Embed Size (px)
Citation preview
Transcriptional control of gene expression
Book recommendation:Christiane Nüsslein-Vollhard:Das Werden des Lebens –Wie Gene die Entwicklung steuern
Book recommendation:Christiane Nüsslein-Vollhard:Das Werden des Lebens –Wie Gene die Entwicklung steuern
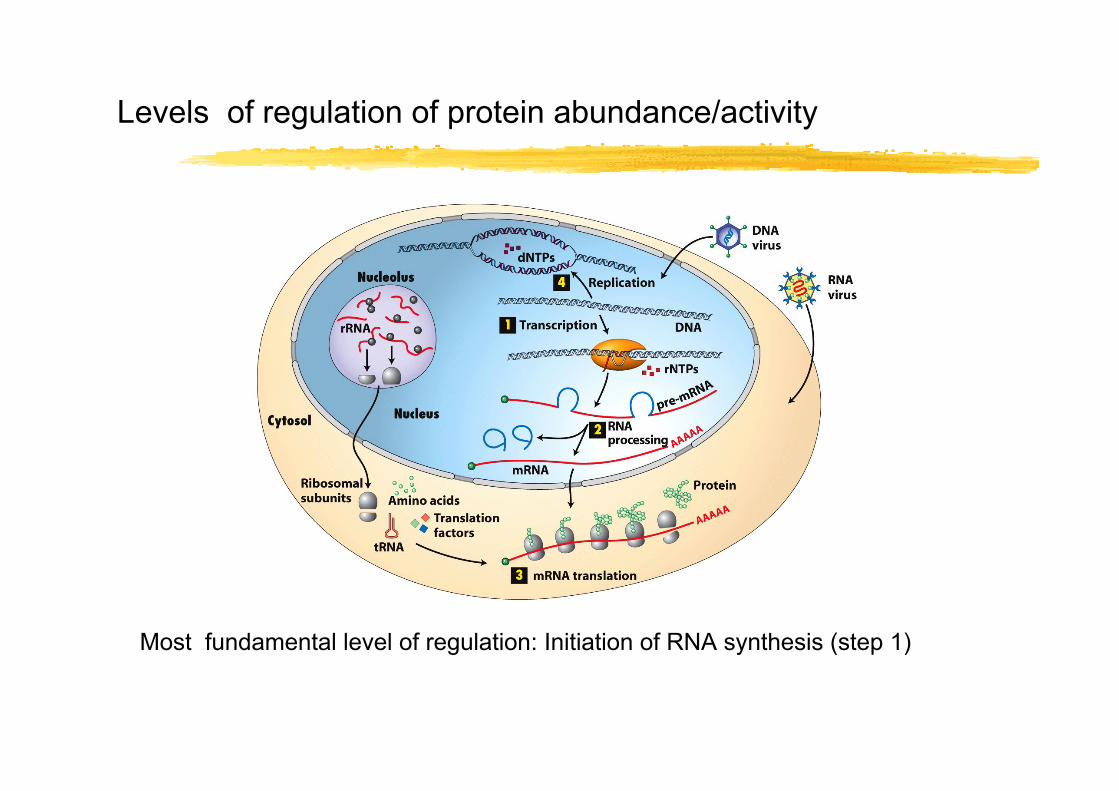
Levels of regulation of protein abundance/activity
Most fundamental level of regulation: Initiation of RNA synthesis (step 1)

Molecular definition of a geneA gene is the entire nucleic acid sequence that is necessary for the controlled production of its final product (RNA or Protein)
In eukaryotes, genes lie amidst a large expanse of noncoding DNA with unknown function and genes may also span regions of DNA unrelated to the gene
If a gene is incapable of producing a final gene-product = pseudogene
Exons =coding region (ORF)
(Introns)
Basal Promoter
Regulatory region (enhancer/prepressor)
PolyA site
Splice site
TSS
In the previous lesson the gene structure was defined, including the macromolecular structure; chromatin
This lesson focuses on regulatory regions + promoters
Repetition from last lesson:

ProcaryotesBacterial regulation of gene expression
Regulation is highly determined by changes in environment (nutritional and physical)Several genes from one metabolic pathway are clustered into operonsOperons are coordinately regulated (one “control-box” for all)RNA polymerase requires σ-factors for binding DNA-sequences at TSSRecruitment of RNA polymerases to TSS is regulated by activating and repressing trans-acting factorsMost cis-acting elements are close to the TSS
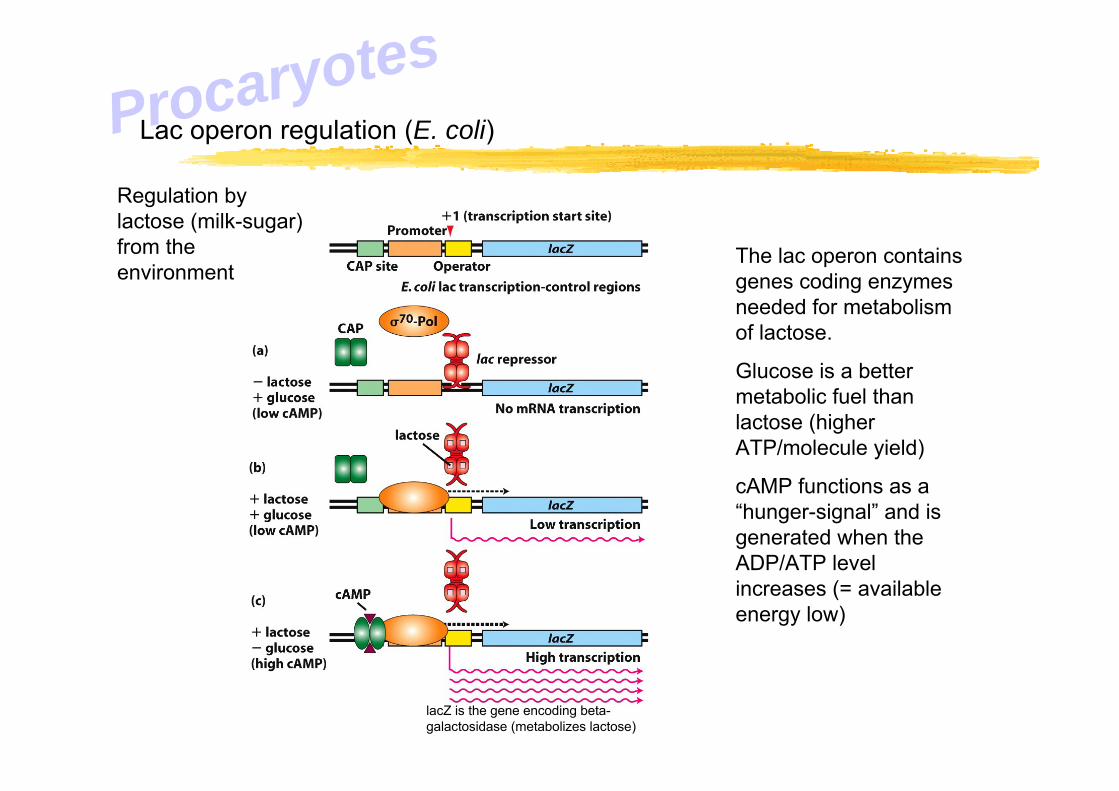
Lac operon regulation (E. coli)
The lac operon contains genes coding enzymes needed for metabolism of lactose.
Glucose is a better metabolic fuel than lactose (higher ATP/molecule yield)
cAMP functions as a “hunger-signal” and is generated when the ADP/ATP level increases (= available energy low)
Regulation by lactose (milk-sugar) from the environment
lacZ is the gene encoding beta-galactosidase (metabolizes lactose)
Procaryotes

ProcaryotesOther regulatory regions can sit further away from the TSS
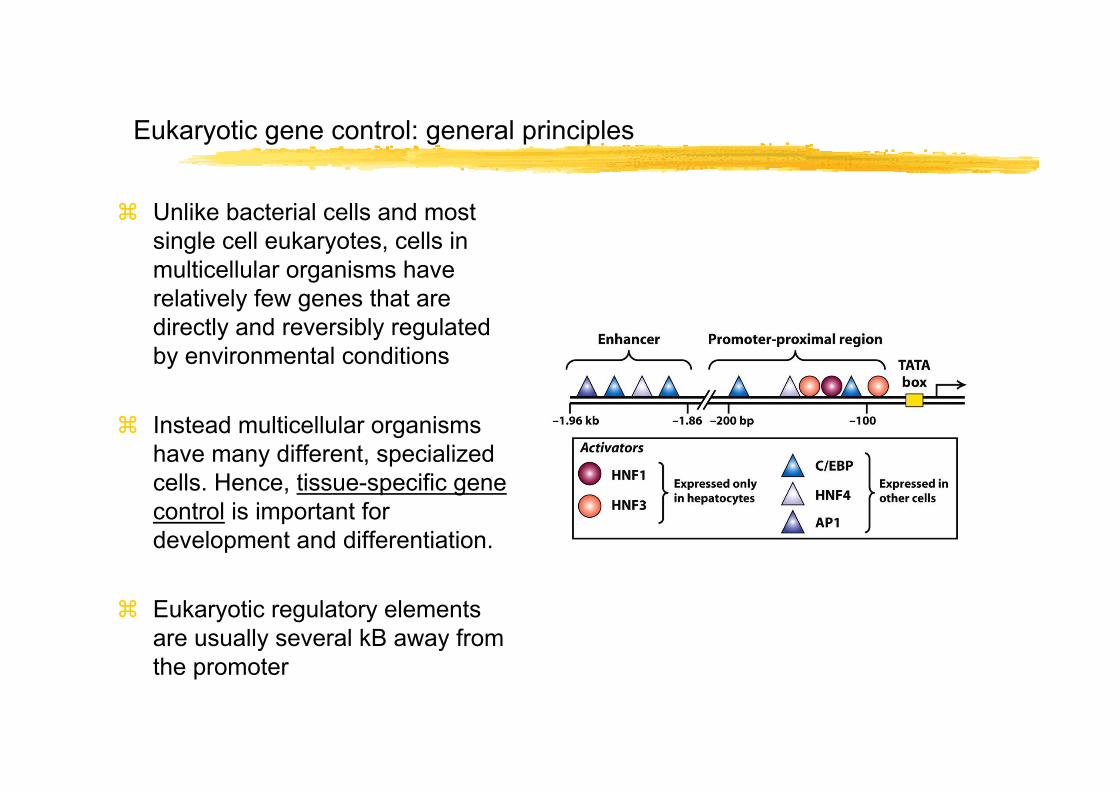
Eukaryotic gene control: general principles
Unlike bacterial cells and most single cell eukaryotes, cells in multicellular organisms have relatively few genes that are directly and reversibly regulated by environmental conditions
Instead multicellular organisms have many different, specialized cells. Hence, tissue-specific gene control is important for development and differentiation.
Eukaryotic regulatory elements are usually several kB away from the promoter
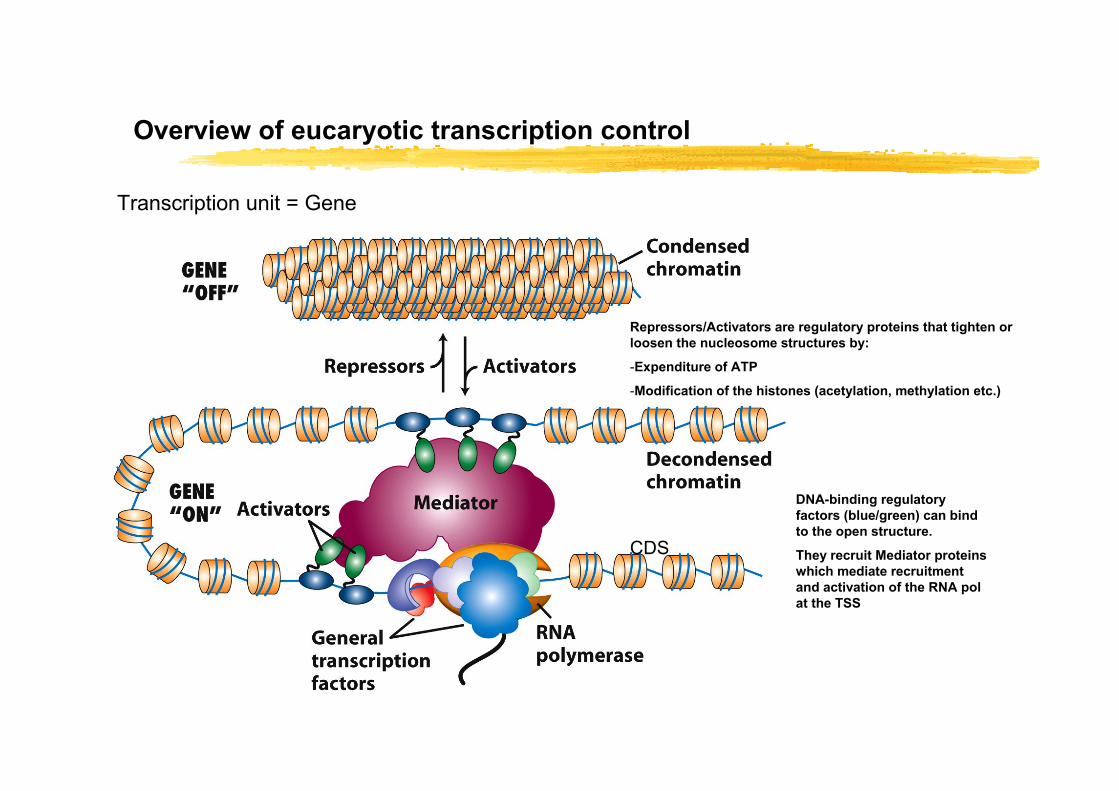
Overview of eucaryotic transcription control
Transcription unit = Gene
Repressors/Activators are regulatory proteins that tighten or loosen the nucleosome structures by:
-Expenditure of ATP
-Modification of the histones (acetylation, methylation etc.)
DNA-binding regulatory factors (blue/green) can bind to the open structure.
They recruit Mediator proteins which mediate recruitment and activation of the RNA pol at the TSS
CDS

Transcription initiation
RNA polymerases initiates transcription of most genes at a unique DNA position lying upstream of the coding sequenceThe base pair where transcription initiates is termed the transcription-initiation start site (TSS)By convention, the TSS in the DNA sequence is designated +1, and base pairs extending in the direction of transcription (downstream) are assigned positive numbers and those extending in the opposite direction (upstream) are assigned negative numbersVarious proteins (RNA polymerase, activators, repressors) interact with DNA at the promoter or several kilobases distant to the promoter to regulate transcription initiationThe DNA sequences (response elements) that bind regulatory proteins are Cis-acting sequences. The regulatory proteins are generally coded by a different gene, hence they are Trans-acting factors

Regulatory sequences in protein-coding genes
Multiple protein-binding control regions are located close to (proximal) or far from (distal) the transcription start site (TSS)
Promoters direct binding of RNA pol II to DNA, determine the site of transcription initiation (the beginning of the pre-mRNA), and influence transcription ratePromoter proximal elements occur within ~ 200 base pairs upstream of TSS. Several such may help regulate a genePromoter proximal elements and enhancers (often distal) often are cell-type specific, functioning only in speific differentiated cell-typesEach gene can be regulated by many different control elements
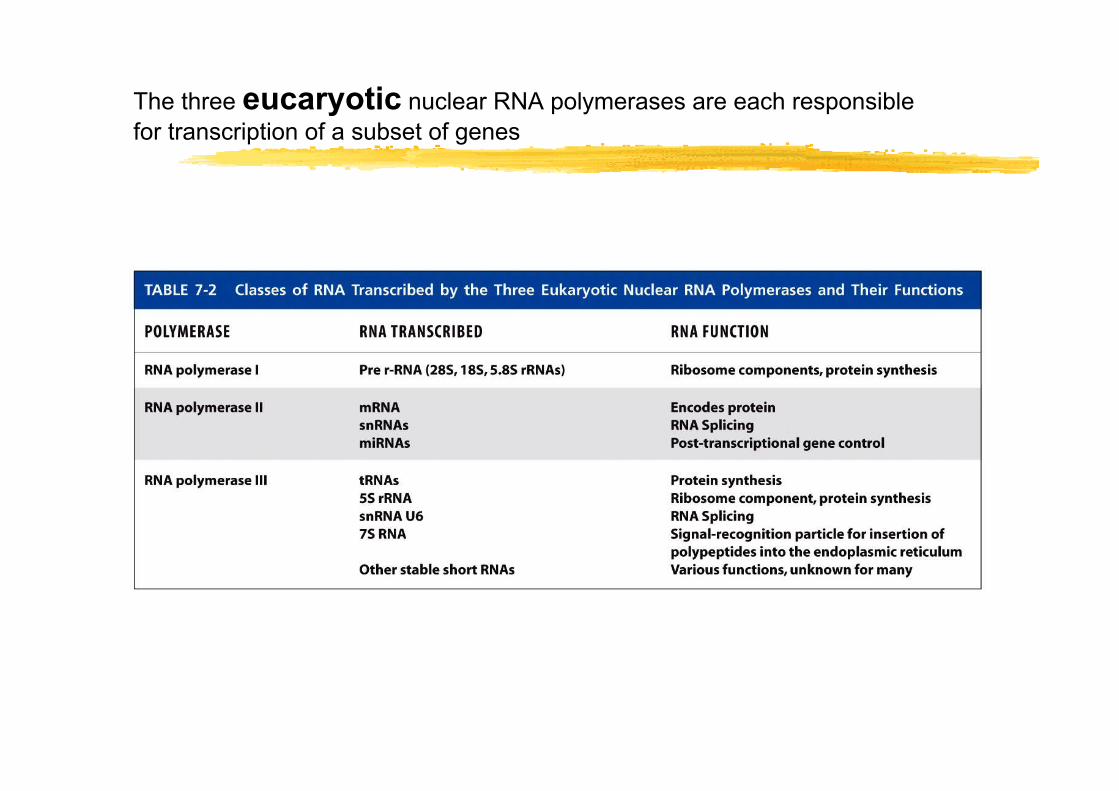
The three eucaryotic nuclear RNA polymerases are each responsible for transcription of a subset of genes

Regulatory sequences in protein coding genes
RNA Pol II binding site(The site where the RNA polymerase pre-initiation complex (PIC) assembles)
•TATA boxes are found in ~ 30% of all protein coding genes
•Other Initiator elements have been found in some of the other genes (current research still identifies new (often developmental specific!) elements
•CpG-islands (G-C rich DNA sequences) are often found in common housekeeping genes
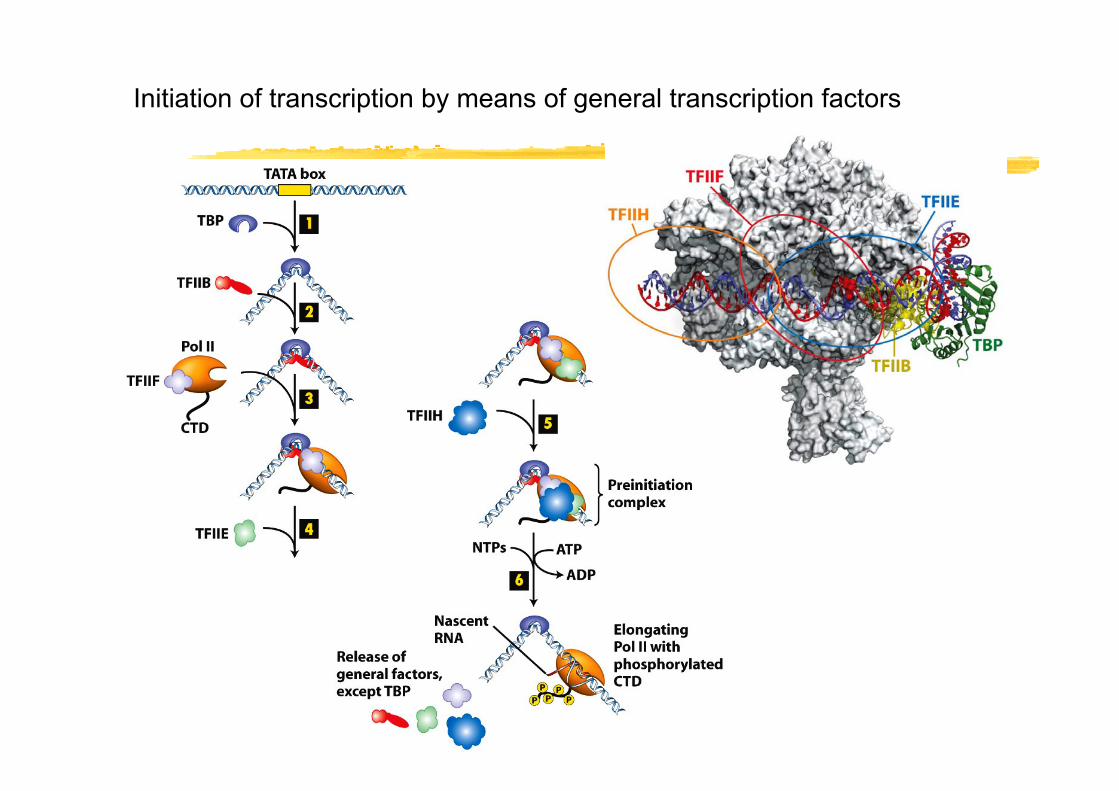
Initiation of transcription by means of general transcription factors

The C-terminal domain (CTD) of the largest subunit of RNA Pol II is phosphorylated during in vivo transcription
Antibody staining of phosphorylated (red) and unphosphorylated (green) Pol II CTD during transcription of a fruitfly chromosome
The CTD contains a repeated aminoacid sequence which can become highly phosphororylated
Repeat sequence:
Tyr-Ser-Pro-Thr-Ser-Pro-Ser

Transcription factors are often controlled by external stimuli
Cells respond to the needs of the body.
Hormones or nutrients (glucose, fatty acids, etc.) are signals that cells can read.
Cells respond by expressing the genes coding for proteins that can fullfill the current need of the body.
Body need signal cell response (gene activation)
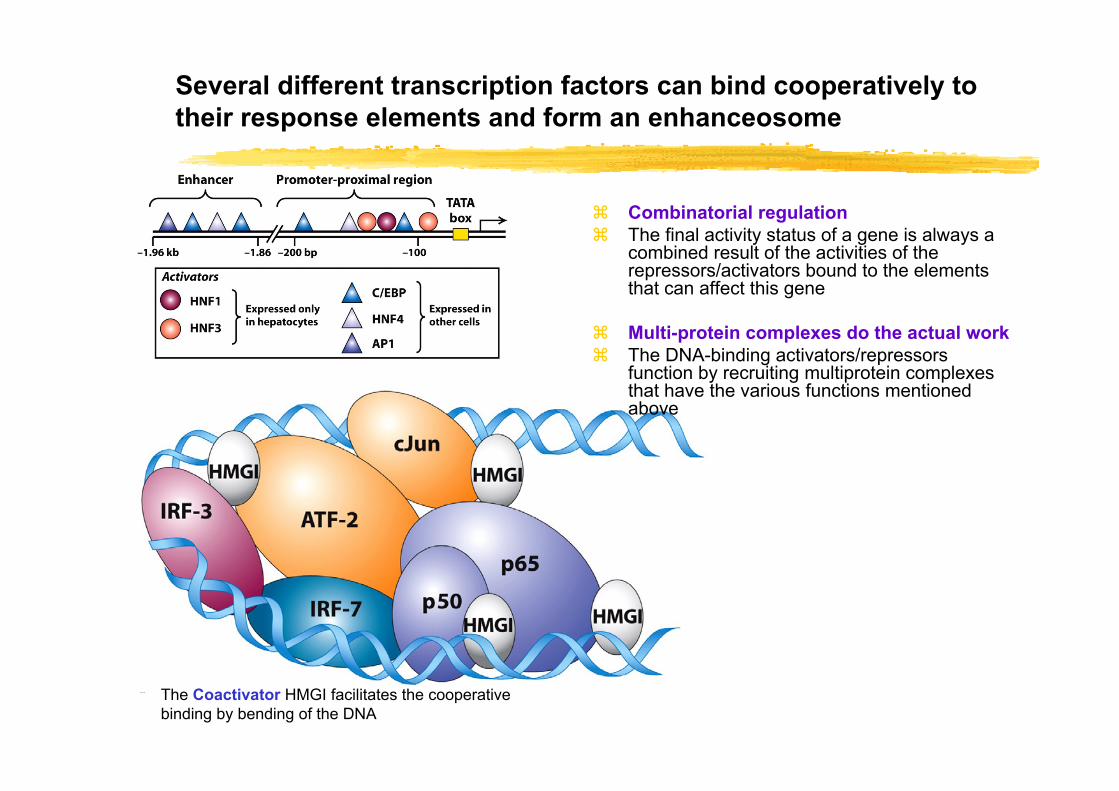
Several different transcription factors can bind cooperatively to their response elements and form an enhanceosome
The Coactivator HMGI facilitates the cooperative binding by bending of the DNA
Combinatorial regulationThe final activity status of a gene is always a combined result of the activities of the repressors/activators bound to the elements that can affect this gene
Multi-protein complexes do the actual workThe DNA-binding activators/repressors function by recruiting multiprotein complexes that have the various functions mentioned above

Transcription factors: Different functions of proteins binding regulator regions
Chromatin condensation/decondensation
•Activators can acetylate and demethylate chromatin to yield opened/loosened euchromatin
•Opened chromatin can be acetylated by other activators. Repressors can de-acetylate chromatin
•Decondensed chromatin is open for binding transcriptional activators and Pol II complex
Transcriptional enhancement
•Activators can mediate increased Pol II pre-initiation assembly
•Activators can then mediate increased transcription rate of the Pol II (through CTD phosphorylation)
•Repressors block the above processes
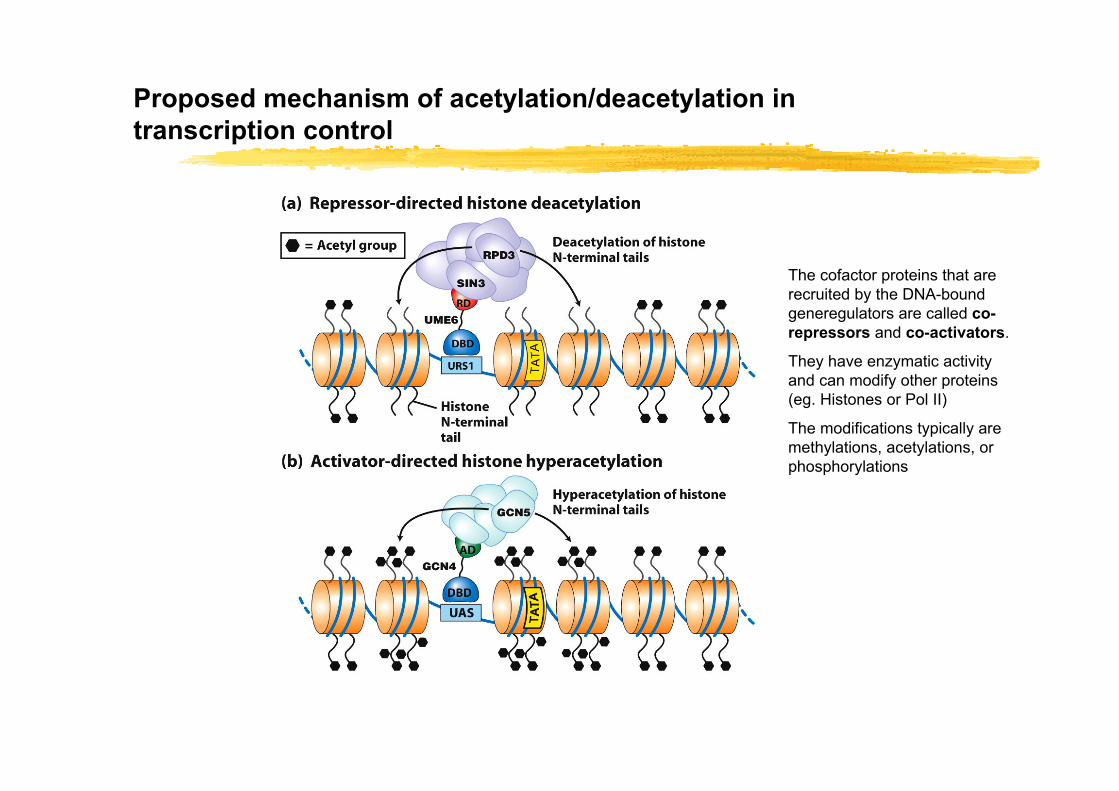
Proposed mechanism of acetylation/deacetylation in transcription control
The cofactor proteins that are recruited by the DNA-bound generegulators are called co-repressors and co-activators.
They have enzymatic activity and can modify other proteins (eg. Histones or Pol II)
The modifications typically are methylations, acetylations, or phosphorylations
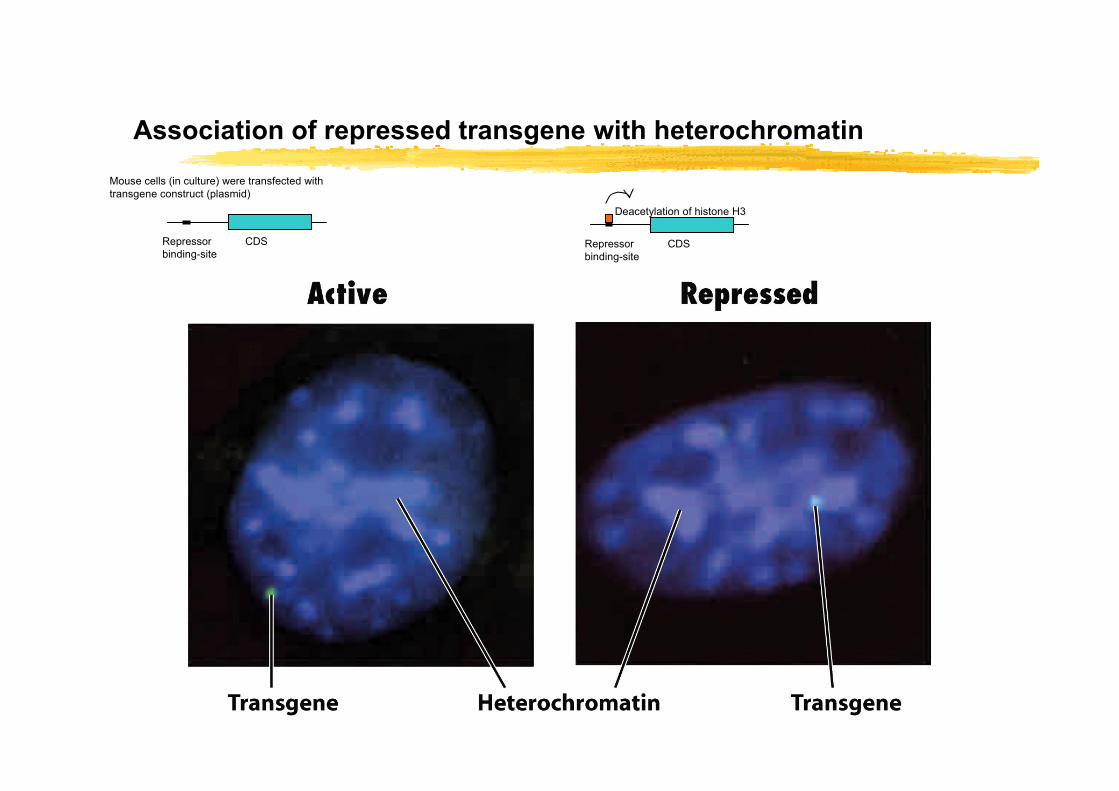
Association of repressed transgene with heterochromatin
CDSRepressor binding-site
Mouse cells (in culture) were transfected with transgene construct (plasmid)
CDSRepressor binding-site
Deacetylation of histone H3
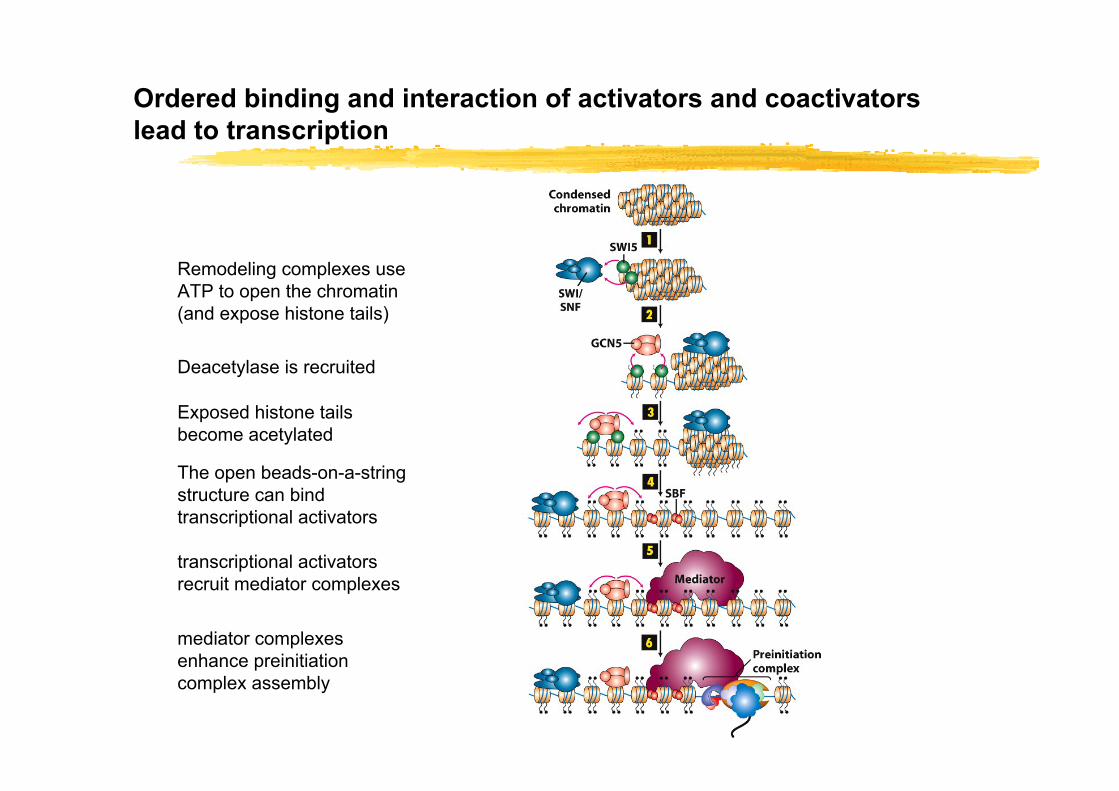
Ordered binding and interaction of activators and coactivators lead to transcription
Remodeling complexes use ATP to open the chromatin (and expose histone tails)
Deacetylase is recruited
Exposed histone tails become acetylated
The open beads-on-a-string structure can bind transcriptional activators
transcriptional activators recruit mediator complexes
mediator complexes enhance preinitiation complex assembly
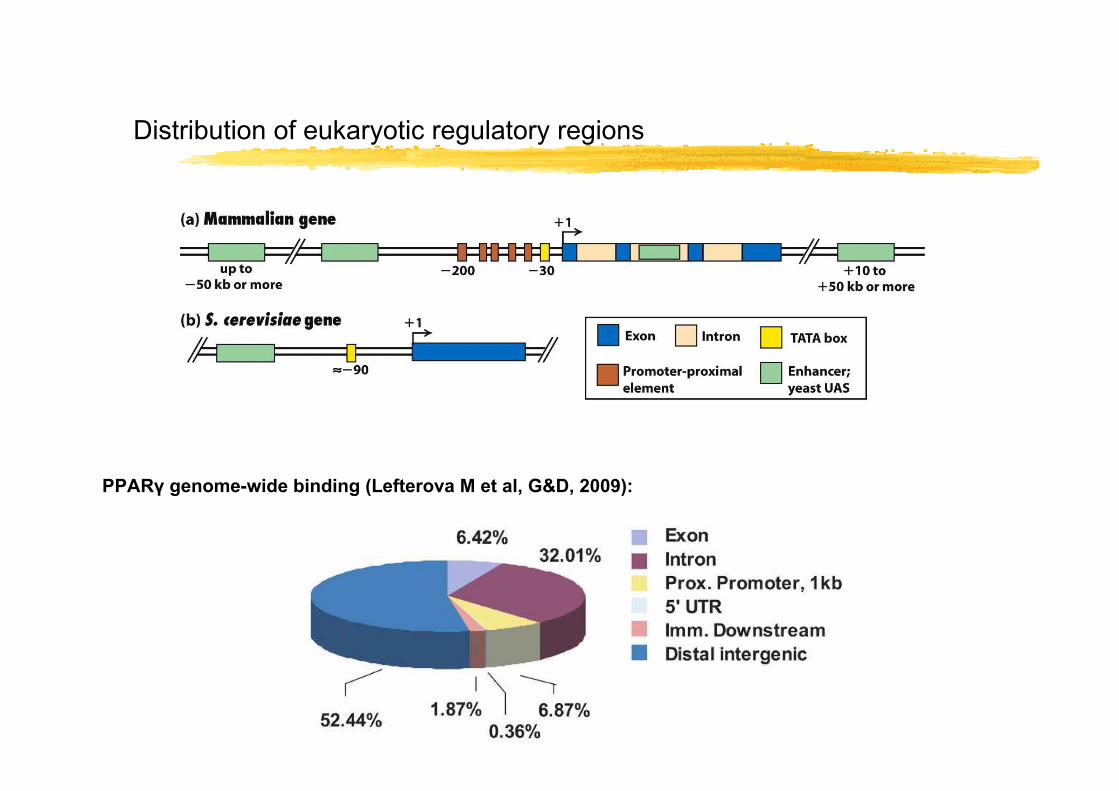
Distribution of eukaryotic regulatory regions
PPARγ genome-wide binding (Lefterova M et al, G&D, 2009):
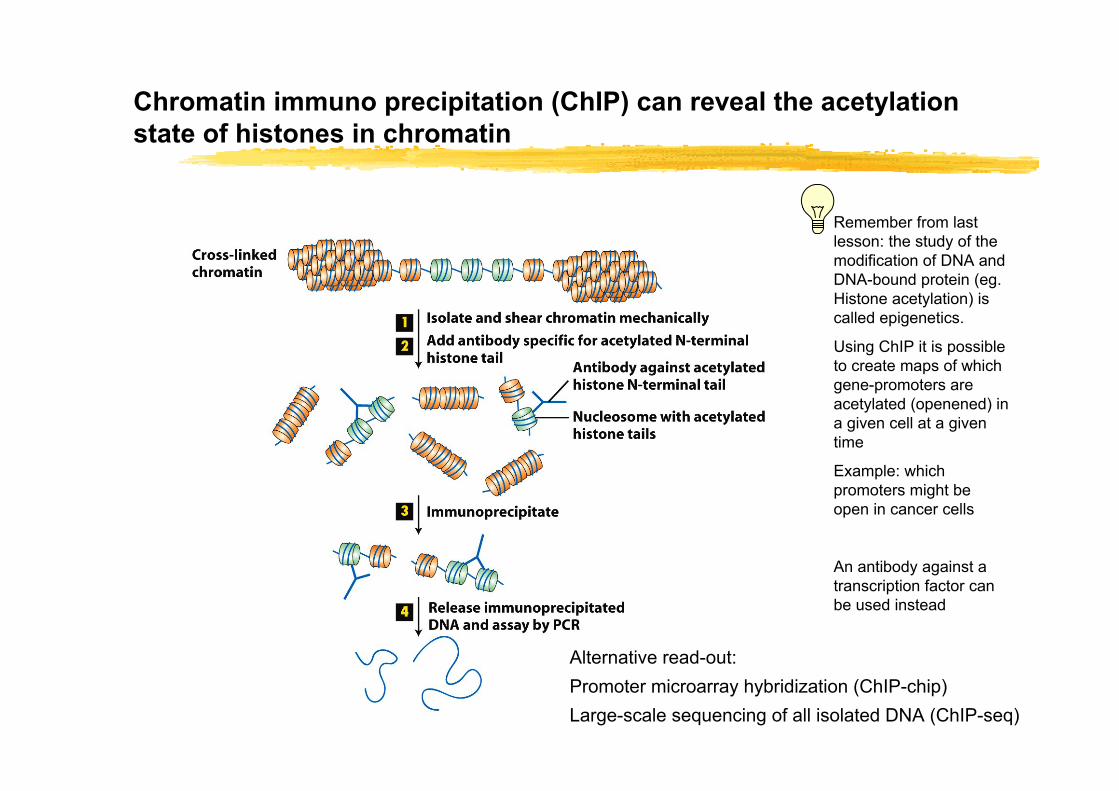
Chromatin immuno precipitation (ChIP) can reveal the acetylationstate of histones in chromatin
Alternative read-out:Promoter microarray hybridization (ChIP-chip)Large-scale sequencing of all isolated DNA (ChIP-seq)
Remember from last lesson: the study of the modification of DNA and DNA-bound protein (eg. Histone acetylation) is called epigenetics.
Using ChIP it is possible to create maps of which gene-promoters are acetylated (openened) in a given cell at a given time
Example: which promoters might be open in cancer cells
An antibody against a transcription factor can be used instead
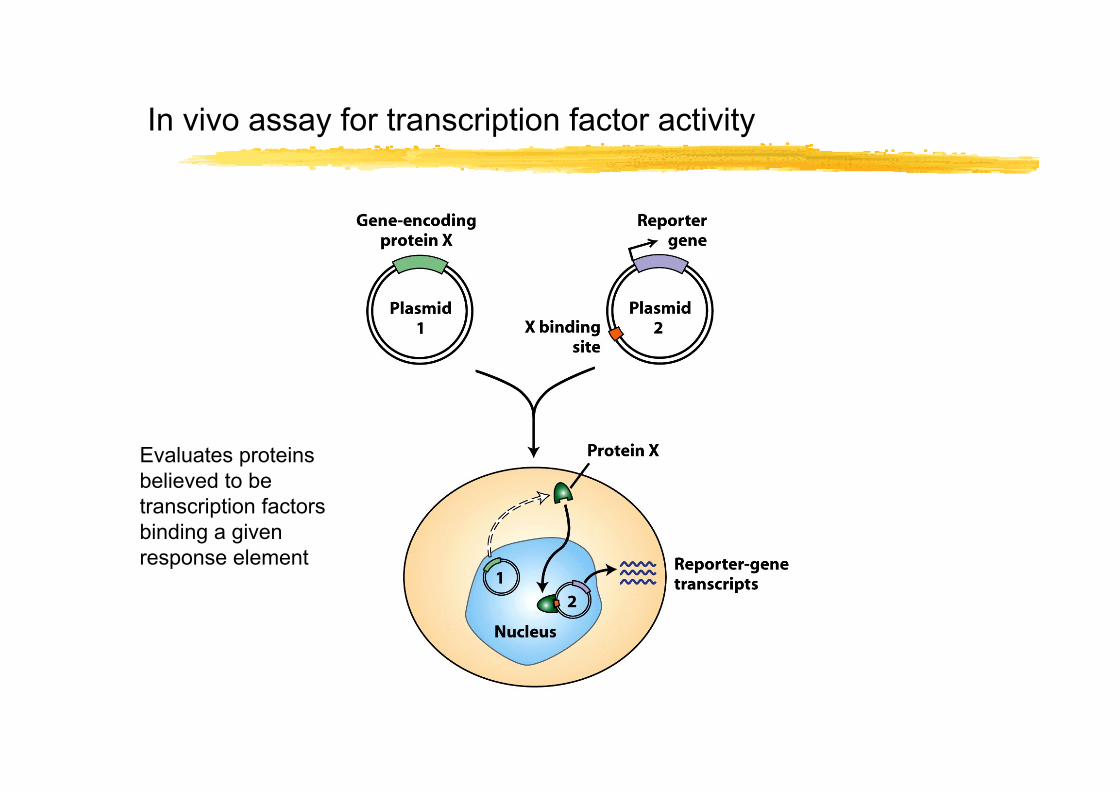
In vivo assay for transcription factor activity
Evaluates proteins believed to be transcription factors binding a given response element

Identification of domains in transcription factors
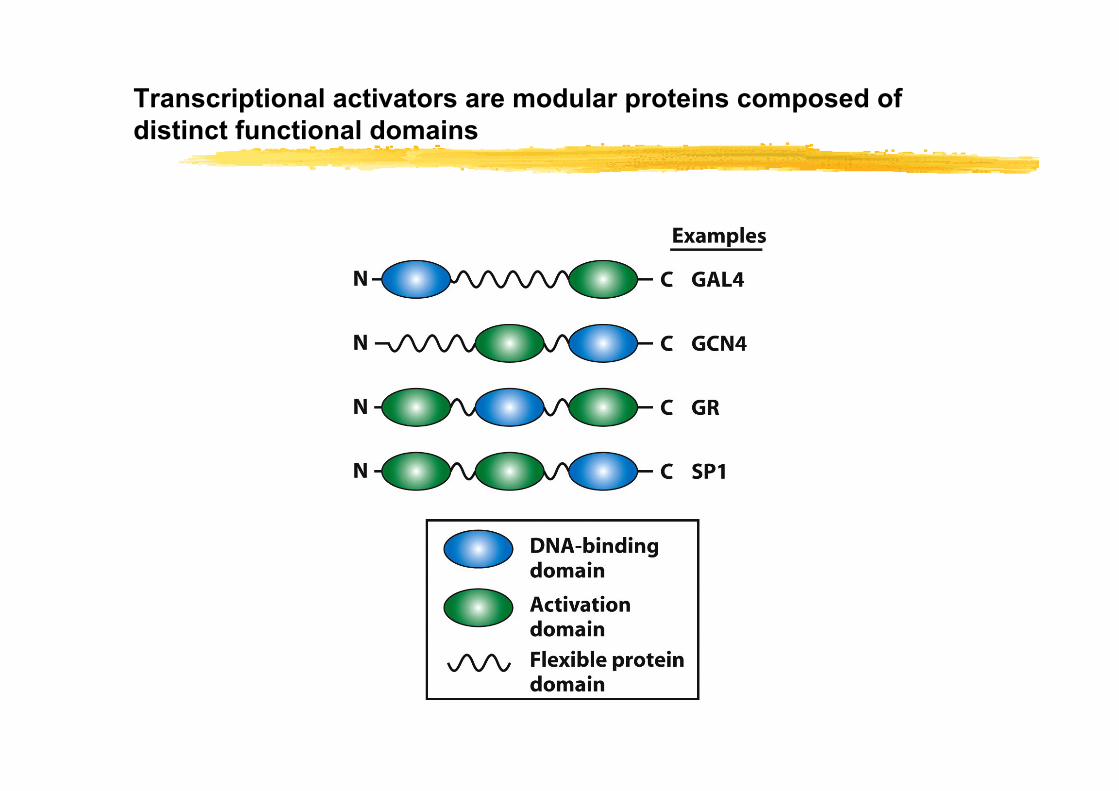
Transcriptional activators are modular proteins composed of distinct functional domains

DNAse I footprinting reveals control element sequencescan be used as first step in transcription factor identification
DNA sequence is labeled with 32P
And incubated +/- protein extract (column fractionated)
DNAse I can not digest DNA protected by a bound protein
Gel electrophoresis of the DNA digest reveals protected region

Electrophoretic mobility shift assay (EMSA) can detect transcription factors during purification
This is applicable to short DNA-sequences such as identified response elements (15-30 basepairs) Principle: radiolabeled 32P-DNA sequence is incubated with protein extract and applied to gel for electrophoretic separation. Retarded 32P-DNA is visualized by autoradiography
DNA-protein complex is bigger than free DNA-probe and therefore migrates slower through the gel
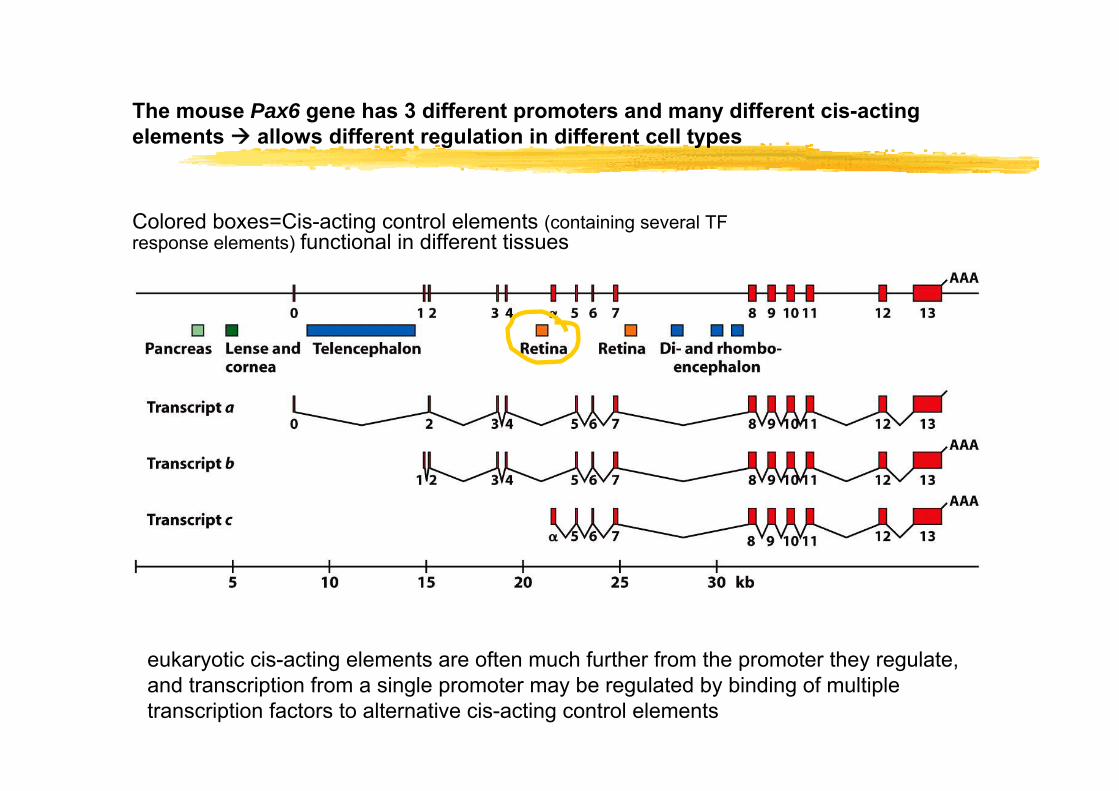
The mouse Pax6 gene has 3 different promoters and many different cis-acting elements allows different regulation in different cell types
eukaryotic cis-acting elements are often much further from the promoter they regulate, and transcription from a single promoter may be regulated by binding of multiple transcription factors to alternative cis-acting control elements
Colored boxes=Cis-acting control elements (containing several TF response elements) functional in different tissues
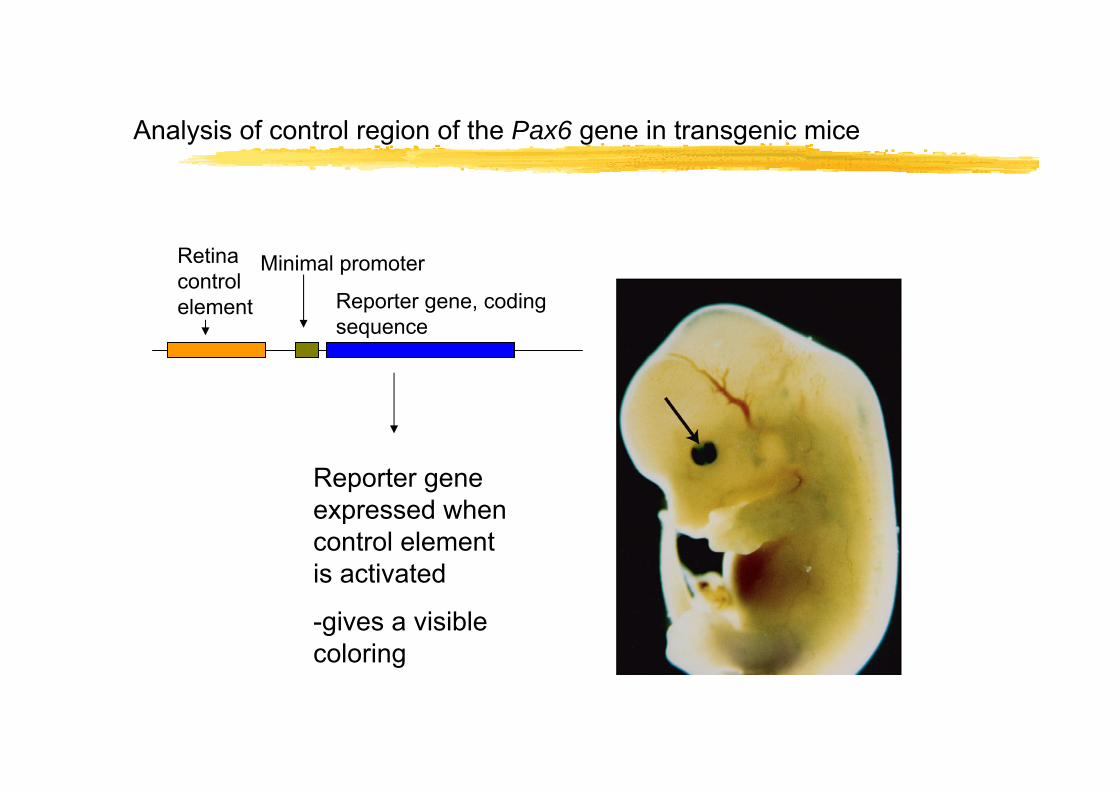
Analysis of control region of the Pax6 gene in transgenic mice
Retina control element
Minimal promoter
Reporter gene, coding sequence
Reporter gene expressed when control element is activated
-gives a visible coloring
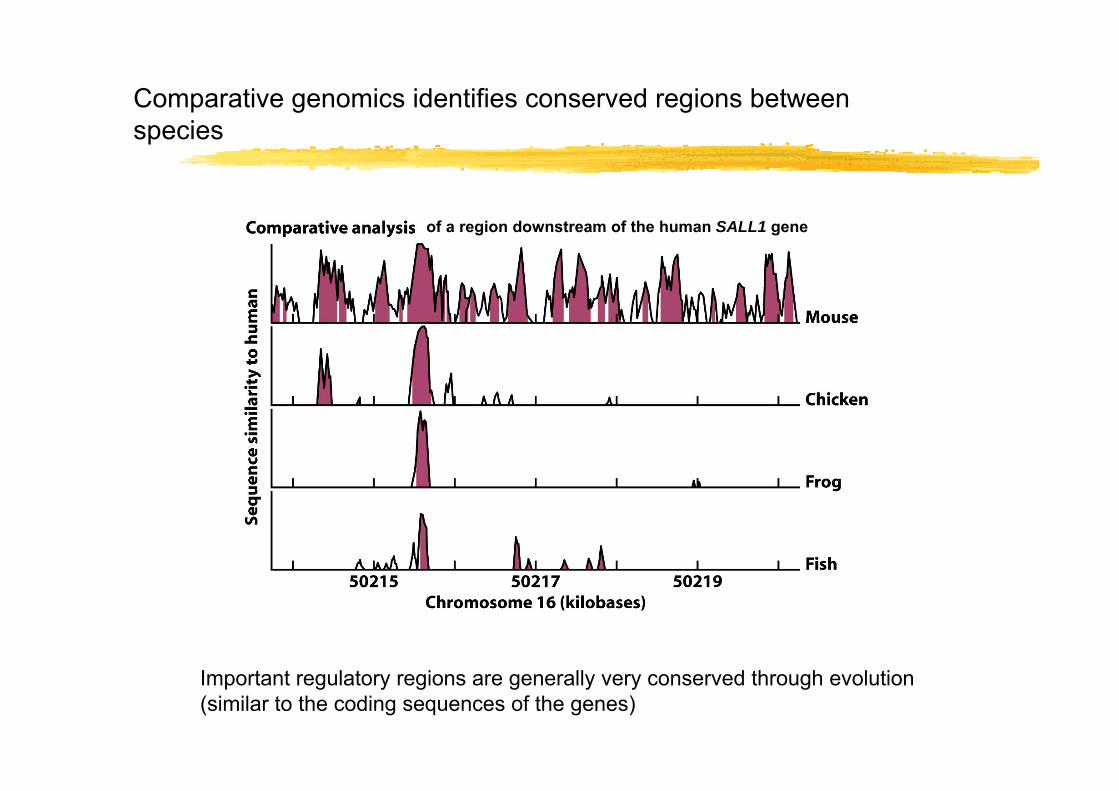
Comparative genomics identifies conserved regions between species
Important regulatory regions are generally very conserved through evolution (similar to the coding sequences of the genes)
of a region downstream of the human SALL1 gene
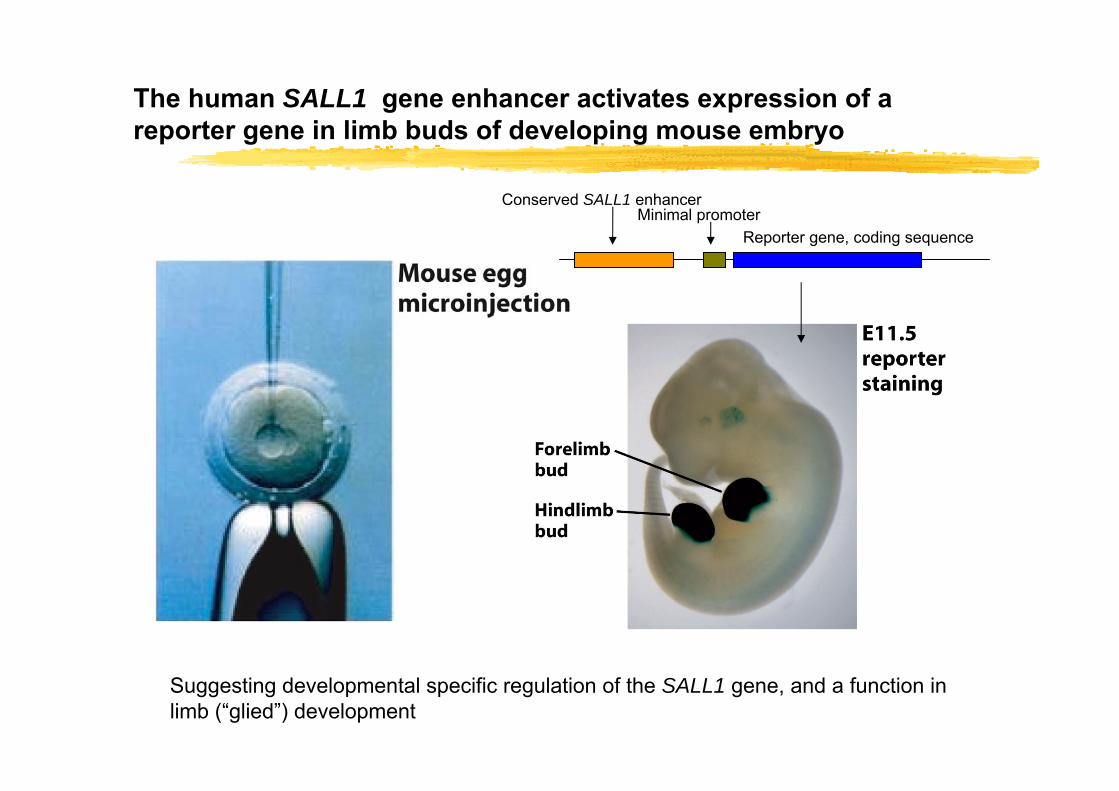
The human SALL1 gene enhancer activates expression of a reporter gene in limb buds of developing mouse embryo
Conserved SALL1 enhancerMinimal promoter
Reporter gene, coding sequence
Suggesting developmental specific regulation of the SALL1 gene, and a function in limb (“glied”) development
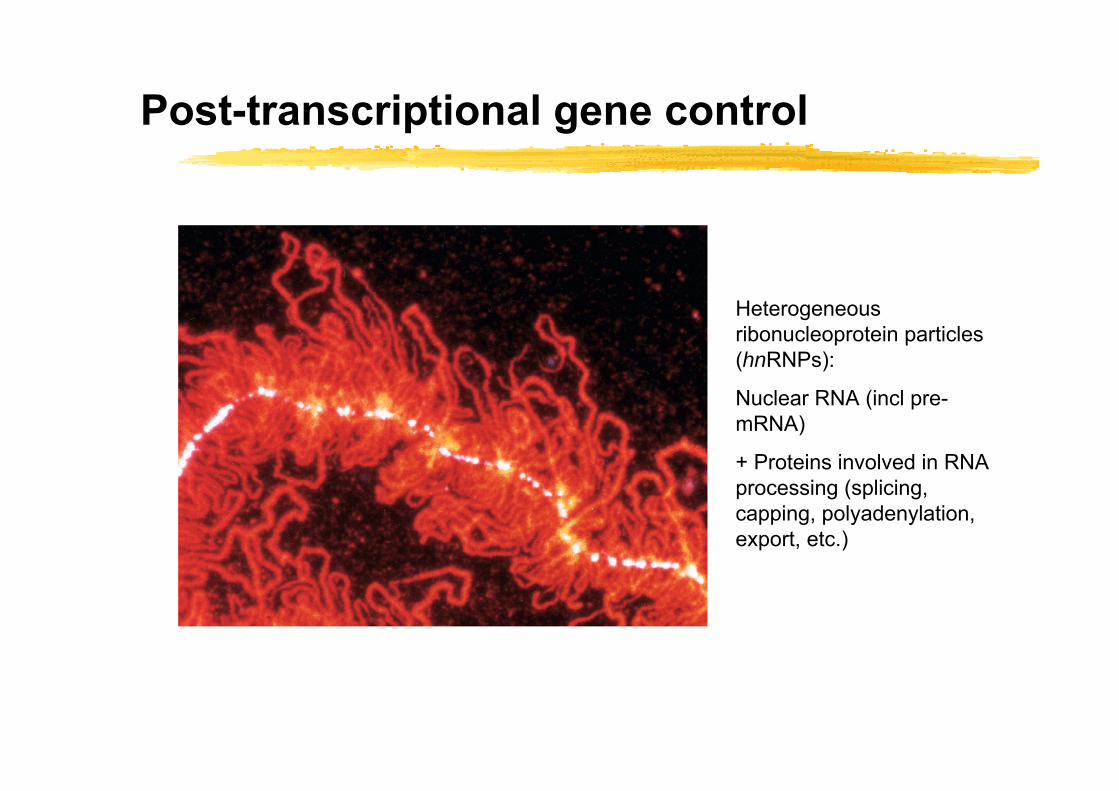
Post-transcriptional gene control
Heterogeneous ribonucleoprotein particles (hnRNPs):
Nuclear RNA (incl pre-mRNA)
+ Proteins involved in RNA processing (splicing, capping, polyadenylation, export, etc.)

Overview
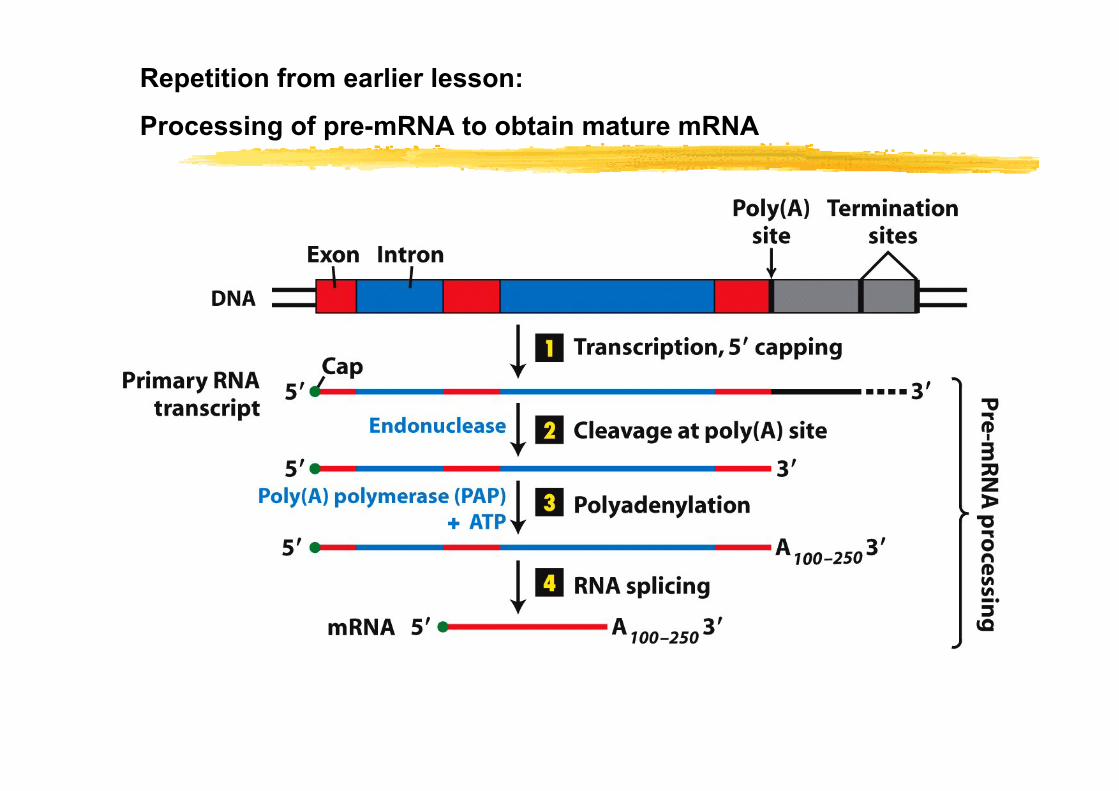
Repetition from earlier lesson:
Processing of pre-mRNA to obtain mature mRNA
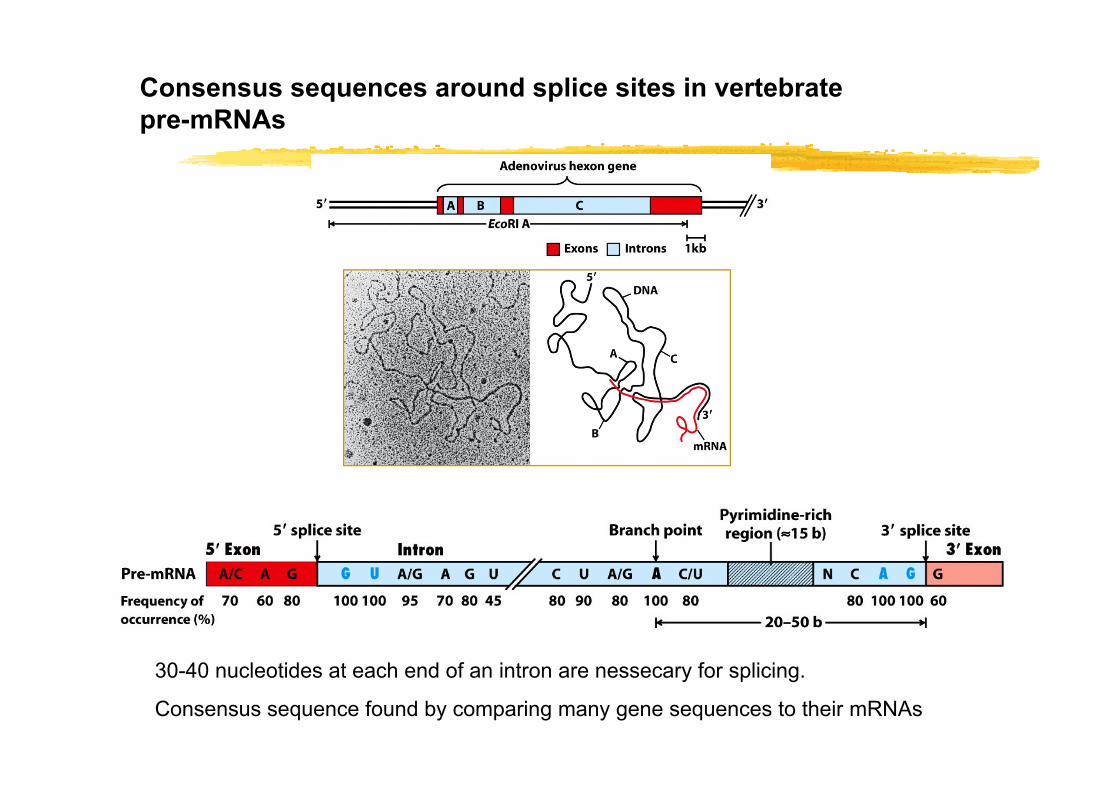
Consensus sequences around splice sites in vertebrate pre-mRNAs
30-40 nucleotides at each end of an intron are nessecary for splicing.
Consensus sequence found by comparing many gene sequences to their mRNAs
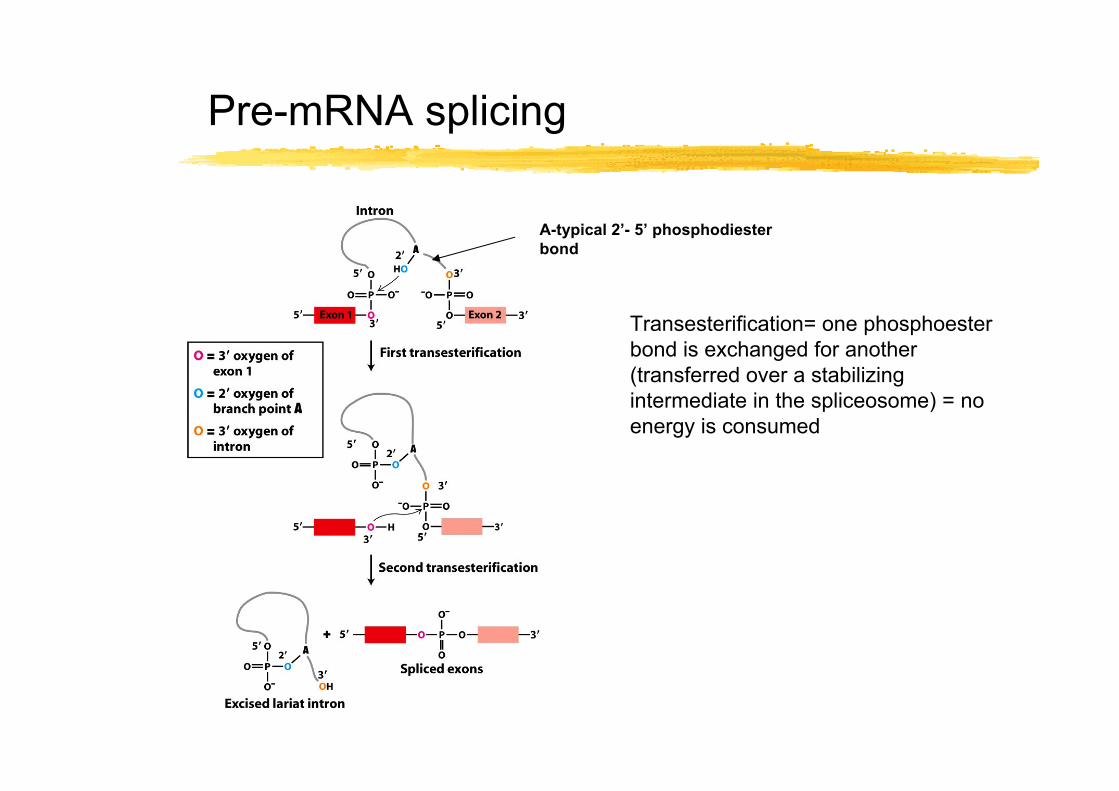
Pre-mRNA splicing
Transesterification= one phosphoester bond is exchanged for another (transferred over a stabilizing intermediate in the spliceosome) = no energy is consumed
A-typical 2’- 5’ phosphodiester bond
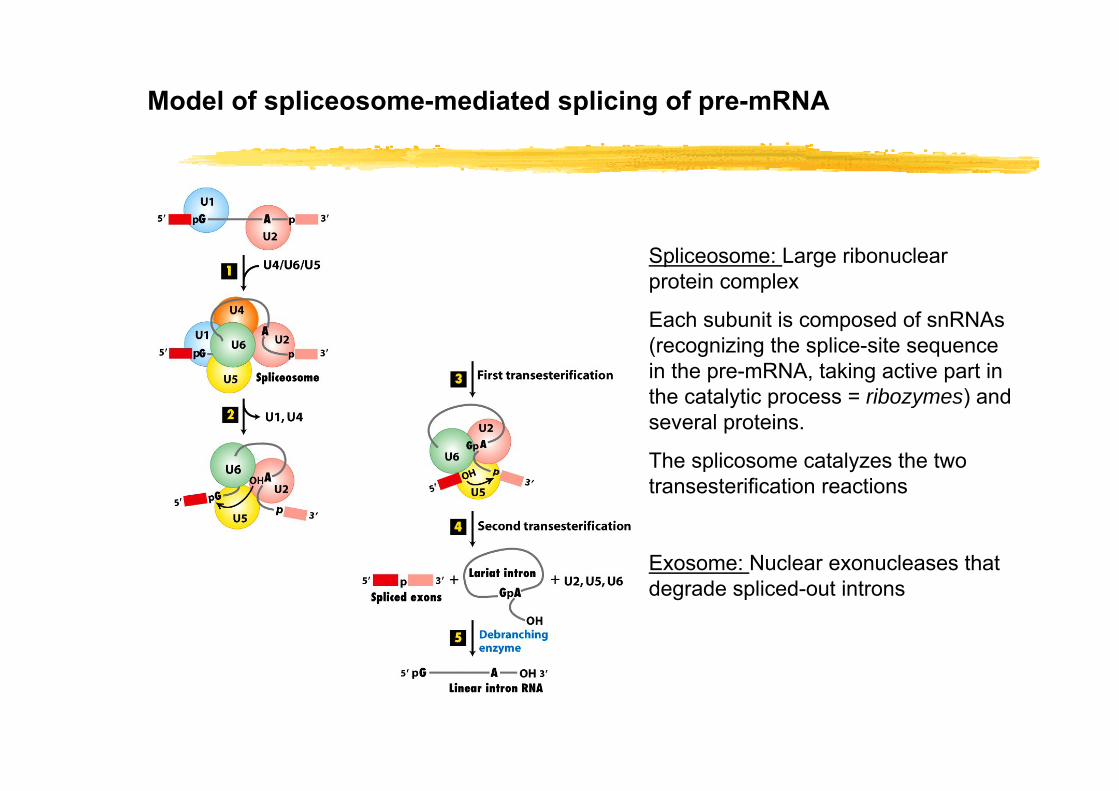
Model of spliceosome-mediated splicing of pre-mRNA
Spliceosome: Large ribonuclear protein complex
Each subunit is composed of snRNAs (recognizing the splice-site sequence in the pre-mRNA, taking active part in the catalytic process = ribozymes) and several proteins.
The splicosome catalyzes the two transesterification reactions
Exosome: Nuclear exonucleases that degrade spliced-out introns

RNA editing can lead to a functionally different protein
Rare or common event? An open question!

Micro RNAs (miRNAs) target mRNAs for degradation
miRNAs are expressed as long pre-miRNAs
They are processed into single or many short miRNAs, which is their active form, by specific proteins called Drosha and Dicer
miRNAs function in a ribo-protein complex called RISC
miRNAs bind (non-perfect match) to the 3’ end of mRNAs and initiate mRNA degradation
Because only non-perfect match is required one miRNA can have more than one target mRNA sequence

Small interfeiring RNAs (siRNAs) can inhibit translation and transcription
siRNAs are expressed as long multicopy RNAs or cleaved from untranslated sequences of normal mRNAs
siRNAs also function in the ribo-protein complex called RISC
siRNAs bind (perfect match) to mRNAs and block translation by inducing cleavage of the mRNA
siRNAs bound to pre-mRNA during transcription, can induce lysine methylation at histones of the parent gene
(NB: previous lesson: methylation induces condensed chromatin formation (heterochromatin)= reduced access for transcription activators and PolII = gene silencing)
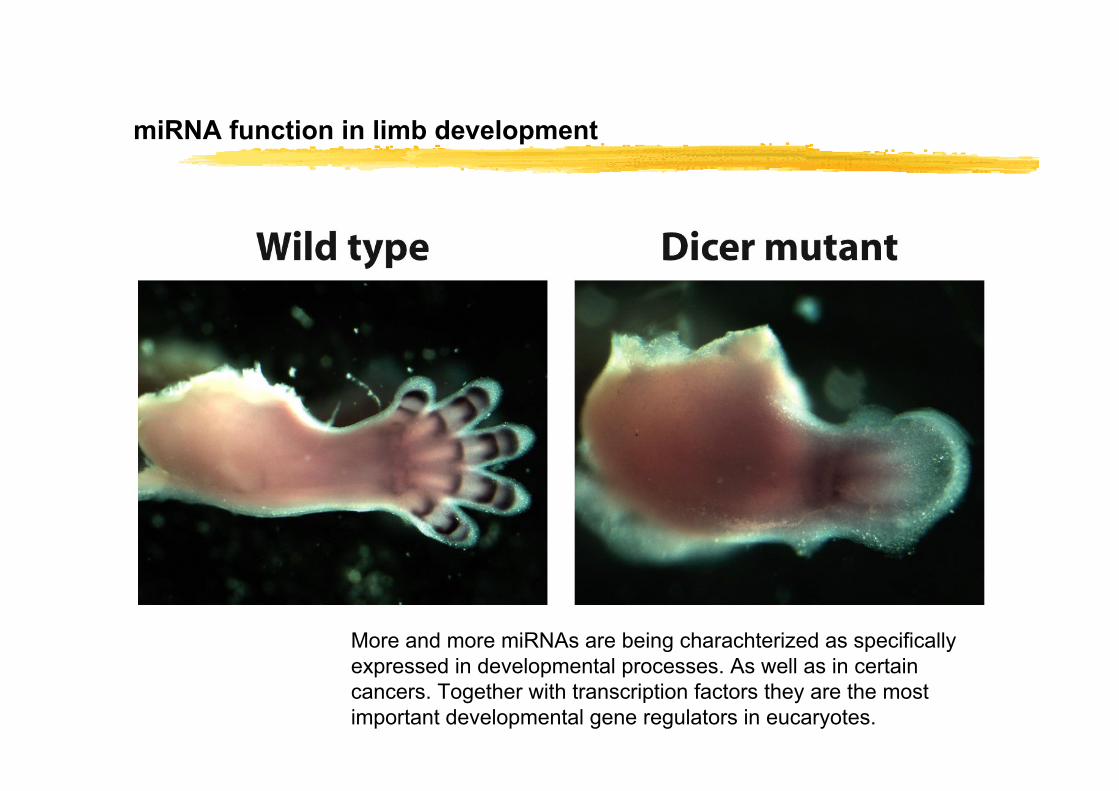
miRNA function in limb development
More and more miRNAs are being charachterized as specifically expressed in developmental processes. As well as in certain cancers. Together with transcription factors they are the most important developmental gene regulators in eucaryotes.
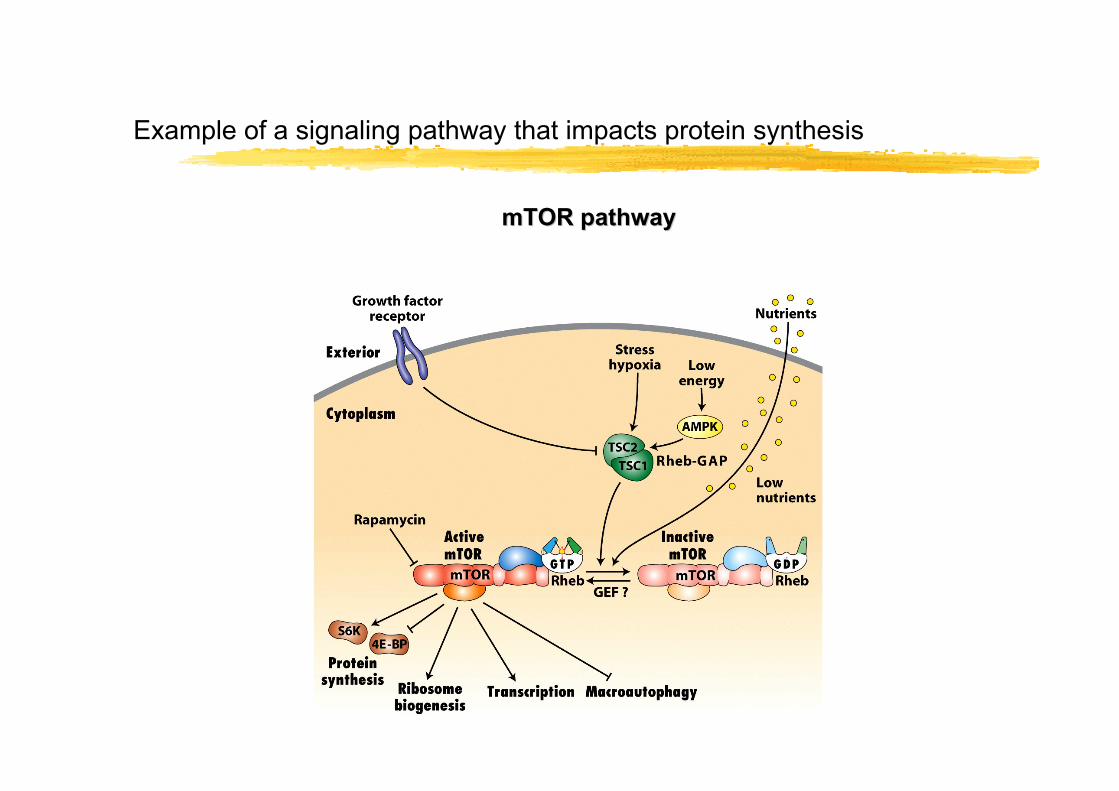
Example of a signaling pathway that impacts protein synthesis
mTOR pathwaymTOR pathway













