8639, JapanTel. �81-3-5449-5375; Fax �81-3-5449-5406e-mail: [email protected]
H. Iwasa · M. KurabayashiSecond Department of Internal Medicine, Gunma University Schoolof Medicine, Gunma, Japan
R. NagaiDepartment of Cardiovascular Medicine, Graduate School ofMedicine, University of Tokyo, Tokyo, Japan
B. Jochimsen et al.: Stetteria hydrogenophila 209T
able
1.
Pol
ymer
ase
chai
n re
acti
on (
PC
R)
prim
ers
used
to
cove
r th
e co
ding
reg
ion
of K
CN
A5,
KC
NIP
2, K
CN
AB
1, a
nd C
AC
NA
1C
Ann
ealin
gR
efer
ence
d cD
NA
Ref
eren
ced
Gen
eE
xona
For
war
d pr
imer
Rev
erse
pri
mer
tem
pera
ture
sequ
ence
geno
me
sequ
ence
KC
NA
51.
1A
CC
CC
A G
TC
TC
C C
CA
GA
GA
CG
AT
G A
AG
AA
G G
GG
TC
G58
NM
_002
234
NT
_024
413
1.2
AG
A C
CC
TG
C C
TG
AG
T T
CA
GC
CA
TT
C C
CT
AC
T C
CA
CT
G C
62N
M_0
0223
4N
T_0
2441
3K
CN
IP2
1G
CT
CA
C C
TG
CT
G C
CT
AG
T G
GC
C C
TG
TG
T C
AC
AT
A C
GC
58A
Y02
6329
NT
_030
094
2G
TC
CC
A T
TA
AA
C C
AA
GT
G C
CC
C T
CC
AC
A C
AC
AT
G C
AG
58A
Y02
6330
NT
_030
094
3C
GC
TG
A T
AG
AC
C T
CG
AA
G A
TG
CC
C C
AG
AA
C C
TC
CA
G C
TA
G58
AY
0263
31N
T_0
3009
44,
5C
CC
TC
C C
AT
CA
C A
TG
AG
CT
TT
GG
C C
TG
GA
G A
TC
CT
G58
AY
0263
31N
T_0
3009
46,
7G
AG
TT
C A
GG
GC
C A
GG
AT
CC
CT
CC
A G
CC
TA
C C
CA
CT
C58
AY
0263
31N
T_0
3009
48
CA
A G
TA
CA
C G
TA
CC
C T
GC
AC
TG
C C
CA
GA
G A
GA
CC
T G
AG
62A
Y02
6331
NT
_030
094
9T
CC
CC
G T
CT
GT
G A
CT
CA
GG
GA
AG
C A
CC
AT
A G
CA
GG
A G
58A
Y02
6331
NT
_030
094
KC
NA
B1
1T
TA
AA
G T
TA
AG
C A
CC
GT
GG
CA
AT
C A
GC
AG
T A
CC
TG
T A
T58
Eng
land
et
al. 1
995
Eng
land
et
al. 1
995c
2G
AC
AA
T A
TG
AG
A C
TG
AG
A A
GC
TG
TG
G G
AG
AG
G G
TC
AT
G A
TG
58E
ngla
nd e
t al
. 199
5N
T_0
0555
33
GG
C T
TC
CA
A T
AG
TA
T G
TA
CA
G T
GC
CA
AA
T T
GT
TT
C A
TT
CT
C T
GA
C58
Eng
land
et
al. 1
995
NT
_005
553
4G
AA
TG
T T
GT
CA
A A
CC
AG
C T
CT
GC
CC
C T
CA
CA
T A
GA
CA
T G
58E
ngla
nd e
t al
. 199
5N
T_0
0555
35
AC
C A
AA
CT
G C
AA
AA
T G
AC
TG
AG
A A
GC
CA
T G
AA
AT
T G
TC
AC
58E
ngla
nd e
t al
. 199
5N
T_0
0555
36
AC
A T
GG
GC
T T
GC
TG
T C
AG
AC
A A
AA
AA
C C
CT
CT
C T
CT
AA
G C
58E
ngla
nd e
t al
. 199
5N
T_0
0555
37
AC
A G
AG
TG
T A
CG
TG
G C
TG
AG
TT
C T
GA
AT
T T
GT
TT
G G
TT
GA
T A
C58
Eng
land
et
al. 1
995
NT
_005
553
8G
TG
TA
T A
TT
TT
C C
TG
TG
A G
AG
TG
AC
A A
AA
GT
G A
AA
TG
A A
CC
TG
58E
ngla
nd e
t al
. 199
5N
T_0
0555
39
AA
T G
GA
AA
A T
GG
AA
T G
TC
AG
TG
T A
TT
CT
G A
AC
GG
A T
GA
GC
58E
ngla
nd e
t al
. 199
5N
T_0
0555
310
CT
T A
GT
GA
A A
TC
GG
T G
GT
GA
CT
GC
TT
G G
CT
TT
C A
TC
AG
T G
58E
ngla
nd e
t al
. 199
5N
T_0
0555
311
TG
C A
CT
CC
C A
AG
AA
C T
GA
CT
TC
TT
C C
CT
CC
T T
CC
TA
C T
C58
Eng
land
et
al. 1
995
NT
_005
553
12A
AC
TT
C T
GT
GC
A G
GC
TG
A T
CT
CA
TC
A A
AC
GT
C A
AA
TA
A C
TC
AC
58E
ngla
nd e
t al
. 199
5N
T_0
0555
313
AT
T G
TC
CA
G C
GC
CT
C T
AT
GA
AT
GA
G C
TT
TG
G G
AA
TA
C T
GC
58E
ngla
nd e
t al
. 199
5N
T_0
0555
314
AT
T A
TA
TT
T C
TG
TT
G G
AT
AG
A G
CT
GC
CA
AA
T G
CC
TG
A A
GG
TA
G T
AC
58E
ngla
nd e
t al
. 199
5N
T_0
0555
3
CA
CN
A1C
b1
TC
C T
CT
TC
G T
GG
CT
G C
TC
GT
G G
AA
AA
G G
CA
GG
G A
GC
58N
M_0
0071
9A
C00
5342
2G
AA
GT
G C
CC
CT
G T
TT
TC
T A
TC
TA
GG
CA
GG
T C
CC
AG
G A
GC
AT
G58
NM
_000
719
AC
0060
51.1
3G
GC
AT
T A
AC
TT
C C
TT
GA
C T
CT
GA
AA
A A
GC
AG
A G
AC
TT
G A
G58
NM
_000
719
AC
0060
51.1
4A
TT
TT
T C
TC
TC
T T
TT
CT
A T
TT
CT
GA
AG
GG
A A
AG
AA
A C
AG
AC
A G
TG
58N
M_0
0071
9N
T_0
0071
95
AA
A A
CC
CA
G T
CC
TG
A C
AG
TC
GT
A C
AC
AA
G A
AC
GG
G C
TT
C58
NM
_000
719
NT
_000
719
6C
TT
GG
T T
CA
GT
G A
GT
GG
C T
CG
GG
AG
C G
AT
AG
A G
GG
CA
G58
NM
_000
719
NT
_000
719
7C
CA
TC
C C
AT
CA
A C
CT
CA
T C
CT
C G
TT
AG
G G
TG
AA
A A
GC
TC
58N
M_0
0071
9N
T_0
0071
98
GG
T C
CC
CT
T C
GG
TC
A C
AG
GC
C A
AT
GG
G T
CA
GA
A T
CT
G58
NM
_000
719
NT
_000
719
9G
GA
GC
T G
TC
TG
T G
GA
AA
G T
AG
GT
C T
GT
GT
G C
AG
CA
A G
TG
AC
58N
M_0
0071
9N
T_0
0071
910
GC
T C
TG
CT
T C
CC
TT
T G
AC
TC
GC
T T
TT
CC
A C
CC
AT
G C
TC
58N
M_0
0071
9N
T_0
0071
911
CC
T C
AG
TC
T C
CA
CA
T T
CA
AA
T A
CT
GG
AT
T A
TT
TG
T C
TG
AT
C T
GA
GC
58N
M_0
0071
9N
T_0
0071
912
GA
G G
AG
TT
C A
GG
GG
C A
TC
GG
T G
TC
TC
C A
GC
AC
A G
CA
C58
NM
_000
719
NT
_000
719
13C
GG
GC
T T
CT
GA
C C
AC
TG
CC
A G
GG
CT
G A
TC
TA
G G
AT
G58
NM
_000
719
NT
_000
719
14C
GG
GC
T T
CT
GA
C C
AC
TG
CC
A G
GG
CT
G A
TC
TA
G G
AT
G58
NM
_000
719
NT
_000
719
210 N. Matsuda et al.: EGF receptor and osteoblastic differentiation
Tab
le 1
.C
onti
nued
Ann
ealin
gR
efer
ence
d cD
NA
Ref
eren
ced
Gen
eE
xona
For
war
d pr
imer
Rev
erse
pri
mer
tem
pera
ture
sequ
ence
geno
me
sequ
ence
15T
CT
AA
G G
AC
AG
T T
CA
AT
A G
CT
CA
GC
CA
GA
T T
AC
GG
C G
AC
AG
C58
NM
_000
719
NT
_000
719
16G
AA
CA
C T
GT
TG
G G
GT
CT
G A
CC
CC
TA
C C
AG
AG
T G
AT
GC
T T
AG
58N
M_0
0071
9N
T_0
0071
917
TT
T T
TT
CT
G C
TG
CT
G A
CT
GC
TC
AC
A G
CT
CC
A G
GA
AG
G58
NM
_000
719
NT
_000
719
18C
CT
TC
T C
CC
CT
G T
GA
CT
GA
TC
CC
A G
GT
TA
G G
CA
GT
G58
NM
_000
719
NT
_000
719
19T
GG
AG
T T
AT
TT
A G
AA
TG
G T
GC
TG
CA
C C
TT
CC
C C
AC
CA
T T
GC
58N
M_0
0071
9N
T_0
0071
920
TT
G A
CT
TG
T C
TC
TC
C T
CC
TG
T C
GT
G C
AG
AA
G A
CA
CA
G G
CG
58N
M_0
0071
9N
T_0
0071
921
AT
C C
CA
TC
C C
CA
CC
C T
GT
GG
A G
GA
GC
A G
AA
GG
G G
AT
G55
Sold
atov
199
2N
T_0
0071
922
GA
G C
CA
CT
A A
TC
CA
A T
TA
TG
C T
GA
T G
CA
TT
G C
CC
AG
G T
TG
58N
M_0
0071
9N
T_0
0071
923
CT
G G
TG
TT
C C
TT
TG
T C
CC
TC
CT
A G
CT
GC
A T
GG
CC
C A
TC
58N
M_0
0071
9N
T_0
0071
924
CA
G G
GT
TA
C A
GG
CA
A G
CT
CC
AT
GC
A G
TG
GG
A C
CA
GT
G58
NM
_000
719
NT
_000
719
25G
CA
TC
T C
CT
GA
A G
CC
AC
G T
CC
CA
CA
A C
GG
AG
C C
CA
GA
C58
NM
_000
719
NT
_000
719
26G
AT
TA
C T
GA
AC
A T
CT
CT
G A
TA
CT
C T
GC
TG
CC
C T
CC
CA
C C
TC
TA
C58
NM
_000
719
NT
_000
719
27G
TG
AA
G G
AA
GA
T G
GG
AG
A T
CG
GC
CT
T G
CT
TT
G C
AT
AC
T C
58N
M_0
0071
9N
T_0
0071
928
CA
A G
GT
CA
C A
CA
GC
C A
GT
AA
GA
AG
AC
T G
AC
AG
C C
CA
CT
C T
G58
NM
_000
719
NT
_000
719
29T
AA
CC
C C
AC
TC
T C
CC
CA
T C
AT
G G
CT
GT
G C
TC
CT
C C
TG
58N
M_0
0071
9N
T_0
0071
930
AG
C T
CC
TC
C C
CT
CT
C C
TG
GG
G A
AT
CA
A A
GG
TC
T T
TC
TC
A G
58N
M_0
0071
9N
T_0
0071
931
CT
G G
GG
AG
G G
CA
GA
G T
AA
TA
CT
GC
CT
G C
CC
TC
C C
TT
AT
G58
Sold
atov
199
2N
T_0
0071
932
CT
C T
CC
CC
G G
CT
GC
T C
TG
GG
T T
AG
GG
A C
AG
AC
G C
CA
TG
58N
M_0
0071
9N
T_0
0071
933
GT
G G
GT
CC
T T
GA
TT
G A
CA
TG
A G
CC
A C
AT
GC
A G
TG
AG
A G
CC
AG
58N
M_0
0071
9N
T_0
0071
934
GC
C T
GG
GC
A C
AG
TC
T A
GC
TC
CC
A C
AC
AG
A G
GA
AG
G A
GC
AG
58N
M_0
0071
9N
T_0
0071
935
TC
A T
GG
GA
G T
CT
CC
T G
CA
CG
GG
AA
C T
GG
GG
A G
AG
AC
A58
NM
_000
719
NT
_000
719
36G
CA
TG
G G
AA
GA
C T
GT
TC
A G
CG
GT
TG
C T
CT
GT
G G
AA
AG
T T
AT
C58
NM
_000
719
NT
_000
719
37T
GA
AC
G T
GG
CT
C T
CC
CT
CG
CT
GC
T G
GC
TG
T T
GA
GT
T C
58N
M_0
0071
9N
T_0
0071
938
AT
T G
TA
CT
G T
TC
CC
C A
CA
GA
T C
TC
T C
AG
CT
T G
GG
GA
A T
CA
G58
NM
_000
719
NT
_000
719
39C
TC
TG
A T
GC
CC
T G
TC
CC
T C
CT
C C
CA
TG
A A
CC
CC
G T
TA
TC
58N
M_0
0071
9N
T_0
0071
940
CT
T T
GC
TT
C T
CT
TC
G C
CT
GC
TG
AG
T G
TC
TC
T C
CC
AG
T G
C58
NM
_000
719
NT
_000
719
41C
AC
CA
A G
GG
GC
T G
AG
GA
T C
GT
G C
GG
GA
G T
CC
AG
G G
AG
58N
M_0
0071
9N
T_0
0071
942
TC
C C
CT
CT
C C
AT
AC
G T
CT
CG
GA
CC
T T
CC
TG
G A
GT
GT
G58
NM
_000
719
NT
_000
719
43T
GG
AG
G A
AA
GG
G A
GC
GT
GG
GT
CG
A G
AG
TG
C G
CC
AT
G58
NM
_000
719
NT
_000
719
44C
CC
AC
C C
CT
CC
T T
CT
TG
CC
TG
TC
T C
CT
GA
G G
GT
TG
C A
G58
NM
_000
719
NT
_000
719
45T
AA
AG
A T
GA
CC
T G
AC
CC
T G
TC
AA
A G
GC
AC
A G
AG
AG
G A
AG
C58
Sold
atov
199
2N
T_0
0071
946
CA
G T
TT
CT
G A
TG
TT
T T
TC
TT
C A
TG
GG
TC
A G
CC
TC
A G
CA
GA
G58
NM
_000
719
NT
_000
719
47G
GA
AC
A A
GC
CC
C A
TG
AG
CA
CA
TC
C A
GA
GA
G C
GA
GC
G58
NM
_000
719
NT
_000
719
48A
TG
GT
G G
CT
CT
C T
GG
CT
GG
CC
TG
T C
CA
AA
A G
TG
TG
A G
58N
M_0
0071
9N
T_0
0071
949
TC
G G
CC
AC
T C
CT
AT
T A
AC
TC
A C
CC
T G
CC
CA
T C
TG
CG
A G
TC
58N
M_0
0071
9N
T_0
0071
950
TT
C C
TT
TG
G T
TC
TT
C A
TG
GC
TA
A A
AA
AA
A G
CC
CG
C A
TC
TG
58N
M_0
0071
9N
T_0
0071
9
aE
xons
fol
low
ed b
y do
ts a
nd s
eque
ntia
l num
bers
wer
e su
bdiv
ided
for
ana
lysi
sbE
xon
num
beri
ng a
nd p
osit
ion
of t
he s
plic
ing
junc
tion
s of
CA
CN
A1C
ref
er t
o in
tron
–exo
n bo
unda
ry s
eque
nces
of
CA
CN
L1A
1 is
ofor
m 1
(So
ldat
ov 1
994)
cP
CR
pri
mer
s fo
r th
is e
xon
wer
e de
sign
ed b
ased
on
the
sequ
ence
des
crib
ed in
the
ref
eren
ce
B. Jochimsen et al.: Stetteria hydrogenophila 211
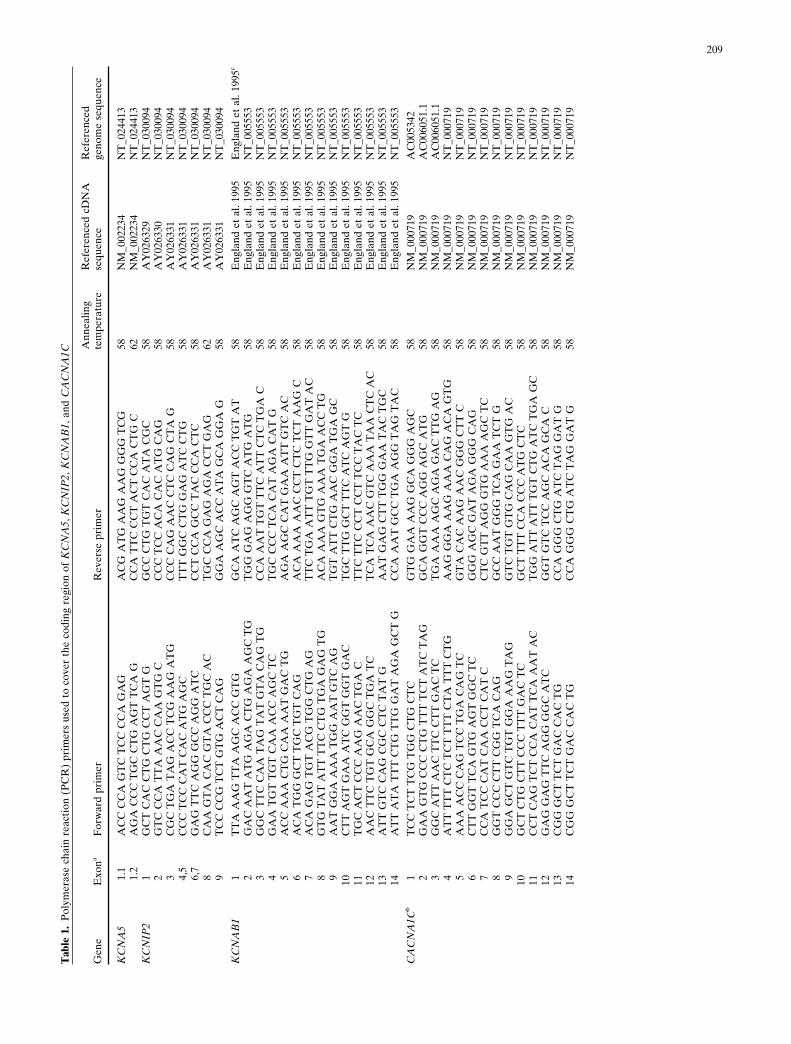
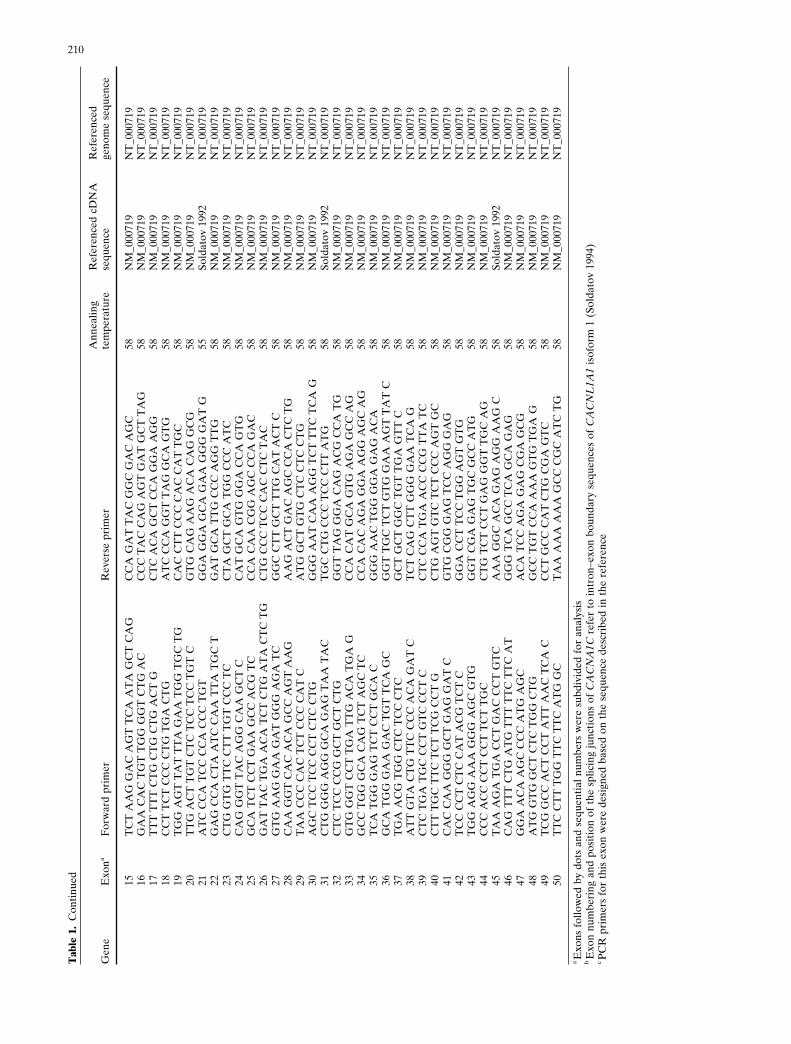
after obtaining informed consent from each participant. Allexons of each gene being scrutinized, as well as flankingintronic sequences, were amplified by newly designed poly-merase chain reaction (PCR) primers to cover the codingregion of each gene (Table 1) and screened by single-strandconformation polymorphism (SSCP) analysis, and aberrantconformers were directly sequenced with ABI 3700 instru-ments (Applied Biosystems, Foster City, CA, USA). Someexons were screened by direct sequencing analysis, depend-ing on the length of the PCR fragment.
To investigate the allelic frequencies for each polymor-phism in our control population, we distinguished allele-specific sequences either by using PCR-RFLP (restrictionfragment length polymorphism) or by hybridizing allele-specific oligonucleotides to DNA from 48 to 96 normal,unrelated individuals in the manner described by Saiki et al.(1986).
Results and discussion
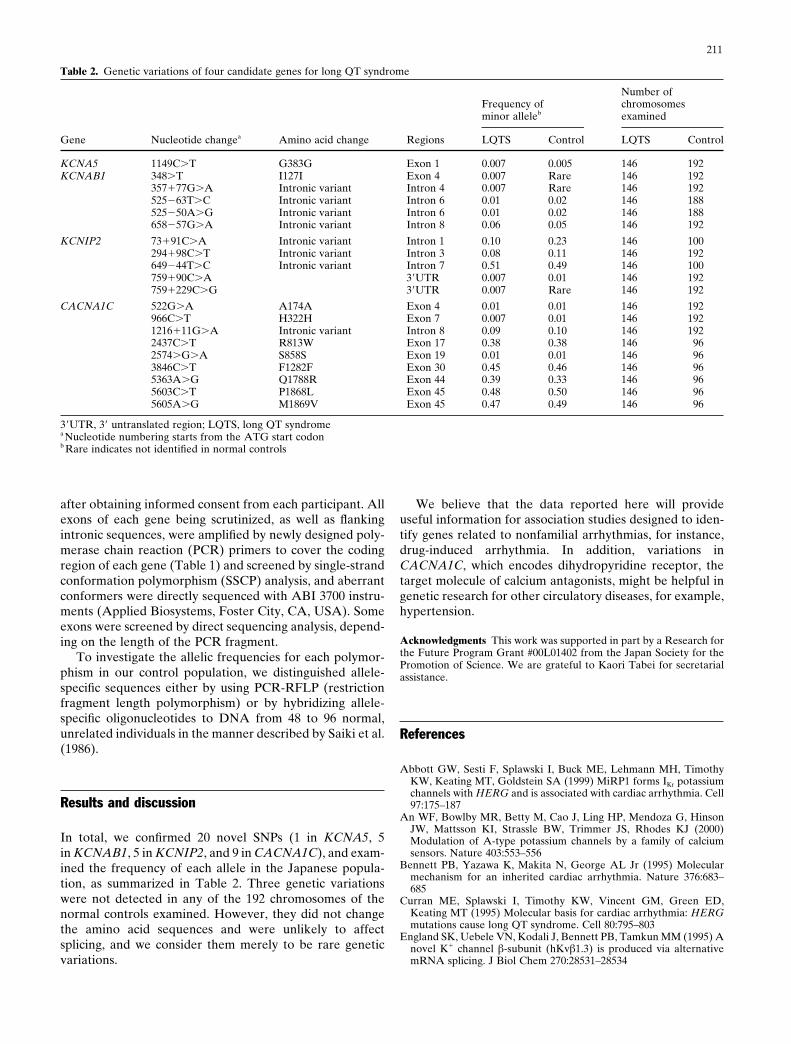
In total, we confirmed 20 novel SNPs (1 in KCNA5, 5in KCNAB1, 5 in KCNIP2, and 9 in CACNA1C), and exam-ined the frequency of each allele in the Japanese popula-tion, as summarized in Table 2. Three genetic variationswere not detected in any of the 192 chromosomes of thenormal controls examined. However, they did not changethe amino acid sequences and were unlikely to affectsplicing, and we consider them merely to be rare geneticvariations.
Table 2. Genetic variations of four candidate genes for long QT syndrome
Number ofFrequency of chromosomesminor alleleb examined
Gene Nucleotide changea Amino acid change Regions LQTS Control LQTS Control
KCNA5 1149C�T G383G Exon 1 0.007 0.005 146 192KCNAB1 348�T I127I Exon 4 0.007 Rare 146 192
357�77G�A Intronic variant Intron 4 0.007 Rare 146 192525�63T�C Intronic variant Intron 6 0.01 0.02 146 188525�50A�G Intronic variant Intron 6 0.01 0.02 146 188658�57G�A Intronic variant Intron 8 0.06 0.05 146 192
KCNIP2 73�91C�A Intronic variant Intron 1 0.10 0.23 146 100294�98C�T Intronic variant Intron 3 0.08 0.11 146 192649�44T�C Intronic variant Intron 7 0.51 0.49 146 100759�90C�A 3�UTR 0.007 0.01 146 192759�229C�G 3�UTR 0.007 Rare 146 192
CACNA1C 522G�A A174A Exon 4 0.01 0.01 146 192966C�T H322H Exon 7 0.007 0.01 146 1921216�11G�A Intronic variant Intron 8 0.09 0.10 146 1922437C�T R813W Exon 17 0.38 0.38 146 962574�G�A S858S Exon 19 0.01 0.01 146 963846C�T F1282F Exon 30 0.45 0.46 146 965363A�G Q1788R Exon 44 0.39 0.33 146 965603C�T P1868L Exon 45 0.48 0.50 146 965605A�G M1869V Exon 45 0.47 0.49 146 96
3�UTR, 3� untranslated region; LQTS, long QT syndromea Nucleotide numbering starts from the ATG start codonb Rare indicates not identified in normal controls
We believe that the data reported here will provideuseful information for association studies designed to iden-tify genes related to nonfamilial arrhythmias, for instance,drug-induced arrhythmia. In addition, variations inCACNA1C, which encodes dihydropyridine receptor, thetarget molecule of calcium antagonists, might be helpful ingenetic research for other circulatory diseases, for example,hypertension.
Acknowledgments This work was supported in part by a Research forthe Future Program Grant #00L01402 from the Japan Society for thePromotion of Science. We are grateful to Kaori Tabei for secretarialassistance.
References
Abbott GW, Sesti F, Splawski I, Buck ME, Lehmann MH, TimothyKW, Keating MT, Goldstein SA (1999) MiRP1 forms IKr potassiumchannels with HERG and is associated with cardiac arrhythmia. Cell97:175–187
An WF, Bowlby MR, Betty M, Cao J, Ling HP, Mendoza G, HinsonJW, Mattsson KI, Strassle BW, Trimmer JS, Rhodes KJ (2000)Modulation of A-type potassium channels by a family of calciumsensors. Nature 403:553–556
Bennett PB, Yazawa K, Makita N, George AL Jr (1995) Molecularmechanism for an inherited cardiac arrhythmia. Nature 376:683–685
Curran ME, Splawski I, Timothy KW, Vincent GM, Green ED,Keating MT (1995) Molecular basis for cardiac arrhythmia: HERGmutations cause long QT syndrome. Cell 80:795–803
England SK, Uebele VN, Kodali J, Bennett PB, Tamkun MM (1995) Anovel K� channel �-subunit (hKv�1.3) is produced via alternativemRNA splicing. J Biol Chem 270:28531–28534

212 N. Matsuda et al.: EGF receptor and osteoblastic differentiation
Itoh T, Tanaka T, Nagai R, Kikuchi K, Ogawa S, Okada S, YamagataS, Yano K, Yazaki Y, Nakamura Y (1998) Genomic organizationand mutational analysis of KVLQT1, a gene responsible for familiallong QT syndrome. Hum Genet 103:290–294
January CT, Riddle JM (1989) Early afterdepolarizations: mechanismof induction and block. A role for L-type Ca2� current. Circ Res64:977–990
Kuo HC, Cheng CF, Clark RB, Lin JJ, Lin JL, Hoshijima M, Nguyen-Tran VT, Gu Y, Ikeda Y, Chu PH, Ross J, Giles WR, Chien KR(2001) A defect in the Kv channel-interacting protein 2 (KChIP2)gene leads to a complete loss of Ito and confers susceptibility toventricular tachycardia. Cell 107:801–813
London B, Jeron A, Zhou J, Buckett P, Han X, Mitchell GF, Koren G(1998) Long QT and ventricular arrhythmias in transgenic mice ex-pressing the N terminus and first transmembrane segment of avoltage-gated potassium channel. Proc Natl Acad Sci U S A 95:2926–2931
Rosati B, Pan Z, Lypen S, Wang HS, Cohen I, Dixon JE, McKinnon D(2001) Regulation of KChIP2 potassium channel � subunit geneexpression underlies the gradient of transient outward current incanine and human ventricle. J Physiol 533.1:119–125
Saiki RK, Bugawan TL, Horn GT, Mullis KB, Erlich HA (1986)Analysis of enzymatically amplified beta-globin and HLA-DQ alphaDNA with allele-specific oligonucleotide probes. Nature 324:163–166
Soldatov NM (1992) Molecular diversity of L-type Ca2� channel tran-scripts in human fibroblasts. Proc Natl Acad Sci U S A 89:4628–4632
Soldatov NM (1994) Genomic structure of human L-type Ca2� channel.Genomics 22:77–87
Splawski I, Tristani-Firouzi M, Lehmann MH, Sanguinetti MC,Keating MT (1997) Mutations in the hminK gene cause long QTsyndrome and suppress IKs function. Nat Genet 17:338–340
Wang Q, Li Z, Shen J, Keating MT (1996a) Genomic organization ofthe human SCN5A gene encoding the cardiac sodium channel.Genomics 34:9–16
Wang Q, Curran ME, Splawski I, Burn TC, Millholland JM, VanRaayTJ, Shen J, Timothy KW, Vincent GM, de Jager T, Schwartz PJ,Towbin JA, Moss AJ, Atkinson DL, Landes GM, Connors TD,Keating MT (1996b) Positional cloning of a novel potassium channelgene: KVLQT1 mutations cause cardiac arrhythmias. Nat Genet12:17–23