Embed Size (px)
Citation preview

Agricultural and Forest Meteorology, 67 (1993) 13-27 13 0168-1923/93/$06.00 © 1993 Elsevier Science Publishers B.V. All rights reserved
Transpiration of well-watered and water-stressed Asian pear trees as determined by lysimetry,
heat-pulse, and estimated by a Penman-Monteith model
H.W. Caspari *'aA, S.R. Green b, W.R.N. E d w a r d s b
aDepartment of Plant Science, Massey University, Palmerston North, New Zealand bThe Horticulture and Food Research Institute of New Zealand Ltd, Palmerston North, New Zealand
Received 18 January 1993; revision accepted 8 July 1993)
Abstract
Early season transpiration of 5-year-old Pyrus serotina trees receiving different amounts of irrigation was measured using lysimetry and the compensation heat-pulse technique. Withholding water resulted in a rapid decline in soil-water content within the lysimeters, and caused a 50% decrease in stomatal conduc- tance and a 25-35% reduction in tree water use. Weekly transpiration rates, as determined by integrated heat-pulse measurements, were comparable with measurements by lysimetry for a well-watered tree, but overestimated water use by a water-stressed tree by 22%. The reason for this discrepancy was thought to be diurnal changes in sapwood water content, leading to an overestimation by the heat-pulse technique. Stomatal conductance and meteorological data obtained over 2 days were used to parameterize a Penman- Monteith model to estimate tree transpiration. On a daily basis, these estimates were within 8% of the sap flow determined by the heat-pulse technique. Substantial sap flow occurred in the stressed tree at night even when the stomata were closed. Obviously, this night-time sap flow could not be predicted by the Penman Monteith model. Instead, it appeared to be a capacitance effect owing to the nocturnal refilling of the tree's internal store of water.
Introduction
Asian pears (Pyrus serotina) are a relatively new horticultural crop, not only in the New Zealand fruit industry, but also in Australia, France and Italy. An expansion of plantings has also taken place in the United States, mainly in the western states of California, Oregon and Washington. Outside of Japan, this high-value fruit crop has been subjected to little scientific study. In particular, little is known about its water relations. Such information is important for irrigation scheduling in order to achieve high yields and fruit quality.
* Corresponding author. i Present address: Institut fiir Ostbau und Gemfiseban, Auf dem Htigel 6, 53121 Bonn, Germany.

14 H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27
A series of studies to investigate the water relations of Asian pears was conducted at Massey University from 1988 to 1992. Initial results on water use, and fruit and shoot growth, of the cultivar Hosui under conditions of limiting and non-limiting irrigation regimes have been reported by Caspari et al. (1993) and Chalmers et al. (1992). In this paper we report on the water use of Hosui Asian pears during the early stages of regulated deficit irrigation (RDI), when the trees were subjected to a short period of water stress. This RDI is an irrigation approach aimed at controlling vegetative growth in perennial fruit crops, with little or no detrimental effect to fruit yield and quality (Chalmers, 1989). Using a well-watered and a deficit-irrigated tree, a comparison is made of the water use as determined by lysimetry and the heat- pulse technique against tree water use estimated by a Penman-Monte i th model. The effect of water stress on the water relations of Asian pear trees was examined and the importance of night-time sap flow, as a means of refilling the tree's water store which has been depleted during the day, was determined.
Materials and methods
Lysimeter facility
The experiment was carried out at the lysimeter facility of Massey University near Palmerston North, New Zealand (latitude 40.2°S, longitude 175.4°E). This facility consisted of a row of 12 drainage lysimeters situated within a 1.1 ha block of Asian pears (Pyrus serotina) that was planted during 1986-1987. The whole block was protected by bird netting, hence the light levels within the block were only about 85% of the light levels outside the block. The row orientation within the block was nor th-south . Each lysimeter consisted of a steel cylinder 1.2-m deep and 1 m in diameter. This assembly was surrounded by a concrete sleeve. The tops of the inner cylinders were about 0.1 m above ground level. The lysimeters were covered with a reflective, opaque plastic cover to eliminate rainfall entry and minimize soil evaporation. The tree spacing within the row was 1.2 m and the rows were 5 m apart. The ground cover was grass, with a 2-m wide herbicide strip along the tree line.
The soil in each lysimeter was carefully packed in 0.1-m layers to a depth of 1 m with the exhumed B horizon Manawatu fine sandy loam. One Asian pear tree (cultivar Hosui), grafted on a seedling rootstock, was planted in each of the lysimeters in September 1987 and trained onto a Tatura Trellis (Chalmers et al., 1978). From 1987 to 1992 irrigation and fertilization of each tree was carried out using a closed nutrient-feed system from two 9100-I tanks (Chalmers et al., 1992). The drainage out of each lysimeter was measured using a tipping-bucket gauge located within a subterranean control room.

H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13-27 15
The soil-moisture content in the lysimeters was measured using a neutron probe. An access tube for the neutron probe was placed 0.2 m from the tree trunk in each lysimeter.
The lysimeter facility was operated automatically by a multi-tasking con- troller/datalogger, which controlled the irrigation supply and measured the drainage from each lysimeter. Accumulated irrigation and drainage volumes were recorded once an hour. Data were transferred automatically to disk once a day using an MS-DOS personal computer. A more-detailed description of the facility can be found in Chalmers et al. (1992).
Soil-moisture measurements
A neutron probe (Model 503DR, CPN, Martinez, CA, USA) was used twice a week to measure changes in the volumetric soil-water content, O, with depth. The vertical profile of O was determined from measurements of O at 0.2-m intervals, covering the depth range from 0.2 to 1.0 m beneath the soil surface. Total soil-water storage was obtained by integrating O with respect to depth. 'Pot capacity' could then be defined exactly as the soil-moisture storage when drainage from the lysimeters had just ceased. All lysimeters were filled to pot capacity prior to the beginning of the experiment.
Irrigation regime
From early September 1991 all trees were irrigated every third day to return the soil moisture to pot capacity. Two different irrigation regimes were then applied from 26 October (42 days after full bloom) onwards. Eight trees were selected for a control treatment and these were irrigated in the way that has just been described. An RDI treatment was applied to induce water stress in the remaining four trees. These trees received only one-third of the amount of irrigation given to the control trees. After the soil-water storage in the driest RDI lysimeter had fallen to about 50% of pot capacity, the irrigation amounts for all the RDI trees were then adjusted to maintain soil water at about this 50% level for a period of 40 days.
Plant measurements
The diurnal course of the leaf-water potential (tI/1) o n the 18 November 1991 was measured using a Scholander pressure bomb (Soilmoisture Equipment, Santa Barbara, CA, USA). The measurements were taken approximately every 2 h from just before sunrise to 20:30 h.
Also on 18 November 1991, and on the 27 November, the leaf stomatal conductance (gs) was measured on fully expanded, sunlit leaves using a

16 H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27
transit-time porometer (Delta-T Devices, Model Mk3, Cambridge, UK). These measurements were taken at approximately 2-h intervals throughout the 2 days. Between four and six readings were taken per tree on three trees per treatment. The stomatal conductance of shaded leaves was not measured. Instead, it was assumed on the basis of previous unpublished measurements that the daytime conductance of shaded leaves was equal to half that of sunlit leaves. Night-time conductances were assumed to be equal for all leaves, having a value equal to 1.0 mm s -1. This night-time value was based on measurements of stomatal conductance obtained on other occasions after nightfall and before stomatal opening the following morning.
For the purpose of calculating tree transpiration, a diurnal course in canopy conductance (go) was calculated by taking the parallel sum of the individual conductances of shaded and lit leaves, weighted by their corresponding leaf areas (e.g. McNaughton and Jarvis, 1983). The proportion of shaded leaves was taken to be 50% of the total leaf area, based on the ratio of ground-projected to total leaf area.
The total leaf area of each tree was measured once at the end of the experiment. All trees were defoliated on 14 April 1992, before the onset of natural leaf drop, and the weight of all leaves recorded. The total projected leaf area (one side only) was estimated from a subsample of 15% of leaves (by fresh weight). This was measured using a LI-COR 3100 leaf area meter (LI-COR, Lincoln, NE, USA). We were unable to measure total leaf areas during the experimental period reported here. Instead, the leaf area for 18 27 November was assumed to be 50% of the total fully developed leaf area measured at the end of the experiment. This estimate was based on a canopy growth curve, similar to that proposed for apples (Cain, 1973).
Tree transpiration
Tree transpiration was determined both by lysimetry and heat-pulse measurements. It was also estimated by a modification of the Penman- Monteith model.
Lysimetry For a hydrologically closed system, such as the lysimeters, the transpiration
(73 can be calculated over some time period At using the water conservation equation, namely
T = I - A W - D (1)
where I is applied irrigation, A W is observed change in the soil-water storage and D is measured drainage. Evaporation from the soil surface was considered negligible since the soil surface was covered. The soil-water storage was determined from twice-weekly neutron probe readings. Consequently,

H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27 17
measurements of T using the lysimeter were calculated over 3- and 4-day periods.
Heat-pulse technique Transpiration rates of two lysimeter trees, one control and one RDI, were
estimated from the rate of sap movement upwards through the stem using the compensation heat-pulse technique (Swanson and Whitfield, 1981). Four sets of heat pulse probes, each comprising a heater and two temperature sensors, were installed into parallel holes drilled radially into the stem of each tree. The probe sets were installed at heights of between 0.2 and 0.4 m above the ground, and were equally spaced around the stem circumference. In order to measure the low rates of sap flow anticipated in these trees, the temper- ature probes were placed 10 mm above and 5 mm below the 1.8 mm diameter stainless-steel heater. Sap velocity was subsequently derived using the theoretical calibrations of Swanson and Whitfield (1981) to account for the probe-induced effects of wounding. These calibrations use sapwood water content in the conversion of heat-pulse velocity to sap velocity. Here the wound size was assumed to be equal to the size of the drill hole, i.e. 2 mm diameter.
Each set of heat pulse probes contained a single set of temperature sensors to measure sap velocity at a given radial depth. A radial profile of sap velocity was obtained by using heat-pulse sensors installed at depths of 5, 10, 15 and 20 mm below the cambium. Volume flow rates were calculated from the integral of the sap-velocity profile over the sapwood cross-section, using a second order least squares regression equation fitted to the sap velocity profile (Edwards and Warwick, 1984; Green and Clothier, 1988), given by
E h = 27r rV(r)dr (2) h
where E h is the measured transpiration (kg h -1) and V(r) is the sap velocity as a function of radial depth, r, in a stem of cambium radius, R, and heartwood radius, h.
Trunk diameters of the well-watered and water-stressed trees were 62.4 mm and 70.0 ram, and the corresponding bark depths were 3.1 mm and 3.0 mm, respectively. The heartwood radius was assumed to be 5 mm, based on laboratory analyses of trunks of cut-off Asian pear trees of similar age. Weekly measurements of trunk circumference 0.1 m above the graft union, indicated a negligible change in trunk cross-sectional area over this time period. Consequently, sapwood areas were assumed to remain constant during the experimental period.
Heat-pulse data were recorded automatically, once every 20 min using a Custom Heat Pulse Logger (HortResearch, Palmerston North, New Zealand).

18 H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27
A continuous record of sap flow was obtained over the time period 30 October-2 December 1991.
Penman- Monteith model The procedure adopted for calculating tree transpiration using the Penman-
Monteith model was similar to that proposed by Thorpe (1978). The tree canopy was assumed to be a single hypostomatous leaf and transpiration calculated as
AE = sRnLp + 0"93pcpDaL/ra (3) s + 0.93"7(2 + rc/ra)
where ~ is the latent heat of vaporisation (2.454 kJ g-l); E is the transpiration rate (g s-l); s is the slope of the saturation vapour pressure curve (Pa °C -1) at the ambient air temperature T a (°C); Rn is the net radiation flux density (W m-2); Lp is the projected ground area covered by the evaporating plant canopy (2.9 m2); p is the air density (kg m 3); Cp is the specific heat capacity of air at constant pressure (J kg -l K-l); Da is the vapour pressure deficit of air (Pa); L is the total projected leaf area (one side only) of the plant canopy (m2); '7 is the psychrometric constant (66.1 Pa); r a and r c are the aerodynamic and canopy resistances (s m 1), respectively. This expression is similar to that of Jarvis and McNaughton (1986), except for the factor 2, which arises because r a is calculated as the parallel sum over both sides of a leaf, and the factor 0.93, which arises from the assumption that the boundary layer resistance for vapour is 0.93 times that for sensible heat. The canopy resistance (rc = 1/g~) was estimated as the parallel sum of the mean leaf resistance for sunlit and shaded leaves, weighted by their corresponding leaf areas. The leaf area index is given by the ratio L/Lp.
Implicit in Eq. (3) is the assumption that the tree canopy absorbs the same amount of radiation as a horizontal plane of area Lp. In this experiment, the tree canopies were trained into a V-shape by tying branches onto a V-shaped wire trellis known as a Tatura Trellis (Chalmers et al., 1978). At a stem height of about 0.5 m above the ground, two scaffold limbs were selected and trained, one up each side of the trellis. The crotch angle between these scaffolds was about 60 ° . Weak secondary limbs and fruiting laterals developing from the main scaffold limbs were tied onto the wires along the tree line in order to fill the canopy. Growth towards the inside of the V was removed. Therefore, as the sun angle varied during the day, it was possible the pattern of radiation absorption by the tree canopy differed from that absorbed by a horizontal surface of area Lp. The results of McNaughton et al. (1992) suggest this approximation for the radiation absorption is reasonable, given the close proximity of neighbouring trees.
Leaf boundary layer resistance was calculated from the empirical relation,

H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13-27 19
derived by Landsberg and Powell (1973), which accounts for the mutual sheltering of clustered leaves, namely
r. = 135(d/u) °5 (4)
where d is a characteristic leaf dimension (m) and u is the mean wind speed (m s -1) across the leaf surface.
Meteorological measurements
Incoming solar radiation, net radiation, air temperature, relative humidity and wind speed were all measured by an automated weather station at the lysimeter site. Mean values were recorded at 20-min intervals using a Campbell CR21X datalogger (Campbell Scientific, Logan, UT, USA). The instruments were placed just above the V-shaped canopy, at a height of approximately 3 m above the ground.
Results
The two trees containing the heat-pulse sensors were focussed. These trees were considered to be representative of the control and RDI treatment. Changes in mean soil-water storage for both the well-watered and the water-stressed tree are shown in Fig. 1. Both lysimeters were maintained close to their pot capacity until the start of the RDI treatment (42 days after full bloom). The pot capacity of each lysimeter was not the same, possibly because of different volumes of soil and root in each lysimeter. After 3 weeks of RDI, the soil-water storage in the RDI lysimeter had
140 0 well-watered tree ]
13o " - ~ 'L 120
110
100 o
"~ 90
"6 80
! 70
o') 60
Sep. Oct. Nov, Dec.
1991
Fig. 1. Changes in soil-water storage per lysimeter for the well-watered (©) and the water-stressed (Q) Asian pear tree. Up-arrows denote 18 and 27 November 1991.

20 H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27
Table 1 Comparison of early season water use (1) of Hosui Asian pear trees calculated from lysimetry and heat-pulse measurements
Observation Well-watered tree Water-stressed tree period
Heat-pulse Lysimetry Heat-pulse Lysimetry
31 Oct. 3 Nov. 14.3 15.7 17.3 12.9 4-10 Nov. 21.4 21.8 25.5 24.6 11 17 Nov. 28.1 30.3 31.5 23.3 18 24 Nov. 33.6 33.3 27.9 19.9 25 Nov. 1 Dec. 27.4 24.4 20.6 19.9
dropped to about 50% of pot capacity. Thereafter, the irrigation schedule of the water-stressed tree was changed to maintain the soil water at about this level.
The weekly transpiration (7) by the two trees, as determined from lysimetry and heat-pulse measurements, is presented in Table 1. For the well-watered tree, the difference in total T by the two methods over the entire 33-day period was less than 0.5%, although individual weekly estimates varied by as much as 10%. For the stressed tree, however, the agreement between lysimetry and the heat-pulse technique was not as good. The heat-pulse technique indicated a 22% higher water use than did lysimetry over the same time period. In a previous experiment, Caspari et al. (1993) found good agreement between the two methods for the first 4 weeks following probe implantation and a more variable behaviour beyond this. This greater variation was attributed to the trees response to wounding. A wound response does not explain our
2.0
~,5
1.0
0.5
0.0
t--
j 0 w e l l - w a t e r e d tree 2o- lo-91 to 1-12-91 • water-stressed tree
0
0
0 0 0 0 • 0 •
10 20 30 40
Days after probe implantation
Fig. 2. The ratio of tree water use determined by the heat-pulse technique (TH) to tree water use determined by lysimetry (TL) as affected by the time following probe implantation. Water use is averaged over 3- and 4-day periods.
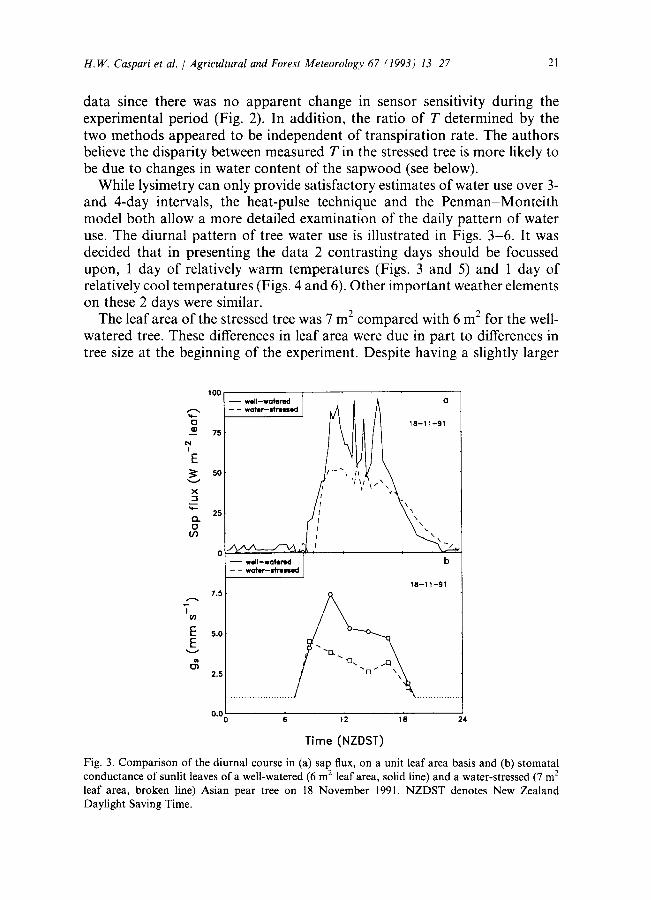
H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27 21
data since there was no apparent change in sensor sensitivity during the experimental period (Fig. 2). In addition, the ratio of T determined by the two methods appeared to be independent of transpiration rate. The authors believe the disparity between measured T in the stressed tree is more likely to be due to changes in water content of the sapwood (see below).
While lysimetry can only provide satisfactory estimates of water use over 3- and 4-day intervals, the heat-pulse technique and the Penman-Monte i th model both allow a more detailed examination of the daily pattern of water use. The diurnal pattern of tree water use is illustrated in Figs. 3-6. It was decided that in presenting the data 2 contrasting days should be focussed upon, 1 day of relatively warm temperatures (Figs. 3 and 5) and 1 day of relatively cool temperatures (Figs. 4 and 6). Other important weather elements on these 2 days were similar.
The leaf area of the stressed tree was 7 m 2 compared with 6 m 2 for the well- watered tree. These differences in leaf area were due in part to differences in tree size at the beginning of the experiment. Despite having a slightly larger
100 ] - - we l l -wa te red | J A o - - w a t l r - - l t r n l e d J ~/~ ~ / /
~ 75
7 E
~ 5o
x
- - w,,-wmertd I b - - w o t t r - s t r n s t d l
i
1 8 - 1 1 - 9 1 ~ ~ 7.5
E 5.0 E
2.5
0.0 0 6 12 18 24
T i m e ( N Z D S T )
Fig. 3. Compar i son of the diurnal course in (a) sap flux, on a unit leaf area basis and (b) s tomatal conductance of sunlit leaves of a well-watered (6 m 2 leaf area, solid line) and a water-stressed (7 m 2 leaf area, broken line) Asian pear tree on 18 November 1991. N Z D S T denotes New Zealand Daylight Saving Time.

22 H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27
0
? E
v
X
m vs-
Q .
0 (/~
I u~
E E
100
75
50
25
7.5
5.0
2.5
0.0
- - well-waterad water-streued
- - wel l -wafered I -- - water-stllned
el
b
2 7 - 1 1 - 9 1
1'2 1'8 24
Time (NZDST)
Fig. 4, Legend as for Fig. 3, using data of 27 November 1991.
leaf area, the stressed tree used less water. This result was not surprising since, as a result of the water-stress, the stomatal conductance of the stressed tree was only about one half that of the well-watered tree for most of the daytime (Figs. 3 and 4). Consequently, sap flow was higher in the well-watered tree with the peak rate occurring around solar noon when net-radiation was highest. Sap flow in the stressed tree peaked at about the same time, but at a lower rate. Sap flow in the well-watered tree responded markedly to changes in net radiation, whereas the stressed tree was much less sensitive to these fluctuations. Thus, the diurnal pattern of sap flow in the stressed tree was relatively constant during the middle part of the day.
For the well-watered tree, the time course in water use predicted by the Penman-Monteith model was in good agreement with measured sap flow on both days (Figs. 5 and 6). The daily totals agreed to within 7% on 18 November and to within 2% on 27 November. In the stressed tree, on the other hand, the daily pattern of predicted water use was less consistent with the pattern of sap flow, particularly during early morning and late afternoon. Reasonable agreement was, however, observed during the middle part of the day (Figs. 5 and 6). On a daily basis, the two methods still agreed to within 8% on 18 November and to within 1% on 27 November. For both trees on both

H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27 23
1.00
I 0 , 7 5 t -
O.S0
_o ,+_
O_ 0 0.25
0 3
0 . 0 0
- - heat -pu lse O P - M model wel l -woterod free
1 8 - 1 1 - 9 1
/ /
- ~ . I /!
- - h e a t - p u l s e / b - - P - M mode( J water -Mr~ssed tree
1 8 - 1 1 - 9 1 I 0 . 7 5 ¢-
0 . 5 0 I / / I
_o r~ 0 0.25 O0 / /
0.00 - r . : : -
- - R . ' ; ' C
- ' 0 . : '
"~" 5oo--r. ¢_,~.;"i, 18-11-9, 20
/ . \
o 60O
200
Cff " " -- 5
0
6 12 18 24
Time (NZDST)
Fig. 5. Comparison of the diurnal course in sap flow as determined by the heat-pulse technique (solid line) and tree transpiration predicted by a Penman-Monteith model (broken line). (a) represents the well-watered tree; (b) represents the water-stressed tree; (c) is the corresponding meteorological data for 18 November 1991. NZDST denotes New Zealand Daylight Saving Time.
days, it appeared that more sap flow was occurring late in the day than was attributed to transpiration via the Penman-Monteith model. It seems unlikely that this sap flow resulted from transpiration because the stomata were closed (Figs. 3 and 4). Also, it appeared that transpiration in the stressed tree was occurring early in the morning before the onset of sap flow.
Except for air temperature (Figs. 5(c) and 6(c)), all other important environ- mental variables were similar on 18 and 27 November. The contrasting tem- peratures had an apparent effect on the stomatal function of both trees, and this was reflected in the different stomatal conductances and water use on these 2 days. In particular, the stomata on both trees opened more than 1 h later on the cooler day (27 November) and values of gs remained about 30%
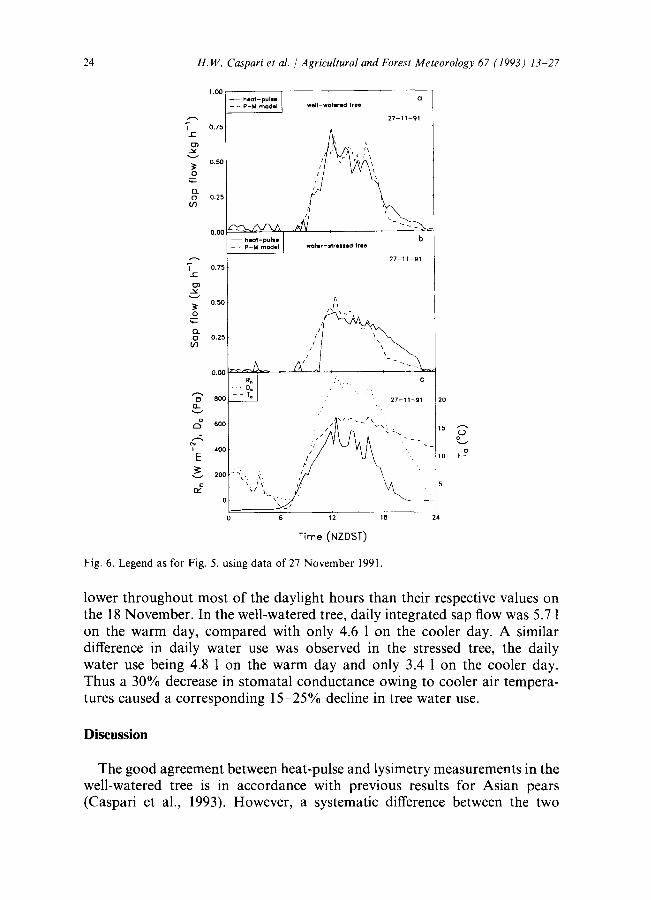
24 H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13-27
I t -
v
?= 0
EL
1.00
0.75
0.50
0.25
0.00
It.. 0.75
0.50 0
13- 0,25
0.00
~ " 800 Q.-
600
I 400 E
200
o
I - - heat -pu lse I a - - P - M model J wel l -watered tree
27-11-91
/ \
- ~ ' 2 " ~ / A Z ~ . - - h e o t - p u l e e
P - M model woter-stressed tree b
27-11-91
ix x ~
Jill / X
, ' " " ' " ' " " ' " ' " " 27-11-91 20
~('" fi'~ / 5
0 6 12 18 24
Time (NZDST)
Fig . 6. Legend as for Fig . 5, using data of 27 November 1991.
lower throughout most of the daylight hours than their respective values on the 18 November. In the well-watered tree, daily integrated sap flow was 5.7 1 on the warm day, compared with only 4.6 1 on the cooler day. A similar difference in daily water use was observed in the stressed tree, the daily water use being 4.8 1 on the warm day and only 3.4 1 on the cooler day. Thus a 30% decrease in stomatal conductance owing to cooler air tempera- tures caused a corresponding 15-25% decline in tree water use.
Discussion
The good agreement between heat-pulse and lysimetry measurements in the well-watered tree is in accordance with previous results for Asian pears (Caspari et al., 1993). However, a systematic difference between the two

H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27 25
methods in the stressed tree was not anticipated. This discrepancy could arise from errors in the sap-flow calculation owing to diurnal changes in the volume fraction of water in the tree trunk. We did not measure the water content of the tree stems during the experiment, but used measured values from well- irrigated Asian pear trees obtained the previous year. Significant changes in sapwood water content have been reported. For example, Waring and Run- ning (1978), and Schnock (1972) measured diurnal changes in sapwood water content of between 20% and 30% for Scots pine and hornbeam, respectively. Similarly, Brough et al. (1986) reported a diurnal change of 15% in the xylem water content of non-irrigated apple trees. We note that a 30% reduction in the water content of the sapwood would lead to a 20% increase in sap flow calculated by the heat-pulse technique. This is of the same order as the discrepancy observed between the heat-pulse technique and lysimetry. These results reiterate the importance of measuring sapwood water contents when applying the heat-pulse technique in trees where significant changes in sapwood water content are likely to occur.
The heat-pulse technique indicated significant sap flow for several hours after the stomata had closed. Similar observations of night-time sap flow in kiwifruit vines have been reported by Green et al. (1989) on nights when the saturation deficit of the air remained elevated. In that case, the stomata remained open at night (gs = 4 mm s-l), so that sap flow could be directly attributed to transpiration. In the case of the Asian pear, however, this night- time sap flow was not the result of transpiration at night, because the stomata were essentially closed (gs = 1 mm s-l).
Nocturnal sap flow occurring in the absence of significant transpiration is compatible with a night-time refilling of the water lost from the tree's different organs, such as leaves, fruit and branches. During daylight hours, water stored in the various plant tissues can be released to the transpiration stream (Powell and Thorpe, 1977; Landsberg and Jones, 1981), but this water even- tually must be replenished. Branches of a tree start to shrink whenever water available through this capacitance becomes exhausted; the degree of shrink- age reflects the water use. Stem shrinkage occurs almost exclusively in tissues external to the xylem (Kozlowski, 1972). In addition, the water content of the xylem declines during the day, the decline being largest in non-irrigated trees (Brough et al., 1986). An increased use of stored water by the stressed tree during the daytime would partly explain subsequent differences in sap flow in the two trees after the stomata had closed at night.
Consistent with this hypothesis are leaf water potentials (~l) measured at 20:30 h of -0 .2 MPa for the well-watered tree and -1.1 MPa for the stressed tree. The former value is typical of the pre-dawn ~l for well-irrigated Asian pear trees (Caspari et al., 1993). It therefore seems likely that the capacitance of the well-watered tree was refilled by about 20:30 h, whereas the stressed tree was still under some degree of water stress. With apple trees the recovery of

26 H.W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13 27
leaf water potential has been observed to mirror the recovery of stem water content (Brough et al., 1986).
Approximately 15% of the daily total sap flow in the stressed tree occurred at night. Landsberg et al. (1976) calculated that an apple tree with 4 m 2 leaf area and a total weight (stem plus roots) of 15 kg would have approximately 0.7 kg of water 'in store'. The sap flow occurring in the stressed tree from 18:00 h to midnight was about 0.65 kg and is comparable with the data for apple. Therefore, our estimates of recharge at night seem reasonable.
Conclusion
The lysimeter facility proved to be an excellent tool for controlling and manipulating the soil water status of our experimental trees. Following 3 weeks of a water-stress treatment, the stomatal conductance of our stressed tree was reduced by about 50% relative to a well-watered tree. This reduction in gs led to a 25-35% reduction in water use. On a daily basis, measured sap flow was in reasonable agreement with transpiration predicted by a Penman- Monteith model. However, significant sap flow continued for several hours after stomata had closed and transpiration had presumably declined. This sap flow is consistent with a capacitance refilling of the tree's water store that had been depleted during the day. This refilling process appears to be an important component in the daily pattern of sap flow in the Asian pear, especially for trees under some degree of water stress. Our results also reiterate the importance of measuring sapwood water content when applying the heat- pulse technique in trees where significant changes in sapwood water content are anticipated.
Acknowledgements
This work was carried out while the senior author was a visiting scientist at the Department of Plant Science, Massey University. The work was supported by a grant to the senior author from the Gottlieb Daimler- und Karl Benz- Stiftung, Ladenburg, Germany.
References
Brough, D.W., Jones, H.G. and Grace, J., 1986. Diurnal changes in water content of the stems of apple trees, as influenced by irrigation. Plant, Cell Environ., 9: l--7.
Cain, J.C., 1973. Foliage canopy development of 'McIntosh' apple hedgerow in relation to mechanical pruning, the interception of solar radiation, and fruiting. J. Am. Soc. Hortic. Sci., 98: 357-360.
Caspari, H.W., Behboudian, M.H., Chalmers, D.C. and Renquist A.R.. 1993. Pattern of seasonal

H, W. Caspari et al. / Agricultural and Forest Meteorology 67 (1993) 13-27 27
water use of Asian pears determined by lysimeters and the heat-pulse technique. J. Am. Soc. Hortic. Sci. (in press).
Chalmers, DJ. , 1989. A physiological examination of regulated deficit irrigation. N.Z.J. Agric. Sci., 23: 44-48.
Chalmers, D.J., van den Ende, B. and van Heek, L., 1978. Productivity and mechanization of the Tatura trellis orchard. HortScience, 13:517 521.
Chalmers, D.J., Andrews, P.K., Harris, K.M., Cameron, E.A. and Caspari, H.W., 1992. Performance of drainage lysimeters for the evaluation of water use by Asian pears. HortScience, 27: 263- 265.
Edwards, W.R.N. and Warwick., N.W.M., 1984. Transpiration from a kiwifruit vine as estimated by the heat-pulse technique and the Penman-Monteith equation. N. Z. J. Agric. Res., 27:537 543.
Green, S.R. and Clothier, B.E., 1988. Water use of kiwifruit vines and apple trees by the heat-pulse technique. J. Exp. Bot., 39:115 123.
Green, S.R., McNaughton, K.G. and Clothier, B.E., 1989. Observations of night-time water use in kiwifruit vines and apple trees. Agric. For. Meteorol., 48:251 261.
Jarvis, P.G. and McNaughton, K.G., 1986. Stomatal control of transpiration: scaling up from leaf to region. Adv. Ecol. Res., 15:1 49.
Kozlowski, T.T., 1972. Shrinkage and swelling of plant tissues. In: T.T. Kozlowski (Editor), Water Deficit and Plant Growth, Vol. 3. Academic Press, New York, pp. 1 64.
Landsberg, J.J. and Powell, D.B.B., 1973. Surface exchange characteristics of leaves subject to mutual interference. Agric. Meteor,-:., 13:169 184.
Landsberg, J.J. and Jones, H.G., 1981. Apple orchards. In: T.T. Kozlowski (Editor), Water Deficit and Plant Growth, Vol. 6. Academic Press, New York, pp. 419-469.
Landsberg, J.J., Blanchard, T.W. and Warrit, B., 1976. Studies on the movement of water through apple trees. J. Exp. Bot., 27: 579-596.
McNaughton, K.G. and Jarvis, P.G., 1983. Predicting effects of vegetation changes on transpiration and evaporation. In: T.T. Kozlowski (Editor), Water Deficit and Plant Growth, Vol. 7. Academic Press, New York, pp. 1 47.
McNaughton, K.G., Green, S.R., Black, T.A., Tynan, B,R. and Edwards, W.R.N., 1992. Direct measurement of net radiation and photosynthetically active radiation absorbed by a single tree. Agric. For. Meteorol., 62:87 107.
Powell, D.B.B. and Thorpe, M.R., 1977. Dynamic aspects of plant water relations. In: J.J. Landsberg and C.V. Cutting (Editors), Environmental Effects on Crop Physiology. Academic Press, London, pp. 259 279.
Schnock, G., 1972. Contenu en eau d'une phytoc6nose et bilan hydrique de 1'6cosyst6me: ch6naie de virelles. Ecol. Plant., 7: 205--226.
Swanson, R.H. and Whitfield, D.W.A., 1981. A numerical analysis of heat pulse velocity theory and practice. J. Exp. Bot., 32:221 239.
Thorpe, M.R., 1978. Net radiation and transpiration of apple trees in rows~ Agric. Meteorol., 19: 41-57.
Waring, R.H. and Running, S.W., 1978. Sapwood water storage: its contribution to transpiration and effect upon water conductance through the stems of old-growth Douglas-fir. Plant, Cell Environ., 1: 131-140.





























