Embed Size (px)
Citation preview

Tropical forest response to climate change Dr. Julián Granados Castellanos Calle 55 – B Número 179 Entre 40 y 42 Fraccionamiento Francisco de Montejo Mérida, Yucatán México [email protected]
November 2006

2
Human disruption on tropical forest ecophysiology
Tropical forest can be found in each of the three land areas, which occur
within the tropical zone and it displays three major tropical regions: The
Neotropical, the Paleotropical, which occupies two African regions, and the South
East Asian tropical forest. Of those regions, Neotropical forest is the most
extensive and diverse in life and portrays a magnificent world biodiversity reservoir,
distributed from Central America to Amazonia. Various vegetation types (i.e.
tropical rain forest, semi evergreen forest, savanna, wetlands, etc.), in which
amalgams of growth forms (trees, lianas, epiphytes, stranglers, herbs, etc.) build
plant communities, which account for the worlds highest arboreal species diversity
number, and even excluding trees, shows the highest world floristic diversity.
Earth biodiversity depends on continuous plant growth resource availability
and solar energy reaching tropical regions has through an evolutionary process
produced a plant productivity-biodiversity binomial, which symbolizes the crown of
atmosphere-biosphere coupling. Atmosphere, through the climate system,
influences Biosphere, and Biosphere reaction due to climate change can be with
water and nutrient cycling changes observed. For exa mple, climatic periodicity
disruptions have substantial effects on precipitation patterns on which ecosystem
process depend. World biomes are sensitive to climate periodicity and Equatorial
tropical forests distribution, formation and species composition has changed with
changing global climates during the last 30,000 years. Since the industrial
revolution a global climate change has been generated as a consequence of
atmospheric greenhouse gases enrichment. Furthermore, pristine tropical forest
has been extensively cleared all over the world (ca. 20 - 30 % of Earth surface),
3
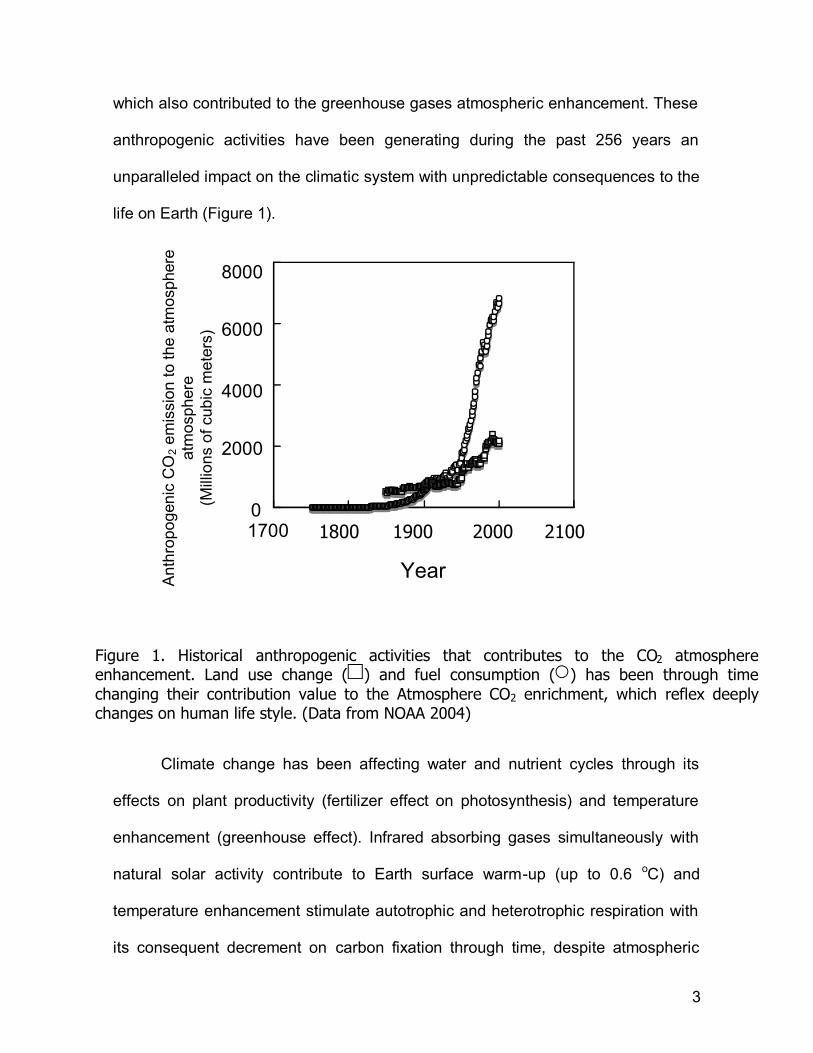
which also contributed to the greenhouse gases atmospheric enhancement. These
anthropogenic activities have been generating during the past 256 years an
unparalleled impact on the climatic system with unpredictable consequences to the
life on Earth (Figure 1).
Climate change has been affecting water and nutrient cycles through its
effects on plant productivity (fertilizer effect on photosynthesis) and temperature
enhancement (greenhouse effect). Infrared absorbing gases simultaneously with
natural solar activity contribute to Earth surface warm-up (up to 0.6 oC) and
temperature enhancement stimulate autotrophic and heterotrophic respiration with
its consequent decrement on carbon fixation through time, despite atmospheric
Figure 1. Historical anthropogenic activities that contributes to the CO2 atmosphere enhancement. Land use change ( ) and fuel consumption ( ) has been through time changing their contribution value to the Atmosphere CO2 enrichment, which reflex deeply changes on human life style. (Data from NOAA 2004)
1700 1800 1900 2000 2100
Year Ant
hrop
ogen
ic C
O2 e
mis
sion
to th
e at
mos
pher
e
atm
osph
ere
(M
illio
ns o
f cub
ic m
eter
s)
0
2000
4000
6000
8000

4
CO2-enrichment fertilization potential. Otherwise, intact tropical forest stores a very
high amount of carbon, accumulated in wood for more than a Century. However,
intact pristine forests are not common anymore and this is a consequence of
intensive change in tropical forest land use, which similarly to CO2-enriched
atmosphere, has the potential to enhance tropical forest dynamics, affecting
tropical forest carbon residence time. A reduction in carbon fores t residence time
involves the following aspects: Tree life span depends on tree species and forest
type, and tree longevity determines carbon residence time, which has major
implications on tropical carbon pool size. About 50% of biomass carbon in tropical
forests is bound in canopy trees that represent a minor proportion of the total
number of tropical trees. However, tropical forest is not like a passive sink of
carbon, and successional processes (gap dynamics in pristine forests and forest
succession in secondary vegetation) may be biased by a CO2-enriched
atmosphere to changes in arboreal species forest dominance due to habitat
productivity determination of plant community structure.
In pristine forest, gap dynamics play a major role in natural regenerat ion
processes, and under gap regeneration conditions CO 2-enhanced liana growth and
competitiveness (i.e. Granados and Korner 2002, Global Change Biology 8:1109-
1117); Granados 2002, dissertation Basel University) have negative influence on
tree growth and CO2-resposivenes which could reduce tree life span and
discrimination against of slow growing, long lived tree species, with obvious
consequences for forest carbon stocking. The relative atmospheric CO 2-
enrichment effect should become stronger as one approaches light compensation
point (where light influence on plant assimilation equals plant respiration) under

5
severe light limitation in pristine tropical forest understory CO2-effect on plant
growth declines with plant age and in forest understory where light availability is
limiting, seedlings will be more CO2-sensitive. Liana CO2-response allows liana
effects on forest dynamics enhancing the possibility of forest succession stall in an
early successional stage with a negative impact on forest structure. Changes in
forest structure toward shorter tree life span could lead to a reduction of carbon
residence time in pristine forest. Whereas, a most common form of tropical forest
stage is secondary vegetation, which shows a very different ecology compared
with pristine forests, due to higher species interactions than in pristine forest. For
example, density and biodiversity of lianas and other fast-growing species
decrease as forest stand age increases. However, atmospheric CO 2-enrichment
effects on the secondary forest regeneration process remain a conundrum.
Regarding the biosphere carbon sink, a main concern is biodiversity, and it
has been disregarded that tropical forests are not homogenous sinks of carbon.
The structural components (species diversity) play a major role in the forests
capacity to store carbon. However, species structure and dominance widely varies
within forest types and with forest life history. Thus, a predictive understanding
needs to account for biodiversity effects on tropical forest carbon storage capacity
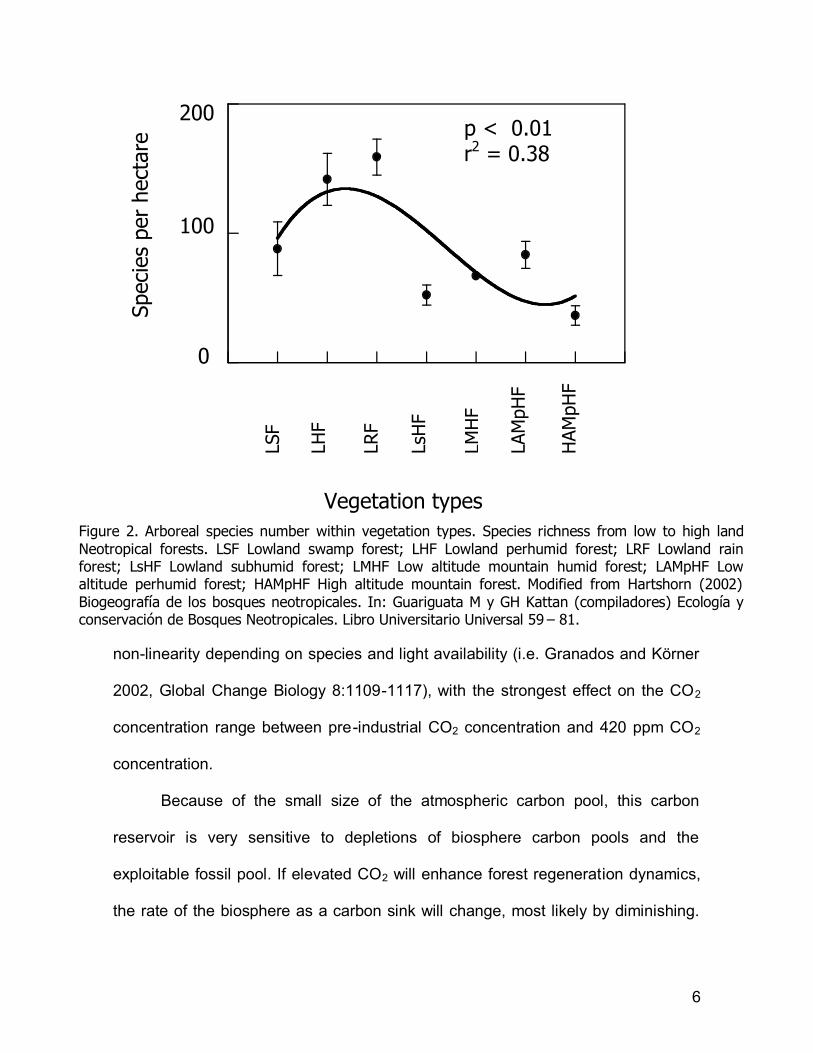
particularly because productivity and species diversity scale dependently (Figure
2). Another important aspect is the temporal character of the elevated CO 2-effect
on the biota. Growth responses to elevated CO2 are non-linear with the degree of
6
Vegetation types
non-linearity depending on species and light availability (i.e. Granados and Körner
2002, Global Change Biology 8:1109-1117), with the strongest effect on the CO2
concentration range between pre-industrial CO2 concentration and 420 ppm CO2
concentration.
Because of the small size of the atmospheric carbon pool, this carbon
reservoir is very sensitive to depletions of biosphere carbon pools and the
exploitable fossil pool. If elevated CO2 will enhance forest regeneration dynamics,
the rate of the biosphere as a carbon sink will change, most likely by diminishing.
p < 0.01 r2 = 0.38
LSF
LHF
LRF
LsH
F
LMH
F
LAM
pHF
HAM
pHF
0
100
200
Figure 2. Arboreal species number within vegetation types. Species richness from low to high land Neotropical forests. LSF Lowland swamp forest; LHF Lowland perhumid forest; LRF Lowland rain forest; LsHF Lowland subhumid forest; LMHF Low altitude mountain humid forest; LAMpHF Low altitude perhumid forest; HAMpHF High altitude mountain forest. Modified from Hartshorn (2002) Biogeografía de los bosques neotropicales. In: Guariguata M y GH Kattan (compiladores) Ecología y conservación de Bosques Neotropicales. Libro Universitario Universal 59 – 81.
Spec
ies
per
hect
are

7
We need forest CO2-enrichment experiments in the tropics to obtain a wide and
predictable understanding of tropical forest response to climate change.
The Neotropics and Global warming
Global warming is an atmospheric greenhouse gas enrichment
consequence. Main greenhouse gases are: Carbon dioxide (CO 2), Methane (CH4),
Nitrogen oxide (NOx), the so called halocarbon compounds (Cfs), water vapor and
other biogenic trace gases known as Volatile Organic Components (VOC). Those
atmospheric components are infrared absorbing gases and since the Industrial
Revolution have increasingly been warming up the Earth surface (up to 0.6 oC).
Greenhouse gases radiative force on climate depends on atmospheric gas
enhancement (source variable) and its atmospheric residence time (atmosphere
molecule stability). According to IPCC (Interguvernamental Panel of Climate
Change), atmospheric life time is defined as: "the burden" (Tg = 1 012 g.) divided by
the mean global sink (Tg/yr) for a gas in steady state (i.e. with unchanging burden)
which denote, if atmospheric burden of gas X is 100 Tg, and the mean global sink
is currently 10 Tg/yr, the life time equals 10 years. However, atmosphe ric lifetime is
difficult to define because source variability depends on reservoir exchange with
reservoirs having a wide range of turnover rates. For example, CH 4 is removed
from the atmosphere by a single process, oxidation by the hydroxyl radical (OH).
This is the reason why it is relatively easy to define it’s atmospheric residence time.
However, atmospheric CH4 concentration increase is to reduce OH concentration,
which in turn, reduces destruction of the additional CH4, lengthening its
atmospheric lifetime. Moreover, industrial atmospheric CO2-enrichment causes

8
biogenic CH4 production through its fertilizer effect on photosynthesis due to
carbon belowground distribution induction (i.e. root exudates enhancement), which
could result in CO2 and CH4 emissions enhancement depending on soil anoxia
conditions. Such mechanisms influences atmospheric greenhouse residence time
and have been disregarded to explain radiative force of atmospheric greenhouse
gases. Nevertheless, there are opposite feedback mechanisms to what has been
explained. For example, an increase of N2O can induce chemical reactions leading
to an increase in ultraviolet radiation available to photolyze the N2O, thereby
shortening its atmospheric lifetime. Otherwise, neglected emissions of volatile
organic compounds (VOCs) from vegetation and soil can lead to substantial
underestimate of carbon release from tropical rain forests to the atmosphere. VOC
fluxes can be important for the global carbon cycle and for the troposphere
chemistry due to its importance in regulating air quality and to explain atmospheric
greenhouse gases lifetimes. Neotropical rainforest influence on lifetime of
atmospheric greenhouse gases has been disregard as a climate change motor,
and this process may explain a major atmosphere-biosphere link. Current carbon
cycle models do not include exchange fluxes of different anthropogenic release
and past records of carbon-isotope ratios (13C) show strong shifts of carbon
reservoirs associated with climate warming. Any study of feedback mechanisms
between global warming and carbon release has to account for biogenic source of
atmospheric enhanced greenhouse gases to reach a predictive understanding of
tropical forests role in climate change models.
Terrestrial ecosystem productivity integrates living organisms and
atmospheric processes. Climate and land use feedbacks on climate mainly through

9
CO2 and H2O cycles and Neotropical forest biodiversity reservoir has been
disregarded on global climate change framework. Particularly, when plant
community theory often invokes competition to explain why species diversity
declines as productivity increases. For example plant-plant and plant-animal
interactions under rising atmospheric temperature has not been seriously explored.
Rising atmospheric temperature due to greenhouse gases enhancement has a
tremendous potential to affect biosphere physiology, because temperature greatly
affects carbon exchange rates and plant growth. Temperature decreases
photosynthesis when leaf temperature rises. Rubisco activase inhibition due to
temperature enhancement strongly interacts with light and water availability due to
changes in CO2 compensation point. The steady state at which net assimilation
rate equals zero (CO2 compensation point) remarkably depends on temperature
and O2 atmospheric concentration. Under enhanced atmospheric CO 2
concentration, water availability changes, with an increasing global temperature
may influence plant respiration (physiologica process associate with plant
maintenance). Atmospheric CO2-enrichment effect on plant respiration and its
temperature interaction have not been yet clarified. The only satisfactory way to
determine plant respiration and atmospheric CO2 interaction is by experiment. This
neglected topic has not been systematically investigated and its role to explain
atmospheric changes in CO2 concentration with its consequent effect on
atmospheric chemistry has also been neglected. For example, elevated CO 2 may
enhance growth, but higher temperatures may diminish it through increased
respiration. How does this tradeoff affect Neotropical forest productivity when we
account for plant diversity productivity links? A lack of information on such

10
temperature per atmospheric CO2-enrichment interaction needs to be filled to
construct a predictive understanding on tropical plant responses under climate
change.
Fertilization effect on photosynthesis by atmospheric elevated CO 2 concentration
Productivity is the rate of resource conversion to biomass per unit area per
unit time and species richness is affected by productivity. Worldwide, Neotropical
forest has no biodiversity parallel and Global climate change due to enhanced
atmospheric CO2 concentration may exert biodiversity effects due to species-
specific differential growth responses which may engage in unpredictable
consequences on global carbon cycle and consequently on climate change. Global
carbon balance responses to global change are highly dependent on factors
limiting primary productivity and net primary production (NPP) linearly increases as
temperature increase, but declines with high precipitation in tropical forest. At an
intermediate spatial scale (within biomes but across communities) the productivity -
diversity relationship is often uni-modal, with diversity peaking at intermediate
productivity. Natural mechanisms research, linking primary production with
biodiversity is a most predictable understanding value to the climate change
problem. For example, factors that change ecosystem composition, such as novel
organism invasion, nitrogen deposition, disturbance frequency, forest
fragmentation, predator decimation, alternative management practices, and global
change itself, are likely to strongly affect ecosystem processes, which represent
changes in biodiversity reservoir.

11
Plants through their photosynthesis depending on CO2, have been growing
during at least 420,000 years under atmospheric CO2 concentrations below 280
ppm. Therefore, atmospheric CO2 anthropogenic enrichment which has been
occurring during 256 years, has been much faster than evolutionary processes in
higher plants, slow growing species in particular. A higher atmospheric CO 2
concentration stimulates plant growth with distinct species growth response due to
inherent lifespan cycles. Within any habitat, an individual plant is more likely to
interact with neighboring plants than within more distant ones. A fast increment on
atmospheric CO2 concentration could be a major determinant of enhanced forest
dynamics due to differential plant growth response within co-occurring competitors.
The existing relationship between primary productivity and species richness
(one aspect of biodiversity concept) can be expressed as follows: productivity
influences species richness. A general agreement of most authors in the literature
express that productivity affects biodiversity. However, no general mechanism has
been described to clarify productivity-biodiversity relationship. Despite that
knowledge gap some major mechanisms, which are susceptible to be by climate
change, has been described. I address three of these mechanisms, pointing out
their capability to affect tropical forest carbon storage capacity due to their
sensitivity to climate change: (1) Neotropical forest vegetation types; (2) Pristine
forest natural regeneration (gap dynamics) and (3) Secondary forest regeneration.
Neotropical forest vegetation types.
Vegetation type is mainly determined by water availability, edaphic
conditions (topography and soil fertility) and altitude. A feature of rainwater

12
availability on Neotropical forest vegetation composition-structure depends on
precipitation in the Yucatan Peninsula. From North to South a precipitation
gradient, annually and geographically ranging from 400 to 3000 mm determines
structural and compositional differences among vegetation types. Recently White
and Hoods (2004 Journal of vegetation science 15:151 – 160) Yucatan Peninsula
study demonstrated a floristic variation consequent to an overall West to East
environmental rainfall gradient similar to what the great Mexican Botanist Miranda
previously described in 1959.
Diverse authors have classified Neotropical forest depending on rainfall
precipitation, temperature, and altitude, which have resulted in eight vegetation
type groups, structurally, floristically and physiognomically different at large.
Arboreal species, which are main forest carbon reservoirs, widely varies in floristic
composition within vegetation types. A wide spectrum of forest responses to
climate change can be expected under such arboreal species variety. Far as I
know, no work had taken such differences in account, and it is a critical aspect to
construct a predictive understanding of climate change and Neotropical forest
growth response. I propose to the scientific community, the construction o f an
understanding framework in which precipitation gradients interacting with soil
fertility factorizes CO2-enrichment arboreal species growth responses. This,
because vegetation types productivity are mainly on those growth factors
dependent.
Climate change in the Pleistocene strongly affected pluvial precipitation
patterns; similar to what has been presently occurring on climatic periodicity
(substantial effects on precipitation patterns) due to the Industrial Revolution

13
influence on atmospheric greenhouse gases concentration. Neotropical forest flora
as we presently know was during Miocene, some 23 to 6 million years ago, more
variegate than contemporary forests. Additionally, the time period during which
Pleistocene climate change occurred was on the order of thousands of years and
generated drastic changes on temperature and precipitation patterns producing a
reduction in tropical forest Earth surface cover. Present Industrial Revolution
greenhouse concentration enhancement was happening during the past 256 years,
and Earth surface temperature warm-up, which has unpredictable consequences
on dynamic, composition and structure of tropical plant communities, is a CO 2
fertilization - greenhouse effect conundrum on Neotropical forest. In figure 3, I
present a hierarchical structure of factors involved in the carbon pool of tropical
forest dynamics. Vegetation type differences and human land use are at most,
determinant of final vegetation recover. A predictable understanding of past,
present and future atmospheric CO2 concentration has to account for such
hierarchical interactions due to atmospheric CO2-enrichment influence on forest
dynamics (i.e. Granados 2002 dissertation Basel University).

14
Tropical Forest (1) Geographical situation
(2) Community vegetation (composition and structure)
Tropical rain forest Tropical wet forest Savanna
(3) Human disruption (a) Natural disruption (Gap dynamics) (b)
Gap
Atmospheric CO2 concentration (planetary scale)
Physical disruption
- Deforestation explains ~ 45 % of increased atmospheric CO2 concentration since 1850 (Malhi et al. 2002)
Ecophysiological disruption
- Elevated CO2 enhances liana growth and competition
- Carbon emission from fossil fuels has surpassed those from defor-estation.
- Forest dynamics enhanced which decreases the probability of carbon sequestration
Figure 3. A hierarchical structure of factors involved in the carbon pool of tropical forest dynamics. (1) Planetary scale; (2) Landscape scale; (3) Regional scale: (a) Human induced disruption, (b) Natural processes. From Granados (2002) Ph.D. Dissertation Basel University

15
Natural regeneration in pristine forest (gap dynamics)
Competition itself induces a set of non-equilibrium species oscillations that
allow coexistence of more species than would normally be expected on the basis
of the number of limiting resources. Whether or not this natural mechanism driv es
Neotropical biodiversity, actual industrial atmospheric CO2 concentration
enrichment powers species bias in a high biodiverse forest Neotropical like. Which
species will profit more than others from the stimulant CO2-fertilizer effect, and
when, is not clear. Yoda’s Self-thinning rule establishes that as the number of plant
competitors decreases, the biomass of competing plants increases. Therefore,
under photosynthetic fertilization effects due to atmospheric CO 2-enrichment, what
kind of competitive mechanisms are able to be change and under what kind of
vegetal communities processes? A wide spectrum of plant-plant and plant-animal
interaction can be listed. I will focus on the following: Natural regeneration in
pristine forest and Secondary forest regeneration.
In 1994, Phillips and Gentry (Science 263:954-958) suggested that
according to large-scale surveys, tropical forest tree turnover seems to have
increased worldwide. All experts do not share this view. However, it was not
disputed that there is a possibility that accelerated tree turnover could be mediated
by increased vigor of climbing plants or other fast growing arboreal species, which
in itself could result from atmospheric CO2-enrichment. The juvenile growth stage
of trees is a critical step in forest regeneration and species-specific interactions can
strongly influence recruitment success.
Tree turnover is determined by a series of factors from which stem size,
plant age, tree size, mechanical and physical damage stand density and light

16
availability are the most important. The forest understory is a very competitive
environment in which light availability varies widely in time and intensity. It is well
known that in general, elevated CO2 improves growth resource use efficiency and
thus may increase plant competitive ability. For example, elevated CO2 reduces
light compensation point of photosynthesis, having a multiplicative effect on plant
biomass production, at severe light limitation. Additionally, the stimulative effect of
CO2 declines with time and thus plant will also decline its CO2 growth response
with age. Consequently, natural forest regeneration at initial conditions due to gap
dynamics are susceptible to bias under atmospheric elevated CO 2 in favor of
plants with a high CO2-growth responsiveness which not necessarily will be fast
grow species because differences in light and CO2 compensation point within plant
life forms will determine differences on CO2-growth responses and
competitiveness.
Because of their dependence on physical support, lianas have the capacity
to influence natural tropical forest regeneration and they represent an important
selective force to tree survival and thus diversity. Initial conditions can be
influenced by liana presence, slowing forest regeneration in an early
developmental stage. At a later stage lianas can enhance tree mortality thus,
having a significant effect on the carbon forest storage capacity. Recent forest
inventories in the Neotropics show that liana dominance has indeed increased over
the last 30 years, which supports Phillips and Gentry original hypothesis and that
has been reinforced by Granados and Körner (2002 Global Change Biology
8:1109-1117) and Granados (2002 dissertation Basel University) data.

17
Intact pristine forest stores very high amounts of carbon, accumulated in
wood for more than a century. However, pristine forests are not common anymore,
and since the Industrial Revolution ca. 20 - 30 % of the original forest area on
Earth has been lost. Additionally, carbon residence time in the forest depends on
tree life span and this depends on arboreal species and forest type. Few studies
consider tree longevity as a main focus. Studies show an age range between 8 to
1400 years, almost depending on species life strategy (pioneer and non-pioneer
arboreal species). Estimates of tree life span for pioneer species are 8 - 40 years
and for non-pioneer tree species between 200 - 1400 years. Remarkably,
atmospheric CO2-enrichment has been happening almost during 255 years and
worldwide forestation criteria have not addressed this aspect of climate change.
Tree life span determines carbon residence time, which has major implications on
forest carbon pool size, because about 50% of biomass carbon is bound in canopy
trees that normally represent a minor proportion of the total number of tropical
trees. As has been demonstrated by Granados and Körner (2002 Global Change
Biology 8:1109-1117) and Granados (2002 dissertation Basel University), under
gap regeneration conditions CO2-enhanced liana growth and competitiveness have
a negative influence on tree growth and CO2-responsivenness, which over
generations could result in reduced tree life span, which in turn will produce a
decrement on carbon forest storage capacity. Figure 4, synthesizes a general
mechanism for CO2 effects on gap dynamics and carbon forest storage capacity. In
a fully structured forest
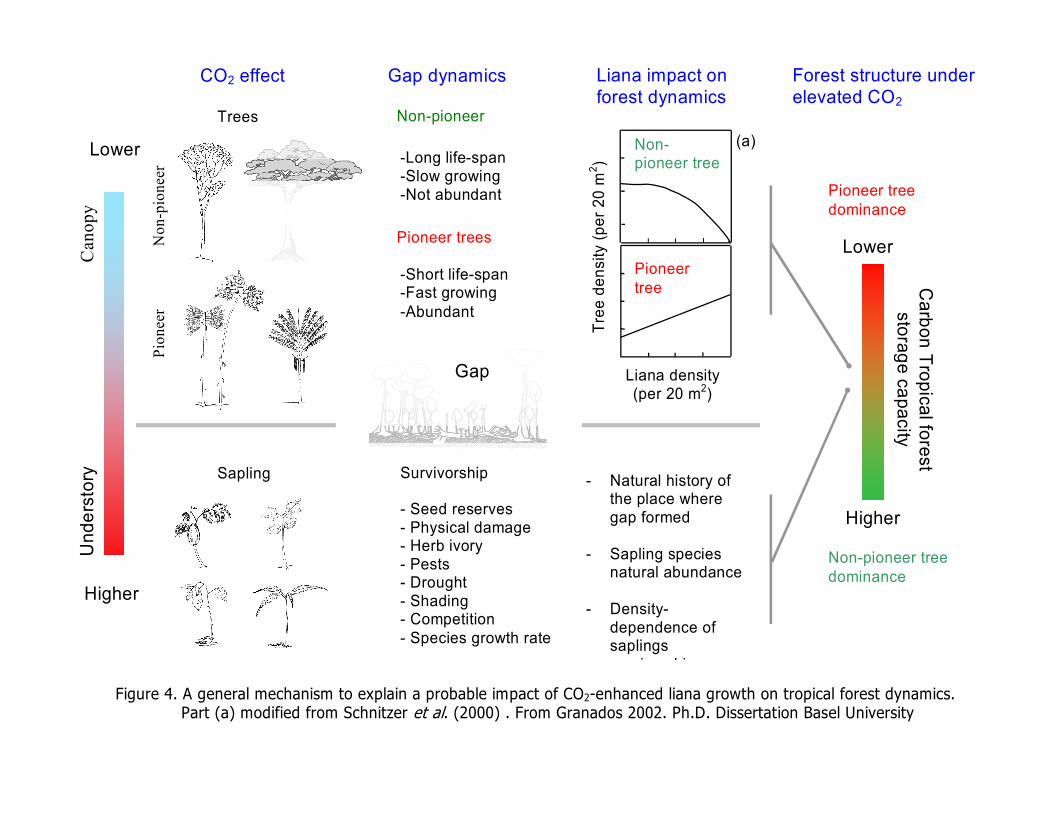
Figure 4. A general mechanism to explain a probable impact of CO2-enhanced liana growth on tropical forest dynamics. Part (a) modified from Schnitzer et al. (2000) . From Granados 2002. Ph.D. Dissertation Basel University
CO2 effect C
ano
py
U
nder
stor
y Gap dynamics Liana impact on
forest dynamics
- Natural history of the place where gap formed
- Sapling species
natural abundance
- Density-dependence of saplings survivorship
Liana density (per 20 m2)
Tree
den
sity
(per
20
m2 )
Forest structure under elevated CO2
Pioneer tree dominance
Carbon Tropical forest
storage capacity
Higher
Lower
Lower
Higher
Saplings
Trees
Pio
nee
r N
on
-pio
nee
r
Pioneer tree
Gap
-Long life-span -Slow growing -Not abundant
-Short life-span -Fast growing -Abundant
Non-pioneer trees
Pioneer trees
Survivorship factors - Seed reserves - Physical damage - Herb ivory - Pests - Drought - Shading - Competition - Species growth rate
Non-pioneer tree dominance
(a) Non-pioneer tree

when a tree falls, a canopy gap opens and a successional event starts being
a major determinant of the new canopy tree species composition. Liana presence
in pristine forest can stall forest succession in early successional stages, favoring
pioneer arboreal species during forest structure build-up, while changes in forest
structure toward shorter tree life span could lead to a reduction of carbon residence
time in forest.
Secondary forest regeneration
Compared with pristine forest, ecology of secondary vegetation is very
different, and this kind of vegetation community is a most common form of the
actual tropical vegetation stage. In general, secondary vegetation forest ecology
differs from pristine forest ecology mainly because interacting species are much
more diverse. In a world with elevated CO2 concentration the fertilization effect on
photosynthesis has the potential to enhance competition among species. A wide
variety of mechanisms can lead to diverse floristical successional processes and
rates of forest recovery. Regarding liana effect, density and biodiversity of lianas
decrease as forest stand ages increase and this seems to be related with light
availability. Because species widely differ in light requirements and light gradients
explain successional dynamics, tropical tree species coexistence will be more
difficult to predict under secondary forest stages. Moreover, despite the fact that
liana abundance is highest in young stands, no liana species are restricted to
younger forest. Thus, liana species presence is not restricting ageing of stands and
slow growing tree species are normally excluded from early successional stages.
Furthermore, pristine forest conversion to secondary vegetation has been shown to

20
result in losses of soil organic matter. Soil organic matter is a main determinant of
soil fertility and a key uncertainty extends to whether the fe rtilization effect is limited
by the availability of other nutrients such as nitrogen and phosphorus. It has been
shown that forest ecosystem responses to CO2-enriched atmosphere are highly
dependent on soil fertility. Depletions on soil organic matter wil l deplete soil fertility
and lianas are equally represented in tropical forest with poor and fertile soils, and
the biggest trees are usually found in tropical forest with fertile soils. Liana impacts
on secondary vegetation are very strong and it has been suggested that by the
protection of such disturbed areas, tropical forest regrowth could be used as a
carbon forest sink. I propose a reassessment of the usefulness of such Kyoto
protocol target.
Forest regeneration confronts plant-plant interactions (in both gap dynamics
and secondary successional processes) that affect the establishment and juvenile
life phase of trees in different forms. Arboreal species juvenile life phase at most
concern tree turnover rates, which took place in the forest understory a nd depend
on local conditions and vegetation type. However, other aspects like latitude,
seasonality, topography heterogeneity and dry periods are also important, but
planetary-scaled processes determine them. For example, long turnover rates are
a function of primary productivity and primary productivity is also a function of
geographical situation due mainly to rainfall variation. Thus, forests that are
intrinsically of low turnover rates are also different in composition and structure and
forest structure-composition is related to biomass fixed and carbon storage
capacity (Figure 3, 4).

21
Human civilization confronts major consequences of its development. Fast
atmospheric CO2-enrichment represents a change in the availability of a growth
factor, which can destabilize associations of plant communities due to its abrupt
character. Current atmospheric CO2-enrichment has been occurring in ecological
time scales, and during past epochs atmospheric CO2 concentration varied
between 200 to 400 ppm, and those changes did not appear to be associated with
collapses in plant communities or species extinction events. On the contrary, those
changes were associated with plant biodiversity increments. However, those CO 2
concentration changes took place over millions of years, which represent
evolutionary time scales, a situation quite different from the current one where CO 2
concentrations change far more rapidly than evolutionary processes in most
species. The link between forest regeneration mechanisms and biodiversity is t he
key process to understand the effect of CO2-enhanced liana growth and
competition on tropical forest dynamics and thus in the global carbon cycle.
Species turnover rates appear to be related with habitat variability, and low
turnover rates of species over great distances in the tropical regions suggest that
population densities of some species are limited by unidentified process that may
be associated with dispersal process. If biodiversity is maintained by a tradeoff
between recruitment ability and competitive ability, a CO2-enhanced arboreal fast
growing species and lianas will have negative effects on tree biodiversity and may
have a major impact on tropical forest composition. Predictions of tropical carbon
fluxes will need to account for the changing composition and dynamics of
supposedly pristine forest. A recently report for undisturbed Amazonian forest
found a non-random change in floristic composition in which many faster-growing

22
genera are increasing in the basal area (i.e. Laurance et al. 2004 N ature 428:171-
175).
The temporal characteristics of atmospheric elevated CO2-fertilization effect
It is completely unknown if and how elevated CO2 will influence the growth
and development of adult tropical trees. Results of the limited number of
experimental studies outside the tropics suggest a reduced sensitivity, as trees get
older. The only test with tall tropical trees was a leaf CO2-enrichment experiment
(Würth et al. 1998 Functional Ecology 12:886-895), which revealed an immediate
accumulation of non-structural carbohydrates (NSC), despite the fact that leaves
where attached to large trees, which could be considered representing a nearly
infinite carbon sink for such leaves. Another aspect to be considered in the adult
trees as a carbon sink is the capacity that each living organism has to change in
size. The growth curve of any living organism presents an inflexion point on which
its growth capacity depletes. Thus, there is a spatial -temporal limit for the trees to
accumulate carbon in its biomass, and for the vegetation community as well due to
density dependence of plant size. Otherwise, the first evidence that elevated CO 2
induces a response which declines with time (where) were presented by Bazzaz et
al. (1995 Proceedings of the nacional academy of sciences of the U.S.A.
98(18):8161-8165). Nevertheless, that work used seedlings; the results obtained in
those experiments are in line with Hattenschwiller et al. (1997 Global Change
Biology 3:436-471) findings, which describe that elevated CO2 effect declines with
tree age. In which period of plant life-span process atmospheric CO2-enrichment
will be critical for final forest regeneration? And how does that critical influence

23
determine plant life form competitiveness? This is a major Science unknown, and a
high research priority due to the worldwide environmental crisis.
Forest growth responses to elevated CO2 are non-linear with the degree of
non-linearity depending on species and light availability. Strongest CO2 effects
occur in the CO2 concentration ranges between pre-industrial CO2 concentration
and 420-ppm CO2 concentration, which is only 60 ppm less than current CO2
concentration. Beyond 420 ppm the growth response to CO 2 declines until 560
ppm CO2 concentration. Therefore, CO2 effects are likely to be underway right
now. A predictive understanding of Neotropical forest responses to climate change
has to account for arboreal species, linear and non-linear growth responses to
atmospheric CO2 enrichment.
The exponential increase of Industrial Revolution atmospheric CO2
concentration has no parallel in the planet’s history, and its effects have not been
clarified because existing work contains few considerations of Biosphere
compensatory features. The experimental approach proposed by Granados and
Körner (2002 Global Change Biology 8:) allows the construction of a global change
framework with high predictive value. For example, Figure 5 (a) shows the
Granados and Körner (2002) experiment. The two of three species mixtures show
a maximum response of plant biomass gain at 560 ppm CO2 concentration, but
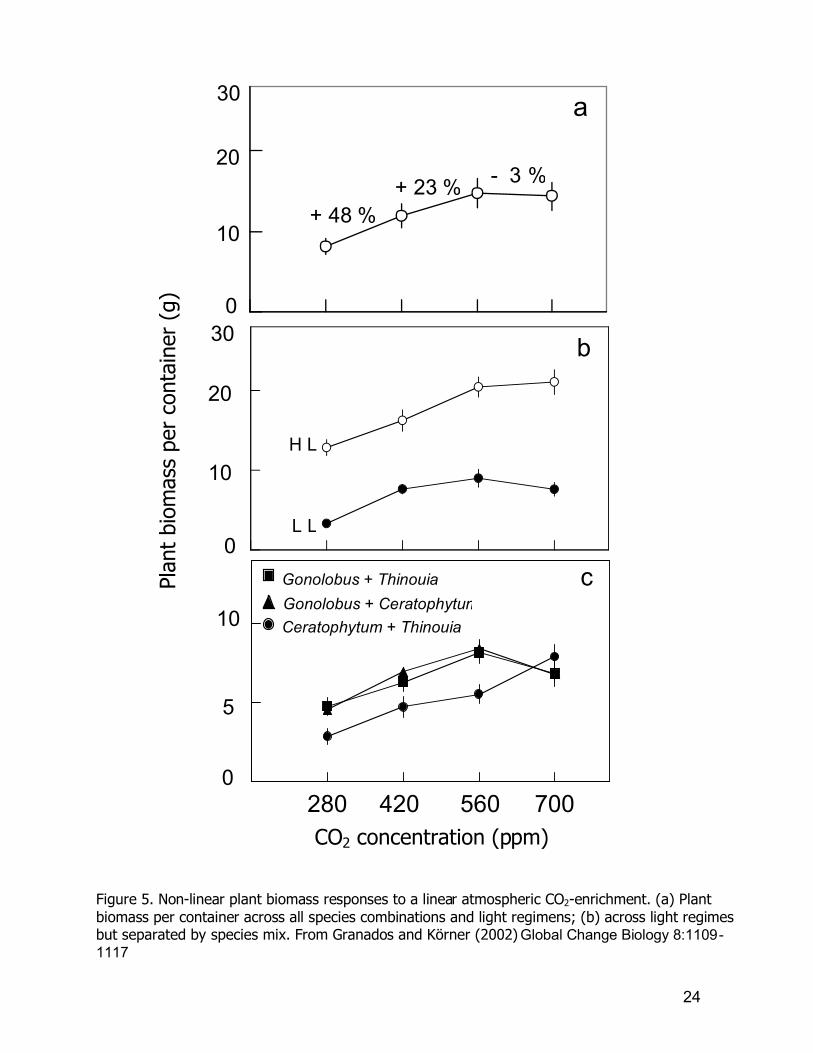
24
H L
L L
b
Gonolobus + ThinouiaGonolobus + CeratophytumCeratophytum + Thinouia
cPlan
t bi
omas
s pe
r co
ntai
ner
(g)
280 0
10
5
30
20
10
0
420 560 700
30
20
10
0
CO2 concentration (ppm)
Figure 5. Non-linear plant biomass responses to a linear atmospheric CO2-enrichment. (a) Plant biomass per container across all species combinations and light regimens; (b) across light regimes but separated by species mix. From Granados and Körner (2002) Global Change Biology 8:1109-1117
a
+ 48 %+ 23 % - 3 %

25
then leveled-off (Figure 5 c). However, graphical linear apperception of third plant
growth mixture (Ceratophytum and Thinouia, the middle and small sized grown
species) is also non-linearly responding, because at 700 ppm CO2 concentration,
no significant differences were found within species mixtures. At the same time this
graphical and statistical particularity shows that although species mixtures fill -up on
growth containers no "pot-bounding" effects were present. I consider that the use
of the Granados and Körner (2002) experimental linearity test can be used in
Neotropical forest response research, due to its capacity to link plant CO 2-growth
responses to atmospheric exponential CO2 enhancement. A predictable
understanding of biosphere climate change effect can be reached using that
experimental-statistical method.