Embed Size (px)
Citation preview

The energetics of transport across membranes
Dr. Denice Bay
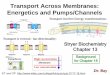
Transport across membranes
Passive Diffusion • 4 types of passive
diffusion: 1. Simple diffusion 2. Facilitated Diffusion 3. Filtration 4. Osmosis
Active Transport • 2 types of active
transport: 1. Primary ATP
driven 3. Secondary
electrochemical chemical gradient driven
H2O, CH3CH2OH, CH3Cl Ions, Metals, Large molecules
Energy membrane
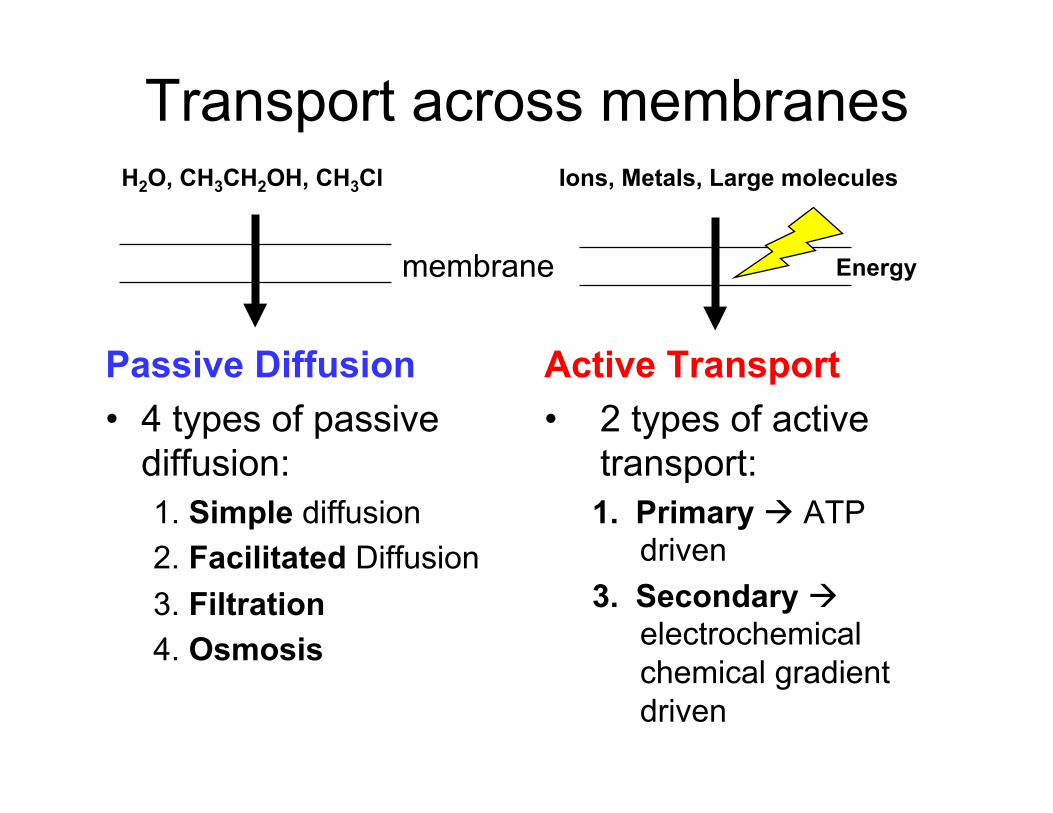
Passive Diffusion 1. Simple Diffusion
2. Facilitated Diffusion
- Solute flux is facilitated by transport through protein channels / pores
- Solute fluxed directly through the membrane
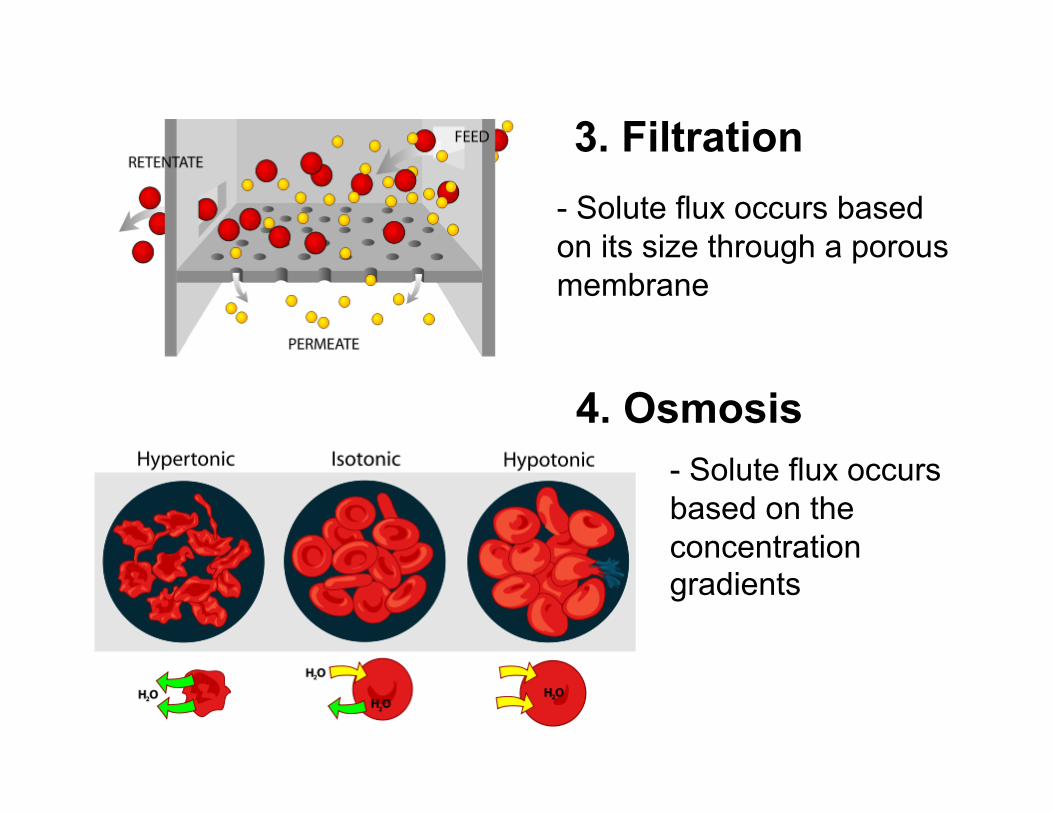
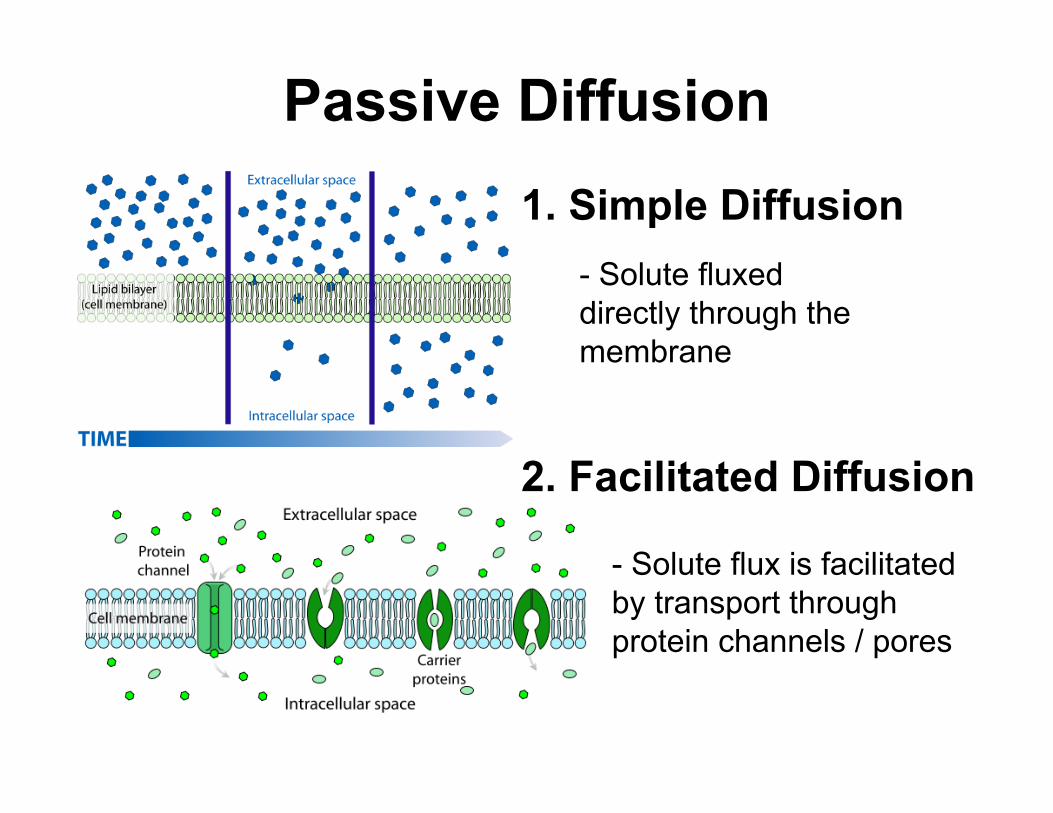
3. Filtration
4. Osmosis
- Solute flux occurs based on its size through a porous membrane
- Solute flux occurs based on the concentration gradients
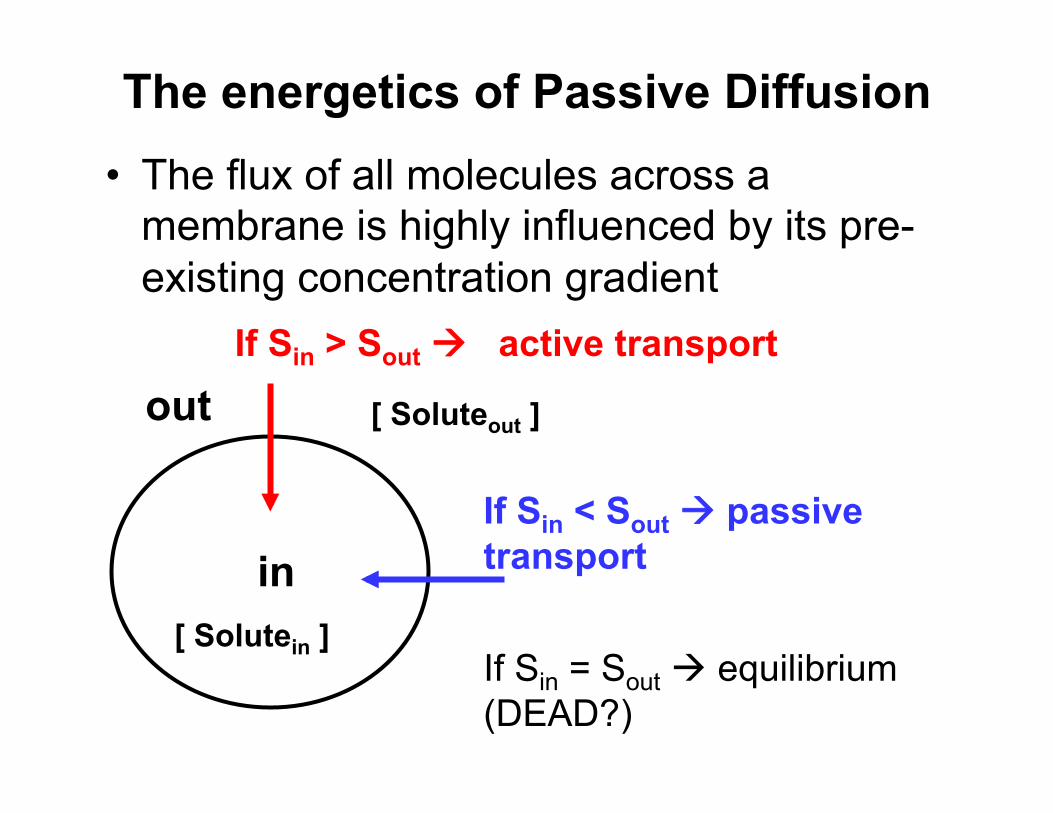
The energetics of Passive Diffusion • The flux of all molecules across a
membrane is highly influenced by its pre-existing concentration gradient
[ Solutein ]
out
in
[ Soluteout ]
If Sin > Sout active transport
If Sin < Sout passive transport
If Sin = Sout equilibrium (DEAD?)

Fick’s first law of passive diffusion Adolf Fick
(1829-1901) Fick’s laws of Diffusion from a biological membrane perspective:
J = - P • A • ΔC
J is the solute flux or the rate of change (dQ/ dt) in solute quantity (Q) over time (t) also represented as “J”
P is the permeability co-efficient of the membrane (m • sec-1) this value is negative due to solute movement towards low [Solute]
A is the surface area (m2) of the membrane where the solute flux is occuring
ΔC is the difference in the concentration across the membrane
(1)
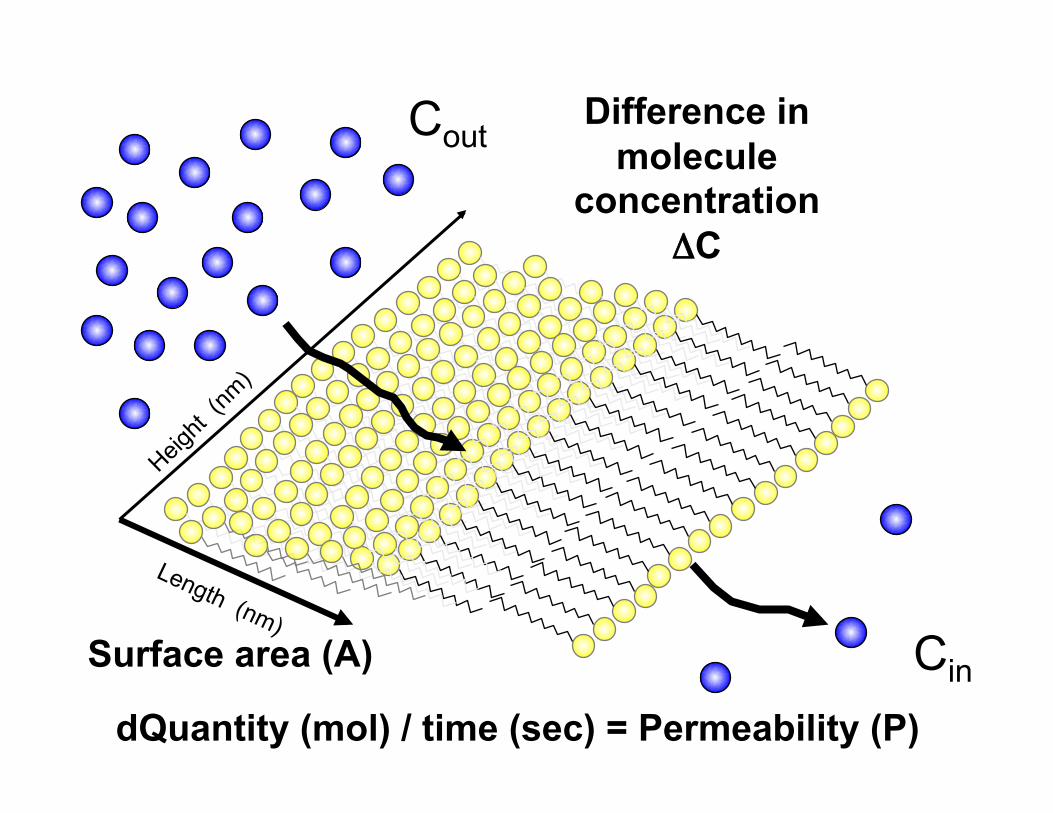
Surface area (A)
dQuantity (mol) / time (sec) = Permeability (P)
Difference in molecule
concentration ΔC
Cout
Cin

Thermodynamics of Passive Diffusion Fick’s law doesn’t allow a quantitative consideration of the energetic requirements that exist during transport.
For an uncharged solute the chemical potential (µ = free energy (G) per mole) can be represented by the following:
µ = chemical potential (G) of the solute µo = standard state chemical potential R = gas constant (8.3 J • mol-1 • deg-1) T = temperature degrees Kelvin (273 oK) lnC = log base e of [solute]
µ = µo + RT • lnC Where,
(2) Modified Nernst equation Walther Nernst
(1864-1941)

• Hence, the difference of µ (Δµ) between two points in space or (more importantly for biologists) two molecules in space separated by a membrane is given by:
• Since µoout = µo
in are equal the equation simplifies to:
Δµout-in = (µoout + RT• lnCout)-( µo
in + RT• lnCin) (3)
Δµout-in = RT•ln (Cout/Cin) (4)
This is an extention of both the van’t Hoff and Gibbs free energy equations.
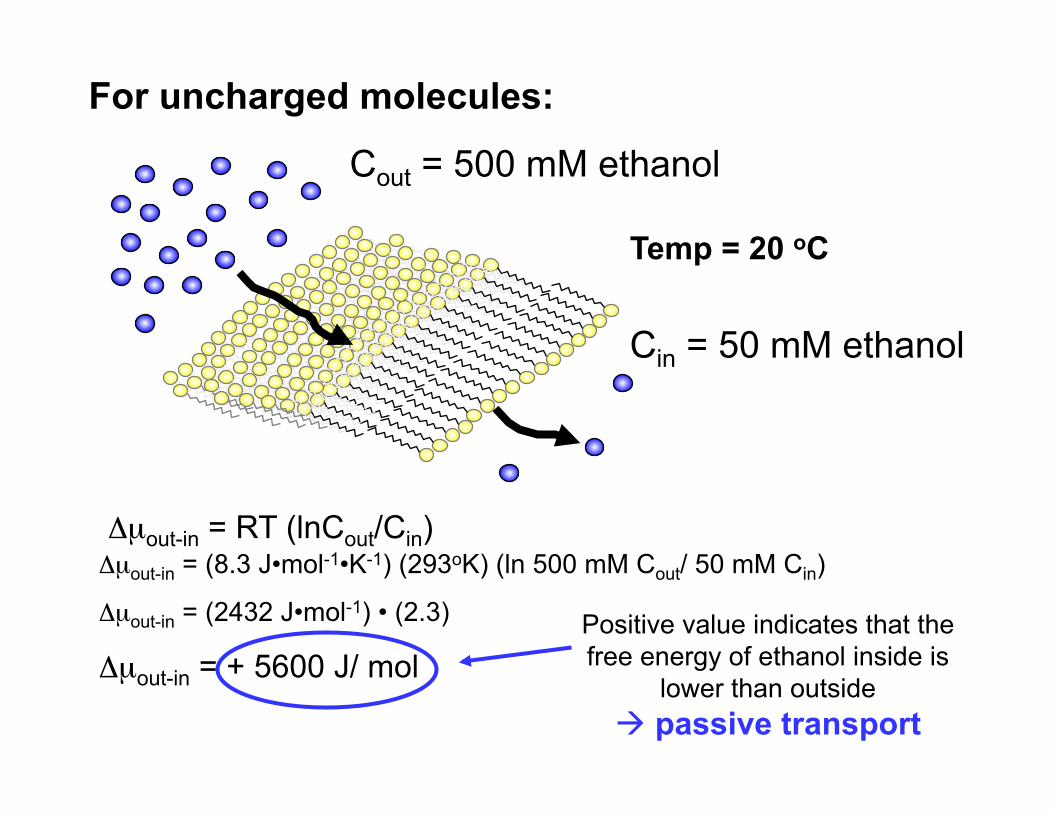
Cout = 500 mM ethanol
Cin = 50 mM ethanol
For uncharged molecules:
Temp = 20 oC
Δµout-in = RT (lnCout/Cin) Δµout-in = (8.3 J•mol-1•K-1) (293oK) (ln 500 mM Cout/ 50 mM Cin)
Δµout-in = (2432 J•mol-1) • (2.3)
Δµout-in = + 5600 J/ mol Positive value indicates that the free energy of ethanol inside is
lower than outside passive transport
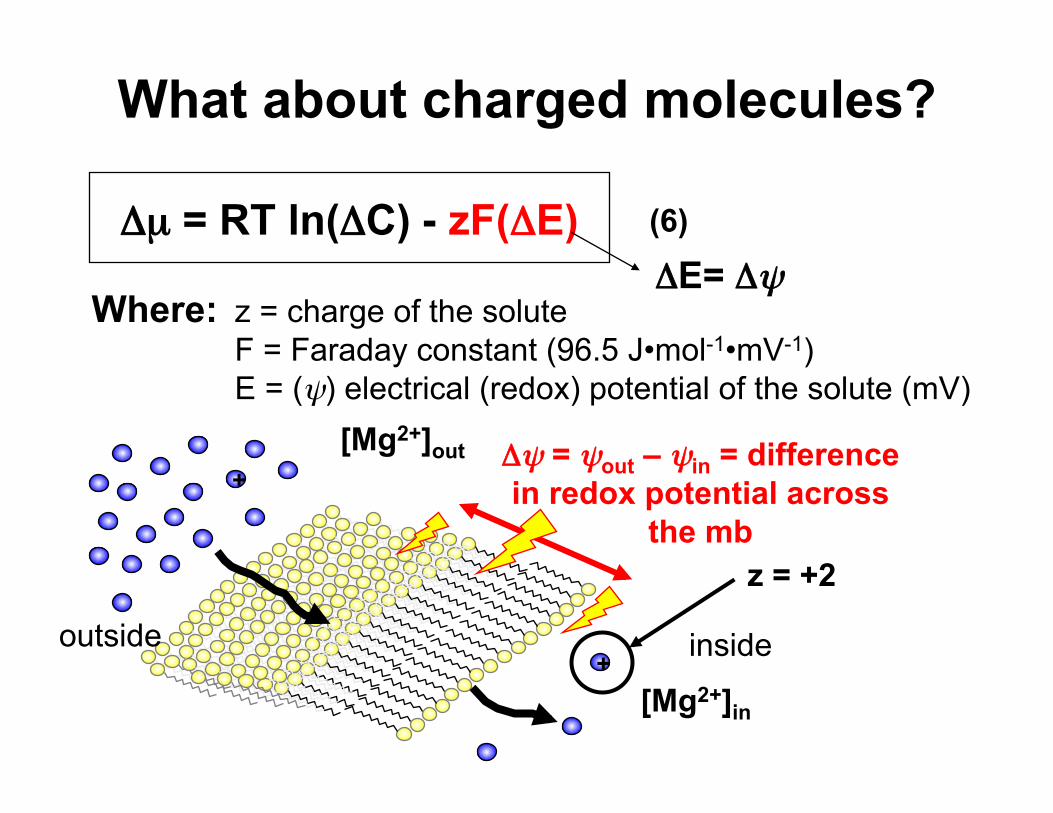
What about charged molecules?
Δµ = RT ln(ΔC) - zF(ΔE) (6)
Where: z = charge of the solute F = Faraday constant (96.5 J•mol-1•mV-1) E = (ψ) electrical (redox) potential of the solute (mV)
+
+ Δψ = ψout – ψin = difference in redox potential across
the mb z = +2
[Mg2+]out
[Mg2+]in
ΔE= Δψ
inside outside
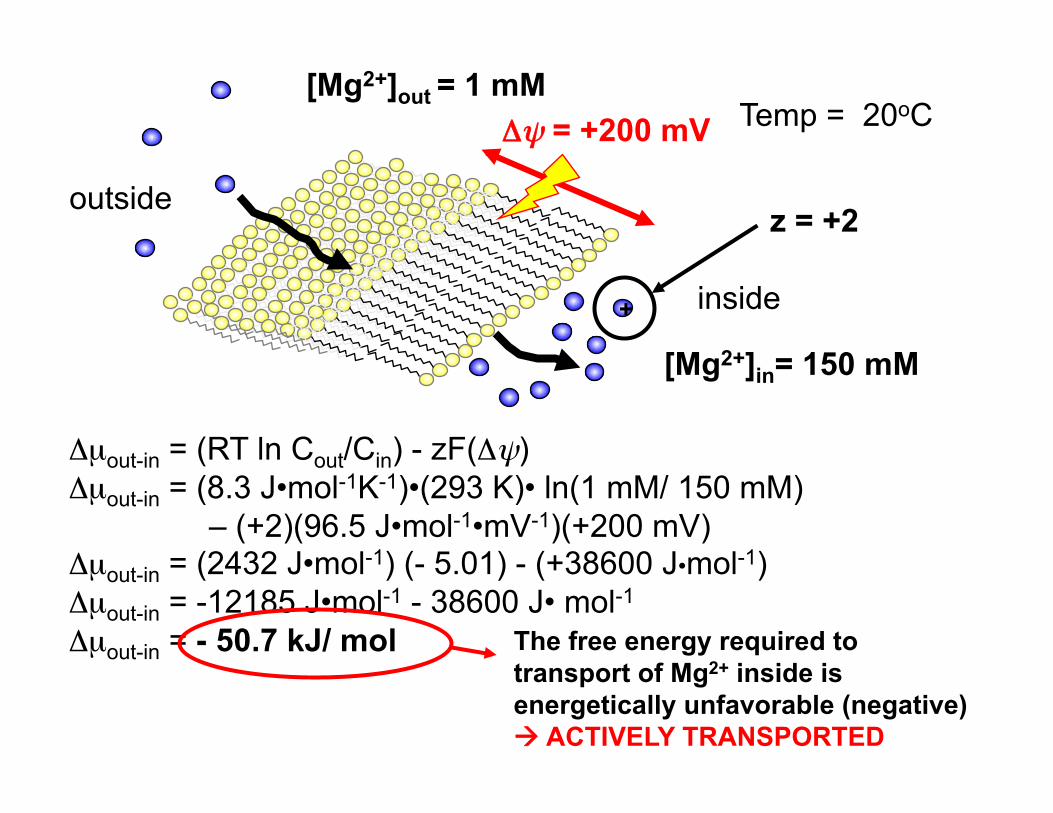
+
Δψ = +200 mV
z = +2
[Mg2+]out = 1 mM
[Mg2+]in= 150 mM
Δµout-in = (RT ln Cout/Cin) - zF(Δψ) Δµout-in = (8.3 J•mol-1K-1)•(293 K)• ln(1 mM/ 150 mM)
– (+2)(96.5 J•mol-1•mV-1)(+200 mV) Δµout-in = (2432 J•mol-1) (- 5.01) - (+38600 J•mol-1) Δµout-in = -12185 J•mol-1 - 38600 J• mol-1
Δµout-in = - 50.7 kJ/ mol The free energy required to transport of Mg2+ inside is energetically unfavorable (negative) ACTIVELY TRANSPORTED
Temp = 20oC
inside
outside

Thermodynamics of Osmosis • The diffusion of solutes across
semi-permeable membranes is also influenced by the pressure within the liquid solution across the membrane
• van’t Hoff determined that differences in [solute] flowed differently across the membrane when pressure varied Jacobus H. van 't Hoff
(1852-1911)
π = - c • RT (7)
c = [solute] (in M) ψπ = osmotic potential (atm or Mpa) R = gas constant (8.31 L kPa K-1 mol-1) T = temperature (Kelvin)
Note similarity to Pascal’s ideal gas law
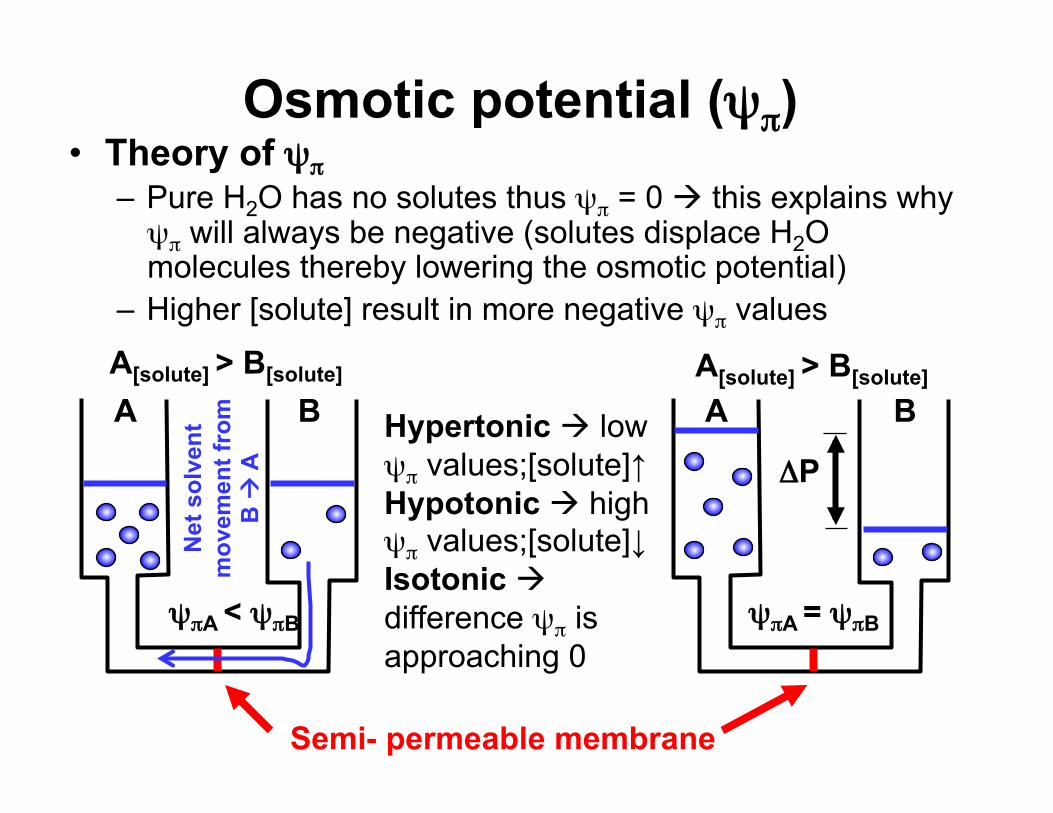
Osmotic potential (ψπ) • Theory of ψπ
– Pure H2O has no solutes thus ψπ = 0 this explains why ψπ will always be negative (solutes displace H2O molecules thereby lowering the osmotic potential)
– Higher [solute] result in more negative ψπ values
Semi- permeable membrane
A B B A
ΔP
A[solute] > B[solute]
ψπA = ψπB ψπA < ψπB
A[solute] > B[solute]
Hypertonic low ψπ values;[solute]↑ Hypotonic high ψπ values;[solute]↓ Isotonic difference ψπ is approaching 0
Net
sol
vent
m
ovem
ent f
rom
B
A

Water potential (Ψw) • Water potential (Ψw) is the algebraic sum of
pressure P and the osmotic potential (ψπ)
• Ψw is essentially describing the free energy (G) in a mass of water which relates how much energy is involved in its movement The Gwater is related in terms of pressure rather than
by J/mol simply based on convention
Ψw = P + ψπ
Ψw = G / Vw Where, Vw = 18 × 10−6 m3 mol−1
(8)
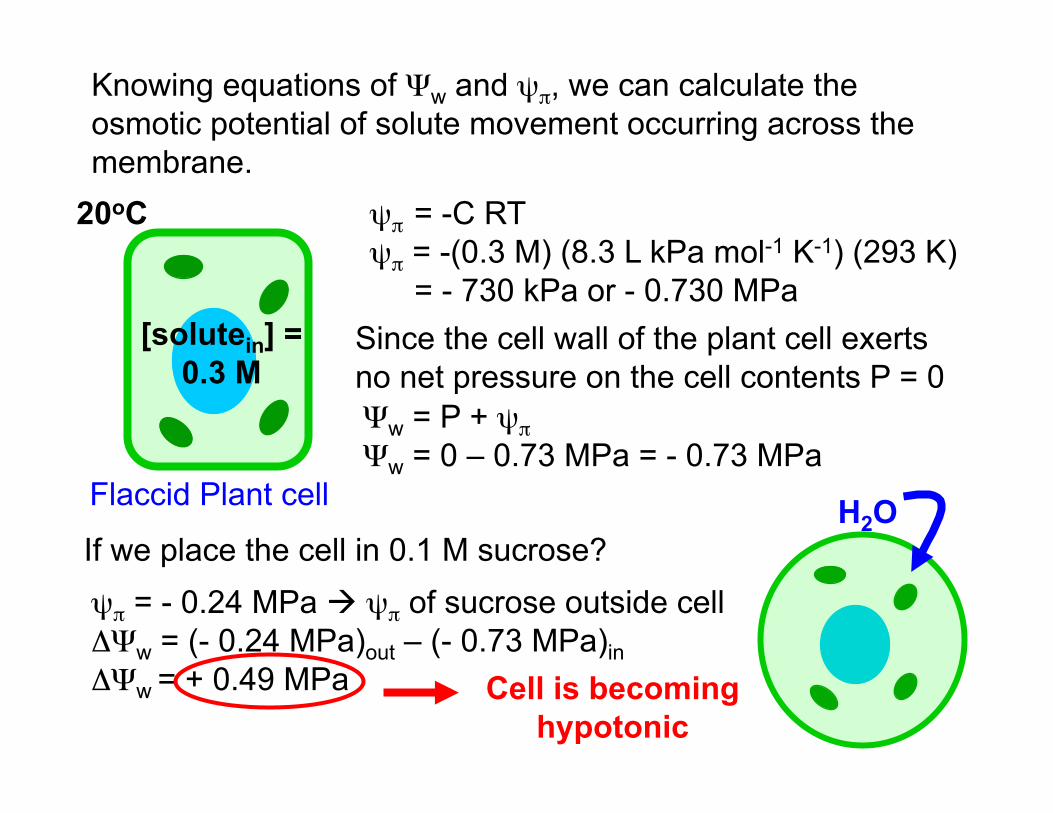
Knowing equations of Ψw and ψπ, we can calculate the osmotic potential of solute movement occurring across the membrane.
[solutein] = 0.3 M
Flaccid Plant cell
Ψw = P + ψπ Ψw = 0 – 0.73 MPa = - 0.73 MPa
ψπ = -C RT ψπ = -(0.3 M) (8.3 L kPa mol-1 K-1) (293 K) = - 730 kPa or - 0.730 MPa
20oC
Since the cell wall of the plant cell exerts no net pressure on the cell contents P = 0
If we place the cell in 0.1 M sucrose? ψπ = - 0.24 MPa ψπ of sucrose outside cell ΔΨw = (- 0.24 MPa)out – (- 0.73 MPa)in ΔΨw = + 0.49 MPa Cell is becoming
hypotonic
H2O

Summary of Passive Transport • 4 types of passive transport • Diffusion of any molecule across a membrane is
influenced by – [solute] inside and outside the membrane – the potential both chemical and electrical that resides
on the given membrane • The free energy required by the electrochemical
potential of the membrane will dictate the type of transport (passive or active) needed to transport it – Positive = passive diffusion – Negative = active transport

• Osmotic transport energetics are influenced by both solute concentration and solvent pressure that exist across the membrane – Osmotic potential (ψπ) is expressed as a unit
of pressure – The additive effect of both ψπ and pressure
(P) combine to give water potential (Ψw) which relates the free energy involvement in H2O movement across the membrane
– If ΔΨw > 0 hypotonic ΔΨw < 0 hypertonic ΔΨw = 0 isotonic
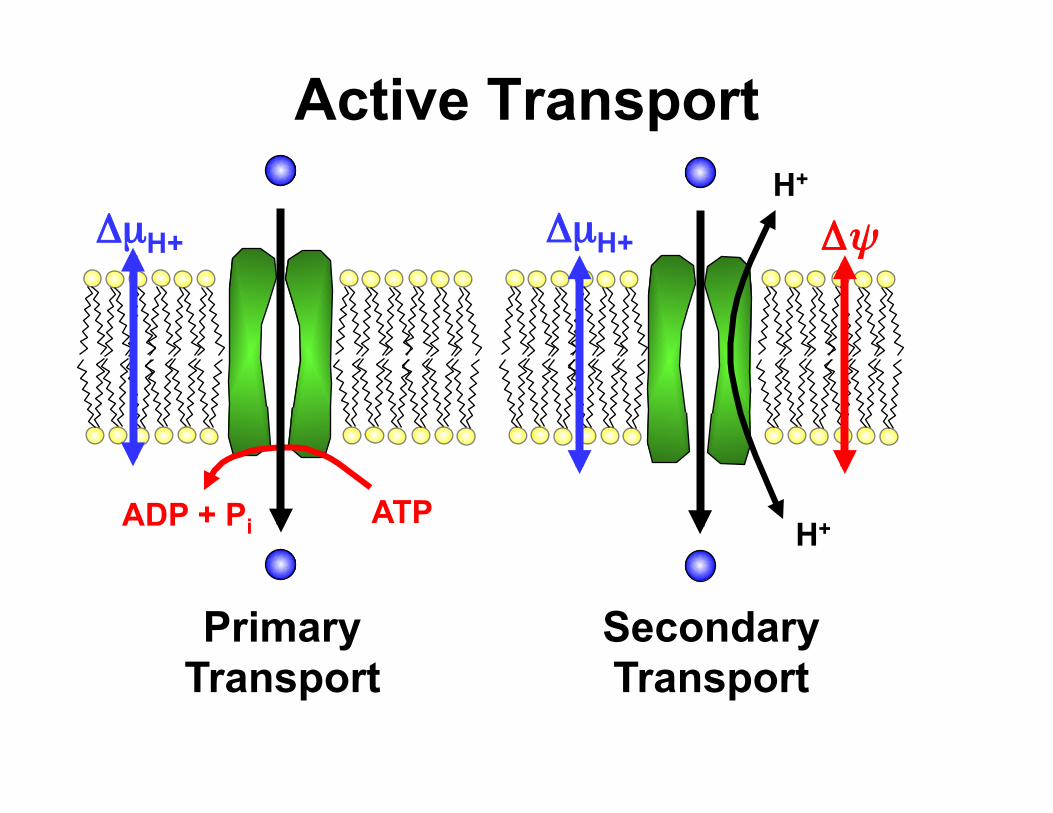
Active Transport
Primary Transport
Secondary Transport
ATP ADP + Pi
H+
H+
Δψ ΔµH+ ΔµH+

Primary (ATP driven) Transport • ATP hydrolysis provides energy for the
movement solutes across membranes under energetically unfavourable conditions
• ATP synthesis is linked to the utilization of the proton electrochemical gradient across the membrane
ATP + H2O + nH+n
ADP + Pi + nH+p
H+p = positive side of the membrane (µH+)
H+n = negative side of the membrane

ΔµATP = ΔGo + RT • lnΔC
• Knowing the standard free energy (ΔGo) of ATP hydrolysis permits the calculation of energy available for molecule transport across the membrane ATP + H2O ADP + Pi ΔGo = -31 kJ/ mol [ATP] = 5 mM , [ADP] = 0.3 mM, [Pi] = 90 mM
Δµ = -3.1x104 J mol-1+(8.3 J mol-1 K-1)•(298 K) • ln [ADP] [Pi] [ATP]
Δµ = -3.1 x104 J mol-1+2.47 J mol-1 • ln [3.0 x10-4 M] [9.0 x10-2 M] [5 x10-3 M]
ΔµATP = -4.4 x104 J/mol or - 44 kJ/mol
(10)

Chemiosmotic Hypothesis • States that ATP synthesis is driven
by ΔµH+
• This means: 1. Membranes must be vesicular, sealed
and impermeable to H+ except pathways and proteins involved in H+/ redox generation
Peter D. Mitchell 1920-1992
2. Energy is stored in a ΔpH gradient or Δµ equivalent to ΔµH+
3. ΔµH+ is formed vectorially by alternating H+ and e- carriers in the electron transport chain transport of e- permit the extrusion of H+ (Hence the ratio of H+/ e- = 1) Actually, H+/ e- > 1 due to the activities of the “Q-cycle”

4. H+ flux is coupled to F0F1 ATPase activity driven by ΔµH+
from the (+) side of the membrane (p-side) = ATP synthesis
the reverse reaction drives H+ translocation to the (-) side (n-side) = ATP hydrolysis (n-side)
NADH
4OH- 2H+
2H+
2H+
2e-
2e-
2e-
ATP
ADP + Pi
3H+
3H+
F0F1 ATPase
ΔµH+
nH+
nH+
ETC NAD+
4H2O
2H+ + ½ O2
H2O p-side
n-side
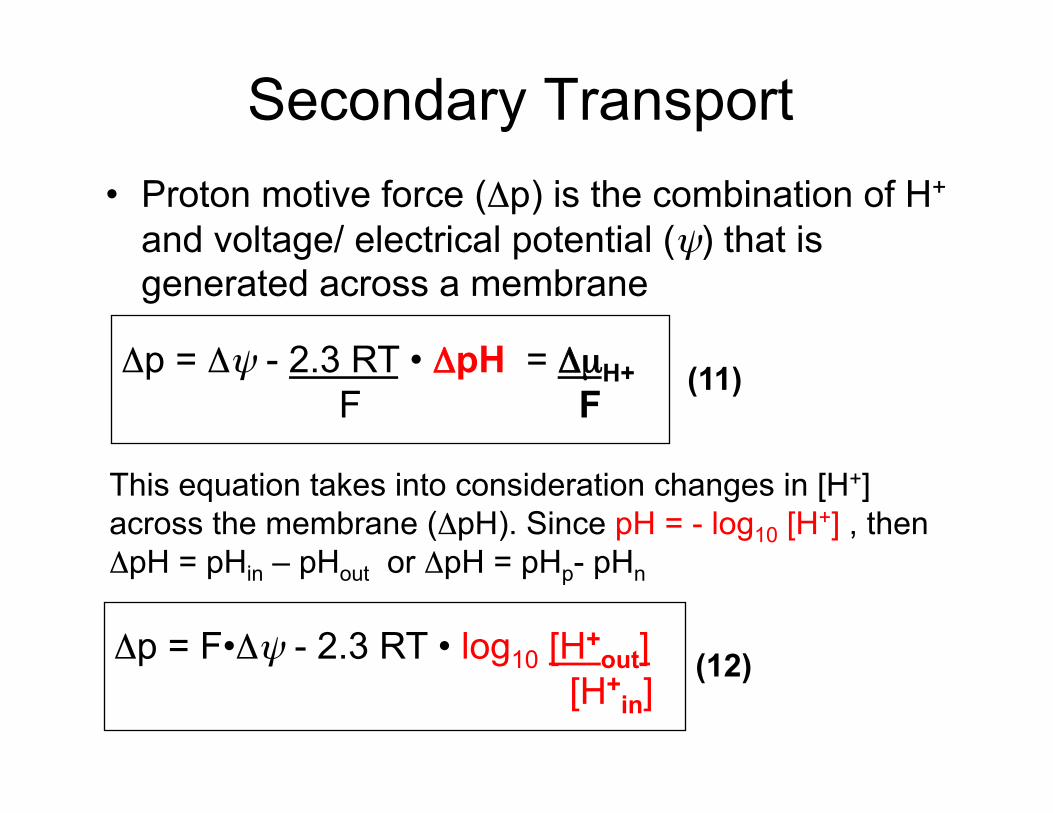
Secondary Transport • Proton motive force (Δp) is the combination of H+
and voltage/ electrical potential (ψ) that is generated across a membrane
Δp = Δψ - 2.3 RT • ΔpH = ΔµH+ F F
(11)
This equation takes into consideration changes in [H+] across the membrane (ΔpH). Since pH = - log10 [H+] , then ΔpH = pHin – pHout or ΔpH = pHp- pHn
Δp = F•Δψ - 2.3 RT • log10 [H+out]
[H+in]
(12)
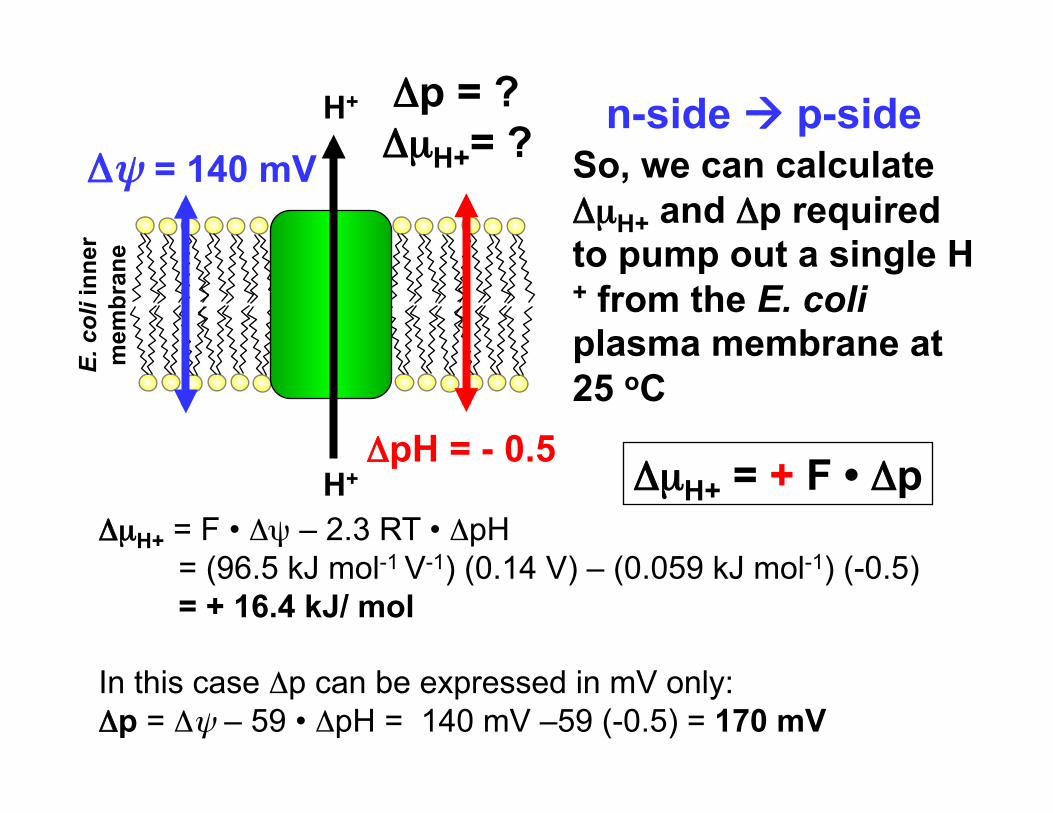
Δψ = 140 mV
H+
H+ ΔpH = - 0.5
So, we can calculate ΔµH+ and Δp required to pump out a single H+ from the E. coli plasma membrane at 25 oC
Δp = ? ΔµH+= ?
ΔµH+ = F • Δψ – 2.3 RT • ΔpH = (96.5 kJ mol-1 V-1) (0.14 V) – (0.059 kJ mol-1) (-0.5) = + 16.4 kJ/ mol
In this case Δp can be expressed in mV only: Δp = Δψ – 59 • ΔpH = 140 mV –59 (-0.5) = 170 mV
E. c
oli i
nner
m
embr
ane
ΔµH+ = + F • Δp
n-side p-side
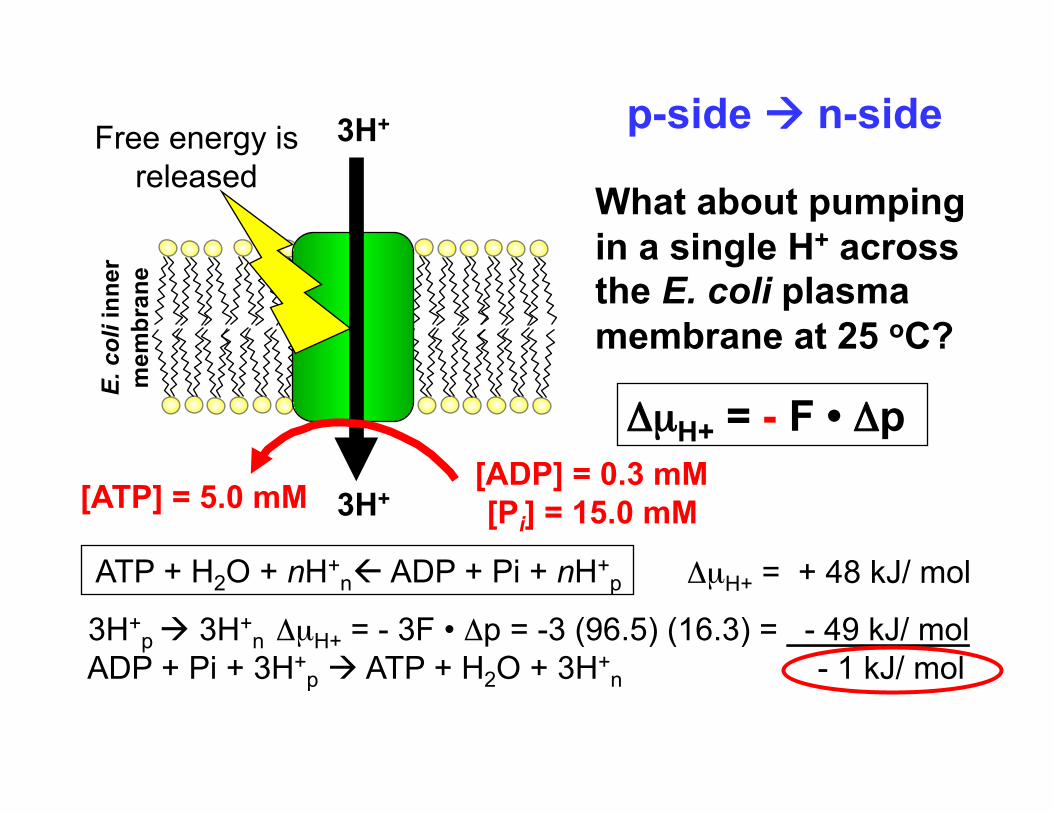
3H+
What about pumping in a single H+ across the E. coli plasma membrane at 25 oC?
E. c
oli i
nner
m
embr
ane
3H+
ΔµH+ = - F • Δp
Free energy is released
ATP + H2O + nH+n ADP + Pi + nH+
p
p-side n-side
[ATP] = 5.0 mM [ADP] = 0.3 mM [Pi] = 15.0 mM
3H+p 3H+
n ΔµH+ = - 3F • Δp = -3 (96.5) (16.3) = - 49 kJ/ mol ADP + Pi + 3H+
p ATP + H2O + 3H+n - 1 kJ/ mol
ΔµH+ = + 48 kJ/ mol
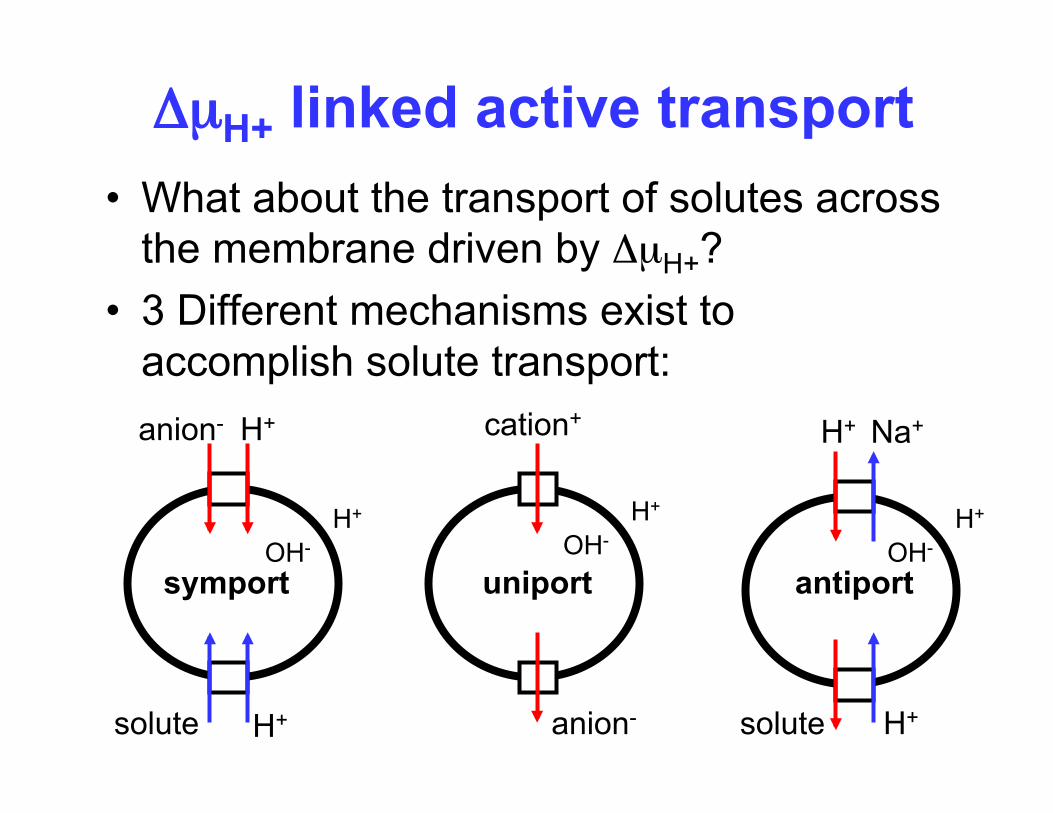
ΔµH+ linked active transport • What about the transport of solutes across
the membrane driven by ΔµH+? • 3 Different mechanisms exist to
accomplish solute transport: anion- H+
solute H+
OH- H+
symport uniport
cation+
anion-
OH- H+
H+ Na+
antiport
solute H+
OH- H+

• If all the free energy available in ΔµH+ is stored in the electrochemical potential of the substrate (Δµs ) then the solute accumulation occurs via each of these 3 mechanisms based on z and n.
Δµs = 2.3 RT log10 [Sin+z] + zF • Δψ
[Sout+z]
(13)
2.3RT log10 [Sin+z] + zF • Δψ = 2.3RT • nΔpH - nFΔψ
[Sout+z]
log10 [Sin+z] = n • ΔpH – (n + z) • Δψ
[Sout+z] 2.3 RT
(14)
Where z = charge on the solute n = number of protons used for transport
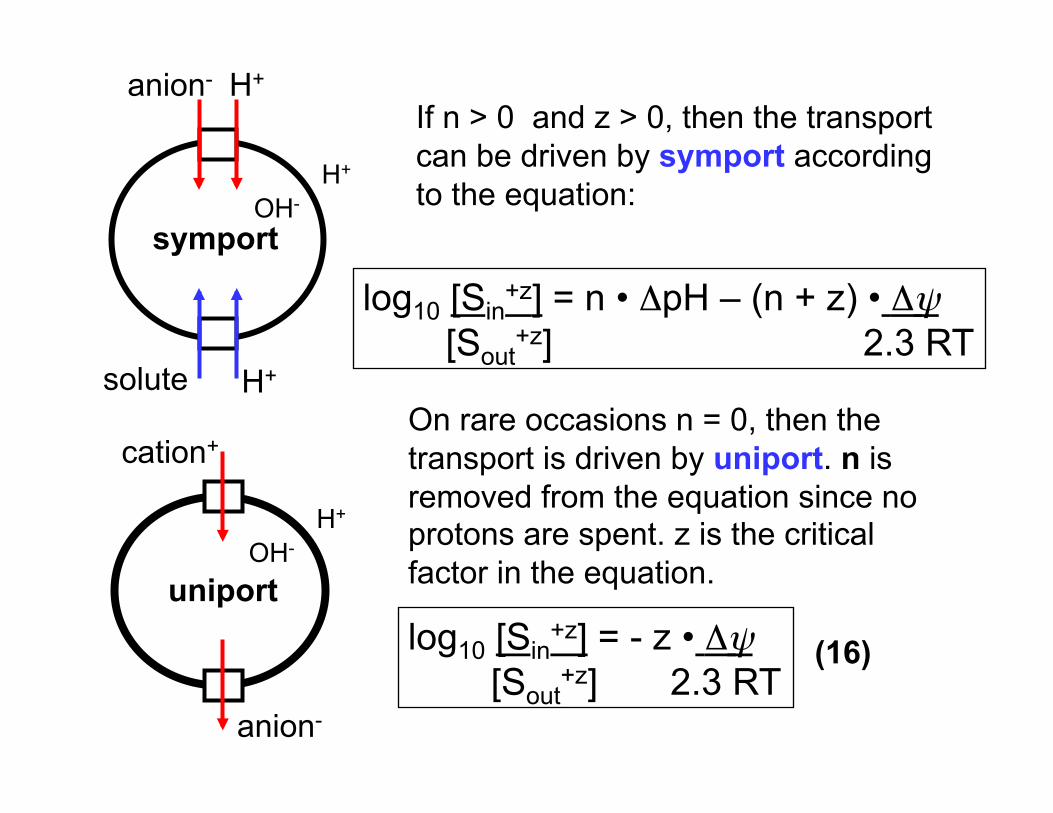
anion- H+
solute H+
OH- H+
symport
If n > 0 and z > 0, then the transport can be driven by symport according to the equation:
log10 [Sin+z] = n • ΔpH – (n + z) • Δψ
[Sout+z] 2.3 RT
On rare occasions n = 0, then the transport is driven by uniport. n is removed from the equation since no protons are spent. z is the critical factor in the equation.
log10 [Sin+z] = - z • Δψ
[Sout+z] 2.3 RT
uniport
cation+
anion-
OH- H+
(16)
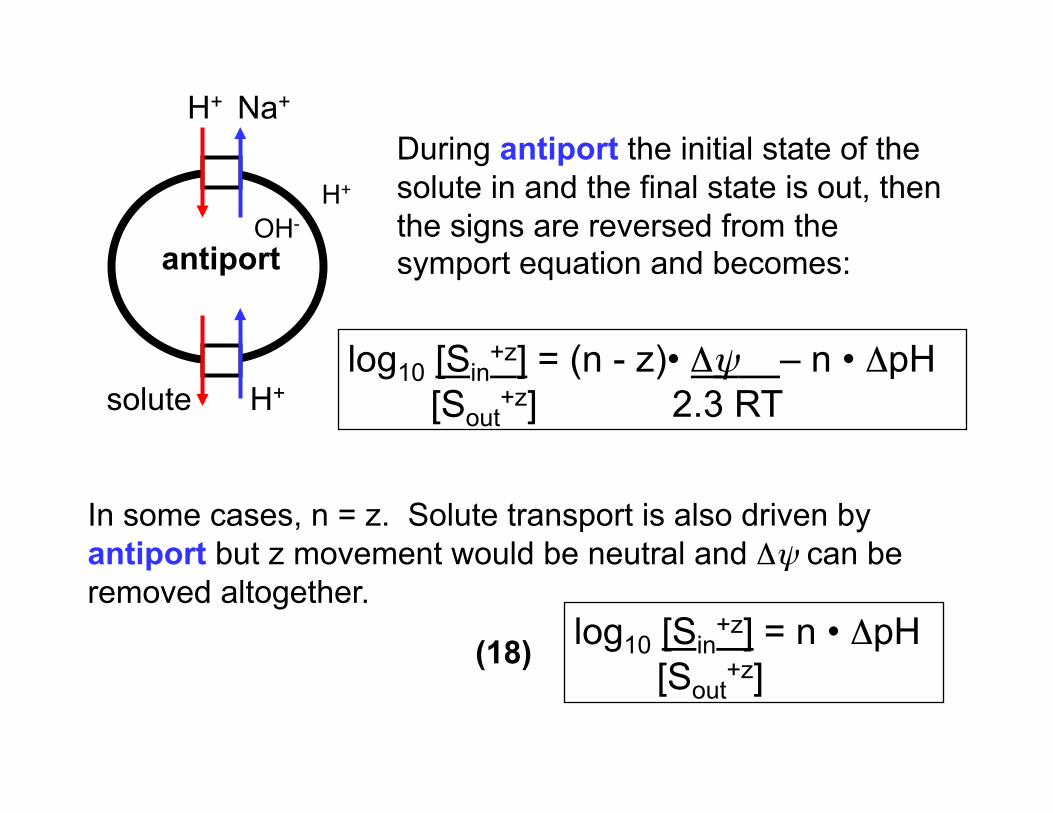
H+ Na+
antiport
solute H+
OH- H+
In some cases, n = z. Solute transport is also driven by antiport but z movement would be neutral and Δψ can be removed altogether.
log10 [Sin+z] = (n - z)• Δψ – n • ΔpH
[Sout+z] 2.3 RT
log10 [Sin+z] = n • ΔpH
[Sout+z]
During antiport the initial state of the solute in and the final state is out, then the signs are reversed from the symport equation and becomes:
(18)

Other methods to generate Δµ • Bacteria and Eukaryotes can produce Δµ and Δψ
using various ions driving the evolution of different electron transport chain components, motility systems, and ATPases
• Na+ and K+ are asymmetrically distributed across membranes due to the activity of Na+, K+ ATPase osmoregulation & cell signalling
• Halophilic microorganisms often use Na+ and K+ in lieu of H+ due to the energetic constraints of their environment costs ATP free energy per mole of Na+ (ΔµNa+)
ΔµNa+ = F • Δψ - 2.3 RT • log10 [Na+out]
[Na+in]
(19)
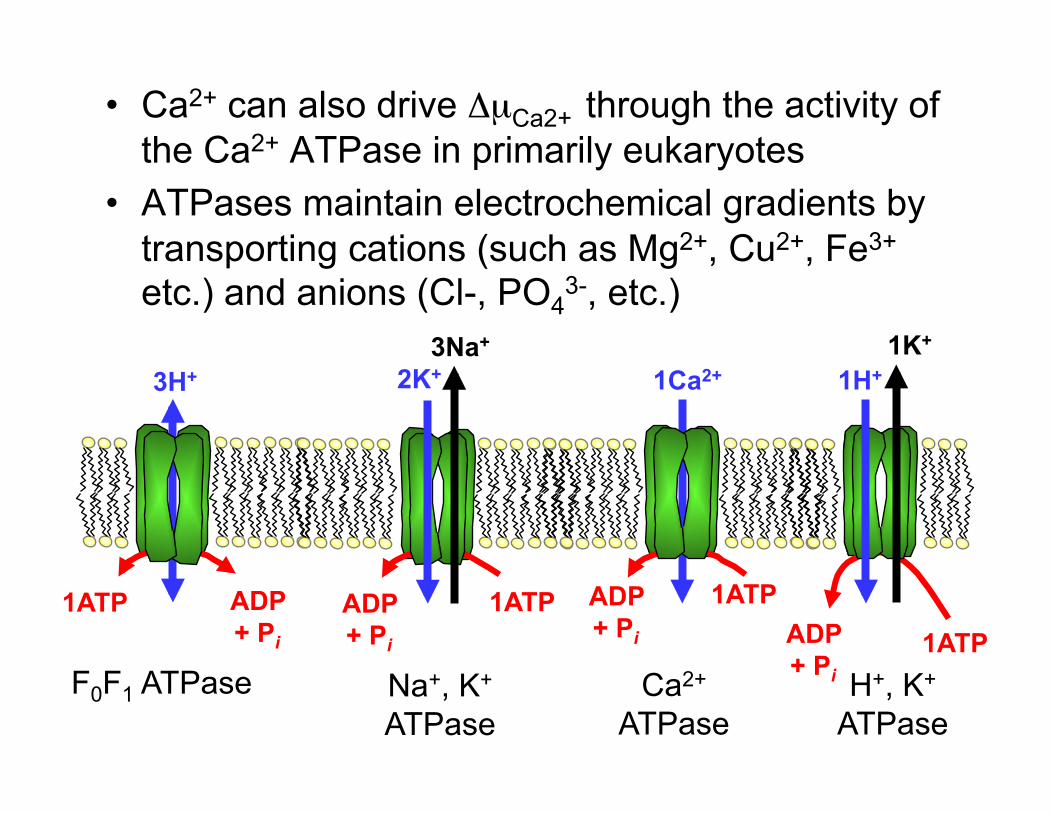
• Ca2+ can also drive ΔµCa2+ through the activity of the Ca2+ ATPase in primarily eukaryotes
• ATPases maintain electrochemical gradients by transporting cations (such as Mg2+, Cu2+, Fe3+
etc.) and anions (Cl-, PO43-, etc.)
ADP + Pi
1ATP 1ATP ADP + Pi
1ATP ADP + Pi
3H+ 3Na+
2K+ 1Ca2+
1ATP ADP + Pi
1H+ 1K+
F0F1 ATPase Na+, K+ ATPase
Ca2+
ATPase H+, K+
ATPase

• Bacteriorhodopsin, from the Archaea Halobacterium salinarium purple membranes pumps H+ across the membrane generating ΔµH+ using energy provided by light
retinal Light

Summary of Active Transport • Active transport requires an input of energy derived from
either ATP hydrolysis (primary) or from Δp (secondary) • Primary transport results in a high yield of free energy
available from ATP + H2O ADP + Pi to transport an energetically prohibited molecule against Δµ of the membrane
• Secondary transport provides energy for primary transport activities (ATP synthesis) as well as for Δp driven reactions – Secondary transport pumps ions (H+) against its natural gradient
to generate energy in the form of an electrochemical potential that facilitates the transport of molecules incapable of energetically diffusing across the membrane
– the solute chemical gradient across a membrane will dictate the type of transport method that can be used ie. symport, uniport, and antiport
– This process is also essential for osmoregulation of other compounds by pumping ions other than H+ across the membrane such as Na+, K+, Ca2+