Embed Size (px)
Citation preview

TRE DISSOCIATION CONSTANTS OF CERTAIN AMINO ACIDS.*
BY PAUL L. KIRK AND CARL L. A. SCHMIDT.
(From the Division of Biochemistry, University of California .%ledical School, Berkeley.)
(Received for publication, October 23, 1928.)
Various attempts (14) have been made to correlate the acid- and base-combining capacity of proteins with the content of certain of the amino acids. As better analytical data have become available, this correlation has become closer, but it is made some- what uncertain by lack of accurate knowledge of the ability of certain amino acids to combine with acid and with base. Ulti- mately, this correlation must also take into consideration the stereochemical structure of the protein molecule. It has been previously pointed out (5) that values for the dissociation constants of oxyproline, &hydroxyglutamic acid, and serine are still lacking. To this list we can add isoleucine and norleucine. To make the list complete there should also be added several other recently discovered amino acids. Our knowledge of these is, at the present time, too fragmentary to warrant attempting work of the nature herein described. Moreover, the dissociation constants for glu- tamic acid were determined by somewhat inadequate methods (6). Harris (7) has published the dissociation constants for valine, but without titration curves, and with an omission in the tables, making it desirable to repeat the determination of the constants for this amino acid.’
*Aided by grants from the Cyrus M. Warren Fund of the American Academy of Arts and Sciences, the Herzstein Fund, and the Research Board of the University of California.
We are indebted to Dr. Kapfhammer for the supply of oxyproline and to the Connecticut Agricultural Experiment Station for the sample of valine.
1 During the preparation of this paper, Simms (8) has published the constants for valine and glutamic acid. We are, however, unable to find the detailed data in the literature.
237
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from

238 Amino Acid Dissociation Constants
We accordingly have attempted to fill these gaps in our knowl- edge of the acid and base strength of the amino acids. It might be pointed out that such knowledge is useful, not only in eluci- dating our concepts of the behavior of the protein molecule, but also in physicochemical studies in which the properties of the amino acid are of direct concern, as for example, the work of Foster and Schmidt (9).
EXPERIMENTAL.
The glutamic acid used in this investigation was prepared from “Ajinomoto” by the method of Schmidt and Foster (10). Racemic valine, racemic norleucine, and d-isoleucine were East- man Kodak products. All were recrystallized before use. A second sample of carefully purified valine, which had been isolated by Dr. T. B. Osborne, was kindly supplied to us by the Con- necticut Agricultural Experimental Station. A second sample of isoleucine was obtained from the Special Chemicals Company. It was recrystallized before use. Oxyproline was supplied to us by Dr. J. Kapfhammer. It was prepared according to the method described by Kapfhammer and Eck (11). The analysis showed it to be a very pure product. Serine was synthesized by the method of Leuchs and Geiger (12) and was recrystallized a number of times. The @hydroxyglutamic acid was prepared from casein by a combination of the transport method of Foster and Schmidt (9) and the method described by Dakin (13). It was further purified by the method suggested by Dakin. The preparation contained a small amount of ash and a correction for this was applied to the titration curve.
All samples were dried over phosphorus pentoxide before weigh- ing. Amino nitrogen estimations on all samples which were pre- pared or purified by us gave, within experimental limits, theo- retical values.
The titration curves were determined by use of the hydrogen electrode. The electrode vessel was of the Clark type. Hydrogen was generated electrolytically from sodium hydroxide solution between nickel electrodes, passed over concentrated sulfuric acid to remove sodium hydroxide spray, then passed over hot plat.in- ized asbestos to remove oxygen, bubbled through water, and led into the electrode vessel. The comparison half-cell was a 1.0
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from
P. L. Kirk and C. L. A. Schmidt 239
N KC1 calomel cell frequently checked against other similar cells of various ages. This was connected to the hydrogen electrode by means of saturated potassium chloride. All cells were kept at 25” in an air thermostat. The potentials were measured by means of a Leeds and Northrup type K potentiometer,2 a sensi- tive light beam galvanometer being used for zero point instrument.
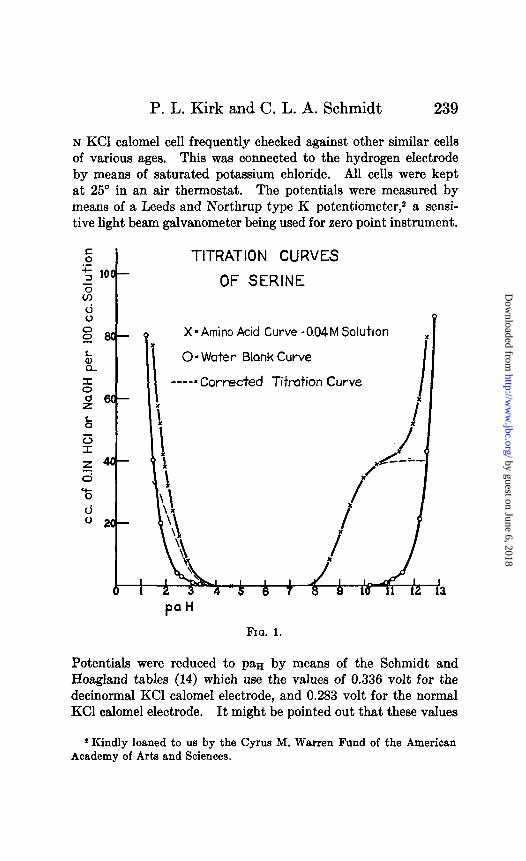
TITRATION CURVES
OF SERINE
I
X=Amino Acid Curve -0.04M Solutton x
I
00 Water Blank Curve
----= Corrected Tit&ion Curve
FIG. 1.
Potentials were reduced to pan by means of the Schmidt and Hoagland tables (14) which use the values of 0.336 volt for the decinormal KC1 calomel electrode, and 0.283 volt for the normal KC1 calomel electrode. It might be pointed out that these values
* Kindly loaned to us by the Cyrus M. Warren Fund of the American Academy of Arts and Sciences.
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from
240 Amino Acid Dissociation Constants
are in much better agreement with recent work (15-17) than the older Sorensen values (18). The pan in all solutions which were acid in reaction was estimated by means of the quinhydrone electrode, with the 1.0 N KC1 calomel electrode as the reference
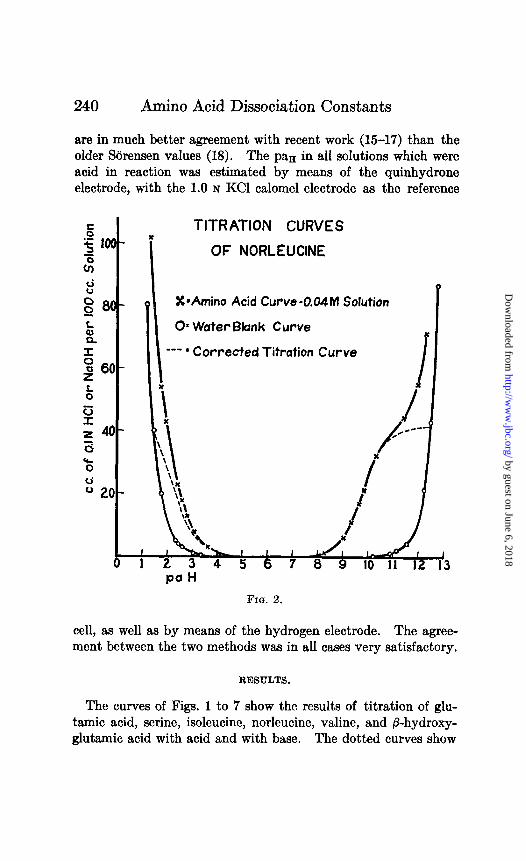
TITRATION CURVES Y
OF NORLEUCINE
X*Amino Acid Curve-0.04M Solution
O* Water Bknk Curve
- * Corrected Titration Curve
FIG. 2.
cell, as well as by means of the hydrogen electrode. The agree- ment between the two methods was in a11 cases very satisfactory.
RESULTS.
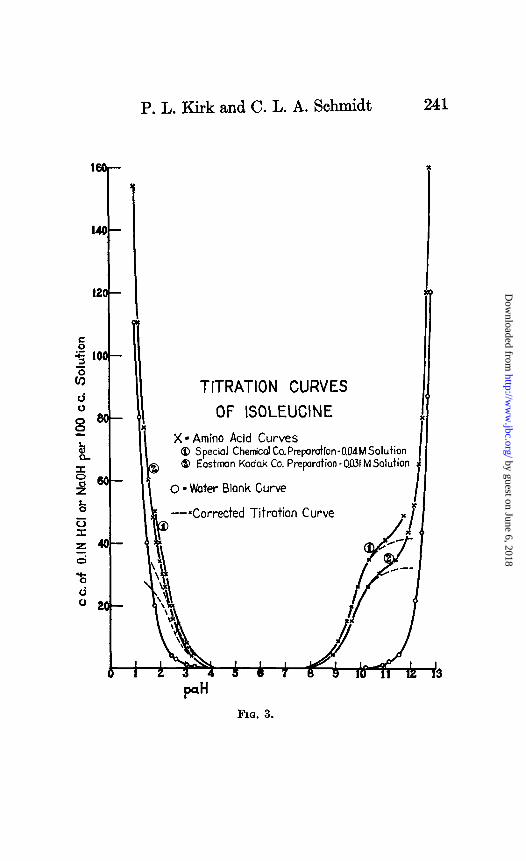
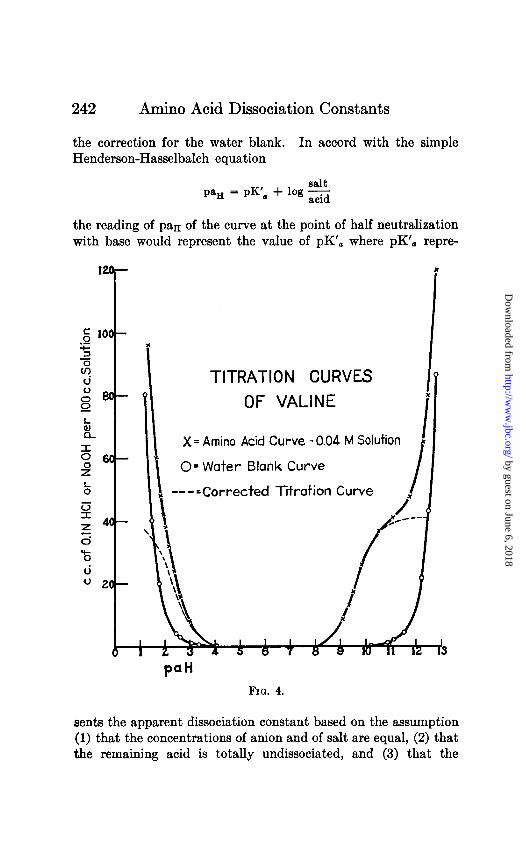
The curves of Figs. 1 to 7 show the results of titration of glu- tamic acid, serine, isoleucine, norleucine, valine, and P-hydroxy- glutamic acid with acid and with base. The dotted curves show
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from
P. L. Kirk and C. L. A. Schmidt 241
--*Corrected Titration Curve
TITRATION CURVES
OF ISOLEUCINE
X= Amino Acid Curves @ Special Chemical Co.Prepamtion-MdMSalution @ Eastman Kodak Co. Prqaration - 0.031 M Solution
o - Woter Blank Curve
P” FIO. 3.
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from
242 Amino Acid Dissociation Constants
the correction for the water blank. In accord with the simple Henderson-Hasselbalch equation
salt psH = PK’, + log --T-&
the reading of pan of the curve at the point of half neutralization with base would represent the value of pK’, where pK’, repre-
!
IO-
3c-
K-
10-
!
TITRATION CURVES
OF VALINE
X = Amino Acid Curve - 0.04 M Solution I x
0= Water Blank Curve
I I( ---=Corrected Titration Curve I .
Pa” Fro. 4.
sents the apparent dissociation constant based on the assumption (1) that the concentrations of anion and of salt are equal, (2) that the remaining acid is totally undissociated, and (3) that the
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from

P. L. Kirk and C. L. A. Schmidt
concentrations of both constituents of the system are equal to their activities. Similarly in accord with the simple equation
pan = pK, - pKb + log ‘s
the reading of the point of half neutralization with acid would represent pK, - pKLa or at that point
pK’* = pK, - paH
These equations obviously do not give the true values for pK’, and pK’b because (1) the concentration of salt is not equal to the concentration of anion (or cation) on account of hydrolysis, (2) the inherent errors in points close to the mid point of the curve have an unduly large effect on the position of the mid point. Moreover, the error of plotting is not entirely negligible.
For these reasons it seems preferable to calculate the dissocia- tion constants directly from the data by means of a proper equa- tion, rather than to read them from the titration mid points. This has the advantage of decreasing the error due to deviations of the individual points from their correct values. The following equation was used in calculating values for pK’..
PK“, = PaH + log c, - CNsC + COH-
c Na+ - ‘OH-
where CA is the total amino acid concentration, Ck,,+ and Con- are the concentrations of sodium and hydroxyl ions, respectively, in mols per liter. In the case of the first acid dissociation constant of glutamic acid and &hydroxyglutamic acid, hydrolysis becomes negligible, but the assumption that all the anion comes from the salt is incorrect. Correction is made for this by modifying the equation, obtaining
PK’, = paH + log ‘A - ‘,a+ - c,+
c,, + c,+
Apparent basic dissociation constants may be calculated from the equation
PK’, = pKw - paH + log c, - Ccl- + c,,
c,,- - c,+
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from
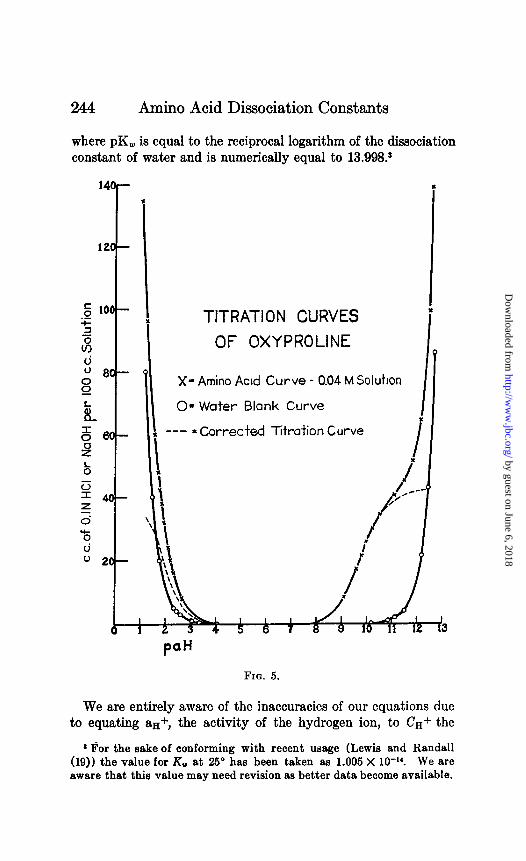
244 Amino Acid Dissociation Constants
where pK, is equal to the reciprocal logarithm of the dissociation constant of water and is numerically equal to 13.998.8
"8 s
TITRATION CURVES r
OF OXYPROLINE
X- Amino Acid Curve - 0.04 M Soluhon
LI II 0= Water Blank Curve
--- = Corrected Titration Curve
FIQ. 5.
We are entirely aware of the inaccuracies of our equations due to equating a=+, the activity of the hydrogen ion, to CH+ the
* F’or the sake of conforming with recent usage (Lewis and Randall (19)) the value for K, at 25” has been taken as 1.005 X 10-l‘. We are aware that this value may need revision as better data become available.
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from
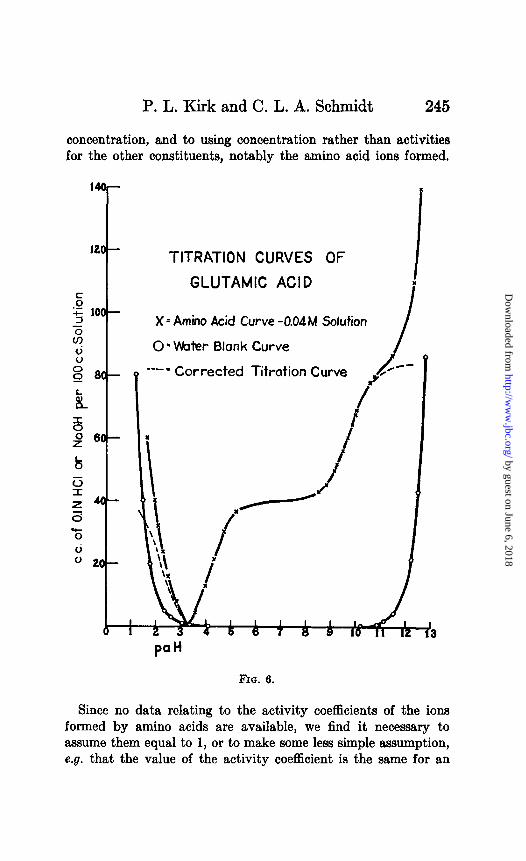
P. L. Kirk and C. L. A. Schmidt 245
concentration, and to using concentration rather than activities for the other constituents, notably the amino acid ions formed.
TITRATION CURVES OF
GLUTAMIC ACID I It
X = Amino Acid Curve -0.04 M Solution
0 = Water Blank Curve
---= Corrected Titration Curve ! /-
/ l
f r
/
3?1a. 6.
Since no data relating to the activity coefiicients of the ions formed by amino acids are available, we find it necessary to assume them equal to 1, or to make some less simple assumption, e.g. that the value of the activity coethcient is the same for an
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from
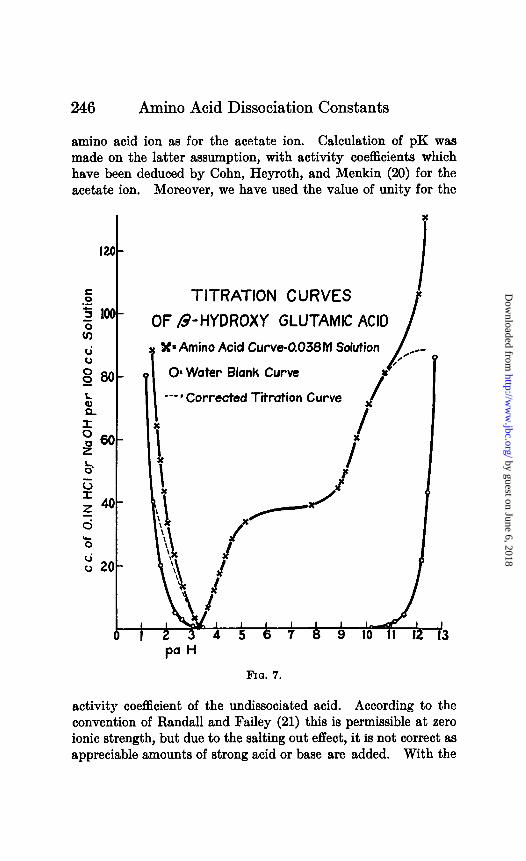
246 Amino Acid Dissociation Constants
ammo acid ion as for the acetate ion. Calculation of pK was made on the latter assumption, with activity coefficients which have been deduced by Cohn, Heyroth, and Menkin (20) for the acetate ion. Moreover, we have used the value of unity for the
TITRATION CURVES I
OF B-HYDROXY GLUTAMIC ACID
Xx Amino Acid Curve-0.038M Solution / ./
,*-- n
0~ Water Blank Curve
---I Corrected Titration Curve
FIO. 7.
activity coefficient of the undissociated acid. According to the convention of Randall and Failey (21) this is permissible at zero ionic strength, but due to the salting out effect, it is not correct as appreciable amounts of strong acid or base are added. With the
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from

P. L. Kirk and C. L. A. Schmidt 247
above assumptions, all values for pK, were found to be slightly higher than for pK’,, the magnitude being roughly 0.05 to 0.1 unit. Due to the fact that the values of pK, thus calculated are less constant than those for pK’,, we consider it preferable to record the latter values obtained by use of the simple incorrect assumption at present. We hope to recalculate the data as soon as values for the activity coefficients of the amino acids become available.4
Table I gives the summarized results of these calculations of the apparent dissociation constants.
TABLE I.
Apparent Dissociation Constants.*
I K’a I K’b PI - B (PK’~ + PK, - PK’I,)
Glutamic acid ............
Serine .................... Valine .................... d-Isoleucine. ............. Oxyproline. .............. @-Hydroxyglutamic acid . .
Norleucine ................
5.62 X lo-& 2.19 x lo-‘0 7.08 x lo-‘0 2.40 x 10-10 2.09 x 10-10 1.86 X lo-‘0 5.82 X 1O-6 2.76 X lo-10 1.72 x lo-lo
1.55 x 10-12 3.22
1.62 x lo-‘* 5.68 2.09 x 10-12 5.97 2.29 x lo-‘* 6.02 8.32 X lo-Is 5.82 2.12 x 10-12 3.28
2.46 X lo-‘* 6.08
* The nomenclature used is in accordance with the classical method. This was done solely for the sake of conforming with established usage and does not imply that the newer concept of ampholyte dissociation as given by Bjerrum (22) is not preferable.
It is seen from these results that the effect of the hydroxyl groups of serine, oxyproline, and &hydroxyglutamic acid are to increase the magnitude of the acid dissociation, themselves having no appreciable power of combining with base. The fact that oxyproline has a basic dissociation, which is lower than an amino group, but still appreciable, ranks it with proline as a possible source of acid binding in the protein molecule.
The values for the isoelectric point, given in the fourth column of Table I, are simply those calculated from the well known equa-
‘Work on the activity coefficients of amino acids is in progress.
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from

248 Amino Acid Dissociation Constants
tion independently of the obvious fact that for all the amino acids except glutamic acid and /3-hydroxyglutamic acid, the iso- electric region is a wide zone, dissociation only taking place in appreciably acid or basic regions.
BIBLIOGRAPHY.
1. Greenberg, D. M., and Schmidt, C. L. A., Proc. Sot. Exp. Biol. and Med., 21, 281 (1923-24). Bracewell, R. S., J. Am. Chem. Sot., 41, 1511 (1919).
2. Cohn, E. J., and Berggren, R. E. L., J. Gen. Physiol., 7, 45 (1924). Cohn, E. J., Physiol. Rev., 6, 349 (1925).
3. Lewis, P. S., Biochem. J., 21, 46 (1927). 4. Redfield, A. C., and Mason, E. D., J. Biol. Chem., 77, 451 (1928). 5. McCay, C. M., and Schmidt, C. L. A., J. Gen. Physiol., 9, 333 (1926). 6. Holmberg, B., Z. p/q&k. Chem., 62, 726 (1908). Walden, P., Z. physik.
Chem., 8, 489 (1891). 7. Harris, L. J., Proc. Roy. Sot. London, Series B, 96, 440 (1923-24). 8. Simms, H. S., J. Gen. Physiol., 11, 629 (1928). 9. Foster, G. L., and Schmidt, C. L. A., Proc. Sot. Exp. Biol. and Med.,
19, 348 (1922); J. Am. Chem. Sot., 46, 1709 (1926); J. Biol. Chem., 66, 545 (1923).
10. Schmidt, C. L. A., and Foster, G. L., Proc. Sot. Exp. Biol. and Med., 18, 205 (1921).
11. Kapfhammer, J., and Eck, R., Z. physiol. Chem., 170, 294 (1927). 12. Leuchs, H., and Geiger, W., Ber. them. Ges., 39, 2644 (1906). 13. Dakin, H. D., Biochem. J., 12,290 (1918); 13, 398 (1919). 14. Schmidt, C. L. A., and Hoagland, D. R., Univ. Calif. Pub. Physiol.,
6, 23 (1919). 16. Kolthoff, J. M., and Bosch, W., Rec. trav. chim. Pays-Bas, 46, 430
(1927). 16. Randall, M., and Young, L. E., J. Am. Chem. Sot., 60, 989 (1928). 17. Scatchard, G., J. Am. Chem. Sot., 47, 696 (1925). 18. Sorensen, S. P. L., Ergebn. Physiol., 12, 393 (1912). 19. Lewis, G. N., and Randall, M., Thermodynamics and the free energy
of chemical substances, New York and London, 1923, 486. 20. Cohn, E. J., Heyroth, F. F., and Menkin, M. F., J. Am. Chem. Sot.,
60, 696 (1928). 21. Randall, M., and Failey, C. F., Chem. Rev., 4, 291 (1927). 22. Bjerrum, N., Z. physik. Chem., 104, 147 (1923).
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Paul L. Kirk and Carl L. A. SchmidtCERTAIN AMINO ACIDS
THE DISSOCIATION CONSTANTS OF
1929, 81:237-248.J. Biol. Chem.
http://www.jbc.org/content/81/2/237.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
ml#ref-list-1
http://www.jbc.org/content/81/2/237.citation.full.htaccessed free atThis article cites 0 references, 0 of which can be
by guest on June 6, 2018http://w
ww
.jbc.org/D
ownloaded from





























