Embed Size (px)
Citation preview

1
1
2
3
4
Running head Metabolic effects of stresses in field grown maize 5
6
7
8
Corresponding author 9
Toshihiro Obata 10
Max-Planck-Institut fuumlr Molekulare Pflanzenphysiologie 11
Am Muumlhlenberg 1 12
14476 Golm 13
Germany 14
Tel +49 (0)331 5678261 15
Fax +49 (0)331 5678408 16
17
18
19
20
Journal Research Article Area Biochemistry and Metabolism 21
Plant Physiology Preview Published on September 30 2015 as DOI101104pp1501164
Copyright 2015 by the American Society of Plant Biologists
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
2
Metabolite profiles of maize leaves in drought heat and combined 22
stress field trials reveal the relationship between metabolism and 23
grain yield1 24
25
Toshihiro Obata Sandra Witt Jan Lisec Natalia Palacios-Rojas Igor Florez-Sarasa 26
Salima Yousfi Jose Luis Araus Jill E Cairns Alisdair R Fernie 27
28 Max-Planck-Institut fuumlr Molekulare Pflanzenphysiologie Am Muumlhlenberg 1 14476 29 Potsdam-Golm Germany (TO SW JL IFS and ARF) CIMMYT Km 45 Carretera 30 Mexico-Veracruz Texcoco Mexico 56130 (NPR) Department de Biologia Vegetal 31 Universitat de Barcelona 08028 Barcelona Spain (SY and JLA) CIMMYT Southern 32 Africa Regional Office Harare Zimbabwe (JEC) 33 34 35 36 One sentence summary 37 38 Folial levels of metabolites including myo-inositol showed correlation with grain yield 39 under drought heat and simultaneous droughtheat stress field trials of tropical maize 40 genotypes 41 42
43
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
3
Footnotes 44 45 1This work was supported by funding from EU FP7 Coordination and Support Action 46 OPTICHINA (FP7-KBBE 26604 TO SW NPR JLA JEC and ARF) the EU integrative 47 project 3TO4 (FP7-KBBE 289582 TO and ARF) the Max Planck Society (TO and ARF) 48 and the Alexander von Humboldt Foundation (IFS) 49 50 51 52
53
54
Author to whom correspondence should be sent Email obatampimp-golmmpgde 55
56
57
The author(s) responsible for distribution of materials integral to the findings presented 58
in this article in accordance with the policy described in the Instructions for Authors 59
(wwwplantphysiolorg) is Toshihiro Obata (e-mail obatampimp-golmmpgde) 60
61
62
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
4
Abstract 63
The development of abiotic stress resistant cultivars is of premium importance for the 64
agriculture of developing countries Further progress in maize performance under 65
stresses is expected by combining marker assisted breeding with metabolite markers In 66
order to dissect metabolic responses and to identify promising metabolite marker 67
candidates metabolite profiles of maize leaves were analysed and compared with grain 68
yield in field trials Plants were grown under well water conditions (control) or exposed 69
to drought heat and both stresses simultaneously Trials were conducted in 2010 and 70
2011 using ten tropical hybrids selected to exhibit diverse abiotic stress tolerance 71
Drought stress invoked accumulation of many amino acids including isoleucine valine 72
threonine and GABA which has been commonly reported in both field and greenhouse 73
experiments in many plant species Two photorespiratory amino acids glycine and 74
serine and myo-inositol also accumulated under drought Combination of drought and 75
heat invoked relatively few specific responses and most of the metabolic changes were 76
predictable from the sum of the responses to individual stresses Statistical analysis 77
revealed significant correlation between levels of glycine and myo-inositol and grain 78
yield under drought Levels of myo-inositol in control conditions were also related to 79
grain yield under drought Furthermore multiple linear regression models very well 80
explained the variation of grain yield via the combination of several metabolites These 81
results indicate the importance of photorespiration and raffinose family oligosaccharide 82
metabolism in grain yield under drought and suggest single or multiple metabolites as 83
potential metabolic markers for breeding of abiotic stress tolerant maize 84
85
86
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
5
Introduction 87
The increasing world population coupled to environmental deterioration is creating ever 88
greater pressure on our capacity for sustainable food productivity Alongside biotic 89
stresses abiotic stresses such as drought heat salinity nutrient deficiency greatly 90
reduce yields in crop fields either when present alone or in combination Breeding for 91
more resilient crops is therefore one of the major approaches to cope with the 92
increasing challenges in world agriculture Considerable research effort has thus been 93
invested in order to dissect plant responses to individual stresses at various levels 94
(reviewed in Urano et al 2010 Lopes et al 2011 Obata and Fernie 2012) but the 95
interaction between different stresses has been far less investigated (Cairns et al 96
2012b Cairns et al 2013 Suzuki et al 2014) In general combination of stresses 97
additively affects plant physiology (ie the symptoms of the individual stresses appear 98
simultaneously) and synergistically diminishes yield and productivity of plants (Keleş 99
and Oumlncel 2002 Giraud et al 2008 Vile et al 2012 Suzuki et al 2014) The 100
molecular responses are however not simply additive and are rarely predicted from the 101
responses to individual stresses (Rizhsky et al 2002 Rizhsky et al 2004 Prasch and 102
Sonnewald 2013 Rasmussen et al 2013) Information from carefully controlled 103
greenhouse experiments has begun to dissect the molecular mechanisms by which 104
plants in particular Arabidopsis respond to drought and temperature stresses (Skirycz 105
et al 2010 Skirycz and Inzeacute 2010 Skirycz et al 2011 Bowne et al 2012 Tardieu 106
2012 Verkest et al 2015) Our knowledge of the molecular basis of the response of 107
crop species in a field environment is however considerably less well advanced (Araus 108
et al 2008 Cabrera-Bosquet et al 2012) That said a large number of genotypes 109
have been generated for on the basis of their resistance to both biotic and abiotic 110
stresses (for a review see Baumlnziger et al 2006 Takeda and Matsuoka 2008 Cooper 111
et al 2014) and the genome sequencing and molecular characterization of a range of 112
stress tolerant plant species have recently been reported (Wu et al 2012 Ma et al 113
2013 Bolger et al 2014 Tohge et al 2014) These studies are not only important as 114
basic research for further studies in crops but also are a prerequisite in the development 115
of molecular marker based approaches to improve crop tolerance to stress 116
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
6
As a first step toward this goal a deeper understanding of the plant response to the 117
stressful environment especially those to multiple stress conditions under field 118
conditions is crucial for improvement of stress-tolerant crops This is important on two 119
levels (i) in the field singular abiotic stresses are rare and (ii) yield and stress adaptation 120
are complex traits which render breeding gains slower than would be expected under 121
optimal conditions (Bruce et al 2002) Recent studies have revealed that the response 122
of plants to combinations of two or more stress conditions is unique and cannot be 123
directly extrapolated from their response to the different stresses when applied 124
individually This would be a result of complex combinations of different and sometimes 125
opposing responses in signalling pathways including those may interact and inhibit one 126
another (Prasch and Sonnewald 2013 Rasmussen et al 2013 Suzuki et al 2014) 127
Maize is grown in over 170 million hectares worldwide of which 130 million hectares are 128
in less-developed countries (FAO 2014) In sub-Saharan Africa (SSA) maize is a staple 129
crop however yields in this region have stagnated at less than 2 t ha-1 while maize 130
yields worldwide have continued to increase (Cairns et al 2012a) Low yields in SSA 131
are largely associated with drought stress and low soil fertility (Baumlnziger and Araus 132
2007) Additionally simulation studies indicate that maize yield in Africa is likely to be 133
significantly impaired by heat stress (Lobell and Burke 2010 Lobell et al 2011) such 134
as that can be anticipated as a result of the changes in climate predicted for the coming 135
decades (Muumlller et al 2011) Moreover sensitivity of maize yield to heat is exacerbated 136
under drought conditions (Lobell et al 2011 Cairns et al 2012ab Cairns et al 2013) 137
Therefore the development of maize germplasm tolerant to drought and heat conditions 138
is of utmost importance to both increase yields and offset predicted yield losses under 139
projected climate change scenarios (Easterling et al 2007) especially in SSA While 140
direct selection for grain yield under drought stress has resulted in admirable gains in 141
grain yield under stress (Baumlnziger et al 2006 Cairns et al 2013) further improvement 142
requires incorporation of additional selection traits (Cairns et al 2012a Cairns et al 143
2012b) In recent years genetic and phenotypic markers have been extensively 144
searched for drought tolerance of maize by high throughput genomic and phenotyping 145
approaches respectively (Tuberosa and Salvi 2006 Wen et al 2011 Araus et al 146
2012 Cairns et al 2013 Prasanna et al 2013 Araus and Cairns 2014 Tsonev et al 147
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
7
2014) Moreover metabolic markers start to draw attention due to their close 148
relationship with yield phenotypes (Fernie and Schauer 2009 Redestig et al 2011 149
Riedelsheimer et al 2012a Witt et al 2012 Riedelsheimer et al 2012b Degenkolbe 150
et al 2013) Accumulation of some metabolites has been reported to be directly related 151
to the performance of potato cultivars in beetle resistance in the field (Tai et al 2014) 152
Additionally identical genomic regions were mapped as both agronomic and metabolic 153
quantitative trait loci (QTL) in field grown maize and wheat indicating the utility of 154
metabolic traits for breeding selection (Riedelsheimer et al 2012b Hill et al 2015) A 155
recent study showed genetic gains in maize grain yield under drought stress were 156
higher using a molecular marker based approach than conventional breeding (Beyene 157
et al 2015) 158
Here we focused on the relationship between leaf metabolites and grain yield under 159
drought heat and simultaneous drought and heat conditions in the field The negative 160
effect of drought stress on maize yield is especially acute during the reproductive stage 161
between tassel emergence and early grain-filling (Grant et al 1989) where it is 162
believed to induce premature seed desiccation and to limit grain filling Grain is more 163
susceptible to drought stress than vegetative tissues and therefore the prediction of 164
grain yield from physiological parameter of leaves is a challenge (Sangoi and Salvador 165
1998 Khodarahmpour and Hamidi 2011) Nevertheless maize yield is dependent on 166
both the assimilate supply to the kernel (source) and the potential of kernel to 167
accommodate this assimilate (sink potential Jones and Simmons 1983) Breeding for 168
modern temperate hybrids has focused more on the sink potential particularly under 169
stress conditions (Tollenaar and Lee 2006) and there should therefore be 170
considerable potential remaining to improve source ability Drought and heat stresses 171
would be anticipated largely to affect leaf metabolism and especially photosynthesis 172
compromising the source capacity of leaves (Chaves et al 2009 Lawlor and Tezara 173
2009 Osakabe et al 2014) In keeping with this drought was found to have the most 174
dramatic effect on the metabolite composition in leaves compare to other organs in our 175
previous greenhouse experiments (Witt et al 2012) Since the source ability is closely 176
related to leaf metabolism the leaf metabolite profile should have close relationship to 177
grain yield particularly under conditions of stress Given that several recent studies have 178
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

2
Metabolite profiles of maize leaves in drought heat and combined 22
stress field trials reveal the relationship between metabolism and 23
grain yield1 24
25
Toshihiro Obata Sandra Witt Jan Lisec Natalia Palacios-Rojas Igor Florez-Sarasa 26
Salima Yousfi Jose Luis Araus Jill E Cairns Alisdair R Fernie 27
28 Max-Planck-Institut fuumlr Molekulare Pflanzenphysiologie Am Muumlhlenberg 1 14476 29 Potsdam-Golm Germany (TO SW JL IFS and ARF) CIMMYT Km 45 Carretera 30 Mexico-Veracruz Texcoco Mexico 56130 (NPR) Department de Biologia Vegetal 31 Universitat de Barcelona 08028 Barcelona Spain (SY and JLA) CIMMYT Southern 32 Africa Regional Office Harare Zimbabwe (JEC) 33 34 35 36 One sentence summary 37 38 Folial levels of metabolites including myo-inositol showed correlation with grain yield 39 under drought heat and simultaneous droughtheat stress field trials of tropical maize 40 genotypes 41 42
43
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
3
Footnotes 44 45 1This work was supported by funding from EU FP7 Coordination and Support Action 46 OPTICHINA (FP7-KBBE 26604 TO SW NPR JLA JEC and ARF) the EU integrative 47 project 3TO4 (FP7-KBBE 289582 TO and ARF) the Max Planck Society (TO and ARF) 48 and the Alexander von Humboldt Foundation (IFS) 49 50 51 52
53
54
Author to whom correspondence should be sent Email obatampimp-golmmpgde 55
56
57
The author(s) responsible for distribution of materials integral to the findings presented 58
in this article in accordance with the policy described in the Instructions for Authors 59
(wwwplantphysiolorg) is Toshihiro Obata (e-mail obatampimp-golmmpgde) 60
61
62
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
4
Abstract 63
The development of abiotic stress resistant cultivars is of premium importance for the 64
agriculture of developing countries Further progress in maize performance under 65
stresses is expected by combining marker assisted breeding with metabolite markers In 66
order to dissect metabolic responses and to identify promising metabolite marker 67
candidates metabolite profiles of maize leaves were analysed and compared with grain 68
yield in field trials Plants were grown under well water conditions (control) or exposed 69
to drought heat and both stresses simultaneously Trials were conducted in 2010 and 70
2011 using ten tropical hybrids selected to exhibit diverse abiotic stress tolerance 71
Drought stress invoked accumulation of many amino acids including isoleucine valine 72
threonine and GABA which has been commonly reported in both field and greenhouse 73
experiments in many plant species Two photorespiratory amino acids glycine and 74
serine and myo-inositol also accumulated under drought Combination of drought and 75
heat invoked relatively few specific responses and most of the metabolic changes were 76
predictable from the sum of the responses to individual stresses Statistical analysis 77
revealed significant correlation between levels of glycine and myo-inositol and grain 78
yield under drought Levels of myo-inositol in control conditions were also related to 79
grain yield under drought Furthermore multiple linear regression models very well 80
explained the variation of grain yield via the combination of several metabolites These 81
results indicate the importance of photorespiration and raffinose family oligosaccharide 82
metabolism in grain yield under drought and suggest single or multiple metabolites as 83
potential metabolic markers for breeding of abiotic stress tolerant maize 84
85
86
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
5
Introduction 87
The increasing world population coupled to environmental deterioration is creating ever 88
greater pressure on our capacity for sustainable food productivity Alongside biotic 89
stresses abiotic stresses such as drought heat salinity nutrient deficiency greatly 90
reduce yields in crop fields either when present alone or in combination Breeding for 91
more resilient crops is therefore one of the major approaches to cope with the 92
increasing challenges in world agriculture Considerable research effort has thus been 93
invested in order to dissect plant responses to individual stresses at various levels 94
(reviewed in Urano et al 2010 Lopes et al 2011 Obata and Fernie 2012) but the 95
interaction between different stresses has been far less investigated (Cairns et al 96
2012b Cairns et al 2013 Suzuki et al 2014) In general combination of stresses 97
additively affects plant physiology (ie the symptoms of the individual stresses appear 98
simultaneously) and synergistically diminishes yield and productivity of plants (Keleş 99
and Oumlncel 2002 Giraud et al 2008 Vile et al 2012 Suzuki et al 2014) The 100
molecular responses are however not simply additive and are rarely predicted from the 101
responses to individual stresses (Rizhsky et al 2002 Rizhsky et al 2004 Prasch and 102
Sonnewald 2013 Rasmussen et al 2013) Information from carefully controlled 103
greenhouse experiments has begun to dissect the molecular mechanisms by which 104
plants in particular Arabidopsis respond to drought and temperature stresses (Skirycz 105
et al 2010 Skirycz and Inzeacute 2010 Skirycz et al 2011 Bowne et al 2012 Tardieu 106
2012 Verkest et al 2015) Our knowledge of the molecular basis of the response of 107
crop species in a field environment is however considerably less well advanced (Araus 108
et al 2008 Cabrera-Bosquet et al 2012) That said a large number of genotypes 109
have been generated for on the basis of their resistance to both biotic and abiotic 110
stresses (for a review see Baumlnziger et al 2006 Takeda and Matsuoka 2008 Cooper 111
et al 2014) and the genome sequencing and molecular characterization of a range of 112
stress tolerant plant species have recently been reported (Wu et al 2012 Ma et al 113
2013 Bolger et al 2014 Tohge et al 2014) These studies are not only important as 114
basic research for further studies in crops but also are a prerequisite in the development 115
of molecular marker based approaches to improve crop tolerance to stress 116
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
6
As a first step toward this goal a deeper understanding of the plant response to the 117
stressful environment especially those to multiple stress conditions under field 118
conditions is crucial for improvement of stress-tolerant crops This is important on two 119
levels (i) in the field singular abiotic stresses are rare and (ii) yield and stress adaptation 120
are complex traits which render breeding gains slower than would be expected under 121
optimal conditions (Bruce et al 2002) Recent studies have revealed that the response 122
of plants to combinations of two or more stress conditions is unique and cannot be 123
directly extrapolated from their response to the different stresses when applied 124
individually This would be a result of complex combinations of different and sometimes 125
opposing responses in signalling pathways including those may interact and inhibit one 126
another (Prasch and Sonnewald 2013 Rasmussen et al 2013 Suzuki et al 2014) 127
Maize is grown in over 170 million hectares worldwide of which 130 million hectares are 128
in less-developed countries (FAO 2014) In sub-Saharan Africa (SSA) maize is a staple 129
crop however yields in this region have stagnated at less than 2 t ha-1 while maize 130
yields worldwide have continued to increase (Cairns et al 2012a) Low yields in SSA 131
are largely associated with drought stress and low soil fertility (Baumlnziger and Araus 132
2007) Additionally simulation studies indicate that maize yield in Africa is likely to be 133
significantly impaired by heat stress (Lobell and Burke 2010 Lobell et al 2011) such 134
as that can be anticipated as a result of the changes in climate predicted for the coming 135
decades (Muumlller et al 2011) Moreover sensitivity of maize yield to heat is exacerbated 136
under drought conditions (Lobell et al 2011 Cairns et al 2012ab Cairns et al 2013) 137
Therefore the development of maize germplasm tolerant to drought and heat conditions 138
is of utmost importance to both increase yields and offset predicted yield losses under 139
projected climate change scenarios (Easterling et al 2007) especially in SSA While 140
direct selection for grain yield under drought stress has resulted in admirable gains in 141
grain yield under stress (Baumlnziger et al 2006 Cairns et al 2013) further improvement 142
requires incorporation of additional selection traits (Cairns et al 2012a Cairns et al 143
2012b) In recent years genetic and phenotypic markers have been extensively 144
searched for drought tolerance of maize by high throughput genomic and phenotyping 145
approaches respectively (Tuberosa and Salvi 2006 Wen et al 2011 Araus et al 146
2012 Cairns et al 2013 Prasanna et al 2013 Araus and Cairns 2014 Tsonev et al 147
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
7
2014) Moreover metabolic markers start to draw attention due to their close 148
relationship with yield phenotypes (Fernie and Schauer 2009 Redestig et al 2011 149
Riedelsheimer et al 2012a Witt et al 2012 Riedelsheimer et al 2012b Degenkolbe 150
et al 2013) Accumulation of some metabolites has been reported to be directly related 151
to the performance of potato cultivars in beetle resistance in the field (Tai et al 2014) 152
Additionally identical genomic regions were mapped as both agronomic and metabolic 153
quantitative trait loci (QTL) in field grown maize and wheat indicating the utility of 154
metabolic traits for breeding selection (Riedelsheimer et al 2012b Hill et al 2015) A 155
recent study showed genetic gains in maize grain yield under drought stress were 156
higher using a molecular marker based approach than conventional breeding (Beyene 157
et al 2015) 158
Here we focused on the relationship between leaf metabolites and grain yield under 159
drought heat and simultaneous drought and heat conditions in the field The negative 160
effect of drought stress on maize yield is especially acute during the reproductive stage 161
between tassel emergence and early grain-filling (Grant et al 1989) where it is 162
believed to induce premature seed desiccation and to limit grain filling Grain is more 163
susceptible to drought stress than vegetative tissues and therefore the prediction of 164
grain yield from physiological parameter of leaves is a challenge (Sangoi and Salvador 165
1998 Khodarahmpour and Hamidi 2011) Nevertheless maize yield is dependent on 166
both the assimilate supply to the kernel (source) and the potential of kernel to 167
accommodate this assimilate (sink potential Jones and Simmons 1983) Breeding for 168
modern temperate hybrids has focused more on the sink potential particularly under 169
stress conditions (Tollenaar and Lee 2006) and there should therefore be 170
considerable potential remaining to improve source ability Drought and heat stresses 171
would be anticipated largely to affect leaf metabolism and especially photosynthesis 172
compromising the source capacity of leaves (Chaves et al 2009 Lawlor and Tezara 173
2009 Osakabe et al 2014) In keeping with this drought was found to have the most 174
dramatic effect on the metabolite composition in leaves compare to other organs in our 175
previous greenhouse experiments (Witt et al 2012) Since the source ability is closely 176
related to leaf metabolism the leaf metabolite profile should have close relationship to 177
grain yield particularly under conditions of stress Given that several recent studies have 178
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

3
Footnotes 44 45 1This work was supported by funding from EU FP7 Coordination and Support Action 46 OPTICHINA (FP7-KBBE 26604 TO SW NPR JLA JEC and ARF) the EU integrative 47 project 3TO4 (FP7-KBBE 289582 TO and ARF) the Max Planck Society (TO and ARF) 48 and the Alexander von Humboldt Foundation (IFS) 49 50 51 52
53
54
Author to whom correspondence should be sent Email obatampimp-golmmpgde 55
56
57
The author(s) responsible for distribution of materials integral to the findings presented 58
in this article in accordance with the policy described in the Instructions for Authors 59
(wwwplantphysiolorg) is Toshihiro Obata (e-mail obatampimp-golmmpgde) 60
61
62
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
4
Abstract 63
The development of abiotic stress resistant cultivars is of premium importance for the 64
agriculture of developing countries Further progress in maize performance under 65
stresses is expected by combining marker assisted breeding with metabolite markers In 66
order to dissect metabolic responses and to identify promising metabolite marker 67
candidates metabolite profiles of maize leaves were analysed and compared with grain 68
yield in field trials Plants were grown under well water conditions (control) or exposed 69
to drought heat and both stresses simultaneously Trials were conducted in 2010 and 70
2011 using ten tropical hybrids selected to exhibit diverse abiotic stress tolerance 71
Drought stress invoked accumulation of many amino acids including isoleucine valine 72
threonine and GABA which has been commonly reported in both field and greenhouse 73
experiments in many plant species Two photorespiratory amino acids glycine and 74
serine and myo-inositol also accumulated under drought Combination of drought and 75
heat invoked relatively few specific responses and most of the metabolic changes were 76
predictable from the sum of the responses to individual stresses Statistical analysis 77
revealed significant correlation between levels of glycine and myo-inositol and grain 78
yield under drought Levels of myo-inositol in control conditions were also related to 79
grain yield under drought Furthermore multiple linear regression models very well 80
explained the variation of grain yield via the combination of several metabolites These 81
results indicate the importance of photorespiration and raffinose family oligosaccharide 82
metabolism in grain yield under drought and suggest single or multiple metabolites as 83
potential metabolic markers for breeding of abiotic stress tolerant maize 84
85
86
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
5
Introduction 87
The increasing world population coupled to environmental deterioration is creating ever 88
greater pressure on our capacity for sustainable food productivity Alongside biotic 89
stresses abiotic stresses such as drought heat salinity nutrient deficiency greatly 90
reduce yields in crop fields either when present alone or in combination Breeding for 91
more resilient crops is therefore one of the major approaches to cope with the 92
increasing challenges in world agriculture Considerable research effort has thus been 93
invested in order to dissect plant responses to individual stresses at various levels 94
(reviewed in Urano et al 2010 Lopes et al 2011 Obata and Fernie 2012) but the 95
interaction between different stresses has been far less investigated (Cairns et al 96
2012b Cairns et al 2013 Suzuki et al 2014) In general combination of stresses 97
additively affects plant physiology (ie the symptoms of the individual stresses appear 98
simultaneously) and synergistically diminishes yield and productivity of plants (Keleş 99
and Oumlncel 2002 Giraud et al 2008 Vile et al 2012 Suzuki et al 2014) The 100
molecular responses are however not simply additive and are rarely predicted from the 101
responses to individual stresses (Rizhsky et al 2002 Rizhsky et al 2004 Prasch and 102
Sonnewald 2013 Rasmussen et al 2013) Information from carefully controlled 103
greenhouse experiments has begun to dissect the molecular mechanisms by which 104
plants in particular Arabidopsis respond to drought and temperature stresses (Skirycz 105
et al 2010 Skirycz and Inzeacute 2010 Skirycz et al 2011 Bowne et al 2012 Tardieu 106
2012 Verkest et al 2015) Our knowledge of the molecular basis of the response of 107
crop species in a field environment is however considerably less well advanced (Araus 108
et al 2008 Cabrera-Bosquet et al 2012) That said a large number of genotypes 109
have been generated for on the basis of their resistance to both biotic and abiotic 110
stresses (for a review see Baumlnziger et al 2006 Takeda and Matsuoka 2008 Cooper 111
et al 2014) and the genome sequencing and molecular characterization of a range of 112
stress tolerant plant species have recently been reported (Wu et al 2012 Ma et al 113
2013 Bolger et al 2014 Tohge et al 2014) These studies are not only important as 114
basic research for further studies in crops but also are a prerequisite in the development 115
of molecular marker based approaches to improve crop tolerance to stress 116
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
6
As a first step toward this goal a deeper understanding of the plant response to the 117
stressful environment especially those to multiple stress conditions under field 118
conditions is crucial for improvement of stress-tolerant crops This is important on two 119
levels (i) in the field singular abiotic stresses are rare and (ii) yield and stress adaptation 120
are complex traits which render breeding gains slower than would be expected under 121
optimal conditions (Bruce et al 2002) Recent studies have revealed that the response 122
of plants to combinations of two or more stress conditions is unique and cannot be 123
directly extrapolated from their response to the different stresses when applied 124
individually This would be a result of complex combinations of different and sometimes 125
opposing responses in signalling pathways including those may interact and inhibit one 126
another (Prasch and Sonnewald 2013 Rasmussen et al 2013 Suzuki et al 2014) 127
Maize is grown in over 170 million hectares worldwide of which 130 million hectares are 128
in less-developed countries (FAO 2014) In sub-Saharan Africa (SSA) maize is a staple 129
crop however yields in this region have stagnated at less than 2 t ha-1 while maize 130
yields worldwide have continued to increase (Cairns et al 2012a) Low yields in SSA 131
are largely associated with drought stress and low soil fertility (Baumlnziger and Araus 132
2007) Additionally simulation studies indicate that maize yield in Africa is likely to be 133
significantly impaired by heat stress (Lobell and Burke 2010 Lobell et al 2011) such 134
as that can be anticipated as a result of the changes in climate predicted for the coming 135
decades (Muumlller et al 2011) Moreover sensitivity of maize yield to heat is exacerbated 136
under drought conditions (Lobell et al 2011 Cairns et al 2012ab Cairns et al 2013) 137
Therefore the development of maize germplasm tolerant to drought and heat conditions 138
is of utmost importance to both increase yields and offset predicted yield losses under 139
projected climate change scenarios (Easterling et al 2007) especially in SSA While 140
direct selection for grain yield under drought stress has resulted in admirable gains in 141
grain yield under stress (Baumlnziger et al 2006 Cairns et al 2013) further improvement 142
requires incorporation of additional selection traits (Cairns et al 2012a Cairns et al 143
2012b) In recent years genetic and phenotypic markers have been extensively 144
searched for drought tolerance of maize by high throughput genomic and phenotyping 145
approaches respectively (Tuberosa and Salvi 2006 Wen et al 2011 Araus et al 146
2012 Cairns et al 2013 Prasanna et al 2013 Araus and Cairns 2014 Tsonev et al 147
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
7
2014) Moreover metabolic markers start to draw attention due to their close 148
relationship with yield phenotypes (Fernie and Schauer 2009 Redestig et al 2011 149
Riedelsheimer et al 2012a Witt et al 2012 Riedelsheimer et al 2012b Degenkolbe 150
et al 2013) Accumulation of some metabolites has been reported to be directly related 151
to the performance of potato cultivars in beetle resistance in the field (Tai et al 2014) 152
Additionally identical genomic regions were mapped as both agronomic and metabolic 153
quantitative trait loci (QTL) in field grown maize and wheat indicating the utility of 154
metabolic traits for breeding selection (Riedelsheimer et al 2012b Hill et al 2015) A 155
recent study showed genetic gains in maize grain yield under drought stress were 156
higher using a molecular marker based approach than conventional breeding (Beyene 157
et al 2015) 158
Here we focused on the relationship between leaf metabolites and grain yield under 159
drought heat and simultaneous drought and heat conditions in the field The negative 160
effect of drought stress on maize yield is especially acute during the reproductive stage 161
between tassel emergence and early grain-filling (Grant et al 1989) where it is 162
believed to induce premature seed desiccation and to limit grain filling Grain is more 163
susceptible to drought stress than vegetative tissues and therefore the prediction of 164
grain yield from physiological parameter of leaves is a challenge (Sangoi and Salvador 165
1998 Khodarahmpour and Hamidi 2011) Nevertheless maize yield is dependent on 166
both the assimilate supply to the kernel (source) and the potential of kernel to 167
accommodate this assimilate (sink potential Jones and Simmons 1983) Breeding for 168
modern temperate hybrids has focused more on the sink potential particularly under 169
stress conditions (Tollenaar and Lee 2006) and there should therefore be 170
considerable potential remaining to improve source ability Drought and heat stresses 171
would be anticipated largely to affect leaf metabolism and especially photosynthesis 172
compromising the source capacity of leaves (Chaves et al 2009 Lawlor and Tezara 173
2009 Osakabe et al 2014) In keeping with this drought was found to have the most 174
dramatic effect on the metabolite composition in leaves compare to other organs in our 175
previous greenhouse experiments (Witt et al 2012) Since the source ability is closely 176
related to leaf metabolism the leaf metabolite profile should have close relationship to 177
grain yield particularly under conditions of stress Given that several recent studies have 178
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

4
Abstract 63
The development of abiotic stress resistant cultivars is of premium importance for the 64
agriculture of developing countries Further progress in maize performance under 65
stresses is expected by combining marker assisted breeding with metabolite markers In 66
order to dissect metabolic responses and to identify promising metabolite marker 67
candidates metabolite profiles of maize leaves were analysed and compared with grain 68
yield in field trials Plants were grown under well water conditions (control) or exposed 69
to drought heat and both stresses simultaneously Trials were conducted in 2010 and 70
2011 using ten tropical hybrids selected to exhibit diverse abiotic stress tolerance 71
Drought stress invoked accumulation of many amino acids including isoleucine valine 72
threonine and GABA which has been commonly reported in both field and greenhouse 73
experiments in many plant species Two photorespiratory amino acids glycine and 74
serine and myo-inositol also accumulated under drought Combination of drought and 75
heat invoked relatively few specific responses and most of the metabolic changes were 76
predictable from the sum of the responses to individual stresses Statistical analysis 77
revealed significant correlation between levels of glycine and myo-inositol and grain 78
yield under drought Levels of myo-inositol in control conditions were also related to 79
grain yield under drought Furthermore multiple linear regression models very well 80
explained the variation of grain yield via the combination of several metabolites These 81
results indicate the importance of photorespiration and raffinose family oligosaccharide 82
metabolism in grain yield under drought and suggest single or multiple metabolites as 83
potential metabolic markers for breeding of abiotic stress tolerant maize 84
85
86
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
5
Introduction 87
The increasing world population coupled to environmental deterioration is creating ever 88
greater pressure on our capacity for sustainable food productivity Alongside biotic 89
stresses abiotic stresses such as drought heat salinity nutrient deficiency greatly 90
reduce yields in crop fields either when present alone or in combination Breeding for 91
more resilient crops is therefore one of the major approaches to cope with the 92
increasing challenges in world agriculture Considerable research effort has thus been 93
invested in order to dissect plant responses to individual stresses at various levels 94
(reviewed in Urano et al 2010 Lopes et al 2011 Obata and Fernie 2012) but the 95
interaction between different stresses has been far less investigated (Cairns et al 96
2012b Cairns et al 2013 Suzuki et al 2014) In general combination of stresses 97
additively affects plant physiology (ie the symptoms of the individual stresses appear 98
simultaneously) and synergistically diminishes yield and productivity of plants (Keleş 99
and Oumlncel 2002 Giraud et al 2008 Vile et al 2012 Suzuki et al 2014) The 100
molecular responses are however not simply additive and are rarely predicted from the 101
responses to individual stresses (Rizhsky et al 2002 Rizhsky et al 2004 Prasch and 102
Sonnewald 2013 Rasmussen et al 2013) Information from carefully controlled 103
greenhouse experiments has begun to dissect the molecular mechanisms by which 104
plants in particular Arabidopsis respond to drought and temperature stresses (Skirycz 105
et al 2010 Skirycz and Inzeacute 2010 Skirycz et al 2011 Bowne et al 2012 Tardieu 106
2012 Verkest et al 2015) Our knowledge of the molecular basis of the response of 107
crop species in a field environment is however considerably less well advanced (Araus 108
et al 2008 Cabrera-Bosquet et al 2012) That said a large number of genotypes 109
have been generated for on the basis of their resistance to both biotic and abiotic 110
stresses (for a review see Baumlnziger et al 2006 Takeda and Matsuoka 2008 Cooper 111
et al 2014) and the genome sequencing and molecular characterization of a range of 112
stress tolerant plant species have recently been reported (Wu et al 2012 Ma et al 113
2013 Bolger et al 2014 Tohge et al 2014) These studies are not only important as 114
basic research for further studies in crops but also are a prerequisite in the development 115
of molecular marker based approaches to improve crop tolerance to stress 116
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
6
As a first step toward this goal a deeper understanding of the plant response to the 117
stressful environment especially those to multiple stress conditions under field 118
conditions is crucial for improvement of stress-tolerant crops This is important on two 119
levels (i) in the field singular abiotic stresses are rare and (ii) yield and stress adaptation 120
are complex traits which render breeding gains slower than would be expected under 121
optimal conditions (Bruce et al 2002) Recent studies have revealed that the response 122
of plants to combinations of two or more stress conditions is unique and cannot be 123
directly extrapolated from their response to the different stresses when applied 124
individually This would be a result of complex combinations of different and sometimes 125
opposing responses in signalling pathways including those may interact and inhibit one 126
another (Prasch and Sonnewald 2013 Rasmussen et al 2013 Suzuki et al 2014) 127
Maize is grown in over 170 million hectares worldwide of which 130 million hectares are 128
in less-developed countries (FAO 2014) In sub-Saharan Africa (SSA) maize is a staple 129
crop however yields in this region have stagnated at less than 2 t ha-1 while maize 130
yields worldwide have continued to increase (Cairns et al 2012a) Low yields in SSA 131
are largely associated with drought stress and low soil fertility (Baumlnziger and Araus 132
2007) Additionally simulation studies indicate that maize yield in Africa is likely to be 133
significantly impaired by heat stress (Lobell and Burke 2010 Lobell et al 2011) such 134
as that can be anticipated as a result of the changes in climate predicted for the coming 135
decades (Muumlller et al 2011) Moreover sensitivity of maize yield to heat is exacerbated 136
under drought conditions (Lobell et al 2011 Cairns et al 2012ab Cairns et al 2013) 137
Therefore the development of maize germplasm tolerant to drought and heat conditions 138
is of utmost importance to both increase yields and offset predicted yield losses under 139
projected climate change scenarios (Easterling et al 2007) especially in SSA While 140
direct selection for grain yield under drought stress has resulted in admirable gains in 141
grain yield under stress (Baumlnziger et al 2006 Cairns et al 2013) further improvement 142
requires incorporation of additional selection traits (Cairns et al 2012a Cairns et al 143
2012b) In recent years genetic and phenotypic markers have been extensively 144
searched for drought tolerance of maize by high throughput genomic and phenotyping 145
approaches respectively (Tuberosa and Salvi 2006 Wen et al 2011 Araus et al 146
2012 Cairns et al 2013 Prasanna et al 2013 Araus and Cairns 2014 Tsonev et al 147
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
7
2014) Moreover metabolic markers start to draw attention due to their close 148
relationship with yield phenotypes (Fernie and Schauer 2009 Redestig et al 2011 149
Riedelsheimer et al 2012a Witt et al 2012 Riedelsheimer et al 2012b Degenkolbe 150
et al 2013) Accumulation of some metabolites has been reported to be directly related 151
to the performance of potato cultivars in beetle resistance in the field (Tai et al 2014) 152
Additionally identical genomic regions were mapped as both agronomic and metabolic 153
quantitative trait loci (QTL) in field grown maize and wheat indicating the utility of 154
metabolic traits for breeding selection (Riedelsheimer et al 2012b Hill et al 2015) A 155
recent study showed genetic gains in maize grain yield under drought stress were 156
higher using a molecular marker based approach than conventional breeding (Beyene 157
et al 2015) 158
Here we focused on the relationship between leaf metabolites and grain yield under 159
drought heat and simultaneous drought and heat conditions in the field The negative 160
effect of drought stress on maize yield is especially acute during the reproductive stage 161
between tassel emergence and early grain-filling (Grant et al 1989) where it is 162
believed to induce premature seed desiccation and to limit grain filling Grain is more 163
susceptible to drought stress than vegetative tissues and therefore the prediction of 164
grain yield from physiological parameter of leaves is a challenge (Sangoi and Salvador 165
1998 Khodarahmpour and Hamidi 2011) Nevertheless maize yield is dependent on 166
both the assimilate supply to the kernel (source) and the potential of kernel to 167
accommodate this assimilate (sink potential Jones and Simmons 1983) Breeding for 168
modern temperate hybrids has focused more on the sink potential particularly under 169
stress conditions (Tollenaar and Lee 2006) and there should therefore be 170
considerable potential remaining to improve source ability Drought and heat stresses 171
would be anticipated largely to affect leaf metabolism and especially photosynthesis 172
compromising the source capacity of leaves (Chaves et al 2009 Lawlor and Tezara 173
2009 Osakabe et al 2014) In keeping with this drought was found to have the most 174
dramatic effect on the metabolite composition in leaves compare to other organs in our 175
previous greenhouse experiments (Witt et al 2012) Since the source ability is closely 176
related to leaf metabolism the leaf metabolite profile should have close relationship to 177
grain yield particularly under conditions of stress Given that several recent studies have 178
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

5
Introduction 87
The increasing world population coupled to environmental deterioration is creating ever 88
greater pressure on our capacity for sustainable food productivity Alongside biotic 89
stresses abiotic stresses such as drought heat salinity nutrient deficiency greatly 90
reduce yields in crop fields either when present alone or in combination Breeding for 91
more resilient crops is therefore one of the major approaches to cope with the 92
increasing challenges in world agriculture Considerable research effort has thus been 93
invested in order to dissect plant responses to individual stresses at various levels 94
(reviewed in Urano et al 2010 Lopes et al 2011 Obata and Fernie 2012) but the 95
interaction between different stresses has been far less investigated (Cairns et al 96
2012b Cairns et al 2013 Suzuki et al 2014) In general combination of stresses 97
additively affects plant physiology (ie the symptoms of the individual stresses appear 98
simultaneously) and synergistically diminishes yield and productivity of plants (Keleş 99
and Oumlncel 2002 Giraud et al 2008 Vile et al 2012 Suzuki et al 2014) The 100
molecular responses are however not simply additive and are rarely predicted from the 101
responses to individual stresses (Rizhsky et al 2002 Rizhsky et al 2004 Prasch and 102
Sonnewald 2013 Rasmussen et al 2013) Information from carefully controlled 103
greenhouse experiments has begun to dissect the molecular mechanisms by which 104
plants in particular Arabidopsis respond to drought and temperature stresses (Skirycz 105
et al 2010 Skirycz and Inzeacute 2010 Skirycz et al 2011 Bowne et al 2012 Tardieu 106
2012 Verkest et al 2015) Our knowledge of the molecular basis of the response of 107
crop species in a field environment is however considerably less well advanced (Araus 108
et al 2008 Cabrera-Bosquet et al 2012) That said a large number of genotypes 109
have been generated for on the basis of their resistance to both biotic and abiotic 110
stresses (for a review see Baumlnziger et al 2006 Takeda and Matsuoka 2008 Cooper 111
et al 2014) and the genome sequencing and molecular characterization of a range of 112
stress tolerant plant species have recently been reported (Wu et al 2012 Ma et al 113
2013 Bolger et al 2014 Tohge et al 2014) These studies are not only important as 114
basic research for further studies in crops but also are a prerequisite in the development 115
of molecular marker based approaches to improve crop tolerance to stress 116
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
6
As a first step toward this goal a deeper understanding of the plant response to the 117
stressful environment especially those to multiple stress conditions under field 118
conditions is crucial for improvement of stress-tolerant crops This is important on two 119
levels (i) in the field singular abiotic stresses are rare and (ii) yield and stress adaptation 120
are complex traits which render breeding gains slower than would be expected under 121
optimal conditions (Bruce et al 2002) Recent studies have revealed that the response 122
of plants to combinations of two or more stress conditions is unique and cannot be 123
directly extrapolated from their response to the different stresses when applied 124
individually This would be a result of complex combinations of different and sometimes 125
opposing responses in signalling pathways including those may interact and inhibit one 126
another (Prasch and Sonnewald 2013 Rasmussen et al 2013 Suzuki et al 2014) 127
Maize is grown in over 170 million hectares worldwide of which 130 million hectares are 128
in less-developed countries (FAO 2014) In sub-Saharan Africa (SSA) maize is a staple 129
crop however yields in this region have stagnated at less than 2 t ha-1 while maize 130
yields worldwide have continued to increase (Cairns et al 2012a) Low yields in SSA 131
are largely associated with drought stress and low soil fertility (Baumlnziger and Araus 132
2007) Additionally simulation studies indicate that maize yield in Africa is likely to be 133
significantly impaired by heat stress (Lobell and Burke 2010 Lobell et al 2011) such 134
as that can be anticipated as a result of the changes in climate predicted for the coming 135
decades (Muumlller et al 2011) Moreover sensitivity of maize yield to heat is exacerbated 136
under drought conditions (Lobell et al 2011 Cairns et al 2012ab Cairns et al 2013) 137
Therefore the development of maize germplasm tolerant to drought and heat conditions 138
is of utmost importance to both increase yields and offset predicted yield losses under 139
projected climate change scenarios (Easterling et al 2007) especially in SSA While 140
direct selection for grain yield under drought stress has resulted in admirable gains in 141
grain yield under stress (Baumlnziger et al 2006 Cairns et al 2013) further improvement 142
requires incorporation of additional selection traits (Cairns et al 2012a Cairns et al 143
2012b) In recent years genetic and phenotypic markers have been extensively 144
searched for drought tolerance of maize by high throughput genomic and phenotyping 145
approaches respectively (Tuberosa and Salvi 2006 Wen et al 2011 Araus et al 146
2012 Cairns et al 2013 Prasanna et al 2013 Araus and Cairns 2014 Tsonev et al 147
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
7
2014) Moreover metabolic markers start to draw attention due to their close 148
relationship with yield phenotypes (Fernie and Schauer 2009 Redestig et al 2011 149
Riedelsheimer et al 2012a Witt et al 2012 Riedelsheimer et al 2012b Degenkolbe 150
et al 2013) Accumulation of some metabolites has been reported to be directly related 151
to the performance of potato cultivars in beetle resistance in the field (Tai et al 2014) 152
Additionally identical genomic regions were mapped as both agronomic and metabolic 153
quantitative trait loci (QTL) in field grown maize and wheat indicating the utility of 154
metabolic traits for breeding selection (Riedelsheimer et al 2012b Hill et al 2015) A 155
recent study showed genetic gains in maize grain yield under drought stress were 156
higher using a molecular marker based approach than conventional breeding (Beyene 157
et al 2015) 158
Here we focused on the relationship between leaf metabolites and grain yield under 159
drought heat and simultaneous drought and heat conditions in the field The negative 160
effect of drought stress on maize yield is especially acute during the reproductive stage 161
between tassel emergence and early grain-filling (Grant et al 1989) where it is 162
believed to induce premature seed desiccation and to limit grain filling Grain is more 163
susceptible to drought stress than vegetative tissues and therefore the prediction of 164
grain yield from physiological parameter of leaves is a challenge (Sangoi and Salvador 165
1998 Khodarahmpour and Hamidi 2011) Nevertheless maize yield is dependent on 166
both the assimilate supply to the kernel (source) and the potential of kernel to 167
accommodate this assimilate (sink potential Jones and Simmons 1983) Breeding for 168
modern temperate hybrids has focused more on the sink potential particularly under 169
stress conditions (Tollenaar and Lee 2006) and there should therefore be 170
considerable potential remaining to improve source ability Drought and heat stresses 171
would be anticipated largely to affect leaf metabolism and especially photosynthesis 172
compromising the source capacity of leaves (Chaves et al 2009 Lawlor and Tezara 173
2009 Osakabe et al 2014) In keeping with this drought was found to have the most 174
dramatic effect on the metabolite composition in leaves compare to other organs in our 175
previous greenhouse experiments (Witt et al 2012) Since the source ability is closely 176
related to leaf metabolism the leaf metabolite profile should have close relationship to 177
grain yield particularly under conditions of stress Given that several recent studies have 178
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

6
As a first step toward this goal a deeper understanding of the plant response to the 117
stressful environment especially those to multiple stress conditions under field 118
conditions is crucial for improvement of stress-tolerant crops This is important on two 119
levels (i) in the field singular abiotic stresses are rare and (ii) yield and stress adaptation 120
are complex traits which render breeding gains slower than would be expected under 121
optimal conditions (Bruce et al 2002) Recent studies have revealed that the response 122
of plants to combinations of two or more stress conditions is unique and cannot be 123
directly extrapolated from their response to the different stresses when applied 124
individually This would be a result of complex combinations of different and sometimes 125
opposing responses in signalling pathways including those may interact and inhibit one 126
another (Prasch and Sonnewald 2013 Rasmussen et al 2013 Suzuki et al 2014) 127
Maize is grown in over 170 million hectares worldwide of which 130 million hectares are 128
in less-developed countries (FAO 2014) In sub-Saharan Africa (SSA) maize is a staple 129
crop however yields in this region have stagnated at less than 2 t ha-1 while maize 130
yields worldwide have continued to increase (Cairns et al 2012a) Low yields in SSA 131
are largely associated with drought stress and low soil fertility (Baumlnziger and Araus 132
2007) Additionally simulation studies indicate that maize yield in Africa is likely to be 133
significantly impaired by heat stress (Lobell and Burke 2010 Lobell et al 2011) such 134
as that can be anticipated as a result of the changes in climate predicted for the coming 135
decades (Muumlller et al 2011) Moreover sensitivity of maize yield to heat is exacerbated 136
under drought conditions (Lobell et al 2011 Cairns et al 2012ab Cairns et al 2013) 137
Therefore the development of maize germplasm tolerant to drought and heat conditions 138
is of utmost importance to both increase yields and offset predicted yield losses under 139
projected climate change scenarios (Easterling et al 2007) especially in SSA While 140
direct selection for grain yield under drought stress has resulted in admirable gains in 141
grain yield under stress (Baumlnziger et al 2006 Cairns et al 2013) further improvement 142
requires incorporation of additional selection traits (Cairns et al 2012a Cairns et al 143
2012b) In recent years genetic and phenotypic markers have been extensively 144
searched for drought tolerance of maize by high throughput genomic and phenotyping 145
approaches respectively (Tuberosa and Salvi 2006 Wen et al 2011 Araus et al 146
2012 Cairns et al 2013 Prasanna et al 2013 Araus and Cairns 2014 Tsonev et al 147
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
7
2014) Moreover metabolic markers start to draw attention due to their close 148
relationship with yield phenotypes (Fernie and Schauer 2009 Redestig et al 2011 149
Riedelsheimer et al 2012a Witt et al 2012 Riedelsheimer et al 2012b Degenkolbe 150
et al 2013) Accumulation of some metabolites has been reported to be directly related 151
to the performance of potato cultivars in beetle resistance in the field (Tai et al 2014) 152
Additionally identical genomic regions were mapped as both agronomic and metabolic 153
quantitative trait loci (QTL) in field grown maize and wheat indicating the utility of 154
metabolic traits for breeding selection (Riedelsheimer et al 2012b Hill et al 2015) A 155
recent study showed genetic gains in maize grain yield under drought stress were 156
higher using a molecular marker based approach than conventional breeding (Beyene 157
et al 2015) 158
Here we focused on the relationship between leaf metabolites and grain yield under 159
drought heat and simultaneous drought and heat conditions in the field The negative 160
effect of drought stress on maize yield is especially acute during the reproductive stage 161
between tassel emergence and early grain-filling (Grant et al 1989) where it is 162
believed to induce premature seed desiccation and to limit grain filling Grain is more 163
susceptible to drought stress than vegetative tissues and therefore the prediction of 164
grain yield from physiological parameter of leaves is a challenge (Sangoi and Salvador 165
1998 Khodarahmpour and Hamidi 2011) Nevertheless maize yield is dependent on 166
both the assimilate supply to the kernel (source) and the potential of kernel to 167
accommodate this assimilate (sink potential Jones and Simmons 1983) Breeding for 168
modern temperate hybrids has focused more on the sink potential particularly under 169
stress conditions (Tollenaar and Lee 2006) and there should therefore be 170
considerable potential remaining to improve source ability Drought and heat stresses 171
would be anticipated largely to affect leaf metabolism and especially photosynthesis 172
compromising the source capacity of leaves (Chaves et al 2009 Lawlor and Tezara 173
2009 Osakabe et al 2014) In keeping with this drought was found to have the most 174
dramatic effect on the metabolite composition in leaves compare to other organs in our 175
previous greenhouse experiments (Witt et al 2012) Since the source ability is closely 176
related to leaf metabolism the leaf metabolite profile should have close relationship to 177
grain yield particularly under conditions of stress Given that several recent studies have 178
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

7
2014) Moreover metabolic markers start to draw attention due to their close 148
relationship with yield phenotypes (Fernie and Schauer 2009 Redestig et al 2011 149
Riedelsheimer et al 2012a Witt et al 2012 Riedelsheimer et al 2012b Degenkolbe 150
et al 2013) Accumulation of some metabolites has been reported to be directly related 151
to the performance of potato cultivars in beetle resistance in the field (Tai et al 2014) 152
Additionally identical genomic regions were mapped as both agronomic and metabolic 153
quantitative trait loci (QTL) in field grown maize and wheat indicating the utility of 154
metabolic traits for breeding selection (Riedelsheimer et al 2012b Hill et al 2015) A 155
recent study showed genetic gains in maize grain yield under drought stress were 156
higher using a molecular marker based approach than conventional breeding (Beyene 157
et al 2015) 158
Here we focused on the relationship between leaf metabolites and grain yield under 159
drought heat and simultaneous drought and heat conditions in the field The negative 160
effect of drought stress on maize yield is especially acute during the reproductive stage 161
between tassel emergence and early grain-filling (Grant et al 1989) where it is 162
believed to induce premature seed desiccation and to limit grain filling Grain is more 163
susceptible to drought stress than vegetative tissues and therefore the prediction of 164
grain yield from physiological parameter of leaves is a challenge (Sangoi and Salvador 165
1998 Khodarahmpour and Hamidi 2011) Nevertheless maize yield is dependent on 166
both the assimilate supply to the kernel (source) and the potential of kernel to 167
accommodate this assimilate (sink potential Jones and Simmons 1983) Breeding for 168
modern temperate hybrids has focused more on the sink potential particularly under 169
stress conditions (Tollenaar and Lee 2006) and there should therefore be 170
considerable potential remaining to improve source ability Drought and heat stresses 171
would be anticipated largely to affect leaf metabolism and especially photosynthesis 172
compromising the source capacity of leaves (Chaves et al 2009 Lawlor and Tezara 173
2009 Osakabe et al 2014) In keeping with this drought was found to have the most 174
dramatic effect on the metabolite composition in leaves compare to other organs in our 175
previous greenhouse experiments (Witt et al 2012) Since the source ability is closely 176
related to leaf metabolism the leaf metabolite profile should have close relationship to 177
grain yield particularly under conditions of stress Given that several recent studies have 178
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

8
indicated the importance of metabolic pre-adaptation to various stress tolerance in 179
plants (Sanchez et al 2011 Benina et al 2013) we also postulate that basal 180
metabolite levels under optimal growth condition could be correlated to stress tolerance 181
In order to test this metabolite profiles of the leaf blades of ten hybrids were analysed in 182
field experiments conducted at CIMMYT subtropical experimental station in 2010 and 183
2011 in which the plants were exposed to singular or combined drought and heat 184
stresses (Cairns et al 2012a Cairns et al 2013) The results are discussed both in the 185
context of current models of stress tolerance as well as with respect to their practical 186
implications for future breeding strategies 187
188
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

9
189
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

10
Results 190
Grain yield was affected by stress treatments 191
Maize plants were grown as part of the CIMMYT field trials in which a collection of 300 192
hybrid lines representing the genetic diversity within the CIMMYT and IITA tropical and 193
subtropical maize improvement programs (Wen et al 2011) were tested for tolerance to 194
drought heat and combination of drought and heat stresses (Cairns et al 2013) Ten 195
maize experimental hybrids (Table I) were chosen to cover a wide range of drought and 196
heat tolerance on grain yield observed in previous field trials (Cairns et al 2013) 197
Drought stress (DS) was imposed by stopping irrigation before flowering to achieve 198
water deficiency at anthesis stage This treatment induces mild drought stress which 199
increase canopy temperature for one to two degrees in the absence of heat stress 200
(Romano et al 2011 Zia et al 2013) However climate conditions varied slightly 201
between the two years of evaluation In 2010 one rainfall event (with 56 mm 202
precipitation) occurred at the start of February one week before drought was applied 203
under optimal temperature (and three weeks before trial mean anthesis) For the 204
combined drought and heat stress one rainfall event of 35 mm occurred two weeks 205
before the start of anthesis and the day before the last irrigation to apply drought stress 206
In 2011 one rainfall event of 13 mm occurred 10 days before the start of anthesis in the 207
combined drought and heat stress trial and 4 days after drought stress was applied 208
(Table II) Drought treatment significantly decreased average grain yields to 458 and 209
352 of well watered plants in 2010 and 2011 seasons respectively (Fig1A) Grain 210
yield across treatments varied in genotypes from 183 to 432 Mg ha-1 in 2010 and from 211
065 to 428 Mg ha-1 in 2011 (Supplemental Fig S1) Grain yield of individual genotypes 212
tended to be in agreement with tolerance levels determined from the previous field 213
experiments (Table I Supplemental FigS1 Supplemental Table S2) Plant height did 214
not differ between tolerant and susceptible genotypes in accordance with occurrence of 215
the stress around flowering when plants have already achieved their maximum height 216
(Supplemental FigS1 Supplemental Table S2) Other parameters did not differ 217
between tolerant and susceptible genotypes whereas tolerant genotypes reached 218
earlier anthesis and silking under HS 219
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

11
Drought treatment did not invoke consistent effects on silking date anthesis date 220
anthesissilking interval ear height and ear number across the two harvests (Fig1B to E 221
and G Supplemental Fig S1) Heat stress (HS) was applied by growing plants in the 222
dry season A group of plants were additionally treated by drought for combined stress 223
(DS+HS) treatment (for precise details see Materials and Methods) HS decreased plant 224
height and shortened the time till silking and anthesis in both years (Fig 1D and E 225
Supplemental Fig S1) However grain yield was significantly decreased in 2010 but not 226
in 2011 (Fig 1A) The grain yield under combined drought and heat condition was also 227
the same level as that in drought condition in 2011(Fig 1A) indicating that the heat 228
treatment in 2011 was not severe enough to affect grain yield Therefore the heat 229
treatment in 2011 was recognized as ldquomild HSrdquo which does not cause yield reduction 230
DS+HS exclusively affected ear number and anthesis silking interval (Fig 1D and E) 231
and lead to a severe yield reduction in 2010 (Fig 1A) Two way ANOVA indicated that 232
grain yield was significantly influenced by genotype treatment and also by their 233
interaction in both years (Supplemental Table S1) Ear number and anthesis date were 234
also influenced by the interaction of genotype and treatment in 2010 but not in 2011 235
(Supplemental Table S1) These results indicate that grain yield is the most suitable 236
parameter to assess genotypic variation of stress tolerance in the current study and 237
were mainly used for correlation analysis with metabolite profile 238
239
Metabolite profiling revealed differential metabolic responses of genotypes to 240
stress conditions 241
Leaf material was harvested for metabolite profiling two weeks after cessation of 242
irrigation Gas chromatography-mass spectroscopy (GC-MS) based metabolite profiling 243
was conducted to analyse the metabolites of central carbon metabolism Metabolite 244
levels were analysed on a per dry weight basis to avoid the effect of differential water 245
contents Forty one metabolites were detected in more than half of the samples Some 246
highly abundant metabolites including sucrose and glucose could not be quantified due 247
to being above the dynamic range limit of the specified settings of the GC-MS The 248
metabolites were clustered into three groups when analysed by hierarchical clustering 249
(Fig 2) The first cluster includes amino acids (eg tryptophan asparagine szlig-alanine 250
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
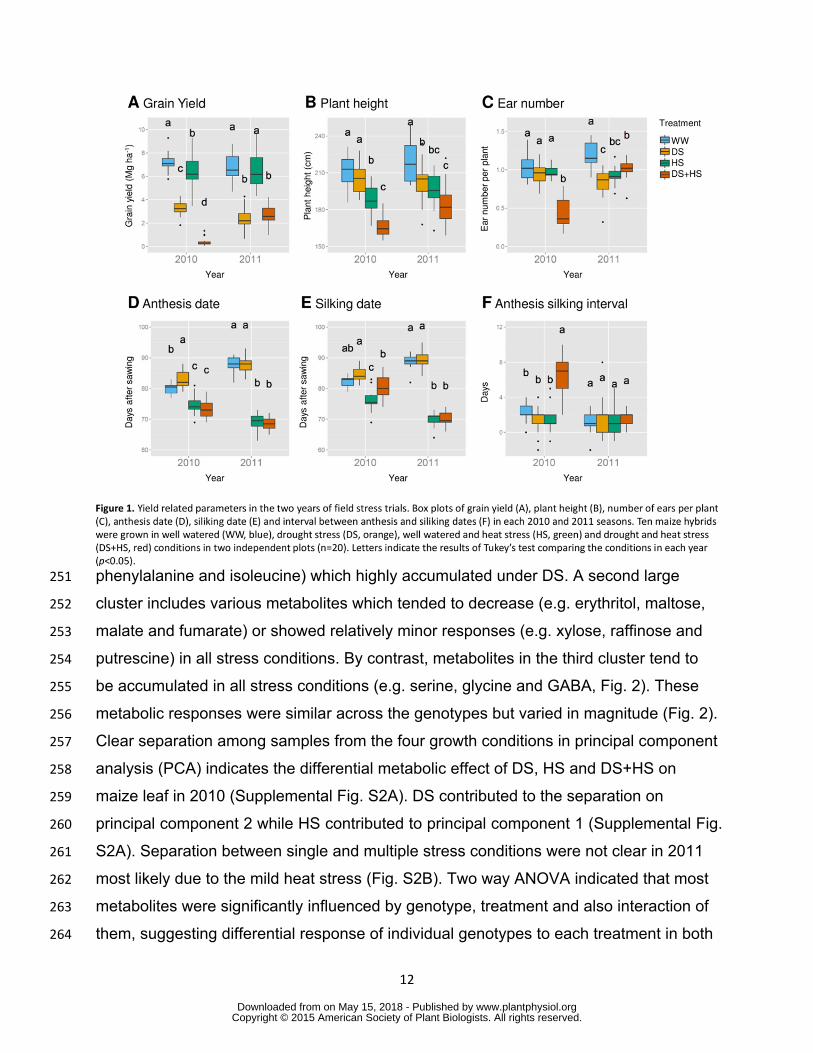
12
phenylalanine and isoleucine) which highly accumulated under DS A second large 251
cluster includes various metabolites which tended to decrease (eg erythritol maltose 252
malate and fumarate) or showed relatively minor responses (eg xylose raffinose and 253
putrescine) in all stress conditions By contrast metabolites in the third cluster tend to 254
be accumulated in all stress conditions (eg serine glycine and GABA Fig 2) These 255
metabolic responses were similar across the genotypes but varied in magnitude (Fig 2) 256
Clear separation among samples from the four growth conditions in principal component 257
analysis (PCA) indicates the differential metabolic effect of DS HS and DS+HS on 258
maize leaf in 2010 (Supplemental Fig S2A) DS contributed to the separation on 259
principal component 2 while HS contributed to principal component 1 (Supplemental Fig 260
S2A) Separation between single and multiple stress conditions were not clear in 2011 261
most likely due to the mild heat stress (Fig S2B) Two way ANOVA indicated that most 262
metabolites were significantly influenced by genotype treatment and also interaction of 263
them suggesting differential response of individual genotypes to each treatment in both 264
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

13
years (Fig 3A and C) The effects on levels of individual metabolites are summarized in 265
Supplemental Table S3 When the effect of treatments was tested ANOVA revealed 266
most metabolites were separately influenced by each stress (Fig 3B and D) However 267
just 13 metabolites were significantly affected by the interaction of HS and DS in 2010 268
(Fig 3B) suggesting limited metabolites responded to combined stresses in a specific 269
manner or the effects of individual stress components compensated each other in some 270
metabolites Some metabolites showed clear tendency of differential accumulation in 271
stress tolerant and sensitive hybrids under stress conditions Among them galactinol 272
levels were lower in tolerant genotypes than in susceptible ones under DS condition in 273
both years (Supplemental Table S4) Accumulation of this metabolite varied between 274
genotypic groups with different tolerance levels under all stress conditions tested 275
(Supplemental Table S4) suggesting a relationship with stress tolerance 276
277
DS and HS conditions invoke increases in the levels of many amino acids 278
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
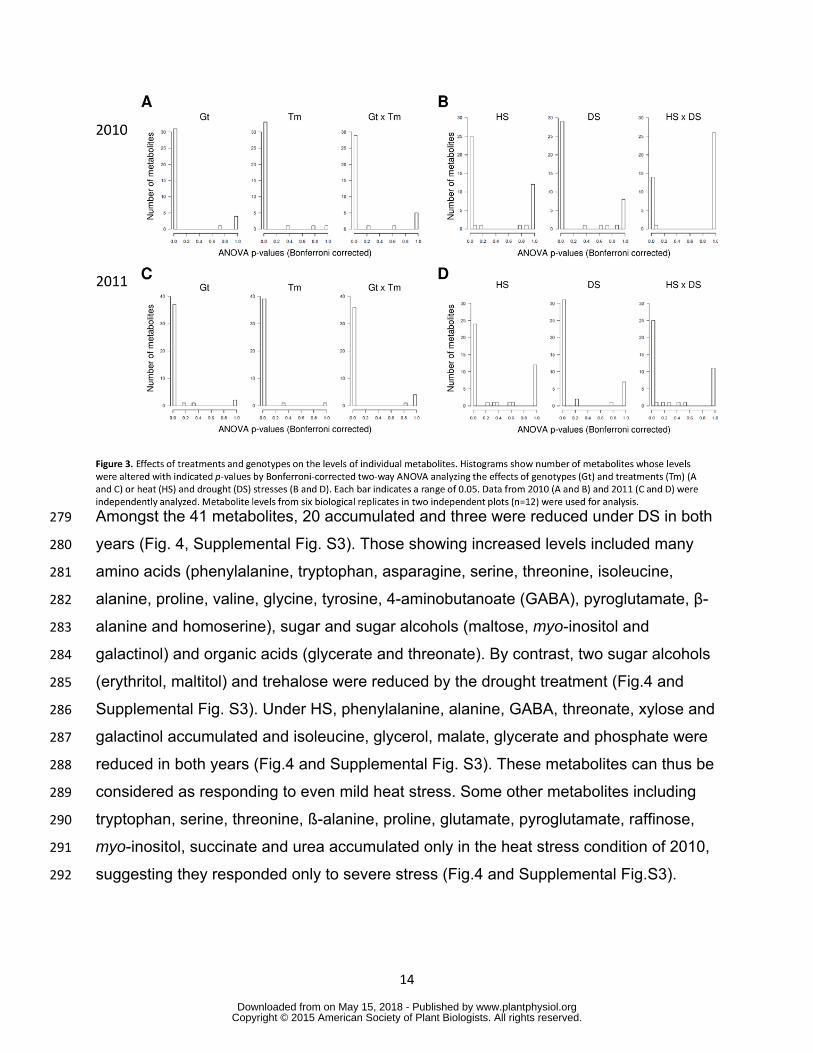
14
Amongst the 41 metabolites 20 accumulated and three were reduced under DS in both 279
years (Fig 4 Supplemental Fig S3) Those showing increased levels included many 280
amino acids (phenylalanine tryptophan asparagine serine threonine isoleucine 281
alanine proline valine glycine tyrosine 4-aminobutanoate (GABA) pyroglutamate β-282
alanine and homoserine) sugar and sugar alcohols (maltose myo-inositol and 283
galactinol) and organic acids (glycerate and threonate) By contrast two sugar alcohols 284
(erythritol maltitol) and trehalose were reduced by the drought treatment (Fig4 and 285
Supplemental Fig S3) Under HS phenylalanine alanine GABA threonate xylose and 286
galactinol accumulated and isoleucine glycerol malate glycerate and phosphate were 287
reduced in both years (Fig4 and Supplemental Fig S3) These metabolites can thus be 288
considered as responding to even mild heat stress Some other metabolites including 289
tryptophan serine threonine szlig-alanine proline glutamate pyroglutamate raffinose 290
myo-inositol succinate and urea accumulated only in the heat stress condition of 2010 291
suggesting they responded only to severe stress (Fig4 and Supplemental FigS3) 292
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
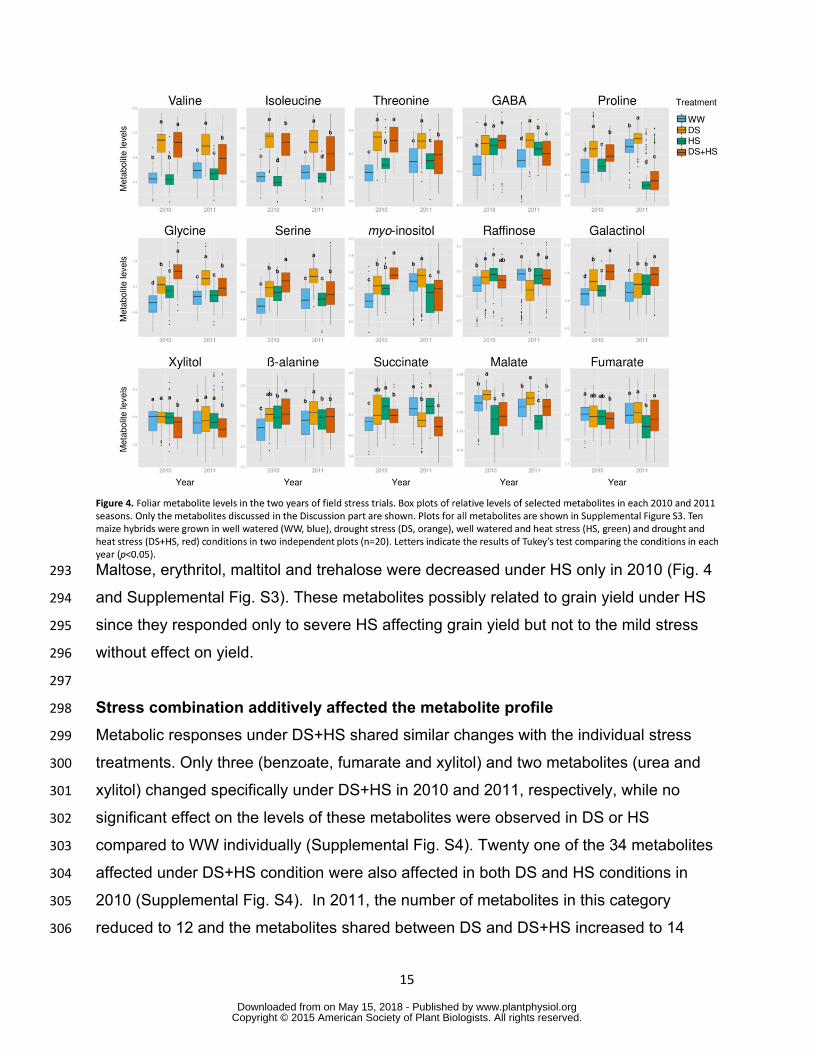
15
Maltose erythritol maltitol and trehalose were decreased under HS only in 2010 (Fig 4 293
and Supplemental Fig S3) These metabolites possibly related to grain yield under HS 294
since they responded only to severe HS affecting grain yield but not to the mild stress 295
without effect on yield 296
297
Stress combination additively affected the metabolite profile 298
Metabolic responses under DS+HS shared similar changes with the individual stress 299
treatments Only three (benzoate fumarate and xylitol) and two metabolites (urea and 300
xylitol) changed specifically under DS+HS in 2010 and 2011 respectively while no 301
significant effect on the levels of these metabolites were observed in DS or HS 302
compared to WW individually (Supplemental Fig S4) Twenty one of the 34 metabolites 303
affected under DS+HS condition were also affected in both DS and HS conditions in 304
2010 (Supplemental Fig S4) In 2011 the number of metabolites in this category 305
reduced to 12 and the metabolites shared between DS and DS+HS increased to 14 306
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

16
most likely due to mild heat stress (Supplemental Fig S4) k-means clustering was 307
performed in order to classify the metabolites according to the responses to stress 308
conditions in 2010 (Supplemental Fig S5) Most of the metabolites fitted well into five 309
clusters with some exceptions in which the response were not clear Those clustered 310
into the first three and the latter two clusters tended to accumulate and decrease under 311
DS+HS respectively Metabolites in cluster 1 and 2 were increased in all stress 312
conditions although those in cluster 1 further accumulated by stress combination 313
Cluster 3 includes many amino acids highly accumulated in both DS and DS+HS 314
conditions The metabolites in cluster 4 are characterized by the reduction in HS while 315
cluster 5 includes those specifically decreased under DS+HS (Supplemental Fig S5) 316
According to the criteria of response modes defined for describing transcript response 317
under stress combination by Rasmussen et al (2013) most of the metabolites in cluster 318
1 2 and some in 4 are assigned to ldquosimilarrdquo response mode Cluster 3 to 5 mostly 319
contain metabolites responded in ldquoindependentrdquo mode whilst just three metabolites 320
namely benzoate in cluster 1 and fumarate and xylitol in cluster 5 could be classified as 321
belonging to the ldquocombinatorialrdquo mode Interestingly metabolic responses in the 2010 322
trial could be well classified into just five of 20 scenarios which were predefined to the 323
responses against stress combinations (Rasmussen et al 2013) It should also be 324
noted that most of the metabolic changes in DS+HS should be predictable from the 325
metabolic responses to each single stress treatment since ldquosimilarrdquo and ldquoindependentrdquo 326
response modes are considered to be predictable (Rasmussen et al 2013) 327
Proline is a metabolite whose function in DS+HS condition has been reported (Rizhsky 328
et al 2004) The accumulation of proline differed between 2010 and 2011 in our maize 329
field trial While proline levels in DS+HS were reduced in comparison to DS in both 330
years it was much lower in 2011 along with the proline level under single HS It should 331
also be noted that the level of proline in DS+HS was still significantly higher than that in 332
WW condition in 2010 (Fig 4) 333
334
Metabolic responses under stress combination could be predicted from the sum 335
of those in single stresses 336
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

17
Many of the metabolic responses especially those in cluster 1 2 and 3 seem to be 337
predictable not only qualitatively but also quantitatively by the simple sum of responses 338
in DS and HS In order to test this hypothesis response factor was calculated by 339
dividing the metabolite level under stress conditions by that in WW Following log2 340
transformation correlations between the sum of the response factors in DS and HS 341
(predicted response factor in DS+HS) and the actual response factor in DS+HS were 342
tested (Table III Supplemental Table S5) Surprisingly the predicted response factors 343
significantly correlated with actual response factors in 17 of 41 metabolites in 2010 344
(Table III) The means of the predicted and actual values were fairly similar in most of 345
the metabolites and t-test showed significant difference between these two values in 346
only 11 of the 41 metabolites (Table III) This analysis suggested that a large part of 347
metabolic response under stress combination could be explained by the additive effects 348
of individual treatments Predicted and actual response factors were correlated in more 349
metabolites in 2011 but this is most likely due to weak effect of HS (Supplemental Table 350
S5) On the other hand the predicted response factor was significantly different from the 351
actual ones in eight metabolites including tyrosine succinate urea GABA raffinose 352
and xylose (Table III) The levels of these metabolites are most likely determined by 353
regulatory mechanisms specifically operating under combined stress conditions 354
355
Correlation analysis revealed a close relationship between leaf metabolite levels 356
and grain yield under stress conditions 357
In order to identify metabolites closely related to grain yield under stress conditions 358
correlation between levels of each metabolite and grain yield in each growth condition 359
was tested by Pearson correlation analysis (Table IV) When the relationship between 360
metabolite levels and grain yield in optimal growth condition was tested only szlig-alanine 361
and maltitol displayed significant correlation (Table IV) Under DS condition levels of 362
glycerol and glycerate showed positive correlation and those of asparagine serine 363
pyroglutamate phosphate itaconate and galactinol were negatively correlated to grain 364
yield Glycine myo-inositol threonate glycerol-3-phosphate and nicotinate showed 365
strong negative correlation to grain yield with the p lt 001 (Table IV Fig 5) Correlation 366
analysis in HS and DS+HS conditions were conducted only for 2010 data since heat 367
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
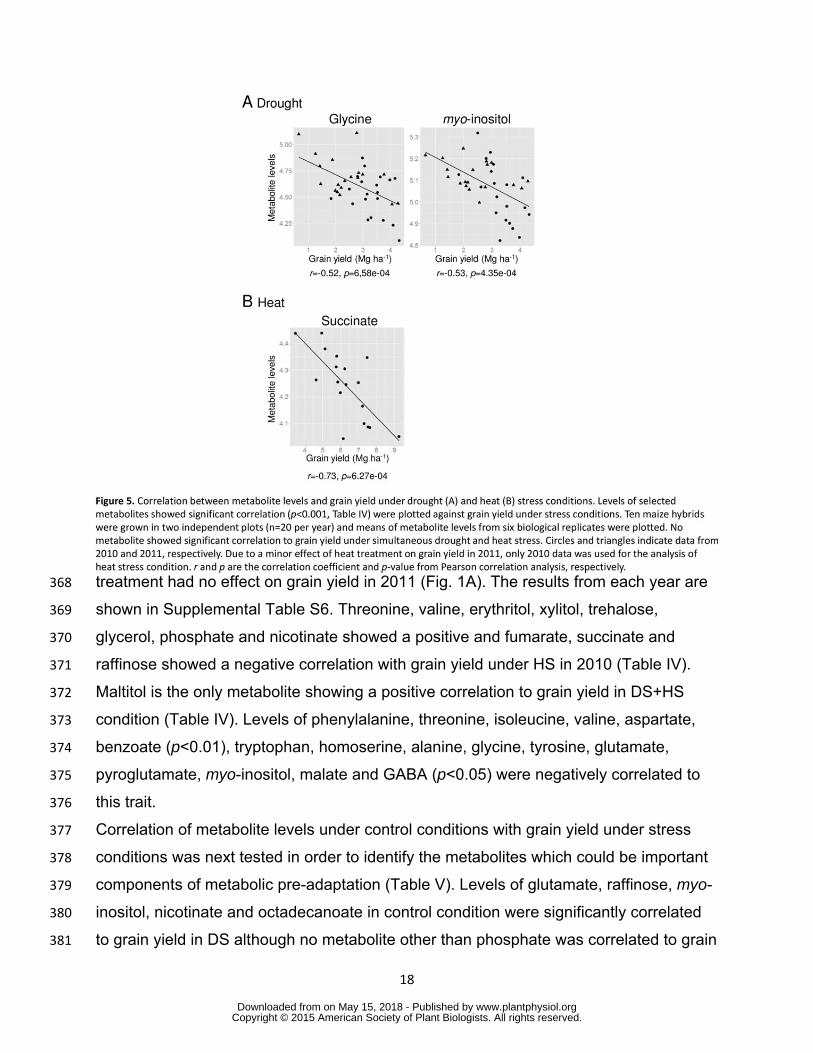
18
treatment had no effect on grain yield in 2011 (Fig 1A) The results from each year are 368
shown in Supplemental Table S6 Threonine valine erythritol xylitol trehalose 369
glycerol phosphate and nicotinate showed a positive and fumarate succinate and 370
raffinose showed a negative correlation with grain yield under HS in 2010 (Table IV) 371
Maltitol is the only metabolite showing a positive correlation to grain yield in DS+HS 372
condition (Table IV) Levels of phenylalanine threonine isoleucine valine aspartate 373
benzoate (plt001) tryptophan homoserine alanine glycine tyrosine glutamate 374
pyroglutamate myo-inositol malate and GABA (plt005) were negatively correlated to 375
this trait 376
Correlation of metabolite levels under control conditions with grain yield under stress 377
conditions was next tested in order to identify the metabolites which could be important 378
components of metabolic pre-adaptation (Table V) Levels of glutamate raffinose myo-379
inositol nicotinate and octadecanoate in control condition were significantly correlated 380
to grain yield in DS although no metabolite other than phosphate was correlated to grain 381
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
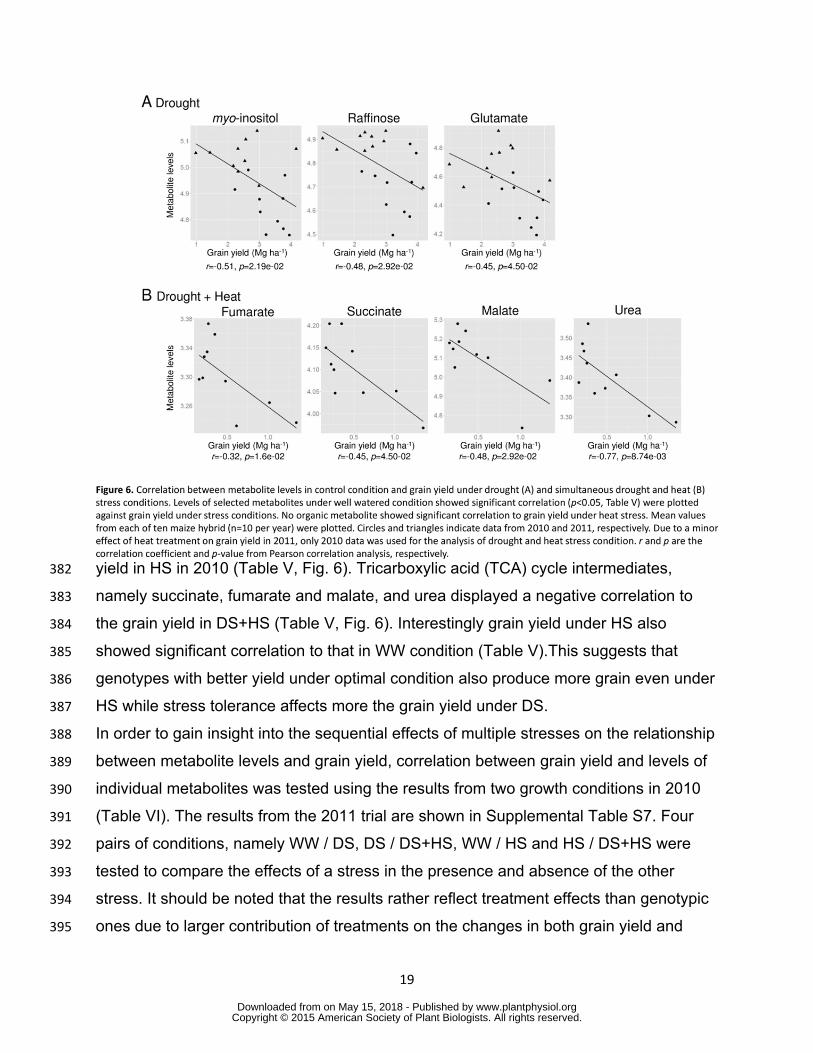
19
yield in HS in 2010 (Table V Fig 6) Tricarboxylic acid (TCA) cycle intermediates 382
namely succinate fumarate and malate and urea displayed a negative correlation to 383
the grain yield in DS+HS (Table V Fig 6) Interestingly grain yield under HS also 384
showed significant correlation to that in WW condition (Table V)This suggests that 385
genotypes with better yield under optimal condition also produce more grain even under 386
HS while stress tolerance affects more the grain yield under DS 387
In order to gain insight into the sequential effects of multiple stresses on the relationship 388
between metabolite levels and grain yield correlation between grain yield and levels of 389
individual metabolites was tested using the results from two growth conditions in 2010 390
(Table VI) The results from the 2011 trial are shown in Supplemental Table S7 Four 391
pairs of conditions namely WW DS DS DS+HS WW HS and HS DS+HS were 392
tested to compare the effects of a stress in the presence and absence of the other 393
stress It should be noted that the results rather reflect treatment effects than genotypic 394
ones due to larger contribution of treatments on the changes in both grain yield and 395
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

20
metabolite levels Twenty metabolites showed significant correlation with grain yield 396
commonly in WWDS and HSDS+HS condition pairs indicating these metabolites were 397
similarly affected by DS regardless the presence of HS (Table VI) Aspartate maltose 398
xylitol and xylose were four metabolites which showed correlation to grain yield only at 399
the presence of HS Contrary to DS only six metabolites were correlated with grain 400
yield commonly when WWHS and HSDS+HS pairs were tested (Table VI) Thirteen 401
and eight metabolites were significantly correlated with grain yield under HS specifically 402
in the presence (DSDS+HS) and absence (WWHS) of DS respectively This suggests 403
that effects of metabolites on grain yield under HS are largely dependent on the 404
presence of DS 405
406
Combination of metabolite levels could explain the variation of grain yield by 407
multiple regression modelling 408
Additionally a multiple linear regression model was constructed in order to identify 409
groups of parameters which coordinately affect grain yield in each growth condition for 410
the 2010 data Grain yield was used as dependent variable and minimum number of 411
independent variables were selected from all parameters measured in this study only 412
metabolite levels and at last metabolite levels under WW which contributed to fully 413
explain the variation of grain yield among genotypes (Table VII) The models requires 414
10 to 15 parameters to explain the variation of grain yield in most cases but only seven 415
were used for DS condition (Table VII) Models were quite similar when all parameters 416
(agronomical variables and metabolites) or only the metabolites were used as 417
independent variables Especially the models selected for DS were identical in both 418
cases (Table VII A B) Similar sets of metabolites under WW condition explained the 419
variation of grain yield in WW and stress conditions (Table VII C) In the case of DS 420
exactly the same sets of metabolites as in WW were selected (Table VII C) 421
Interestingly galactinol is selected for all models (Table VII) When the same analysis 422
was conducted for 2011 data galactinol was again selected as a parameter in all 423
models whereas the models required more independent variables than for the 2010 424
data to explain yield variation (Supplemental Table S8) 425
426
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

21
427
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

22
Discussion 428
Comparison of drought stress responses in field and greenhouse experiments 429
Large scale metabolite analyses under stress conditions in the field remains rare To 430
our best knowledge this is the first study reporting metabolic effects of simultaneous 431
abiotic stresses in field grown plants Heat stress was applied by altering the planting 432
date to ensure the reproductive phase coincided with high temperatures (Craufurd et al 433
2013) Despite limitations in fine climate control large scale field trials are still valuable 434
since it is often reported that important agronomical traits are masked in greenhouse 435
grown crops (Alexandersson et al 2014) In our previous study in controlled 436
greenhouse conditions genotypes chosen to cover wide range of DS tolerance based 437
on field results did not display differential effects of DS on physiological traits (Witt et al 438
2012) Further field studies showed these contrasting genotypes to show differential 439
physiological responses to DS (Cairns et al 2012a Cairns et al 2013) Additionally in 440
the current study genotypes showed differential physiological responses to DS albeit all 441
six genotypes tested in the greenhouse experiment were also included and other four 442
were selected by the same criteria These results reaffirmed the importance of 443
conducting field experiments to understand effects of abiotic stresses on crops On the 444
other hand some metabolic responses were shared in both greenhouse and field trials 445
Accumulation of amino acids including isoleucine valine threonine and GABA is a 446
metabolic response common in many abiotic stress environments in Arabidopsis (Obata 447
and Fernie 2012)These metabolites were also accumulated in maize in both 448
greenhouse and field trials under all stress conditions tested although threonine and 449
GABA were not annotated in greenhouse samples (Sicher and Barnaby 2012 Witt et 450
al 2012 Barnaby et al 2013) Other amino acids such as proline phenylalanine and 451
tryptophan also accumulated under DS in both conditions (Sicher and Barnaby 2012 452
Witt et al 2012 Barnaby et al 2013) as well as in Arabidopsis (Urano et al 2009) 453
The accumulation of these metabolites was much lower in this field study than the other 454
greenhouse studies (Sicher and Barnaby 2012 Witt et al 2012 Barnaby et al 2013) 455
most likely depending on the severity of drought stress due to the soil structure and 456
coincident rainfalls As accumulation of amino acids under DS has been reported in 457
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

23
various plant species (Evers et al 2010 Degenkolbe et al 2013 Barchet et al 2014 458
Hatzig et al 2014 Suguiyama et al 2014) it can be considered as a well-conserved 459
and robust metabolic response to DS in plants This response might be due to less 460
dilution effect caused by diminished growth under stress conditions (Geacutenard et al 461
2014) However our field study was performed in fully grown plants and DS was 462
imposed near flowering which is the most sensitive stage of maize grain production to 463
drought stress but leaf expansion had finished at that period It should also be noted the 464
plant height was not significantly affected by DS in 2010 indicating the dilution effect 465
played a minor role in amino acid accumulation Proline is one of many well-known 466
compatible solutes in plants (Hare and Cress 1997) Branched chain amino acids 467
(BCAAs valine leucine and isoleucine) and other amino acids sharing synthetic 468
pathways with BCAA (lysine threonine and methionine) accumulate in various abiotic 469
stress conditions (Obata and Fernie 2012) and have also been proposed as compatible 470
solutes (Joshi et al 2010) or alternative electron donors for respiratory electron 471
transport chain (Arauacutejo et al 2011) although comparative assessment of these 472
functions under drought stress conditions remain elusive 473
Metabolic effects of individual drought and heat stresses 474
In addition to the metabolites described in the previous paragraph many metabolites 475
accumulated under DS in both years Although some of them including GABA and 476
galactinol have been suggested to function in abiotic stress tolerance (Fait et al 2008 477
Nishizawa et al 2008) we focus mainly on glycine and serine here and on myo-inositol 478
in a later paragraph Apart from other amino acids glycine and serine are closely related 479
to photorespiration (Bauwe et al 2010) Even in C4 plants like maize in which RubisCO 480
oxygenation reaction should take place at a lower rate than in C3 plants recent studies 481
have indicated the essentiality of photorespiration for growth under normal air (Zelitch et 482
al 2009 Maurino and Peterhansel 2010) Both glycine and serine were accumulated 483
under DS in the current study suggesting altered photorespiratory flux This might be 484
related to yield performance under DS since these metabolites especially glycine 485
correlated to grain yield Photorespiration has actually been proposed to function in 486
protection from photoinhibition under drought salt and high light stresses as a sink of 487
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

24
excess reducing equivalent (Wingler et al 2000) andor by preventing excess 488
accumulation of reactive oxygen species (Voss et al 2013) It has also been shown to 489
contribute in tolerance to moderate water deficiency in tomato plants by ameliorating 490
nitrogen use efficiency reduced by lower nitrogen assimilation (Saacutenchez-Rodriacuteguez et 491
al 2011) It should also be noted that glycine and serine are principal sources of one-492
carbon unit largely consumed to synthesize an osmoprotectant glycine betaine in 493
some plant tissues (Hanson and Roje 2001) The levels of this osmolite differ among 494
maize varieties (Brunk et al 1989) and positively correlate to the degree of salt 495
tolerance (Saneoka et al 1995) Negative correlations between grain yield in DS and 496
levels of glycine and serine are possibly related to the levels of consumption of these 497
metabolites to synthesize glycine betaine leading to the variation of yield performance 498
under drought in maize genotypes Trehalose is another well-known osmoprotectant in 499
some insects plants and yeast but its accumulation and function are species specific 500
(Iturriaga et al 2009) The decrease of trehalose under DS in this study suggests its 501
function other than as an osmoprotectant in maize 502
Among the metabolites those responded to HS succinate accumulated and its level 503
negatively correlated to grain yield under severe HS in 2010 This is a novel observation 504
to our knowledge although the functional background is hardly explained Since 505
succinate is a metabolite connecting TCA cycle and GABA shunt (Fait et al 2008) the 506
balance between these two pathways might affect succinate level Interestingly GABA 507
shunt related metabolites namely GABA and glutamate and TCA cycle organic acid 508
malate and fumarate were increased and decreased under HS respectively 509
510
Effects of simultaneous drought and heat stresses 511
The effects of simultaneous application of drought and heat have been relatively well 512
studied in comparison to other stress combinations due to its economic impact and 513
increasing risk by global climate change in the near future (Suzuki et al 2014) There 514
are two studies so far in which metabolite profiles under DS+HS conditions in 515
Arabidopsis were examined with application of relatively mild (Prasch and Sonnewald 516
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

25
2013) and severe (Rizhsky et al 2004) heat stresses In both studies proline was 517
accumulated under DS but not in DS+HS (Rizhsky et al 2004 Prasch and Sonnewald 518
2013) This is explained as a consequence of avoidance of the toxic effect of proline 519
under HS (Rizhsky et al 2004) Such clear regulation was not observed in this study 520
and proline levels were differentially affected by stresses in the two years Whilst this 521
might be related to varied environmental conditions between the two years including 522
stress levels it is more likely due to different adaptation strategies of DS between maize 523
and Arabidopsis It is becoming clear that plant species have specific preferences on 524
the selection of compatible solutes to accumulate under stress conditions (Gong et al 525
2005 Benina et al 2013) While proline is one of the well-known compatible solute in 526
Arabidopsis (Hare and Cress 1997) this species accumulates only small amounts of 527
glycine-betaine (Missihoun et al 2011) which has been proven to be involved in stress 528
tolerance in maize (Brunk et al 1989) It is therefore conceivable that the degree of 529
dependence on proline for DS tolerance andor cellular proline concentration are 530
different between Arabidopsis and maize (Spoljarević et al 2011 Sperdouli and 531
Moustakas 2012) resulting in the different regulation of proline level under DS+HS 532
condition 533
Similar to both Arabidopsis studies only few metabolites specifically responded to 534
DS+HS in our field maize experiment Interestingly most of the metabolic changes in 535
DS+HS were quantitatively predictable from the sum of responses to each single stress 536
in contrast to transcript responses in Arabidopsis (Rasmussen et al 2013) In fact the 537
Arabidopsis metabolite profiling results from milder stress treatments show similar 538
tendency (Prasch and Sonnewald 2013) but not in the severe stress experiment 539
(Rizhsky et al 2004) It is possible that metabolic pathways are regulated to meet the 540
metabolic demands under each stress condition resulting in additive metabolite profile 541
under stress combination unless the metabolic network is collapsed by severe stress 542
treatments Given that the naturally feasible stresses are imposed more mildly than 543
typical stress treatments in greenhouse experiments (Romano et al 2011 Zia et al 544
2013) the general metabolic response in stress combination should be considered as 545
the sum of individual stresses in the field This is also supported by PCA in which 546
drought and heat contribute the majority of the variance observed in the metabolic data 547
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

26
with PC1 separating DS from WW PC2 separating H from no H and DS+H being 548
separated from WW in an additive fashion Another result supporting this argument 549
comes from the correlation analysis using two conditions in which the effect of DS was 550
well conserved regardless the presence of HS However HS treatments were differently 551
affected in the presence or the absence of DS indicating specific effects of HS under 552
stress combination on both metabolite levels and the grain yield This might be due to 553
stomatal closure which would be anticipated to occur under DS and which would be 554
expected to induce effects of HS on plant metabolism Indeed the negative effect of 555
heat on photosynthesis has been reported to be apparent only at the presence of DS in 556
European oak tobacco and wheat (Suzuki et al 2014) 557
558
myo-inositol as a potential metabolic marker for breeding of drought tolerant 559 maize 560
One of our main goals was to find metabolic markers useful for the selection of maize 561
genotypes giving better grain yield under abiotic stress conditions The preferential 562
choice of metabolite levels in the variable selection by multiple linear regression 563
analysis suggests metabolic traits to be promising markers which might behave stronger 564
than classical agronomical yield components for explaining variability in grain yield 565
Although further validation of the results and efficient methods for screening are 566
required for the actual use of candidate metabolite markers in breeding new strategies 567
of molecular breeding such as marker-assisted recurrent selection (MARS) which 568
require only one cycle of phenotyping and subsequently focus on selection based on 569
genotypic data potentially open up new avenues for high cost low throughput 570
phenotyping options (Jannink et al 2010 Bohra 2013) Selection markers which can 571
be determined in optimal growth condition are desired since it is very difficult to control 572
stress conditions in the field Metabolic markers would be a promising target because 573
the species specific metabolite profile under non-stress condition has been recognized 574
to be closely related to stress tolerance and the adaptation strategy of plant species 575
(Benina et al 2013) myo-inositol is the most promising candidate of a single marker 576
metabolite for yield performance under drought found in this study It was accumulated 577
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

27
and its level was negatively correlated to grain yield under DS Additionally its level in 578
WW condition was also negatively correlated to grain yield in DS These results suggest 579
a possibility of marker assisted breading to choose maize genotype raising better grain 580
yield under DS by the selection of genotype containing less myo-inositol in well-watered 581
leaves myo-inositol itself is implicated to function as an osmolite (Kaur et al 2013) like 582
other sugar alcohols However the importance of myo-inositol in plant stress tolerance is 583
rather related to its function as a precursor of many metabolites involved in abiotic 584
stress tolerance Raffinose family oligosaccharides (RFOs) especially raffinose are 585
ubiquitous in plant kingdom and contribute to stress tolerance likely by membrane 586
stabilization and antioxidative functions (Van den Ende 2013) Raffinose is synthesized 587
by adding a galactose residue from galactinol to sucrose and myo-inositol is used to 588
synthesize galactinol Therefore cellular myo-inositol metabolism is closely related to 589
the accumulation of RFOs and further to stress tolerance (Elsayed et al 2014) Actually 590
galactinol and raffinose were accumulated under DS as myo-inositol although raffinose 591
reduced in 2011 season Galactinol levels exhibited a relationship with the tolerance 592
levels of the genotypes and levels of galactinol and raffinose negatively correlated to the 593
grain yield in DS and HS respectively Galactinol level was chosen for all models 594
explaining the genotypic variation of grain yield in all growth conditions by multiple linear 595
regression analysis Additionally the raffinose level under WW condition showed 596
correlation to grain yield in DS as well as myo-inositol These observations indicate a 597
close relationship between metabolism of myo-inositol and RFOs and yield performance 598
of maize under drought stress The negative correlation between levels of these 599
metabolites and grain yield under DS indicates that the yield performance is not due to 600
the osmoprotective functions of these compounds but rather to the metabolism of these 601
compounds One possible explanation is that the genotypes showing lower 602
accumulation of myo-inositol and raffinose synthesize higher degree RFOs such as 603
stachyose The pathways of RFO metabolism reconstructed from genomic information 604
revealed most of the key enzymes are encoded by multiple gene members with different 605
expression patterns (Zhou et al 2012) indicating the operation and importance of RFO 606
metabolism in maize RFOs can also serve as mobile and storage carbon sources with 607
advantages in osmolitic and mobile flexibility over sucrose and starch respectively (Van 608
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

28
den Ende 2013) Therefore it is also possible that the lower myo-inositol and raffinose 609
level in tolerant genotypes is due to their use of RFOs as carbon sources Altogether 610
metabolite profiles from field drought stress experiments indicate the importance of RFO 611
metabolism in yield performance under drought in field grown maize although the 612
mechanism underlying this remains to be investigated 613
Possible relationship between basal respiration and yield performance under the 614
drought and heat stress combination 615
Levels of three TCA cycle related metabolites in WW condition showed negative 616
correlation to grain yield in DS+HS stress combination Although this result should be 617
considered with special caution due to the uneven distribution of grain yield among 618
genotypes this is an interesting observation since these metabolites showed completely 619
different responses against each stress treatment It might be considerable that the 620
basal operation of TCA cycle is related to the yield performance under DS+HS and 621
therefore these metabolites can also be used as metabolic markers The down 622
regulation of respiratory pathways including TCA cycle is reported under DS+HS 623
condition in a previous Arabidopsis study (Prasch and Sonnewald 2013) supporting 624
this possibility 625
It should be noted that the correlation coefficient in our current study is relatively low 626
however this is most likely due to the fact that the data are highly variable since they 627
were obtained from field grown samples where the control of growth condition is difficult 628
There are some previously reported correlation analysis in field studies on metabolite 629
levels which corroborate this statement (Robinson et al 2007 Degenkolbe et al 630
2013) That said the coefficient values of the most correlations discussed here range 631
from -077 to -045 with p values lower than 005 or 0001 which can be considered as 632
highly reliable 633
634
Multiple metabolic features as biochemical markers 635
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

29
The use of multiple metabolites as biochemical markers is another possible way to 636
improve grain yield under stress conditions Indeed a strategy of employing multiple 637
markers has been proposed for molecular marker assisted breeding (Jannink et al 638
2010) The results of current study indicate the potentiality of this approach since the 639
combination of metabolites explained the variation of grain yield very well especially 640
under stress conditions in the multiple linear regression models The metabolic traits 641
showed performance in yield prediction superior to conventional agronomical 642
parameters which have shown to be correlated to grain yield suggesting the potential of 643
metabolite profiling in breeding program GC-MS based metabolite profiling is especially 644
promising due to its highthroughput robust nature and compatibility to analyse wide 645
range of primary metabolites (Obata and Fernie 2012) We employed the multiple linear 646
regression for model establishment in this study but other regression methods including 647
multi-level response analyses random forest model and correlation network analyses 648
should also be useful This study also indicated the potential of the metabolite levels in 649
WW to predict grain yield under stress conditions Interestingly galactinol was selected 650
in all models as the variable contributing to yield prediction As described above 651
galactinol functions as galactosyl donor especially for the synthesis of RFOs including 652
raffinose and stachyose (Loewus and Murthy 2000) The contribution of galactinol in 653
yield explanation emphasizes the importance of RFO metabolism in grain yield 654
performance 655
656
Conclusions 657
Metabolite profile of maize leaves from field DS HS and DS+HS combination trials was 658
analysed in the current study The metabolite profiling study using field samples is still 659
rare and this is the first study reporting metabolite responses to stress combination in 660
field grown crops making our results a good reference for future studies One of the 661
interesting findings is that the metabolic responses to DS+HS combination were rather 662
the sum of the effects by two individual stresses than novel or divergent effects This is 663
likely due to progressive nature of field stress treatments and needs to be considered as 664
a general trend under field conditions In contrast to phenotypic and transcriptomic 665
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

30
profiles monitored in previous studies (Witt et al 2012 Alexandersson et al 2014) 666
some typical stress responses of primary metabolism in the field grown plants are fairly 667
similar to those of greenhouse grown plants and seem well conserved between growth 668
conditions and even among species This robustness of metabolic change renders it a 669
good candidate for marker assisted breeding The metabolite profiling of field stress 670
samples successfully identified metabolite signatures closely related to grain yield under 671
abiotic stress conditions It highlights the importance of photorespiration and RFO 672
metabolism for yield performance under DS Especially myo-inositol and RFO levels are 673
quite promising metabolic markers for maize breeding since those in WW condition 674
were correlated to grain yield in DS allowing the selection under normal growth 675
condition There are some conventional high performance liquid chromatography 676
(HPLC) based methods available to analyse myo-inositol and RFOs and recent 677
optimization of high performance anion exchange chromatography coupled with pulsed 678
amperometric detection (HPAECndashPAD) would allow higher throughput analysis for 679
biochemical marker assisted breeding (Gangola et al 2014) Additionally multiple linear 680
regression analysis suggested the possible interplay between metabolic pathways in 681
stress tolerance and potential use of multiple metabolic markers for yield prediction 682
Further trials should be conducted to confirm the relationship between these metabolic 683
traits and yield performance under stress and to test the effectiveness of metabolites for 684
biochemical maker assisted breeding 685
686
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

31
Materials and Methods 687
Plant materials and experimental conditions 688
Ten maize lines were chosen based on their contrasting response to drought and 689
combined drought and heat stresses (Cairns et al 2013)(Table I) Single cross hybrids 690
were generated by crossing lines with the tropical tester CML-539 691
Experiments were conducted at the CIMMYT experimental station in Tlaltizapaacuten 692
Meacutexico (18deg41rsquoN 99deg07rsquoW 940 masl) A total of four experiments were planted each 693
year comprised of two different water and temperature regimes Optimal temperature 694
experiments were planted at the end of the wet season (late November) and higher 695
temperature experiments were planted at the start of the dry season (mid-February) 696
Due to the low latitude of the experimental station this experimental design facilitated 697
application of different temperature conditions without large effects on day length and 698
irradiation Two different water treatments were used at each temperature regime a 699
well-watered control and anthesis stage drought stress Drought stress was imposed by 700
stopping irrigation before flowering to ensure stress at anthesis In 2010 trials under 701
WW DS HS and DS+HS received a total of 1037 mm 520 mm 790 mm and 576 mm 702
of irrigation respectively In 2011 trials under WW DS HS and DS+HS received a total 703
of 1151 mm 550 mm 639 mm and 600 mm of irrigation respectively Rainfall 704
temperature data during experiments are presented in Table II Experiments were 705
planted in two-row plots with a final plant density of 667 plants m-2 An alpha-lattice 706
design was used replicated two times All plots received 80 kg N ha-1 (as urea) 80 kg P 707
ha-1 (as triple calcium superphosphate Ca (H2PO4)middot2H20) at sowing A second 708
application of N (80 kg N ha-1) was applied 5 weeks after sowing (V6 stage Ritchie et 709
al 1998) Recommended plant weed and insect control measures were used 710
711
Field measurements 712
Days to anthesis and silking were recorded when 50 of the plants had shed pollen 713
and 50 of the plants had silks respectively The ASI was calculated as days to silking 714
ndash days to anthesis At physiological maturity plant height was measured on two 715
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

32
representative plants per plot then all plants were hand harvested and grain yield 716
measured Grain weights were adjusted to 125 moisture content 717
718
Metabolite profiling 719
Metabolite profiling was performed as described in Witt et al (2012) Leaf blades were 720
harvested two weeks after the water stress was applied at the flowering stage Samples 721
were collected from six individual plants from each plot and treated as biological 722
replicates Materials were snap frozen in liquid nitrogen and liophilized for one week 723
One hundred milligram of lyophilized powder was used for gas chromatography-mass 724
spectrometry (GC-MS) based metabolite profiling following the protocol of Lisec et al 725
(2006) Peaks were manually annotated and ion intensity was determined by the aid of 726
TagFinder (Luedemann et al 2012) using a reference library derived from Golm-727
Metabolome-Database (GMD Kopka et al 2005) The parameters used for peak 728
annotation were shown in the Supplemental Table S9 following the recommended 729
reporting format (Fernie et al 2011) 730
731
Data analysis 732
Metabolite levels were represented by the observed ion intensity of a selected unique 733
ion Ion intensity was log10 transformed and normalized using an ANOVA based model 734
for removal of measurement bias (Lisec et al 2011) The normalized dataset used for 735
the analysis was shown in Supplemental Table S10 736
Statistical analyses and graphical representations (ANOVA Tukeyrsquos HSD test heatmap 737
hierarchical clustering Bonferroni-correction PCA boxplots Venn diagram correlation 738
analysis Studentrsquos t-test) were performed using the R-software environment 311 739
(httpcranr-project org) The box plots were drawn by ggplot function in ggplot2 740
package The PCA was conducted after pareto-normalization using the rnipals-741
algorithm of the pcaMethods package (Stacklies et al 2007) ANOVA was conducted 742
using the anova-function on linear models incorporating either genotype and treatment 743
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

33
or HS and DS status as factors and allowing interactions Resulting p-values were 744
corrected for multiple testing using the stringent Bonferroni method Tukey HSD test 745
was conducted by glht function in multcomp package Correlation was tested by cortest 746
function and scatter plots were drawn by ggplot function Venn diagram was drawn 747
using venndiagram function in VennDiagram package Effects of stress combination on 748
metabolite levels were predicted from response in single stresses as follows Response 749
factors were calculated by dividing metabolite levels transformed to linear values and 750
divided by those from well-watered plants grown in the same field replicate followed by 751
log2 transformation Predicted response factors in DS+HS condition are the sum of 752
those in DS and HS conditions The values calculated from each genotype and field 753
was considered as a replicate (n=20) and used for the calculation of the mean and SEM 754
Correlation between actual and predicted response factors were tested by cor test 755
function in R Further the metabolic and agronomic data set was subjected to multiple 756
linear regression analysis to ascertain which combination of metabolites alone or 757
metabolites plus agronomic traits fully explained differences in grain yield across the 758
whole set of genotypes and replications within each growing condition Models were 759
selected using ldquoLinear regressionrdquo function of SPSS (SPSS Inc Chicago IL USA) with 760
ldquoEntrerdquo method to gain a R2 value approaching 1000 Multiple linear regression analysis 761
using the levels of metabolites under WW as independent variables and grain yield 762
under the three different stress conditions as dependent variables were also performed 763
764
Acknowledgements 765
The authors appreciate the help of Ciro Sanchez for coordinating the field experiments 766
in Mexico 767
768
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

34
Figure legends 769
Figure 1 Yield related parameters in the two years of field stress trials Box plots of 770 grain yield (A) plant height (B) number of ears per plant (C) anthesis date (D) siliking 771 date (E) and interval between anthesis and siliking dates (F) in each 2010 and 2011 772 seasons Ten maize hybrids were grown in well watered (WW blue) drought stress (DS 773 orange) well watered and heat stress (HS green) and drought and heat stress (DS+HS 774 red) conditions in two independent plots (n=20) Letters indicate the results of Tukeyrsquos 775 test comparing the conditions in each year (plt005) 776
Figure 2 Heatmap of metabolic responses to stress conditions in each genotype 777 Metabolite levels in each stress condition were normalized to that in well watered 778 condition and log2 transformed Values are the means of up to 12 biological replicates 779 Red and blue colors represent increase and decrease of metabolite using a false-color 780 scale respectively Samples from drought stress (DS) drought and heat stress 781 (DS+HS) and well watered and heat stress (HS) are arranged from the left Genotypes 782 are ordered by the grain yield in the corresponding stress condition (smallest at the left) 783 in each year 784
Figure 3 Effects of treatments and genotypes on the levels of individual metabolites 785 Histograms show number of metabolites whose levels were altered with indicated p-786 values by Bonferroni-corrected two-way ANOVA analyzing the effects of genotypes (Gt) 787 and treatments (Tm) (A and C) or heat (HS) and drought (DS) stresses (B and D) Each 788 bar indicates a range of 005 Data from 2010 (A and B) and 2011 (C and D) were 789 independently analyzed Metabolite levels from six biological replicates in two 790 independent plots (n=12) were used for analysis 791
Figure 4 Foliar metabolite levels in the two years of field stress trials Box plots of 792 relative levels of selected metabolites in each 2010 and 2011 seasons Only the 793 metabolites discussed in the Discussion part are shown Plots for all metabolites are 794 shown in Supplemental Figure S3 Ten maize hybrids were grown in well watered (WW 795 blue) drought stress (DS orange) well watered and heat stress (HS green) and 796 drought and heat stress (DS+HS red) conditions in two independent plots (n=20) 797 Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 798 (plt005) 799
Figure 5 Correlation between metabolite levels and grain yield under drought (A) and 800 heat (B) stress conditions Levels of selected metabolites showed significant correlation 801 (plt0001 Table IV) were plotted against grain yield under stress conditions Ten maize 802 hybrids were grown in two independent plots (n=20 per year) and means of metabolite 803 levels from six biological replicates were plotted No metabolite showed significant 804 correlation to grain yield under simultaneous drought and heat stress Circles and 805
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

35
triangles indicate data from 2010 and 2011 respectively Due to a minor effect of heat 806 treatment on grain yield in 2011 only 2010 data was used for the analysis of heat stress 807 condition r and p are the correlation coefficient and p-value from Pearson correlation 808 analysis respectively 809
Figure 6 Correlation between metabolite levels in control condition and grain yield 810 under drought (A) and simultaneous drought and heat (B) stress conditions Levels of 811 selected metabolites under well watered condition showed significant correlation 812 (plt005 Table V) were plotted against grain yield under stress conditions No organic 813 metabolite showed significant correlation to grain yield under heat stress Mean values 814 from each of ten maize hybrid (n=10 per year) were plotted Circles and triangles 815 indicate data from 2010 and 2011 respectively Due to a minor effect of heat treatment 816 on grain yield in 2011 only 2010 data was used for the analysis of drought and heat 817 stress condition r and p are the correlation coefficient and p-value from Pearson 818 correlation analysis respectively 819
820
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

36
Table I Summary of maize genotypes Tolerance toa Genotype Entryb DS DS+HS HS La Posta Seq C7-F64-2-6-2-2 147 Tolerant Tolerant - DTPWC9-F31-1-3-1-1 102 Tolerant Tolerant Tolerant DTPYC9-F143-5-4-1-2 8 Moderate Tolerant Tolerant La Posta Seq C7-F18-3-2-1-1 37 Tolerant Moderate Tolerant CML-486 72 Moderate Tolerant Moderate La Posta Seq C7-F96-1-2-1-3 89 Susceptible Tolerant Tolerant CML311MBR C3 Bc F95-2-2-1 87 Susceptible Moderate Moderate CML311MBR C2 Bc F41-2 34 Tolerant Tolerant Susceptible DTPYC9-F69-3-5-1-1 91 Tolerant Susceptible Susceptible [GQL5[GQL5[MSRXPOOL9]C1F2-205-1(OSU23i)-5-3-X-X-1-BB]F2-4sx]-11-3-1-1
117 Moderate Susceptible Susceptible
aBased on Cairns et al (2013) bThe numbers used to identify the genotypes in this manuscript 821
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
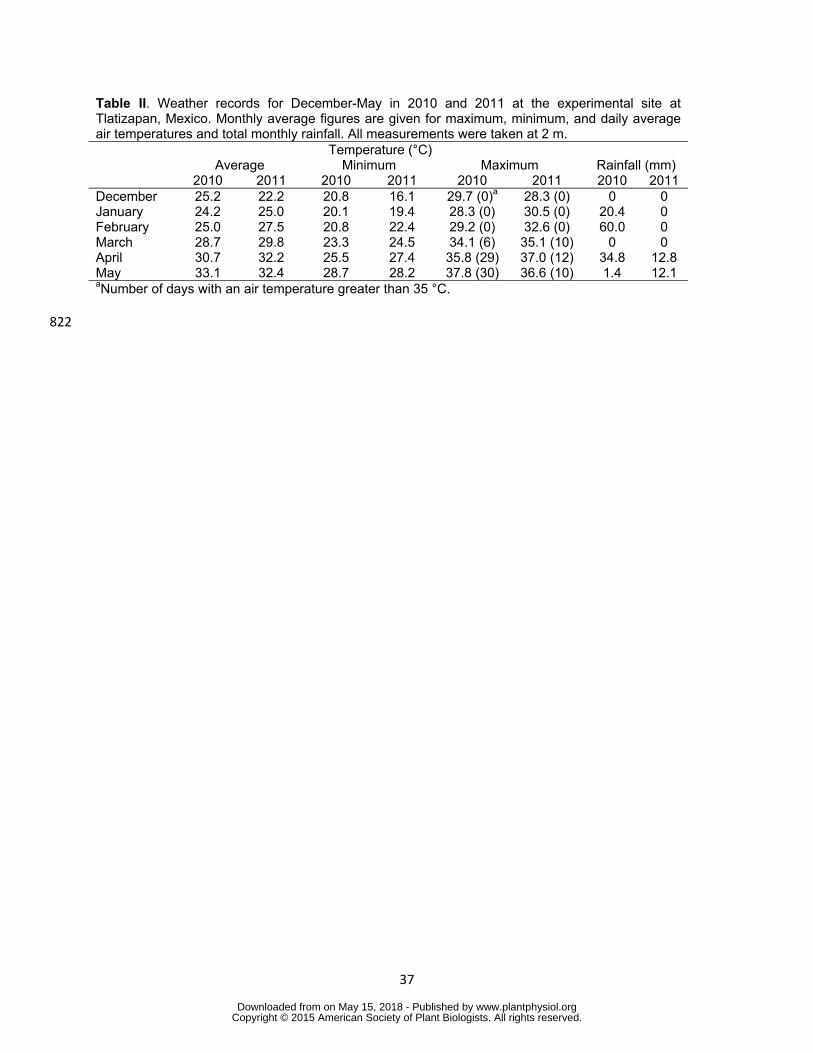
37
Table II Weather records for December-May in 2010 and 2011 at the experimental site at Tlatizapan Mexico Monthly average figures are given for maximum minimum and daily average air temperatures and total monthly rainfall All measurements were taken at 2 m Temperature (degC) Average Minimum Maximum Rainfall (mm) 2010 2011 2010 2011 2010 2011 2010 2011 December 252 222 208 161 297 (0)a 283 (0) 0 0 January 242 250 201 194 283 (0) 305 (0) 204 0 February 250 275 208 224 292 (0) 326 (0) 600 0 March 287 298 233 245 341 (6) 351 (10) 0 0 April 307 322 255 274 358 (29) 370 (12) 348 128 May 331 324 287 282 378 (30) 366 (10) 14 121 aNumber of days with an air temperature greater than 35 degC
822
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
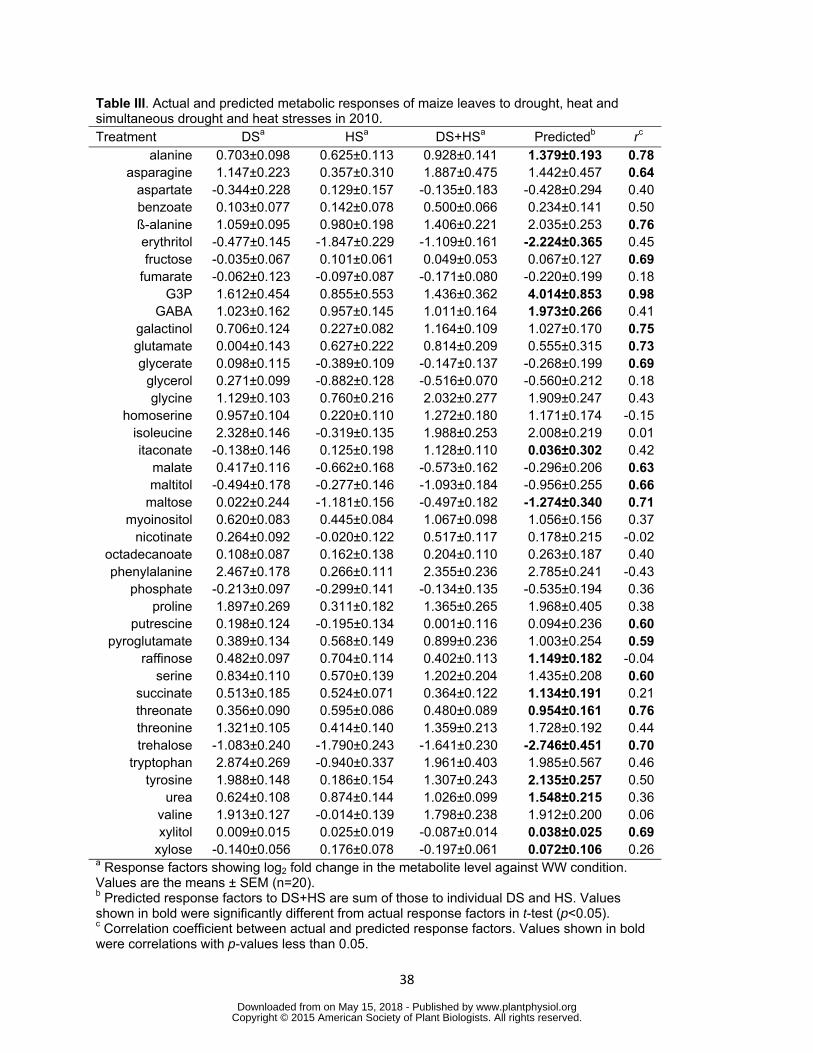
38
Table III Actual and predicted metabolic responses of maize leaves to drought heat and simultaneous drought and heat stresses in 2010 Treatment DSa HSa DS+HSa Predictedb rc
alanine 0703plusmn0098 0625plusmn0113 0928plusmn0141 1379plusmn0193 078asparagine 1147plusmn0223 0357plusmn0310 1887plusmn0475 1442plusmn0457 064
aspartate -0344plusmn0228 0129plusmn0157 -0135plusmn0183 -0428plusmn0294 040benzoate 0103plusmn0077 0142plusmn0078 0500plusmn0066 0234plusmn0141 050szlig-alanine 1059plusmn0095 0980plusmn0198 1406plusmn0221 2035plusmn0253 076erythritol -0477plusmn0145 -1847plusmn0229 -1109plusmn0161 -2224plusmn0365 045fructose -0035plusmn0067 0101plusmn0061 0049plusmn0053 0067plusmn0127 069
fumarate -0062plusmn0123 -0097plusmn0087 -0171plusmn0080 -0220plusmn0199 018G3P 1612plusmn0454 0855plusmn0553 1436plusmn0362 4014plusmn0853 098
GABA 1023plusmn0162 0957plusmn0145 1011plusmn0164 1973plusmn0266 041galactinol 0706plusmn0124 0227plusmn0082 1164plusmn0109 1027plusmn0170 075glutamate 0004plusmn0143 0627plusmn0222 0814plusmn0209 0555plusmn0315 073glycerate 0098plusmn0115 -0389plusmn0109 -0147plusmn0137 -0268plusmn0199 069
glycerol 0271plusmn0099 -0882plusmn0128 -0516plusmn0070 -0560plusmn0212 018glycine 1129plusmn0103 0760plusmn0216 2032plusmn0277 1909plusmn0247 043
homoserine 0957plusmn0104 0220plusmn0110 1272plusmn0180 1171plusmn0174 -015isoleucine 2328plusmn0146 -0319plusmn0135 1988plusmn0253 2008plusmn0219 001itaconate -0138plusmn0146 0125plusmn0198 1128plusmn0110 0036plusmn0302 042
malate 0417plusmn0116 -0662plusmn0168 -0573plusmn0162 -0296plusmn0206 063maltitol -0494plusmn0178 -0277plusmn0146 -1093plusmn0184 -0956plusmn0255 066
maltose 0022plusmn0244 -1181plusmn0156 -0497plusmn0182 -1274plusmn0340 071myoinositol 0620plusmn0083 0445plusmn0084 1067plusmn0098 1056plusmn0156 037
nicotinate 0264plusmn0092 -0020plusmn0122 0517plusmn0117 0178plusmn0215 -002octadecanoate 0108plusmn0087 0162plusmn0138 0204plusmn0110 0263plusmn0187 040phenylalanine 2467plusmn0178 0266plusmn0111 2355plusmn0236 2785plusmn0241 -043
phosphate -0213plusmn0097 -0299plusmn0141 -0134plusmn0135 -0535plusmn0194 036proline 1897plusmn0269 0311plusmn0182 1365plusmn0265 1968plusmn0405 038
putrescine 0198plusmn0124 -0195plusmn0134 0001plusmn0116 0094plusmn0236 060pyroglutamate 0389plusmn0134 0568plusmn0149 0899plusmn0236 1003plusmn0254 059
raffinose 0482plusmn0097 0704plusmn0114 0402plusmn0113 1149plusmn0182 -004serine 0834plusmn0110 0570plusmn0139 1202plusmn0204 1435plusmn0208 060
succinate 0513plusmn0185 0524plusmn0071 0364plusmn0122 1134plusmn0191 021threonate 0356plusmn0090 0595plusmn0086 0480plusmn0089 0954plusmn0161 076threonine 1321plusmn0105 0414plusmn0140 1359plusmn0213 1728plusmn0192 044trehalose -1083plusmn0240 -1790plusmn0243 -1641plusmn0230 -2746plusmn0451 070
tryptophan 2874plusmn0269 -0940plusmn0337 1961plusmn0403 1985plusmn0567 046tyrosine 1988plusmn0148 0186plusmn0154 1307plusmn0243 2135plusmn0257 050
urea 0624plusmn0108 0874plusmn0144 1026plusmn0099 1548plusmn0215 036valine 1913plusmn0127 -0014plusmn0139 1798plusmn0238 1912plusmn0200 006xylitol 0009plusmn0015 0025plusmn0019 -0087plusmn0014 0038plusmn0025 069
xylose -0140plusmn0056 0176plusmn0078 -0197plusmn0061 0072plusmn0106 026a Response factors showing log2 fold change in the metabolite level against WW condition Values are the means plusmn SEM (n=20) b Predicted response factors to DS+HS are sum of those to individual DS and HS Values shown in bold were significantly different from actual response factors in t-test (plt005) c Correlation coefficient between actual and predicted response factors Values shown in bold were correlations with p-values less than 005
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
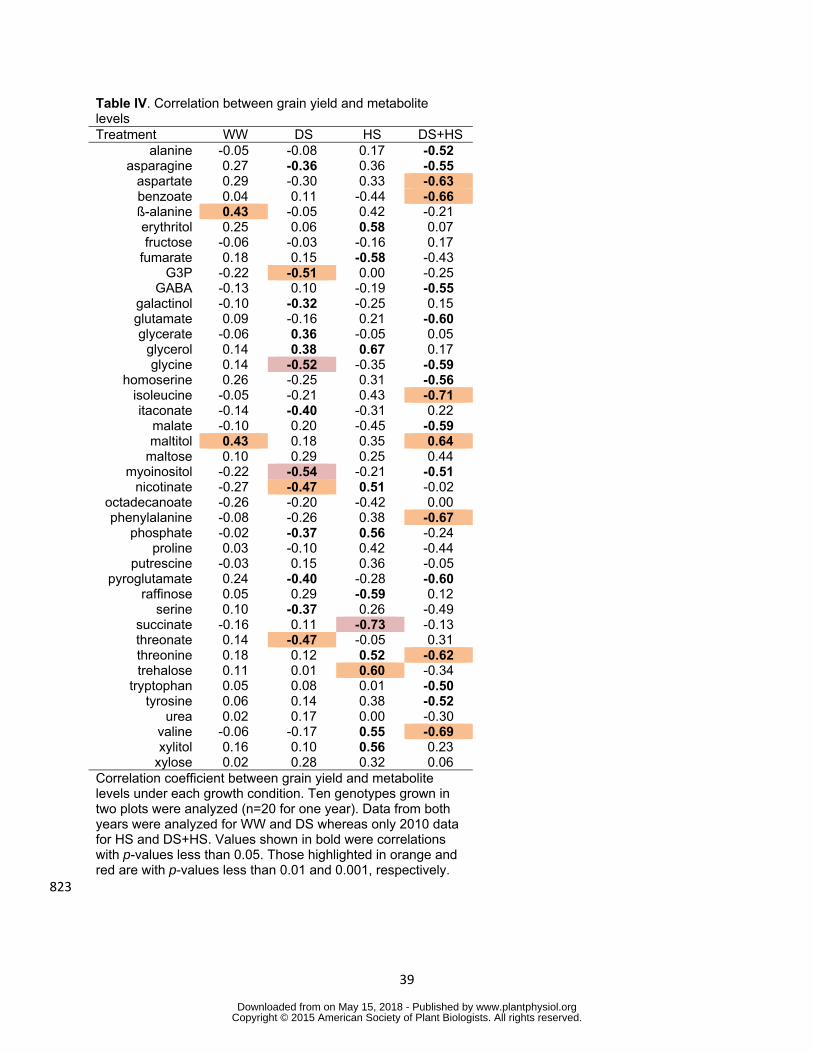
39
Table IV Correlation between grain yield and metabolite levels Treatment WW DS HS DS+HS
alanine -005 -008 017 -052 asparagine 027 -036 036 -055
aspartate 029 -030 033 -063 benzoate 004 011 -044 -066 szlig-alanine 043 -005 042 -021 erythritol 025 006 058 007 fructose -006 -003 -016 017
fumarate 018 015 -058 -043 G3P -022 -051 000 -025
GABA -013 010 -019 -055 galactinol -010 -032 -025 015 glutamate 009 -016 021 -060 glycerate -006 036 -005 005
glycerol 014 038 067 017 glycine 014 -052 -035 -059
homoserine 026 -025 031 -056 isoleucine -005 -021 043 -071 itaconate -014 -040 -031 022
malate -010 020 -045 -059 maltitol 043 018 035 064
maltose 010 029 025 044 myoinositol -022 -054 -021 -051
nicotinate -027 -047 051 -002 octadecanoate -026 -020 -042 000 phenylalanine -008 -026 038 -067
phosphate -002 -037 056 -024 proline 003 -010 042 -044
putrescine -003 015 036 -005 pyroglutamate 024 -040 -028 -060
raffinose 005 029 -059 012 serine 010 -037 026 -049
succinate -016 011 -073 -013 threonate 014 -047 -005 031 threonine 018 012 052 -062 trehalose 011 001 060 -034
tryptophan 005 008 001 -050 tyrosine 006 014 038 -052
urea 002 017 000 -030 valine -006 -017 055 -069 xylitol 016 010 056 023
xylose 002 028 032 006 Correlation coefficient between grain yield and metabolite levels under each growth condition Ten genotypes grown in two plots were analyzed (n=20 for one year) Data from both years were analyzed for WW and DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively 823
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
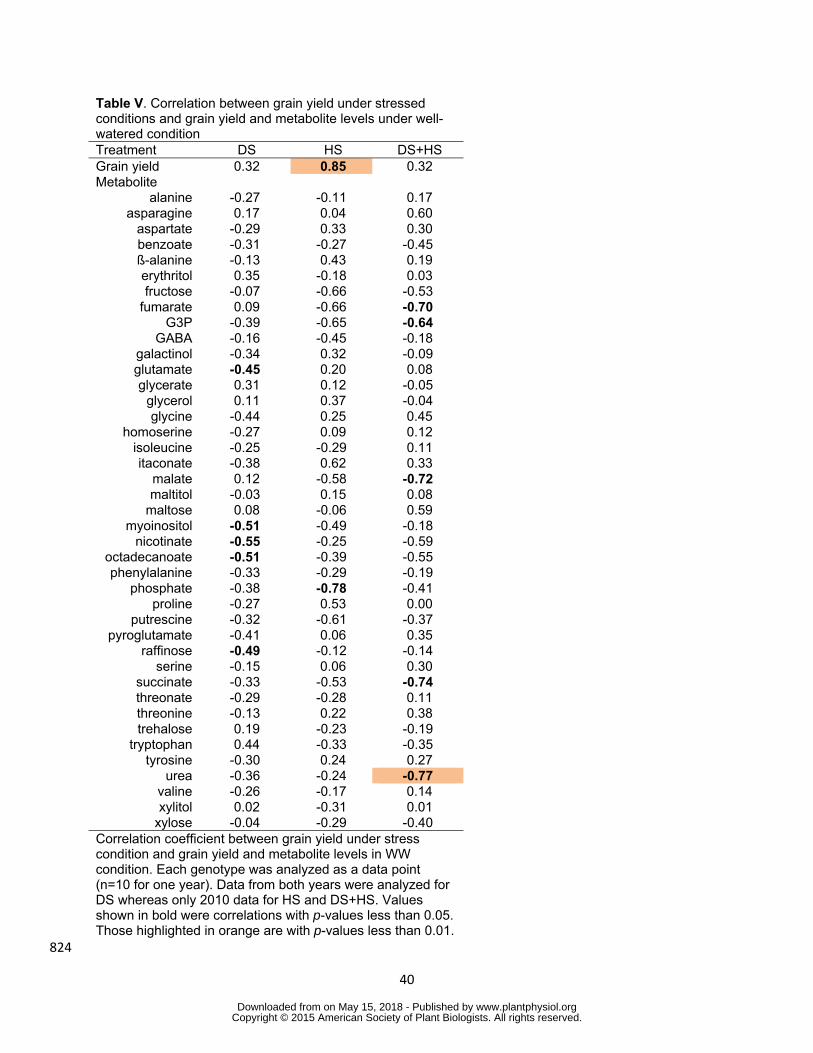
40
Table V Correlation between grain yield under stressed conditions and grain yield and metabolite levels under well-watered condition Treatment DS HS DS+HS Grain yield 032 085 032 Metabolite
alanine -027 -011 017 asparagine 017 004 060
aspartate -029 033 030 benzoate -031 -027 -045 szlig-alanine -013 043 019 erythritol 035 -018 003 fructose -007 -066 -053
fumarate 009 -066 -070 G3P -039 -065 -064
GABA -016 -045 -018 galactinol -034 032 -009 glutamate -045 020 008 glycerate 031 012 -005
glycerol 011 037 -004 glycine -044 025 045
homoserine -027 009 012 isoleucine -025 -029 011 itaconate -038 062 033
malate 012 -058 -072 maltitol -003 015 008
maltose 008 -006 059 myoinositol -051 -049 -018
nicotinate -055 -025 -059 octadecanoate -051 -039 -055 phenylalanine -033 -029 -019
phosphate -038 -078 -041 proline -027 053 000
putrescine -032 -061 -037 pyroglutamate -041 006 035
raffinose -049 -012 -014 serine -015 006 030
succinate -033 -053 -074 threonate -029 -028 011 threonine -013 022 038 trehalose 019 -023 -019
tryptophan 044 -033 -035 tyrosine -030 024 027
urea -036 -024 -077 valine -026 -017 014 xylitol 002 -031 001
xylose -004 -029 -040 Correlation coefficient between grain yield under stress condition and grain yield and metabolite levels in WW condition Each genotype was analyzed as a data point (n=10 for one year) Data from both years were analyzed for DS whereas only 2010 data for HS and DS+HS Values shown in bold were correlations with p-values less than 005 Those highlighted in orange are with p-values less than 001 824
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
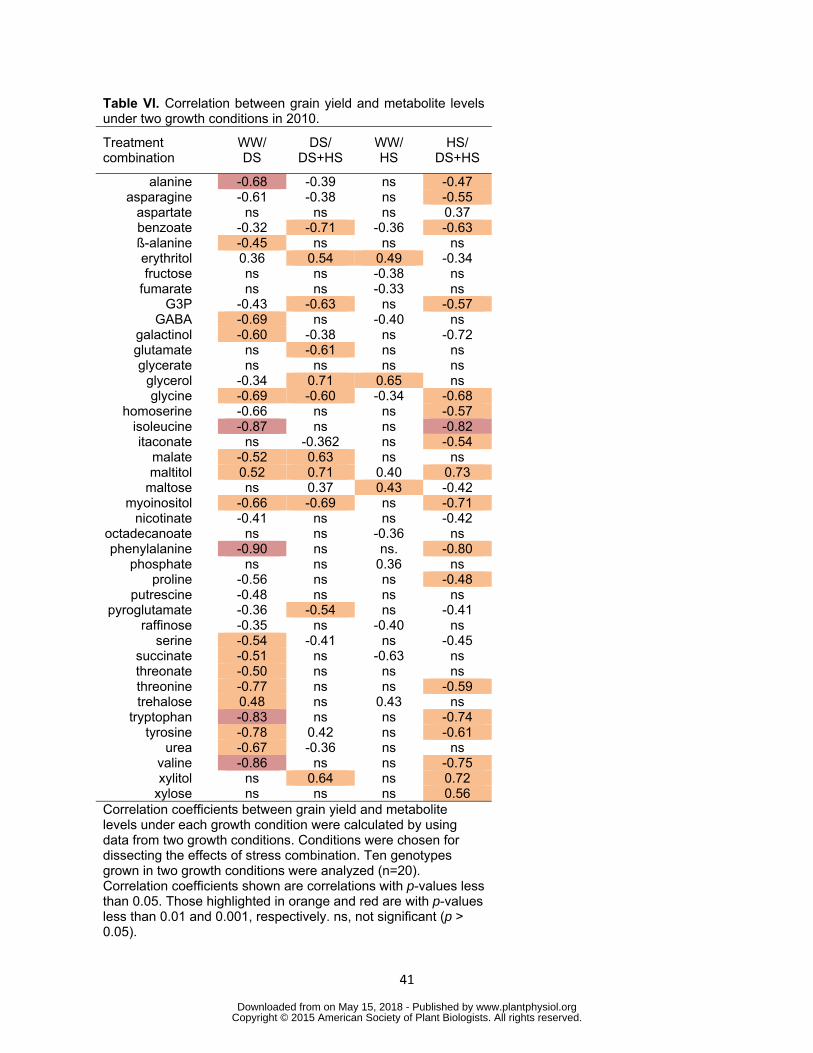
41
Table VI Correlation between grain yield and metabolite levels under two growth conditions in 2010
Treatment combination
WW DS
DS DS+HS
WW HS
HS DS+HS
alanine -068 -039 ns -047 asparagine -061 -038 ns -055
aspartate ns ns ns 037 benzoate -032 -071 -036 -063 szlig-alanine -045 ns ns ns erythritol 036 054 049 -034 fructose ns ns -038 ns
fumarate ns ns -033 ns G3P -043 -063 ns -057
GABA -069 ns -040 ns galactinol -060 -038 ns -072 glutamate ns -061 ns ns glycerate ns ns ns ns
glycerol -034 071 065 ns glycine -069 -060 -034 -068
homoserine -066 ns ns -057 isoleucine -087 ns ns -082 itaconate ns -0362 ns -054
malate -052 063 ns ns maltitol 052 071 040 073
maltose ns 037 043 -042 myoinositol -066 -069 ns -071
nicotinate -041 ns ns -042 octadecanoate ns ns -036 ns phenylalanine -090 ns ns -080
phosphate ns ns 036 ns proline -056 ns ns -048
putrescine -048 ns ns ns pyroglutamate -036 -054 ns -041
raffinose -035 ns -040 ns serine -054 -041 ns -045
succinate -051 ns -063 ns threonate -050 ns ns ns threonine -077 ns ns -059 trehalose 048 ns 043 ns
tryptophan -083 ns ns -074 tyrosine -078 042 ns -061
urea -067 -036 ns ns valine -086 ns ns -075 xylitol ns 064 ns 072
xylose ns ns ns 056 Correlation coefficients between grain yield and metabolite levels under each growth condition were calculated by using data from two growth conditions Conditions were chosen for dissecting the effects of stress combination Ten genotypes grown in two growth conditions were analyzed (n=20) Correlation coefficients shown are correlations with p-values less than 005 Those highlighted in orange and red are with p-values less than 001 and 0001 respectively ns not significant (p gt 005)
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
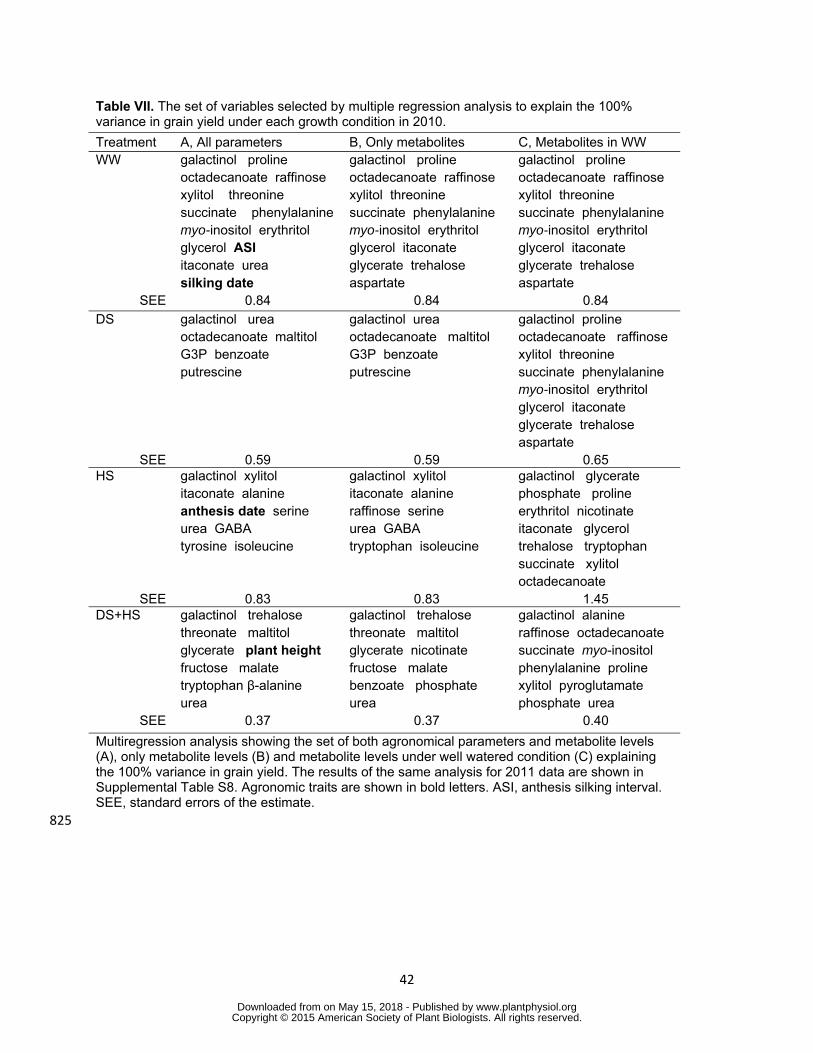
42
Table VII The set of variables selected by multiple regression analysis to explain the 100 variance in grain yield under each growth condition in 2010 Treatment A All parameters B Only metabolites C Metabolites in WW WW galactinol proline
octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol ASI itaconate urea silking date
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 084 084 084 DS galactinol urea
octadecanoate maltitol G3P benzoate putrescine
galactinol urea octadecanoate maltitol G3P benzoate putrescine
galactinol proline octadecanoate raffinose xylitol threonine succinate phenylalanine myo-inositol erythritol glycerol itaconate glycerate trehalose aspartate
SEE 059 059 065 HS galactinol xylitol
itaconate alanine anthesis date serine urea GABA tyrosine isoleucine
galactinol xylitol itaconate alanine raffinose serine urea GABA tryptophan isoleucine
galactinol glycerate phosphate proline erythritol nicotinate itaconate glycerol trehalose tryptophan succinate xylitol octadecanoate
SEE 083 083 145 DS+HS galactinol trehalose
threonate maltitol glycerate plant height fructose malate tryptophan β-alanine urea
galactinol trehalose threonate maltitol glycerate nicotinate fructose malate benzoate phosphate urea
galactinol alanine raffinose octadecanoate succinate myo-inositol phenylalanine proline xylitol pyroglutamate phosphate urea
SEE 037 037 040 Multiregression analysis showing the set of both agronomical parameters and metabolite levels (A) only metabolite levels (B) and metabolite levels under well watered condition (C) explaining the 100 variance in grain yield The results of the same analysis for 2011 data are shown in Supplemental Table S8 Agronomic traits are shown in bold letters ASI anthesis silking interval SEE standard errors of the estimate 825
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

43
Supplemental Figures and Tables 826
Supplemental Table S1 Effects of treatments and genotypes on the yield parameters 827
Supplemental Table S2 Effect of treatments on yield parameters of the subsets of 828 maize genotypes grouped by stress tolerance 829
Supplemental Table S3 Genotype and treatment effects on metabolite levels in each 830 year 831
Supplemental Table S4 Effect of treatments on metabolite levels of the subsets of 832 maize genotypes grouped by stress tolerance 833
Supplemental Table S5 Actual and predicted metabolic responses of maize leaves to 834 drought heat and simultaneous drought and heat stresses in 2011 835
Supplemental Table S6 Correlation between grain yield and metabolite levels under 836 each growth condition in each year 837
Supplemental Table S7 Correlation between grain yield and metabolite levels under 838 two growth conditions in 2011 839
Supplemental Table S8 The set of variables selected by multiregression analysis to 840 explain the 100 variance in grain yield under each growth condition in 2011 841
Supplemental Table S9 (xlsx) Parameters used for peak annotation in GC-MS 842 analysis 843
Supplemental Table S10 (csv) Normalized dataset used for the statistical analysis 844
845
846
Supplemental Figure S1 (PDF) Yield related parameters in the two years of field 847 stress trials Grain yield plant height number of ears per plant anthesis date silking 848 date and interval between anthesis and silking dates in each 2010 and 2011 seasons 849 Ten maize hybrids were grown in well watered (WW blue) drought stress (DS orange) 850 well watered and heat stress (HS green) and drought and heat stress (DS+HS red) 851 conditions in two independent field replicates and the data from each of them was 852 plotted Genotypes are ordered according to the grain yield under each stress treatment 853 in previous field trials (Cairns et al 2013) The letters following entry number of 854 genotype represents the tolerant level described in Table I t tolerant m moderate s 855 susceptible 856
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

44
Supplemental Figure S2 Principal component analysis (PCA) of metabolite profiles 857 under field stress conditions PCA score plot of foliar metabolite profiles of ten maize 858 genotypes subjected to well-watered (WW blue circle) drought stress (DS orange 859 square) heat stress (HS green triangle) and simultaneous drought and heat (DS+HS 860 red triangle) conditions in 2010 (A) and 2011 (B) field trials Percentage of the variance 861 captured by each principal component (PC) is given close to each respective axis Each 862 point represents one biological sample 863
Supplemental Figure S3 (PDF) Foliar metabolite levels in the two years of field stress 864 trials Box plots of relative levels of all metabolites in each 2010 and 2011 seasons Ten 865 maize hybrids were grown in well watered (WW blue) drought stress (DS orange) well 866 watered and heat stress (HS green) and drought and heat stress (DS+HS red) 867 conditions in two independent plots (A) boxplots showing metabolite levels under each 868 growth condition Mean values from each plotgenotype were treated as replicates 869 (n=20) Letters indicate the results of Tukeyrsquos test comparing the conditions in each year 870 (plt005) (B) boxplots showing metabolite levels in each genotype under each growth 871 condition The result from an individual sample was treated as a replicate Genotypes 872 are ordered according to the grain yield under each stress treatment in previous field 873 trials (Cairns et al 2013) The letters following entry number of genotype represents the 874 tolerant level described in Table I t tolerant m moderate s susceptible 875
Supplemental Figure S4 Number of metabolites which altered its level by either heat 876 drought or simultaneous heat and drought treatments in 2010 (A) and 2011 (B) field 877 trials Venn diagram showing number of metabolites which altered levels in a stress 878 condition significantly different from that in the well-watered condition in Tukeyrsquos test 879 (plt005 Supplemental Figure S3) 880
Supplemental Figure S5 Clustering of metabolites according to the responses to 881 drought heat and simultaneous drought and heat treatments in the 2010 trial All 882 detected metabolites were clustered into five groups by k-means clustering function in 883 MultiExperiment Viewer (httpwwwtm4orgmev Saeed et al 2003) under the 884 Pearson correlation coefficient The number of clusters was chosen by both rule of 885 thumb (N=41) and elbow method Means of metabolite levels were centered and plotted 886 for each of the well-watered (WW) drought stress (DS) heat stress (HS) and 887 simultaneous drought and heat (DS+HS) treatments Metabolites included in each 888 cluster are indicated in the plots Gray and magenta lines show the metabolite levels of 889 individual metabolites and average of those in the cluster 890
891
892
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

Parsed CitationsAlexandersson E Jacobson D Vivier MA Weckwerth W Andreasson E (2014) Field-omics-understanding large-scale moleculardata from field crops Front Plant Sci 5 286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Arauacutejo WL Tohge T Ishizaki K Leaver CJ Fernie AR (2011) Protein degradation - an alternative respiratory substrate for stressedplants Trends Plant Sci 16 489-498
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Cairns JE (2014) Field high-throughput phenotyping the new crop breeding frontier Trends Plant Sci 19 52-61Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Serret MD Edmeades GO (2012) Phenotyping maize for adaptation to drought Front Physiol 3 305Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Araus JL Slafer GA Royo C Serret MD (2008) Breeding for Yield Potential and Stress Adaptation in Cereals CRC Crit Rev PlantSci 27 377-412
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Araus J (2007) Recent advances in breeding maize for drought and salinity stress tolerance In MA Jenks PMHasegawa SM Jain eds Adv Mol Breed Towar Drought Salt Toler Crop Springer Netherlands Dordrecht pp 587-601
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Baumlnziger M Setimela PS Hodson D Vivek B (2006) Breeding for improved abiotic stress tolerance in maize adapted to southernAfrica Agric Water Manag 80 212-224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barchet GLH Dauwe R Guy RD Schroeder WR Soolanayakanahally RY Campbell MM Mansfield SD (2014) Investigating thedrought-stress response of hybrid poplar genotypes by metabolite profiling Tree Physiol 34 1203-1219
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barnaby JY Kim M Bauchan G Bunce J Reddy V Sicher RC (2013) Drought responses of foliar metabolites in three maizehybrids differing in water stress tolerance PLoS One 8 e77145
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bauwe H Hagemann M Fernie AR (2010) Photorespiration players partners and origin Trends Plant Sci 15 330-336Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benina M Obata T Mehterov N Ivanov I Petrov V Toneva V Fernie AR Gechev TS (2013) Comparative metabolic profiling ofHaberlea rhodopensis Thellungiella halophyla and Arabidopsis thaliana exposed to low temperature Front Plant Sci 4 499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Beyene Y Semagn K Mugo S Tarekegne A Babu R Meisel B Sehabiague P Makumbi D Magorokosho C Oikeh S et al (2015)Genetic gains in grain yield through genomic selection in eight bi-parental maize populations under drought stress Crop Sci 55154-163
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bohra A (2013) Emerging paradigms in genomics-based crop improvement ScientificWorldJournal 2013 585467Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bolger A Scossa F Bolger ME Lanz C Maumus F Tohge T Quesneville H Alseekh S Soslashrensen I Lichtenstein G et al (2014)The genome of the stress-tolerant wild tomato species Solanum pennellii Nat Genet 46 1034-1038
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bowne JB Erwin TA Juttner J Schnurbusch T Langridge P Bacic A Roessner U (2012) Drought responses of leaf tissues fromwheat cultivars of differing drought tolerance at the metabolite level Mol Plant 5 418-429
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bruce WB Edmeades GO Barker TC (2002) Molecular and physiological approaches to maize improvement for drought toleranceJ Exp Bot 53 13-25
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Brunk DG Rich PJ Rhodes D (1989) Genotypic variation for glycinebetaine among public inbreds of maize Plant Physiol 91 1122-1125
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cabrera-Bosquet L Crossa J von Zitzewitz J Serret MD Araus JL (2012) High-throughput phenotyping and genomic selectionthe frontiers of crop breeding converge J Integr Plant Biol 54 312-320
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Crossa J Zaidi PH Grudloyma P Sanchez C Araus JL Thaitad S Makumbi D Magorokosho C Baumlnziger M et al (2013)Identification of Drought Heat and Combined Drought and Heat Tolerant Donors in Maize Crop Sci 53 1335-1346
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sanchez C Vargas M Ordontildeez R Araus JL (2012a) Dissecting maize productivity ideotypes associated with grain yieldunder drought stress and well-watered conditions J Integr Plant Biol 54 1007-1020
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cairns JE Sonder K Zaidi PH Verhulst N Mahuku G Babu R Nair SK Das B Govaerts B Vinayan MT et al (2012b) Maizeproduction in a changing climate impacts adaptation and mitigation strategies Adv Agron 114 1-58
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chaves MM Flexas J Pinheiro C (2009) Photosynthesis under drought and salt stress regulation mechanisms from whole plant tocell Ann Bot 103 551-560
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cooper M Messina CD Podlich D Totir LR Baumgarten A Hausmann NJ Wright D Graham G (2014) Predicting the future ofplant breeding complementing empirical evaluation with genetic prediction Crop Pasture Sci 65 311-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Craufurd PQ Vadez V Jagadish SVK Prasad PVV Zaman-Allah M (2013) Crop science experiments designed to inform cropmodeling Agric For Meteorol 170 8-18
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Degenkolbe T Do PT Kopka J Zuther E Hincha DK Koumlhl KI (2013) Identification of drought tolerance markers in a diversepopulation of rice cultivars by expression and metabolite profiling PLoS One 8 e63637
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Easterling W Aggarwal P Batima P Brander K Erda L Howden M Kirilenko A Morton J Soussana J Schmidhuber J et al (2007)Food fibre and forest products In M Parry O Canziani J Palutikof P van der Lindin C Hanson eds Clim Chang 2007 ImpactsAdapt Vulnerability Cambridge University Press CambridgeUK pp 273-313
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Elsayed AI Rafudeen MS Golldack D (2014) Physiological aspects of raffinose family oligosaccharides in plants protection againstabiotic stress Plant Biol (Stuttg) 16 1-8
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

Van den Ende W (2013) Multifunctional fructans and raffinose family oligosaccharides Front Plant Sci 4 247Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Evers D Lefegravevre I Legay S Lamoureux D Hausman J-F Rosales ROG Marca LRT Hoffmann L Bonierbale M Schafleitner R(2010) Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metaboliteapproach J Exp Bot 61 2327-2343
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fait A Fromm H Walter D Galili G Fernie AR (2008) Highway or byway the metabolic role of the GABA shunt in plants TrendsPlant Sci 13 14-19
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
FAO (2014) FAO statistical database httpfaostat3faoorgPubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Aharoni A Willmitzer L Stitt M Tohge T Kopka J Carroll AJ Saito K Fraser PD DeLuca V (2011) Recommendations forreporting metabolite data Plant Cell 23 2477-2482
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fernie AR Schauer N (2009) Metabolomics-assisted breeding a viable option for crop improvement Trends Genet 25 39-48Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gangola MP Jaiswal S Khedikar YP Chibbar RN (2014) A reliable and rapid method for soluble sugars and RFO analysis inchickpea using HPAEC-PAD and its comparison with HPLC-RI Food Chem 154 127-133
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Geacutenard M Baldazzi V Gibon Y (2014) Metabolic studies in plant organs dont forget dilution by growth Front Plant Sci 5 85Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Giraud E Ho LHM Clifton R Carroll A Estavillo G Tan Y-F Howell KA Ivanova A Pogson BJ Millar AH et al (2008) The absenceof ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress Plant Physiol 147595-610
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gong Q Li P Ma S Indu Rupassara S Bohnert HJ (2005) Salinity stress adaptation competence in the extremophile Thellungiellahalophila in comparison with its relative Arabidopsis thaliana Plant J 44 826-839
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Grant RF Jackson BS Kiniry JR Arkin GF (1989) Water deficit timing effects on yield components in maize Agron J 81 61-65Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hanson AD Roje S (2001) One-carbon metabolism in higher plants Annu Rev Plant Physiol Plant Mol Biol 52 119-137Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hare PD Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants Plant Growth Regul 21 79-102Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hatzig S Zaharia LI Abrams S Hohmann M Legoahec L Bouchereau A Nesi N Snowdon RJ (2014) Early osmotic adjustmentresponses in drought-resistant and drought-sensitive oilseed rape J Integr Plant Biol 56 797-809
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hill CB Taylor JD Edwards J Mather D Langridge P Bacic A Roessner U (2015) Detection of QTL for metabolic and agronomic wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

traits in wheat with adjustments for variation at genetic loci that affect plant phenology Plant Sci 233 143-154Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iturriaga G Suaacuterez R Nova-Franco B (2009) Trehalose metabolism from osmoprotection to signaling Int J Mol Sci 10 3793-810Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jannink J-L Lorenz AJ Iwata H (2010) Genomic selection in plant breeding from theory to practice Brief Funct Genomics 9 166-177
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jones RJ Simmons SR (1983) Effect of altered source-sink ratio on growth of maize kernels Crop Sci 23 129-134Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Joshi V Joung J-G Fei Z Jander G (2010) Interdependence of threonine methionine and isoleucine metabolism in plantsaccumulation and transcriptional regulation under abiotic stress Amino Acids 39 933-947
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaur H Verma P Petla BP Rao V Saxena SC Majee M (2013) Ectopic expression of the ABA-inducible dehydration-responsivechickpea L-myo-inositol 1-phosphate synthase 2 (CaMIPS2) in Arabidopsis enhances tolerance to salinity and dehydration stressPlanta 237 321-335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Keles Y Oumlncel I (2002) Response of antioxidative defence system to temperature and water stress combinations in wheatseedlings Plant Sci 163 783-790
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Khodarahmpour Z Hamidi J (2011) Evaluation of drought tolerance in different growth stages of maize (Zea mays L) inbred linesusing tolerance indices African J Biotechnol 10 13482-13490
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kopka J Schauer N Krueger S Birkemeyer C Usadel B Bergmuumlller E Doumlrmann P Weckwerth W Gibon Y Stitt M et al (2005)GMDCSBDB the Golm Metabolome Database Bioinformatics 21 1635-1638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lawlor DW Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells a criticalevaluation of mechanisms and integration of processes Ann Bot 103 561-579
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Roumlmisch-Margl L Nikoloski Z Piepho H-P Giavalisco P Selbig J Gierl A Willmitzer L (2011) Corn hybrids display lowermetabolite variability and complex metabolite inheritance patterns Plant J 68 326-336
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lisec J Schauer N Kopka J Willmitzer L Fernie AR (2006) Gas chromatography mass spectrometry-based metabolite profiling inplants Nat Protoc 1 387-396
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Baumlnziger M Magorokosho C Vivek B (2011) Nonlinear heat effects on African maize as evidenced by historical yieldtrials Nat Clim Chang 1 42-45
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lobell DB Burke MB (2010) On the use of statistical models to predict crop yield responses to climate change Agric For Meteorol150 1443-1452
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

Loewus F Murthy PPN (2000) myo-Inositol metabolism in plants Plant Sci 150 1-19Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lopes MS Araus JL van Heerden PDR Foyer CH (2011) Enhancing drought tolerance in C4 crops J Exp Bot 62 3135-3153Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luedemann A von Malotky L Erban A Kopka J (2012) TagFinder preprocessing software for the fingerprinting and the profilingof gas chromatography-mass spectrometry based metabolome analyses Methods Mol Biol 860 255-286
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ma T Wang J Zhou G Yue Z Hu Q Chen Y Liu B Qiu Q Wang Z Zhang J et al (2013) Genomic insights into salt adaptation in adesert poplar Nat Commun 4 2797
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maurino VG Peterhansel C (2010) Photorespiration current status and approaches for metabolic engineering Curr Opin PlantBiol 13 249-256
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Missihoun TD Schmitz J Klug R Kirch H-H Bartels D (2011) Betaine aldehyde dehydrogenase genes from Arabidopsis withdifferent sub-cellular localization affect stress responses Planta 233 369-382
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muumlller C Cramer W Hare WL Lotze-Campen H (2011) Climate change risks for African agriculture Proc Natl Acad Sci U S A 1084313-4315
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nishizawa A Yabuta Y Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidativedamage Plant Physiol 147 1251-1263
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Obata T Fernie AR (2012) The use of metabolomics to dissect plant responses to abiotic stresses Cell Mol life Sci 69 3225-3243Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Osakabe Y Yamaguchi-Shinozaki K Shinozaki K Tran L-SP (2014) ABA control of plant macroelement membrane transportsystems in response to water deficit and high salinity New Phytol 202 35-49
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasanna BM Araus JL Crossa J Cairns JE (2013) High-Throughput and Precision Phenotyping for Cereal Breeding Programs InPK Gupta RK Varshney eds Cereal Genomics II Springer Netherlands Dordrecht pp 341-374
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Prasch CM Sonnewald U (2013) Simultaneous application of heat drought and virus to Arabidopsis plants reveals significantshifts in signaling networks Plant Physiol 162 1849-1866
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rasmussen S Barah P Suarez-Rodriguez MC Bressendorff S Friis P Costantino P Bones AM Nielsen HB Mundy J (2013)Transcriptome responses to combinations of stresses in Arabidopsis Plant Physiol 161 1783-1794
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Redestig H Kusano M Ebana K Kobayashi M Oikawa A Okazaki Y Matsuda F Arita M Fujita N Saito K (2011) Exploringmolecular backgrounds of quality traits in rice by predictive models based on high-coverage metabolomics BMC Syst Biol 5 176
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

Riedelsheimer C Czedik-Eysenberg A Grieder C Lisec J Technow F Sulpice R Altmann T Stitt M Willmitzer L Melchinger AE(2012a) Genomic and metabolic prediction of complex heterotic traits in hybrid maize Nat Genet 44 217-220
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Riedelsheimer C Lisec J Czedik-Eysenberg A Sulpice R Flis A Grieder C Altmann T Stitt M Willmitzer L Melchinger AE (2012b)Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize Proc Natl Acad Sci U S A 1098872-8877
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ritchie J Singh U Godwin D Bowen W (1998) Cereal growth development and yield In GY Tsuji G Hoogenboom PK Thorntoneds Underst Options Agric Prod Springer Netherlands Dordrecht pp 79-98
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Mittler R (2002) The combined effect of drought stress and heat shock on gene expression in tobacco PlantPhysiol 130 1143-1151
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rizhsky L Liang H Shuman J Shulaev V Davletova S Mittler R (2004) When defense pathways collide The response ofArabidopsis to a combination of drought and heat stress Plant Physiol 134 1683-1696
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Robinson AR Ukrainetz NK Kang K-Y Mansfield SD (2007) Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trialsreveals strong environmental and weak genetic variation New Phytol 174 762-773
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Romano G Zia S Spreer W Sanchez C Cairns J Araus JL Muumlller J (2011) Use of thermography for high throughput phenotypingof tropical maize adaptation in water stress Comput Electron Agric 79 67-74
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saeed AI Sharov V White J Li J Liang W Bhagabati N Braisted J Klapa M Currier T Thiagarajan M et al (2003) TM4 a freeopen-source system for microarray data management and analysis Biotechniques 34 374-378
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez DH Pieckenstain FL Escaray F Erban A Kraemer U Udvardi MK Kopka J (2011) Comparative ionomics andmetabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesisPlant Cell Environ 34 605-617
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saacutenchez-Rodriacuteguez E Rubio-Wilhelmi M del M Riacuteos JJ Blasco B Rosales MAacute Melgarejo R Romero L Ruiz JM (2011) Ammoniaproduction and assimilation its importance as a tolerance mechanism during moderate water deficit in tomato plants J PlantPhysiol 168 816-823
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Saneoka H Nagasaka C Hahn DT Yang WJ Premachandra GS Joly RJ Rhodes D (1995) Salt tolerance of glycinebetaine-deficient and -containing maize lines Plant Physiol 107 631-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sangoi L Salvador RJ (1998) Maize susceptibility to drought at flowering a new approach to overcome the problem Ciecircncia Rural28 699-706
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sicher RC Barnaby JY (2012) Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites towater stress Physiol Plant 144 238-253
Pubmed Author and TitleCrossRef Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

Google Scholar Author Only Title Only Author and Title
Skirycz A De Bodt S Obata T De Clercq I Claeys H De Rycke R Andriankaja M Van Aken O Van Breusegem F Fernie AR et al(2010) Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves toprolonged mild osmotic stress Plant Physiol 152 226-244
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Inzeacute D (2010) More from less plant growth under limited water Curr Opin Biotechnol 21 197-203Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Skirycz A Vandenbroucke K Clauw P Maleux K De Meyer B Dhondt S Pucci A Gonzalez N Hoeberichts F Tognetti VB et al(2011) Survival and growth of Arabidopsis plants given limited water are not equal Nat Biotechnol 29 212-214
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sperdouli I Moustakas M (2012) Interaction of proline sugars and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress J Plant Physiol 169 577-585
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Spoljarevic M Agic D Lisjak M Gumze A Wilson ID Hancock JT Teklic T (2011) The relationship of proline content andmetabolism on the productivity of maize plants Plant Signal Behav 6 251-257
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Stacklies W Redestig H Scholz M Walther D Selbig J (2007) pcaMethods--a bioconductor package providing PCA methods forincomplete data Bioinformatics 23 1164-1167
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suguiyama VF Silva EA Meirelles ST Centeno DC Braga MR (2014) Leaf metabolite profile of the Brazilian resurrection plantBarbacenia purpurea Hook (Velloziaceae) shows two time-dependent responses during desiccation and recovering Front PlantSci 5 96
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Suzuki N Rivero RM Shulaev V Blumwald E Mittler R (2014) Abiotic and biotic stress combinations New Phytol 203 32-43Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tai HH Worrall K Pelletier Y De Koeyer D Calhoun LA (2014) Comparative metabolite profiling of Solanum tuberosum against sixwild Solanum species with Colorado potato beetle resistance J Agric Food Chem 62 9043-9055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Takeda S Matsuoka M (2008) Genetic approaches to crop improvement responding to environmental and population changesNat Rev Genet 9 444-457
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tardieu F (2012) Any trait or trait-related allele can confer drought tolerance just design the right drought scenario J Exp Bot 6325-31
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tohge T de Souza LP Fernie AR (2014) Genome-enabled plant metabolomics J Chromatogr B Analyt Technol Biomed Life Sci966 7-20
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tollenaar M Lee EA (2006) Dissection of physiological processes underlying grain yield in maize by examining geneticimprovement and heterosis Maydica (Italy) 51 399-408
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tsonev S Todorovska E Avramova V Kolev S Abu-Mhadi N Christov NK (2014) Genomics Assisted Improvement of Drought wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved
Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

Tolerance in Maize QTL Approaches Biotechnol Biotechnol Equip 23 1410-1413Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tuberosa R Salvi S (2006) Genomics-based approaches to improve drought tolerance of crops Trends Plant Sci 11 405-412Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Kurihara Y Seki M Shinozaki K (2010) Omics analyses of regulatory networks in plant abiotic stress responses CurrOpin Plant Biol 13 132-138
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Urano K Maruyama K Ogata Y Morishita Y Takeda M Sakurai N Suzuki H Saito K Shibata D Kobayashi M et al (2009)Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics Plant J 57 1065-1078
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Verkest A Byzova M Martens C Willems P Verwulgen T Slabbinck B Rombaut D Van de Velde J Vandepoele K Standaert E etal (2015) Selection for Improved Energy Use Efficiency and Drought Tolerance in Canola Results in Distinct Transcriptome andEpigenome Changes Plant Physiol (in press)
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vile D Pervent M Belluau M Vasseur F Bresson J Muller B Granier C Simonneau T (2012) Arabidopsis growth under prolongedhigh temperature and water deficit independent or interactive effects Plant Cell Environ 35 702-718
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Voss I Sunil B Scheibe R Raghavendra AS (2013) Emerging concept for the role of photorespiration as an important part ofabiotic stress response Plant Biol (Stuttg) 15 713-722
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wen W Araus JL Shah T Cairns J Mahuku G Baumlnziger M Torres JL Saacutenchez C Yan J (2011) Molecular Characterization of aDiverse Maize Inbred Line Collection and its Potential Utilization for Stress Tolerance Improvement Crop Sci 51 2569-2581
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wingler A Lea PJ Quick WP Leegood RC (2000) Photorespiration metabolic pathways and their role in stress protection PhilosTrans R Soc Lond B Biol Sci 355 1517-1529
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Witt S Galicia L Lisec J Cairns J Tiessen A Araus JL Palacios-Rojas N Fernie AR (2012) Metabolic and phenotypic responses ofgreenhouse-grown maize hybrids to experimentally controlled drought stress Mol Plant 5 401-417
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Wu H-J Zhang Z Wang J-Y Oh D-H Dassanayake M Liu B Huang Q Sun H-X Xia R Wu Y et al (2012) Insights into salt tolerancefrom the genome of Thellungiella salsuginea Proc Natl Acad Sci U S A 109 12219-12224
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zelitch I Schultes NP Peterson RB Brown P Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize innormal air Plant Physiol 149 195-204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou M-L Zhang Q Zhou M Sun Z-M Zhu X-M Shao J-R Tang Y-X Wu Y-M (2012) Genome-wide identification of genes involvedin raffinose metabolism in Maize Glycobiology 22 1775-1785
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zia S Romano G Spreer W Sanchez C Cairns J Araus JL Muumlller J (2013) Infrared Thermal Imaging as a Rapid Tool for IdentifyingWater-Stress Tolerant Maize Genotypes of Different Phenology J Agron Crop Sci 199 75-84
Pubmed Author and Title wwwplantphysiolorgon May 15 2018 - Published by Downloaded from
Copyright copy 2015 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon May 15 2018 - Published by Downloaded from Copyright copy 2015 American Society of Plant Biologists All rights reserved

![AN OBATA TYPE RESULT FOR THE FIRST …vassilev/CRLichObata.pdf2 S. IVANOV AND D. VASSILEV 1. Introduction The classical theorems of Lichnerowicz [29] and Obata [33] give correspondingly](https://img.pdfslide.us/doc/110x75/5f1062037e708231d448d66e/an-obata-type-result-for-the-first-vassilev-2-s-ivanov-and-d-vassilev-1-introduction.jpg)



























