Embed Size (px)
Citation preview

Journal of Zmmunogenetics (1986) 13,187-194.
T R A N S C R I P T I O N A L C O N T R O L O F M H C CLASS I1 A N T I G E N E X P R E S S I O N O N MOUSE T C E L L
L I N E S
s. K. S I N G H , K. A. D O N O V A N A N D c. S . DAVID
Department of Immunology, Mayo Clinic, Rochester, MN 55905 U S A .
S U M M A R Y
We have analysed the factors which regulate MHC class I1 expression in mouse T cell lines. Two such lines, BW 5147 and PLT-24.2, were used in this study. Using 5-azacytidine (5 AzaC) we have shown that hypomethylation of DNA can induce class I1 antigen synthesis in BW 5147. The expression of class I1 in PLT-24.2 cells seems to be under a different control mechanism. Southern blot analysis of I-Ap gene in PLT-24.2 suggests that the expression of class I1 in this cell line is probably the outcome of a gene rearrangement. We hypothesise that insertion of viral long terminal repeats (LTR) next to the class I1 genes in transformed T cell lines can act as a promoter for the expression of class I1 antigens.
I N T R O D U C T I O N
I-region associated (Ia) antigens are class I1 major histocompatibility complex (MHC) encoded polypeptides. The four class I1 polypeptides in mice, A,, Ap, E,, E,, are expressed predominantly on B lymphocytes and certain macrophage populations that mediate a wide variety of immunological phenomena, including the activation of lymphocytes in mixed lymphocyte culture (Me0 et al., 1975), the generation of cytotoxic lymphocytes (Klein, 1978) as well as in MHC restriction (Klein & Nagy, 1982). In contrast to the presence of class I antigens on all nucleated cells, class I1 antigens are limited in their tissue distribution (David et al., 1976). Several studies have shown that expression of class I1 molecules is under regulatory control. Cell surface expression of class I1 antigens is under both positive and negative control. Class I1 antigens are inducible in some cell lines by gamma interferon (IFN-y) (King & Jones, 1983). This effect appears to act at the level of transcription, since addition of IFN-y into cultures of mouse macrophage lines results in an increase in class I1 mRNA (Nakamura et al., 1984). Conversely, substances such as prostaglandins have been shown to negatively regulate expression of Ia antigens on cultured mouse macrophages (Snyder et al., 1982). More recently, work from our laboratory and others has suggested a role for DNA methylation in regulation of class I1 gene expression (Peterlin et al., 1984; S i g h et a/., 1985).
Address for reprints: Dr S. K. Singh, Department of Immunology, Mayo Clinic, Rochester, MN 55905 U.S.A.
187

188 S. K . Singh, K . A . Donovan and C . S . David Studies to date on the regulation of class I1 gene expression suggest that it is under the
control of multiple mechanisms. In the present study we have explored the regulation of class I1 expression, since these molecules are important in mediating various immune functions and play a role in autoimmune diseases. The present study analyses the transcription of class I1 mRNA and expression of class I1 antigens by two mouse T cell lines BW 5 147 (Gruetzmann & Hammerling, 1978) and PLT-24.2 (Singh et al., 1984).
M A T E R I A L S A N D M E T H O D S
Cell lines BW 5147 was obtained from Dr Chris Krco, Mayo Clinic. PLT-24.2 was generated in
our laboratory and has been described earlier (Singh et al., 1984). Both cell lines were maintained in RPMI 1640,5% (v/v) fetal calf serum and antibiotics.
Induction of hypomethylation BW 5147 was cultured in the presence of 10-20 pg/ml of 5-azacytidine (5 AzaC) for
2 4 4 8 h. The cells were harvested and washed in phosphate-buffered saline (PBS) and cultured for another 72 h in RPMI 1640 medium. The cells were harvested and analysed for the expression of class I1 antigens by indirect immunofluorescence.
Indirect immunofluorescence Exponentially dividing cells were harvested, reacted with anti-class I1 monoclonal
antibodies (mAbs) and subjected to analysis using the fluorescence-activated cell sorter as described elsewhere (Singh et al., 1985).
Isolation of chromosomal DNA Cells from culture were washed three times with PBS, and resuspended in 15 ml of
10 m~ Tris and 55 mM ethylenediamine tetraacetic acid (pH 7.5). Protease K (500 pg/ml), followed by 20% (w/v) sodium dodecyl sulphate (final concentration 0.5%) was added. The suspension was incubated 3 h overnight at 37OC. The DNA was extracted once with phenol, twice with chloroform/isoamyl alcohol (24 : l), ethanol precipitated and resuspended in distilled water. The DNA was then digested with RNase (50 pg/ml) at 37OC for 1 h. The DNA was again extracted once with phenol, once with chloroform/isoamyl alcohol and then ethanol precipitated.
Restriction enzyme analysis of chromosomal DNA Restriction endonuclease digestions were performed according to the manufacturers’
recommendations for 6 h with 4 U of enzyme per pg of DNA. Fifteen micrograms of completely digested DNA were electrophoresed in agarose gel, transferred onto nylon membranes (Amersham Co.) and hybridized to the class II specific probe. Hybridizations were performed for 16 h at 65°C in a solution containing 6 x SSC, 5 x Denhardt’s, 10% (w/v) dextran sulphate, 100 &mi salmon sperm DNA and 0.5% (w/s) SDS. Filters were washed extensively at 65OC in 2 x SSC and 0.5% SDS for 1 h and exposed to x-ray film at -7OOC. In some cases higher stringency washes in 0.1 x SSC and 0.5% SDS were performed.
MHC class 11 expression on T cell lines 189
The class I1 probe was a generous gift from Dr Lee Hood. pCA 12 is a 2.4 kb Hind I11 DNA probes
fragment and recognizes the 5'-end of the I-A, gene.
Monoclonal antibodies
Hammerling and Dr D. Sachs, respectively. Anti-class I1 mAb 17-227 (anti-Ak) and 13-4 (anti-Ek) were kindly provided by Dr G.
RESULTS
Induction of hypomethylation and class ZZ antigen expression In order to study the role of DNA methylation, BW 5 147, a T cell line, was treated with
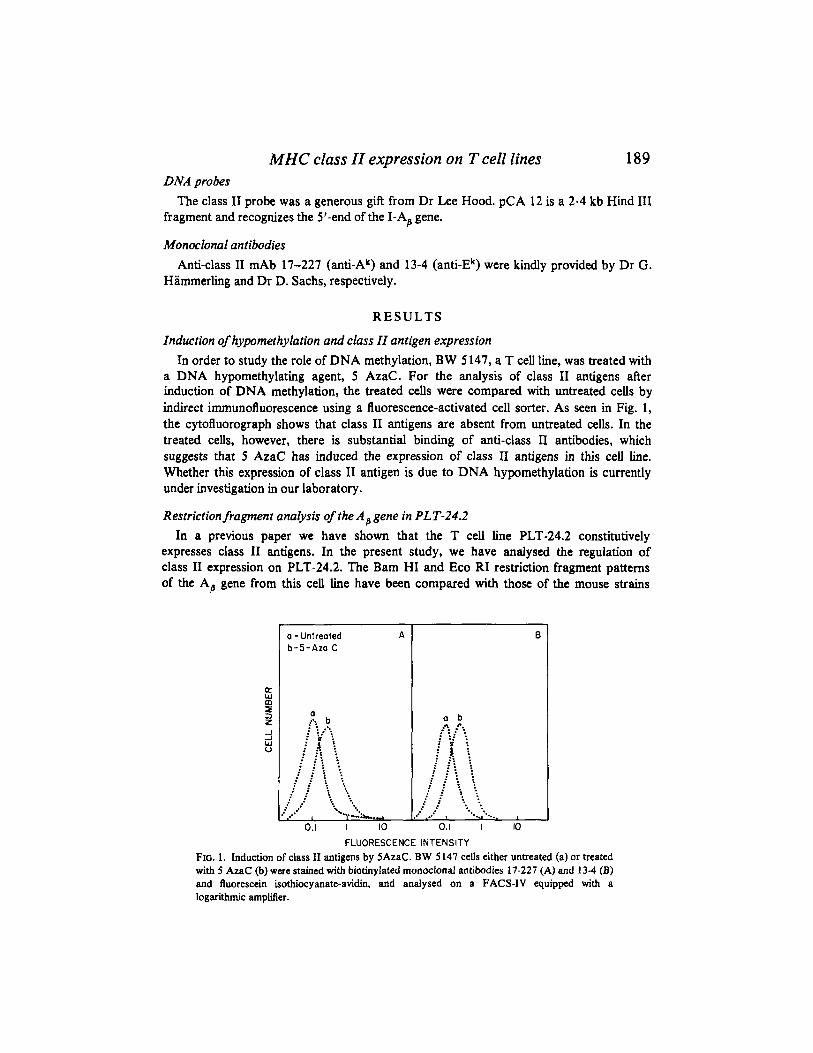
a DNA hypomethylating agent, 5 AzaC. For the analysis of class I1 antigens after induction of DNA methylation, the treated cells were compared with untreated cells by indirect immunofluorescence using a fluorescence-activated cell sorter. As seen in Fig. 1, the cytofluorograph shows that class I1 antigens are absent from untreated cells. In the treated cells, however, there is substantial binding of anti-class I1 antibodies, which suggests that 5 AzaC has induced the expression of class I1 antigens in this cell line. Whether this expression of class I1 antigen is due to DNA hypomethylation is currently under investigation in our laboratory.
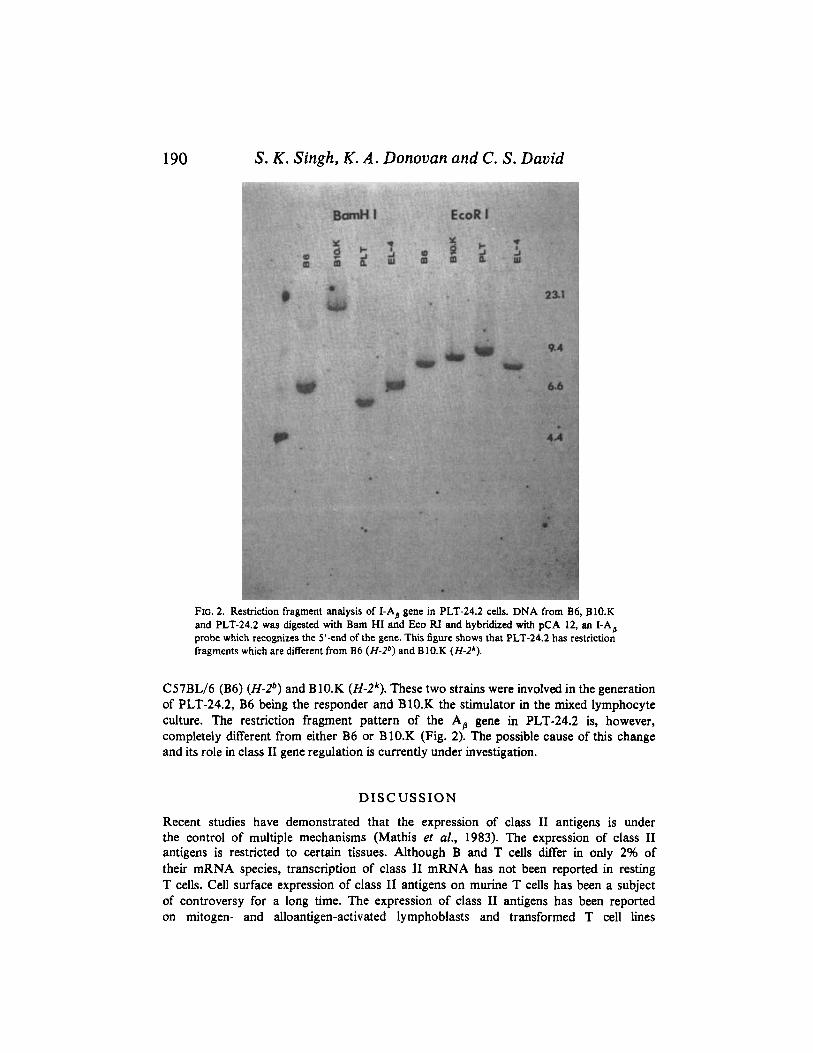
Restriction fragment analysis of the A , gene in PLT-24.2 In a previous paper we have shown that the T cell line PLT-24.2 constitutively
expresses class I1 antigens. In the present study, we have analysed the regulation of class I1 expression on PLT-24.2. The Bam HI and Eco RI restriction fragment patterns of the A,, gene from this cell line have been compared with those of the mouse strains
____ ~~
a -Untreated A b - 5 - A z a C
B
FLUORESCENCE INTENSITY FIG. 1. Induction of class I1 antigens by 5AzaC. BW 5 147 cells either untreated (a) or treated with 5 AzaC (b) were stained with biotinylated monoclonal antibodies 17-227 (A) and 13-4 (B) and fluorescein isothiocyanate-avidi, and analysed on a FACS-IV equipped with a logarithmic amplifier.
190 S. K . Singh, K . A . Donovan and C . S . David
FIG. 2. Restriction fragment analysis of LAB gene in PLT-24.2 cells. DNA from B6, B1O.K and PLT-24.2 was digested with Bam HI and Eko RI and hybridized with pCA 12, an LAp probe which recognizes the 5’-end of the gene. This figure shows that PLT-24.2 has restriction fragments which are different from B6 (H-2”) and B 10.K (H-29.
C57BL/6 (B6) (H-2”) and B1O.K (H-29. These two strains were involved in the generation of PLT-24.2, B6 being the responder and BIO.K the stimulator in the mixed lymphocyte culture. The restriction fragment pattern of the A, gene in PLT-24.2 is, however, completely different from either B6 or B1O.K (Fig. 2). The possible cause of this change and its role in class I1 gene regulation is currently under investigation.
D I S C U S S I O N
Recent studies have demonstrated that the expression of class I1 antigens is under the control of multiple mechanisms (Mathis et al., 1983). The expression of class I1 antigens is restricted to certain tissues. Although B and T cells differ in only 2% of their mRNA species, transcription of class I1 mRNA has not been reported in resting T cells. Cell surface expression of class I1 antigens on murine T cells has been a subject of controversy for a long time. The expression of class I1 antigens has been reported on mitogen- and alloantigen-activated lymphoblasts and transformed T cell lines

MHC class II expression on T cell lines 191 (David el al., 1976; Lorber et al., 1982). The expression of HLA-DR antigens, however, is readily observed on a population of T cells (De Wolfet al., 1979; Indiveri et al., 1980).
In the present study we have attempted to address the question of what controls the transcription of class I1 genes in T cells. It is becoming increasingly apparent that most eukaryotic genes are under multiple control. The expression of immunoglobulins in B lymphocytes is preceded by gene rearrangement (Maki et al., 1980). A similar observation has been made for T cell receptor genes in activated T cells (Hedrick el al., 1984). The rearrangement of class I1 genes has not been reported, however. The structure of class I1 genes as studied by restriction fragment analyses is the same in different tissues (Steinmetz et al., 1982). Recently, DNA methylation as a controlling factor in MHC gene expression has been under investigation. Peterlin and colleagues (1984) have shown that in U 937, a monocyte cell line, the expression of HLA-DR genes is controlled by two factors, One is the methylation of genes, which can be reversed by the DNA demethylating agent 5 AzaC, and the other is IFN-y, which promotes the appearance of HLA-DR transcripts. Similar observations have been reported by Reitz and co-workers (1984), where the expression of HLA-DR antigens in HTLV-infected T cells is the result of DNA hypomethylation. In the present study, we have looked at the relationship of DNA methylation and class I1 antigen expression in a mouse T cell line, BW 5147. The pyrimidine analogue 5 AzaC is a potent effector of decreased DNA methylation in mammalian cells. 5 AzaC has been shown to substitute for cytosine residues in DNA and to interfere with the maintenance of methylation by binding the methyltransferase enzyme irreversibly at the AzaC sites and thus, in effect, blocking further activity (Santi et al., 1983). In the present experiment, BW 5147 was cultured in vitro in the presence of 5 AzaC. As has been described earlier, 5 AzaC induces class I1 antigen expression in the BW 5 147 cell line. In order to be certain that the 5 AzaC causes demethylation of class I1 genes in BW 5147, we are currently analysing DNA isolated from treated and untreated cells by Southern blot analysis. In this study, we use the Hpa II/Msp I endonuclease system to examine the correlation between methylation of C,G sites and class I1 antigen expression. Restriction enzymes Hpa I1 and Msp I recognize the same sequence CCGG, but differ in that Hpa I1 will not cut if the internal cytosine is methylated. These studies will help us to understand the role of methylation in class I1 gene expression. A possible role of DNA methylation in the regulation of class I1 antigen expression has been depicted in Fig. 3.
The expression of class I1 antigens in PLT-24.2 seems to be under a different control
A gene B
5'-3i + No protein
No m-RNA 3' -5,
5-azacytidine
3 3'
+7+
+ chain
5' 1 3' 5' m-RNA
FIG. 3. A model which describes the role of DNA methylation in class I1 antigen expression on T cell tumours. MC = methylcytosine; C = cytosine.

192 S. K . Singh, K . A . Donovan and C. S . David mechanism. As mentioned earlier, PLT-24.2 was derived from an alloreactive cell population (Singh et al., 1984). This cell line grows autonomously in culture without any exogenous stimulation and has the characteristics of a transformed phenotype, as evident from the soft-agar colony assay. Previously, using indirect immunofluorescence and immunochemical techniques, we have demonstrated that this cell line is of T cell origin and expresses class I1 antigens on its surface. Tryptic peptide profiles of A, and A, chains of PLT-24.2 were found to be identical to those of syngeneic B lymphocytes (H-29 . However, when we analyzed genomic restriction enzyme fragments by Southern blotting, a different pattern was observed. When Bam HI and Eco RI digested fragments were hybridized to an A,, probe, we observed that PLT-24.2 gave rise to bands which were different from either B6 (H-2*) or B10.K (H-zk) . This was rather puzzling. This cell line was derived from a B6 anti-B 10.K mixed lymphocyte culture. Based on the serological and biochemical data, PLT-24.2 was assumed to be derived from B6 responder cells. As mentioned earlier, the tryptic peptide maps of H-21-A antigen chains were identical to B lymphocytes of B6 strain. Since the tryptic peptide map was performed using radio- labelled leucine, however, we might not have detected all the peptides.
In this paper we propose a mechanism for class I1 gene expression in the PLT-24.2 cell line. It has been demonstrated by several authors that long terminal repeats (LTR) of the retrovirus genome can act as the promoter for the expression of cellular genes (Wong er aZ., 1985). We hypothesise that the transformation status of PLT-24.2 is due to viral infection during long-term culture in uitro. An LTR sequence inserted near to the class I1 genes might act as an enhancer element for the transcription of class I1 mRNA. Alternatively, endogenous LTR sequences might have been transposed next to the class I1 genes of PLT-24.2. This assumption is based on the finding that novel Eco RI and Bam HI bands are observed for the A, gene in PLT-24.2. We have proposed a model of how the insertion of viral LTR can activate the class I1 genes in PLT 24.2 (Fig. 4). The insertion of new sequences codd affect DNA topology and produce local conformations that allow access of RNA polymerase to the DNA template, or unwinding the double helix at the site of transcription. We have constructed a genomic library of PLT-24.2 in a bacterophage A vector to isolate the A, gene for further characterization by nucleotide sequencing. This will enable us to know whether any LTR sequence has been inserted near the A, gene.
MODEL II--0EREPRESSION OF Ap GENE BY VIRAL PROMOTER
Viral gene A&b
+ 4 Eco R 1 Eco R 1 - -
P PLT-Apb
EcoR1 EcoR1 4 + 3 a o o c
FIG. 4. Possible role of viral genes in the activation of class I1 genes in transformed T cells. Insertion of a viral promoter (P) can activate class I1 genes in PLT-24.2 cell line.

MHC class 11 expression on T cell lines 193 A wealth of data has been accumulated on the role of mitogen and IFN-y in the induction
of class I1 antigens on B cells. How these exogenous factors act on the expression of class I1 antigens has not been explored, however. Recent evidence suggests that the enhancer sequence might require a tissue-specific trans-acting factor. It has been demonstrated in the immunoglobulin gene system, that induction by external stimuli during the pre-B cell maturation stage provides this trans-acting effect, which becomes constitutive in later stages (Nelson et al., 1984). The expression of class I1 antigens in activated T cells probably depends upon external stimuli, e.g. mitogen, IFN. These external stimuli might confer DNaseI hypersensitive sites in.or around the class I1 genes, which in turn makes the DNA accessible to RNA polymerase. This has been shown in other systems, e.g. T cell receptor genes (Bier et al., 1985). Whether this mechanism is responsible for differential expression of class I1 antigens in different tissue is under investigation in our laboratory.
A C K N O W L E D G M E N T S
This work was supported in part by USNIH A1 Grant no. 14764 and CA grant no. 24473.
R E F E R E N C E S
BIER, E., HASHIMOTO, Y., GREENE, M.I. & MAXAM, A.M. (1985) Active T-cell receptor genes have intron deoxyribonuclease hypersensitive sites. Science, 229,528.
DAVtD, C.S., MEO, T., MCCORMICK, J. & SHREFFLER, D.C. (1976) Expression of individual Ia specificities on T and B cells. 1. Studies with mitogen induced blast cells. Journal ofExperimenta1 Medicine, 143,218.
DE WOLF, W.C., SCHLOSSMAN, S.F. & YUNIS, E.J. (1979) DRW antisera react with activated T cells. Journal of Immunology, 122,1780.
GRUETZMANN, R. & HAMMERLING, GJ. (1978) Characterization and functional analysis of T cell hybrids. Current Topics in Microbiology and Immunology, 81. 188.
HEDRICK, S.M., NIELSON, E.A., KAVALER, 1. COmN, D.I. & DAVIS, M.M. (1984) Sequence relationships between putative-T cell receptor polypeptides and immunoglobulins. Nature, 304 153.
INDIVERI, F., WESON, B.S., RWSSO, C., QUAWA, V., PELLJXRINO. M.A. & FERRONE. S. (1980) Ia like antigens on human T lymphocytes. Relationship to other surface markers, role in mixed lymphocyte reactions, and structural profile. Journal of Immunology, 125, 2673.
KING, D.P. & JONES, P.P. (1983) Induction of Ia and H-2 antigens on a macrophage cell line by immune interferon. Journal of Immunology, 13 1, 3 15.
KLEIN, J. (1978) Genetics of cell mediated lymphocytotoxicity in the mouse. Springer Seminars in Immunoparhology, I, 3 1.
KLEIN, J . & NAGY, Z. (1982) Mhc Restriction and Ir genes. Aduances in Cancer Research, 37,233. LORBER, M.I., LOKEN, M.R., STALL, A.M. & Frrcq F.W. (1982) I-A antigens on cloned alloreactive murine
T lymphocytes are acquired passively. Journal oflmmunology, 128,73 1. MAKI, R., KEARNEY, J., PAIGE, C. & TONEGAWA, S. (1980) Immunoglobulin gene rearrangement in immature
B cells. Science, 209, 1366. MATHIS, D.J., BENOIST, C., WILLIAMS, V.E. 11, KANTER, M. & MCDEVI-IT, H.O. (1983) Several mechanisms
can account for defective E, gene expression in different mouse haplotypes. Proceedings of the National Academy of Sciences, U.SA., 80,273.
MEO, T., DAVID, C. S., RUNBEEK, A. M., NABHOW M., MIAGGIANO, V.C. & SHREFFLER, D.C. (1975) Inhibition of mouse MLR by anti-Ia sera. Transplanfation Proceedings, 7, 127.
NAKAMURA, M., MANSER, T., PEARSON, G., DALEY, D.N. & GEFTER. M.L. (1984) Effect of IFN-y on the immune response in uiuo and on gene expression in uifro. Nature, 307,381.
NELSON, K.J., MATHER, E.L. & PERRY, R.P. (1984) Lipopolysaccharide-induced transcription of the kappa immunoglobulin locus occurs on both alleles and is independent of methylation status. Nucleic Acids Research, 12, 19 1 1.
PEWFUN, B.M., GONWA, T.A. & STOBO, J.D. (1984) Expression of HLA-DR by a human monocyte cell line is under transcriptional control. Journal of Molecular and Cellular Immunology, 1, 19 1.

194 S. K . Singh, K . A . Donovan and C . S . David REG M.S., MANN, D.L., EIDEN, M., TRAINOR, C.D. & CUR=, M.F. (1984) DNA methylation and
expression of HLA-DR a. Molecular and Cellular Biology, 4, 890. SANTI, D.V., GARREIT, C.E. C BARR, P.J. (1983) On the mechanisms of inhibition of DNA-cytosine methyl
transferase by cytosine analogs. Cell, 33,9. SINGH, S.K., ABRAMSON, E.J., KRCO, C.J. & DAVID, C.S. (1984) Endogenous synthesis of Ia antigens by a
cloned T cell line. Journal of Molecular and Cellular Immunology, 1, 147. SXNGH, S.K., DONOVAN, K. A., WOLOSCHAK, G.E. & DAVID, C.S. (1985) DNA methylation and expression of
Ia antigens in murine T cell lines. Advances in Gene Technology; Molecular Biology of the Immune System, ICSU Short Reports, 2,307.
SNYDER, D.S., BELLER, D.I. & UNANUE, E.R. (1982) Prostaglandins modulate macrophage Ia expression. Nature, 299, 163.
STEINMETZ, M., MINARD, K., HORVARTH, S . el al. (1982) A molecular map of the immune response region from the major histocompatibility complex of the mouse. Nature, 300,35.
WONG. T.C., GOODENOW, R.S., SHER, B.T. & DAYIDSON, N. (1985) The promoter of the long terminal repeat of feline leukemia virus is effective for expression of a mouse H-2 histocompatibility gene in mouse and human cells. Gene, 34, 27.