Embed Size (px)
Citation preview

Plant, Cell and Environment
(2003)
26
, 1515–1523
© 2003 Blackwell Publishing Ltd
1515
Blackwell Science, LtdOxford, UKPCEPlant, Cell and Environment0016-8025Blackwell Science Ltd 2003? 2003
26?15151523Original Article
Metabolic changes in P-deficient rice rootsJ. Wasaki
et al.
Correspondence: Jun Wasaki, Tel. & Fax:
+
81 11 706 3845; e-mail:[email protected]
Transcriptomic analysis of metabolic changes by phosphorus stress in rice plant roots
J. WASAKI
1
, R. YONETANI
1
, S. KURODA
1
, T. SHINANO
1,2
, J. YAZAKI
3
, F. FUJII
4
, K. SHIMBO
4
, K. YAMAMOTO
4
, K. SAKATA
3
, T. SASAKI
3
, N. KISHIMOTO
3
, S. KIKUCHI
3
, M. YAMAGISHI
1
& M. OSAKI
1
1
Graduate School of Agriculture, Hokkaido University, N9W9, Kita-ku, Sapporo, 060–8589, Japan,
2
Creative Research Initiative ‘Sousei’ (CRIS), Hokkaido University, N8W5, Kita-ku, Sapporo, 060-0808, Japan,
3
National Institute of Agrobiological Sciences, 2-1-2, Kannondai, Tsukuba, Ibaraki, 305–8602, Japan and
4
Society for Techno-innovation of Agriculture, Forestry and Fisheries, 446–1, Ippaizuka, Kamiyokoba, Tsukuba, Ibaraki, 305–0854, Japan
ABSTRACT
As most soil phosphates exist as insoluble inorganic phos-phate and organic phosphates, higher plants have devel-oped several strategies for adaptation to low phosphorus(P). These include the secretion of acid phosphatase andorganic acids, induction of the inorganic phosphate (Pi)transporter and the substitution of some enzyme activitiesas alternative pathways to increase P utilization efficiency.It has been proposed that plants also have a ‘
pho
regulon’system, as observed in yeast and
Escherichia coli
; however,the detail of the regulation system for gene expression onP status is still unclear in plants. To investigate the alter-ation of gene expression of rice roots grown under P-defi-cient conditions, a transcriptomic analysis was conductedusing a cDNA microarray on rice. Based on the changes ofgene expression under a –P treatment, the up-regulation ofsome genes due to P deficiency was confirmed. Some newimportant metabolic changes are suggested, namely: (1)acceleration of carbon supply for organic acid synthesisthrough glycolysis; (2) alteration of lipid metabolism; (3)rearrangement of compounds for cell wall; and (4) changesof gene expression related to the response for metallic ele-ments such as Al, Fe and Zn.
Key-words
: phosphorus deficiency; rice, cDNA microarray;transcriptome.
INTRODUCTION
As most of phosphorus (P) compounds exist as insolubleinorganic phosphate or organic phosphate, the acquisitionratio of P is lower than other essential elements. With theincrease in world population, an efficient use of P resourcesis required for sustainable food production. To achieve thisaim, the strategies for adaptation to low P need to be wellknown.
When plants are grown under P-deficient conditions,their roots secrete acid phosphatase (APase) and organic
acids to release inorganic phosphate (Pi) from organic com-pounds and insoluble inorganic phosphate compounds,respectively (Gardner, Barber & Parbey 1983; Tadano &Sakai 1991). Several related genes have been isolated andcharacterized such as
LASAP2
encoding S-APase of whitelupin (Wasaki
et al
. 2000). It has also been reported thatgene expression of high affinity Pi transporter wasincreased in roots of P-deficient plants (Mucchal, Pardo &Raghothama 1996; Mucchal & Raghothama 1999; Liu
et al
.2001; Kai
et al
. 2002). The efficient utilization of absorbedP in plant tissues is also known. Many papers describe thebypass pathway contributing to save Pi rather than produc-ing energy (Duff
et al
. 1989; Duff, Plaxton & Lefebvre1991). Production of ribonucleases also increases under Pdeficiency, and its function is to mobilize Pi from internalRNA pools (Nürnberger
et al
. 1990; Löffler
et al
. 1992;Green 1994; Bosse & Köck 1998). Goldstein, Baretlein &Danon (1989) proposed that plant has a ‘
pho
regulon’, asobserved in yeast (Bergman
et al
. 1986) and
Escherichiacoli
(Torriani & Ludtke 1985), which may regulate theexpression of these low P adaptation-related genes. How-ever, the detail of the regulation mechanism of the geneexpression system responding on P status has not yet beenexplained.
In recent years, genomic studies have been undertaken.For example, the genome sequence of
Arabidopsis thaliana
has been completed (The Arabidopsis Genome Initiative2000), and the draft sequence of the rice genome deter-mined (Goff
et al
. 2002; Yu
et al
. 2002). Further researchinvolving the genome sequence includes the developmentof post-genomic studies that include transcriptomic analysisusing cDNA arrays. Recently, several articles describing theresults of transcriptomic analysis for inorganic nutrientmetabolisms of higher plants have been published (Wang
et al
. 2000; Thimm
et al
. 2001; Wang, Gravin & Kochian2001, 2002; Negishi
et al
. 2002). These analyses have con-tributed notably to the understanding of how changes ofmetabolism respond on nutrient status.
In the present study, we used the cDNA microarray ofrice made by the microarray project in Japan (Kishimoto
et al
. 2002). The all-inclusive expression analysis was per-formed to investigate the changes of gene expression under

1516
J. Wasaki
et al
.
© 2003 Blackwell Publishing Ltd,
Plant, Cell and Environment,
26,
1515–1523
P-deficient conditions and to understand the function of thelow P adaptation mechanism of plant.
MATERIALS AND METHODS
Plant material
Rice (
Oryza sativa
L. ssp. japonica cv. Michikogane) seedswere sterilized with 70% ethanol and germinated for 36 hin 1 m
M
CaCl
2
with aeration in dark conditions. Germinatedplants were transferred to pots containing 1 m
M
CaCl
2
andgrown for 4 d to reduce the level of stored phosphorus inthe seed. They were then transferred to a nutrient solutionand treated with or without P. All culture was conducted ina growth chamber at 28
∞
C, 24 h light, approximately130
m
mol photons m
-
2
s
-
1
at leaf level and 50% relativehumidity. The composition of the nutrient solution was asfollows: 0.83 m
M
NH
4
NO
3
, 0 (–P solution) or 32
m
M
(
+
Psolution) NaH
2
PO
4
, 0.38 m
M
KCl, 0.19 m
M
K
2
SO
4
, 0.75 m
M
CaCl
2
, 0.82 m
M
MgSO
4
, 36
m
M
Fe (III)-EDTA, 9.1
m
M
MnSO
4
, 46
m
M
H
3
BO
3
, 3.1
m
M
ZnSO
4
, 0.16
m
M
CuSO
4
and7.4 n
M
(NH
4
)
6
Mo
7
O
24
. The pH was adjusted to 5.2 using0.1
M
NaOH and the solution was changed every 24 h. Halfof the plants grown on the
+
P solution were transferred tothe –P solution after 8 d and all plants were collected thenext day. The treatments were defined as
+
P treatment,short-term –P treatment (transferred from the
+
P solutionafter 8 d) and long-term –P treatment (continuously grownin the –P solution for 9d). The fresh weights of shoots androots were measured, then the samples were frozen imme-diately using liquid nitrogen and stored at
-
80
∞
C.
Measurement of total P content
The shoots were freeze-dried and ground, then approxi-mately 50 mg of each sample was digested with H
2
SO
4
–H
2
O
2
. The P content in the digested solution was measuredusing the method of Saheki, Takeda & Shimizu (1985).
RNA extraction and labelling
Total RNAs were extracted by sodium dodecyl sulphate(SDS)–phenol method (Palmiter 1974) and purified byCsCl gradient ultracentrifuge method (Sambrook, Fritsch& Maniatis 1989). Forty micrograms of purified RNAs wereused per array and set as the targets for microarrays. Thesewere reverse-transcribed and labelled using Superscript II(Gibco BRL, Rockville, MD, USA) with Cy5-dCTP(Amersham Bioscience, Piscataway, NJ, USA). Unlabelledprimers and dyes were removed with GFP columns (Amer-sham Bioscience).
Microarray analysis
Rice cDNA microarrays were prepared by the microarrayproject in Japan (Kishimoto
et al
. 2002), with a systemdeveloped by Amersham Bioscience. The arrays contained8987 cDNA clones from several organs of rice (
Oryza sativa
L. ssp. japonica cv. Nipponbare) and were prepared on twoslide glasses. Half of the cDNAs were spotted on each slideas duplicates, and hybridization was performed at 60
∞
C for4 h using a humidified chamber and an ExpressHyb solu-tion (Clontech, Palo Alto, CA, USA). After hybridization,the glasses were washed and dried. The signal intensity ofeach spot was obtained using an array scanner (FLA8000;Fuji Film, Tokyo, Japan) and the scanned signals were anal-ysed using Array Gauge software from Fuji Film.
The values gained from each spot were standardized toremove influences among slide glasses caused by uneven-ness of hybridization and differences in labelling efficiency.Since the distribution of the logarithmic value for each spotwas similar to normal distribution, the deviation valuesfrom all signal intensities were calculated using logarithmicvalues. Each spotted field consisted of 384 spots and wasregarded as a population for normalization. Changes of theexpression mass were estimated by the differences of thedeviation values between treatments. If the deviation valuein the –P treatment increased or decreased by more thanfive among all replications, they were defined as ‘up-regu-lated genes’ or ‘down-regulated genes’, respectively. Thenormalization and statistical analyses were performedusing Microsoft Excel 2001 (Microsoft, Redmond, WA,USA) on personal computers (Apple Computer, Cuper-tino, CA, USA). Each EST was annotated by the superiorknown gene identified in a B
LAST
search, and the signifi-cance of annotation was estimated by whether the e-valuewas under 1.0e
-
10.
Investigation of mRNA accumulation using quantitative real-time polymerase chain reaction
Isolated RNA was treated with DNase (RT grade; NipponGene, Tokyo, Japan) to digest the contaminated genomicDNA, then reverse-transcribed using the 1st Strand cDNASynthesis Kit for reverse transcription (RT)-polymerasechain reaction (PCR) (AMV) (Roche Diagnostics, Basel,Switzerland). The first stranded cDNAs were used as tem-plates for quantitative real-time PCR using the LightCycler system (Roche Diagnostics). The TaqStart antibody(Clontech) was used for repression of non-specific amplifi-cation and the
RAc1
gene was selected as the control genebecause it was reported that expression of this gene is rel-atively stable among actin isoforms in rice (McElroy
et al
.1990). Calibration curves were drawn with two replicationsbased on the specific plasmids.
RESULTS AND DISCUSSION
The P concentration in leaves was markedly decreased afterthe long-term –P treatment [7.1
±
0.05 mg
P
g
-
1
dry weight(DW) in the
+
P treatment, 2.0
±
0.20 mg
P
g
-
1
DW in thelong-term –P treatment]. In the long-term –P treatment,the fresh weight of shoots decreased significantly (160
±
1.5 mg plant
-
1
in the
+
P treatment, 107
±
1.7 mg plant
-
1
inthe long-term –P treatment), whereas the root fresh weightwas similar among the treatments (111
±
2.7 mg plant
-
1
in

Metabolic changes in P-deficient rice roots
1517
© 2003 Blackwell Publishing Ltd,
Plant, Cell and Environment,
26,
1515–1523
the
+
P treatment, 110
±
4.0 mg plant
-
1
in the long-term –Ptreatment).
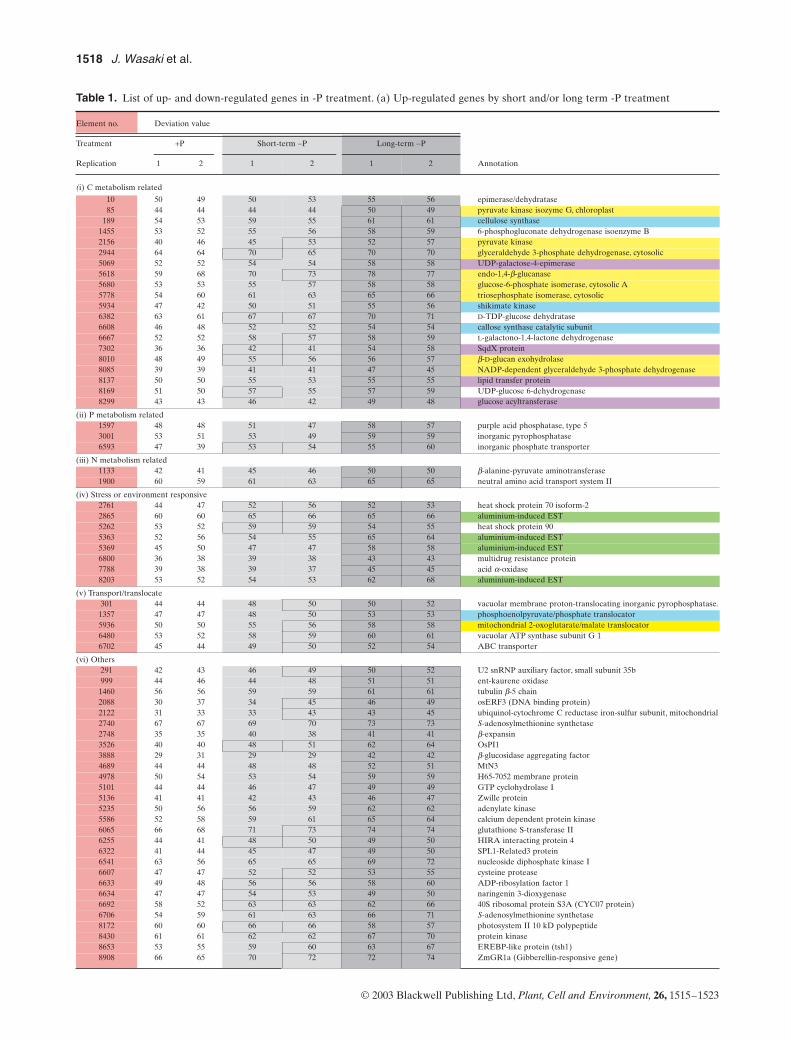
The genes up- and down-regulated by the –P treatmentare listed in Table 1. There were 15 up-regulated genes inthe short-term (24 h) and 86 in the long-term (9 d) –P treat-ment, whereas there were 23 and 97 down-regulated genesin the two treatments, respectively. Nine of 15 genes up-regulated, and 14 of 23 genes down-regulated in the short-term –P treatment showed similar regulation to the long-term –P treatment. Element no. 3526 showed the mostsignificant increase of its transcription in the long-term –Ptreatment. Its function was unknown, and was designated
OsPI1
(
Oryza sativa p
hosphate-limitation
i
nducible gene
1
;Wasaki
et al
. 2003). The functions of 46 (47%) of the down-regulated genes were known, whereas those of 63 (73%)were known for the up-regulated genes.
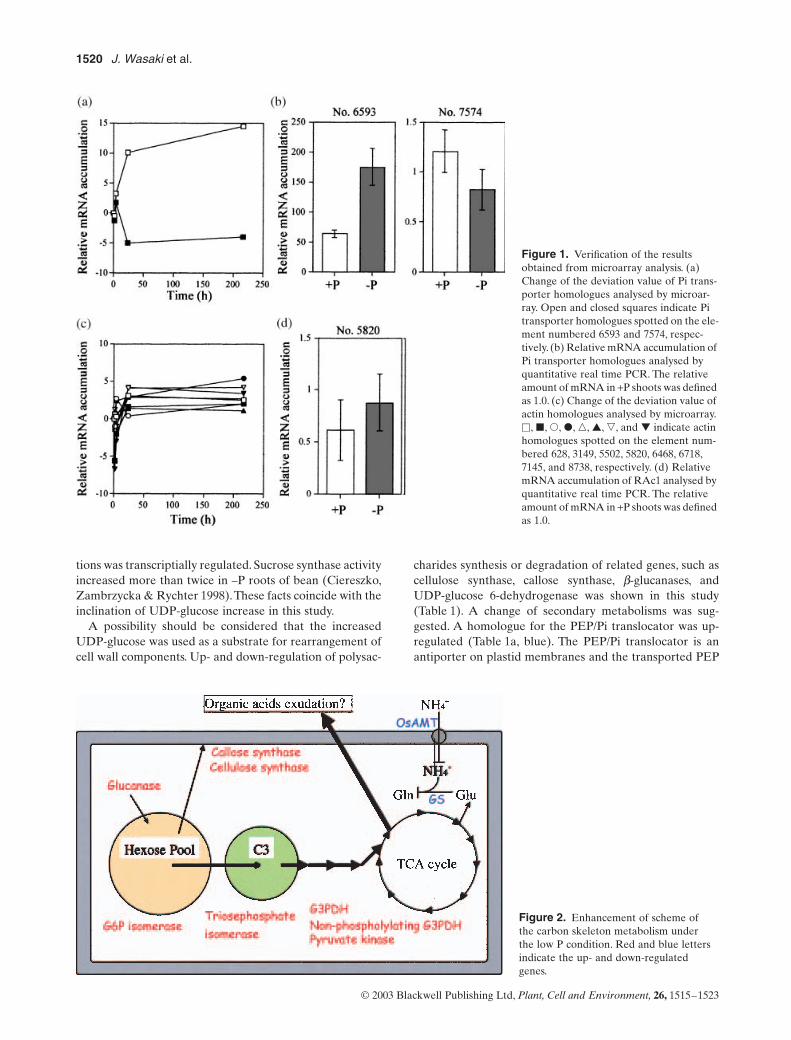
To evaluate the validity of microarray analysis, themRNA level of two genes encoding high affinity Pi trans-porters was quantified by RT-PCR. One of them was up-regulated, and the level of another decreased (Fig. 1a). Theresults of quantitative real-time PCR indicated similar pat-terns, that is, significant increase in no. 6593 and slightdecrease in no. 7574 (Fig. 1b). High affinity Pi transportergenes were isolated and characterized in several plants,such as
Catharanthus roseus
(Kai et al. 1997), Arabidopsisthaliana (Mucchal et al. 1996; Smith et al. 1997; Mudge et al.2002), Lycopersicon esculentum (Mucchal & Raghothama1999), Nicotiana tobaccum (Kai et al. 2002) and Lupinusalbus (Liu et al. 2001), some of which have been suggestedto play a role in high Pi absorption efficiency in roots underP-deficient conditions. It was also known that some Pitransporters are more constitutive in root tissues(Mucchal& Raghothama 1999; Liu et al. 2001; Mudge et al. 2002).
Microarray analysis suggested that the slope of theincrease of actin mRNA was similar in all isoforms(Fig. 1c). The reproducibility of the result was confirmed byquantitative real time PCR analysis about one of actin iso-forms, designated as RAc1 (Fig. 1d). It was reported thatthe RNase activities in plant cells increased under P limita-tion (Nürnberger et al. 1990; Löffler et al. 1992; Green 1994;Bosse & Köck 1998) and, thus, RNA pool may act as aphosphorus reservoir in the cell. As the result of a decreaseof total RNA pool, important mRNA molecules, such asactin, may increase relatively. The decreases of expressionof some photosynthesis-related genes are also consideredto present a similar phenomenon, because it is consideredthat these genes are not essential for root function.
del Pozo et al. (1999) reported that the expression of type5 acid phosphatase (APase) in shoots and roots of Arabi-dopsis increases under low P conditions. A homologue oftype 5 APase spotted on the microarray also increased inour assay (Table 1a, element no. 1597), indicating that thehomologue played a role in the efficient use of P in riceplants, similar to the Arabidopsis gene. However, anotherhomologue of type 5 APase decreased in the long-term –Ptreatment (Table 1b, element no. 8064). Further studiesfocused on the localization and enzymatic properties arerequired to clarify their function.
Genes up-regulated in the long-term –P treatment con-tained pyrophosphatase (PPiase) homologues (Table 1a,element no. 301, 3001). Pyrophosphate (PPi) is not onlyhydrolysed to two inorganic phosphate molecules, but alsohas a high-energy phosphate ester bond, which can replaceATP as an energy source. Palma, Blumwald & Plaxton(2000) reported that the activity and protein mass of vacu-olar type H+-PPiase increases under P-deficient conditions.In this study, both vacuolar H+-ATPase (element no. 301)and H+-PPiase (element no. 3001) were up-regulated, sug-gesting that their functions were related to the active trans-port between the vacuole and cytosol. The cDNAmicroarray used in this study does not include all genes ofrice. Nevertheless, alteration of numerous genes related tocarbon skeleton metabolism was indicated by the arrayanalysis. When the alteration fits on the metabolism map,important information is found (Fig. 2).
Several genes related to glycolysis increased their expres-sion level under the –P condition (Table 1a, yellow, Fig. 2);this provides a larger amount of carbon sources for thetricarboxylic acid (TCA) cycle. Uhde-Stone et al. (2003a)also reported that some glycolytic pathway related geneswere induced in –P proteoid roots, which were formed inwhite lupin lateral roots under the P-deficient conditions.Furthermore, because the level of OsAMT1, which encodesthe ammonia transporter, and glutamine synthase homo-logues were down-regulated in the –P roots (Table 1b, yel-low, Fig. 2), it seems that the carbon flow for amino acidsynthesis is repressed. Many plants secrete organic acidsunder P-deficient conditions to use the sparingly solubleinorganic phosphate compounds from roots (Gardner et al.1983). It has been reported that rice roots also secretecitrate (Kirk, Santos & Findenegg 1999). The necessity ofcarbon supply by the acceleration of glycolysis seemsappropriate for organic acid exudation. In the long-term–P treatment, some up- or down-regulated genes arerelated to lipid metabolism (Table 1, pink). It is interestingthat a rice homologue of the sqdX gene, required for sul-folipid synthesis, was up-regulated. The sqdX gene was iso-lated from a cyanobacterium with sqdA, sqdB, sqdC andsqdD, and consists of an operon for sulfoquinovosyl dia-cylglycerol (SQDG) synthesis (Benning & Somerville1992a,b; Rossak et al. 1995; Güler et al. 1996). It was pre-dicted that sqdX product catalyses the SQDG synthesizingreaction from UDP-sulfoquinovose and diacylglycerol,because of its homology with glycosyltransferase (Güler,Essigmann & Benning 2000). In higher plants, SQD1,homologous with sqdB of cyanobacteria, was isolated fromArabidopsis (Essigmann et al. 1998). P deficiency enhancesSQD1 expression and SQDG accumulation in leaves(Essigmann et al. 1998). As SQDG has the ability to substi-tute for phospholipids, it was suggested that the increase ofSQGD synthesis is available for the efficient use of P in themembrane (Essigmann et al. 1998). Gniazdowska, Szal &Rychter (1999) reported that the phospholipids of kidneybean roots were reduced by approximately half under a –Pcondition, although the total lipid content per protein ofplasma membrane fraction was not changed. It was under-
1518 J. Wasaki et al.
© 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 1515–1523
Table 1. List of up- and down-regulated genes in -P treatment. (a) Up-regulated genes by short and/or long term -P treatment
Element no. Deviation value
Annotation
Treatment +P Short-term -P Long-term -P
Replication 1 2 1 2 1 2
(i) C metabolism related
10 50 49 50 53 55 56 epimerase/dehydratase85 44 44 44 44 50 49 pyruvate kinase isozyme G, chloroplast
189 54 53 59 55 61 61 cellulose synthase1455 53 52 55 56 58 59 6-phosphogluconate dehydrogenase isoenzyme B2156 40 46 45 53 52 57 pyruvate kinase2944 64 64 70 65 70 70 glyceraldehyde 3-phosphate dehydrogenase, cytosolic5069 52 52 54 54 58 58 UDP-galactose-4-epimerase5618 59 68 70 73 78 77 endo-1,4-b-glucanase5680 53 53 55 57 58 58 glucose-6-phosphate isomerase, cytosolic A5778 54 60 61 63 65 66 triosephosphate isomerase, cytosolic5934 47 42 50 51 55 56 shikimate kinase6382 63 61 67 67 70 71 D-TDP-glucose dehydratase6608 46 48 52 52 54 54 callose synthase catalytic subunit6667 52 52 58 57 58 59 L-galactono-1,4-lactone dehydrogenase7302 36 36 42 41 54 58 SqdX protein8010 48 49 55 56 56 57 b-D-glucan exohydrolase8085 39 39 41 41 47 45 NADP-dependent glyceraldehyde 3-phosphate dehydrogenase8137 50 50 55 53 55 55 lipid transfer protein8169 51 50 57 55 57 59 UDP-glucose 6-dehydrogenase8299 43 43 46 42 49 48 glucose acyltransferase
(ii) P metabolism related1597 48 48 51 47 58 57 purple acid phosphatase, type 53001 53 51 53 49 59 59 inorganic pyrophosphatase6593 47 39 53 54 55 60 inorganic phosphate transporter
(iii) N metabolism related1133 42 41 45 46 50 50 b-alanine-pyruvate aminotransferase1900 60 59 61 63 65 65 neutral amino acid transport system II
(iv) Stress or environment responsive2761 44 47 52 56 52 53 heat shock protein 70 isoform-22865 60 60 65 66 65 66 aluminium-induced EST5262 53 52 59 59 54 55 heat shock protein 905363 52 56 54 55 65 64 aluminium-induced EST5369 45 50 47 47 58 58 aluminium-induced EST6800 36 38 39 38 43 43 multidrug resistance protein7788 39 38 39 37 45 45 acid a-oxidase8203 53 52 54 53 62 68 aluminium-induced EST
(v) Transport/translocate301 44 44 48 50 50 52 vacuolar membrane proton-translocating inorganic pyrophosphatase.1357 47 47 48 50 53 53 phosphoenolpyruvate/phosphate translocator5936 50 50 55 56 58 58 mitochondrial 2-oxoglutarate/malate translocator6480 53 52 58 59 60 61 vacuolar ATP synthase subunit G 16702 45 44 49 50 52 54 ABC transporter
(vi) Others291 42 43 46 49 50 52 U2 snRNP auxiliary factor, small subunit 35b999 44 46 44 48 51 51 ent-kaurene oxidase1460 56 56 59 59 61 61 tubulin b-5 chain2088 30 37 34 45 46 49 osERF3 (DNA binding protein)2122 31 33 33 43 43 45 ubiquinol-cytochrome C reductase iron-sulfur subunit, mitochondrial2740 67 67 69 70 73 73 S-adenosylmethionine synthetase2748 35 35 40 38 41 41 b-expansin3526 40 40 48 51 62 64 OsPI13888 29 31 29 29 42 42 b-glucosidase aggregating factor4689 44 44 48 48 52 51 MtN34978 50 54 53 54 59 59 H65-7052 membrane protein5101 44 44 46 47 49 49 GTP cyclohydrolase I5136 41 41 42 43 46 47 Zwille protein5235 50 56 56 59 62 62 adenylate kinase5586 52 58 59 61 65 64 calcium dependent protein kinase6065 66 68 71 73 74 74 glutathione S-transferase II6255 44 41 48 50 49 50 HIRA interacting protein 46322 41 44 45 47 49 50 SPL1-Related3 protein6541 63 56 65 65 69 72 nucleoside diphosphate kinase I6607 47 47 52 52 53 55 cysteine protease6633 49 48 56 56 58 60 ADP-ribosylation factor 16634 47 47 54 53 49 50 naringenin 3-dioxygenase6692 58 52 63 63 62 66 40S ribosomal protein S3A (CYC07 protein)6706 54 59 61 63 66 71 S-adenosylmethionine synthetase8172 60 60 66 66 58 57 photosystem II 10 kD polypeptide8430 61 61 62 62 67 70 protein kinase8653 53 55 59 60 63 67 EREBP-like protein (tsh1)8908 66 65 70 72 72 74 ZmGR1a (Gibberellin-responsive gene)
Metabolic changes in P-deficient rice roots 1519
© 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 1515–1523
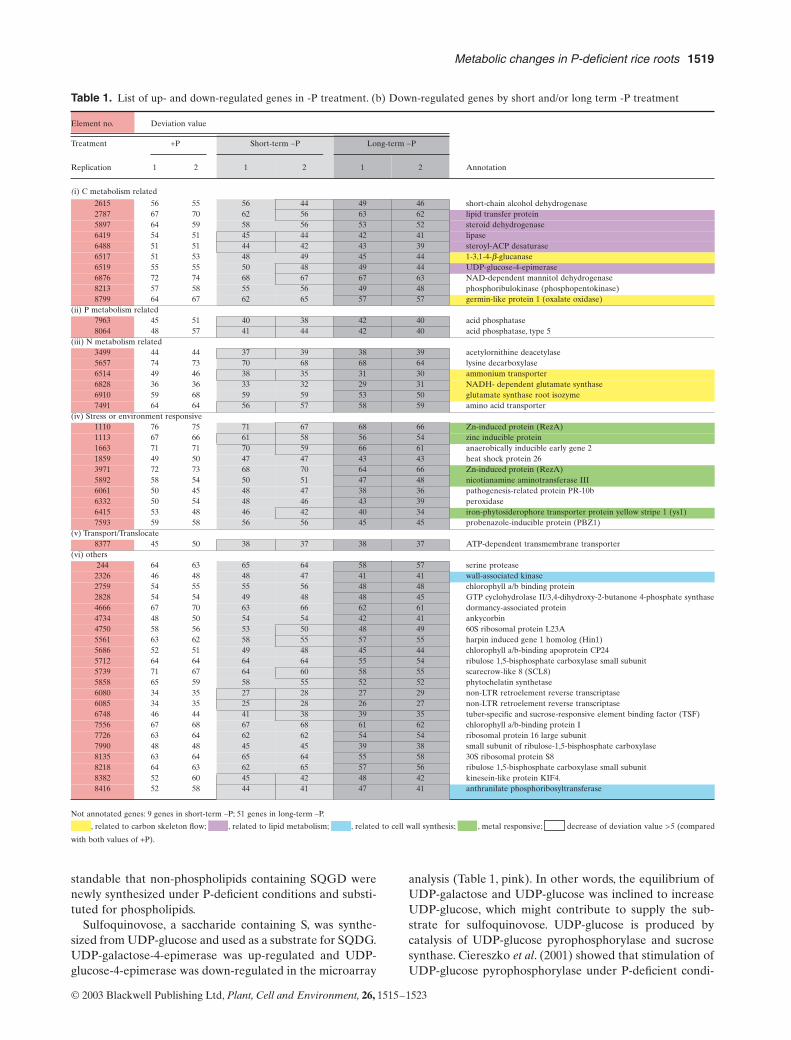
Table 1. List of up- and down-regulated genes in -P treatment. (b) Down-regulated genes by short and/or long term -P treatment
Element no. Deviation value
Annotation
Treatment +P Short-term -P Long-term -P
Replication 1 2 1 2 1 2
(i) C metabolism related
2615 56 55 56 44 49 46 short-chain alcohol dehydrogenase2787 67 70 62 56 63 62 lipid transfer protein5897 64 59 58 56 53 52 steroid dehydrogenase6419 54 51 45 44 42 41 lipase6488 51 51 44 42 43 39 steroyl-ACP desaturase6517 51 53 48 49 45 44 1-3,1-4-b-glucanase6519 55 55 50 48 49 44 UDP-glucose-4-epimerase6876 72 74 68 67 67 63 NAD-dependent mannitol dehydrogenase8213 57 58 55 56 49 48 phosphoribulokinase (phosphopentokinase)8799 64 67 62 65 57 57 germin-like protein 1 (oxalate oxidase)
(ii) P metabolism related7963 45 51 40 38 42 40 acid phosphatase8064 48 57 41 44 42 40 acid phosphatase, type 5
(iii) N metabolism related3499 44 44 37 39 38 39 acetylornithine deacetylase5657 74 73 70 68 68 64 lysine decarboxylase6514 49 46 38 35 31 30 ammonium transporter6828 36 36 33 32 29 31 NADH- dependent glutamate synthase6910 59 68 59 59 53 50 glutamate synthase root isozyme7491 64 64 56 57 58 59 amino acid transporter
(iv) Stress or environment responsive1110 76 75 71 67 68 66 Zn-induced protein (RezA)1113 67 66 61 58 56 54 zinc inducible protein1663 71 71 70 59 66 61 anaerobically inducible early gene 21859 49 50 47 47 43 43 heat shock protein 263971 72 73 68 70 64 66 Zn-induced protein (RezA)5892 58 54 50 51 47 48 nicotianamine aminotransferase III6061 50 45 48 47 38 36 pathogenesis-related protein PR-10b6332 50 54 48 46 43 39 peroxidase6415 53 48 46 42 40 34 iron-phytosiderophore transporter protein yellow stripe 1 (ys1)7593 59 58 56 56 45 45 probenazole-inducible protein (PBZ1)
(v) Transport/Translocate8377 45 50 38 37 38 37 ATP-dependent transmembrane transporter
(vi) others244 64 63 65 64 58 57 serine protease2326 46 48 48 47 41 41 wall-associated kinase2759 54 55 55 56 48 48 chlorophyll a/b binding protein2828 54 54 49 48 48 45 GTP cyclohydrolase II/3,4-dihydroxy-2-butanone 4-phosphate synthase4666 67 70 63 66 62 61 dormancy-associated protein4734 48 50 54 54 42 41 ankycorbin4750 58 56 53 50 48 49 60S ribosomal protein L23A5561 63 62 58 55 57 55 harpin induced gene 1 homolog (Hin1)5686 52 51 49 48 45 44 chlorophyll a/b-binding apoprotein CP245712 64 64 64 64 55 54 ribulose 1,5-bisphosphate carboxylase small subunit5739 71 67 64 60 58 55 scarecrow-like 8 (SCL8)5858 65 59 58 55 52 52 phytochelatin synthetase6080 34 35 27 28 27 29 non-LTR retroelement reverse transcriptase6085 34 35 25 28 26 27 non-LTR retroelement reverse transcriptase6748 46 44 41 38 39 35 tuber-specific and sucrose-responsive element binding factor (TSF)7556 67 68 67 68 61 62 chlorophyll a/b-binding protein I7726 63 64 62 62 54 54 ribosomal protein 16 large subunit7990 48 48 45 45 39 38 small subunit of ribulose-1,5-bisphosphate carboxylase8135 63 64 65 64 55 58 30S ribosomal protein S88218 64 63 62 65 57 56 ribulose 1,5-bisphosphate carboxylase small subunit8382 52 60 45 42 48 42 kinesein-like protein KIF4.8416 52 58 44 41 47 41 anthranilate phosphoribosyltransferase
Not annotated genes: 9 genes in short-term –P; 51 genes in long-term –P.
, related to carbon skeleton flow; , related to lipid metabolism; , related to cell wall synthesis; , metal responsive; decrease of deviation value >5 (compared
with both values of +P).
standable that non-phospholipids containing SQGD werenewly synthesized under P-deficient conditions and substi-tuted for phospholipids.
Sulfoquinovose, a saccharide containing S, was synthe-sized from UDP-glucose and used as a substrate for SQDG.UDP-galactose-4-epimerase was up-regulated and UDP-glucose-4-epimerase was down-regulated in the microarray
analysis (Table 1, pink). In other words, the equilibrium ofUDP-galactose and UDP-glucose was inclined to increaseUDP-glucose, which might contribute to supply the sub-strate for sulfoquinovose. UDP-glucose is produced bycatalysis of UDP-glucose pyrophosphorylase and sucrosesynthase. Ciereszko et al. (2001) showed that stimulation ofUDP-glucose pyrophosphorylase under P-deficient condi-
1520 J. Wasaki et al.
© 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 1515–1523
tions was transcriptially regulated. Sucrose synthase activityincreased more than twice in –P roots of bean (Ciereszko,Zambrzycka & Rychter 1998). These facts coincide with theinclination of UDP-glucose increase in this study.
A possibility should be considered that the increasedUDP-glucose was used as a substrate for rearrangement ofcell wall components. Up- and down-regulation of polysac-
charides synthesis or degradation of related genes, such ascellulose synthase, callose synthase, b-glucanases, andUDP-glucose 6-dehydrogenase was shown in this study(Table 1). A change of secondary metabolisms was sug-gested. A homologue for the PEP/Pi translocator was up-regulated (Table 1a, blue). The PEP/Pi translocator is anantiporter on plastid membranes and the transported PEP
Figure 1. Verification of the results obtained from microarray analysis. (a) Change of the deviation value of Pi trans-porter homologues analysed by microar-ray. Open and closed squares indicate Pi transporter homologues spotted on the ele-ment numbered 6593 and 7574, respec-tively. (b) Relative mRNA accumulation of Pi transporter homologues analysed by quantitative real time PCR. The relative amount of mRNA in +P shoots was defined as 1.0. (c) Change of the deviation value of actin homologues analysed by microarray. �, �, �, �, �, ▲, �, and ▼ indicate actin homologues spotted on the element num-bered 628, 3149, 5502, 5820, 6468, 6718, 7145, and 8738, respectively. (d) Relative mRNA accumulation of RAc1 analysed by quantitative real time PCR. The relative amount of mRNA in +P shoots was defined as 1.0.
Figure 2. Enhancement of scheme of the carbon skeleton metabolism under the low P condition. Red and blue letters indicate the up- and down-regulated genes.

Metabolic changes in P-deficient rice roots 1521
© 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 1515–1523
into the plastids could be used as a substrate of shikimatekinase (Hausler et al. 2000), whose homologue was up-reg-ulated (Table 1a, blue). The Trp synthesis pathway wasassumed to be repressed by the down-regulation of anthra-nilate phosphoribosyltransferase homologue (Table 1b,blue). On the other hand, members of the Tyr/Phe synthesispathway were not influenced. Phe is one of the substratesfor anthocyanin synthesis, and the accumulation of antho-cyanin is known under P starvation (Bariola, MacIntosh &Green 1999). Another research using macroarray of whitelupin revealed up-regulation of lignin synthesis-relatedgenes in –P proteoid roots of white lupin (Uhde-Stone et al.2003b). An increase of the lignin content may relate to therearrangement of cell wall components.
Generally, P is over-accumulated under Zn-deficientand P-sufficient conditions (Huang et al. 2000). Huanget al. (2000) explained that this accumulation is caused byover-expression of a high affinity Pi transporter, which isup-regulated in Zn-deficient barley independently of theP status. It was considered that Zn has a role in regulat-ing the P starvation inducible molecular mechanism withsome molecules, such as RezA of rice (the function is notstill clarified; accession No. AAA87049), which wasdown-regulated in our long-term –P treatment (Table 1b,green).
When Poaceae plants are grown under Fe deficiency,their roots exude phytosiderophore to obtain Fe frominsoluble forms (Takagi 1976; Takagi, Nomoto & Take-moto 1984). In this study, homologues for nicotianamineaminotransferase and yellow stripe1 (transporter ofFe(III)-phytosiderophore in maize; Curie et al. 2001)encoding a transporter of Fe (III)-phytosiderophore inmaize were down-regulated in the long-term –P treatment(Table 1b, green). It is possible that Fe and Pi will forminsoluble compounds in the +P nutrient solution. This indi-cates that the +P plants would initially fail under a slightiron deficiency, then their phytosiderophore system wouldrespond.
Wang et al. (2001, 2002) made a macroarray spotted1280 mineral nutrient-related cDNAs of tomato and anal-ysed the responses on P, K and Fe deficiencies. They iden-tified some genes previously not associated with P, K andFe nutrition, and suggested the co-ordination and co-regulation of uptake of these mineral nutrients. Althoughthere is a qualitative difference between our study andtheirs in aspects of the period of treatments and light con-dition, P responsible genes associated with N, Zn, Fe andAl were also identified in this study. To clarify the wholemolecular mechanism of plant strategy to adapt to P defi-ciency, it will be necessary to consider the cross talk amongresponses to different elements.
ACKNOWLEDGMENTS
This study was supported by a grant from the Ministry ofAgriculture, Forestry and Fisheries of Japan (Rice GenomeProject MA-2111).
REFERENCES
Bariola P.A., MacIntosh G.C. & Green P.J. (1999) Regulation ofS-like ribonuclease levels in Arabidopsis. Antisense inhibitionof RNS1 or RNS2 elevates anthocyanin accumulation. PlantPhysiology 119, 331–342.
Benning C. & Somerville C.R. (1992a) Isolation and genetic com-plementation of a sulfolipid-deficient mutant of Rhodobactersphaeroides. Journal of Bacteriology 174, 2352–2360.
Benning C. & Somerville C.R. (1992b) Identification of an operoninvolved in sulfolipid biosynthesis in Rhodobacter sphaeroides.Journal of Bacteriology 174, 6479–6487.
Bergman L.W., McClinton D.C., Madden S.L. & Peris L.H. (1986)Molecular analysis of the DNA sequences involved in the tran-scriptional regulation of the phosphate-repressible acid phos-phatase (PHO5) of Saccharomyces cerevisiae. Proceedings of theNational Academy of Sciences of USA 83, 6070–6074.
Bosse D. & Köck M. (1998) Influence of phosphate starvation onphosphohydrolases during development of tomato seedlings.Plant, Cell and Environment 21, 325–332.
Ciereszko I., Johansson H., Hurry V. & Kleczkowski L.A. (2001)Phosphate status affects the gene expression, protein contentand enzymatic activity of UDP-glucose pyrophosphorylase inwild-type and pho mutants of Arabidopsis. Planta 212, 598–605.
Ciereszko I., Zambrzycka A. & Rychter A. (1998) Sucrose hydrol-ysis in bean roots (Phaseolus vulgaris L.) under phosphate defi-ciency. Plant Science 133, 139–144.
Curie C., Panaviene Z., Loulergue C., Dellaporta S.L., Briat J.F.& Walker E.L. (2001) Maize yellow stripe1 encodes a mem-brane protein directly involved in Fe (III) uptake. Nature 409,346–349.
Duff S.M.G., Moorhead G.B.G., Lefevbre D.D. & Plaxton W.C.(1989) Phosphate starvation inducible bypasses of adenylate andphosphate dependent glycolytic enzymes in Brassica nigra sus-pension cells. Plant Physiology 90, 1275–1278.
Duff S.M.G., Plaxton W.C. & Lefebvre D.D. (1991) Phosphate-starvation response in plant cells: De novo synthesis anddegradation of acid phosphatases. Proceedings of the NationalAcademy of Sciences of USA 88, 9538–9542.
Essigmann B., Güler S., Narang R.A., Linke D. & Benning C.(1998) Phosphate availability affects the thylakoid lipid compo-sition and the expression of SQD1, a gene required for sulfolipidbiosynthesis in Arabidopsis thaliana. Proceedings of the NationalAcademy of Sciences of USA 95, 1950–1955.
Gardner W.K., Barber D.A. & Parbey D.G. (1983) The acquisitionof phosphorus by Lupinus albus L. III. The probable mechanismby which phosphorus movement in the soil/root interface isenhanced. Plant and Soil 70, 107–124.
Gniazdowska A., Szal B. & Rychter A.M. (1999) The effect ofphosphate deficiency on membrane phospholipid compositionof bean (Phaseolus vulgaris L.) roots. Acta Physiologiae Plan-tarum 21, 263–269.
Goff S.A., Ricke D., Lan T.-H., et al. (2002) A draft sequence ofthe rice genome (Oryza sativa L. ssp. japonica). Science 296, 92–100.
Goldstein A.H., Baretlein D.A. & Danon A. (1989) Phosphatestarvation stress an experimental system for molecular analysis.Plant Molecular Biology Reporter 7, 7–16.
Green P.J. (1994) The ribonucleases of higher plants. AnnualReview of Plant Physiology and Plant Molecular Biology 45,421–445.
Güler S., Essigmann B. & Benning C. (2000) A cyanobacterialgene, sqdX, required for biosynthesis of the sulfolipid sulfouino-vosyldiacylglycerol. Journal of Bacteriology 182, 543–545.
Güler S., Seelinger A., Härtel H., Renger G. & Benning C. (1996)A null mutant of Synechococcus sp. PCC7942 deficient in the

1522 J. Wasaki et al.
© 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 1515–1523
sulfolipid sulfoquinovosyldiacylglycerol. Journal of BiologicalChemistry 271, 7501–7507.
Hausler R.E., Baur B., Scharte J., Teichmann T., Eicks M., FischerK.L., Flugge U.I., Schubert S., Weber A. & Fischer K. (2000)Plastidic metabolite transporters and their physiological func-tions in the inducible crassulacean acid metabolism plant Mes-sembryanthemum crystallinum. Plant Journal 24, 285–296.
Huang C., Barker S.J., langridge P., Smith F.W. & Graham R.D.(2000) Zinc deficiency up-regulates expression of high-affinityphosphate transporeter genes in both phosphate-sufficientand – deficient barley roots. Plant Physiology 124, 415–422.
Kai M., Masuda Y., Kikuchi Y., Osaki M. & Tadano T. (1997)Isolation and characterization of a cDNA from Catharanthusroseus which is highly homologous with phosphate transporter.Soil Science and Plant Nutrition 43, 227–235.
Kai M., Takazumi K., Adachi H., Wasaki J., Shinano T. & OsakiM. (2002) Cloning and characterization of four phosphate trans-porter cDNAs in tobacco. Plant Science 163, 837–846.
Kirk G.J.D., Santos E.E. & Findenegg G.R. (1999) Phosphatesolubilization by organic anion excretion from rice (Oryza sativaL.) growing in aerobic soil. Plant and Soil 211, 11–18.
Kishimoto N., Yazaki J., Fujii F., Shimbo K., Ohta T., ShimataniZ., Nagata Y., Hashimoto A. & Kikuchi S. (2002) Rice cDNAmicroarray: a powerful tool for transcriptome analysis in ricefunctional genomics. Recent Research Developments in PlantBiology 2, 49–59.
Liu J., Uhde-Stone C., Li A., Vance C. & Allan D. (2001) Aphosphate transporter with enhanced expression in proteoidroots of white lupin (Lupinus albus L.). Plant and Soil 237, 257–266.
Löffler A., Abel S., Jost W., Beintema J.J. & Glund K. (1992)Phosphate-regulated induction of intracellular ribonuclease incultured tomato (Lycopersicon esculentum) cells. Plant Physiol-ogy 98, 1472–1478.
McElroy D., Rothenberg M., Reece K.S. & Wu R. (1990) Charac-terization of the rice (Oryza sativa) actin gene family. PlantMolecular Biology 15, 257–268.
Mucchal U.S., Pardo J.M. & Raghothama K.G. (1996) Phosphatetransporters from the higher plant Arabidopsis thaliana. Pro-ceedings of the National Academy of Sciences of USA 93, 10519–10523.
Mucchal U.S. & Raghothama K.G. (1999) Transcriptional regula-tion of plant phosphate transporters. Proceedings of the NationalAcademy of Sciences of USA 96, 5868–5872.
Mudge S.R., Rae A.L., Diatloff E. & Smith F.W. (2002) Expres-sion analysis suggests novel roles for members of the Pht1 familyof phosphate transporters in Arabidpsis. Plant Journal 31, 341–353.
Negishi T., Nakanishi H., Yazaki J., Kishimoto N., Fujii F.,Shimbo K., Yamamoto K., Sakata K., Sasaki T., Kikuchi S.,Mori S. & Nishizawa N.K. (2002) cDNA microarray analysis ofgene expression during Fe-deficiency stress in barley suggeststhat polar transport of vesicles is implicated in phytosidero-phore secretion in Fe-deficient barley roots. Plant Journal 30,83–94.
Nürnberger T., Abel S., Jost W. & Glund K. (1990) Induction ofan extracellular ribonuclease in cultured tomato cells uponphosphate starvation. Plant Physiology 92, 970–976.
Palma D.A., Blumwald E. & Plaxton W.C. (2000) Upregulation ofvacuolar H+-translocating pyrophosphatase by phosphate star-vation of Brassica napus (rapeseed) suspension cell cultures.FEBS Letter 486, 155–158.
Palmiter R.D. (1974) Magnesium precipitation of ribonucleopro-tein complexes. Expedient techniques for the isolation of under-graded polysomes and messenger ribonucleic acid. Biochemistry13, 3606–3615.
del Pozo J.C., Allona I., Rubio V., Leyva A., de la Peña A.,Aragoncillo C. & Paz-Ares J. (1999) A type 5 acid phosphatasegene from Arabidopsis thaliana is induced by phosphate starva-tion and by some other types of phosphate mobilising/oxidativestress conditions. Plant Journal 19, 579–589.
Rossak M., Tietje C., Heinz E. & Benning C. (1995) Accumulationof UDP-sulfoquinovose in a sulfolipid-deficient mutant ofRhodobacter sphaeroides. Journal of Biological Chemistry 270,25792–25797.
Saheki S., Takeda A. & Shimizu T. (1985) Assay of inorganicphosphate in the mild pH range, suitable for measurement ofglycogen phosphorylase activity. Analytical Biochemistry 13,3605–3615.
Sambrook J., Fritsch E.F. & Maniatis T. (1989) Molecular Cloning,2nd edn. Cold Spring. Harbor Laboratory Press, Cold SpringHarbor, NY, USA.
Smith F.W., Ealing P.M., Dong B. & Delhaize E. (1997) Thecloning of two Arabidopsis genes belonging to a phosphatetransporter family. Plant Journal 11, 83–92.
Tadano T. & Sakai H. (1991) Secretion of acid phosphatase by theroots of several crop species under phosphorus-deficient condi-tions. Soil Science and Plant Nutrition 37, 129–140.
Takagi S. (1976) Naturally occurring iron-chelating compounds inoat- and rice-root washing. I. Activity measurement and prelim-inary characterization. Soil Science and Plant Nutrition 22, 4232–4233.
Takagi S., Nomoto K. & Takemoto S. (1984) Physiological aspectof mugineic acid, a possible phytosiderophore of graminaceousplants. Journal of Plant Nutrition 7, 469–477.
The Arabidopsis Genome Initiative (2000) Analysis of the genomesequence of the flowering plant Arabidopsis thaliana. Nature408, 796–815.
Thimm O., Essigmann B., Kloska S., Altmann T. & Buckhout T.J.(2001) Response of Arabidopsis to iron deficiency stress asrevealed by microarray analysis. Plant Physiology 127, 1030–1043.
Torriani A. & Ludtke D.N. (1985) The Pho regulon of Escherichiacoli. In The Molecular Biology of Bacterial Growth (eds M.Shaechter, F.C. Neidhardt, J. Ingraham & N.O. Kjeldgaard), pp.224–242. Jones & Bartlett, Boston, MA, USA.
Uhde-Stone C., Gilbert G., Johnson J.M.-F., Litjens R., Zinn K.E.,Temple S.J., Vance C.P. & Allan D.L. (2003a) Acclimation ofwhite lupin to phosphorus deficiency involves enhanced expres-sion of genes related to organic acid metabolism. Plant and Soil248, 99–116.
Uhde-Stone C., Zinn K.E., Ramirez-Yáñez M., Li A., Vance C.P.& Allan D.L. (2003b) Nylon filter arrays reveal differential geneexpression in proteoid roots of white lupin in response to phos-phorus deficiency. Plant Physiology 131, in press.
Wang Y.-H., Gravin D.F. & Kochian L.V. (2001) Nitrate-inducedgenes in tomato roots. Array analysis reveals novel genes thatmay play a role in nitrogen nutrition. Plant Physiology 127, 345–359.
Wang Y.-H., Gravin D.F. & Kochian L.V. (2002) Rapid inductionof regulatory and transporter genes in response to phosphorus,potassium, and iron deficiencies in tomato roots. Evidence forcross talk and root/rhizosphere-mediated signals. Plant Physiol-ogy 130, 1361–1370.
Wang R., Guegler K., LaBrie S.T. & Crawford N.M. (2000)Genomic analysis of a mutant response in Arabidopsisreveals diverse expression patterns and novel metabolic andpotential regulatory genes induced by nitrate. Plant Cell 12,1491–1509.
Wasaki J., Omura M., Ando M., Dateki H., Shinano T., Osaki M.,Ito H., Matsui H. & Tadano T. (2000) Molecular cloning androot specific expression of secretory acid phosphatase from

Metabolic changes in P-deficient rice roots 1523
© 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 1515–1523
phosphate deficient lupin (Lupinus albus L.). Soil Science andPlant Nutrition 46, 427–437.
Wasaki J., Yonetani R., Shinano T., Kai M. & Osaki M. (2003)Expression of the OsPI1 gene, cloned from rice roots usingcDNA microarray, rapidly responds to phosphorus status. NewPhytologist 158, 239–248.
Yu J., Hu S., Wang J., et al. (2002) A draft sequence of the ricegenome (Oryza sativa L. ssp. indica). Science 296, 79–92.
Received 19 March 2003; accepted for publication 4 May 2003





























