Embed Size (px)
Citation preview

TRD 01: Early Life History Descriptions
THE REGIONAL TRAINING WORKSHOP ON LARVAL FISH IDENTIFICATIONAND FISH EARLY LIFE HISTORY SCIENCE
SEAFDEC/TD, SAMUT PRAKAN, THAILAND16 – 31 MAY 2007

TRD01 page1
From : Leis and Carson-Ewart (2000). The Larvae of Indo-Pacific Coastal Fishes : An Indentificationguide to marine fish larvae. Fauna Malesiana Handbook 2
Terminology adopted
Terminology of the developmental stages of fishes has been the subject of much debate, andmany attempts have been made to define a universal system. Mostly this has resulted in confusion.Because there is so much diversity in the way in which fishes develop, it is unlikely that any onesystem of terminology will ever be accepted by a majority of workers.
The terminology used here is based on the widely used system of Ahlstrom and co-workers(Kendall et al.,1984). We define larval stage to end with the attainment of full external meristiccharacters, the arrival of any mobile structure (for instance, eye of flatfishes or dorsal fin of clupeiformfishes) at its ultimate position, and the loss of temporary specializations to pelagic life (not just theattainment of full fin counts as many workers have done). This definition is adopted for two reasons.In the tropics, the larvae of many benthic species attain the full complement of fin rays but are stilllarvae in the sense of being ‘self-supporting immature forms which differ fundamentally from theparents in structure’ (Tweney&Hughes, 1961). These young stages remain pelagic, are oftentransparent, and they frequently lack scales. Secondly, many tropical fishes have a stages characterizedby striking and often bizarre, temporary, morphological specializations for pelagic life (called theprejuvenile stage by Hubbs, 1958). These stages are often scaled and heavily pigmented but differmarkedly from the adult. It is our feeling that these stages are appropriately termed larval. The larvaland pelagic stages are not synonymous; the young of many benthic fishes remain pelagic for a timefollowing transition as juveniles as, of course, do the young of pelagic fishes. However, for themajority of benthic fishes, the end of larval stage does approximately coincide with settlement. Formost of these, the morphological transition from larva to juvenile is abrupt and takes place over asmall size range. For most pelagic taxa transition is gradual and may take place over an extendedsize range, probably because there is not such a marked change in habitats, and hence selectiveregimes, at the end of the larval stage. Individuals in transition would generally be considered to belarval. The larval stage is further divided into three segments that are defined by formation of thecaudal fin and concomitant flexion of the notochord.
Terminology for developmental stages as used here (Fig. 1)
“egg” : spawning to hatching“larval stage” : hatching to attainment of complete fin ray counts and beginning of
squamation (arrangement of scales on the skin) Yolk sac larva : Development stage beginning with hatching and ending
with exhausting of yolk reserves and characterized presenceof a yolk sac.
Preflexion larva ab : Developmental stage beginning at hatching and ending atthe start of upward flexion of the notochord.
Flexion larva ab : Development stage beginning with flexion of thenotochord and ending with the hypural bones assuming avertical position.
Postflexion larva ab : Development stage from formation of the caudal fin(hypural element vertical) to attainment of full externalmeristic complements (fin rays).
Transition larva b

TRD01 page2
“juvenile b” : completion of fin ray counts and beginning of squamation until fishenters adult population or attain sexual maturity.
“adult b”
a Yolk sac may be present.b Settlement may occur during this stage or not at all if the adults are pelagic.
This system is flexible and has the virtue of simplicity in both concept and terminology. Forexample, fishes that are live bearers, or never develop a caudal fin, or hatch with the caudal fin fullydeveloped, can be accommodated by skipping the inappropriate stages. Our goal in this terminologyhas been to minimize the number and complexity of terms without becoming too general. To theseends we have not used terms that etymologically contradict their stated definitions (for instance,prolarva and postlarva) that have either been unclearly defined or misunderstood and used in severalways (for instance, post larva, prejuvenile) or that are exceeding complex (for instance,protopterygiolarva). A further reason for abandonment of ‘prejuvenile’ is the difficulty often presentin deciding what constitutes a ‘more or less strikingly – often bizarrely – modified pelagic lifehistory stage… not appropriately termed either postlarval or juvenile’ (Hubb,1958). Acanthuridsqualify, and perhaps serranids, but do atherinids (which have a strikingly short gut) or gobiids(which have a strikingly large gas bladder)? The system used here works reasonably well for taxonomicand ecological investigations of tropical marine fish larvae of pelagic, reef, and soft-bottom demersalspecies and is consistent with the biological concept of the larva.

TRD01 page3
Fig. 1 Development stages (Early life history stage of Trachurua symmetricus from Ahlstrom fromand Ball ,1954 cited in Kendall, Ahlstrom and Moser, 1983)

TRD01 page4
Abbreviations
These abbreviations are used throughout the text. The terms are defined in the Glossary, and illustratedin Fig. 2. A few infrequently used abbreviations are defined in the text.
A anal fin P1L pectoral-fin lengthBD* body depth P2 pelvic finBL* body length P2L pelvic-fin lengthBW body width PAL* preanal lengthC caudal fin PDL* pre dorsal-fin lengthD dorsal fin PGBL pre gas-bladder lengthDSL dorsal-fin-spine length POSL preopercular-spine lengthED* eye diameter PP2L pre pelvic-fin lengthHL* head length SL standard lengthHW head width SnL* snout lengthMW mouth width TL total lengthNL notochord length UL unspecified lengthP1 pectoral fin VAFL* vent to anal-fin length
*Measurements taken routinely, see Fig.2D
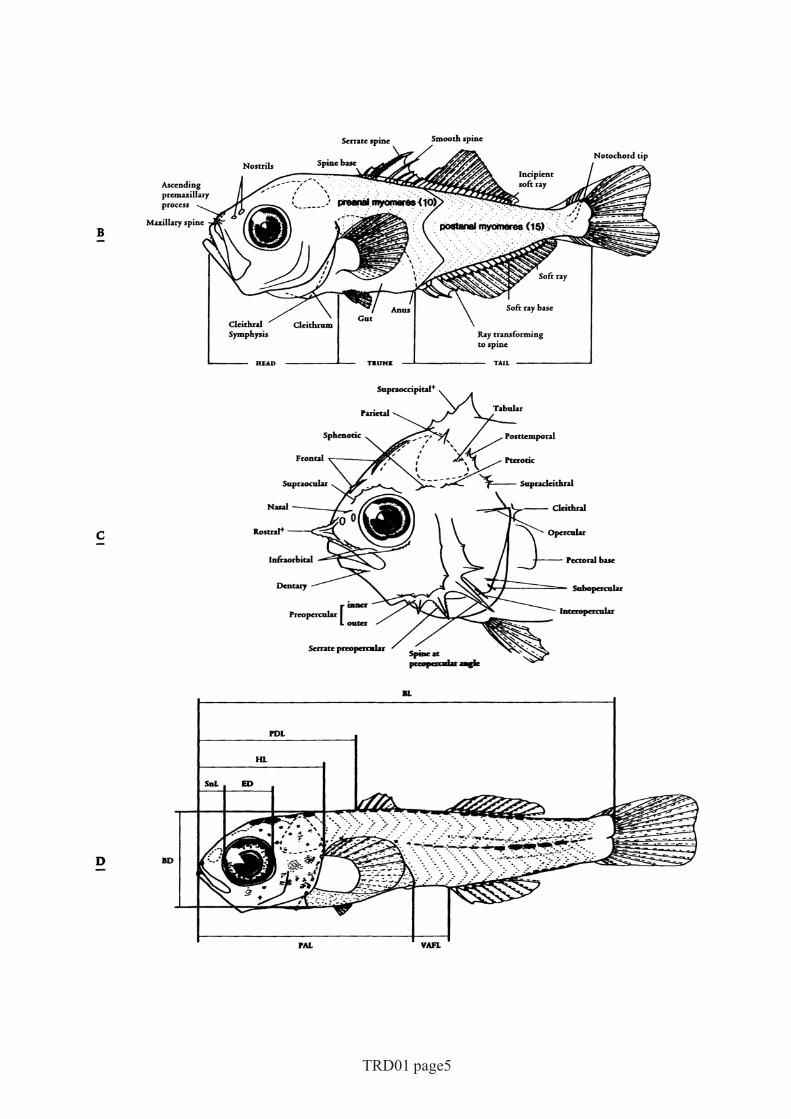
Fig 2 The major morphological characters and measurements of fish larvae used in this book.Many of the terms shown are defined in the glossary.A hypothetical preflexion larvaB hypothetical postflexion larvaC head of a hypothetical larva showing head and pectoral girdle spination; spines marked
with ‘+’ are unpaired medial spinesD postflexion mullid larva showing the measurements taken routinely. Abbreviations
are defined in the text.
TRD01 page5
TRD01 page6
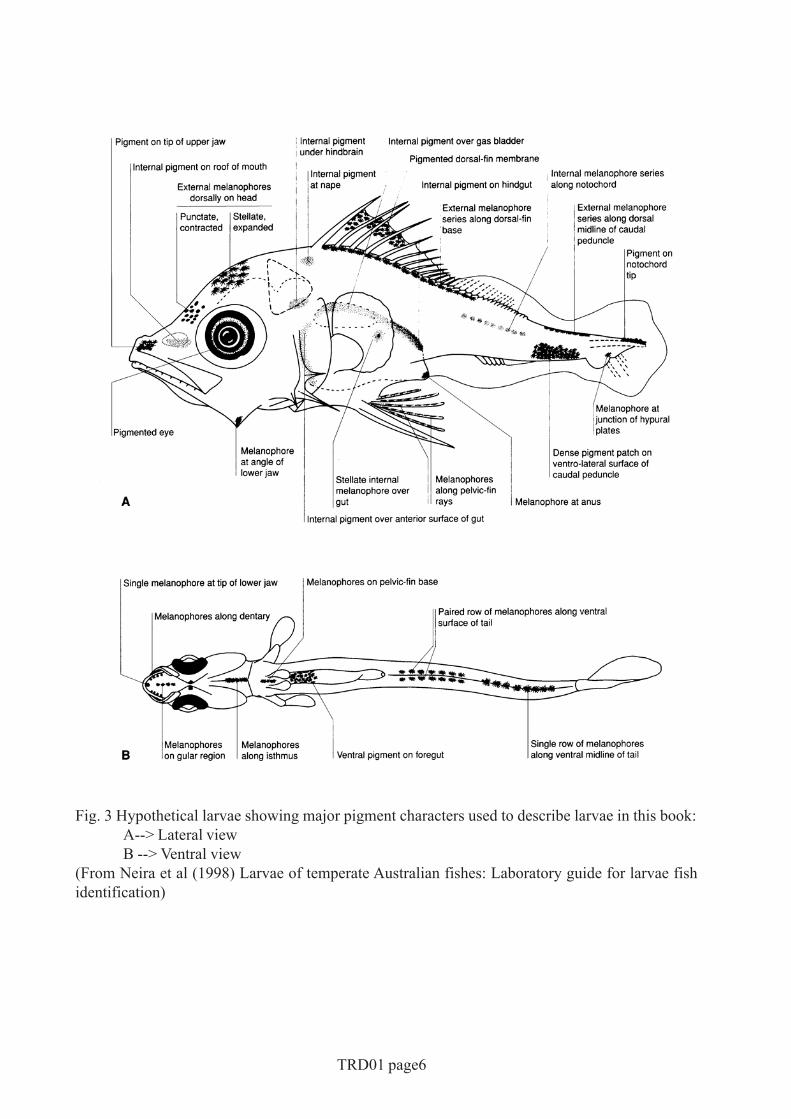
Fig. 3 Hypothetical larvae showing major pigment characters used to describe larvae in this book:A--> Lateral viewB --> Ventral view
(From Neira et al (1998) Larvae of temperate Australian fishes: Laboratory guide for larvae fishidentification)

TRD01 page7
Characters as used in descriptions of larvae
Body ShapeIn our descriptions of the general shape of the body, we use the following broadcategories that relate body depth (BD) to body length (BL):Very Elongate: BD < 10% BLElongate: BD 10-20% BLModerate: BD 20-40% BLDeep: BD 40-70% BLVery Deep: BD > 70% BL
Categories used to define head size relate head length (HL) to BL;Small Head: HL < 20% BLModerate Head: HL 20-33% BLLarge Head: HL > 33% BL
Categories used to define eye size relate eye diameter (ED) to HL;Small Eye: ED < 25% HLModerate Eye: ED25-33% HLLarge Eye: ED>33% HL
All references to size of body parts are to relative size. Thus, a structure that does not changein absolute size may be said to reduce, or to have become relatively smaller, because thebody as a whole has grown. Similarly, if growth of a given structure is positive but does notkeep pace with overall growth, it will reduce in relative size, perhaps from large to moderate.Few structures except very large fin or head spines actually become absolutely smaller duringdevelopment.
MyomeresMyomere counts include all myomeres bounded anteriorly by a myoseptum, and are dividedinto pre-and postanal element (per + post). This is not necessarily equivalent to the divisionof vertebrae between precaudal and caudal centra because the position of the anus maychange ontogenetically or may be anterior to the posterior edge of the abdominal cavity.There is a near one-to-one correspondence between total number of myomeres and numbersof vertebrae. In preflexion larvae notochord segmentation posterior to the terminal myomeremay be mistaken for additional myomeres.
GutThe gut of fish larvae always starts as a straight tube. This subsequently differentiates intofunctionally different sections which may be visually discernible: a portion of the gut maybe striated, for example. The gut usually folds or coils into loops, thereby increasing itslength without increasing body length. The folding may take place before hatching, butmore commonly it happens during or shortly after the yolk-sac stage and in some cases aftertransformation to the juvenile stage. The timing of folding and its extent are species-specificand therefore useful taxonomic characters.
Gas bladderThe buoyancy-regulating gas bladder is present in most larvae but is lost in the adults ofsome taxa (for instance, gobiids). In these species the gas bladder is not really a temporaryspecialization to pelagic larval life: it is the loss of the gas bladder in adults that is the

TRD01 page8
specialization. Two factors alter the size or degree of inflation of the gas bladder. Larvaecaptured at depth and brought rapidly to the surface may have expanded gas bladders due tothe pressure change. Secondly, many taxa have a small, inconspicuous gas bladder duringthe day but a strongly inflated, conspicuous gas bladder at night (for instance, clupeiformes,sillaginids). Thus, larvae collected at night may appear different from those collected duringthe day because of nocturnal gas bladder inflation. However, Larvae that normally haveinflated gas bladders at night may have deflated bladders if collected at night around a light.Finally, in some taxa, the gas bladder migrates ontogenetically. For example, in clupeiformes,anterior migration of the gas bladder is common, whereas in many gobioid larvae, the gasbladder migrates poateriorly.
Head spinationOne of the most striking larval specializations is the head spination found in many speciesthat lack head spines as adults. The most widespread type of head spination is that on theinner and outer preopercular borders. However, spines may appear on any part of the head(Fig.2). The sequence of development, degree of elongation, placement, number, andornamentation of the spines are important characters for identification, and we note these inthe descriptions. Most of these spines are resorbed, overgrown or incorporated into sensorycanals by the end of the larval stage, but some remain well into the juvenile stage, and somespines may be retained by the adults. In describring the preopercular spines, the followingdefinitions of length apply: small – shorter than the pupil diameter; moderate – larger thanpupil diameter, but do not extend over the border of the opercular series (opercle, subopercleand interopercle); large – extend past the border of the opercular series. The head spinesprobably have a defensive function (Moser, 1981). We include among head spines the spinesof the upper pectoral girdle (supracleithrum, cleithrum, postcliethrum). Nomenclature ofthe borders (or edges) of the preopercle can be confusing. What some authors refer to as theposterior border or the medial border or ridge, we call the outer border because, radially, it isoutside the more enclosed inner border (called anterior, or lateral border or ridge by some).Almost without exception, the spines of the outer border are larger and more numerous thanthose of the inner border.
EyesMost fish have round or nearly round eyes. A large number have eyes shaped like roundedsquares or rectangles (squarish) or than are slightly elongate (longer than high). Finally, arelatively few have eyes than are much smaller laterally than they are vertically (narroweyes, Weihs & Moser, 1981). Choroid tissue is a mass of apparently undifferentiated tissuethat clings to the ventral margin of the eye of the larvae of some species. It may be involvedin metamorphosis of the eye (Moser & Ahlstrom, 1970), and it is usually unpigmentedalthough in a few species it may be nearly as dark as the eye. Choroid tissue seems to bemost common in larvae with narrow eyes.
Fin formationThe fist stage in development of the medial fins is the undifferentiated finfold. The finfold isinitially continuous (or nearly so) from occiput around the tail to near the cleithral symphysis,but it becomes discontinuous and eventually disappears as the fins differentiate. Thickenings,or anlagen, appear in the finfold, usually along the finfold/body interface, but at the peripheryof the finfold in a few species. The anlagen soon begin to segment into discrete finbases.The soft-ray bases are typically massive and elongate, rounded at the distal end, and havelittle or no space between them. The rounded end of the base articulates with the soft ray viaa ball and socket joint. The incipient rays are thickenings oriented more or less perpendicularto the body axis in the finfold and extending the width of the finfold. The rays are considered

TRD01 page9
to have formed (i.e. no longer incipient) when their ossification begins. One ray forms oneach base, although the last ray may be divided nearly to the base. After formation the rayssegment and may branch.
Spines form from triangular bases than are usually much smaller than ray bases. The spinesbases are generally separated from one another, and the finfold may disappear in theinterspaces while the spines are still incipient. Again, there is one spine per base. It is notunusual for the last spine of the dorsal or anal fin to form as a soft ray (complete withsegments) that subsequently hardens into a spine. In this case, the base of this element isintermediate in shape and size between a spine base and ray base. Because of the differencesbetween the types of base, it is usually possible to distinguish between spines and raysbefore these elements are fully formed. Even after the spines and rays are present, it may beeasier to use bases rather than the spines or rays to obtain fin formulate. This is especiallytrue in species with weak or flexible spines such as labrids.
The caudal fin has some developmental peculiarities. In most teleosts (there are exceptionsamong the Gadiformes and Ophidiiformes), the caudal fin forms from a ventral anlage nearthe tip of the notochord. The anlage breaks up into only a few segments, so there is not a 1:1ratio of basal segments to fin rays. While the rays form, the notochord flexes upward, bringingthe rays parallel to the body axis (see Ahlstrom & Moser, 1976). The basal segments becomethe supporting bones of the caudal complex.
The paired fins form from buds, do not form separate bases for each element, and passthrough an incipient-ray stage. The pectoral bud forms very early, usually in the yolk-sacstage, but the pectoral rays may form very late. The pelvic fin often develops a spine (this istaxon-dependent), but the pectoral fin does not in any of the fish considered here.
The usual pattern of fin development is for the caudal fin to form fist. Near the end ofnotochord flexion, the soft rays of the dorsal and anal fins begin to ossify. The spines usuallyossify after the soft rays. The pectoral rays form next, and the pelvic fin is usually last. Thereare many species-dependent exceptions to this generalized pattern, however, and any finmay be the first or last to form.
Certain fin spines and rays or even whole fins may become very long or become ornamentedin a taxon-specific manner with various hard (for instance, serrations) or soft (for instance,fleshy bulbs) structures. These are nearly always tempolary specializations to the pelagiclarval stage and probably serve in defence against predators (Moser, 1981), although it ispossible some of the soft structures aid in flotation or camouflage.
SizeFor each family, we give a table of size at which certain developmental events occur andsize of the smallest and largest specimen examined. The ranges given reflect variation amongtaxa, while the size of the smallest and largest specimens examined give the size range overwhich our descriptions apply and how large the larvae or juveniles may become prior tosettlement. Where a range of sizes is given for smallest or largest, the sizes of more than onespecies and included. The range given for size at flexion is the largest preflexion specimenand the smallest postflexion specimen.

TRD01 page10
MorphometricsIn the morphometrics tables measurements are expressed as a proportion of body length.Body length for preflexion and flexion larvae is notochord length. Body length for postflexionlarvae is standard length. Certain measurements were made routinely on all specimens (seelist of abbreviations on page 5); others were made when needed. Unless noted other otherwisein the text, all lengths are body length as specified here.
The variation in morphometrics of larval fishes is often high because of diversity in, andontogenetic changes to, body shape, damage or distortions suffered during collection, orshrinkage. Shrinkage is probably a result of water loss from hypertonic tissues of the larvaafter death and is particularly severe in small larvae before the vertebral column is ossified.Changes in length of 33% following death and preservation have been reported for newlyhatched, yolk-sac larvae (Miller & Sumida, 1974). Several factors can influence the degreeof shrinkage including time between death and preservation, type of preservative, and methodof collection (Theilacker, 1980). Because larvae usually shrink when preserved, one oftenencounters preserved larvae of a smaller size than that reported for newly-hatched larvae(which are often measured alive).
In the Morphometrics tables we include flexion-stage larvae with preflexion larvae. Alllarvae were measured under a dissection microscope equipped with an eyepiece micrometeror a digitizing pad. Magnification varied from 6 to 50 X depending on the size of the specimen,and precision of measurements ranged from 0.02 to 0.13 mm depending on magnification.Measurements in the text were usually rounded to the nearest 0.1 mm if less than 10 mm, tothe nearest 0.5 mm between 10 and 20 mm, and to the nearest 1 mm if greater than 20 mm.
PigmentWe are concerned here only with melanin. Although live fish larvae may have a number ofpigments, the preservatives and fixatives commonly used quickly bleach the reds, yellows,blues and silvers, leaving only the browns and blacks of melanin. The terms melanophoresand pigment are thus used interchangeably, and all references are to pigment in preservedspecimens. In the illustrations, pigment on the surface of the body (external pigment) isportrayed as shown in Fig.2A. Internal pigment is portrayed with stipples. We depart fromthis rule by using stipples to portray surface pigment that is widespread and of relativelyuniform density (usually on late larvae or early juvenile specimens).
Meristics tableNearly all meristic data were derived from the literature. Many sources were used, and wehave not cited these to save space; however, reference to the papers cited in the adult sectionor one of the major faunal works will give a good entry to the literature on the family. In afew cases we made our own counts from specimens or radiographs. We attempted tostandardize literature counts taken by different methods (see below), but in some cases themethods (for instance, if the urostyle is included in vertebral counts) were not recorded sosome errors may have been introduced. Other possible sources of error were erroneouscounts (often perpetuated from source to source) and artificially truncated ranges based ontoo few specimens or species. We attempted to include counts from all recognized speciesunder each genus, but in most cases complete counts were not available for all species.Therefore, the Meristics Tables should be used with caution. In these tables, a ‘_’ indicatesno data.

TRD01 page11
Fin-ray and vertebral counts
Spines are indicated by Roman numerals, and soft rays by Arabic numerals. A commaindicates an undivided fin, and a ‘+’ indicates a divided fin with the exception of the caudalfin where a ‘+’ indicates the division between dorsal and ventral primary rays. A particularproblem concerns caudal rays: the counts given are for principal (used interchangeably withprimary) rays. This count is defined in the literature in at least three ways:
1. the number of branched rays plus two (Hubbs & Lagler, 1964);2. the rays which articulate with the hypural bones (Miller & Jorgenson, 1973) however,
these authors note ‘our judgment, based upon our interpretation of the literature,was used to determine this count’; and
3. the rays supported by the hypurals and parahypural (Moser et al., 1977).
It was often impossible to determine in published accounts which method was used, andwhile definitions 1 and 3 often give identical counts, this is not always true. Definition 1 isthe least useful for larvae, as fin rays branch late in ontogeny. If counts were given asbranched rays rather than as principal, we have so indicated. We frequently used the bookon caudal fin osteology by Fujita (1990) to obtain the required information.
Preferred counts (please note caution above):
- Caudal (C): the rays supported by the hypurals and parahypural.- Dorsal (D), and (A): each element with a separate base was counted.- Pectoral (P1): including all elements, usually without regard to segmentation or
branching.- Pelvic (P2): all elements- Vertebrae: all elements including the urostyle, divided if possible into precaudal and
caudal vertebrae.

TRD01 page12
Glossary
We assume the user will have a working knowledge of adult fishes, so we define onlythose terms of special application to fish larvae. We refer the reader without such a workingknowledge to a modern ichthyology text such as Helfman et al. (1971), or to a work on fish osteologysuch as Rojo (1991). General terminology mostly follow that of Hubbs & Lagler (1964). Forspines of the head we follow the terminology of Fig. 2C. Some infrequently occurring spines notshown in Fig. 2C are named after the bone from which they are derived.
AAnlage – Rudimentary form of an anatomical structure; primordium; a German German word the
plural of which is Anlagen.
Bbody depth – The vertical distance between body margins (exclusive of fins) through the anterior
margin of the pectoral-fin base: not necessarily the greatest body depth.body length – Size of the larva; corresponds to notochord length in preflexion and flexion larvae
and to standard length after flexion.body width – The transverse distance between body margins at the pectoral-fin base.
Cchoroids fissure – Line of juncture of invaginating borders of the optic cup; apparent in young fish
as a trough-like area below lens.choroids tissue – An undifferentiated mass of (usually) unpigmented tissue adhering to the ventral
surface of the eye (see Moser & Ahlstrom 1970).cleithral symphysis – The cartilaginous joint between the two cleithra where they join ventrally
Often forms a prominent point along the ventral profile.cleithrum – Prominent bone of pectoral girdle, clearly visible in many fish larvae.
Ddemersal egg – An egg which remains on the bottom either free or attached to the substratum.
Eexterilium larva – A term referring to the larvae of some unidentified species of the ophidiid subfamily
Neobythinae characterized by an elongate, compressed body and a very long, trailinggut ornamented with flaps and streamers (illustrations are found in Gordon et al., 1984C); Okiyama, 1988d (p. 340).
eye diameter – The horizontal distance across the midline of the pigmented region of the eye.
Ffin elements – Fin supports; spines and soft rays.finfold – Medial fold of integument that extends along body of developing fishes and from which
media fins arise.fin length – Length of the longest soft ray of designated fin.flexion – Bending upward of the notochord tip as part of the process of caudal-fin formation.flexion larva – Development stage beginning with flexion of the notochord and ending with the
hypural bones assuming a vertical position.Ggas bladder – Membranous, gas-filled organ located between the kidneys and alimentary canal in
teleost fishes; air bladder or swim bladder.

TRD01 page13
Hhead length – The horizontal distance from the tip of snout to posterior-most part of opercular
membrane; prior to development of operculum, measured to the posterior margin ofcleithrum.
head width - The transverse distance between margins at the widest area of the head. Not applicableto larvae with flared opercles.
Hypural bones – The basal bones of the caudal fin that support the caudal-fin rays.
Iincipient fin-ray – Early stage in fin-ray development; and unossified thickening in finfold articulating
with the fin base.initially – When used in the Morphology section, meaning in the smallest available specimens.interopercle – A bone of the skull that may bear spination (see Fig. 2).
Jjuvenile – Developmental stage from attainment of full external meristic complements and loss of
temporary specializations for pelagic life to sexual maturity.
Klachrymal – The anterior bone of the infraorbital series. Frequently overlaps the maxilla when the
mouth is closed.larva- Developmental stage between hatching (or birth) and attainment of full external meristic
complements (fins and scales) and loss of temporary specializations for pelagic life;yolk-sac through postflexion stage inclusive.
Mmelanin – A black pigment.melanophores – Melanin-bearing cells (brown to black); frequently capable of expansions and
contractions which change their size and shape.mouth width – The gape; transverse distance between corners of the mouth.myomeres – Serial muscle bundles of the body.myosepta – Connective tissue partitions separating adjacent myomeres.
Nnotochord - Longitudinal supporting axis of body which is eventually replaced as a support by the
vertebral column in teleostean fishes.notochord length – Straight line distance from tip of snout to posterior tip of notochord; used prior
to and during flexion.
Oolfactory pit – A shallow depression on the snout from which olfactory organ develops.opercle (operculum) – Bone of the skull that may bear spination (see Fig. 2).ossification – Hardening of bony parts through deposition of calcium salts; usually detected by
staining with alizarinotic capsule – Sensory anlage from which the ear develops; clearly visible during early development
Pparahypural – Modified haemal spine of the penultimate vertebral centrum. Supports some caudal-
fin rays.
TRD01 page14
pectoral bud – Swelling at site of future pectoral fin; anlage of pectoral fin.pelagic – In the water column as distrinct from substrate-associated; neither necessarily planktonic
nor oceanic.pelagic egg – Egg which floats freely in the water column, often slightly positively buoyant.pelagic juvenile – Pre-settlement juvenile of a species that is benthic or reef-associated as an adult.pelvic bud – Swelling at site of future pelvic (ventral) fin; anlage of pelvic fin.postanal mymeres – Myomeres posterior to the posterior margin of the anus; includes terminal
myomere from which urostyle forms.postcleithrum – Bone of the pectoral girdle that may bear a small spine just dorsal to the pectoral-fin
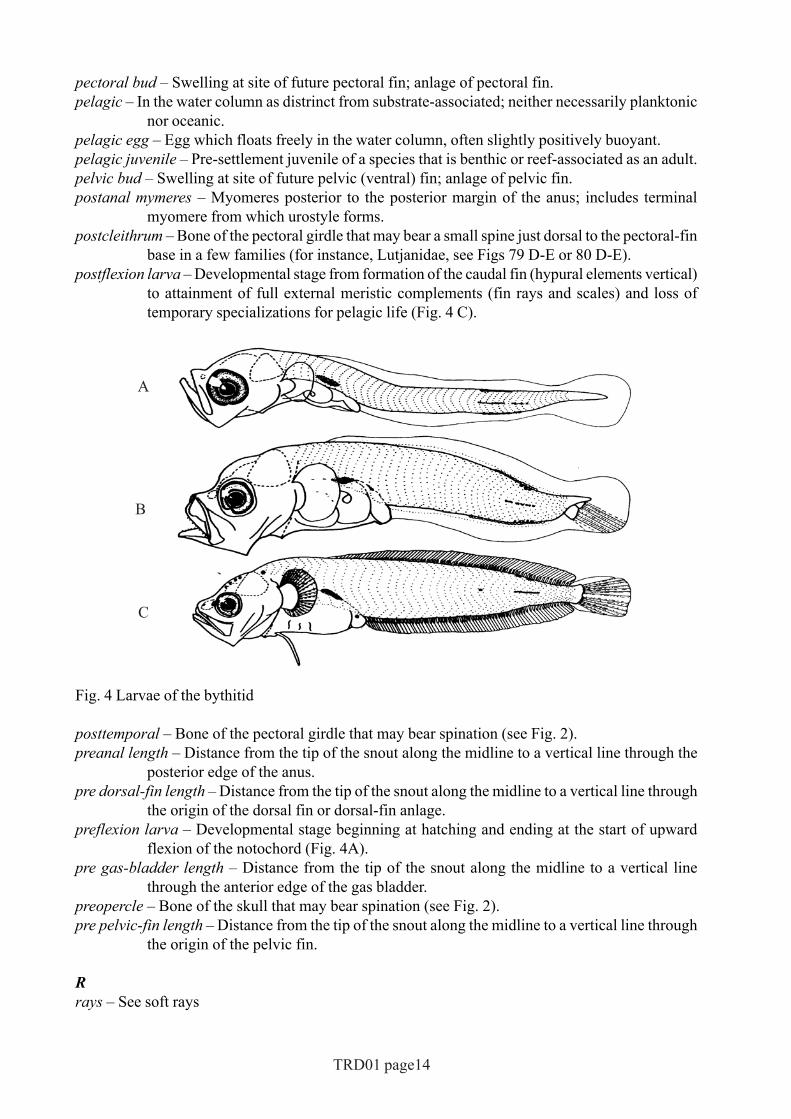
base in a few families (for instance, Lutjanidae, see Figs 79 D-E or 80 D-E).postflexion larva – Developmental stage from formation of the caudal fin (hypural elements vertical)
to attainment of full external meristic complements (fin rays and scales) and loss oftemporary specializations for pelagic life (Fig. 4 C).
Fig. 4 Larvae of the bythitid
posttemporal – Bone of the pectoral girdle that may bear spination (see Fig. 2).preanal length – Distance from the tip of the snout along the midline to a vertical line through the
posterior edge of the anus.pre dorsal-fin length – Distance from the tip of the snout along the midline to a vertical line through
the origin of the dorsal fin or dorsal-fin anlage.preflexion larva – Developmental stage beginning at hatching and ending at the start of upward
flexion of the notochord (Fig. 4A).pre gas-bladder length – Distance from the tip of the snout along the midline to a vertical line
through the anterior edge of the gas bladder.preopercle – Bone of the skull that may bear spination (see Fig. 2).pre pelvic-fin length – Distance from the tip of the snout along the midline to a vertical line through
the origin of the pelvic fin.
Rrays – See soft rays
A
B
C

TRD01 page15
Ssettlement – Process by which a larva or juvenile leaves the pelagic environment and adopts a
substrate-associated lifestyle; not applicable to species pelagic as adults.settlement stage – Development stage where the pelagic larva or juvenile is morphologically and
physiologically ready (competent) to adopt a substrate- associated life style. Often, butnot always, associated with larva to juvenile transition. Applicable only to species thatare not pelagic as adults.
snout length – Horizontal distance from the tip of the snout to the anterior margin of the pigmentedregion of the eye.
soft rays – Bilaterally paired, usually segmented fin supports; often referred to as rays.spine length – Straight line distance of a fin or body spine from base to tip.spines – 1 Unpaired, unsegmented, unbranched fin supports, usually (but not always) stiff and
pungent; may initially form as bilaterally paired structures prior to ossification.2 Bony, pointed, elongate projections on the body, usually the head, that are often
temporary specializations for pelagic life.standard length – Distance from tip of the snout along the midline to a vertical line through the
posterior edge of the hypural plate.subopercle – Bone of the skull that may bear spination (see Fig. 2)supracleithrum – Bone of the pectoral girdle that may bear spination (see Fig. 2)Ttail – Portion of body posterior to the anus.teeth – Larvae of some species develop ‘larval teeth’ prior to the appearance of definitive teeth.
Larval teeth are little exserted spines on the premaxilla or dentary, and they are lostduring the larval phase (Baldwin, 1990). The descriptions herein do not generallydistinguish between ‘larval teeth’ and definitive teeth.
temporary specializations for pelagic life – Morphological (not pigment) characters such as trailingguts, prepercular spines, or elongate fin spines that are not retained in adults but arepresent during the pelagic phase. Often these are lost after settlement, but if present insettled individuals, they are clearly reduced and are disappearing.
total length – Distance from the tip of the snout along the midline to the posterior edge of the caudalfinfold; body length is traditionally expressed as total length in the Japanese literature.
transition – Change from the larval to the juvenile stage. May take place over an extended period oftime. Especially used for pelagic taxa where there is not a change in habitat at or nearthe end of the larval phase. Individuals in transition are considered larval. In the text,we avoid the use of the term metamorphosis because of its implications of abrupt andextensive morphological change over a short period.
trunk – Body between head and anus.
Uunspecified length – Undefined measurement of body length derived from the literature.urostyle – the posterior-most vertebral centrum.
Vvent to anal-fin length – Straight line distance from posterior edge of the vent (anus) to the anterior
origin of the anal fin or anal-fin anlage.
Yyolk sac – A bag-like, ventral extension of the primitive gut containing the yolk.Yolk-sac larva – Developmental stage beginning with hatching and ending with exhaustion of yolk
reserves and characterized by presence of a yolk sac.

TRD01 page16
Literature often used for identification of fish larvae
1. Fahay, M.P. 1983. Guide to the early stages of marine fishes occurring in the Western NorthAtlantic Ocean, Cape Hatteras to the Southern Scotian Shelf. J. Northw. Atl. Fish. Sci. 4: 423 pp.
2. Fritzsche, R.A. 1978. Development of fish of the Mid-Atlantic Bight, and atlas of eggs, larvaland juvenile stages. Vol. V. Chaetodontidae through Ophidiidae, U.S. Fish. Wild. Serv.,Biol. Serv. Prog. FWS/OBS-78/12.
3. Leis, J.M. and B.M. Carson-Ewart. 2000. The Larvae of Indo-Pacific Coastal Fishes: Anidentification guide to marine fish larvae. Fauna Malesiana; Vol. 2. Leiden; Boston;Koln: Brill. 850 pp.
4. Moser, H.G., W.J. Richards, D.M. Cohen, M.P. Fahay, A.W. Kendall Jr. and S.L. Richardson.(eds.). 1984. Ontogeny and Systematic of Fishes. An International Symposium Dedicatedto the Memory of E.H. Ahlstrom, Special Publication no. 1, American Society ofIchthyologists and Herpetologists. 760 pp.
5. Niera, F.J., A.G. Miskiewicz and T. Trnski. 1998. Larvae of Temperate Australian Fishes:Laboratory Guide for Larval Fish Identification. University of Western Australia Press.474 pp.
6. Nishikawa, Y. and D.W Rimmer. 1987. Identification of larval tunas, billfishes and otherscombroid fishes (suborder Scombroidei): an illustrated guide. CSIRO MarineLaboratories Report 186, 20 pp.
7. Okiyama, M. 1988. An Atlas of the Early Stage Fishes in Japan. Tokai University Press. 1157pp. (in Japanese).
8. Ozawa, T. 1986. Studies on the oceanic ichthyoplankton in the Western North Pacific KyushuUniversity Press. 430 pp.



















![One fish [Режим совместимости] fish.pdf · Dr. Seuss One fish two fish red fish blue fish. One fish Two fish . Blue fish Red fish. Blue fish Black fish. Old fish](https://img.pdfslide.us/doc/110x75/5fce8df40415697f677cef57/one-fish-fishpdf-dr-seuss-one-fish-two.jpg)









