Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 93, pp. 3319-3324, April 1996Plant Biology
Association of glycolate oxidation with photosynthetic electrontransport in plant and algal chloroplasts
(carbon concentration mechanism/dissolved inorganic carbon pump/photorespiration/quinone oxidoreductase)
ARUN GOYAL AND N. EDWARD TOLBERT
Department of Biochemistry, Michigan State University, East Lansing, MI 48824-1319
Contributed by N. Edward Tolbert, December 18, 1995
ABSTRACT Photosynthetic carbon metabolism is initi-ated by ribulose-bisphosphate carboxylase/oxygenase(Rubisco), which uses both CO2 and 02 as substrates. One2-phosphoglycolate (P-glycolate) molecule is produced foreach 02 molecule fixed. P-glycolate has been considered to bemetabolized exclusively via the oxidative photosynthetic car-bon cycle. This paper reports an additional pathway forP-glycolate and glycolate metabolism in the chloroplasts.Light-dependent glycolate or P-glycolate oxidation by osmot-ically shocked chloroplasts from the algae Dunaliella or spin-ach leaves was measured by three electron acceptors, methylviologen (MV), potassium ferricyanide, or dichloroindophe-nol. Glycolate oxidation was assayed with 3-(3,4)-dichloro-phenyl)-1,1-dimethylurea (DCMU) as oxygen uptake in thepresence ofMV at a rate of 9 ,umol per mg of chlorophyll perh. Washed thylakoids from spinach leaves oxidized glycolateat a rate of 22 ,jmol per mg of chlorophyll per h. Thislight-dependent oxidation was inhibited completely by SHAM,an inhibitor of quinone oxidoreductase, and 75% by 2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone (DBMIB),which inhibits electron transfer from plastoquinone to thecytochrome b6f complex. SHAM stimulated severalfold gly-colate excretion by algal cells, Dunaliella or Chlamydomonas,and by isolated Dunaliella chloroplasts. Glycolate and P-glycolate were oxidized about equally well to glyoxylate andphosphate. On the basis of results of inhibitor action, thepossible site which accepts electrons from glycolate or P-glycolate is a quinone after the DCMU site but before theDBMIB site. This glycolate oxidation is a light-dependent,SHAM-sensitive, glycolate-quinone oxidoreductase systemthat is associated with photosynthetic electron transport inthe chloroplasts.
All plants and algae have the same bifunctional enzyme,ribulose bisphosphate carboxylase/oxygenase (Rubisco; EC4.1.1.39) for photosynthetic carbon metabolism. The carbox-ylase reaction initiates CO2 fixation and reduction by the C3reductive photosynthetic cycle, and the oxygenase reactioninitiates the C2 oxidative photosynthetic carbon cycle. The twocycles coexist as competitive reactions for the two gaseoussubstrates CO2 and 02, and together they represent photosyn-thetic carbon metabolism (1). 2-Phosphoglycolate (P-glycolate) synthesis in the chloroplasts by the oxygenase ac-tivity of Rubisco and its metabolism via the C2 oxidativephotosynthetic carbon cycle in the peroxisomes and mitochon-dria is equated with photorespiratory carbon metabolism. Inhigher plants and many multicellular algae, glycolate is oxi-dized by glycolate oxidase in the peroxisomes. However, inunicellular green algae, glycolate oxidase and leaf-type per-oxisomes are absent, and instead these algae contain a lowlevel of a mitochondrial glycolate dehydrogenase ["1 ,tmolper mg of chlorophyll (mg-1 Chl).h-1] (1-4). The glycolate-
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
dependent dichloroindophenol (DCIP, also known as
DCPIP)-reduction activity of unicellular algae has beentermed a dehydrogenase, because the electron acceptor fromthe partially purified enzyme is not directly linked to oxygen(4). The flux of carbon via glycolate, which is about 10-15,Lmol.mg-I Chl-h-l, is considered to be equal to its excretionin the presence of the aminotransferase inhibitor aminooxy-acetate (AOA), which blocks its metabolism (5). Since the fluxof excreted glycolate is substantially higher than the measur-
able capacity of the dehydrogenase, it is possible that theglycolate dehydrogenase cannot be isolated or measured ac-
curately or that glycolate may also be metabolized by anothersystem. The later hypothesis is consistent with high activity ofRubisco and P-glycolate phosphatase for glycolate synthesis inthe chloroplasts (4, 6).
Previous investigators had studied glycolate formation andexcretion by chloroplasts, but a system for glycolate metabo-lism in the chloroplasts has not been described (7-10). Theblue green alga Anabaena variabilis has been reported toexhibit photosystem I (PS I)-dependent oxidation of glycolate(11). We find that salicylhydroxamic acid (SHAM) and AOAgreatly stimulate glycolate excretion by algal cells or isolatedalgal chloroplasts (refs. 12 and 13; see Fig. 1). This suggestsinhibition of an active glycolate metabolism system in thechloroplasts, because in the absence of inhibitors glycolate didnot accumulate in large amounts. SHAM or AOA does notinhibit or change the activity of Rubisco or P-glycolate phos-phatase for glycolate synthesis in the chloroplasts. In this paperevidence is presented for a light-dependent glycolate oxidationwhich is associated with the photosynthetic electron transportsystem from algae (Dunaliella) or a higher plant (spinach).Preliminary data were reported in an abstract (13).
MATERIALS AND METHODS
Organism, Plant Material, Preparation of Chloroplasts andThylakoids, and Electron Transport Assay. Axenic cultures ofDunaliella tertiolecta were grown photoautotrophically andconcentrated as described elsewhere (14). Photosyntheticallyactive chloroplasts were isolated from either high-C02-grownor air-adapted cells, which must be in the logarithmic phase ofgrowth (15). The chloroplasts had to be used within 2 h afterpreparation, because activity for the Hill reaction and theglycolate-dependent reduction of ferricyanide decreased rap-idly (data not shown). Spinach (Spinacia oleracea L. cv. SavoyHybrid 612; Harris Seed Company, Rochester, NY) was grownin a greenhouse for 4 weeks, and young leaves were picked inthe morning. Washed spinach chloroplasts were obtained by
Abbreviations: AOA, aminooxyacetate; CCCP, carbonylcyanide3-chlorophenylhydrazone; Chl, chlorophyll; Cyt, cytochrome; DB-MIB, 2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone; DCIP (alsoknown as DCPIP), dichloroindophenol; DCMU, 3-(3,4-dichlorophe-nyl)-1,1-dimethylurea; DIC, dissolved inorganic carbon; E, einstein;FCCP, carbonylcyanide 4-trifluoromethoxyphenylhydrazone; MV,methyl viologen; PS I or II, photosystem I or II; Rubisco, ribulosebisphosphate carboxylase/oxygenase; SHAM, salicylhydroxamic acid.
3319

3320 Plant Biology: Goyal and Tolbert
the method of Cerovic and Plesnicar (16), and chloroplastswere further purified on a Percoll gradient (15) and resus-pended to a concentration of 1 mg of Chl-ml-1.
Thylakoid membranes were prepared from fresh deveinedspinach leaves. Leaves (125 g) were homogenized in a Waringblender for 2 periods of 5 sec in 125 ml of isolation mediumcontaining 40 mM Tricine, 400 mM NaCl, 2 mM MgCl2, and0.2% bovine serum albumin (BSA), pH adjusted to 8. Thehomogenate was filtered through four layers of cheeseclothand centrifuged at 300 x g for 1 min to remove unbroken cellsand debris. The resultant supernatant was centrifuged at 4000x g for 5 min and the pellet was resuspended in 50 ml of"high-salt" medium containing 20 mM Tricine, 150 mM NaCl,5 mM MgCl2, and 0.2% BSA, pH adjusted to 8.0. Thissuspension was centrifuged at 4000 x g for 5 min and the pelletwas resuspended in the same manner in a "low-salt" mediumcontaining 20 mM Tricine, 15 mM NaCl, 400 mM sucrose, and0.2% BSA, pH adjusted to 8.0. The resuspended thylakoids (1mg Chl-ml-1) were packaged in 0.5-ml aliquots and stored at-80°C until use. Thylakoids exhibited rates of gramicidin-(final concentration 5 tLg.ml-1) uncoupled electron transportof 340 ,tmol of O2-mg- Chlh- 1.
Electron transport was measured by changes in oxygenconcentration at 25°C in a temperature-regulated cuvette of aRank Brothers oxygen electrode. Light intensity at the surfaceof the cuvette was 1200 taE-m-2-s-1 [1 einstein (E) = 1 mol ofphotons]. The assay mixture contained, in 1.5 ml: 50 tag of Chl,50 mM Tricine, 50 mM NaCl, 3 mM MgCl2, and 5 mMNaH2PO4, and the pH was adjusted to 8.
Glycolate Excretion by Whole Cells or by Isolated Chloro-plasts. Five milliliters of high-CO2-grown Dunaliella cell sus-pensions (containing 100 ,tg of Chl) in 10 mM sodiumphosphate buffer with 170 mM NaCl (pH 7) was stirred andilluminated (800 ptE.m-2.s-1). One millimolar NaHCO3 wasadded at zero time and every 10 min thereafter. For chloro-plasts, 5 ml of chloroplast suspension [40 jig Chl.ml-~ inKCl/Hepes buffer (15) in which sorbitol was replaced with 175mM KC1] with 3-phosphoglyceric acid (1 mM) was placed in a25-ml beaker in a water bath at 25°C and illuminated (400iE-m-2.s-1) with an incandescent 300-W reflector lamp. After10 min of preillumination a 0.5-ml sample was withdrawn aszero time and 0.5 mM NaHCO3 was added; this procedure wasrepeated at every 10 min thereafter. Aliquots (1 ml) of cell orchloroplast suspensions were withdrawn every 15 min and,after centrifugation, glycolate was measured in the superna-tant by the Calkins method (17).
Methylviologen (MV)- and Light-Dependent 02 Uptake.Intact chloroplasts containing 50 ,tg of Chl were resuspendedin 2 ml of assay buffer (50 mM Hepes/7 mM MgCl2/7 mMsodium phosphate, pH 7.5) in the 02 electrode, so that theywere immediately broken by osmotic shock. To inhibit cata-lase, 1 mM KCN was added. Because it was not possible toprepare high-quality washed thylakoids from Dunaliella, os-motically shocked chloroplasts were.used. MV (0.2 mM) wasadded to the buffer after the addition of chloroplasts. Afterincubation in the dark for 2 min, the light was turned on and theMV-dependent 02 uptake was measured for 2-3 min, before 1,tM 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU) wasadded, which immediately inhibited 02 uptake due to inhibitionof electron flow from PS II to the plastoquinone pool. Twominutes after DCMU addition, 3 mM glycolate or other organicacid was added. 02 uptake resumed immediately (see Fig. 1). Thesubstrate-dependent oxygen uptake was followed for at least 5min, during which time inhibitors could be used to block electrontransfer to MV. SHAM (2.5-5.0 mM) was used to inhibit aproposed "glycolate-quinone oxidoreductase reaction"; 2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone (DBMIB)(10-20 ,tM), to inhibit flow of electrons from plastoquinone tothe cytochrome (Cyt) b6f complex; and 2 juM carbonylcyanide4-trifluoromethoxyphenylhydrazone (FCCP) or 20 ,AM carbon-
ylcyanide 3-chlorophenylhydrazone (CCCP), to uncouple theelectron transport. Uncouplers (FCCP or CCCP) stimulated (upto 40%) substrate-independent or endogenous flow of electronsbut not the organic acid-dependent 02 uptake (results notshown). Fresh stock solutions of DBMIB (1 mM) and SHAM(500 mM) were prepared daily in 95% ethanol. The maximum PSI noncyclic electron transport as MV-dependent 02 uptake with1 mM ascorbate as the electron donor and 100 tuM DCIP was 160tamol of 02-mg-1 Chl-h-1.
Light- and Glycolate-Dependent Reduction of DCIP orFerricyanide. Glycolate-dependent reduction of DCIP wasassayed anaerobically (18). The reaction mixture (2.5 ml)contained 100 mM potassium phosphate buffer (pH 8.3), 150LiM DCIP, chloroplasts containing 100 ,tg of Chl, and appro-priate inhibitors in a Thunburg cuvette with 5 mM glycolate inthe side arm. The reaction was incubated for 30 min and acontrol was run in parallel without added glycolate. After-wards, a 1-ml aliquot was centrifuged to remove thylakoids andA600 of the supernatant was measured for the remaining DCIP(E of 21.9 M-1 cm-1 at pH 8.3). The AA was multiplied by afactor of 3.95 for maximum activity, if a saturating dyeconcentration could have been used (18). SHAM (2.5 mM)was used to inhibit light-dependent glycolate oxidation, buthigher concentrations of SHAM caused nonenzymatic reduc-tion of DCIP.
Potassium ferricyanide reduction by glycolate in the lightwas assayed aerobically in a 2.5-ml mixture, containing 1.5 mMK3Fe(CN)6 in 50 mM sodium phosphate buffer (pH 8.3),chloroplasts containing 50 or 100 Ag of Chl, inhibitors, and 5mM glycolate. After 10 min, a 1-ml aliquot was centrifuged toremove thylakoids, and A420 of the supernatant was measured.Potassium ferricyanide (1.5 mM) gave an A420 of 1.5, whichdropped to 1.4 in a complete reaction mixture. An A420increase of 1.0 would be equivalent to the reduction of 1 timolof ferricyanide. Uncouplers (FCCP and CCCP) caused in-creased reduction of ferricyanide (results not shown).
Determination of Inorganic Phosphate and Glyoxylate.These were measured after the oxidation of glycolate orP-glycolate by osmotically shocked chloroplasts or thylakoidsfrom algae or spinach. The 2.1-ml reaction mixture contained50 mM Tricine (pH 8.0), 5 mM MgCl2, 5 mM NaCl, chloro-plasts or thylakoids containing 100-150 jtg of Chl, and 5 mMsubstrate. To prevent nonenzymatic decarboxylation of glyoxy-late by H202, 100 units of catalase was added to each assay.When catalase was omitted, the amount of glyoxylate produc-tion was underestimated by up to 50% after 20-30 min (datanot shown). Aliquots (250 ual) were withdrawn and centrifugedat 0, 5, 10, 15, and 30 min and were assayed immediately forinorganic phosphate with malachite green (19) and for glyoxy-late as the phenylhydrazone by increase in A324 (20).
RESULTSGlycolate excretion from the whole cells or chloroplasts of aunicellular green alga in the light was increased severalfold bythe presence of 2.5 mM SHAM (Fig. 1). SHAM has been usedas an inhibitor of an alternative respiration/quinone oxi-doreductase reaction in plant mitochondria (21). Conse-quently our working hypothesis has been that chloroplasticglycolate oxidation may be linked with the electron transportsystem by a glycolate-quinone oxidoreductase system that isalso SHAM inhibited (see Fig. 3). To further test this hypoth-esis for glycolate metabolism in the chloroplasts, MV, DCIP,and potassium ferricyanide were used as electron acceptors,along with DCMU, to block electron transport from PS II toPS I.
Effect of Glycolate on the MV-Coupled 02 Uptake andInhibition by SHAM and DBMIB. The flow of electronsoriginating from the oxidation of glycolate or other organicacids was coupled to MV, which accepts the electrons from PS
Proc. Natl. Acad. Sci. USA 93 (1996)
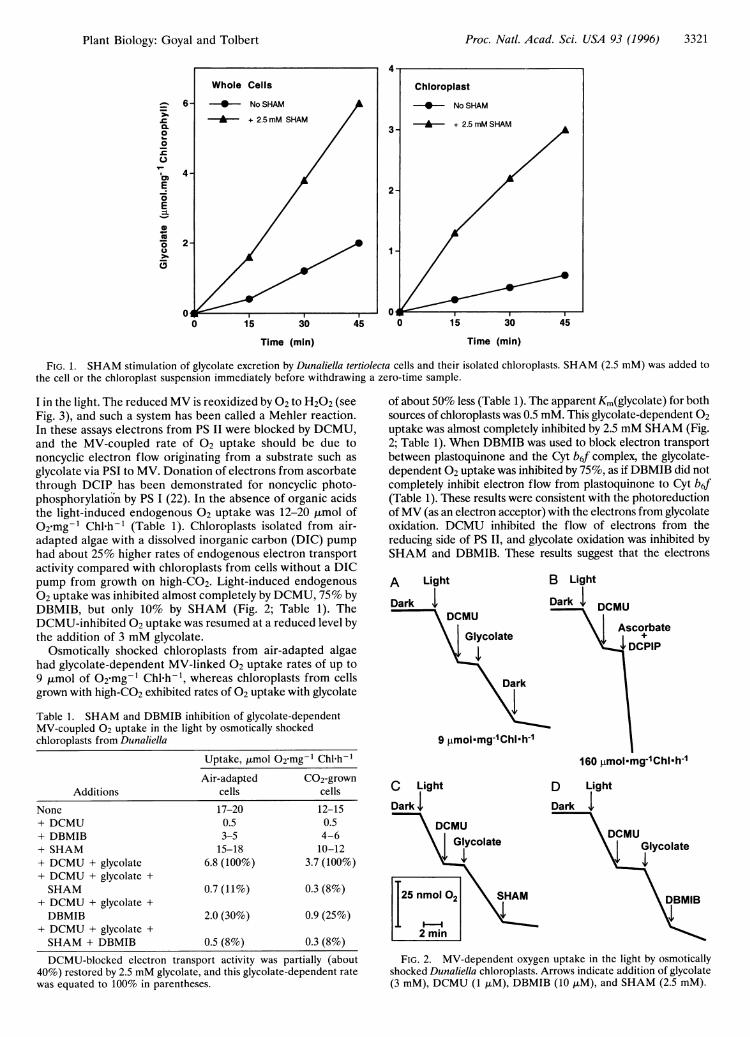
Proc. Natl. Acad. Sci. USA 93 (1996) 3321
-_--- No SHAM --- No SHAM
0. 3+ 2.5 mM SHAM + 2.5 mM SHAM
0 1O) 4-
E
0 15 30 45 0 1'5 3'0 45
Time (min) Time (min)
FIG. 1. SHAM stimulation of glycolate excretion by Dunaliella tertiolecta cells and their isolated chloroplasts. SHAM (2.5 mM) was added tothe cell or the chloroplast suspension immediately before withdrawing a zero-time sample.
I in the light. The reduced MV is reoxidized by 02 to H202 (seeFig. 3), and such a system has been called a Mehler reaction.In these assays electrons from PS II were blocked by DCMU,and the MV-coupled rate of 02 uptake should be due tononcyclic electron flow originating from a substrate such asglycolate via PSI to MV. Donation of electrons from ascorbatethrough DCIP has been demonstrated for noncyclic photo-phosphorylation by PS I (22). In the absence of organic acidsthe light-induced endogenous 02 uptake was 12-20 gtmol ofO2.mg-1 Chl-h-1 (Table 1). Chloroplasts isolated from air-adapted algae with a dissolved inorganic carbon (DIC) pumphad about 25% higher rates of endogenous electron transportactivity compared with chloroplasts from cells without a DICpump from growth on high-CO2. Light-induced endogenous02 uptake was inhibited almost completely by DCMU, 75% byDBMIB, but only 10% by SHAM (Fig. 2; Table 1). TheDCMU-inhibited 02 uptake was resumed at a reduced level bythe addition of 3 mM glycolate.
Osmotically shocked chloroplasts from air-adapted algaehad glycolate-dependent MV-linked 02 uptake rates of up to9 ,umol of O2.mg-1 Chl-h-1, whereas chloroplasts from cellsgrown with high-CO2 exhibited rates of 02 uptake with glycolate
Table 1. SHAM and DBMIB inhibition of glycolate-dependentMV-coupled 02 uptake in the light by osmotically shockedchloroplasts from Dunaliella
Uptake, tamol 02-mg-1 Chl-h-1
Air-adapted C02-grownAdditions cells cells
None 17-20 12-15+ DCMU 0.5 0.5+ DBMIB 3-5 4-6+ SHAM 15-18 10-12+ DCMU + glycolate 6.8 (100%) 3.7 (100%)+ DCMU + glycolate +SHAM 0.7 (11%) 0.3 (8%)
+ DCMU + glycolate +DBMIB 2.0 (30%) 0.9 (25%)
+ DCMU + glycolate +SHAM + DBMIB 0.5 (8%) 0.3 (8%)DCMU-blocked electron transport activity was partially (about
40%) restored by 2.5 mM glycolate, and this glycolate-dependent ratewas equated to 100% in parentheses.
of about 50% less (Table 1). The apparent Km(glycolate) for bothsources of chloroplasts was 0.5 mM. This glycolate-dependent 02uptake was almost completely inhibited by 2.5 mM SHAM (Fig.2; Table 1). When DBMIB was used to block electron transportbetween plastoquinone and the Cyt b6f complex, the glycolate-dependent 02 uptake was inhibited by 75%, as ifDBMIB did notcompletely inhibit electron flow from plastoquinone to Cyt b6f(Table 1). These results were consistent with the photoreductionofMV (as an electron acceptor) with the electrons from glycolateoxidation. DCMU inhibited the flow of electrons from thereducing side of PS II, and glycolate oxidation was inhibited bySHAM and DBMIB. These results suggest that the electrons
A Light B Light
Dark [ Dark I DCMU\ DCMU \DCU\|\ Ascorbate
Glycolate +
DCPIP
4 Dark
9 tmol-mg-1Chl-h-1160 gmol-mg-1Chl-h-1
C Light D LightDarkj Dark
Glycolate DCMUolateGlycolate
25 nmol 02 SHAM BMIB
2 min
FIG. 2. MV-dependent oxygen uptake in the light by osmoticallyshocked Dunaliella chloroplasts. Arrows indicate addition of glycolate(3 mM), DCMU (1 ,tM), DBMIB (10 ,tM), and SHAM (2.5 mM).
Plant Biology: Goyal and Tolbert

3322 Plant Biology: Goyal and Tolbert
from glycolate oxidation were donated to the electron transportchain at a site between the primary acceptor of PS II and Cyt bf.Since SHAM did not inhibit electron transport between PS II andPS I, electrons from glycolate oxidation apparently did not godirectly through the plastoquinone pool. We propose that thepossible site which accepts electrons from glycolate is a "SHAM-sensitive glycolate-quinone oxidoreductase system" that is linkedwith the electron transport chain as shown in Fig. 3. In boiledchloroplast preparations, no glycolate-dependent 02 uptake was
observed, as if electrons from glycolate oxidation were linked tothe electron transport chain by a heat-inactivatable enzyme.
Eight other organic acids (acetate, citrate, formate, D- andL-lactate, malate, oxaloacetate, and succinate) were much lesseffective as electron donors to the noncyclic electron transport.Similar low levels of succinate stimulation of chlororespirationhave been reported (23).
Light- and Glycolate-Dependent Reduction of DCIP andFerricyanide. Osmotically shocked Dunaliella chloroplasts re-
duced DCIP anaerobically in the presence of glycolate andlight at a rate of about 6 tLmol'mg- Chlh- 1 (Table 2). Becausethis reaction was light dependent, the DCIP should have beenacting as a Hill acceptor from PS I during glycolate oxidation.DCMU did not inhibit glycolate- and light-dependent reduc-tion of DCIP, suggesting that PS II was not involved and thatelectrons from glycolate oxidation entered the electron trans-port chain after the DCMU inhibition site at the quencher,"Q." SHAM (2.5 mM) inhibited glycolate-dependent reduc-tion of DCIP. SHAM alone at this concentration did notinhibit electron transport. SHAM inhibition of a light-dependent glycolate oxidation by thylakoids suggests that it islikely that a quinone-oxidoreductase type of oxidation systemis present which can feed electrons to PS I.
Using the DCIP assay for glycolate-oxidizing enzymes, our
group and others have reported an FMN-linked glycolateoxidase in the peroxisomes (24, 25) and an alternate form thatwas thought to be linked to a quinone (26) or to ferridoxin (27).The latter form had been purified from etiolated tissue whichcontained little activity of the flavin-linked form (28). Thequinone form of glycolate oxidase seems to have propertiesmore like the chloroplastic glycolate-oxidizing system that isdescribed in this paper.
In osmotically shocked chloroplasts, ferricyanide as a Hillreagent can accept electrons in the vicinity of plastoquinone as
well as on the reducing side of PS I. The light- and glycolate-dependent rate of ferricyanide reduction was 27 + 8/tmol-mg-1 Chl-h-1 (Table 2), and this rate was inhibited(90%) by DCMU. The rate of ferricyanide reduction withglycolate was about 4-fold higher than the measured rate ofDCIP reduction. An increase of 2-fold could be expectedbecause two electrons are required per DPIP, compared withone electron in the case of ferricyanide. Possibly the DCIPassay is an underestimation from reoxidation of the reducedDCIP by the electron transport chain. Similar to Hill activity,
glycolateSHAM,I glyoxylate MV H202
Q QH2 MVH2 02DCMU PQQ
DBMIB Cyt fPC
FIG. 3. Proposed scheme for the light-dependent glycolate oxida-tion by the chloroplast thylakoids. "Q," primary electron acceptor(quencher); Q, quinone; PQ, plastoquinone; PC, plastocyanin.
Table 2. Anaerobic DCIP reduction and aerobic ferricyanidereduction by osmotically shocked chloroplasts with glycolate
Reduction
,umol.mg-1Treatment Additions Acceptor Chlh- 1
Light Glycolate DCIP 5.6 + 1.3Dark Glycolate DCIP 1.2 ± 0.7Light Glycolate + DCMU DCIP 4.7 + 0.6Dark Glycolate + DCMU DCIP 0.9 ± 0.5Light Glycolate + SHAM DCIP 1.7 + 0.9Dark Glycolate + SHAM DCIP 1.1 + 0.6Light None K3Fe(CN)6 83 + 5Light Glycolate K3Fe(CN)6 110 + 8
Light- and glycolate-dependent reduction 27 ± 8
Values are mean of at least six independent assays. DCIP experi-ments could not be run aerobically because reduced DCIP was rapidlyoxidized.
the glycolate-dependent reduction of ferricyanide decreasedas the age of isolated chloroplasts increased (data not shown).
Substrate-Dependent 02 Uptake in the Light by Osmoti-cally Shocked Chloroplasts or Thylakoids from SpinachLeaves. These chloroplasts without MV had little or no 02uptake. Upon addition of MV and 3 mM glycolate or P-glycolate, in the light, spinach chloroplasts had rates of 02uptake of about 22 ,Lmol-mg-1 Chlh-l. The apparent Km(glycolate or P-glycolate) was 0.5 mM (data not shown). Thesubstrate-dependent 02 uptake ceased in the dark. Glyoxylate(1 mM) exhibited only one-half the rate of 02 uptake, andhigher concentrations promoted less 02 uptake. L-Lactate (3mM), a substrate analog of glycolate, also resulted in 02uptake at about half the rate as with glycolate. Other organicacids (acetate, citrate, formate, D-lactate, oxaloacetate, andsuccinate) were not effective (results not shown). When cata-lase was added to the reaction after a few min of 02 uptakewith glycolate, half the amount of 02 uptake was released, andthe renewed rate of 02 uptake was half of that in the absenceof catalase. These results suggest that the 02 uptake was dueto the formation of H202 as in a Mehler reaction.
Electron transport uncouplers (CCCP, FCCP, and NH4C1)did not increase the rate of glycolate-dependent 02 uptake.AOA, hydroxylamine, SHAM, glyoxylate, glycolate, and P-glycolate do not uncouple electron transport as do NH4C1 andgramicidin. Hydroxypyridinemethanesulfonate (HPMS), aninhibitor of peroxisomal glycolate oxidase in higher plants, didnot inhibit glycolate or P-glycolate metabolism by chloroplastsor thylakoids. These results indicate that it is unlikely that thespecific light- and glycolate-dependent 02 uptake by spinachthylakoids was due to peroxisomal glycolate oxidase or algalmitochondrial glycolate dehydrogenase. The latter are notlight dependent and are inhibited by the sulfonate.Because thylakoids from spinach had a very high rate of 02
uptake in the presence of light, MV, and gramicidin, it wasdifficult to measure accurately increases in 02 uptake due toglycolate or P-glycolate, within the limitation of the 02 elec-trode and chart recorder. To observe substrate-dependentoxidation, 02 uptake was measured without gramicidin. Thy-lakoids metabolized glycolate at a rate of about 22 ,gmol of02-mg-1 Chl.h-1, and the rate of P-glycolate oxidation wasabout half the rate with glycolate. Two washings of thethylakoids should have removed nearly all the P-glycolatephosphatase of the stroma, yet P-glycolate was oxidized rapidlywithout a time lag to hydrolyze it to glycolate, whose Km was0.5 mM. Glyoxylate (1 mM) inhibited the endogenous rate of02 uptake by 30%. Manometric observations (N.E.T., unpub-lished results) have also revealed high rates of oxygen uptakein the light by isolated chloroplasts from spinach leaves with
Proc. Natl. Acad. Sci. USA 93 (1996)

Proc. Natl. Acad. Sci. USA 93 (1996) 3323
either P-glycolate or glycolate without a lag for possiblehydrolysis of P-glycolate.
Products of Glycolate and P-Glycolate Oxidation. Osmoti-cally shocked chloroplasts from both spinach and Dunaliellaformed glyoxylate from either glycolate or P-glycolate in thelight, but the rate of glyoxylate formation from P-glycolate wasabout 40% less than from the glycolate (Table 3). In the darkthese chloroplasts formed 10-15% as much glyoxylate as in thelight. Glycolate and P-glycolate oxidation was light dependent;therefore their oxidation should not have been by contami-nating peroxisomal glycolate oxidase or the mitochondrialdehydrogenase. With P-glycolate as a substrate, phosphate wasreleased rapidly in the dark and the light (Table 4), but the rateof phosphate release was higher in the light. Broken spinachchloroplasts converted P-glycolate to phosphate and glyoxylateat twice the rate as Dunaliella chloroplasts. The phosphaterelease from P-glycolate in the light may have been due to anoxidative dephosphorylation process.
DISCUSSIONEarlier work on glycolate metabolism in isolated chloroplastswas summarized by Gibbs (7), who showed an increase in 14Cfound in glycolate during 14CO2 fixation with increasing 02and/or decreasing CO2. Such data are consistent with glyco-late synthesis by the oxygenase activity of Rubisco (4) and with02 inhibition of CO2 fixation (Warburg effect) and photore-spiration (1, 6). The earlier reports probably underestimatedglycolate production, since they did not measure total glycolateformation or the formation of unlabeled glycolate from unla-beled carbohydrate reserves, especially in the presence of lowlevels of 14CO2. and high levels of 02. Gibbs (7) proposed ascheme for chloroplastic synthesis of glycolate, but his reportdid not consider the oxidation of glycolate by the chloroplasts.Rather, all the glycolate was thought to have been excreted bythe chloroplasts and oxidized in the peroxisomes (4). Ourpaper proposes that some of the glycolate is oxidized in thechloroplasts and that only excess glycolate may be excreted foroxidation in the peroxisomes (4) or mitochondria (29).
This paper demonstrates the existence of a SHAM-inhibited, light-dependent glycolate-quinone oxidoreductasesystem that is associated with thylakoid membranes of thechloroplasts (Fig. 3). DBMIB, which is a plastoquinone an-tagonist preventing transfer of electrons from plastoquinoneto Cyt b6f, also inhibited glycolate oxidation. Because chloro-plasts were able to oxidize glycolate in the presence ofDCMU,but not in the presence of DBMIB, electron donation isproposed to be in the plastoquinone region of the photosyn-thetic electron transport chain. Minor oxygen uptake in thepresence ofDBMIB may be due to its photoreduction, as it canbe reduced by chloroplasts to produce H202 (30). SHAM is aninhibitor of alternative respiration or a quinone-oxidoreduc-tase reaction in the plant mitochondria (21). SHAM alsospecifically increases the excretion of glycolate by the cells and
Table 3. Glyoxylate production from glycolate or P-glycolate byisolated osmotically shocked chloroplasts from spinachand Dunaliella
Glyoxylate produced,tLmol.mg-1 Chl.h-1
Substrate Treatment Spinach Dunaliella
Glycolate Light 17 6P-glycolate Light 11 4Glycolate Dark 2 1P-glycolate Dark 2 1
Table 4. Release of inorganic phosphate from 5 mM P-glycolatePi released, tamolmg- 1 Chlh- 1
Spinach Spinach DunaliellaTreatment chloroplasts thylakoids chloroplasts
Light 54 49 18Dark 36 47 11
The chloroplasts from spinach or Dunaliella were osmoticallyshocked. Thylakoids from spinach were washed twice, but thylakoidsfrom Dunaliella could not be prepared.
chloroplasts (Fig. 1; ref. 13). Thus stimulated glycolate excre-tion by chloroplasts would seem to be due to SHAM inhibitionof this chloroplastic glycolate metabolism. This appears to bea side-reaction from the complete photosynthetic electrontransport system, because SHAM did not inhibit 02 evolutionor CO2 fixation by algae or isolated chloroplasts. Heat treat-ment of thylakoids diminished this activity, suggesting a systeminvolving a proteinaceous component. This component seemsto be tightly bound to the thylakoids, as activity for glycolateor P-glycolate oxidation remained after two washings. Theglycolate-oxidizing system is designated as a "SHAM-sensitiveglycolate-quinone oxidoreductase" in the thylakoids betweenPS II and PS I. This site is likely to be a special quinone passingelectrons in the region of plastoquinone.
In the last decade, growing evidence has been presented forchlororespiration (23, 31), although its terminal oxidase hasnot been identified. By using chlorophyll fluorescence kinetics,Bennoun (31) also demonstrated SHAM inhibition of chlo-rorespiration. Though chlororespiration is considered to be aprocess distinct from photorespiration or the Mehler reaction,it is conceptually similar to oxidation of glycolate by thethylakoid membranes. Treatment of a chloroplast suspensionwith 5 iJM antimycin a, an inhibitor of mitochondrial respira-tion, had no effect on light-induced 02 uptake with glycolate.Although succinate may donate electrons to the photosyn-thetic electron transport chain of Chlamydomonas duringchlororespiration, light inhibited this oxidation (23). However,the glycolate-oxidizing system in the chloroplast is dependenton light. Therefore the proposed SHAM-sensitive glycolate-quinone oxidoreductase system is different from chlororespi-ration. In addition, neither mitochondrial glycolate dehydro-genase nor chlororespiration is inhibited by SHAM.
In the complete photosynthetic electron transport process,electrons come from the quencher of PS II after the photolysisof water. During noncyclic photophosphorylation in vivo,ferredoxin and NADP should be reduced by PS I with electronscoming from the oxidation of substrates such as glycolate. If PSI is coupled to oxygen uptake in a Mehler reaction or someother Hill acceptor such as ferric salts, it could generate ATPbut not reduce NADPH. Noncyclic phosphorylation by PS Ishould generate two ATP molecules per glycolate oxidized.This type of noncyclic phosphorylation occurs in anaerobicphotosynthetic bacteria or algae without PS II (32). In thisconcept, electrons from the reduced end of PS I are cycledback through Cyt f This cyclization has been postulated toinvolve a NADPH-ascorbate reductase (22). If glycolate oxi-dation were the electron donor instead, the glyoxylate productcould be reduced back to glycolate by the NADPH-glyoxylatereductase in the chloroplast (33). The combination of bothreactions creates a cyclic photophosphorylation system forATP synthesis from glycolate oxidation without NADPHaccumulation. Since CO2 fixation requires more ATP (18molecules per 6 CO2 molecules) than NADPH (12 moleculesper 6 CO2 molecules), abundant glycolate and an active PS Ioxidation system for it would be an essential part of photo-synthetic carbon metabolism. This would justify the oxygenaseactivity of Rubisco. These anaerobic reactions may haveevolved early in photosynthesis, as did Rubisco, but the process
The assay mixture contained 50 mM Tricine (pH 8.0), 5 mM MgC12,100 units of catalase, and chloroplasts equivalent to 100 gg ofChl-ml-1.
Plant Biology: Goyal and Tolbert

3324 Plant Biology: Goyal and Tolbert
would have required internal 02 from PS II for P-glycolatesynthesis.The proposed scheme in Fig. 3 explains how glycolate
oxidation by the thylakoids may be a regulatory process forcyclic photophosphorylation linked to photosynthetic carbonmetabolism. The Km(glycolate) for the glycolate-quinone ox-idoreductase system in the chloroplast is 0.5 mM. If thisoccurred in plant chloroplasts, it should promote more cyclicphosphorylation for the extra ATP for fixing CO2. Glycolatemetabolism by PS I for cyclic photophosphorylation could alsoprovide ATP for the DIC-algal pump (14, 34) and explain whythe pump is inhibited by SHAM or darkness (14). SHAM, aquinone inhibitor, blocks glycolate metabolism by isolatedchloroplasts and stimulates glycolate excretion (Fig. 1; ref. 13),and thus SHAM seems to reduce the total ATP synthesis forthe DIC pump. When the external CO2 concentration is low,increased oxygenase activity of Rubisco would form moreglycolate and, to restore the internal CO2 level, the CO2 pumpwould become more active with ATP generated by glycolateoxidation by PS I.The presence of a glycolate-oxidizing system associated with
the photosynthetic electron transport in the chloroplast may bea way nature can efficiently recycle carbon from glycolate whileregenerating energy under extreme environments. It makesuse of the glycolate, which is inevitably synthesized, by gen-erating ATP for CO2 fixation. Under low-CO2 conditions itmay also result in survival of the organism by maintaining somebasal rates of metabolism from the utilization of sugar reservesfor the regeneration of products of the C3 cycle.Although this research was started with Dunaliella chloro-
plasts because these cells do not contain a peroxisomal gly-colate oxidase, it was extended to spinach chloroplasts, whichcan also metabolize glycolate and phosphoglycolate in the lightby using the electron transport system. The oxidation ofglycolate by chloroplasts or thylakoids from spinach leavescannot be due to contaminating peroxisomal glycolate oxidase,which is not light dependent. Thus we have presented evidencefor a light-dependent glycolate oxidation, measured as 02uptake by the Mehler reaction, via the PS I electron transportchain of chloroplasts from either Dunaliella or spinach leaves.
We thank John Hosler and Charles F. Yocum for discussions duringthis investigation.
1. Tolbert, N. E. (1994) in Regulation ofAtmospheric C02 and 02 byPhotosynthetic Carbon Metabolism, eds. Tolbert, N. E. & Preiss,J. (Oxford Univ. Press, New York), pp. 8-33.
2. Nelson, E. B. & Tolbert, N. E. (1970) Arch. Biochem. Biophys.141, 102-110.
3.
4.
5.
6.
7.8.9.
10.11.12.
13.14.15.
16.17.18.19.
20.
21.
22.
23.
24.25.
26.
27.
28.
29.
30.
31.32.
33.34.
Frederick, S. E., Gruber, P. J. & Tolbert, N. E. (1973) PlantPhysiol. 52, 318-323.Husic, D. W., Husic, H. D. & Tolbert, N. E. (1987) Crit. Rev.Plant Sci. 5, 45-100.Tolbert, N. E., Harrison, M. & Selph, N. (1983) Plant Physiol. 72,1075-1083.Tolbert, N. E. (1992) in Phylogenetic Changes in Peroxisomes ofAlgae: Phylogeny of Plant Peroxisomes, ed. Stabenau, H. (Univ. ofOldenburg Press, Oldenburg, Germany), pp. 204-232.Gibbs, M. (1969) Ann. N. Y. Acad. Sci. 168, 356-368.Takabe, T. & Akazawa, T. (1981) Plant Physiol. 68, 1093-1097.Howitz, K. T. & McCarty, R. E. (1983) FEBS Lett. 254, 339-342.Tolbert, N. E. (1958) Brookhaven Symp. Biol. 11, 271-275.Murai, T. & Katoh, T. (1975) Plant Cell Physiol. 16, 789-797.Goyal, A. & Tolbert, N. E. (1992) in Phylogenetic Changes inPeroxisomes of Algae: Phylogeny of Plant Peroxisomes, ed.Stabenau, H. (Univ. of Oldenburg Press, Oldenburg, Germany),pp. 187-203.Goyal, A. & Tolbert, N. E. (1996) Plant Physiol. 96, S343.Goyal, A. & Tolbert, N. E. (1989) Plant Physiol. 89, 1264-1269.Goyal, A., Betsche, T. & Tolbert, N. E. (1988) Plant Physiol. 88,543-546.Cerovic, Z. G. & Plesnicar, M. (1984) Biochem. J. 223, 543-545.Calkins, V. P. (1943) Ind. Eng. Chem. 15, 762-764.Tolbert, N. E. (1971) Methods Enzymol. 23, 665-682.Lanzetta, P. A., Alvarez, L. J., Reinnach, P. S. & Candia, 0. A.(1975) Anal. Biochem. 100, 95-97.Baker, A. L. & Tolbert, N. E. (1967) Methods Enzymol. 9,338-342.Schonbaum, G. R., Bonner, W. D., Storey, B. T. & Bahr, J. T.(1971) Plant Physiol. 47, 124-128.Asada, K., Takahashi, M., Tanaka, K. & Nakano, Y. (1977) inBiochemical and Medical Aspects ofActive Oxygen, eds. Hayaishi,0. & Asada, K. (Japan Sci. Soc. Press, Tokyo), pp. 45-64.Willieford, K. O., Gombos, A. & Gibbs, M. (1989) Plant Physiol.90, 1084-1087.Tolbert, N. E. & Burris, R. H. (1950)J. Biol. Chem. 186, 791-804.Tolbert, N. E., Oeser, A., Kisaki, T., Hageman, R. H. &Yamazaki, R. K. (1968) J. Biol. Chem. 248, 5179-5184.Kolesnikov, P. A., Petrochenko, E. I. & Zore, S. V. (1958) Dokl.Akad. Nauk. S.S.S.R. 123, 729-732.Baker, A. L. & Tolbert, N. E. (1967) Biochim. Biophys. Acta 131,169-178.Kuczmak, P. A. & Tolbert, N. E. (1962) Plant Physiol. 37, 729-734.Beezley, B. B., Gruber, P. J. & Frederick, S. E. (1976) PlantPhysiol. 58, 315-319.Izawa, S. & Pan, R. L. (1978) Biochem. Biophys. Res. Commun.83, 1171-1177.Bennoun, P. (1982) Proc. Natl. Acad. Sci. USA 79, 4352-4356.Lascelles, J. (1973) Microbial Photosynthesis (Dowden, Hutchin-son, & Ross, Stroudsburg, PA).Zelitch, I. & Gotto, A. M. (1962) Biochem. J. 84, 541-546.Ogawa, T., Miyano, A. & Inoue, Y. (1985) Biochim. Biophys. Acta808, 77-84.
Proc. Natl. Acad. Sci. USA 93 (1996)





























