Embed Size (px)
Citation preview

www.elsevier.com/locate/ygcen
General and Comparative Endocrinology 153 (2007) 189–197
Tracking the evolution of the proenkephalin gene in tetrapods
Erin Roberts, Pezhman Shoureshi, Katarzyna Kozak, Laura Szynskie,Andrea Baron, Stephanie Lecaude, Robert M. Dores *
Department of Biological Sciences, University of Denver, 2190 E. Iliff, Olin Hall 102, Denver, CO 80210, USA
Received 11 December 2006; accepted 21 February 2007Available online 3 March 2007
Abstract
In gnathostomes there is remarkable consistency in the organization of the proenkephalin gene. This opioid precursor encodes sevenopioid (YGGF) sequences: five pentapeptide sequences, a met-enkephalin-7 sequence and a met-enkephalin-8 sequence. Yet, within ver-tebrate lineages there can be distinct sets of pentapeptide opioids (YGGFM or YGGFL). In the Sarcopterygii, the sixth opioid positionin lungfishes and anuran amphibian proenkephalin genes encodes a met-enkephalin (YGGFM) sequence. However, in mammalian pro-enkephalin there is a leu-enkephalin (YGGFL) sequence at this position. This study was done to test the hypothesis that the presence ofthe leu-enkephalin sequence in mammals is a feature common to amniote vertebrates, but not present in anamniote vertebrates. Toresolve this issue, proenkephalin cDNAs were cloned from the urodele amphibians, Amphiuma means and Necturus maculosus, andtwo amniote vertebrates, the turtle, Chrysemys scripta, and the brown snake, Storeria dekayi. As predicted, a met-enkephalin sequenceis present at the sixth opioid position in urodele amphibians; whereas, a leu-enkephalin sequence is present at this opioid site in the reptileproenkephalin sequences. These data are consistent with the conclusion that the transition from a met-enkephalin sequence to aleu-enkephalin sequence at the sixth opioid position in tetrapod proenkephalins occurred in the ancestral proto-reptiles. Phylogeneticanalyses, using the Maximum Parsimony and Neighbor-Joining algorithms, of the amphibian proenkephalin sequences supported theposition that anuran and urodele amphibians are a monophyletic assemblage. The same analysis of reptile-related proenkephalinsequences, including the deduced amino acid sequence of a partially characterized alligator proenkephalin cDNA, could not conclusivelyresolve the phylogeny of the major reptilian orders.� 2007 Elsevier Inc. All rights reserved.
Keywords: Proenkephalin; Reptiles; Urodele amphibians; Evolution
1. Introduction
Within the opioid/orphanin gene family proenkephalinis the common precursor for pentapeptide opioids(YGGFX), and two C-terminally extended forms of met-enkephalin; met-enkephalin-7 (YGGFMXX) and met-enkephalin-8 (YGGFMXXX) (Douglass et al., 1984).Among the gnathostomes, the organizational plan for theproenkephalin precursor is remarkably conserved. Forexample in lineages as diverse as mammals, cartilaginousfish and bony fish there are usually seven opioid sequences
0016-6480/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygcen.2007.02.023
* Corresponding author. Fax: +1 303 871 3471.E-mail address: [email protected] (R.M. Dores).
present in this precursor (for review see: Khalap et al.,2005). These opioid sites can be labeled A–G (Kilpatricket al., 1981), and in all gnathostomes that have been stud-ied to date, opioid positions A, B, C and E encode a met-enkephalin (YGGFM) sequence, whereas opioid positionD usually encodes the met-enkephalin-8 sequence, and opi-oid position G encodes the met-enkephalin-7 sequence (seeFig. 1; Khalap et al., 2005). The met-enkephalin-8 sequenceappears to be subclass specific. For example, in the non-tel-eost ray-finned fishes, Acipenser transmontanus (sturgeon),Polypterus senegalus (bicher), and Amia calva (bowfin),the sequence for met-enkephalin-8 is YGGFMRSI (Kha-lap et al., 2005), whereas in two species of lungfishes thesequence of this opioid is YGGFMRSL (Sollars et al.,2000; Dores et al., 2000). However, for all the bony fish
AUSLUNG MALL--VKCSCLLLVLCACLILAVWAECSKDCAHCTYHLGQQAEINPLSCTLECEGKLSST 61AMPHI MALLLLRHCVCLILVLATCHVSTVRAECSKDCAACTYRPGLRADINPLACTLECEGKLPSS NECT MALLLLKHCCCLLLVLAACLIGAVRSECSEDCAACTFRLGHQADINPLTCTLECEGKLPSS TARICHA MALLLLRNCCRLLLVLATCHIAAVLGECSKDCASCAYRQGARTDINPLTCTLECEGKLPSS BOMBINA MQLVL--RHCCLALALTTYLFVAVRADCSKDCATCAYHLGQQAELNSLACTLECEGKLPLA SPEA MHTTVGSKHCCLVLAITTSLSVGIQAESSKDCASCTYHLGQQTEINSLACTLECEGKLSSA TURTLE MALLL-RLCGLLLLALSSCLVWTVQADCSKDCASCSYRLGHHAEINPLACTLECEGKLPSA SNAKE MTLLL-RCYFLLL--AFAYLILTIQAECTKDCASCAHQLAYHADINPLACTLECEGKLPSA CHICKEN MALLL-RLGCSLLALSTCLLPTRARADCGRDCAACAYRLGPRADIHPLACTLECEFKLPSA HUMAN MARFL--TLCTWLLLLGPGLLATVRAECSQDCATCSYRLVRPADINFLACVLECEGKLPSL
** A ** B ** AUSLUNG KTWDMCKELLQAGKSEGTQE-GESTSTENEKESLERLLAKRYGGFMKRYGGFMKKMDELYH 122AMPHI KAWDTCKELLQIIKLDIAQQ----DGEKDPDRDENHLLAKKYGGFMKRYGGFMKKMDELHH NECT QAWDTFKEFLQVIKLDLVQQ----DGQKDPAGDEKHLLAKKYGGFMKRYGGFMKKMDELYT TARICHA KAWDTCKELLQLIKLDLVQQ----DGESD-QMDANHLLAKKYGGFMKRYGGFMKKMDELYR BOMBINA KAWGTCKELLQAAKEDNVQ-------EEKRTKATIALIAKKYGGFMKRYGGFMKKMDELYH SPEA KAWGPCRELLQLAKVDVCQ-----EAEKEPDNNDSHFIAKKYGGFMKRYGGFMKKMDELYR TURTLE KAWETCKELVQLTKLDLSEEGNSAPGDNKKEQDENHLLAKKYGGFMKRYGGFMKKMDELYH SNAKE KAWETCKELLQITKLDSP------------KEDESHLLAKKYGGFMKRYGGFMKKMDELYH CHICKEN KAWETCKELLQLTKLDLPEDGNAAPGD-KKELDENHLLAKKYGGRMKRYGGFMKKMDELYH HUMAN KIWETCKELLQLSKPELPQDG-TSTLRENSKPEESHLLAKRYGGFMKRYGGFMKKMDELYP
** C ** AUSLUNG LEPESENNGREILAKRYGGFMKKDPETGSLT-DSSDLLRDLLFGG---DNREGDYYMENTG 183AMPH VEPEEDYK-CEILAKRYGGFMKKDSDGE-----SADLLRELLGTVDGDNSEAGPNRDSNSE NECT VEPEEEYN-GEILAKRYGGFMKKDSDGE-----ATDLLRELLGTADGGNPEAGRYRDSANE TARICHA VEPEEENNNGEILAKRYGGFMKKDSDAE-----AADLLRELLGTGNGDNPEAGHYRDSAND BOMBINA VEPEDGENGGEILAKKYGGFMKKEADTD-----TSDLLRELLAT---------NGDPENND SPEA PEAEEDNNGGEILAKKYGGFMKKEFDSD-----TADLLRELLGS----NPEA-ISYHDNNE TURTLE VEPEEETNGGEILTKRYGGFMKKDSNDD-ALANSSDLLKELLGTG---ANAEASHYRQIND SNAKE VEPERKPNVGEILAKRYGGFMKKDSDDD-GLANSSDLLKELLATE---ENAEIGRYQDLSD CHICKEN PESEDEANGGEILAKRYGGFMKKDSDDD-ALANSSDLLKELLGAG---DNPEAAHYRGVNE HUMAN MEPEEEANGSEILAKRYGGFMKKDAEEDDSLANSSDLLKELLETG---DNRERSHHQDGSD
** D ** ** E ** ** F ** AUSLUNG KENNVMKRYGGFMRSLKRSTDQEDMAKDLQKRYGGFMRRVGRPEWKLDNQKRYGGFMRRFT 244AMPHI SQGDVSKRYGGFMRGIKRSPEMEEGAKDLHKRYGGFMRRVGRPEWWQDYQKRYGGFMRRFT NECT SQGEVSKRYGGFMREVKRSPEMEEGAKDLHKRYGGFMRRVGRPEWWLDYQKRYGGFMRRFA TARICHA SQGEVSKRYGGFMRGVKRSPEMEEGAKDLHKRYGGRMRRVGRPEWWLDYQKRYGGFMRRFA BOMBINA TPEEMSKRYGGFMRGYRRSPEAEDDTRVLQKRYGGFMRRVGRPEWWQDYQKRYGGFMRRFA SPEA IPGEINKRYGGFMRNYRRSPDVEDEAREMHKRYGGFMRRVGRPEWWQDYQKRYGGFMRRFS TURTLE NDGEVSKRYGGRMRSFKRSPELEDEAKELQKRYGGFMRRVGRPEWWLDYQKRYGGFLKRFT SNAKE SDSEVLKRYGGFMRSIKRSPELEGDAKELQKRYGGFMRRVGRPEWWMDYQKRYGGFLKRFA CHICKEN NDGDVSKRYGGFMRSVKRSPELEDEAKELQKRYGGFMRRVGRPEWWLDYQKRYGGFLKRFA HUMAN NEEEVSKRYGGFMRGLKRSPQLEDEAKELQKRYGGFMRRVGRPEWWMDYQKRYGGFLKRFA
** GAUSLUNG DP-LFSEEDSEMNSTEDPDTEKRYGGFMGY 273AMPHI DYSAPSDEDGESYSKEIPEMEKRYGGFMRFNECT DYSAPSDEDGESYSKEIPEMEKRYGGFMRITARICHA DSSAPSEEEAESYSKEIPEMEKRYGGFMRYBOMBINA DSLLPSDEDGESYSKEVPEVEKRYGGFMRYSPEA DSVLPSDEDGESYSKEIPEMDKRYGGFMRFTURTLE DSFLPSDEDGESYSKEVPAMEKRYGGFMRFSNAKE DTLLPSDDDGENYSKEAPEMEKRYGGFMRFCHICKEN DSILPSEEDGETYSKEVPEMEKRYGGFMRFHUMAN EA-LPSDEEGESYSKEVPEMEKRYGGFMRF
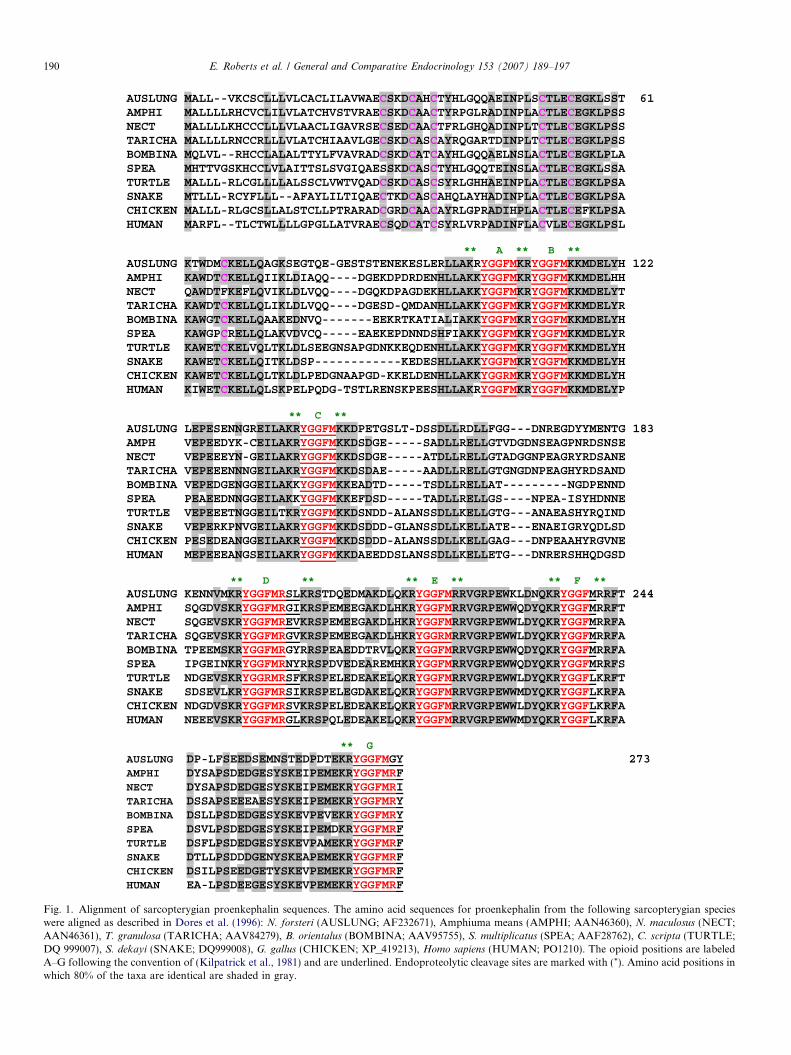
Fig. 1. Alignment of sarcopterygian proenkephalin sequences. The amino acid sequences for proenkephalin from the following sarcopterygian specieswere aligned as described in Dores et al. (1996): N. forsteri (AUSLUNG; AF232671), Amphiuma means (AMPHI; AAN46360), N. maculosus (NECT;AAN46361), T. granulosa (TARICHA; AAV84279), B. orientalus (BOMBINA; AAV95755), S. multiplicatus (SPEA; AAF28762), C. scripta (TURTLE;DQ 999007), S. dekayi (SNAKE; DQ999008), G. gallus (CHICKEN; XP_419213), Homo sapiens (HUMAN; PO1210). The opioid positions are labeledA–G following the convention of (Kilpatrick et al., 1981) and are underlined. Endoproteolytic cleavage sites are marked with (*). Amino acid positions inwhich 80% of the taxa are identical are shaded in gray.
190 E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197

E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197 191
proenkephalin sequences that have been characterized thusfar the met-enkephalin-7 sequence is always found to beYGGFMGY (Khalap et al., 2005). Thus, the met-enkeph-alin-7 and met-enkephalin-8 sequences could be useful forphylogenetic studies.
By far the most variable opioid site in proenkephalin isopioid site F. In mammals a leu-enkephalin sequence isfound at this penultimate opioid position (Douglass et al.,1984). However, in several species of anuran amphibians amet-enkephalin sequence is found at this site (Martens andHerbert, 1984; Wong et al., 1991; Lecaude et al., 1999). Amet-enkephalin sequence is also present at this position intwo species of lungfishes (Dores et al., 2000; Sollars et al.,2000) and in the proenkephalin sequence of a shark (Khalapet al., 2005). Based on these observations it has been pro-posed that the presence of a met-enkephalin sequence at opi-oid position F may represent the ancestral condition forgnathostomes (Khalap et al., 2005). However, the evolutionof amino acid substitutions at this opioid position is morecomplex when ray-finned fish are factored into the analysis.In a proenkephalin cDNA cloned from the brain of the stur-geon Acipenser transmontanus a relic opioid sequence wasdetected at this position (Danielson and Dores, 1999),whereas in a proenkephalin cDNA cloned from the brainof the bicher, Polypterus senegalus, a YGGFF sequencewas present at this position (Khalap et al., 2005). Finally,in neopterygian ray-finned fish like Amia calva and Danio
rerio there is a leu-enkephalin sequence at this position(Gonzalez Nunez et al., 2003; Khalap et al., 2005). Collec-tively these observations indicate that variations in the opi-oid sequence at position F have been influenced byreversions resulting in secondary changes at this position.Hence, the phylogeny of amino acid substitutions at opioidsites F appears to be lineage specific. Although recent studieshave focused on variation in the proenkephalin-derived opi-oids in ray-finned fish (Khalap et al., 2005), there are severalunresolved issues with respect to the radiation of the proen-kephalin gene in the Sarcopterygii.
Since a met-enkephalin sequence is found at opioid site Fin lungfish, the oldest extant lineage within the Sarcopterygii,and in anuran amphibians, the operating hypothesis for thisstudy was that in the early radiation of this class, as repre-sented by infraclasses Dipnoi and Amphibia, the presenceof a met-enkephalin sequence at opioid site F is the ancestralcondition. Hence, the presence of a leu-enkephalin sequenceat this position in mammals would be the derived condition,and this change must have occurred at some point in the radi-ation of the ancestral amniotes. If this hypothesis is correctthen the proenkephalin gene of urodele amphibians shouldencode a met-enkephalin sequence at opioid site F, and theproenkephalin gene in extant reptiles should encode a leu-enkephalin sequence at opioid site F. To test this hypothesis,proenkephalin cDNAs were cloned from the urodeleamphibians, Amphiuma means and Necturus maculosus,and two species of reptiles, the turtle, Chrysemys scripta
(Order Chelonia), and the brown snake, Storeria dekayi
(Suborder Serpentes).
2. Methods
2.1. Animals
Adult turtles (C. scripta), and adult urodele amphibians (A. means andN. maculosus) were purchased from Carolina Biological Co. (Burlington,NC) and kept in a free-running freshwater aquarium at 25 �C. The animalswere sacrificed by decapitation following the guidelines provided by theUniversity of Denver Animal Care committee. The brain from each ani-mal was removed intact, flash frozen in liquid nitrogen and stored in a�80 �C freezer. Adult brown snake (S. dekayi) brains were kindly pro-vided by Dr. Valantine Lance (San Diego Zoo), and adult alligator brains(Alligator mississippiensis) were provided by Dr. Louis Guillette (Univer-sity of Florida). The animals were sacrificed by decapitation followingthe animal care guidelines of their respective institutions and the brain tis-sue was collected from each species. The tissue was flash frozen in liquidnitrogen and stored at �80 �C for later use.
2.2. Cloning of proenkephalin cDNAs
Polyadenylated mRNA was isolated from brain extracts of each spe-cies by direct capture onto oligo dT25 paramagnetic beads (Novagen,Madison, WI) using the procedure of Jakobsen et al. (1994). First strandsynthesis was done on each set of mRNAs using superscript II reversetranscriptase (Gibco BRL, Grand Island, NY) and the first strand synthe-sis was primed off the poly A+ tail of the mRNA using the primer anchordT17 (50-GACTCGAGTCGGATCCATCGATTTTTTTTTTTTTTTTT-30)(1 lM). Finally, a homopolymeric guanine tail was added to the3 0-end of all first-strand cDNAs using terminal deoxynucleotidyl transfer-ase (Gibco, Gaithersburg, MD).
A combination of 3 0-RACE and 5 0-RACE polymerase chain reaction(PCR) procedures (Frohman et al., 1988) were used to amplify the proen-kephalin cDNAs from each species. For each species overlapping3 0-RACE and 5 0-RACE products were cloned and sequenced. The detailsof the RACE procedures will be provided for each species. FollowingPCR, the amplified products were purified on Wizard PCR prep columns(Promega, Madison, WI), and then separately fractionated on a 2% lowmelting point agarose (Sigma) gel that was run in 1· TAE running buffer(0.04 M Tris–acetic acid, 0.001 M EDTA, pH 8.0). The PCR productswere extracted from the agarose gel using Wizard PCR Prep columns(Promega, Madison, WI), and were separately ligated into the pGEM-Tvector (Promega, Madison, WI) at 4 �C for 12 h. Following incubation,the ligated vector was ethanol precipitated, electroporated into DH5aEscherichia coli bacterial cells, and plated onto a carbenicillin and X-galpositive LB/CARB agar plate for approximately 18 h. Colonies that con-tained the appropriate plasmids were cultured for 16–18 h at 37 �C in 2 mlof LB/CARB broth. Afterwards, the plasmids were isolated using WizardPlus SV minipreps (Promega) in 50 ll of sterile water. Each purified plas-mid was sequenced on a Beckman-Coulter CEQ 2000 Autosequencer.Sequencing results were then analyzed online using the BLAST algorithm(Altschuyl et al., 1997) from the National Center for BiotechnologyInformation.
Maximum Parsimony analysis and Neighbor-Joining analysis weredone using the PAUP 4.3 (Swofford, 1998) program. Amino acid sequencealignment was done as described in Dores et al. (1996). The Australianlungfish proenkephalin sequence was used as the outgroup for theseanalyses.
2.3. Turtle proenkephalin cDNA
For 3 0-RACE procedure the degenerate forward opioid primer,POMC 1024 (5 0-AA[A/G][A/C]GITA[C/T]GGIGGITT[C/T]ATG-3 0)was used with the reverse primer, Anchor dT17 (5 0-GACTCGAGTCGGATCCATCGA(T)17-3 0). The thermal profile for this PCR was: initial dena-turation at 94 �C for 10 min followed by 31 cycles of denaturing at 94 �Cfor 1.5 min, primer annealing at 56 �C for 1.5 min, and extension at 72 �Cfor 3 min. Two identical 3 0-RACE clones were sequenced.

192 E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197
The nucleotide sequence of the consensus 3 0-RACE product was usedas a template to design the gene specific reverse primer, Turtle PROENKREV (5 0-GCCTCCATTTGTTTCCTCCTC-3 0) for the 5 0-RACE proce-dure. This primer was used in conjunction with the forward primer,DC10 (5 0-GAATTCGCGGCCGCTTCAGTCCCCCCCCCC-3 0) thatwas targeted to the 5 0-G-tail on each cDNA that was added after first-strand synthesis. The thermal profile for the 5 0-RACE procedure was: ini-tial denaturation at 94 �C for 5 min followed by 31 cycles of denaturing at94 �C for 1 min, primer annealing at 55.9 �C for 50 s, and extension at72 �C for 1.5 min. Three identical 5 0-RACE clones were sequenced toobtain the consensus sequence for the 5 0-RACE product.
2.4. Urodele amphibian proenkephalin cDNAs
For the 3 0-RACE procedure used for both the A. means and N. macu-
losus brain samples the forward primer was the degenerate opioid primer,POMC 1024, and the reverse primer was Anchor dT17. The thermal pro-file for the 3 0-RACE procedure was: initial denaturation at 94 �C for5 min, followed by 32 cycles of denaturing at 94 �C for 1 min, primerannealing (7 cycles at 56 �C; 27 cycles at 58 �C) for 1.5 min, and extensionat 72 �C for 2.5 min. For the A. means 3 0-RACE procedure two identicalclones were sequenced, and for the N. maculosus four identical clones weresequenced.
For the A. means 5 0-RACE procedure, the sequence of the 3 0-RACEproduct was used as template to design the gene specific reverse primerAmpProenkR1 (5 0-CCTTGATGGGCCTCCTCTTCC-3 0). This primerwas used in conjunction with the forward primer DC10. The thermal pro-file for the 5 0-RACE reaction was: initial denaturation at 94 �C for 5 min,then 32 cycles of denaturing at 94 �C for 1 min, primer annealing at 58 �Cfor 1 min, and extension at 72 �C for 2.5 min. For the A. means 5 0-RACEprocedure four identical clones were sequenced.
For the N. maculosus 5 0-RACE procedure, the sequence of the3 0-RACE product was used as template to design the gene specific reverseprimer NecENK-R2 (5 0-TCTGGGCTCCTCTTGACCT-3 0). This primerwas used in conjunction with the forward primer DC10. The thermal pro-file for the 5 0-RACE reaction was: initial denaturation at 94 �C for 5 min,then 32 cycles of denaturing at 94 �C for 1 min, primer annealing at 58 �Cfor 1 min, and extension at 72 �C for 3 min. For the N. maculosus
5 0-RACE procedure five identical clones were sequenced.
2.5. Snake proenkephalin cDNA
The snake 3 0-RACE procedure utilized the opioid-specific degenerateforward primer POMC-288: (5 0-AA[G/A][A/C]G[A/C/T]TA[C/T]GGIGG[A/C/G]TTCATG-30), and the reverse primer Anchor dT17. Thethermal profile for this PCR was: initial denaturation at 94 �C for 4 minfollowed by 33 cycles of the following: denaturing at 94 �C for 1 min,primer annealing at 56.4 �C for 1 min, extension at 72 �C for 3 min. Threeidentical clones were sequenced to obtain the consensus sequence forsnake proenkephalin 3 0-RACE product.
For the 5 0-RACE procedure the snake 3 0-RACE race proenkephalinproduct was used as a template to design the reverse primer, Brown SnakeproenkR1 (5 0-TCTGGAGCCTCTTTGGAATA-30). This primer wasused in conjunction with the forward primer, DC10. The thermal profilefor the 5 0-RACE procedure was: initial denaturation at 94 �C for 4 minfollowed by 33 cycles of the following: denaturing at 94 �C for 1 min, pri-mer annealing at 58.9 �C for 1 min, and extension at 72 �C for 3 min.Three identical clones were sequenced to obtain the consensus sequencefor snake proenkephalin 5 0-RACE product.
2.6. Alligator proenkephalin 3 0-RACE product
For the alligator 3 0-RACE reaction the degenerate opioid primer,POMC 1024 was used in conjunction with reverse primer AnchordT17. The thermal profile for this PCR was: initial denaturation at95 �C for 5 min followed by 33 cycles of the following: denaturingat 95 �C for 1 min, primer annealing at 56 �C for 1 min, and extension
at 72 �C for 3 min. Eight identical clones were sequenced to obtain theconsensus sequence for the alligator 3 0-RACE proenkephalin product.Repeated attempts to clone the 5 0-end of the alligator proenkephalincDNA using a variety of reverse primers designed from the alligator3 0-RACE proenkephalin sequence were not successful.
3. Results
By using the 3 0-RACE and 5 0-RACE proceduresdescribed in METHODS it was possible to obtain thesequences of proenkephalin cDNAs individually clonedfrom brain extracts of the urodele amphibians, A. means
(AAN46360) and N. maculosus (AAN46361), and the rep-tiles, C. scripta (DQ 999007) and S. dekayi (DQ999008).The nucleotide sequences for these cDNA are posted inGenBank, and the amino acid sequences of these proen-kephalin precursors appears in Fig. 1 along with the proen-kephalin sequences for the following sarcopterygianspecies: the Australian lungfish, Neoceratodus forsteri (Sol-lars et al., 2000), the newt, Taricha granulosa (Walthers andMoore, 2005), the toads, Bombina orientalus (Dores et al.,2001) and Spea multiplicatus (Lecaude et al., 1999), thechicken, Gallus gallus, (XP_419213), and human (Nodaet al., 1982). A total of seven gaps were required to alignthe seven opioid sites in the sarcopterygian proenkephalinsequences. Overall, 56% of the amino positions in the sar-copterygian proenkephalin sequences are conserved. In theN-terminal region of these precursors there are six con-served cysteine residues (C28, C32, C35, C50, C54, C67),and all of the precursors contained five pentapeptidesequences (sites A, B, C, E, F), along with a single met-enkephalin-8 sequence (site D) and a single met-enkepha-lin-7 sequence (site G).
Focusing first on the amphibian proenkephalinsequences presented in Fig. 1, the following trends wereobserved. In both the urodele (A. means, N. maculosus,
T. granulosa) and the anuran (B. orientalus, S. multiplica-
tus) proenkephalins there were met-enkephalin sequencesat all of the pentapeptide sites (A, B, C, E, F). At themet-enkephalin-8 site (D) all of the amphibian opioidshad the core sequence, YGGFMR. Yet there was consider-able primary sequence variability at amino acid positionsseven and eight, with no clear phylogenetic trend event.In a similar manner, at the met-enkephalin-7 site (G) allof the amphibian opioids had the core sequence,YGGFMR, with amino acid substitutions at position 7that also did not follow any phylogenetic trend. In spiteof these differences, Maximum Parsimony analysis of theamphibian proenkephalin sequences, using the Australianlungfish proenkephalin sequence as the outgroup, indicatedthat the anuran and urodele proenkephalin sequencescould be separated into two clades (Fig. 2a) that had robustbootstrap values. The anuran amphibian proenkephalinsequences formed a clade with a bootstrap value of 99%and the urodele amphibian sequences formed a clade witha bootstrap value of 97%. The urodele sequences could befurther divided into a clade consisting of the N. maculosus

AUS LUNGFISH
BOMBINA
SPEA
AMPHIUMA
NECTURUS
TARICHA
99%
97%
91%
a
b
c
AUS. LUNGFISH
TURTLE
SNAKECHICKEN
HUMAN
AUS. LUNGFISH
TURTLE
CHICKENSNAKE
HUMAN
99%
55%
Fig. 2. Phylogenetic analysis of tetrapod proenkephalin sequences. (a) Maximum Parsimony analysis, using the exhaustive search mode, was done on thefollowing amphibian proenkephalin sequences: A.means (AMPHIUMA), N. maculosus (NECTURUS), T. granulosa (TARICHA), B. orientalus
(BOMBINA), and S. multiplicatus (SPEA). Australian lungfish (N. forsteri) was used as the outgroup. Bootstrap analysis was done on 1000 replicates andthe bootstrap values are shown at the branch points. (b) Maximum Parsimony analysis, using the exhaustive search mode, was done on the followingamniote proenkephalin sequences: C. scripta ( TURTLE) and S. dekayi (SNAKE), G. gallus, (CHICKEN), and H.sapiens (HUMAN). Australian lungfish(Neoceratodus forsteri) was used as the outgroup. Bootstrap analysis was done on 1000 replicates and the bootstrap values are shown at the branch points.(c) Neighbor-Joining analysis of the data set analyzed in (b).
E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197 193
and T. granulosa sequences (bootstrap value of 91%) withthe A. means proenkephalin sequence as the sistersequence. The same branching pattern was obtained whenthe amphibians sequences were analyzed using the Neigh-bor-Joining method (data not shown).
When the same comparison was done for the amnioteproenkephalin sequences presented in Fig. 1 (C. scripta,
S. dekayi, G. gallus, human), a somewhat different set oftrends became apparent. Although all of the amniote pro-enkephalins had met-enkephalin sequences at opioid sitesA, B, C, and E, a leu-enkephalin sequence was present atopioid position F. In addition, unlike the amphibians, allthe amniotes had the same opioid sequence at position G(YGGFMRF). However, at the met-enkephalin-8 sequence(position D) although all species had the same opioid coressequence (YGGFMR) there was considerable species spe-cific variability at amino acid positions seven and eight inboth the amniote proenkephalin sequences and theamphibian proenkephalin sequences. Maximum Parsimonyanalysis of the amniote proenkephalin sequences yielded a
clade that included the turtle, snake and chicken proen-kephalin sequences with human proenkephalin as the sistersequence (Fig. 2b). Within this clade turtle proenkephalinwas the sister sequence to a clade consisting of the snakeand chicken sequences, but the bootstrap value forsnake/chicken was weak (55%). When the same data setwas analyzed using the Neighbor-Joining method(Fig. 2c), although human proenkephalin was still the sistersequence to a clade consisting of the turtle, snake, andchicken proenkephalin sequences, the branching patternfor this clade was not the same as observed in the Maxi-mum Parsimony analysis.
4. Discussion
Among the gnathostomes, the organizational scheme forproenkephalin appears to have undergone the fewest num-ber of lineage specific changes as compared to the otherprecursor-coding genes in the opioid/orphanin gene family(Dores et al., 2002). In the POMC gene there have been

194 E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197
lineage specific changes in the number of melanocortin-related end-products present in this precursor as reviewedpreviously (Dores and Lecaude, 2005). In the prodynor-phin gene there has been a reduction in the number of opi-oid sequences present in this precursor during the radiationof the Sarcopterygii (Dores et al., 2004). A similar trendmay also have occurred during the radiation of the pro-orphanin gene in gnathostomes (Danielson et al., 2001).By contrast, the number and position of opioid end-prod-ucts within the proenkephalin gene have remained stableamong the several lineages of gnathostome vertebrates.For example, the presence of seven opioid sites in the sar-copterygian proenkephalin sequences presented in Fig. 1 isalso seen in cartilaginous fish and ray-finned fish proen-kephalin sequences (Khalap et al., 2005.). Given these com-mon gnathostome features, the objectives for this studywere to evaluate trends in the organization of sarcoptery-gian proenkephalin sequences with a focus on the opioidsites D, F and G (Fig. 1).
With respect to opioid site F, the operating hypothesiswas that the presence of a met-enkephalin sequence at thisposition in lungfishes and anuran amphibians representedthe ancestral condition, whereas the presence of a leu-enkephalin sequence at this position in mammals repre-sented the derived condition. As observed in Fig. 1, amet-enkephalin sequence is present at this position in uro-dele proenkephalin sequences; whereas, a leu-enkephalinsequence is present at this position in two species of reptilesand one avian species. Collectively these observations sup-port the conclusion that the appearance of a leu-enkephalinsequence at opioid position F is a feature common to theamniotes. Furthermore, the transition from a met-enkeph-alin sequence at this position (the dipnoi/anamniote condi-tion) to a leu-enkephalin sequence (the amniote condition)must have occurred in the ancestral proto-reptiles at somepoint in the Carboniferous Period (Carroll, 1988).
The rather clear cut differences between opioidsequences at position F in anamniote tetrapods and amni-ote tetrapods is less apparent for opioid position D, themet-enkephalin-8 sequence (Fig. 1). Although both anam-niote and amniote tetrapods have the core sequence,YGGFMR at this position, there is a considerable degreeof primary sequence variability at amino positions sevenand eight in both the anamniote and amniote met-enkeph-alin-8 sequences. However, there does appear to be a ten-dency for a hydrophobic residue at amino acid positioneight in both the anamonite and amniote met-enkephalin-8 sequences. Similar trends were observed in the tetrapodmet-enkephalin-7 sequences presented in Fig. 1. All ofthe tetrapod met-enkephalin-7 sequences had the coresequence, YGGFMR, and all of these sequences had ahydrophobic residue at the C-terminal amino acid position.
Although the physiological significance of the primarysequence differences observed in the C-terminal regions ofvertebrate met-enkephalin-7 and met-enkephalin-8 hasnot been investigated, studies have been done on the opioidreceptor binding properties of met-enkephalin and
leu-enkephalin. While it is generally accepted that met-enkephalin and leu-enkephalin bind with equal affinity tothe delta opioid receptor (Kieffer et al., 1992), there appearto be differences in the manner in which these two opioidsbind to mu and kappa opioid receptors (Neilson, 2003). Inthe latter study, it was observed that while met-enkephalinbound to delta and mu opioid receptors with equal affinity,this ligand bound to the kappa opioid receptor with muchless affinity (d � l >> j). When the same analysis was donefor leu-enkephalin, this ligand was found to have the high-est binding affinity for the delta receptor. However, leu-enkephalin bound to the mu and kappa receptors withlower affinities, respectively (d > l >> j). The differencesin binding efficacy for met-enkephalin and leu-enkephalinappears to be a function of the proximity of the side chainrings on the tyrosine reside at position 1 and phenylalaninereside at position 4 in these ligands. The aromatic ringproximity in leu-enkephalin is predicted to be greater thanthe aromatic ring proximity in met-enkephalin because ofthe presence of the leucine residue at amino acid position5. This difference appears to influence the binding efficacyof these two ligands to delta and mu opioid receptors (Neil-son, 2003). Since the terminal fields for some enkephaliner-gic neurons are at postsynaptic neurons that expressmultiple opioid receptor subtypes (Mansour et al., 1988),the relative binding affinities of met-enkephalin and leu-enkephalin may influence the physiological response ofthe postsynaptic neuron. As a result there could be subtledifferences in the signal transduction events that occur insome anamniote enkephalinergic circuits as compared tothe same enkephalinergic circuits in amniotes. To date,comparative analyses to investigate this possibility havenot been done.
Phylogenetic analysis of the proenkephalin sequencespresented in Fig. 1, using the Maximum Parsimony algo-rithm, yielded a single most parsimonious tree in whichthe anamniote proenkephalin sequences and the amnioteproenkephalin sequences were segregated into two distinctclades (data not shown). A separate analysis of the anam-niote sequences (Fig. 2a) produced a single most parsimo-nious tree in which the anuran proenkephalin sequencesand the urodele proenkephalin sequences formed twoclades that each had robust bootstrap values. The topogra-phy of the Maximum Parsimony analysis presented inFig. 2a was supported by a Neighbor-Joining analysis doneon the same data set (data not shown). These observationsare consistent with the hypothesis that urodele amphibiansand anuran amphibians are a monophyletic assemblage(Carroll, 1988).
When the same phylogenetic analyses were performedon the amniote proenkephalin sequences presented inFig. 1 the outcomes of the Maximum Parsimony analysisand the Neighbor-Joining analysis were not identical(Fig. 2b). In both analyses human proenkephalin wasthe sister sequence to a clade that included the turtle,snake and chicken proenkephalin sequences. However,in the Maximum Parsimony tree turtle proenkephalin

a [ OPIOID-288 ]K K M D E L Y H V E P E E 20aaacgttacggggggttcatgaagaaaatggatgagctttaccatgtagaaccagaggag 60 * * [C] * *
E T T G G E I L A K R Y G G F M K K D S 40gaaaccactggaggagaaattcttgctaagcggtatggaggattcatgaagaaagactca 120
D D D T L A N S S D L L K E L L G T G D 60gatgatgacaccttggctaattcttctgatctgctgaaggagcttctgggaacaggagac 180 * *
N N E A G H Y R E I S E N D G E V S K R 80Aacaatgaagctggacattatcgagaaataagtgagaatgatggggaggtcagcaaaaga 240 [D] * *
Y G G F M R S I K R S P E L E D E A K E 100tatggcggcttcatgagaagcataaagcgcagtccagaactggaagatgaagccaaagag 300 * * [E] * *
L Q K R Y G G F M R R V G R P E W W L D 120ttgcaaaagagatatggtggctttatgagaagagtaggtaggccagaatggtggctagac 360 * * [F] * *
Y Q K R Y G G F L K R F T D S V L P S E 140taccagaaacgatatggtggatttcttaagcgcttcactgattctgttctgccttcagaa 420
* * [G]E D G E S Y S K E V P E M E K R Y G G F 160gaagatggggaaagttattccaaagaggttccagagatggaaaagagatatggtggattt 480
M R F STOP 163atgagattttaacaccctccccttttatcccttttgtcccaaatgagagagactgcctca 540ttgatcataatttattgtaatgtgttgcttgtactgtacagttttttactgtcctagtta 600atgatgcaatctgcaatctgttgtttcagggtccgtgttcttttaaatcaataagtaggt 660tctgttttagtccaatttaaatctatattgtagttttcatgccaaaaatattttggttac 720ctttatcccttttctttaaacagaacaacacatcttcagtaaaattgcttaactgtacat 780ataataaattcttcatcttaactgc 803 ++++++
AUS. LUNGFISH
HUMAN
TURTLE
ALLIGATOR
SNAKE
99%61%
b
Fig. 3. Partial sequence of an alligator proenkephalin cDNA. (a) The partial amino acid sequence of a proenkephalin cDNA cloned from brain extracts ofthe alligator, A. mississippiensis is present. The opioid positions are labeled C–G following the convention of (Kilpatrick et al., 1981) and are underlined.Endoproteolytic cleavage sites are marked with (*). (b) Maximum Parsimony analysis, using the exhaustive search mode, was done on the followingproenkephalin sequences: C. scripta (TURTLE) and S. dekayi (SNAKE), A. mississippiensis, (ALLIGATOR), and H. sapiens (HUMAN). Australianlungfish (Neoceratodus forsteri) was used as the outgroup. Bootstrap analysis was done on 1000 replicates and the bootstrap values are shown at thebranch points.
E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197 195
was the sister sequence to a snake/ chicken clade;whereas in the Neighbor-Joining tree snake proenkepha-lin was the sister sequence to turtle/chicken clade. Thesnake/chicken clade in the Maximum Parsimony tree isconsistent with an early view of reptile phylogeny thatplaced turtles (Order Chelonia) in Subclass Anapsida, areptilian group that has been viewed as the sister group
to squamate reptiles (lizards and snakes) and archeosaurs(crocodilian reptiles and birds; Carroll, 1988). However,the bootstrap value for the snake/chicken clade in theMaximum Parsimony tree was extremely weak (55%),and based on this data set it would be difficult to statewith conviction that members of Order Chelonia arethe sister group to all extant reptiles.

196 E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197
The topography of the Neighbor-Joining tree presentedin Fig. 2b is consistent with the hypothesis that Order Che-lonia should be placed into Subclass Diapsida (squamatereptiles and archeosaurs) and that the modern turtles aremore closely related to the archeosaurs than to the squa-mate reptiles. The latter hypothesis has received consider-able support from recent molecular phylogeny studies(Hedges and Poling, 1999; Kumazawa and Nishida, 1999;Iwabe et al., 2004). However, the proenkephalin data setpresented in Fig. 1 was not sufficient to support or refutethe Chelonia/Diapsida hypothesis. An important sequencenot present in Fig. 1 is the sequence of a crocodillian pro-enkephalin. In an effort to rectify this situation, the partialsequence of a proenkephalin cDNA cloned from brainextracts of the American alligator (A. mississippiensis) ispresented in Fig. 3a. Although repeated attempts to char-acterize the complete open reading frame of the alligatorproenkephalin cDNA were not successful, five of the pre-dicted seven opioid sequences were detected in this partialcDNA sequence. Note that the following features commonto amniotes were detected in the partial sequence of thealligator proenkephalin: (a) met-enkephalin sequences werepresent at opioid sites C and E; (b) a leu-enkephalinsequence was present at opioid site F; and (c) anYGGFMRF sequence is present at opioid site G. Maxi-mum Parsimony analysis of the turtle, alligator, snake,and human proenkephalin sequences using a data set thatcorresponded to the partial alligator proenkephalinsequences (positions 117–273 in Fig. 1) yielded the singlemost parsimonious tree presented in Fig. 3b. Using the cor-responding partial sequence of Australian lungfish proen-kephalin as the outgroup, the reptile proenkephalinsequences formed a clade with human proenkephalin asthe sister sequence. Within the reptilian clade, the snakeproenkephalin sequence was the sister sequence to a tur-tle/alligator clade. Although the bootstrap value (61%)for the turtle/alligator clade is still weak, Neighbor-Joininganalysis yielded a tree with the same topography (data notshown). Furthermore, it is anticipated that if the full lengthsequence for the amniote proenkephalins could have beenused in this phylogenetic analysis, the bootstrap value forthe turtle/alligator clade would improve. This conclusionis tempered by the realization that the amino acid sequenceof proenkephalin may not contain enough informativecharacters to resolve the phylogeny of the various reptil-ian-related orders. However, the data presented in Fig. 3would suggest that turtle genes may be more closely relatedto archeosaur genes, and that Chelonia/Diapsida hypothe-sis is the most reasonable way to interpret the phylogeny ofextant reptiles.
Acknowlegments
We thank Valentine Lance and Louis Gillette for provid-ing the snake and alligator tissues, respectively. This researchwas supported by NSF Grant IOB 0516958 (R.M.D.).
References
Altschuyl, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.I.I., 1997.Basic logical alignment search tool. Mol. Biol. 215, 403–410.
Carroll, R.L., 1988. Vertebrate paleontology and evolution. FreemanPress, New York.
Danielson, P.B., Dores, R.M., 1999. The evolution of the opioid/orphaningene family. Gen. Comp. Endocrinol. 113, 169–186.
Danielson, P., Hoverstein, M.T., Fitzpatrick, M., Schreck, C., Akil, H.,Dores, R.M., 2001. Sturgeon orphanin: a molecular ‘‘Fossil’’ thatbridges the gap between the opioids and orphanin FQ/N. J. Biol.Chem. 276, 22114–22119.
Dores, R.M., Lecaude, S., 2005. Trends in the evolution of theproopiomelanocortin gene. Gen. Comp. Endocrinol. 142, 81–93.
Dores, R.M., Rubin, D.A., Quinn, T.W., 1996. Mini-review: is it possibleto construct phylogenetic trees using polypeptide hormone sequences.Gen. Comp. Endocrinol. 103, 1–12.
Dores, R.M., Lee, J., Sollars, C., Danielson, P., Lihrmann, I., Vallarino,M., Vaudry, H., 2000. In the African lungfish met-enkephalin and Leu-enkephalin are derived from separate genes: cloning of a proenkeph-alin cDNA. Neuroendocrinology 72, 224–230.
Dores, R.M., Costantino, D., Walnutt, J., Danielson, P., Lecaude, S.,2001. Analyzing the radiation of the proenkephalin gene in tetrapods:cloning of a Bombina orientalis proenkephalin cDNA. Peptides 22,2021–2025.
Dores, R.M., Lecaude, S., Bauer, D., Danielson, P.B., 2002. Analyzing theevolution of the opioid/orphanin gene family. Mass Spectrom. Rev.21, 220–243.
Dores, R.M., Sollars, C., Lecaude, S., Lee, J., Danielson, P., Alrubaian, J.,Lihrman, I., Joss, J.M.P., Vaudry, H., 2004. Reconstructing theevolution of the prodynorphin gene: cloning of prodynorphin cDNAsfrom the brain of the Australian lungfish and the African lungfish.Neuroendocrinology 79, 185–196.
Douglass, I., Civelli, O., Herbert, H., 1984. Polyprotein gene expression:generation of diversity of neuroendocrine peptides. Annu. Rev.Biochem. 53, 665–715.
Frohman, M.A., Dush, M.K., Martin, G.R., 1988. Rapid production offull-length cDNAs from rare transcripts: amplification using a singlegene-specific oligonucleotide primer. Proc. Natl. Acad. Sci. USA 85,8998–9002.
Gonzalez Nunez, V., Gonzalez, S., Sarmiento, R., Rodriguez, R.E., 2003.Characterization of zebrafish proenkephalin reveals novel opioidsequences. Brain Res. Mol. Brain Res. 114, 31–39.
Hedges, S.B., Poling, L.L., 1999. A molecular phylogeny of reptiles.Science 283, 998–1001.
Iwabe, N., Hara, Y., Kumazawa, Y., Shibamoto, K., Saito, Y., Miyata,T., Katoh, K., 2004. Sister group relationship of turtles to the bird-crocodilian clade revealed by nuclear DNA-coded proteins. Mol. Biol.Evol. 22, 810–813.
Jakobsen, K.S., Haugen, M., Saeboe-Larsen, S., Hollung, K., Espell-und, M., Hornes, E., 1994. In: Hornes, M., Olvisk, O. (Eds.),Advances in Biomagnetic Separation. Eaton Publishing, Oslo, pp.61–71.
Khalap, A., Bagrosky, B., Lecaude, S., Youson, J., Danielson, P., Dores,R.M., 2005. Trends in the evolution of the proenkephalin andprodynorphin genes in gnathostomes. Trends Comp. Endocrinol.Neurobiol. 1040, 22–37.
Kieffer, B.L., Befort, K., Gaveriaux-Fuff, C., Hirth, C.G., 1992. The delta-opioid receptor: Isolation of a cDNA by expression cloning andpharmacological characterization. Proc. Natl. Acad. Sci. USA 89,12045–12052.
Kilpatrick, D.L., Toniguichi, T., Jones, B.N., Shirley, J.E., Hullihan, J.,Kimura, S., Udenfriend, S., 1981. A highly potent 3200 dalton adrenalopioid peptide that contains both a [Met] and [Leu]enkephalinsequence. Proc. Nat. Acad. Sci. USA 78, 3261–3268.
Kumazawa, Y., Nishida, S., 1999. Complete mitochondrial DNAsequences of the green turtle and blue-tailed mole skink: statistical

E. Roberts et al. / General and Comparative Endocrinology 153 (2007) 189–197 197
evidence for archosaurian affinity of turtles. Mol. Biol. Evol. 16, 784–792.
Lecaude, S., Alrubaian, J., Sollars, C., Propper, C., Danielson, P., Dores,R.M., 1999. Organization of proenkephalin in amphibians: cloning aproenkephalin cDNA from the brain of the anuran amphibian, Spea
multiplicatus. Peptides 21, 339–344.Mansour, A., Khaturian, H., Lewis, M.E., Akil, H., Watson, S.J., 1988.
Anatomy of CNS opioid receptors. Trends Neurosci. 11, 308–314.Martens, G.J., Herbert, E., 1984. Polymorphism and absence of Leu-
enkephalin sequences in proenkephalin genes in Xenopus laevis. Nature310, 251–254.
Neilson, B.G., 2003. The probability distribution of side-chain confor-mations in [Leu] and [Met] enkephalin determines the potency andselectivity to mu and delta opiate receptors. Biopolymers 71, 577–592.
Noda, M., Teranishi, Y., Takahashi, H., Toyosato, M., Notake, M.,Nakanishi, S., Numa, S., 1982. Isolation and structural organization ofthe human preproenkephalin gene. Nature 297, 431–434.
Sollars, C., Danielson, P., Joss, J.M.P., Dores, R.M., 2000. Decipheringthe origin of Met-enkephalin and Leu-enkephalin in lobe-finned fish:cloning of Australian lungfish proenkephalin. Brain Res. 874, 131–136.
Swofford, D.L., 1998. PAUP: Phylogenetic analysis using parsimony,version beta 4.0, Illinois Natural History Survey, Champaign, IL.
Walthers, E.A., Moore, F.L., 2005. Cloning proenkephalin from the brainof a urodele amphibian (Taricha granulosa) using a DOR-specificprimer in a 3 0RACE reaction. Gen. Comp. Endocrinol. 142, 364–370.
Wong, M., Rius, R.A., Loh, Y.P., 1991. Characterization of Xenopus
laevis proenkephalin gene. Brain Res. Mol. Brain Res. 11, 197–205.

























