Embed Size (px)
Citation preview

JOURNAL OF EXPERIMENTAL ZOOLOGY 283:675–688 (1999)
© 1999 WILEY-LISS, INC.
Transport of Organic Molecules byReptilian Nephrons
W.H. DANTZLER*Department of Physiology, College of Medicine, University of Arizona,Tucson, Arizona 85724-5051
ABSTRACT This paper considers transport of glucose, amino acids, organic anions, urate, andorganic cations by reptilian renal tubules. Glucose undergoes net reabsorption involving entryacross the luminal membrane by a Na+-dependent, carrier-mediated process. However, entry mayoccur down an electrochemical gradient and exit at the basolateral membrane, against an electro-chemical gradient. Amino acids may undergo both net reabsorption and net secretion. Taurinereabsorption involves entry across the luminal membrane against an electrochemical gradient viatwo independent electrogenic, Na+ and Cl–-dependent co-transport systems. Organic anions (e.g.,PAH) undergo net secretion. This process involves entry into the cells at the basolateral mem-brane against an electrochemical gradient via a tertiary active process, the final step involvingcountertransport for α-ketoglutarate. Exit at the luminal membrane is down an electrochemicalgradient, probably via anion exchange or carrier-mediated diffusion. Urate undergoes net secre-tion. This process involves transport into the cells against an electrochemical gradient via anunknown, Na+-independent, K+-sensitive, mechanism that may involve anion exchange and exitacross the luminal membrane down an electrochemical gradient via what appears to be simplediffusion. Some organic cations, such as tetraethylammonium (TEA), undergo net secretion andsome, such as N1-methylnicotinamide (NMN), undergo net reabsorption. Net TEA secretion in-volves entry across the basolateral membrane, down an electrochemical gradient via carrier-medi-ated diffusion or organic anion exchange, and exit at the luminal membrane against anelectrochemical gradient via countertransport for H+. Net NMN reabsorption involves entry acrossthe luminal membrane, down an electrochemical gradient via Na+-dependent, carrier-mediateddiffusion, with exit across the basolateral membrane against an electrochemical gradient via anunknown process that probably involves countertransport for an organic cation. J. Exp. Zool.283:675–688, 1999. © 1999 Wiley-Liss, Inc.
Reptilian nephrons transport glucose, amino ac-ids, organic anions (or acids), and organic cations(or bases). Although the nephrons undoubtedlytransport other organic substances as well, theseare the only ones about which significant trans-port information is available. Moreover, almost allthe information on these transport processescomes from work with snakes. In the present dis-cussion, I review the current state of knowledgefor each of these transport processes and then dis-cuss recent work from our laboratory. The recentwork relates almost entirely to the transport oforganic anions and cations. I provide basic descrip-tive models for these transport processes, butmany mechanistic and most regulatory aspectshave yet to be elucidated.
GLUCOSEIn reptiles, as in other vertebrates, glucose is
freely filtered at the glomerulus. In view of itsapparent absence from ureteral urine, the renal
tubules entirely reabsorb glucose. The reab-sorptive mechanism has been studied with iso-lated perfused proximal tubules and brush-borderand basolateral membrane vesicles (BBMV andBLMV) from garter snake kidneys (Thamnophissp.) (Barfuss and Dantzler, ’76; Benyajati andDantzler, ’88; Benyajati and Bay, ’90). Studies ofboth types indicate that transport across the lu-minal membrane occurs via a saturable, Na+-de-pendent, phlorizin-sensitive process. However,whereas the BBMV studies indicate that, as inmammals, transport across this membrane isagainst an electrochemical gradient (Na+-depen-dent, electrogenic development of a transientintravesicular concentration above equilibrium)(Benyajati and Dantzler, ’88), the isolated tubule
Grant sponsor: National Science Foundation; Grant number: IBN-9513892.
*Correspondence to: William H. Dantzler, Department of Physiol-ogy, College of Medicine, University of Arizona, Tucson, AZ 85724-5051. E-mail: [email protected]

676 W.H. DANTZLER
studies indicate that transport is down an elec-trochemical gradient (intracellular concentrationduring transport, always below that in the tubulelumen and bath) (Barfuss and Dantzler, ’76). It isunclear whether these differences reflect a simpletechnical difference between preparations or atrue difference between what occurs in an isolatedmembrane and an intact tubule. Transport againsta concentration gradient at the luminal membranemay have been obscured in the intact tubulepreparation because the intracellular concentra-tion was measured only when there was sufficientglucose in the lumen to saturate the transport sys-tem (Barfuss and Dantzler, ’76). At present, it isnot possible to decide whether transport of glu-cose into the cells involves secondary active trans-port or facilitated diffusion.
Transport of glucose out of the cells across thebasolateral membrane during the transepithelialreabsorptive process appears to involve a carrier-mediated process against a concentration gradi-ent (Barfuss and Dantzler, ’76). This concept issupported by three observations with isolated per-fused tubules: (1) When glucose entry across theluminal membrane is blocked with phlorizin, theintracellular concentration of glucose falls and,thus, the concentration gradient across the baso-lateral membrane increases. (2) When tubulesspontaneously stop transporting glucose, the in-tracellular concentration rises to equal the con-centration in the bathing medium, eliminating theconcentration gradient across the basolateralmembrane. (3) The apparent passive permeabil-ity of the basolateral membrane to glucose (~4.5× 10–6 cm s–1) is too low to permit transport acrossthis membrane by simple passive diffusion at theobserved rate. Preliminary data with BLMV fromsnake tubules also support the concept of a Na+-independent transport system that may occuragainst an electrochemical gradient at this mem-brane (Benyajati and Bay, ’90).
In summary, filtered glucose is completely re-absorbed by reptilian renal proximal tubules.Studies with snake kidneys indicate that trans-epithelial reabsorption involves entry into the cellsacross the luminal membrane via a Na+-depen-dent, carrier-mediated process and exit across thebasolateral membrane via a Na+-independent, car-rier-mediated process. However, whereas all dataare compatible with transport out of the cells atthe basolateral side against an electrochemicalgradient, data from isolated tubules and brush-border membrane vesicles provide conflicting evi-dence about whether entry into the cells across
the luminal membrane is down or up an electro-chemical gradient. No information is yet availableabout the molecular characteristics of these trans-port steps.
AMINO ACIDSTransport has been evaluated in reptiles by
whole-kidney clearance and BBMV and BLMVstudies. In alligators (Alligator mississippiensis),clearance data indicate that all filtered amino ac-ids undergo net reabsorption that is about 99%complete (Hernandez and Coulson, ’67). However,in marine and terrestrial snakes (olive sea snake,Aipysurus laevis; garter snake, Thamnophissirtalis), such studies indicate that both net reab-sorption and net secretion of taurine, other β-aminoacids (β-alanine and β-aminoisobutyric acid), andan endogenous analog of the β-amino acids (L-cys-teic acid) occur (Benyajati and Dantzler, ’86a,b).However, the physiological significance of net se-cretion of these amino acids for snakes is unknown.Moreover, although other endogenous amino acidsundergo net reabsorption in these snakes, the ex-tent of the reabsorption is variable and often lessthan that observed in other animals (Benyajati andDantzler, ’86a,b). As an example, in both gartersnakes and sea snakes only about 80% of the fil-tered histidine and, frequently, the filtered serineis reabsorbed. In garter snakes adapted to cold, thefraction of filtered glutamate and phenylalanine re-absorbed falls to about 60%.
The differences in amino acid transport betweenalligators and snakes observed in these clearancestudies may relate to the site of urine collection,from the ureters in snakes (Benyajati and Dant-zler, ’86a,b) and from the bladder in alligators(Hernandez and Coulson, ’67). Reabsorption oreven secretion of amino acids may occur in thebladder or cloaca of reptiles as well as in the re-nal tubules (Benyajati and Dantzler, ’86a). If thisis the case, the almost complete reabsorption ob-served in alligators may be the result of reabsorp-tion beyond the renal tubules. The possibility ofcloacal reabsorption is supported by preliminarystudies on sea snakes in which cloacal and ure-teral urine were collected from the same animals(Benyajati and Dantzler, ’86a), but this possibil-ity requires more direct confirmation.
No direct studies have been made of the tubulesite or sites of amino acid transport in reptiles. How-ever, as in other vertebrates, the proximal tubulemust be involved. In snakes, both net reabsorptionand net secretion probably occur in the proximaltubule. Moreover, evaluation of the transport rates

RENAL TRANSPORT OF ORGANIC MOLECULES IN REPTILES 677
as a function of filtered load in clearance studies inthese animals suggests that net reabsorption oc-curs proximal to net secretion in this tubule region(Benyajati and Dantzler ’86b). This requires confir-mation by a more direct approach.
In reptiles, the mechanism of amino acid trans-port has been studied only for taurine and onlywith BBMV and BLMV from garter snake (Tham-nophis sp.) kidneys (Benyajati and Johnson, ’91;Benyajati and Bay, ’92, ’94). However, the trans-port of this amino acid is of particular interestbecause it undergoes both net reabsorption andnet secretion in these animals.
Taurine is transported into BBMV against anelectrochemical gradient by two independent elec-trogenic, Na+ and Cl–-dependent cotransport sys-tems: (1) a high-affinity, low-capacity system (Kt≅ 70 µM; Jmax ≅ 22 pmol mg protein–1) that func-tions at taurine concentrations below ~0.8 mM;and (2) a low-affinity, high-capacity system (Kt ≅1.1 mM; Jmax ≅ 250 pmol mg protein–1) that func-tions at taurine concentrations above ~0.8 mM(Benyajati and Johnson, ’91; Benyajati and Bay,’92). The high-affinity system apparently trans-ports 2–3 Na+, 1 Cl–, and 1 taurine (Benyajati andBay, ’92). This coupling suggests that this systemfunctions only in the reabsorptive direction anddoes not play any role in the secretory process fortaurine. In this regard, the efflux of taurine fromBBMV is extremely slow, despite the absence ofevidence for intracellular binding, and is notstimulated by extravesicular taurine or NaCl(Benyajati and Johnson, ’91; Benyajati and Bay,’94). Because the normal concentration of taurinein the plasma, and thus in the glomerular filtrate,is ~1 µM, far below the Kt of the high-affinity sys-tem, it seems likely that this system functions un-der most physiological circumstances to reabsorbtaurine from the filtrate (Benyajati and Bay, ’92).However, when these animals are acclimated tocold (e.g., winter hibernation), the plasma taurineconcentration rises to ~0.5–~1.8 mM, suggestingthat under these circumstances the low-affinity,high-capacity system may function to reclaim fil-tered taurine (Benyajati and Bay, ’92).
Transport of taurine at the basolateral mem-brane is of particular importance because taurinecan undergo net secretion as well as net reabsorp-tion. Taurine is transported into BLMV by anelectroneutral cotransport system with Na+ andCl– (Benyajati and Bay, ’94). This system is of sub-stantially smaller capacity than the one identi-fied in BBMV. Moreover, unlike the transportsystem in the BBMV, that in the BLMV is driven
only by the Na+ gradient, not the Cl– gradient(Benyajati and Bay, ’94). Instead, Cl– apparentlyacts as a catalytic activator of the transporter(Benyajati and Bay, ’94). Also, there is only 1 Na+
transported per taurine molecule (Benyajati andBay, ’94). Finally, efflux of taurine from the BLMV,in contrast to that from the BBMV, is quite rapidand enhanced by extravesicular taurine and NaCl(Benyajati and Bay, ’94). The latter data suggestthat at least part of the taurine exit across thebasolateral membrane occurs by a carrier-medi-ated process, perhaps the reversal of the uptakestep (Benyajati and Bay, ’94). The combined datafrom the studies with BBMV and BLMV showinga luminal high-affinity, electrogenic, 2–3Na-1Cl–-1taurine cotransporter, a basolateral electro-neutral 1Na+-1Cl–-1taurine cotransporter, and amore marked efflux of taurine across the baso-lateral than the luminal membrane appear to re-flect an overall net reabsorptive process in theintact tubule (Benyajati and Bay, ’94). The inte-grated process involved in net secretion remainsto be determined.
In summary, for reptiles, in general, filteredamino acids largely are reabsorbed by the renaltubules. However, some of the apparent reabsorp-tion may occur in the cloaca or bladder distal tothe kidney. This has yet to be demonstrated di-rectly. Moreover, in some ophidian species, the de-gree of tubular reabsorption varies and nettubular secretion of taurine and some other β-amino acids can occur. The factors regulating thedegree of reabsorption or secretion are unknown,but studies of cold adaptation in species that nor-mally undergo hibernation suggest that controlmay relate to the metabolic state of the animals.As in other vertebrate classes, transport clearlyoccurs in the proximal tubule but no informationis available on other possible tubule sites of trans-port. Possible tubular transport mechanisms havebeen studied only for taurine, and only withBBMV and BLMV from snake nephrons. Thesestudies reveal Na+- and Cl–-dependent transport-ers that appear to be involved primarily in thereabsorptive process but provide no informationabout the secretory process. The low and variablereabsorption and the net secretion of amino acidsdemonstrated in snakes are highly unusual amongvertebrates, and the transport mechanisms in-volved and their regulation merit critical study.
ORGANIC ANIONS (OR ACIDS)A general system for the net secretion of a wide
range of endogenous and exogenous hydrophobic

678 W.H. DANTZLER
organic anions (or weak organic acids that existas anions at physiological pH), including manyenvironmental toxins, exists in the renal tubulesof reptiles as well as other vertebrates. There arecertainly other systems for the transport of spe-cific organic anions (or weak acids) in reptilianrenal tubules. However, except for the amino ac-ids (which are discussed above) and urate (whichis discussed below), nothing is known about thedetails of these transport systems in reptiles andthey are not considered in this discussion exceptas they relate to the general organic anion trans-port system.
The prototype organic anion for the study of thistransport system is p-aminohippurate (PAH).Clearance studies with this substance indicatethat the transport system is saturable and that,below the saturation level, the rate of secretion isa function of the load delivered to the peritubulartransporter (Dantzler, ’67). At the cellular andmembrane levels, almost all information availablehas come from studies of PAH transport in iso-lated, perfused snake (Thamnophis sp.) renal tu-
bules (Dantzler, ’88). These studies showed thatnet secretion from bath to lumen against a con-centration gradient occurs primarily in the distalportion of the proximal tubule, saturates at abathing medium concentration of ~6 × 10–5 M, andhas an apparent Kt of ~10 µM and Jmax of about325 fmol min–1 mm–1 (Dantzler, ’74a, ’76) (Fig. 1).
Further studies with this preparation have fo-cused on the possible mechanisms of transport atthe basolateral and luminal membranes of the tu-bule cells and the interactions that may occur be-tween them. Nearly 25 years ago, the basicpattern of transport at the two membranes wasestablished (Dantzler, ’74a,b). During the processof transepithelial transport of PAH from the bathto lumen against a concentration gradient, theconcentration inside the cells is greater than thatin either the bath or the lumen (Dantzler, ’74a).Because the inside of the cells is ~60 mV nega-tive compared to either the bath or lumen (Kimand Dantzler, ’95a), PAH is transported as an an-ion and there is no evidence of significant bind-ing within the cells, these original concentration
Fig. 1. Descriptive functional model for organic anion(PAH) secretion by snake proximal tubule cells. Kinetic con-stants (Kt and Jmax) and apparent permeabilities of luminal(PL) and peritubular (PP) membranes are shown. Broken ar-rows with open circles indicate carrier-mediated movementdown an electrochemical gradient. Solid arrows with opencircles indicate movement against an electrochemical gradi-
ent by some form of secondary active transport. Solid arrowsand solid circle indicate primary active transport. A– indi-cates anion of unknown type; αKG2– represents α-ketoglut-arate. Broken arrow with question mark indicates some formof possible feedback from the transport step across the lumi-nal membrane to the transport step across the basolateralmembrane.

RENAL TRANSPORT OF ORGANIC MOLECULES IN REPTILES 679
measurements established the basic pattern ofPAH transport into the cells against an electro-chemical gradient at the peritubular side andmovement from the cells into the lumen down anelectrochemical gradient (Fig. 1). Moreover, mea-surements of the apparent passive permeabilitiesof the membranes to PAH gave a value for theluminal membrane (~3.5 × 10–5 cm s–1) about seventimes that of the basolateral membrane (~0.5 ×10–5 cm s–1) (Dantzler, ’74a,b). These measure-ments too are compatible with the basic model(Fig. 1), if PAH transported into the cells at thebasolateral side is to move readily into the lumen,not back into the bath.
The transepithelial permeability measured di-rectly from PAH flux from lumen to bath (0.67 ×10–5 cm s–1) is virtually the same as that calcu-lated from the individually measured membranepermeabilities (0.44 × 10–5 cm s–1). The PAH fluxfrom lumen to bath is readily predicted from theindividual membrane permeabilities (Dantzler,’76). These data strongly suggest that the smallbackflux from lumen to bath moves across thecells, not between them. Moreover, this very smalllumen-to-bath flux and the low transepithelialpermeability are clearly reflected in the observa-tion that net PAH transport from bath to lumenis essentially independent of perfusion rate(Dantzler, ’74a).
Most studies since the initial ones have exploredthe transport step into the cells at the basolateralmembrane. Although transport occurs against anelectrochemical gradient and is readily eliminatedby general inhibitors of metabolism and energyproduction, there is no evidence of direct couplingof transport to hydrolysis of ATP (Dantzler, ’76).Therefore, it rapidly became apparent that thistransport step had to involve some form of sec-ondary active transport. Because of this and be-cause other secondary active transport systemsinvolving the coupling of inorganic ion transportto organic molecule transport were becoming wellaccepted, the initial studies of this process ex-plored the relationship of inorganic ions—Na+, K+,Ca2+—to the basolateral PAH transport step. Re-moval of Na+ from the medium bathing isolated,perfused snake renal tubules was shown to re-versibly depress net PAH secretion, apparently byinhibiting concentrative transport into the cellsacross the basolateral membrane and by increas-ing the apparent passive permeability of thatmembrane to PAH (Dantzler and Bentley, ’76).These effects do not result from an increase inthe concentration of cytosolic Ca2+ in response to
the removal of Na+ (Dantzler and Brokl, ’84a,b).Moreover, the requirement for Na+ clearly doesnot involve a directly coupled Na+-PAH cotrans-port system. However, the basolateral transportstep was shown to be inhibitable by the anion-exchange inhibitor, 4-acetamido-4′-isothiocyano-2,2′-disulfonic stilbene (SITS) (Dantzler andBentley, ’80), thereby suggesting that transportof PAH into the cells might involve counter-trans-port for some other anion whose transport intothe cells might be directly coupled to Na+ trans-port (Fig. 1). Based on studies performed withBLMV from rat kidneys, Burckhardt and co-work-ers (Shimada, et al., ’87) and Pritchard (’87, ’88)proposed such a tertiary transport system (Fig.1). In this proposal, the final step involves thecounter-transport of PAH (or other anions thatshare this system) into the cells against an elec-trochemical gradient, in exchange for a dicar-boxylate (physiologically, α-ketoglutarate [αKG])moving out of the cells down its electrochemicalgradient (Fig. 1). In the original model, the out-wardly directed gradient for αKG is maintained,in turn, by uptake across the basolateral mem-brane by the Na+-dicarboxylate cotransport sys-tem that exists in this membrane (Fig. 1). Thedirect input of energy for this tertiary system in-volves the Na+-K+-ATPase that resides in thebasolateral membrane and that maintains the lowintracellular concentration of Na+ by transport-ing it out of the cells (Fig. 1). These studies alsosuggested that the stoichiometry of the PAH/αKGexchange is 1:1, so that inside negative basolateralmembrane potential favors the outward movementof αKG (two negative charges) and the inwardmovement of PAH (one negative charge).
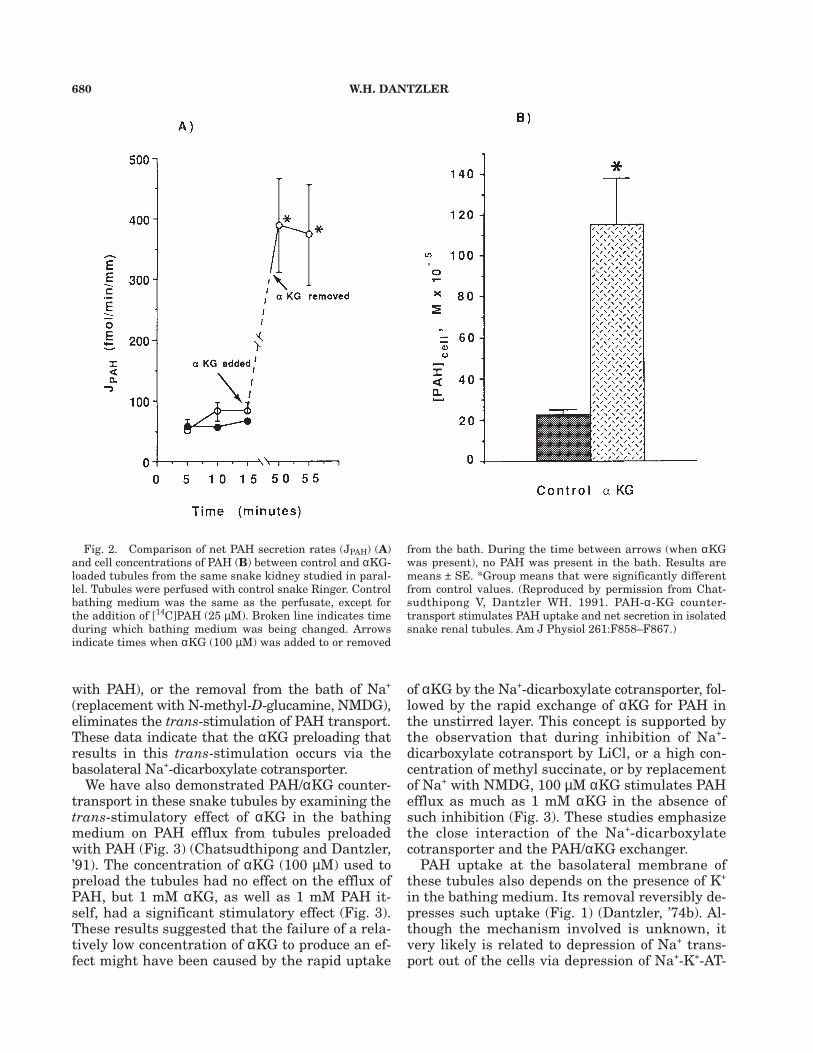
Although this model was developed from stud-ies with rat BLMV, we have demonstrated that itholds for PAH transport in intact perfused tubulesisolated from snake kidneys (Chatsudthipong andDantzler, ’91). When these tubules are preloadedwith αKG, both uptake into the cells and nettransepithelial transport increase to a comparableextent (Fig. 2). Moreover, the trans-stimulationshown in Fig. 2 is inhibited by the addition of SITSto the bathing medium, along with PAH after thetubules are preloaded with αKG, thereby support-ing the concept of PAH/αKG exchange. Also, in-hibition of the basolateral Na+-dicarboxylatecotransporter during the αKG preloading periodby the addition to the bath of LiCl (a relativelyspecific inhibitor of this transporter), or a highconcentration of methyl succinate (a competitive in-hibitor of αKG transport that does not exchange
680 W.H. DANTZLER
with PAH), or the removal from the bath of Na+
(replacement with N-methyl-D-glucamine, NMDG),eliminates the trans-stimulation of PAH transport.These data indicate that the αKG preloading thatresults in this trans-stimulation occurs via thebasolateral Na+-dicarboxylate cotransporter.
We have also demonstrated PAH/αKG counter-transport in these snake tubules by examining thetrans-stimulatory effect of αKG in the bathingmedium on PAH efflux from tubules preloadedwith PAH (Fig. 3) (Chatsudthipong and Dantzler,’91). The concentration of αKG (100 µM) used topreload the tubules had no effect on the efflux ofPAH, but 1 mM αKG, as well as 1 mM PAH it-self, had a significant stimulatory effect (Fig. 3).These results suggested that the failure of a rela-tively low concentration of αKG to produce an ef-fect might have been caused by the rapid uptake
of αKG by the Na+-dicarboxylate cotransporter, fol-lowed by the rapid exchange of αKG for PAH inthe unstirred layer. This concept is supported bythe observation that during inhibition of Na+-dicarboxylate cotransport by LiCl, or a high con-centration of methyl succinate, or by replacementof Na+ with NMDG, 100 µM αKG stimulates PAHefflux as much as 1 mM αKG in the absence ofsuch inhibition (Fig. 3). These studies emphasizethe close interaction of the Na+-dicarboxylatecotransporter and the PAH/αKG exchanger.
PAH uptake at the basolateral membrane ofthese tubules also depends on the presence of K+
in the bathing medium. Its removal reversibly de-presses such uptake (Fig. 1) (Dantzler, ’74b). Al-though the mechanism involved is unknown, itvery likely is related to depression of Na+ trans-port out of the cells via depression of Na+-K+-AT-
Fig. 2. Comparison of net PAH secretion rates (JPAH) (A)and cell concentrations of PAH (B) between control and αKG-loaded tubules from the same snake kidney studied in paral-lel. Tubules were perfused with control snake Ringer. Controlbathing medium was the same as the perfusate, except forthe addition of [14C]PAH (25 µM). Broken line indicates timeduring which bathing medium was being changed. Arrowsindicate times when αKG (100 µM) was added to or removed
from the bath. During the time between arrows (when αKGwas present), no PAH was present in the bath. Results aremeans ± SE. *Group means that were significantly differentfrom control values. (Reproduced by permission from Chat-sudthipong V, Dantzler WH. 1991. PAH-α-KG counter-transport stimulates PAH uptake and net secretion in isolatedsnake renal tubules. Am J Physiol 261:F858–F867.)

RENAL TRANSPORT OF ORGANIC MOLECULES IN REPTILES 681
Pase activity. This would, in turn, interfere withαKG transport into the cells across the basolateralmembrane.
Although our data clearly indicate that αKGtaken up by the cells via the basolateral Na+-dicarboxylate cotransporter can trans-stimulatePAH uptake, αKG produced in the cells by me-tabolism will also function to trans-stimulate PAHuptake via the PAH/αKG exchanger. The relativeimportance of basolateral uptake and metabolismin establishing the outwardly directed gradient ofαKG in snake tubules has not been determined.As far as metabolism is concerned, however, insnake tubules neither the basal level nor thetrans-stimulated PAH transport is influenced bythe buffer used in perfusing and bathing solutions.This may simply mean that the metabolic produc-tion of αKG in snake tubules is unaffected by thebuffer system. Moreover, the importance, underphysiological conditions, of αKG uptake via the
basolateral Na+-dicarboxylate cotransporter forreclaiming cellular αKG that has been exchangedfor PAH is not known.
In addition to uptake at the basolateral mem-brane, αKG is filtered at the glomerulus and isreabsorbed in the proximal tubule by the luminalNa+-dicarboxylate cotransporter. This transporterin mammals apparently has a lower affinity, buthigher capacity for substrate, than the basolateraltransporter (Wright and Wunz, ’87). It appearedpossible that reabsorption of αKG by the luminaltransporter might play a role in establishing theoutwardly directed gradient for αKG at thebasolateral membrane. We examined this possi-bility in preliminary experiments with perfusedtubules by measuring net PAH secretion in thepresence and absence of 100 µM αKG in the per-fusate. This concentration is about 10 times theconcentration normally present in the filtrate, butits presence produced about a two-fold stimula-
Fig. 3. Summary of effects of test substrates in bathingmedium (left) on efflux of [14C]PAH across the basolateralmembrane of snake tubules. Bars represent mean values ofefflux coefficients as percents of their own controls (with con-trol as 100%). Number of determinations is shown in paren-theses at end of each bar. Line at end of each bar is SE. MSindicates methyl succinate; NMDG indicates N-methyl-D-
glucamine. Mean control value for all experiments was 3.57± 0.19 × 10–6 cm/s. *Significant difference from control. (Re-produced by permission from Chatsudthipong V, Dantzler WH.1991. PAH-α-KG countertransport stimulates PAH uptakeand net secretion in isolated snake renal tubules. Am J Physiol261:F858–F867.)

682 W.H. DANTZLER
tion of net PAH secretion (Dantzler and Evans,unpublished observations). Glutarate (100 µM),which is not readily metabolized by renal tubulecells but does exchange for PAH at the basolateralmembrane, produced an identical effect whenadded to the lumen (Dantzler and Evans, unpub-lished observations). Thus, this stimulation was notsimply the result of increased energy productionfrom αKG metabolism. The stimulatory effect wasat least partially blocked by the simultaneous ad-dition of LiCl to the lumen (Dantzler and Evans,unpublished observations). These data suggest thattransport of αKG into the cells via the luminal Na+-dicarboxylate cotransporter can contribute to theoutwardly directed gradient for PAH/αKG exchangeat the basolateral membrane (Fig. 1). However, thephysiological significance of this process for normalorganic anion transport in snake renal tubules hasyet to be determined.
Although the data on snake renal tubules sup-port the presence of a PAH/αKG countertrans-porter at the basolateral membrane, it has notyet been determined if the molecular characteris-tics of such a carrier in snake tubules are the sameas those of the basolateral transporter recentlycloned from mammalian tissue (Sweet et al., ’97).However, the functional similarity between rep-tilian and mammalian transporters makes nearmolecular identity likely.
During net secretion, PAH transported into thecells against an electrochemical gradient at thebasolateral membrane moves out of the cells downan electrochemical gradient, across the luminalmembrane (Fig. 1). This luminal transport stepmust be mediated in some fashion because it canbe inhibited and because the permeability of theluminal membrane to PAH is much too high to beexplained by simple passive diffusion (see aboveand Fig. 1) (Dantzler, ’88). In isolated, perfusedsnake renal tubules, SITS inhibits PAH transportacross the luminal membrane, suggesting that itcould involve anion exchange (Fig. 1) (Dantzlerand Bentley, ’80). However, the presence of PAHor phenol red in the lumen not only fails to stimu-late PAH flux from the cells, but actually inhibitsit (Dantzler and Bentley, ’79). Transport of PAHinto the lumen does not require the presence ofCl– in the lumen, but appears to require a substi-tute for Cl– to which the membrane is highly per-meable (Dantzler and Bentley, ’81). Although theselast observations could reflect the presence of aluminal anion exchanger, they could also reflectthe effects of Cl– substitutions on the luminalmembrane potential; a point yet to be examined.
If they do reflect an effect on membrane poten-tial, transport from cell to lumen might occur bycarrier-mediated diffusion driven by the electro-chemical potential (Fig. 1). This transport processmay involve sulfhydral groups (Dantzler andBentley, ’83) and be dependent on Ca2+ entry intothe cells (Dantzler and Brokl, ’84a,b). In additionto these data obtained with isolated, perfusedsnake tubules, data obtained with snake renalBBMV suggest that the transport step for PAHacross the luminal membrane may be Na+-depen-dent (Benyajati and Dantzler, ’88). In any case,in snake renal tubules, as in the renal tubules ofother vertebrates, luminal transport of organicanions is much less well understood than baso-lateral transport.
Regardless of the nature of the luminal andbasolateral transporters in snake renal tubules,they may interact with one another. Any inhibi-tion of PAH movement from the cells to the lu-men of isolated, perfused snake renal tubulesresults in a reduction of PAH uptake into the cellsat the basolateral side (Dantzler and Bentley, ’79,’80, ’81; Dantzler and Brokl, ’84a,b). These datasuggest that there may be some type of feedbackcommunication between the luminal and baso-lateral transporters (Fig. 1).
URATEUrate, in its various forms, is the major excre-
tory end product of nitrogen metabolism in all rep-tiles except some chelonians. It is freely filteredat the glomerulus and undergoes net secretion bythe renal tubules (Dantzler, ’78). Almost all thedetailed information on the secretory processcomes from studies with isolated, perfused snake(Thamnophis sp.) renal tubules. Net secretionfrom bath to lumen against a concentration gra-dient occurs to a similar extent throughout theproximal tubule, not largely in one segment likePAH (Dantzler, ’73). Like net PAH secretion, how-ever, net urate secretion in these tubules involvestransport into the cells against an electrochemicalgradient at the peritubular side, followed by move-ment from the cells down an electrochemical at theluminal side (Fig. 4) (Dantzler, ’73). An inhibitoryeffect of SITS on the basolateral transporter sug-gests that it involves anion exchange, but what theexchanged anion might be is completely unknown(Fig. 4) (Mukherjee and Dantzler, ’85).
This overall basic model is similar to that forPAH, but there are significant differences betweenthe transport processes for urate and PAH, in ad-dition to the differences in anatomical localiza-

RENAL TRANSPORT OF ORGANIC MOLECULES IN REPTILES 683
tion noted above. First, the apparent permeabil-ity of the basolateral membrane to urate (~3.1 ×10–5 cm s–1) is about four times the permeabilityof luminal membrane (~0.75 × 10–5 cm s–1) (Fig.4) (Dantzler, ’76), a relationship that appears tobe quite inefficient if urate transported into thecells at the peritubular side is to move into thelumen, not back into the bath. Second, basolateraltransport into the cells increases when there isfiltrate (or a similar artificial solution) in the lu-men and decreases when it is absent (Dantzler,’73). Third, although there is no evidence of netreabsorption, substantial passive unidirectionalreabsorption can occur (Dantzler, ’73). Moreover,this backflux apparently occurs between the cells(Fig. 4) because the apparent permeability of theentire epithelium determined directly from theefflux measurements (~ 2.4 × 10–5 cm s–1) is aboutfour times the value (~0.60 × 10–5 cm s–1) calculatedfrom the independently measured permeabilities ofthe luminal and basolateral membranes (Dantzler,’76). Because reptilian nephrons filter intermittently,these three characteristics of urate transport—thehigh permeability of the basolateral membrane, thedependence of basolateral transport on filtrate inthe lumen, and the large paracellular backflux—
may have evolved to prevent accumulation of in-soluble urates in the cells and lumens of neph-rons that are not filtering.
Fourth, for net urate secretion, the apparent Kt(~150 µM) is about 15 times and the apparentJmax (~140 fmol min–1 mm–1) is about 0.5 timesthe corresponding values for net PAH secretion(Figs. 1 and 4) (Dantzler, ’73, ’74a, ’82). However,although the Kt for urate is much higher than thatfor PAH, it still is well below the physiologicalplasma urate concentration (~400–500 µM) inthese snakes. This suggests that the transport sys-tem is normally nearly saturated and that the rateof net secretion is largely determined by the rateof backdiffusion and, thus, by the rate of flowalong the lumen. Fifth, in contrast to PAH, neturate secretion shows no dependence on Na+
(Randle and Dantzler, ’73), making it quite unlikelythat the basolateral transport step involves counter-transport for dicarboxylates such as αKG. Sixth,also in contrast to PAH, net urate secretion showsno sensitivity to Ca2+ entry into the cells (Dantzlerand Brokl, ’84c). Seventh, however, like PAH, neturate secretion is K+-dependent. Removal of K+ fromthe bathing medium reversibly depresses net uratesecretion by completely eliminating transport into
Fig. 4. Descriptive functional model for urate secretion by snake proximal tubule cells.Symbols are the same as in Fig. 1.

684 W.H. DANTZLER
the cells against an electrochemical gradient at thebasolateral membrane (Randle and Dantzler, ’73).In view of the lack of effect of Na+ removal, it isdifficult at present to postulate an appropriatemechanism for this effect.
Eighth and finally, in very marked contrast toPAH transport, there is no evidence the movementof urate from the cells into the lumen during netsecretion is carrier-mediated. In isolated, perfusedsnake proximal tubules, neither SITS, probenecid,nor even unlabelled urate itself has any effect onthe movement of radioactive urate across the lu-minal membrane or on the apparent permeabil-ity of that membrane to urate (Dantzler andBentley, ’79; Mukherjee and Dantzler, ’85). More-over, there is no evidence of mediated urate trans-port in BBMV from snake kidneys (Benyajati andDantzler, ’88). These data all are compatible withurate movement across the luminal membrane bypurely passive diffusion. However, there is no di-rect evidence for passive diffusion. Moreover, theabsence of evidence for carrier-mediated transportdoes not prove its absence, and the normal secre-tory flux of urate across this membrane is suffi-ciently large that the process deserves additionalexamination.
ORGANIC CATIONS (OR BASES)Less is known about renal transport of organic
cations than organic anions in reptiles, but it ap-pears that such transport must be important inregulating the internal concentrations of suchcompounds. These include endogenous naturalcompounds (e.g., choline or N1-methylnicotina-mide [NMN]) and exogenous natural or syntheticcompounds (e.g., tetraethylammonium [TEA]). Asin the case of organic anion transport, almost allthe information on renal organic cation transportin reptiles comes from studies with isolated, per-fused snake (Thamnophis sp.) renal tubules(Hawk and Dantzler, ’84; Dantzler and Brokl, ’86,’87, ’88; Kim and Dantzler, ’95a, ’96, ’97a,b). Thesestudies involved TEA and NMN, quaternary am-monium compounds of quite different structure.They indicate that either net secretion or net re-absorption, depending on the cation, can occurthroughout the proximal tubule (Dantzler, unpub-lished observations; Hawk and Dantzler, ’84;Dantzler and Brokl, ’86).
Initial studies indicated that TEA undergoes netsecretion resulting from the difference between asmall saturable flux from lumen to bath and amuch larger saturable flux from bath to lumen(Fig. 5) (Hawk and Dantzler, ’84). Later studies
concentrated on the transport steps at the baso-lateral and luminal membranes, particularly asthey apply to the large transepithelial secretoryflux. The Kt for both the transepithelial secretoryflux (~20 µM) and the initial rate of basolateraltransport into the cells (~18 µM) are essentiallythe same, suggesting that this transport step israte limiting for the transepithelial flux (Hawkand Dantzler, ’84; Kim and Dantzler, ’95a). TEAuptake into the cells across the basolateral mem-brane is decreased by ~25% when the basolateralmembrane potential is reduced from –60 to about–20 mV (by increasing bath K+ concentration from3 to 65 mM or adding 10 mM Ba2+ to the bath)(Kim and Dantzler,’95a, ’97b). These data are com-patible with electrogenic transport into the cellsacross the basolateral membrane (Fig. 5). How-ever, in addition to entry driven by the membranepotential, studies of radiolabeled TEA efflux acrossthe basolateral membrane of tubules with oil-filledlumens indicate that transport can be driven byorganic cation/organic cation exchange (e.g., ex-change for unlabeled TEA or choline) (Dantzlerand Brokl, ’88). Electrogenic-facilitated diffusionand electroneutral organic cation/organic cationexchange appear to be functions of a single trans-porter operating in different modes (Fig. 5).
A number of other features of the basolateralTEA transporter have also been defined. First,during both transepithelial bath-to-lumen flux andbasolateral uptake, steady-state intracellular con-centrations of TEA at any membrane potentialstudied are higher than those predicted for pas-sive distribution at electrochemical equilibrium(Hawk and Dantzler, ’84; Kim and Dantzler, ’95a).These observations suggest that transport into thecells across the basolateral membrane can occuragainst an electrochemical gradient. However, nei-ther electrogenic facilitated diffusion nor electro-neutral organic cation/organic cation exchange cantransport TEA against an electrochemical gradi-ent. Moreover, the high intracellular concentra-tion can be reduced, but not eliminated, bymetabolic inhibitors such as cyanide. It seemsmost likely that these high intracellular concen-trations are the result of binding to cytoplasmiccomponents, or sequestration in intracellular com-partments (Pritchard and Miller, ’93). Second,specificity studies of the basolateral transporterinvolving inhibition of TEA uptake by a series ofn-tetraalkylammonium (n-TAA) compounds (n =1–5) (including TEA itself) indicate that inhibi-tory potency increases with increasing alkyl chainlength (Fig. 6) (Kim and Dantzler, ’96). Thus, the

RENAL TRANSPORT OF ORGANIC MOLECULES IN REPTILES 685
apparent affinity of the carrier for n-TAA com-pounds increases with their increasing hydropho-bicity (or lipophilicity). These findings are virtuallyidentical to those obtained in studies of mamma-lian proximal tubules (Fig. 6) (Groves, et al., ’94).
Third, and of particular significance, a consid-erable body of data now demonstrates that trans-port of TEA across the basolateral membrane ofsnake proximal tubules is asymmetrical in a fash-ion that favors uptake into the cells over effluxinto the peritubular fluid. These studies involvedcomparisons of the basolateral efflux of radiola-beled TEA from tubules with oil-filled lumens withbasolateral uptake of radiolabeled TEA. The effluxof TEA across the basolateral membrane, like up-take, occurs by a saturable process that is de-scribed by Michaelis-Menten kinetics (Kim andDantzler, ’96). However, the Kt for efflux (~100µM) is about six times the Kt for uptake (~18 µM),suggesting that the affinity of the carrier for TEAis greater in the uptake direction than in the efflux
Fig. 5. Descriptive functional model for tetraethylammo-nium (TEA+) and N1-methylnicotinamide (NMN+) in snake
proximal renal tubules. OC+ indicates organic cation of un-known type. Other symbols are the same as in Fig. 1.
direction (Kim and Dantzler, ’96). The reductionin basolateral membrane potential describedabove, instead of increasing efflux of positivelycharged TEA as it reduces uptake, has no effecton efflux (Kim and Dantzler, ’95a). Thus, mem-brane potential only influences carrier-mediatedentry, not exit across the basolateral membrane.
An additional aspect of the asymmetry of baso-lateral transport involves the effects of H+ con-centration. An initial study indicated that effluxof TEA across the basolateral membrane couldbe trans-stimulated very slightly by a low pHin the bathing medium, thereby suggesting thatTEA/H+ exchange might occur at this membrane(Dantzler and Brokl, ’88). In this study, how-ever, not only was the apparent trans-stimula-tion very slight, we also had no measurementsof intracellular pH (pHi) and, thus, of the pos-sible H+ gradient across the basolateral mem-brane. Nevertheless, because we had determinedthat physiological pHi normally was lower than

686 W.H. DANTZLER
some type of direct effect on basolateral transportindependent of any effect on membrane potential.Of particular interest, there appears to be an op-timal intracellular H+ concentration for TEA up-take that corresponds to the one found at thephysiological pHi of 7.1. It also appears that thereis an optimal extracellular H+ concentration forTEA efflux that corresponds to the one found inthe extracellular fluid at the physiological pH of7.4. The mechanism involved in this control is un-known, but it certainly supports the concept ofasymmetrical carrier-mediated transport of or-ganic cations across the basolateral membrane ofsnake renal tubules.
If, indeed, entry of TEA into the tubule cells acrossthe basolateral membrane is down an electrochemi-cal gradient, as appears to be the case despite thehigh intracellular concentrations developed, thentransport out of the cells into the lumen must beagainst an electrochemical gradient. This now ap-pears to be the case in mammalian tubules, andtransport into the lumen appears to be driven bycountertransport for H+ (Dantzler and Wright, ’97).That also appears to be the case for reptilian renaltubules. Our first studies on TEA efflux from thecells across the luminal membrane of tubules cov-ered with oil indicated that efflux could be substan-tially accelerated by a low pH in the lumen, as wellas by the addition of unlabeled TEA or choline tothe lumen (Dantzler and Brokl, ’88). This first studyclearly indicated that TEA/H+ exchange could oc-cur across the luminal membrane in intact tubules.To examine this more closely, we evaluated TEA/H+ countertransport in BBMV from these snake re-nal tubules (Dantzler, et al., ’91). The data showthat an outwardly directed H+ gradient (inside pH6.0, outside pH 7.5) not only stimulates uptake intothese BBMV, but also supports concentrative accu-mulation (transient intravesicular concentrationabove equilibrium). These data support the earlierones and indicate that TEA/H+ countertransport iscapable of moving TEA into the lumen against anelectrochemical gradient (Fig. 5). In BBMV, the ini-tial rate of this H+-gradient driven TEA uptake is asaturable process that is adequately described byMichaelis-Menten kinetics, yielding a Kt of ~500 µM(Dantzler et al., ’91). These studies also demonstratethat TEA uptake is independent of membrane po-tential, thereby indicating that TEA/H+ exchangeis electroneutral. Finally, the studies show that, asin mammalian proximal renal tubules, a Na+/H+ ex-changer exists in the luminal membrane of thesesnake proximal tubules.
In summary, these data indicate that in snakes,
Fig. 6. Relationship between alkyl chain length and in-hibitory potency of n-tetraalkylammonium compounds for iso-lated snake and rabbit proximal renal tubules. Open circlesrepresent data from snake tubules. Each point is mean ± SEfor 5–6 experiments. Solid circles represent data from rabbittubules. Each of these points is mean ± SE for three experi-ments. Line is fitted to all points by the method of leastsquares, but simply is added to indicate general relationshipbetween alkyl chain length and inhibitory potency. The rab-bit data are from Groves et al. (’94). The snake data are fromKim and Dantzler (’96). (Reproduced by permission from KimYK, Dantzler WH. 1996. Specificity of basolateral organic cat-ion transport in snake renal proximal tubules. Am J Physiol270:R1025–R1030.)
the extracellular peritubular pH (pHo) (Kim andDantzler, ’95b), it appeared that TEA/H+ exchangecould be physiologically important if it functionedto stimulate transport into the cells. Therefore,we evaluated the effects of altering both pHo andpHi (and, thus, the H+ gradient across the mem-brane) on basolateral uptake and efflux of TEA(Kim and Dantzler, ’97a). We altered pHo by ad-justing the buffer of the bathing medium and pHiby the short-term addition of either NH4Cl orpropionic acid to the bathing medium (Kim andDantzler, ’97a). These studies gave no indicationof trans-stimulation of either uptake or efflux ofTEA. Therefore, TEA/H+ exchange does not ap-pear to occur across the basolateral membrane.However, we found evidence for trans-inhibitionof both TEA uptake and efflux, as well as cis-in-hibition of TEA uptake, by increasing H+ concen-trations. A high H+ concentration appears to have

RENAL TRANSPORT OF ORGANIC MOLECULES IN REPTILES 687
as in mammals, transport of TEA out of the cellsacross the luminal membrane involves a tertiaryactive transport process (Fig. 5). The final step inthis process is the transport of TEA out of the cellsagainst its electrochemical gradient in exchange forH+ moving into the cells down its electrochemicalgradient (Fig. 5). The inwardly directed H+ gradi-ent is established in turn by the luminal Na+/H+
exchanger in which H+ is transported out of the cellsagainst its electrochemical gradient, in exchange forNa+ moving into the cells down its electrochemicalgradient (Fig. 5). Finally, the direct energy expen-diture in this process involves the maintenance ofa low intracellular Na+ concentration by the trans-port of Na+ out of the cells against its electro-chemical gradient via Na+-K+-ATPase at thebasolateral membrane (Fig. 5).
Transport of NMN by snake proximal tubulesappears to be distinct from the transport of TEA(Dantzler and Brokl, ’86). This is in marked con-trast to mammalian proximal tubules in whichmost monovalent organic cations, including NMN,share a common process for net secretion (Wrightand Dantzler, ’97). Indeed, in studies on mammals,NMN has been used as frequently as TEA as aprototypical organic cation for the study of thistransport system. In these snake renal tubules,however, there is no inhibition of NMN transportby even very high concentrations of TEA (Dantzlerand Brokl, ’86), and only minimal inhibition ofTEA transport by concentrations of NMN highenough to cause nonspecific effects (Hawk andDantzler, ’84).
For NMN, as for TEA, there are saturable lu-men-to-bath and bath-to-lumen fluxes (Fig. 5)(Dantzler and Brokl, ’86). However, both fluxes aremuch larger (about three times) than even the bath-to-lumen flux for TEA (Fig. 5) (Hawk and Dantzler,’84; Dantzler and Brokl, ’86). Moreover, the lumen-to-bath flux is slightly larger than the bath-to-lu-men flux, resulting in apparent net reabsorptionrather than net secretion (Fig. 5) (Dantzler andBrokl, ’86). Indeed, direct measurements of net fluxshowed that there is always a small net reabsorptiveflux that is quantitatively equal to the differencebetween the two separately measured unidirectionalfluxes (Dantzler and Brokl, ’86). Thus, for NMN,relatively large unidirectional fluxes differ onlyslightly, producing a small net reabsorptive flux;whereas for TEA, much smaller unidirectionalfluxes differ greatly, producing a relatively large netsecretory flux (Fig. 5).
In the isolated, perfused snake tubules, NMNenters the cells during each unidirectional flux
down an electrochemical gradient apparently viaNa+-dependent, carrier-mediated diffusion (Fig. 5)(Dantzler and Brokl, ’86). It is then transportedout of the cells across the opposite membrane,against an electrochemical gradient (Fig. 5)(Dantzler and Brokl, ’86). This pattern is appar-ent even during measurements of net reabsorp-tion, or during stop-flow measurements with equalconcentrations of NMN in lumen and bath, be-cause the intracellular concentration of NMN atthese times is always far below that expected forpassive distribution at electrochemical equilibrium(Dantzler and Brokl, ’86). Studies of inhibitionwith structural analogues of NMN indicate that,at a minimum, a ring configuration containing aquaternary ammonium is essential for interactionwith the transporters at either membrane (Dant-zler and Brokl, ’86).
Studies of efflux of NMN across the basolateralmembrane of tubules with oil-filled lumens, andacross the luminal membrane of tubules coveredwith oil, indicate that transport out of the cells atthe basolateral, but not the luminal, membrane caninvolve countertransport for other organic cations(Fig. 5) (Dantzler and Brokl, ’87). This basolateraltransport step, which is probably the dominant onein the net reabsorptive process, may also be acti-vated by H+ or possibly involve NMN/H+ exchange(Fig. 5) (Dantzler and Brokl, ’87). Preliminary stud-ies with BBMV from snake kidneys, like those withintact tubules, provide no evidence for any counter-transport process for NMN at the luminal mem-brane (Dantzler, et al., ’91). Thus, there is noinformation on the mechanism of NMN transportfrom the cells at this membrane. There is also noinformation available at present as to whether thecarrier for basolateral entry of either NMN or TEAinto the cells resembles the one cloned from mam-malian tissue (Gründemann et al., ’94). It is clearthat much remains to be done to understand thedetails of the NMN transport process in reptilianrenal tubules.
LITERATURE CITEDBarfuss DW, Dantzler WH. 1976. Glucose transport in iso-
lated perfused proximal tubules of snake kidney. Am JPhysiol 231:1716–1728.
Benyajati S, Bay SM. 1990. Isolation and characterization ofbasolateral membranes from snake kidney cells: compari-son with brush-border membranes. Physiologist 33:A95.
Benyajati S, Bay, SM. 1992. Sodium-taurine cotransport inreptilian renal brush-border membrane vesicles. PflügersArch 421:168–175.
Benyajati S, Bay SM. 1994. Basolateral taurine transport sys-tem in reptilian renal cells. Am J Physiol 266:F439–F449.
Benyajati S, Dantzler WH. 1986a. Plasma levels and renal

688 W.H. DANTZLER
handling of endogenous amino acids in snakes: a compara-tive study. J Exp Zool 238:17–28.
Benyajati S, Dantzler WH. 1986b. Renal secretion of aminoacids in ophidian reptiles. Am J Physiol 250:R712–R720.
Benyajati S, Dantzler WH. 1988. Enzymatic and transportcharacteristics of isolated snake renal brush-border mem-branes. Am J Physiol 255:R52–R60.
Benyajati S, Johnson JL. 1991. Characteristics of renal trans-port of taurine in reptilian brush-border membrane vesicles.Am J Physiol 260:R879–R888.
Chatsudthipong V, Dantzler WH. 1991. PAH-a-KG counter-transport stimulates PAH uptake and net secretion in iso-lated snake renal tubules. Am J Physiol 261:F858–F867.
Dantzler WH. 1967. Glomerular and tubular effects of argin-ine vasotocin in water snakes (Natrix sipedon). Am J Physiol212:83–91.
Dantzler WH. 1973. Characteristics of urate transport by iso-lated perfused snake proximal renal tubules. Am J Physiol224:445–453.
Dantzler WH. 1974a. PAH transport by snake proximal re-nal tubules: differences from urate transport. Am J Physiol226:634–641.
Dantzler WH. 1974b. K+ effects on PAH transport and mem-brane permeabilities in isolated snake renal tubules. Am JPhysiol 117:1361–1370.
Dantzler WH. 1976. Comparison of uric acid and PAH trans-port by isolated, perfused snake renal tubules. In: SilbernaglS, Lang F, Greger R, editors. Amino acid and uric acid trans-port. Stuttgart: Thieme. p 169–180.
Dantzler, WH. 1978. Urate excretion in nonmammalian ver-tebrates. In: Kelley WN, Weiner IM, editors. Handbook ofexperimental pharmacology, vol 51: uric acid. Heidelberg:Springer-Verlag. p 185–210.
Dantzler WH. 1982 Studies on nonmammalian nephrons. Kid-ney Int 22:560–570.
Dantzler WH. 1988. Comparative physiology of the vertebratekidney. Springer-Verlag, Heidelberg. p 198.
Dantzler WH, Bentley SK. 1976. Low Na+ effects on PAHtransport and permeabilities in isolated snake renal tubules.Am J Physiol 230:256–262.
Dantzler WH, Bentley SK. 1979. Effects of inhibitors in lu-men on PAH and urate transport by isolated renal tubules.Am J Physiol 236:F379–F386.
Dantzler WH, Bentley SK. 1980. Bath and lumen effects ofSITS on PAH transport by isolated perfused renal tubules.Am J Physiol 238:F16–F25.
Dantzler WH, Bentley SK. 1981. Effects of chloride substi-tutes on PAH transport by isolated perfused renal tubules.Am J Physiol 241:F632–F644.
Dantzler WH, Bentley SK. 1983. Effects of sulfhydral re-agent, p-chloromercuribenzoate, on p-aminohippuratetransport by isolated, perfused snake renal tubules. RenalPhysiol 6:209–217.
Dantzler WH, Brokl OH. 1984a. Effects of low [Ca2+] andLa3+ on PAH transport by isolated perfused renal tubules.Am J Physiol 236:F175–F187.
Dantzler WH, Brokl OH. 1984b. Verapamil and quinidine ef-fects on PAH transport by isolated perfused renal tubules.Am J Physiol 246:F188–F200.
Dantzler WH, Brokl OH. 1984c. Lack of effect of low [Ca2+],La3+, and pyrazinoate on urate transport by isolated, per-fused snake renal tubules. Pflügers Arch 401:262–265.
Dantzler WH, Brokl OH. 1986. N1-methylnicotinamide trans-port by isolated perfused snake proximal renal tubules. AmJ Physiol 250:F407–F418.
Dantzler WH, Brokl OH. 1987. NMN transport by snake re-nal tubules: choline effects, countertransport, H+-NMN ex-change. Am J Physiol 253:F656–F663.
Dantzler WH, Brokl OH. 1988. TEA transport by snake re-nal tubules: choline effects, countertransport and H+-TEAexchange. Am J Physiol 255:F167–F176.
Dantzler, WH, Wright, SH. 1997. Renal tubular transportof organic anions and cations. In: Sipes IG, McQueenCA, Gandolfi AJ, Goldstein R, editors. Comprehensivetoxicology vol 7: renal toxicology. Oxford: Pergamon. p61–75.
Dantzler WH, Wright SH, Brokl OH. 1991. Tetraethylammo-nium transport by snake renal brush-border membranevesicles. Pflügers Arch 418:325–332.
Groves CE, Evans KK, Dantzler WH, Wright SH. 1994.Peritubular organic cation transport in isolated rabbit proxi-mal tubules. Am J Physiol 266:F450–F458.
Gründemann D, Gorboulev V, Gambaryan S, Veyl M, KoepsellH. 1994. Cloning of an organic cation transporter whichmediates renal and hepatic excretion of multiple drugs. Na-ture 372:549–522.
Hawk CT, Dantzler WH. 1984. Tetraethylammonium trans-port by isolated perfused snake renal tubules. Am J Physiol246:F476–F487.
Hernandez T, Coulson RA. 1967. Amino acid excretion in thealligator. Comp Biochem Physiol 23:775–784.
Kim YK, Dantzler WH. 1995a. Relation of membrane poten-tial to basolateral TEA transport in isolated snake proxi-mal renal tubules. Am J Physiol 268:R1539–R1545.
Kim YK, Dantzler WH. 1995b. Intracellular pH in snake re-nal proximal tubules. Am J Physiol 269:R822–R829.
Kim YK, Dantzler WH. 1996. Specificity of basolateral or-ganic cation transport in snake renal proximal tubules. AmJ Physiol 270:R1025–R1030.
Kim YK, Dantzler WH. 1997a. Effects of pH on basolateraltetraethylammonium transport in snake renal proximal tu-bules. Am J Physiol 272:R955–R961.
Kim YK, Dantzler WH. 1997b. Ca2+ and Ba2+ effects onbasolateral tetraethylammonium transport in isolated snakerenal proximal tubules. Pflügers Arch 435:28–33.
Mukherjee SK, Dantzler WH. 1985. Effects of SITS on uratetransport by isolated, perfused snake renal tubules. PflügersArch 403:35–40.
Pritchard JB. 1987. Luminal and peritubular steps in renaltransport of p-aminohippurate. Biochim Biophys Acta906:295–308.
Pritchard JB. 1988. Coupled transport of p-aminohippurateby rat kidney basolateral membrane vesicles. Am J Physiol255:F597–F604.
Pritchard JB, Miller MS. 1993. Mechanisms mediating re-nal secretion of organic anions and cations. Physiol Rev73:765–796.
Randle HW, Dantzler WH. 1973. Effects of K+ and Na+ onurate transport by isolated perfused snake renal tubules.Am J Physiol 225:1206–1214.
Shimada H, Moewes B, Burckhardt G. 1987. Indirect cou-pling to Na+ of p-aminohippuric acid uptake into ratbasolateral membrane vesicles. Am J Physiol 253:F795–F801.
Sweet DH, Wolff NA, Pritchard JB. 1997. Expression cloningand characterization of ROAT1. J Biol Chem 272:30088–30095.
Wright SH, Wunz TM. 1987. Succinate and citrate transportin renal basolateral, brush-border membranes. Am J Physiol253:F432–F439.