Embed Size (px)
Citation preview

1585Journal of Cell Science 109, 1585-1596 (1996)Printed in Great Britain © The Company of Biologists Limited 1996JCS4142
Transport pathway, maturation, and targetting of the vesicular stomatitis virus
glycoprotein in skeletal muscle fibers
Paavo Rahkila, Anne Alakangas, Kalervo Väänänen, and Kalervo Metsikkö*
Biocenter and Department of Anatomy, University of Oulu, Kajaanintie 52A, FIN-90220 Oulu, Finland
*Author for correspondence
We have infected isolated skeletal muscle fibers with thevesicular stomatitis virus or the mutant tsO45, whose gly-coprotein is blocked in the endoplasmic reticulum at 39°C.Immunofluorescence analysis for the viral glycoproteinindicated that the fibers were infected over their entirelength at a virus dose of 109/ml. When we infected themyofibers with the tsO45 mutant at 39°C, the viral glyco-protein appeared to be localised to the terminal cisternaeof the sarcoplasmic reticulum. Upon shifting the culturesto the permissive temperature, 32°C, in the presence ofdinitrophenol, which blocks vesicular transport, the viralglycoprotein proceeded to completely fill the sarcoplasmicreticulum. Thus, both the endoplasmic reticulum located atthe terminal cisternae of the sarcoplasmic reticulum, andthe entire endoplasmic and sarcoplasmic reticulumappeared to be continuous. Shifting the culture tempera-ture from 39°C to 20°C, resulted in prominent perinuclearstaining throughout the fibers, accompanied by the appear-ance of distinct bright dots between the nuclei. Electron
microscopic immunoperoxidase labeling indicated thatthese bright structures represented the Golgi apparatus.When either the tsO45-infected or wild-type virus-infectedfibers were incubated at 32°C, the viral glycoproteinshowed a staining pattern that consisted of double rows ofpunctate fluorescence. Immunogold labeling showed thatthe viral glycoprotein was present in both the transversetubules as well as the endoplasmic/sarcoplasmic reticulumendomembranes. In addition, extensive viral budding wasobserved in the transverse tubules. Metabolic labelingexperiments revealed that only half of the glycoprotein wasprocessed in the Golgi, and this processed form had becomeincorporated into the budding viral particles. Thus, theprocessed viral glycoprotein was targeted to the transversetubules. The other half of the glycoprotein remained endo-glycosidase H-sensitive, suggesting its retention in the endo-plasmic/sarcoplasmic reticulum endomembranes.
Key words: Myofiber, Sarcoplasmic reticulum, Transverse tubule
SUMMARY
INTRODUCTION
Muscle cells are produced by the fusion of mononucleatedmyoblasts, thereby forming multinucleated myotubes. Duringthe fusion process, intracellular organelles reorganize andextensive differentiation occurs. The filamentous contractileapparatus develops together with a complex intracellularmembrane network consisting of the sarcoplasmic reticulum(SR) and the transverse (T)-tubulus system (reviewed byFlucher, 1992). In developing myotubes, these are tubulovesic-ular structures with a longitudinal orientation. In adult musclefibers, however, they are highly organized into transversecross-striated structures.
The SR of skeletal muscle fibers contains ER componentssuch as BiP, protein disulfide isomerase and calnexin, stronglysuggesting a continuity between these two structures (Volpe etal., 1992; Villa et al., 1993). Certain ER components, such asthe Ca2+-dependent ATPase, are probably transported directlyfrom the ER to the SR, since they could not be found in theGolgi (Jorgensen et al., 1977). However, calsequestrin, a majorprotein of the SR lumen, was found in the Golgi, suggesting atransport route for this protein through the organelle (Thomas
et al., 1989). Calsequestrin clearly traveled through the Golgiin transfected mononucleated cells (Raichman et al., 1995),while Ca2+-ATPase did not (Karin and Settle, 1992). Thesefindings suggest that proteins are translocated from the ER tothe SR either directly or via the Golgi. It is, however, notknown where the ER is situated in the SR.
In cultured myotubes, the reorganization and synthesis oforganelles is reflected by the protein transport pathways andmaturation processes. Thus, in chick embryo muscle cultures,acetylcholine esterase is not secreted correctly and most of itremains Endo H-sensitive, indicating a lack of Golgi process-ing (Rotundo et al., 1989). Furthermore, a major transport routefrom the Golgi to the SR was observed. In L6 myoblast-derivedmyotubes a fraction of the Golgi stacks were disassembled, anddid not process glycoproteins normally (Kellokumpu et al.,1995). Multinucleated myotubes derived from a C2C12 cell lineseem to secrete glycoproteins efficiently (Yao and Kurachi,1992). A number of studies have shown that transgene productsare secreted in skeletal muscle (Barr and Leiden, 1991; Dhawanet al., 1991; Dai et al., 1992), but the efficiency of the secretionwas not analyzed and it is not known whether the secretionproducts were processed in the muscle fibers. It is not known
1586 P. Rahkila and others
for sure whether skeletal myofibers exhibit a functional Golgiand where it is localized. Jasmin et al. (1989, 1995) havereported that Golgi structures are found only in the vicinity ofthe neuromuscular junction but Ralston (1993), on the contrary,has found Golgi elements throughout the muscle fiber.
Here, we have analyzed organelle localization and glycopro-tein trafficking and processing in isolated skeletal myofibers,using the vesicular stomatitis virus (VSV) as a tool. We showthat the isolated myofibers support VSV infection. Using the ther-mosensitive VSV mutant, tsO45, whose glycoprotein (G protein)is blocked in the ER at the restrictive temperature, we localizedthe ER at the A-I borders. At lower temperatures, the mutant Gprotein had direct access from the ER sites to the whole SR. Sur-prisingly, only a fraction of the viral G protein matured normallyin the Golgi although the latter was present around every nucleusthroughout the muscle fibers. The processed form of the G proteinwas targeted to the T-tubules, but not to the sarcolemma.
MATERIALS AND METHODS
Isolation of myofibersFlexor digitorum brevis muscles from the feet of adult Spraque-
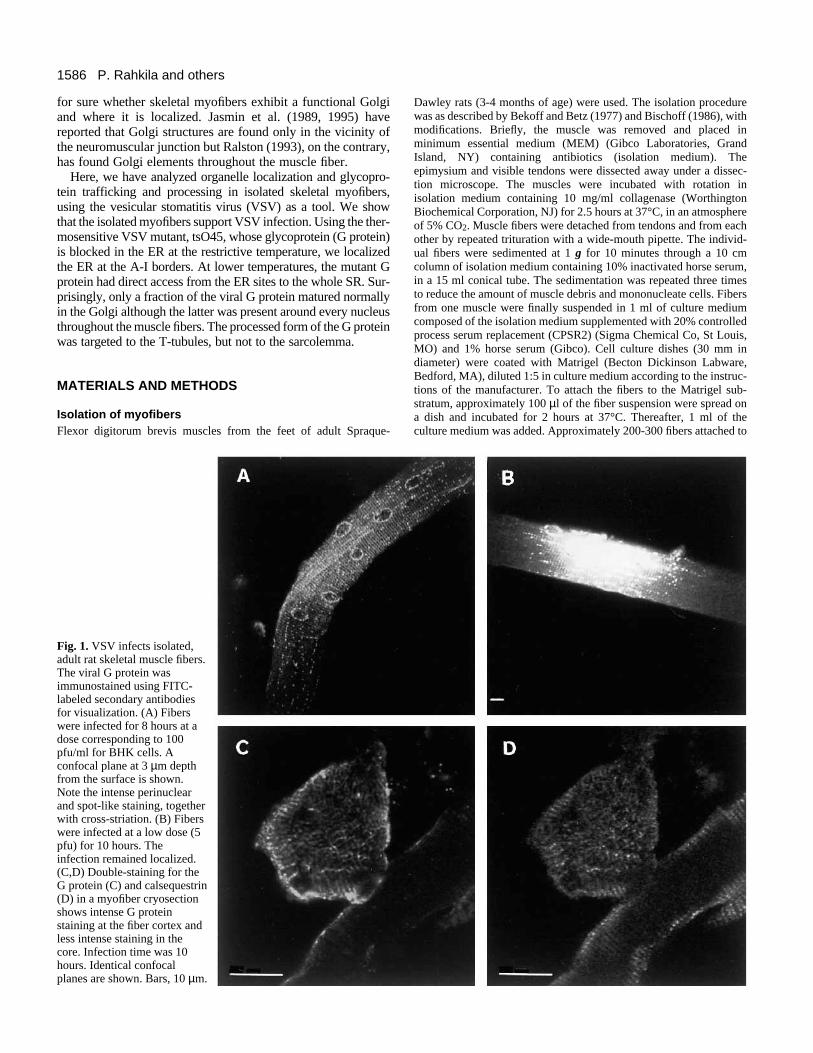
Fig. 1. VSV infects isolated,adult rat skeletal muscle fibers.The viral G protein wasimmunostained using FITC-labeled secondary antibodiesfor visualization. (A) Fiberswere infected for 8 hours at adose corresponding to 100pfu/ml for BHK cells. Aconfocal plane at 3 µm depthfrom the surface is shown.Note the intense perinuclearand spot-like staining, togetherwith cross-striation. (B) Fiberswere infected at a low dose (5pfu) for 10 hours. Theinfection remained localized.(C,D) Double-staining for theG protein (C) and calsequestrin(D) in a myofiber cryosectionshows intense G proteinstaining at the fiber cortex andless intense staining in thecore. Infection time was 10hours. Identical confocalplanes are shown. Bars, 10 µm.
Dawley rats (3-4 months of age) were used. The isolation procedurewas as described by Bekoff and Betz (1977) and Bischoff (1986), withmodifications. Briefly, the muscle was removed and placed inminimum essential medium (MEM) (Gibco Laboratories, GrandIsland, NY) containing antibiotics (isolation medium). Theepimysium and visible tendons were dissected away under a dissec-tion microscope. The muscles were incubated with rotation inisolation medium containing 10 mg/ml collagenase (WorthingtonBiochemical Corporation, NJ) for 2.5 hours at 37°C, in an atmosphereof 5% CO2. Muscle fibers were detached from tendons and from eachother by repeated trituration with a wide-mouth pipette. The individ-ual fibers were sedimented at 1 g for 10 minutes through a 10 cmcolumn of isolation medium containing 10% inactivated horse serum,in a 15 ml conical tube. The sedimentation was repeated three timesto reduce the amount of muscle debris and mononucleate cells. Fibersfrom one muscle were finally suspended in 1 ml of culture mediumcomposed of the isolation medium supplemented with 20% controlledprocess serum replacement (CPSR2) (Sigma Chemical Co, St Louis,MO) and 1% horse serum (Gibco). Cell culture dishes (30 mm indiameter) were coated with Matrigel (Becton Dickinson Labware,Bedford, MA), diluted 1:5 in culture medium according to the instruc-tions of the manufacturer. To attach the fibers to the Matrigel sub-stratum, approximately 100 µl of the fiber suspension were spread ona dish and incubated for 2 hours at 37°C. Thereafter, 1 ml of theculture medium was added. Approximately 200-300 fibers attached to
1587Protein trafficking in muscle fibers
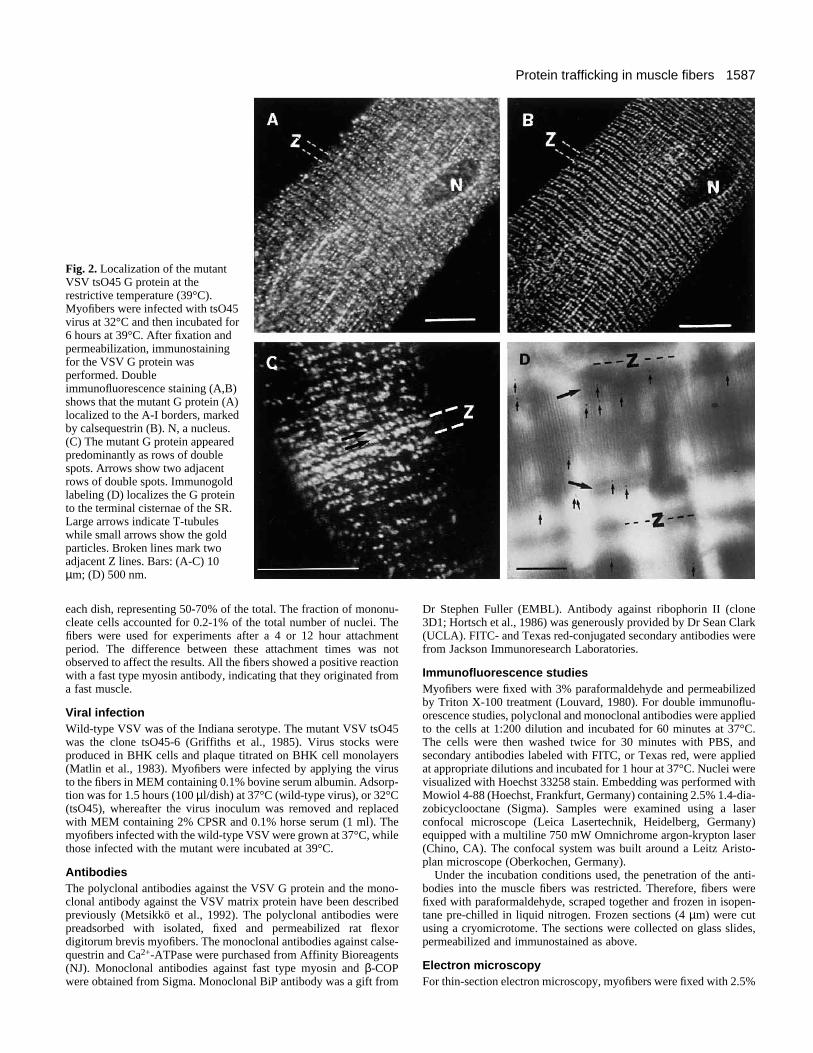
Fig. 2. Localization of the mutantVSV tsO45 G protein at therestrictive temperature (39°C).Myofibers were infected with tsO45virus at 32°C and then incubated for6 hours at 39°C. After fixation andpermeabilization, immunostainingfor the VSV G protein wasperformed. Doubleimmunofluorescence staining (A,B)shows that the mutant G protein (A)localized to the A-I borders, markedby calsequestrin (B). N, a nucleus.(C) The mutant G protein appearedpredominantly as rows of doublespots. Arrows show two adjacentrows of double spots. Immunogoldlabeling (D) localizes the G proteinto the terminal cisternae of the SR.Large arrows indicate T-tubuleswhile small arrows show the goldparticles. Broken lines mark twoadjacent Z lines. Bars: (A-C) 10µm; (D) 500 nm.
each dish, representing 50-70% of the total. The fraction of mononu-cleate cells accounted for 0.2-1% of the total number of nuclei. Thefibers were used for experiments after a 4 or 12 hour attachmentperiod. The difference between these attachment times was notobserved to affect the results. All the fibers showed a positive reactionwith a fast type myosin antibody, indicating that they originated froma fast muscle.
Viral infectionWild-type VSV was of the Indiana serotype. The mutant VSV tsO45was the clone tsO45-6 (Griffiths et al., 1985). Virus stocks wereproduced in BHK cells and plaque titrated on BHK cell monolayers(Matlin et al., 1983). Myofibers were infected by applying the virusto the fibers in MEM containing 0.1% bovine serum albumin. Adsorp-tion was for 1.5 hours (100 µl/dish) at 37°C (wild-type virus), or 32°C(tsO45), whereafter the virus inoculum was removed and replacedwith MEM containing 2% CPSR and 0.1% horse serum (1 ml). Themyofibers infected with the wild-type VSV were grown at 37°C, whilethose infected with the mutant were incubated at 39°C.
AntibodiesThe polyclonal antibodies against the VSV G protein and the mono-clonal antibody against the VSV matrix protein have been describedpreviously (Metsikkö et al., 1992). The polyclonal antibodies werepreadsorbed with isolated, fixed and permeabilized rat flexordigitorum brevis myofibers. The monoclonal antibodies against calse-questrin and Ca2+-ATPase were purchased from Affinity Bioreagents(NJ). Monoclonal antibodies against fast type myosin and β-COPwere obtained from Sigma. Monoclonal BiP antibody was a gift from
Dr Stephen Fuller (EMBL). Antibody against ribophorin II (clone3D1; Hortsch et al., 1986) was generously provided by Dr Sean Clark(UCLA). FITC- and Texas red-conjugated secondary antibodies werefrom Jackson Immunoresearch Laboratories.
Immunofluorescence studiesMyofibers were fixed with 3% paraformaldehyde and permeabilizedby Triton X-100 treatment (Louvard, 1980). For double immunoflu-orescence studies, polyclonal and monoclonal antibodies were appliedto the cells at 1:200 dilution and incubated for 60 minutes at 37°C.The cells were then washed twice for 30 minutes with PBS, andsecondary antibodies labeled with FITC, or Texas red, were appliedat appropriate dilutions and incubated for 1 hour at 37°C. Nuclei werevisualized with Hoechst 33258 stain. Embedding was performed withMowiol 4-88 (Hoechst, Frankfurt, Germany) containing 2.5% 1.4-dia-zobicyclooctane (Sigma). Samples were examined using a laserconfocal microscope (Leica Lasertechnik, Heidelberg, Germany)equipped with a multiline 750 mW Omnichrome argon-krypton laser(Chino, CA). The confocal system was built around a Leitz Aristo-plan microscope (Oberkochen, Germany).
Under the incubation conditions used, the penetration of the anti-bodies into the muscle fibers was restricted. Therefore, fibers werefixed with paraformaldehyde, scraped together and frozen in isopen-tane pre-chilled in liquid nitrogen. Frozen sections (4 µm) were cutusing a cryomicrotome. The sections were collected on glass slides,permeabilized and immunostained as above.
Electron microscopyFor thin-section electron microscopy, myofibers were fixed with 2.5%
1588 P. Rahkila and others
glutaraldehyde. Samples were postfixed with 4% OsO4, dehydratedand embedded in Epon. For immunoperoxidase labeling, fibers werefixed with 3% paraformaldehyde for 20 minutes, followed by 0.5%saponin treatment. The fibers were blocked with fetal calf serum andthen incubated with pre-adsorbed anti-VSV G protein antibodies,followed by incubation with secondary antibodies conjugated to per-oxidase. These incubations were carried out in the presence ofsaponin. The fibers were fixed with 2.5% glutaraldehyde before thediaminobenzidine reaction. Sections (100 nm) were examined withthe Philips 410 electron microscope. For immuno-gold labeling, fiberswere fixed for 30 minutes with 6% paraformaldehyde containing 0.1%glutaraldehyde and cryoprotected with 2.3 M sucrose for 10 minutesbefore cryosectioning. The sections (100 nm) were blocked with fetalcalf serum and then incubated with anti-VSV G protein antibodies,followed by incubation with Protein A conjugated to 10 nm goldparticles. Background was evaluated using noninfected fibers.
Metabolic labelingThe fibers were allowed to attach for 4 hours prior to infection. Pulse-chase labeling with [35S]methionine (Amersham Corp., Bucks,England) was performed at 10 hour post-infection. A 15 minute pulselabeling (50 µCi/ml) was performed, followed by various chase times.Overnight labeling (10 hours) was performed in MEM containing 5µM methionine, using 100 µCi/ml of [35S]methionine.
Other methodsIn pulse-chase experiments, fibers were solubilized with PBS con-taining 1% Triton X-100 and 1% deoxycholate. The viral G proteinwas immunoprecipitated using specific antibodies and immobilized
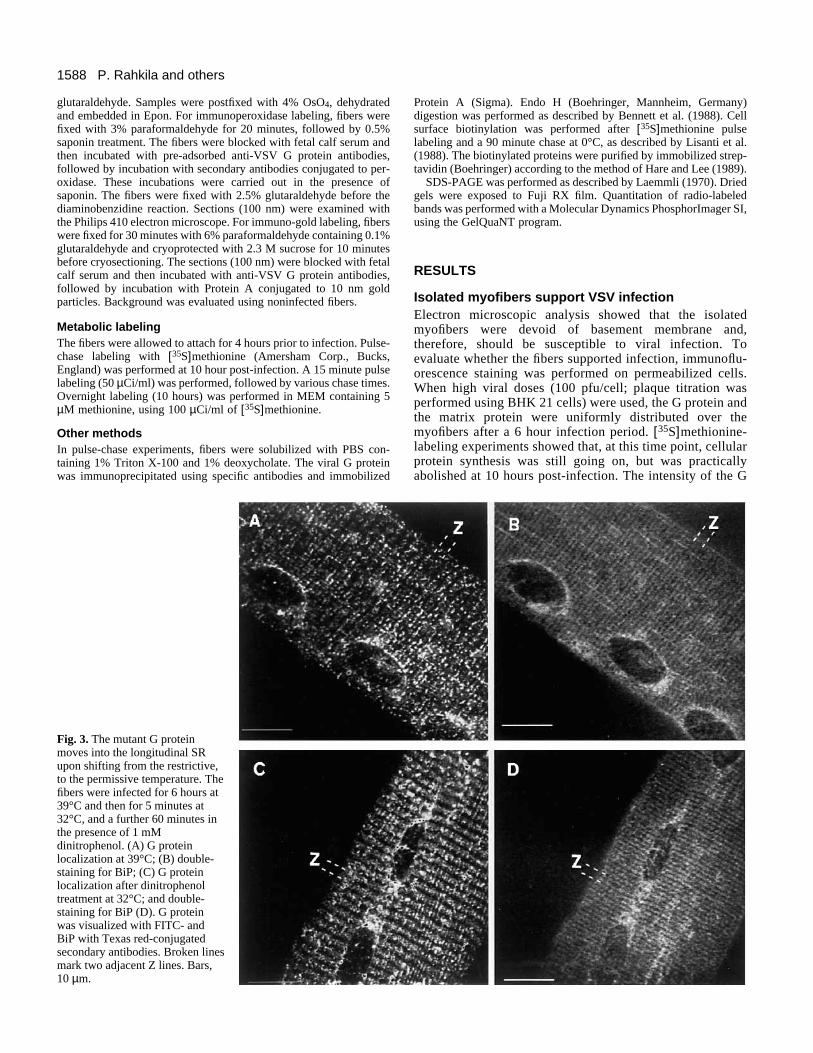
Fig. 3. The mutant G proteinmoves into the longitudinal SRupon shifting from the restrictive,to the permissive temperature. Thefibers were infected for 6 hours at39°C and then for 5 minutes at32°C, and a further 60 minutes inthe presence of 1 mMdinitrophenol. (A) G proteinlocalization at 39°C; (B) double-staining for BiP; (C) G proteinlocalization after dinitrophenoltreatment at 32°C; and double-staining for BiP (D). G proteinwas visualized with FITC- andBiP with Texas red-conjugatedsecondary antibodies. Broken linesmark two adjacent Z lines. Bars,10 µm.
Protein A (Sigma). Endo H (Boehringer, Mannheim, Germany)digestion was performed as described by Bennett et al. (1988). Cellsurface biotinylation was performed after [35S]methionine pulselabeling and a 90 minute chase at 0°C, as described by Lisanti et al.(1988). The biotinylated proteins were purified by immobilized strep-tavidin (Boehringer) according to the method of Hare and Lee (1989).
SDS-PAGE was performed as described by Laemmli (1970). Driedgels were exposed to Fuji RX film. Quantitation of radio-labeledbands was performed with a Molecular Dynamics PhosphorImager SI,using the GelQuaNT program.
RESULTS
Isolated myofibers support VSV infectionElectron microscopic analysis showed that the isolatedmyofibers were devoid of basement membrane and,therefore, should be susceptible to viral infection. Toevaluate whether the fibers supported infection, immunoflu-orescence staining was performed on permeabilized cells.When high viral doses (100 pfu/cell; plaque titration wasperformed using BHK 21 cells) were used, the G protein andthe matrix protein were uniformly distributed over themyofibers after a 6 hour infection period. [35S]methionine-labeling experiments showed that, at this time point, cellularprotein synthesis was still going on, but was practicallyabolished at 10 hours post-infection. The intensity of the G
1589Protein trafficking in muscle fibers
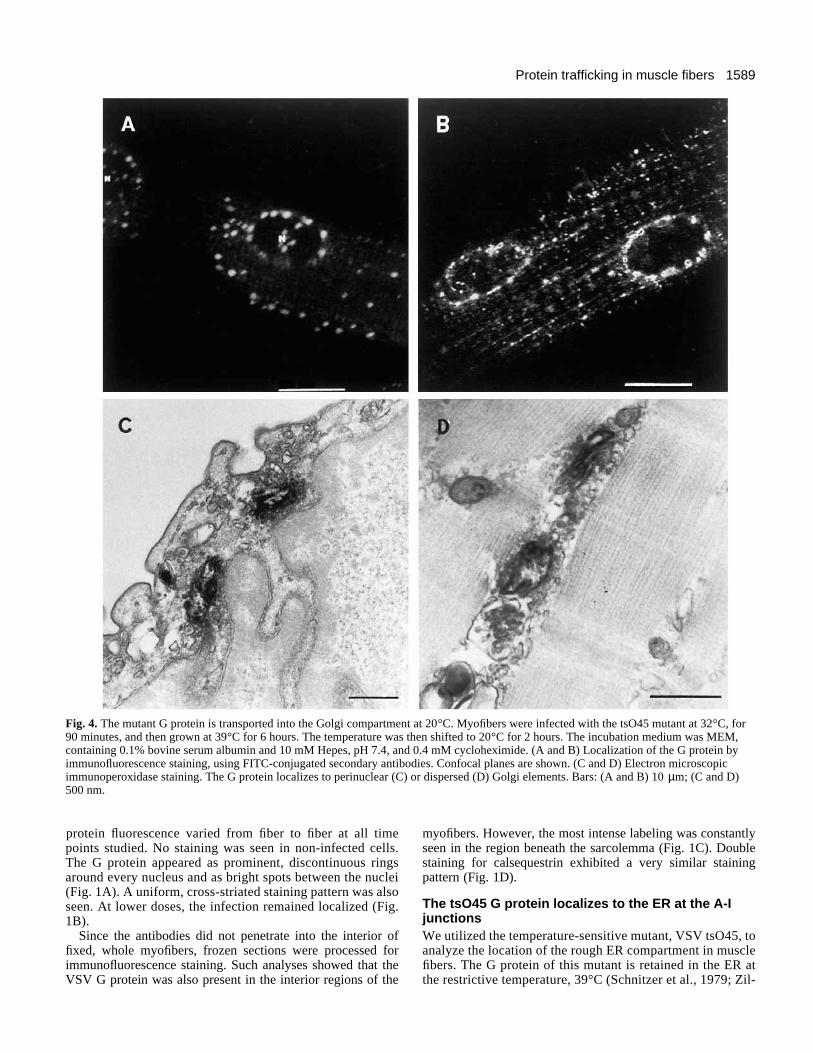
Fig. 4. The mutant G protein is transported into the Golgi compartment at 20°C. Myofibers were infected with the tsO45 mutant at 32°C, for90 minutes, and then grown at 39°C for 6 hours. The temperature was then shifted to 20°C for 2 hours. The incubation medium was MEM,containing 0.1% bovine serum albumin and 10 mM Hepes, pH 7.4, and 0.4 mM cycloheximide. (A and B) Localization of the G protein byimmunofluorescence staining, using FITC-conjugated secondary antibodies. Confocal planes are shown. (C and D) Electron microscopicimmunoperoxidase staining. The G protein localizes to perinuclear (C) or dispersed (D) Golgi elements. Bars: (A and B) 10 µm; (C and D)500 nm.
protein fluorescence varied from fiber to fiber at all timepoints studied. No staining was seen in non-infected cells.The G protein appeared as prominent, discontinuous ringsaround every nucleus and as bright spots between the nuclei(Fig. 1A). A uniform, cross-striated staining pattern was alsoseen. At lower doses, the infection remained localized (Fig.1B).
Since the antibodies did not penetrate into the interior offixed, whole myofibers, frozen sections were processed forimmunofluorescence staining. Such analyses showed that theVSV G protein was also present in the interior regions of the
myofibers. However, the most intense labeling was constantlyseen in the region beneath the sarcolemma (Fig. 1C). Doublestaining for calsequestrin exhibited a very similar stainingpattern (Fig. 1D).
The tsO45 G protein localizes to the ER at the A-IjunctionsWe utilized the temperature-sensitive mutant, VSV tsO45, toanalyze the location of the rough ER compartment in musclefibers. The G protein of this mutant is retained in the ER atthe restrictive temperature, 39°C (Schnitzer et al., 1979; Zil-
1590 P. Rahkila and others
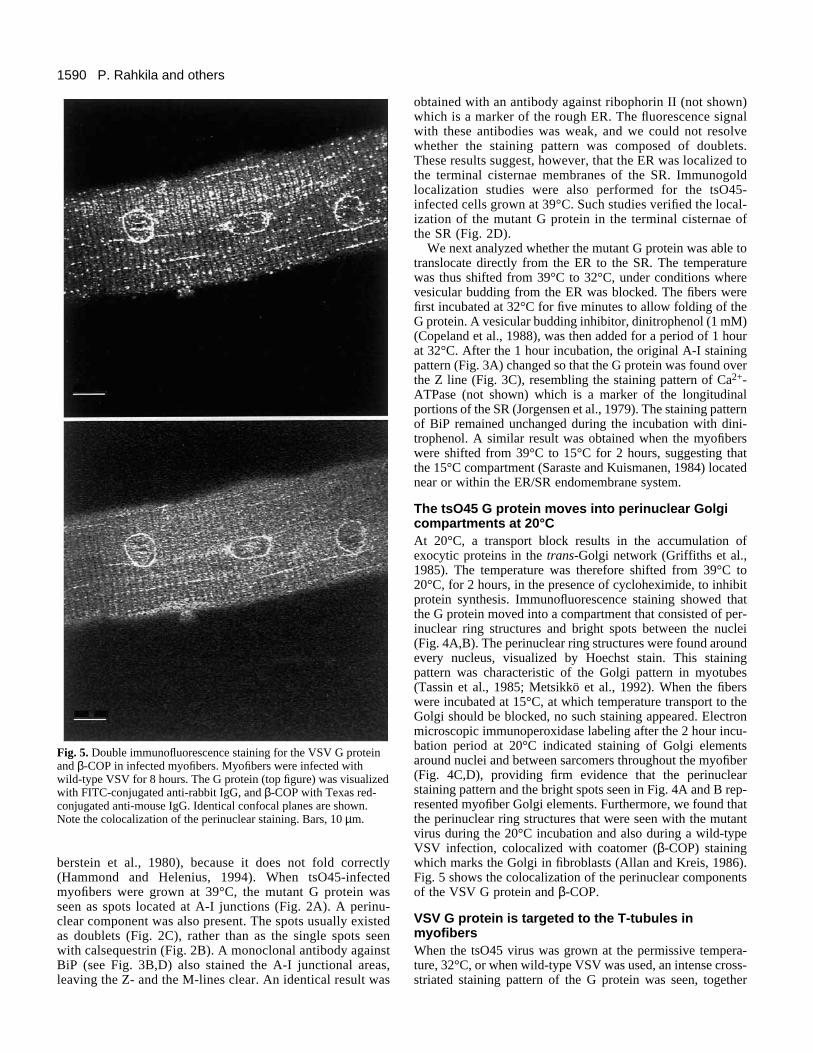
Fig. 5. Double immunofluorescence staining for the VSV G proteinand β-COP in infected myofibers. Myofibers were infected withwild-type VSV for 8 hours. The G protein (top figure) was visualizedwith FITC-conjugated anti-rabbit IgG, and β-COP with Texas red-conjugated anti-mouse IgG. Identical confocal planes are shown.Note the colocalization of the perinuclear staining. Bars, 10 µm.
berstein et al., 1980), because it does not fold correctly(Hammond and Helenius, 1994). When tsO45-infectedmyofibers were grown at 39°C, the mutant G protein wasseen as spots located at A-I junctions (Fig. 2A). A perinu-clear component was also present. The spots usually existedas doublets (Fig. 2C), rather than as the single spots seenwith calsequestrin (Fig. 2B). A monoclonal antibody againstBiP (see Fig. 3B,D) also stained the A-I junctional areas,leaving the Z- and the M-lines clear. An identical result was
obtained with an antibody against ribophorin II (not shown)which is a marker of the rough ER. The fluorescence signalwith these antibodies was weak, and we could not resolvewhether the staining pattern was composed of doublets.These results suggest, however, that the ER was localized tothe terminal cisternae membranes of the SR. Immunogoldlocalization studies were also performed for the tsO45-infected cells grown at 39°C. Such studies verified the local-ization of the mutant G protein in the terminal cisternae ofthe SR (Fig. 2D).
We next analyzed whether the mutant G protein was able totranslocate directly from the ER to the SR. The temperaturewas thus shifted from 39°C to 32°C, under conditions wherevesicular budding from the ER was blocked. The fibers werefirst incubated at 32°C for five minutes to allow folding of theG protein. A vesicular budding inhibitor, dinitrophenol (1 mM)(Copeland et al., 1988), was then added for a period of 1 hourat 32°C. After the 1 hour incubation, the original A-I stainingpattern (Fig. 3A) changed so that the G protein was found overthe Z line (Fig. 3C), resembling the staining pattern of Ca2+-ATPase (not shown) which is a marker of the longitudinalportions of the SR (Jorgensen et al., 1979). The staining patternof BiP remained unchanged during the incubation with dini-trophenol. A similar result was obtained when the myofiberswere shifted from 39°C to 15°C for 2 hours, suggesting thatthe 15°C compartment (Saraste and Kuismanen, 1984) locatednear or within the ER/SR endomembrane system.
The tsO45 G protein moves into perinuclear Golgicompartments at 20°CAt 20°C, a transport block results in the accumulation ofexocytic proteins in the trans-Golgi network (Griffiths et al.,1985). The temperature was therefore shifted from 39°C to20°C, for 2 hours, in the presence of cycloheximide, to inhibitprotein synthesis. Immunofluorescence staining showed thatthe G protein moved into a compartment that consisted of per-inuclear ring structures and bright spots between the nuclei(Fig. 4A,B). The perinuclear ring structures were found aroundevery nucleus, visualized by Hoechst stain. This stainingpattern was characteristic of the Golgi pattern in myotubes(Tassin et al., 1985; Metsikkö et al., 1992). When the fiberswere incubated at 15°C, at which temperature transport to theGolgi should be blocked, no such staining appeared. Electronmicroscopic immunoperoxidase labeling after the 2 hour incu-bation period at 20°C indicated staining of Golgi elementsaround nuclei and between sarcomers throughout the myofiber(Fig. 4C,D), providing firm evidence that the perinuclearstaining pattern and the bright spots seen in Fig. 4A and B rep-resented myofiber Golgi elements. Furthermore, we found thatthe perinuclear ring structures that were seen with the mutantvirus during the 20°C incubation and also during a wild-typeVSV infection, colocalized with coatomer (β-COP) stainingwhich marks the Golgi in fibroblasts (Allan and Kreis, 1986).Fig. 5 shows the colocalization of the perinuclear componentsof the VSV G protein and β-COP.
VSV G protein is targeted to the T-tubules inmyofibersWhen the tsO45 virus was grown at the permissive tempera-ture, 32°C, or when wild-type VSV was used, an intense cross-striated staining pattern of the G protein was seen, together
1591Protein trafficking in muscle fibers
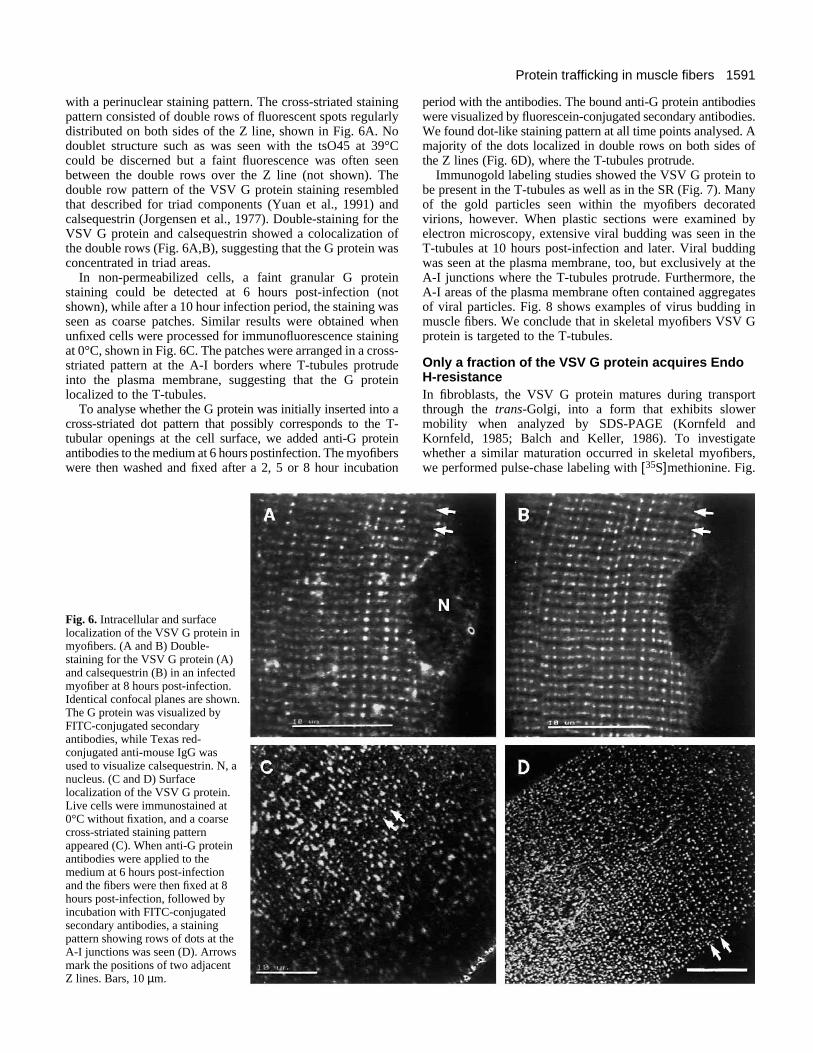
with a perinuclear staining pattern. The cross-striated stainingpattern consisted of double rows of fluorescent spots regularlydistributed on both sides of the Z line, shown in Fig. 6A. Nodoublet structure such as was seen with the tsO45 at 39°Ccould be discerned but a faint fluorescence was often seenbetween the double rows over the Z line (not shown). Thedouble row pattern of the VSV G protein staining resembledthat described for triad components (Yuan et al., 1991) andcalsequestrin (Jorgensen et al., 1977). Double-staining for theVSV G protein and calsequestrin showed a colocalization ofthe double rows (Fig. 6A,B), suggesting that the G protein wasconcentrated in triad areas.
In non-permeabilized cells, a faint granular G proteinstaining could be detected at 6 hours post-infection (notshown), while after a 10 hour infection period, the staining wasseen as coarse patches. Similar results were obtained whenunfixed cells were processed for immunofluorescence stainingat 0°C, shown in Fig. 6C. The patches were arranged in a cross-striated pattern at the A-I borders where T-tubules protrudeinto the plasma membrane, suggesting that the G proteinlocalized to the T-tubules.
To analyse whether the G protein was initially inserted into across-striated dot pattern that possibly corresponds to the T-tubular openings at the cell surface, we added anti-G proteinantibodies to the medium at 6 hours postinfection. The myofiberswere then washed and fixed after a 2, 5 or 8 hour incubation
Fig. 6. Intracellular and surfacelocalization of the VSV G protein inmyofibers. (A and B) Double-staining for the VSV G protein (A)and calsequestrin (B) in an infectedmyofiber at 8 hours post-infection.Identical confocal planes are shown.The G protein was visualized byFITC-conjugated secondaryantibodies, while Texas red-conjugated anti-mouse IgG wasused to visualize calsequestrin. N, anucleus. (C and D) Surfacelocalization of the VSV G protein.Live cells were immunostained at0°C without fixation, and a coarsecross-striated staining patternappeared (C). When anti-G proteinantibodies were applied to themedium at 6 hours post-infectionand the fibers were then fixed at 8hours post-infection, followed byincubation with FITC-conjugatedsecondary antibodies, a stainingpattern showing rows of dots at theA-I junctions was seen (D). Arrowsmark the positions of two adjacentZ lines. Bars, 10 µm.
period with the antibodies. The bound anti-G protein antibodieswere visualized by fluorescein-conjugated secondary antibodies.We found dot-like staining pattern at all time points analysed. Amajority of the dots localized in double rows on both sides ofthe Z lines (Fig. 6D), where the T-tubules protrude.
Immunogold labeling studies showed the VSV G protein tobe present in the T-tubules as well as in the SR (Fig. 7). Manyof the gold particles seen within the myofibers decoratedvirions, however. When plastic sections were examined byelectron microscopy, extensive viral budding was seen in theT-tubules at 10 hours post-infection and later. Viral buddingwas seen at the plasma membrane, too, but exclusively at theA-I junctions where the T-tubules protrude. Furthermore, theA-I areas of the plasma membrane often contained aggregatesof viral particles. Fig. 8 shows examples of virus budding inmuscle fibers. We conclude that in skeletal myofibers VSV Gprotein is targeted to the T-tubules.
Only a fraction of the VSV G protein acquires EndoH-resistanceIn fibroblasts, the VSV G protein matures during transportthrough the trans-Golgi, into a form that exhibits slowermobility when analyzed by SDS-PAGE (Kornfeld andKornfeld, 1985; Balch and Keller, 1986). To investigatewhether a similar maturation occurred in skeletal myofibers,we performed pulse-chase labeling with [35S]methionine. Fig.

1592 P. Rahkila and others
Fig. 7. Electron microscopic immunogold labeling for theVSV G protein. Myofibers were infected with VSV for 12hours, fixed and subjected to immunogold labeling asdescribed in Materials and Methods. A-I junctional areasand Z-lines show labeling while the A band contained abackground density of gold particles. Arrows indicate T-tubules. Broken lines indicate Z lines. Bars, 100 nm.
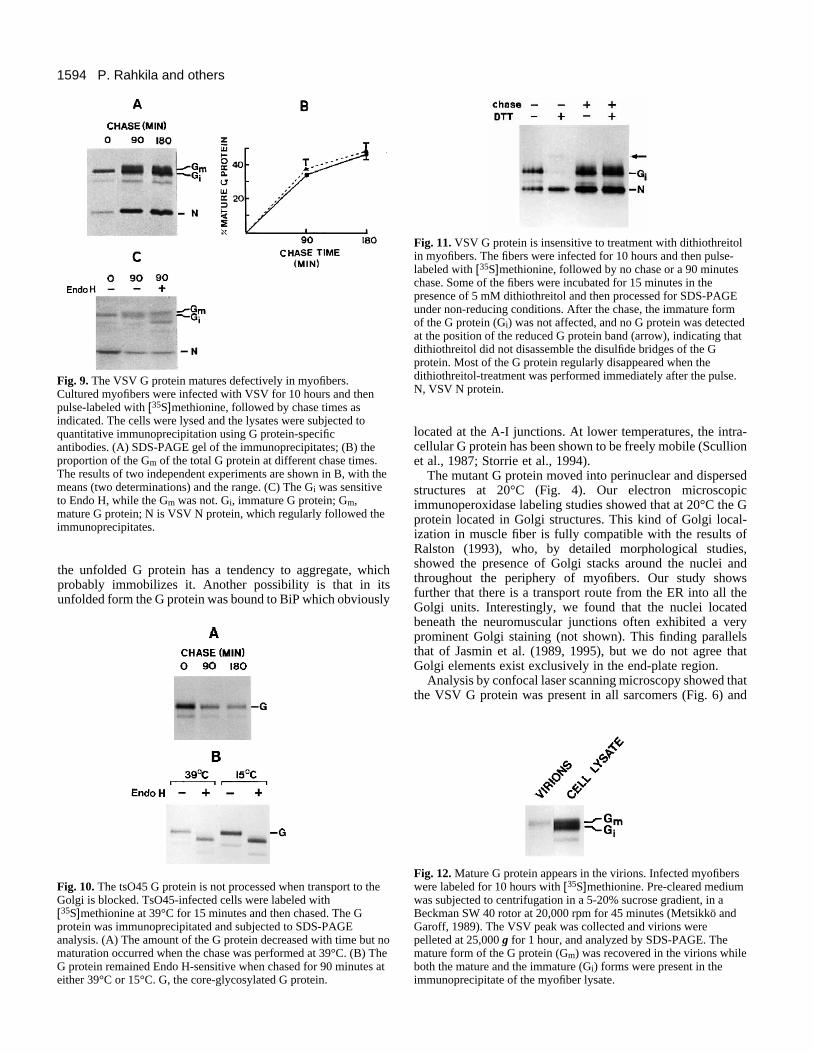
9A shows that the maturation of the wild-type G protein wasdefective in the myofibers. Only 46% of the total labeled Gprotein was transformed into a less mobile form during a 180minute chase period (Fig. 9B). Similar results were obtainedwhen the labeling was performed at 6 hours post-infection,when cellular protein synthesis was still going on (not shown).We constantly found that the G protein band that was not trans-formed to the less mobile form was processed into a slightlymore mobile form during the chase. Similar defective matura-tion of the G protein was observed with tsO45-infectedmyofibers grown at 32°C.
The conversion of the G protein from an Endo H-sensitiveform into an Endo H-resistant form is a Golgi-specific event(Davidson and Balch, 1993). The Endo H-sensitive andresistant forms can be distinguished by their mobilities in SDS-PAGE gels (Dunphy et al., 1985; Balch and Keller, 1986). Inmyofibers, the more mobile G protein form remained Endo H-sensitive during the chase, while the slower form becameresistant to the enzyme (Fig. 9C). It can be concluded that thefaster form was not processed in the medial Golgi. WhentsO45-infected cells were pulsed at 39°C and then chased at39°C, the amount of the G protein decreased, with a half timeof about 2 hours, suggesting degradation (Fig. 10A). When the
chase was performed either at 39°C, or 15°C, the G proteinremained totally Endo-H sensitive and no mobility shiftsoccurred (Fig. 10B).
The finding that the more mobile G protein remained EndoH-sensitive suggests that it was retained in the ER/SRendomembrane system, possibly due to a folding defect.Treatment of cells with dithiothreitol has been shown to be anindicator of disulfide bridge formation and folding (Braakmanet al., 1992; Hammond and Helenius, 1994). We thereforeanalyzed whether the G protein became resistant to dithiothre-itol during the chase. Fig. 11 shows that the G protein wastotally unaffected by this treatment. It thus seems that properfolding occurred.
Mature G protein appears in the viral particlesPlaque assays of the conditioning medium of the myofibers at10 hours post-infection, using wild-type VSV, indicated asmall amount of infective particles (9×105 pfu/ml). To analyzewhich form of the G protein appeared in the virions, weinfected myofibers for 6 hours and then labeled the fibers with[35S]methionine for 10 hours. The labeling medium wassubjected to sucrose gradient centrifugation to purify thevirions. SDS-PAGE showed that the virions contained the

1593Protein trafficking in muscle fibers
mature form of the G protein (Fig. 12). About 6% of the totalG protein was recovered in the form of virions. Cell surfacebiotinylation experiments, performed after pulse-labeling anda 180 minute chase period, also detected a faint band of themature form of the G protein (not shown).
DISCUSSION
We employed enveloped viruses to analyze protein transportin isolated, multinucleate muscle cells. The results show thatrat myofibers were readily infected with VSV after removal ofthe basement membrane. At low doses, the infection remainedlocalized, as occurs with myotubes (Metsikkö et al., 1992),while at high virus doses, the infection occupied the wholelength of the myofibers. We found that other viruses, such asSemliki Forest virus, and the Influenza virus, could also infectmyofibers (unpublished).
Fig. 8. Budding and accumulation of VSV particles inthe A-I junctional areas on the plasma membrane (A)and in T-tubules (B). Plastic sections (100 nm) wereanalysed by electron microscopy. Bars, 1 µm (insets,100 nm).
A
B
ER-retention signal-containing chaperones, such as BiP andcalnexin, have been reported to be present in the whole SR ofskeletal muscle fibers (Volpe et al., 1992; Villa et al., 1993).Where the rough ER is situated in the ER/SR endomembranenetwork has remained obscure. When we used the tsO45mutant at the restrictive temperature, A-I junctions werelabeled (Fig. 2). An antibody against BiP also labeled the A-Ijunctions, suggesting localization to the terminal cisternae butnot to the whole SR. On shifting to the permissive temperaturebut with budding from the ER blocked, we saw movement ofthe G protein to the longitudinal portions of the SR (Fig. 3).This finding is compatible with the proposal that ER and SRmembranes are continuous (Volpe et al. 1992). However, ourresults show that the ER and the SR are not identical. We donot know why the mutant G protein remained immobile in themyofibers at 39°C, since the mutant G protein rapidly movedto the smooth ER at 39°C in UT-1 cells (Bergman and Fusco,1990). However, Hammond and Helenius (1994) showed that
1594 P. Rahkila and others
Fig. 9. The VSV G protein matures defectively in myofibers.Cultured myofibers were infected with VSV for 10 hours and thenpulse-labeled with [35S]methionine, followed by chase times asindicated. The cells were lysed and the lysates were subjected toquantitative immunoprecipitation using G protein-specificantibodies. (A) SDS-PAGE gel of the immunoprecipitates; (B) theproportion of the Gm of the total G protein at different chase times.The results of two independent experiments are shown in B, with themeans (two determinations) and the range. (C) The Gi was sensitiveto Endo H, while the Gm was not. Gi, immature G protein; Gm,mature G protein; N is VSV N protein, which regularly followed theimmunoprecipitates.
Fig. 11. VSV G protein is insensitive to treatment with dithiothreitolin myofibers. The fibers were infected for 10 hours and then pulse-labeled with [35S]methionine, followed by no chase or a 90 minuteschase. Some of the fibers were incubated for 15 minutes in thepresence of 5 mM dithiothreitol and then processed for SDS-PAGEunder non-reducing conditions. After the chase, the immature formof the G protein (Gi) was not affected, and no G protein was detectedat the position of the reduced G protein band (arrow), indicating thatdithiothreitol did not disassemble the disulfide bridges of the Gprotein. Most of the G protein regularly disappeared when thedithiothreitol-treatment was performed immediately after the pulse.N, VSV N protein.
the unfolded G protein has a tendency to aggregate, whichprobably immobilizes it. Another possibility is that in itsunfolded form the G protein was bound to BiP which obviously
Fig. 10. The tsO45 G protein is not processed when transport to theGolgi is blocked. TsO45-infected cells were labeled with[35S]methionine at 39°C for 15 minutes and then chased. The Gprotein was immunoprecipitated and subjected to SDS-PAGEanalysis. (A) The amount of the G protein decreased with time but nomaturation occurred when the chase was performed at 39°C. (B) TheG protein remained Endo H-sensitive when chased for 90 minutes ateither 39°C or 15°C. G, the core-glycosylated G protein.
located at the A-I junctions. At lower temperatures, the intra-cellular G protein has been shown to be freely mobile (Scullionet al., 1987; Storrie et al., 1994).
The mutant G protein moved into perinuclear and dispersedstructures at 20°C (Fig. 4). Our electron microscopicimmunoperoxidase labeling studies showed that at 20°C the Gprotein located in Golgi structures. This kind of Golgi local-ization in muscle fiber is fully compatible with the results ofRalston (1993), who, by detailed morphological studies,showed the presence of Golgi stacks around the nuclei andthroughout the periphery of myofibers. Our study showsfurther that there is a transport route from the ER into all theGolgi units. Interestingly, we found that the nuclei locatedbeneath the neuromuscular junctions often exhibited a veryprominent Golgi staining (not shown). This finding parallelsthat of Jasmin et al. (1989, 1995), but we do not agree thatGolgi elements exist exclusively in the end-plate region.
Analysis by confocal laser scanning microscopy showed thatthe VSV G protein was present in all sarcomers (Fig. 6) and
Fig. 12. Mature G protein appears in the virions. Infected myofiberswere labeled for 10 hours with [35S]methionine. Pre-cleared mediumwas subjected to centrifugation in a 5-20% sucrose gradient, in aBeckman SW 40 rotor at 20,000 rpm for 45 minutes (Metsikkö andGaroff, 1989). The VSV peak was collected and virions werepelleted at 25,000 g for 1 hour, and analyzed by SDS-PAGE. Themature form of the G protein (Gm) was recovered in the virions whileboth the mature and the immature (Gi) forms were present in theimmunoprecipitate of the myofiber lysate.

1595Protein trafficking in muscle fibers
Fig. 13. Plausible transport routes of the VSV G protein in skeletalmyofibers. The model postulates that the ER and the SR membranesare continuous. The immature G protein (open rectangle) is assumedto diffuse directly from the ER to the SR. A transport route via theGolgi to the SR remains possible. A fraction of the G protein travelsthrough the Golgi and undergoes processing (filled rectangle). Theprocessed form is transported to the T-tubules but not to thesarcolemma.
immunogold labeling located the G protein to the T-tubules, aswell as to the ER/SR endomembranes (Fig. 7). The G proteindid not show defects in disulfide bridging (Fig. 11), suggest-ing that folding occurred. Therefore, the G protein was notsimply retained in the ER due to defective folding. A plausibleexplanation for the SR location of the G protein is that afraction of it translocated from the ER to the SR.
A dual maturation of the VSV G protein was observed inthe isolated skeletal myofibers (Fig. 9). A portion of the Gprotein matured normally. Therefore, mannosidase II activityis present in muscle fibers, although the enzyme is undetectedby a monoclonal antibody that is reactive with myotubes(Ralston, 1993). Another portion of the virus glycoproteinremained Endo H-sensitive, indicating that processing in themedial Golgi did not occur (Davidson and Balch, 1993). Onepossibility is that the VSV G protein was produced in such highamounts that the folding and transport machinery becamesaturated. This seems unlikely, however, since similar dualmaturation occurred during early infection and the immatureG protein folded correctly (Fig. 11). We suggest that half ofthe G protein passed through a muscle-specific pathway thattranslocates proteins from the ER to the SR. Our laboratoryrecently demonstrated that L6 myoblast-derived myotubesexhibit a similar dual processing of the VSV G protein (Kel-lokumpu et al. 1995). Both the Endo H-resistant and -sensitiveforms were transported to the Golgi. In chick primarymyotubes, calsequestrin, a resident protein of the terminalcisternae of the SR, seems to be transported to the cis-Golgiand back to the SR (Thomas et al., 1989). In contrast, anotherSR component, the Ca2+-ATPase, never exits the ER/SRsystem (Karin and Settle, 1992). It remains to be resolvedwhether or not the fraction of the G protein remaining EndoH-sensitive was transported to the Golgi and possibly back tothe ER/SR endomembranes in the skeletal myofibers.
We could show that virions collected from the mediumpossessed the mature form of the G protein. Our electronmicroscopic data strongly suggest that these virions originatedfrom the T-tubules and their orifices. Furthermore, immuno-fluorescence staining of non-permeabilized cells revealed rowsof dots at the A-I junctions, leaving other areas of the plasmamembrane clear. Similar results were obtained when primaryantibodies were added to the medium during the viral infection.Therefore, we conclude that the mature form of the G proteinwas targeted to the T-tubules and was absent at the sar-colemma. It has been shown earlier that T-tubular proteinsdiffer from those of the plasma membrane throughout muscledevelopment (Yuan et al., 1990; Flucher et al., 1991). A 50kDa sarcolemmal protein was not present in T-tubules, and a28 kDa T-tubular protein was not found at the sarcolemma(Jorgensen et al., 1990). While our results indicate a pathwayfrom the Golgi to the T-tubules, it remains unresolved how sar-colemmal proteins are targeted.
In conclusion, our results locate the ER at the A-I junctionalarea of the SR membrane system and confirm the view(Ralston, 1993) that Golgi elements are found throughout themyofibers. Furthermore, they show that skeletal myofibersprocess a fraction of the VSV G protein and target theprocessed form to the T-tubulus system. The sarcolemmareceived little, if any, G protein. A large fraction of the Gprotein remained Endo H-sensitive, suggesting that it was nottransported beyond the cis-Golgi and that it was retained in theER/SR endomembrane system. Fig. 13 depicts the hypotheti-cal transport routes for the VSV G protein in muscle fibers.
We thank Ms Sari Seinijoki for expert assistance, and Dr KaiSimons (EMBL) for valuable comments and critically reading themanuscript. This study was supported by The Ministry of Education,Finland.
REFERENCES
Allan, V. J. and Kreis, T. E. (1986). A microtubule-binding protein associatedwith membranes of the Golgi apparatus. J. Cell Biol. 103, 2229-2239.
Balch, W. E. and Keller, D. S. (1986). ATP-coupled transport of vesicularstomatitis virus G protein. J. Biol. Chem. 261, 14690-14696.
Barr, E. and Leiden, J. M. (1991). Systemic delivery of recombinant proteinsby genetically modified myoblasts. Science 254, 1507-1509.
Bekoff, A. and Betz, W. (1977). Properties of isolated adult rat muscle fibersmaintained in tissue culture. J. Physiol. 271, 537-547.
Bennett, M. K., Wandinger-Ness, A. and Simons, K. (1988). Release ofputative exocytotic transport vesicles from perforated MDCK cells. EMBO J.7, 4075-4085.
Bergmann, J. E. and Fusco, P. J. (1990). The G protein of vesicular stomatitisvirus has free access into and egress from the smooth endoplasmic reticulumof UT-1 cells. J. Cell Biol. 110, 625-635.
Bischoff, R. (1986). A satellite cell mitogen from crushed adult muscle. Dev.Biol. 115, 140-147.
Braakman, I., Helenius, J. and Helenius, A. (1992). Manipulating disulfidebond formation and protein folding in the endoplasmic reticulum. EMBO J.11, 1717-1722.
Copeland, C. S., Zimmer, K.-P., Wagner, K. R., Healey, G. A., Mellman, I.and Helenius, A. (1988). Folding, trimerization, and transport are sequentialevents in the biogenesis of influenza virus hemagglutinin. Cell 53, 197-209.
Dai, Y., Roman M., Naviaux, R. K. and Verma, I. M. (1992). Gene therapyvia primary myoblasts: Long-term expression of factor IX protein followingtransplantation in vivo. Proc. Nat. Acad. Sci. USA 89, 10892-10895.
Davidson, H. W. and Balch, W. E. (1993). Differential inhibition of multiplevesicular transport steps between the endoplasmic reticulum and trans Golginetwork. J. Biol. Chem. 268, 4216-4226.

1596 P. Rahkila and others
Dhawan, J., Pan, L. C., Pavlath, G. K., Travis, M. A., Lanctot, A. M. andBlau, H. M. (1991). Systemic delivery of human growth hormone byinjection of genetically engineered myoblasts. Science 254, 1509-1512.
Dunphy, W. G., Brands, R. and Rothman, J. E. (1985). Attachment ofterminal N-acetylglucosamine to asparagine-linked oligosaccharides occursin central cisterna of the Golgi stack. Cell 40, 463-472.
Flucher, B. E., Terasaki, M., Chin, H., Beeler, T. J. and Daniels, M. P.(1991). Biogenesis of transverse tubules in skeletal muscle in vitro. Dev.Biol. 145, 77-90.
Flucher, B. E. (1992). Structural analysis of muscle development: Transversetubules, sarcoplasmic reticulum, and the triad. Dev. Biol. 154, 245-260.
Griffiths, G., Pfeiffer, S., Simons, K. and Matlin, K. (1985). Exit of newlysynthesised membrane proteins from the trans cisterna of the Golgi complexto the plasma membrane. J. Cell Biol. 101, 949-964.
Hammond, C. and Helenius, A. (1994). Quality control in the secretorypathway: Retention of a misfolded viral membrane glycoprotein involvescycling between the ER, intermediate compartment, and Golgi apparatus. J.Cell Biol. 126, 41-52.
Hare, J. F. and Lee, E. (1989). Metabolic behavior of cell surface biotinylatedproteins. Biochemistry 28, 574-580.
Hortsch, M., Avossa, D. and Meyer, D. I. (1986). Characterization ofsecretory protein translocation: Ribosome-membrane interaction inendoplasmic reticulum. J. Cell Biol. 103, 241-253.
Jasmin, B. J., Cartaud, J., Bornens, M. and Changeux, J. P. (1989). Golgiapparatus in chick skeletal muscle: changes in its distribution during endplate development and denervation. Proc. Nat. Acad. Sci. USA 86, 7218-7222.
Jasmin, B. J., Antony, C., Changeux, J.-P. and Cartaud, J. (1995). Nerve-dependent plasticity of the Golgi complex in skeletal muscle fibres:compartmentalization within the subneural sarcoplasm. Eur. J. Neurosci. 7,470-479.
Jorgensen, A. O., Kalnins, V. I., Zubrzycka, E. and MacLennan, D. H.(1977). Assembly of the sarcoplasmic reticulum. J. Cell Biol. 74, 287-298.
Jorgensen, A. O., Kalnins, V. and MacLennan, D. H. (1979). Localization ofsarcoplasmic reticulum proteins in rat skeletal muscle byimmunofluorescence. J. Cell Biol. 80, 372-384.
Jorgensen, A. O., Arnold, W., Shen, A. C.-Y., Yuan, S., Gaver, M. andCampbell, K. P. (1990). Identification of novel proteins unique to eithertransverse tubules (ts28) or the sarcolemma (SL 50) in rabbit skeletal muscle.J. Cell Biol. 110, 1173-1185.
Karin, N. J. and Settle, V. J. (1992). The sarcoplasmic reticulum Ca2+-ATPase, SERCA1A, contains endoplasmic reticulum targeting information.Biochem. Biophys. Res. Commun. 186, 219-227.
Kellokumpu, S., Sormunen, R., Väänänen, K. and Metsikkö, K. (1995).Defective maturation of a viral glycoprotein and partial loss of the Golgistack structure during in vitro myogenesis. Exp. Cell. Res. 220, 101-111.
Kornfeld, R. and Kornfeld, S. (1985). Assembly of asparagine-linkedoligosaccharides. Annu. Rev. Biochem. 54, 631-664.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly ofbacteriophage T4. Nature 227, 680-685.
Lisanti, M. P., Sargiacomo, A., Greave, L., Saltiel, A. R. and Rodriguez-Boulan, E. (1988). Polarized apical distribution of glycosylphosphatidylinositol-anchored proteins in a renal epithelial cell line. Proc.Nat. Acad. Sci. USA 85, 9557-9561.
Louvard, D. (1980). Apical membrane aminopeptidase appears at sites of cell-cell contact in cultured epithelial cells. Proc. Nat. Acad. Sci. USA 77, 4132-4136.
Matlin, K. S., Bainton, D. F., Pesonen, M., Louvard, D., Genty, N. andSimons, K. (1983). Transepithelial transport of a viral membraneglycoprotein implanted into the apical membrane of MDCK cells. I.Morphological evidence. J. Cell Biol. 97, 627-637.
Metsikkö, K. and Garoff, H. (1989). Role of heterologous and homologousglycoproteins in phenotypic mixing between Sendai virus and vesicularstomatitis virus. J. Virol. 63, 5111-5118.
Metsikkö, K., Hentunen, T. and Väänänen, K. (1992). Local expression andexocytosis of viral glycoproteins in multinucleated muscle cells. J. Cell Biol.117, 987-995.
Raichman, M., Panzeri, M. C., Clementi, E., Papazafiri, P., Eckley, M.,Clegg, D. O., Villa, A. and Meldolesi, J. (1995). Differential localizationand functional role of calsequestrin in growing and differentiated myoblasts.J. Cell Biol. 128, 341-354.
Ralston, E. (1993). Changes in architecture of the Golgi complex and othersubcellular organelles during myogenesis. J. Cell Biol. 120, 399 409.
Rotundo, R., Thomas, K., Porter-Jordan, K., Benson, R. J. J., Fernandez-Valle, C. and Fine, R. E. (1989). Intracellular transport, sorting, andturnover of acetylcholinesterase. J. Biol. Chem. 264, 3146-3152.
Saraste, J. and Kuismanen E. (1984). Pre- and post-Golgi vacuoles operate inthe transport of Semliki Forest virus membrane glycoproteins to the cellsurface. Cell 38, 535-549.
Schnitzer, T. J., Dickson, C. and Weiss, R. A. (1979). Morphological andbiochemical characterization of viral particles produced by the tsO45 mutantof vesicular stomatitis virus at restrictive temperature. J. Virol. 29, 185-195.
Scullion, B. F., Hou, Y., Puddington, L., Rose, J. K. and Jacobson, K.(1987). Effects of mutations in three domains of the vesicular stomatitis viralglycoprotein on its lateral diffusion in the plasma membrane. J. Cell Biol.105, 69-75.
Storrie, B., Pepperkok, R., Stelzer, E. H. K. and Kreis, T. E. (1994). Theintracellular mobility of a viral membrane glycoprotein measured byconfocal microscope fluorescence recovery after photobleaching. J. Cell Sci.107, 1309-1319.
Tassin, A. M., Paintrand, M., Berger, E. G. and Bornens, M. (1985). TheGolgi apparatus remains associated with microtubule organizing centersduring myogenesis. J. Cell Biol. 101, 630-638.
Thomas, K., Navarro, J., Benson, R. J. J., Campbell, K. P., Rotundo, R. L.and Fine, R. E. (1989). Newly synthesised calsequestrin, destined for thesarcoplasmic reticulum, is contained in early/intermediate Golgi-derivedclathrin-coated vesicles. J. Biol. Chem. 264, 3140-3145.
Villa, A., Podini, P., Nori, A., Panzeri, M. C., Martini, A., Meldolesi, J. andVolpe P. (1993). The endoplasmic reticulum-sarcoplasmic reticulumconnection. II. Postnatal differentiation of the sarcoplasmic reticulum inskeletal muscle fibers. Exp. Cell Res. 209, 140-148.
Volpe, P., Villa, A., Podini, P., Martini, A., Nori, A., Panzeri, M. C. andMeldolesi, J. (1992). The endoplasmic reticulum-sarcoplasmic reticulumconnection: distribution of endoplasmic reticulum markers in thesarcoplasmic reticulum of skeletal muscle fibers. Proc. Nat. Acad. Sci. USA89, 6142-6146.
Yao, S.-N. and Kurachi, K. (1992). Expression of human factor IX in miceafter injection of genetically modified myoblasts. Proc. Nat. Acad. Sci. USA89, 3357-3361.
Yuan, S., Arnold, W. and Jorgensen, A. O. (1990). Biogenesis of transversetubules: immunocytochemical localization of a transverse tubular protein(TS28) and a sarcolemmal protein (SL50) in rabbit skeletal muscle in situ. J.Cell Biol. 110, 1187-1198.
Yuan, S. H., Arnold, W. and Jorgensen, A. O. (1991). Biogenesis oftransverse tubules and triads: immunolocalization of the 1.4,-dihydropyridine receptor, TS28, and the ryanodine receptor in rabbit skeletalmuscle developing in situ. J. Cell Biol. 112, 289-301.
Zilberstein A., Snider, M. D., Porter, M. and Lodish, H. F. (1980). Mutantsof vesicular stomatitis virus blocked at different stages of maturation of theviral glycoprotein. Cell 21, 417-427.
(Received 22 November 1995 - Accepted 5 March 1996)





























