Embed Size (px)
Citation preview

Molecular Microbiology (1995) 17(6), 1085-1092
Transcriptional activation of the Escherichia coil bgl operon: negative regulation by DNA structural elements near the promoter
Jatinder Singh, t$ Mitali Mukerji $ and S. Mahadevan* Developmental Biology and Genetics Laboratory, Indian Institute of Science, Bangalore 560 012, India.
Summary
The bgl operon of Escherichia coil is transcriptionally inactive in wild-type cells. DNA insertion sequences (IS) constitute a major class of spontaneous muta- tions that activate the cryptic bgl promoter. In an attempt to study the molecular mechanism of activa- tion mediated by insertion sequences, transcription of the bgl promoter was carried out in vitro. Stimula- tion of transcription is observed when a plasmid con- taining an insertionally activated bgl promoter is used as a template in the absence of proteins other than RNA polymerase. Deletions that remove sequences upstream of the bgl promoter, and insertion of a 1.2kb DNA fragment encoding resistance to kana- mycin, activate the promoter. Point mutations within a region of dyad symmetry upstream of the pro- moter, which has the potential to extrude into a cruci- form structure under torsional stress, also lead to activation. Introduction of a sequence with dyad sym- metry, upstream of an activated bgl promoter carrying a deletion of upstream sequences, results in a fourfold reduction in transcription. These results suggest that the cryptic nature of the bgl promoter is because of the presence of DNA structural elements near the pro- moter that negatively affect transcription.
Introduction
Cryptic genes are enigmas in terms of function and evolu- tion, One of the best-studied cryptic genetic systems is the bgloperon of Escherichia coliwhich encodes the functions necessary for the uptake and metabolism of 13-glucoside sugars. The bgl operon in wild-type cells is expressed at a low level which is insufficient for growth on 13-glucoside
Received 31 October, 1994; revised 8 May, 1995; accepted 17 May, 1995. ?Present address: Department of Molecular Biology and Micro- biology, Tufts University Medical School, Boston, Massachusetts 02111, USA. ~These authors made an equal contribution to this work. *For correspondence. E-mail [email protected]; Tel. (80) 3092607; Fax (80) 3341683; Telex 084-8349.
substrates. In wild-type populations, spontaneous mutants arise that show a Bgl ÷ phenotype as a result of the tran- scriptional activation of the operon. Most of the activating mutations are because of DNA transpositions, predomi- nantly of the insertion sequences IS1 and IS5, into the regulatory region of the operon termed bglR (Reynolds et aL, 1981). This is in contrast to the mutational inactivation of genes by transposable elements that is commonly observed. Initially four such insertions were mapped at the molecular level to a 47 bp target region upstream of the bgl promoter (Reynolds et aL, 1986). Subsequent analysis has extended the target region to 233 bp which include insertions downstream of the transcription start site (Schnetz and Rak, 1992).
In addition to insertion sequences, unlinked mutations that affect DNA supercoiling have been shown to activate the bgl operon (Di Nardo et aL, 1982; Higgins et aL, 1988). The operon is also subject to catabolite repres- sion. The only point mutations that activate the operon, iso- lated after extensive mutagenesis, are transitions within the CAP-cAMP binding region (Reynolds et aL, 1986; Lopilato and Wright, 1990). CAP-cAMP has been shown to bind with higher affinity to the mutant CAP site in vitro (Reynolds et aL, 1986). Upon activation, the bgl genes are subject to regulation by 13-glucosides via a mechanism involving antitermination of transcription (Mahadevan and Wright, 1987; Schnetz and Rak, 1988; Houman et aL, 1990).
We have attempted to elucidate the molecular mechan- ism of mutational activation of the bgl operon in order to explore the reasons for its crypticity. Transcriptional acti- vation of the operon may involve interaction of a specific protein with a recognition sequence present within the inser- tion sequence. Alternatively, the sequences upstream of the bgl promoter may contain structural elements that have a negative effect on transcription, as previously proposed by Lopilato and Wright (1990). Insertion sequences may be activating the operon by overcoming the negative effect of these structural elements either by disrupting the elements or by isolating them from the promoter. Transcription of the bgl promoter in vitro, deletion analysis of the target region, and mutagenesis of upstream sequences, reported here, implicate DNA structural elements involved in negative regulation.

1086 J. Singh, M. Mukerji and S. Mahadevan
Results
Transcription from the bgl promoter in vitro
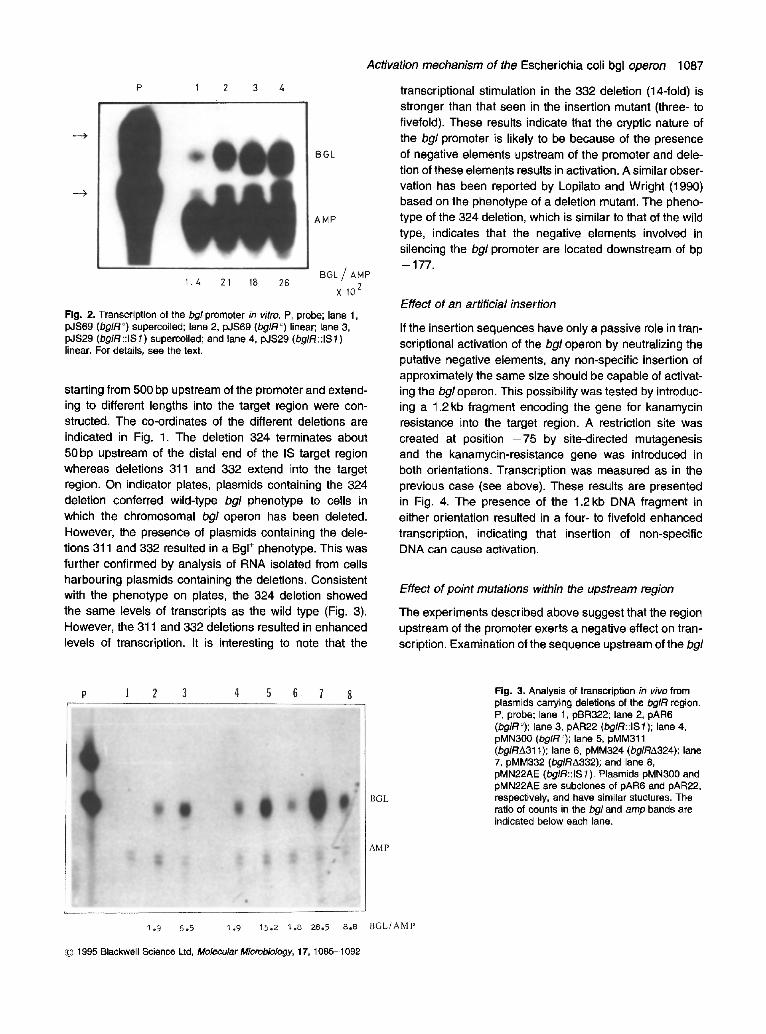
One possible mechanism by which insertion sequences may activate the bgl operon is by providing a binding sequence for an activator protein in a manner analogous to eukaryotic enhancer elements. Such a model has been reported previously (Schnetz and Rak, 1992). Alter- natively, insertion sequences may be disrupting DNA structural elements near the promoter that negatively affect transcription (Lopilato and Wright, 1990). To distin- guish between these two possibilities, we measured tran- scription from the wild-type (silent) bgl promoter and an IS/ activated bgl promoter in vitro. The activating IS 1 insertion is located at position - 9 2 (Fig. 1). Plasmids pJS29 and pJS69, containing the mutant and wild-type bgl promoter, respectively, were used as templates for tran- scription. The plasmids carried approximately 6kb DNA upstream of the bgl promoter in addition to the bgl regula- tory region and a part of the bglG gene.
Transcription reactions were carried out with either supercoiled or linear templates and RNA polymerase. The transcripts were analysed by hybridization to a 5'- labelled 52 bp oligonucleotide complementary to the first 42 nucleotides of the bgltranscript. The hybridization mix- ture also contained a 38 bp oligonucleotide complementary to 30 bases of the 13-1actamase transcript. The hybrids
were digested with $1 nuclease and analysed on a urea-acrylamide gel. The results of these experiments are shown in Fig. 2. Transcription from the wild-type pro- moter in the supercoiled state results in low levels of the bgl message. However, transcription from the template containing the mutant promoter showed a 13-fold enhance- ment. Upon linearization by digestion with a restriction enzyme, the wild-type promoter showed 15-fold enhanced transcription. Thus IS 1 can activate the bgl promoter in the absence of proteins other than RNA polymerase. Activa- tion is seen even with the wild-type promoter when the tem- plate DNA is in a relaxed state. This is consistent with the observation that gyr mutations that reduce negative super- coiling activate the operon (Di Nardo et aL, 1982). Hence, the inactive state of the wild-type promoter is most likely because of the presence of structural elements that are sensitive to the superhelical state of the DNA. However, it cannot be ruled out that, in vivo, this activation is modu- lated by specific proteins such as CAP-cAMP or non- specific proteins such as H-NS.
Deletion of the target region results in activation
The in vitro transcription experiments described above suggest that the cryptic nature of the bgl promoter is likely to be due to the presence of negative elements near the promoter. To test this possibility further, deletions
-230 - 1 9 0 ~ 3 2 4 I
5 ' TATCTI'CTACTACGTGAAGGGGCAGGATTTCCGTCACGTCGGTGGCGATGAGCTGGATAAACTGCTGGC
3 ' ATAGAAGATGATGCACTTCCCCGTCCTAAAGGCAGTGCAGCCACCGCTACTCGACCTATTTGACGACCG
-150 GG c ~311 -110
GGGGAAAGATAGCGACAAATAATTCACCAGACAAATCCCAATAACTTAATTATTGGGATTTGTTATATA D 4
CCCCTTTCTATCGCTGTTTATTAAGTGGTCTGTTTAGGGTTATTGAATTAATAACCCTAAACAATATAT
A332 I c-70 i '
TAACTTTATAAATTCCTAAAATTACACAAAG TTAATAA C ['G C GAG CATG GTCAT~TTTTTATCAATAGC
ATTG AAATATTTAAG G ATTTTAATG TG TTTC AA~TATTG AC G CTC G TAC C AG TA1]AAAAATAG TTAT--C-~
t . KAN
-30 -10 ,^x +1 +10
GC~TTTCTCTGCACGCAA'I~,ATTTCCGA?,C('TGGAT(;TTCGTTATAAAAACCATTA C G ~-X A C G A T J~ A A A (; A (; A (' (; T (; (' (; "r T ,xl:x m T:r..~ :~{r T A :x :x (; (; ( " r T G (; A C C "r A (' A A G C A A T A T T ; T T (; (; T A A T
+50 +90
ATAAATGACTGGATTGTTACTGCAT'rCGCAGGCAAA:gCCTGAUATAACCAGAGAATACTGGTGAAGTCGG 3' TATTTACTGACCTAACAATG AC GTAAG C GTCC GTTTTG GACTGTATTG GTCTCTTATGACCACTTCAG CC 5 '
Fig. 1. Features of the bgl regulatory region (bglR). The position of the activating IS1 insertion used in this study is shown by an upward arrow. The end-points of deletions are indicated by vert ical lines. The arrow at +1 indicates the direction of transcription. The C A P - c A M P binding site, - 3 5 and - 10 sequences are boxed. The T --, C transition at - 7 3 creates a Hpal site. The position of insertion of the kan gene in both orientation is also shown. The mutat ions within the region of dyed symmetry ( inverted arrows) creating an Mscl site are also indicated.
© 1995 Blackwell Science Ltd, Molecular Microbiology, 17, 1085-1092
Activation mechanism of the Escherichia coil bgl operon 1087
transcriptional stimulation in the 332 deletion (14-fold) is stronger than that seen in the insertion mutant (three- to fivefold). These results indicate that the cryptic nature of the bgl promoter is likely to be because of the presence of negative elements upstream of the promoter and dele- tion of these elements results in activation. A similar obser- vation has been reported by Lopilato and Wright (1990) based on the phenotype of a deletion mutant. The pheno- type of the 324 deletion, which is similar to that of the wild type, indicates that the negative elements involved in silencing the bgl promoter are located downstream of bp
- 177.
Fig. 2. Transcription of the bgl promoter in vitro. P, probe; lane 1, pJS69 (bg/R °) supercoiled; lane 2, pJS69 (bg/R °) linear; lane 3, pJS29 (bg/R ::IS 1 ) supercoiled; and lane 4, pJS29 (bg/R ::IS 1 ) linear. For details, see the text.
starting from 500 bp upstream of the promoter and extend- ing to different lengths into the target region were con- structed. The co-ordinates of the different deletions are indicated in Fig. 1. The deletion 324 terminates about 50bp upstream of the distal end of the IS target region whereas deletions 311 and 332 extend into the target region. On indicator plates, plasmids containing the 324 deletion conferred wild-type bgl phenotype to cells in which the chromosomal bgl operon has been deleted. However, the presence of plasmids containing the dele- tions 311 and 332 resulted in a Bgl + phenotype. This was further confirmed by analysis of RNA isolated from cells harbouring plasmids containing the deletions. Consistent with the phenotype on plates, the 324 deletion showed the same levels of transcripts as the wild type (Fig. 3). However, the 311 and 332 deletions resulted in enhanced levels of transcription. It is interesting to note that the
Effect of an artificial insertion
If the insertion sequences have only a passive role in tran- scriptional activation of the bgl operon by neutralizing the putative negative elements, any non-specific insertion of approximately the same size should be capable of activat- ing the bgl operon. This possibility was tested by introduc- ing a 1.2 kb fragment encoding the gene for kanamycin resistance into the target region. A restriction site was created at position - 7 5 by site-directed mutagenesis and the kanamycin-resistance gene was introduced in both orientations. Transcription was measured as in the previous case (see above). These results are presented in Fig. 4. The presence of the 1.2 kb DNA fragment in either orientation resulted in a four- to fivefold enhanced transcription, indicating that insertion of non-specific DNA can cause activation.
Effect of point mutations within the upstream region
The experiments described above suggest that the region upstream of the promoter exerts a negative effect on tran- scription. Examination of the sequence upstream of the bgl
Fig. 3. Analysis of transcription in vivo from plasmids carrying deletions of the bglR region. P, probe; lane 1, pBR322; lane 2, pAR6 (bg/R°); lane 3, pAR22 (bg/R::lS 1 ); lane 4, pMN300 (bglR °); lane 5, pMM311 (bg/RA311); lane 6, pMM324 (bg/RA324); lane 7, pMM332 (bglRA332); and lane 8, pMN22AE (bglR::lS 1). Plasmids pMN300 and pMN22AE are subclones of pAR6 and pAR22, respectively, and have similar stuctures. The ratio of counts in the bg/and amp bands are indicated below each lane.
© 1995 Blackwell Science Ltd, Molecular Microbiology, 17, 1085-1092
1088 J. Singh, M. Mukerji and S. Mahadevan
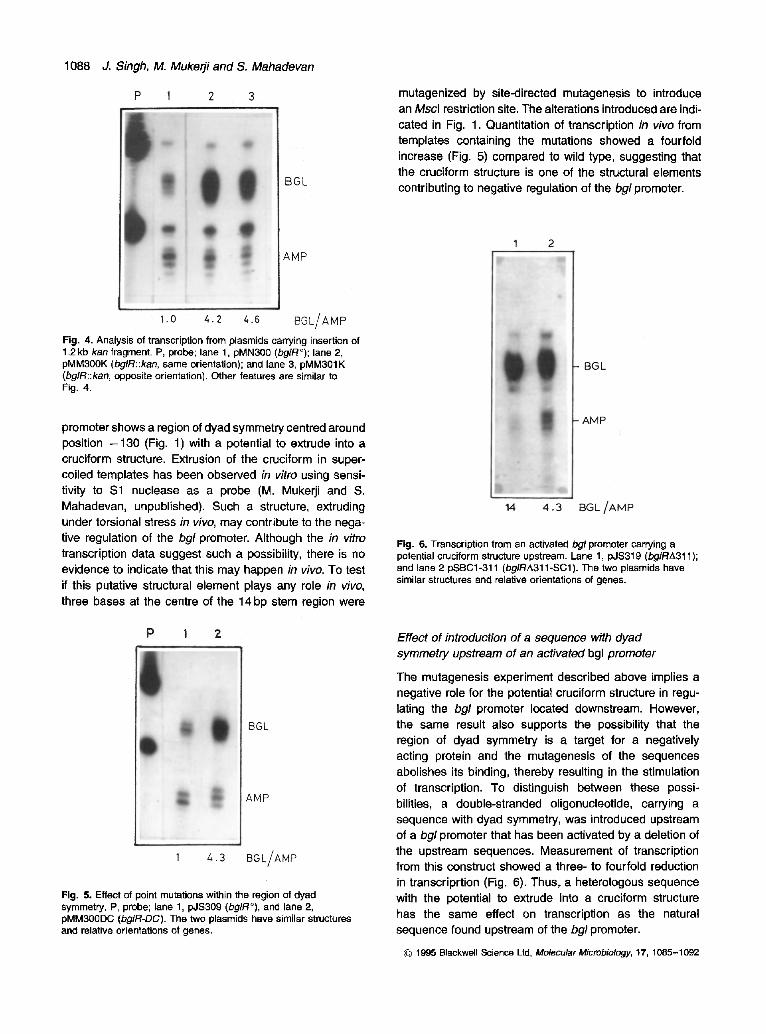
mutagenized by site-directed mutagenesis to introduce an Mscl restriction site. The alterations introduced are indi- cated in Fig. 1. Quantitation of transcription in vivo from templates containing the mutations showed a fourfold increase (Fig. 5) compared to wild type, suggesting that the cruciform structure is one of the structural elements contributing to negative regulation of the bgl promoter.
Fig. 4. Analysis of transcription from plasmids carrying insertion of 1.2 kb kan fragment. P, probe; lane 1, pMN300 (bglR°); lane 2, pMM300K (bglR::kan, same orientation); and lane 3, pMM301K (bglR::kan, opposite orientation). Other features are similar to Fig. 4.
promoter shows a region of dyad symmetry centred around position -130 (Fig. 1) with a potential to extrude into a cruciform structure. Extrusion of the cruciform in super- coiled templates has been observed in vitro using sensi- tivity to $1 nuclease as a probe (M. Mukerji and S. Mahadevan, unpublished). Such a structure, extruding under torsional stress in vivo, may contribute to the nega- tive regulation of the bgl promoter. Although the in vitro transcription data suggest such a possibility, there is no evidence to indicate that this may happen in vivo. To test if this putative structural element plays any role in vivo, three bases at the centre of the 14 bp stem region were
Fig. 5. Effect of point mutations within the region of dyad symmetry. P, probe; lane 1, pJS309 (bglR°), and lane 2, pMM300DC (bglR-DC). The two plasmids have similar structures and relative orientations of genes.
Fig. 6. Transcription from an activated bgl promoter carrying a potential cruciform structure upstream. Lane 1, pJS319 (bglRA311); and lane 2 pSBC1-311 (bglRA311-SC1). The two plasmids have similar structures and relative orientations of genes.
Effect of introduction of a sequence with dyad symmetry upstream of an activated bgl promoter
The mutagenesis experiment described above implies a negative role for the potential cruciform structure in regu- lating the bgl promoter located downstream. However, the same result also supports the possibility that the region of dyad symmetry is a target for a negatively acting protein and the mutagenesis of the sequences abolishes its binding, thereby resulting in the stimulation of transcription. To distinguish between these possi- bilities, a double-stranded oligonucleotide, carrying a sequence with dyad symmetry, was introduced upstream of a bgl promoter that has been activated by a deletion of the upstream sequences. Measurement of transcription from this construct showed a three- to fourfold reduction in transcriprtion (Fig. 6). Thus, a heterologous sequence with the potential to extrude into a cruciform structure has the same effect on transcription as the natural sequence found upstream of the bgl promoter.
© 1995 Blackwell Science Lid, Molecular Microbiology, 17, 1085-1092

Activation mechanism of the Escherichia coil bgl operon 1089
Discussion
The bgl operon has the unusual feature of possessing a cryptic promoter that is activated by spontaneous muta- tions. We have shown that the silent nature of the wild- type promoter is the result of the presence of DNA struc- tural elements upstream of the promoter. This conclusion is supported by several lines of evidence: in vitro trans- cription, deletion analysis, insertion of non-specific DNA sequence upstream of the promoter, and mutagenesis of a potential cruciform structure near the promoter.
One of the possible ways insertion sequences can medi- ate transcriptional activation is by providing sites of inter- action for a cellular protein that causes enhancement of transcription. However, the in vitro transcription studies using templates containing the bgl promoter show that the wild-type promoter is transcriptionally inactive when the template DNA is supercoiled. The presence of the insertion sequence is sufficient to cause a 13-fold activa- tion of the promoter in vitro. The in vitro transcription studies also show that activation can be observed even with the wild-type promoter if the template is relaxed. Therefore, the silent nature of the wild-type promoter is likely to be because of the presence of structural elements that are sensitive to DNA supercoiling. The activation seen in vitro is higher (13-fold) compared to the activation seen in vivo (three- to fivefold). The negative effect of DNA struc- tural elements is possibly enhanced in vivo by the binding of proteins such as H-NS. Alternatively, the increased tran- scriptional activation seen in vitro is due to the possibility that the superhelical density of the DNA template used in the in vitro experiments is higher than that found in vivo. This may help reduce the transcription from the wild-type promoter to a level which is lower compared to that seen in vivo. In the absence of exact measurements of super- helical densities of the DNA templates used, this possibi- lity cannot be ruled out.
Schnetz and Rak (1992) reported evidence for the role of sequences within the insertion sequence IS5 for tran- scriptional activation. The product of the ins5A gene was postulated to function as a transcriptional activator by bind- ing to a specific sequence within the IS5 element. The model proposed by Schnetz and Rak (1992) for the activa- tion of the bgl promoter also invokes the negative effect of DNA structural elements. However, in this case, the bind- ing of the Ins5A protein to sequences within the IS5 element is postulated to alter the topology of DNA near the promoter, thus overcoming the effect of the negative elements. Since the bgl operon is also activated by other insertion sequences, the effect seen with IS5 is likely to be specific for IS5 and may not constitute a general mechanism of activation. This is further supported by our observation that the gene encoding resistance to kana- mycin, in either orientation, is capable of activating the
bgl promoter in vivo when present upstream. Similar acti- vation is seen when the upstream region carries dele- tions. The deletion that extends close to the CAP-binding site results in a much stronger stimulation of the promoter in vivo compared to a naturally occuring IS 1.
The extent of activation reported by Schnetz and Rak (1992) is of the order of 40-60-fold. The maximum in vivo activation mediated by a natural IS 1 insertion that leads to a Bgl ÷ phenotype is only fivefold, as detected by our assay. It is difficult to make a direct comparison of the two quantifications because Schentz and Rak (1992) measured activation in the absence of an internal con- trol, whereas the extent of activation reported here is after normalization to an internal control.
Lopilato and Wright (1990) have also reported that the cryptic nature of the bgl promoter is due to negative elements present near the promoter. Insertion of linkers within the target region was shown to activate the bg/pro- moter. Interestingly, the extent of activation seen is less when the number of linkers is reduced. The linkers may either disrupt the negative elements or isolate them from the promoter. Therefore, insertion of a larger number of linkers is expected to produce a more pronounced effect. A deletion of the target region was also shown to activate the operon. Activation was quantified by measuring the enzymatic activity of a reporter gene. In this case, the effect is lower compared to that produced by an IS 1 inser- tion. The reduced activation seen by Lopilato and Wright (1990) may be due to the assay used. As we have assayed for the transcription from the bgl promoter by measuring the bgl transcript upstream of the attenuator in the leader region, influence of the attenuator is excluded. A comparison of the basal level of transcription reported by Lopilato and Wright (1990) shows that this is a likely possibility.
The deletion analysis presented in this report helps to define the upstream boundary of the sequences involved in negative regulation. The deletion 324, which starts at bp - 5 0 0 and removes all sequences upstream of the bgl promoter up to bp - 177, results in a phenotype similar to that of the wild type. This result shows that the negative elements are located downstream of bp -177. Deletions that extend further downstream result in strong transcrip- tional activation in vivo.
How does the region upstream of the promoter exert a negative effect on bg/transcription? One possibility is that the target region assumes a different secondary struc- ture when the template DNA is torsionally stressed. The region contains a sequence with dyad symmetry that can assume a cruciform structure when the DNA is negatively supercoiled (M. Mukerji and S. Mahadevan, unpublished). The deletions and insertions disrupt this sequence or iso- late it from the promoter. A substitution of three bases within the region of dyad symmetry, which is expected to
© 1995 Blackwell Science Ltd, Molecular Microbiology, 17, 1085-1092

1090 J. Singh, M. Mukerji and S. Mahadevan
destabilize the potential cruciform structure, produced a fourfold effect on transcription from the bgl promoter. There is a corresponding decrease in transcription when a synthetic sequence with a potential to form a cruciform structure is introduced upstream of an activated pro- moter. These results indicate that the cruciform structure is one of the negative elements involved in the regulation of the bgl promoter. The cruciform structure, when present upstream of the promoter, may exclude proper interaction of the polymerase to the promoter. Alternatively, it may inhibit the binding of other activators such as CAP- cAMP. There is increasing evidence that the presence of paranemic structural elements near genes and within coding sequences can affect gene expression (Brahma- chari et aL, 1990; Yagil, 1991). In most cases, such para- nemic structures are formed when the template DNA is torsionally stressed. Given the observation that the bgl operon is activated by mutation in the gyrgene, it is attrac- tive to speculate that in the mutant background, such struc- tures are not formed. However, additional evidence is needed to substantiate such a model.
Mutations at the locus osmZ which encodes the histone- like protein H-NS (Hulton et aL, 1990), also have been shown to activate the bgloperon. In this case, the mutations have been shown to increase negative supercoiling of a reporter plasmid. As the effect of the osmZ mutation is the opposite of that of the gyr mutations in terms of the superhelical density of DNA, one has to postulate a differ- ent mechanism for the effect of the osmZ mutation on bgl expression. The H-NS protein may stabilize paranemic structures by interacting directly with DNA upstream of the bgl promoter. Alternatively, the H-NS protein may act as a non-specific repressor of bgl expression. This has been shown in vitro in the case of the proV promoter (Ueguchi and Mizuno, 1993). The direct effect of the loss of negative regulation in the osmZ mutant may be more significant than the indirect effect of increased negative supercoiling.
In addition to the potential cruciform structure upstream of the bgl promoter, the region may also contain other structural elements that contribute to negative regulation. One possibility is that the DNA in this region has an intrin- sic curvature. Curved DNA has been implicated in positive and negative regulation (see Perez-Martin etaL, 1994 for a review). Curvature in an orientation opposite to that seen in the active promoter can result in negative regulation. Examination of restriction fragments carrying the bgl pro- moter region on acrylamide gels has not shown any evi- dence for bending (Lopilato and Wright, 1990). Therefore, the nature of additional negative elements, if any, is unknown at this time.
The cryptic bgl operon is thus activated by different mechanisms. The basic reason for its crypticity is appa- rently due to the presence of negative elements
upstream of the promoter. Insertion sequences either dis- rupt these elements or isolate them from the promoter leading to transcriptional activation. Tighter binding of the CAP-cAMP complex in strains carrying point mutations in the CAP site may also result in reducing the effect of the negative elements by facilitating interaction of the promoter with RNA polymerase. Thus, the two general schemes considered for the activation of the bgl pro- moter, namely negative regulation by DNA structural elements and positive regulation by activators, are not necessarily mutually exclusive. Mutations that decrease negative supercoiling of DNA, such as gyr mutations, may function by destabilizing the negative elements. Low levels of activation could be achieved under specific situ- ations by modulation of DNA supercoiling. However, the physiological significance of such a regulation with respect to the bgl operon is not clear at this time. In addi- tion, DNA-binding proteins such as H-NS may function by acting as non-specific repressors. Since DNA insertion sequences constitute the predominant class of sponta- neous mutations that activate the bgl operon, disruption of negative elements by DNA transposition appears to be the most common method of de-cryptification of the operon.
Experimental procedures
Plasmid constructions
Plasmid pMN300, which contains the wild-type bgl operon, was constructed by subcloning a 6.5kb EcoRI-Aval frag- ment from pAR6 (Reynolds et aL, 1986) into the Aval- EcoRI site of pBR322. The resulting 9.5 kb plasmid contains all of the bgl structural genes and the wild-type promoter (bglR°). The Pvul site upstream of the promoter was con- verted to an Xhol site by ligating with Xhol linkers after partial digestion with Pvul and treatment with Klenow fragment to remove the protruding ends. Plasmids pMN324, 311, and 332 were constructed by treatment with Ba131 nuclease (NEB) after linearizing pMN300 by Xhol digestion; this gene- rated deletions that originate at position -500 and terminate at -177, -123, and -76, respectively. The sequences upstream of the deletions in all the cases were made uniform by ligating 5 kb XhoI-EcoRI fragments from the deletion deri- vatives downstream of the Xhol site of pMN300. The resulting plasmids are pMM311, pMM324, and pMM332, respectively. The deletion in pMM311 extended to 1 base downstream of the point of insertion of IS 1 in pSAL6 and in pMM332, to 1 base downstream of IS5 in pAR13 (Reynolds et al., 1986).
The templates used in the in vitro transcription, pJS69 and pJS29, were constructed by ligating EcoRI-Hindlll fragments from pAR6 and pAR22 (Reynolds et aL, 1986), respectively, with pUC19 DNA digested with EcoRI and Hindlll. The result- ing plasmids contained the bgl promoter with 5.5kb of upstream DNA sequences and a portion of the bglG gene.
The piasmids pMM300K and pMM301K, containing, as an insertion, the 1.2kb kan gene fragment, were constructed by introducing an Hpal site at position -75 by site-directed
© 1995 Blackwell Science Ltd, Molecular Microbiology, 17, 1085-1092

Activation mechanism of the Escherichia coli' bgl operon 1091
mutagenesis (Kunkel, 1985). A 1 kb XhoI-Hpal fragment con- taining the bglR ° allele from pMN300 was subcloned into the vector pBluescript SK+ (Stratagene). The kan gene fragment was obtained from plasmid pUC4K (Pharmacia) as a 1.2 kb fragment. After making the ends flush using the Klenow frag- ment, the 1.2 kb fragment was ligated with the plasmid con- taining the Hpal site at position - 7 5 within the bglR region, subsequent to linearization by Hpal digestion. Recombinants containing insertion of the 1.2 kb fragment in both orientations were obtained by screening kanamycin resistant (Kan a) trans- formants. The bglR region containing the kan insertion was reintroduced into pMN300 using Mlul and EcoRV sites.
The plasmid pMM3OODC, containing the point mutations within the potential cruciform structure, was also constructed by site-directed mutagenesis as described above. Subclones containing the bglR ° (wild type) and the bglR-DC (disrupted cruciform) alleles were used in transcription analysis as described below. The plasmid pSBC1-311, carrying the synthetic sequence with dyad symmetry, was derived by ligating the 1 kb XhoI-Hindll l fragment from pMN311 down- stream of the Hindlll site in plasmid pSBC1 (Sarkar et aL, 1991) subsequent to making the ends flush using Klenow fragment. This introduces the sequence 5'-TGCTCGACGA- CACTCGAGAATCCTCGAGTGCCGTCGAGCA-3' upstream of the bglRA311 allele.
Analysis of transcripts
The plasmids containing deletions and insertions within the bglR region were introduced into the E. coil strain JF201 (Reynolds et aL, 1986) which is deleted for the chromosomal bgl operon. Analysis of transcripts was carried out as described below. The following oligonucleotides were used as probes after labelling the 5' end using [y-32p]-ATP (BARC) and T4 polynucleotide kinase (Bangalore Genei): oligonucleotide I (bgl) 5'-CCAGTCATTI'A'I-FAATGG'ITFFTA- TAACGAACATCCAGGTTCGGAAATrAAT-3'; and oligo- nucleotide II (amp), 5'-GCGGCGACCGAGTTGCTC'I-I-GCC- CGGCGTCATTAGCAT-3'.
Oligonucleotide I is complementary to region - 10 to +42 of the bgl sequence (Fig. 1). The 5' end of oligonucleotide II is complementary to 30 bases of the 13-1actamase (amp) tran- script. After $1 digestion, the protected fragment correspond- ing to the bgl transcript will be shorter than the full-length oligonucleotide. This was designed in order to differentiate between the protected fragment and the undigested probe, and also to detect specific transcription originating from the +1 site of bgl.
Isolation of RNA
E. coil strain JF201, in which the bgl operon is deleted, was transformed with various plasmids. The transformants were grown to exponential phase in minimal medium supple- mented with 0.4% w/v casamino acid, 0.4% succinic acid and 1001~gm1-1 ampicillin. The cells were washed with a buffer (4°C) containing 50mM glucose, 100mM Tris-HCI pH 8.0, and 50 mm EDTA, resuspended in 3 ml of the buffer containing 1001~g m1-1 lysozyme and incubated at 37°C for 5 min. To the samples, SDS to a final concentration of 0.1% was added. After adding an equal volume of acid phenol
(65°C), the tubes were vortexed briefly and placed in a 65°C water bath for 10rain. The samples were centrifuged at 1 2 0 0 0 x g for 15rain and the aqueous phase was re- extracted with acid phenol followed by extraction with a mix- ture of chloroform:isoamyl alcohol (23:1). RNA was stored under ethanol.
Sl nuclease mapping
RNA (10 i~g) was precipitated and dissolved in 30 #1 of hybrid- ization buffer (40 mm PIPES, pH 6.4, 1 mM EDTA, pH 8.0 and 0.4 M NaCI) containing 50 000 c.p.m. (0.2 pmoles) each of the bgl and amp oligonucleotides. Probe concentrations were standardized to have excess of probe in each reaction. The mixture was incubated at 90°C for 10 min and then at 55°C for 16 h to allow hybridization. To the hybridization mixtures, 300111 of cold $1 buffer (280 mM NaCI, 50 mM sodium acetate pH 4.5, and 4.5 mM ZnSO4) containing 30 units of $1 nuclease (Sigma Chemicals) was added. The samples were incubated at 37 °C for 45 min. The reactions were terminated by addition of 80 rtl of $1 stop buffer containing 4 M ammonium acetate and 50 mM EDTA. After precipitation with 2vol. of ethanol in the presence of 50 I~g ml-1 of yeast tRNA as carrier, the pel- lets were rinsed with 70% ethanol, resuspended in forma- mide dye and electrophoresed on 12% urea-acrylamide gel. The bands were visualized by autoradiography.
Quantification of transcripts
Bands corresponding to the bgl and amp transcripts were cut from the gel and counts taken using a scintillation counter (Beckman). The ratio of bgl to amp counts was taken in each case to normalize the values. To avoid the possibility that the amp transcription itself may be influenced by the supercoiling status of the template (Aleixandre and Blanco 1987), bgl transcription was also normalized to the expres- sion of the kan gene by repeating the $1 experiments using similar plasmids carrying the Kan a marker. These experi- ments showed that the ratios of activation seen with the Kan a plasmids are similar to that seen with the ampicillin resistant (Amp R) plasmids (data not shown).
Preparation of supercoiled template for in vitro transcription
Supercoiled template for in vitro transcription was prepared by subjecting pJS69 and pJS29 DNA to a neutral sucrose- gradient centrifugation to remove nicked molecules. The DNA samples were loaded on a neutral sucrose gradient (5-20%) and centrifuged using an SW28 rotor at 26000 rpm for 18 h at 4°C. Fractions of 1 ml were collected from the gradient. To check the mobility of the plasmid DNA, 5 I~1 of each fraction was loaded on a 1% agarose gel. Fractions containing 100% supercoiled plasmid DNA were pooled and the DNA was precipitated. Sucrose was removed from DNA by dialysis against Tr is-EDTA (TE) pH 8.0.
In vitro transcription
Transcription reactions were carried out in a final volume of
© 1995 Blackwell Science Ltd, Molecular Microbiology, 17, 1085-1092

1092 J. Singh, M. Mukerji and S. Mahadevan
25 p.I. The reaction mixture consisted of 40 mM Tris-HCI pH 7.9, 10mM MgCI2, 1 mM EDTA, 40mM NaCI, 0.04mg m1-1 BSA, 1 mM DTF, 50 units RNase inhibitor (Sigma Chemicals), 10% glycerol and 1 I~g of template DNA. Ribonucleotide triphos- phates were used at a final concentration of 0.3 mM, and 8 units of E. coli RNA polymerase was used in each reaction. Whenever necessary, the supercoiled DNA template was linearized by digestion with Hindlll. The reactions were incu- bated at 37°C for 45 min and stopped by the addition of an equal volume of stop buffer (0.6 M sodium acetate, 10mM EDTA, 501~g ml-1 carrier tRNA). RNA was precipitated with 2.5vo1. of 95% ethanol and was probed for bgl-specific tran- scripts as described above. Hybridizations were carried out at 40°C in the presence of 80% formamide. The transcripts were quantified by scanning an autoradiograph, exposed for the optimal time, using a densitometer (LKB).
Acknowledgements
We thank A, Wright for the gift of many strains and plasmids, D. Chatterji for the gift of RNA polymerase, S. K, Brahmachari for providing the pSBC1 plasmid, and A. Wright, V, Brahma- chari, V. Nanjundiah and the two referees for comments on the earlier version of the manuscript. We also thank L, Antony for expert technical assistance and S. Bharathi for typing the manuscript, We are also grateful to our colleagues in the sister departments at the Indian Institute of Science for making their facilities available during various stages of this work, The oligonucleotides used in this study were made available by the facility supported by the Department of Bio- technology, This work was funded by Grant BT/ReD/15/10/ 93 from the Department of Biotechnology. Financial support from the Director of the Indian Institute of Science is also gratefully acknowledged, J,S, was a recipient of a post- doctoral fellowship from the Department of Biotechnology, and M,M. received a Senior Research Fellowship from the University Grants Commission,
References
Aleixandre, V., and Blanco, M. (1987) Heterogeneity in the level of ampicillin resistance conferred by pBR322 deriv- atives with different supercoiling. Mol Gen Genet 209: 56- 60.
Brahmachari, S.K., Ramesh, N., Shouche, Y.S., Mishra, R.K., Bagga, R., and Meera, G. (1990) Unusual DNA structure: sequence requirements and role in transcription control. In Structure and Methods. Vol. 2: DNA Protein Complexes and Proteins. Sarma, R.H., and Sarma, M.H. (eds). New York: Adenine Press, pp. 33-49.
Di Nardo, S., Voelkel, K.A., Sternglanz, R., Reynolds, A.E.,
and Wright, A. (1982) Escherichia coil DNA toposiomerase I mutants have compensatory mutations in DNA gyrase genes. Cell 31: 43-51.
Higgins, C.F., Dorman, C.J., Stirling, D.A., Waddel, L., Broth, I.R., May, G., and Bremer, E. (1988) Physiological role of DNA supercoiling in the osmotic regulation of gene expression in S. typhimurium and E. coli. Cell52: 569-584.
Houman, F., Diaz-Torres, M.R., and Wright, A. (1990) Transcriptional antitermination in the bgl operon of E. coli is modulated by a specific RNA binding protein. Cell 62: 1153-1163.
Hulton, C.S.J., Seirafi, A., Hinton, C.J.D., Sidebotham, J.M., Waddel, L., Pavitt, G.D., Owen-Hughes, T., Spassky, A., Buc, H., and Higgins, C.F., (1990) Histone-like protein HI (H-NS), DNA supercoiling, and gene expression in bacteria. Cell 63:631-642.
Kunkel, T.A., (1985) Rapid and efficient site-specific muta- genesis without phenotypic selection. Proc Natl Acad Sci USA 82: 488-492.
Lopilato, J., and Wright, A. (1990) Mechanisms of activation of the cryptic bgl operon of E. coil K-12. In The Bacterial Chromosome. Drlica, K., and Riley, M. (eds). Washington, D.C.: ASM, pp. 435-444.
Mahadevan, S., and Wright, A. (1987) A bacterial gene involved in transcription antitermination: regulation at a rho- independent terminator in the bgl operon of E. coil Cell50: 485-494.
Perez-Martin, J., Rojo, F., and de Lorenzo, V. (1994) Promoters responsive to DNA bending: a common theme in pro- karyotic gene expressin. Microbiol Rev 58: 268-290.
Reynolds, A.E., Felton, J., and Wright, A. (1981) Insertion of DNA activates the cryptic bgl operon of E. coli. Nature 293: 625-629.
Reynolds, A.E., Mahadevan, S., Le Grice, S.F.J., and Wright, A. (1986) Enhancement of bacterial gene expression by insertion elements or by mutation in a CAP-cAMP binding site. J Mol Bio1191: 85-95.
Sarkar, P.S., Bagga, R., Balagurumurthy, P., and Brahma- chari, S.K. (1991) A novel approach to design cis-acting DNA structural elements for regulation of gene expression in vivo. Curr Sci 60: 586-591.
Schnetz, K., and Rak, B. (1988) Regulation of the bgloperon of Escherichia coil by transcription antitermination. EMBO J 7: 3271-3277.
Schnetz, K., and Rak, B. (1992) A mobile enhancer of transcription in Escherichia coil Proc Natl Aca Sci USA 89: 1244-1248.
Ueguchi, C., and Mizuno, T. (1993) The Escherichia coil nucleoid protein H-NS functions directly as a transcriptional repressor. EMBO J 12: 1039-1046.
Yagil, G. (1991) Paranemic structures of DNA and their role in DNA unwinding. Crit Rev Biochem Mol Bio126: 475-559.
© 1995 Blackwell Science Ltd, Molecular Microbiology, 17, 1085-1092





























