Embed Size (px)
Citation preview

Topography of Spiral GanglionProjections to Cochlear Nucleus
During Postnatal Development in Cats
RUSSELL L. SNYDER* AND PATRICIA A. LEAKE
Epstein Laboratory, Department of Otolaryngology, University of California, San Francisco,San Francisco, California 94143-0526
ABSTRACTA fundamenntal organizational principle of the central auditory system is that virtually
all areas are tonotopically organized. However, we know very little about the timing ormechanisms that are responsible for the development of this organization. When cats areborn, their auditory nervous systems are extremely immature, and their hearing thresholdsare very high. Until postnatal days 7–10 (P7–10), cats have behavioral and physiologicalthresholds which are near or above the pain threshold for adults and also have poor frequencyselectivity. Physiological thresholds for auditory nerve fibers and cochlear nucleus neuronsare typically above 100–120 dB SPL (sound pressure level re 20 µPa). Three weeks later (atapproximately P31), the sensitivity and frequency discrimination (tuning) of these neuronsapproximate adult values. This study examines the development of the tonotopic projectionsfrom the spiral ganglion to the cochlear nucleus during the period in cat development in whichthe auditory system undergoes the transition from being essentially nonfunctional to havingadult-like function.
With the animals heavily anesthetized, the cochleas were surgically exposed in kittensranging in age from P6 to P45. Focal injections of Neurobiotin (NB) were made intoRosenthal’s canal, labeling a small cluster of cells in the spiral ganglion of each cochlea. Theprojections of these labeled cells were visualized as frequency-specific bands of labeled axonsand terminals in all major subdivisions of the cochlear nucleus. The thickness of these bands(i.e., the dimension of the bands orthogonal to the isofrequency representation and across thefrequency gradient) were measured and compared to similar projections in adults. As in adultcats, the thickness of the bands varied only slightly with the location of the injection site(frequency representation) over a range of 1–7 mm from the cochlear base (45–13 kHz).Moreover, band thickness did not vary significantlywith age. These data indicate that the tonotopicorganization of spiral ganglion projections to the cochlear nucleus is as precise in kittens as youngasP6 as it is in adults. J. Comp. Neurol. 384:293–311, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: tonotopic representation; auditory nerve; primary afferents; topographic maps
In almost every area of the auditory system, the neuronsare arrayed in an orderly topographic arrangement accord-ing to the frequencies to which they are most sensitive.This fundamental tonotopic organization is established atthe level of the sensory receptors (hair cells), which arearrayed along the cochlear spiral such that those sensitiveto high frequencies are located in the basal cochlea, andthose most sensitive to low frequencies are located in theapex. This tonotopic organization is maintained through-out the auditory system by a series of spatially restrictedprojections from each successive level of organization tothe next. The most thoroughly studied of these projectionsis the primary afferent projection from the spiral ganglionneurons in the cochlear to the cochlear nucleus (CN). Each
inner hair cell (IHC) is innervated by ten to 30 spiralganglion neurons situated at a specific location along thecochlear spiral (Keithley and Cronin-Schreiber, 1987).Spiral ganglion cells in adjacent cochlear sectors innervateadjacent hair cells in the organ of Corti peripherally, andcentrally, they innervate offset but overlapping regions of
Grant sponsor: NIDCD; Grant number: NS-R01-DC00160; Grant spon-sor: Hearing Research, Inc.*Correspondence to: Russell L. Snyder, Epstein Laboratory, Dept. of
Otolaryngology, Univ. of California, San Francisco, CA94143-0526.E-mail: [email protected] 14 October 1996; Revised 20 February 1997; Accepted 11 March
1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 384:293–311 (1997)
r 1997 WILEY-LISS, INC.

the CN forming bandsor laminae in each of the major CNsubdivisions, the anteroventral cochlear nucleus or AVCN,the posteroventral cochlear nucleus or PVCN, and thedorsal cochlear nucleus or DCN (Leake and Snyder, 1988,1989; Leake et al., 1992; Ryugo and May, 1993; Sando,1965; Snyder et al., 1997). Axons from more basal cochleasectors project more dorsally in each subdivision, whereasthose frommore apical sectors project more ventrally. Thisorderly arrangement of primary afferent projections formsthe basis of the well-known tonotopic organization of thesenuclear subdivisions (Rose et al., 1960; Osen, 1970; Nodaand Pirsig, 1974; Webster et al., 1978; Bourk et al., 1981;Feng and Vater, 1985; Snyder and Leake, 1988; Leake andSnyder, 1989; Snyder et al., 1997). The organization of theCN, the first and obligatory synaptic station in the centralauditory pathway, underlies the tonotopic organization ofthe rest of the central auditory system.Despite the detailed descriptions of the tonotopic organi-
zation of the primary afferent projections to the cochlearnucleus, very little information is available about when orhow this organization is achieved. The available datasuggest that it might become established relatively late,i.e., during the late prenatal and/or early postnatal period.At birth, the cat’s auditory system is relatively immatureand essentially nonfunctional. Behavioral thresholds areextremely high—greater than 110 dB SPL (sound pressurelevel re 20 µPa)—(Foss and Flottorp, 1974; Clements andKelly, 1978; Olmstead and Villablanca, 1980; Ehret andRomand, 1981). Ensemble and single unit physiologicalthresholds in the auditory nerve are also elevated. Walshet al. (1986) were unable to record auditory brainstemresponses (ABRs) by using 135 dB SPL clicks until afterpostnatal day 7 (P7), and in individual animals, ABRthresholds are often greater than 100 dB SPL until as lateas P10–P14. Thresholds of single auditory nerve fibers arecomparable to the behavioral thresholds, ranging from 100to 120 dB SPL during the 1st postnatal week (Kettner etal., 1985; Walsh and McGee, 1986), and decrease to adultlevels over the next 14 to 21 days.In addition to their high thresholds during the first 2
postnatal weeks, auditory nerve fibers and neurons in theCN discharge rhythmically in response to constant acous-tic stimulation (Carlier et al., 1975; Romand andMarty, 1975;Brugge and O’Connor, 1984; Walsh and McGee, 1986, 1987,1988). Acoustically driven responses become adult-like overthe course of the 2nd and 3rd postnatal weeks. Approxi-mately 90% of all recorded neurons respond rhythmicallyto constant stimulation until P7, and rhythmic spontane-ous activity in the interstimulus intervals persisted insome neurons until P22 (Walsh and McGee, 1987).During the 1st postnatal week, the cat auditory system
also shows poor frequency selectivity, which persists until
late in the 2nd postnatal week (Walsh and Romand, 1992).Although most fibers have some spontaneous activity asearly as P3, most of these spontaneous rates are below 10spikes/second (s/s) during the first 6 postnatal days (Walshand McGee, 1987). In contrast, the spontaneous activityobserved in adult auditory nerve fibers can exceed 100 s/swith most fibers having rates above 10 s/s (Liberman,1978; Kim and Molnar, 1972).During the 2nd and 3rd postnatal week, auditory nerve
fibers undergo a series of major developmental changesculminating in responses that are nearly adult-like. Thesechanges include increased sensitivity, increased frequencydiscrimination (tuning), decreased first spike latency, lossof rhythmic firing, increased dynamic range, and increaseddischarge synchronization (Brugge et al., 1978; Romand,1983; Kettener et al., 1985; Walsh and McGee, 1986, 1988;Brugge, 1988, 1992). With the exception of synchroniza-tion (phase-locking), most response properties of auditorynerve fibers are mature after the 4th postnatal week(P28–P32). However, phase-locking in primary afferentsmay remain immature up to P45 (Brugge et al., 1978), andthe cochlear nuclei do not reach their adult size untilapproximately 50 days after birth (Larsen, 1984).In summary, the cat auditory system can be character-
ized as altricial, i.e., developing relatively late in thepostnatal period. In this study, our goal was to describequantitatively the development of the tonotopic projec-tions of the primary afferents (the spiral ganglion neuronsof the cochlea) to the cochlear nucleus in kittens from P6 toP45. This encompasses the period over which the catauditory system makes the transition from being virtuallynonfunctional to having adult-like responses.Our hypothesis was that the primary afferent projec-
tions would show substantial developmental maturationin their topographic organization. We have examinedthese projections by making focal extracellular injectionsof the neural tracer, Neurobiotin (NB), into the spiralganglion of kittens and adults. The results of the presentquantitative study in kittens suggest that this topographicprecision is established very early. Spiral ganglion projec-tions appear to be at least as precise by P6 as they are inadults. The absolute thickness of the average P6 frequency-band lamina is actually less than that of the average adultlamina. However, if the thickness is normalized for theincrease in the size of the cochlear nucleus with age, themean normalized thickness of the P6 laminae is notsignificantly different from that of the adult laminae inany of the three major CN subdivisions. These resultsindicate that the tonotopic organization of the primaryafferent projections into each CN subdivision is fullydeveloped by the 6th postnatal day.
MATERIALS AND METHODS
Injections
Fifteen kittens ranging in age from P6 to P45 wereobtained from a closed colony maintained at the Univer-sity of California at San Francisco. All pregnancies weretimed and lasted for 64 to 68 days with an average of 65.76 2.2. Additionally, the five youngest animals had gesta-tion periods of 65 or 66 days. All animals were free ofmiddle ear disease as judged by direct examination of themiddle ear. Each animal was tranquilized with an intra-muscular injection of ketamine HCl (25 mg/kg) andacepromazine (0.2 mg/kg). An intravenous catheter was
Abbreviations
ABR auditory brainstem responseAN auditory nerveAVCN anteroventral cochlear nucleusCF characteristic frequencyCN cochlear nucleus (i.e., AVCN 1 PVCN 1 DCN)DCN dorsal cochlear nucleusHRP horseradish peroxidaseIHC inner hair cellNB neurobiotinPVCN posteroventral cochlear nucleusVCN ventral cochlear nucleus (i.e., AVCN 1 PVCN)
294 R.L. SNYDER AND P.A. LEAKE

inserted into the cephalic vein, and a surgical level ofanesthesia was induced and maintained by infusion ofsodium pentobarbital (20 mg/kg). The respiratory andheart rates were monitored throughout the experimentalprocedures. Body temperature was maintained by using awarm water recirculating blanket. In older kittens thehead was positioned and immobilized by using a mouth-bar head-holder mounted in a magnetic base. The auditorybulla on each side was surgically exposed and opened topermit access to the cochlea. The round window wasexcised to allow direct visualization of Rosenthal’s canal inthe hook region and lower basal turn of the cochlea and themost basal sectors of the spiral ganglion. Injections intomore apical portions of the spiral ganglion are possible butthey require extensive cochlear dissections. In older kit-tens, a small hole into Rosenthal’s canal was carefullymade through the bone of themodiolus by using the tip of a30-gauge needle as a curette. In younger kittens, the spiralganglion was injected directly through the thin and par-tially ossified bone of the modiolus.Thick-walled glass micropipettes were pulled, and the
tips were broken to a diameter of 10–30 µm. The pipetteswere mounted on the needle tip of a 1-µl microsyringe(Unimetrics), sealed in place by using melted dental wax,and filled with a solution of 5% NB, (Vector Laboratories,Burlingame, CA) dissolved in distilled water. A smallquantity of the vital stain Trypan blue (,0.5%) was addedto the clear NB solution so that injections could be visuallyconfirmed. The microsyringe was mounted in a microma-nipulator, and the pipette tip was then positioned over theopening in Rosenthal’s canal and inserted into the spiralganglion. A small quantity of the tracer (,0.1 µl) wasinjected manually. The tip of the pipette was left in placefor a minimum of 10 minutes after the injection.In most cases, two injections were made in each cochlea.
The location of each injection was recorded by capturing avideo image generated by a color video camera (PanasonicKS102) mounted on the beam splitter of an operatingmicroscope (Zeiss OPMI). Images were captured by a colorvideo-capture card (ComputerEyes/RT, Digital Vision,Dedham, MA) mounted within a Macintosh Quadra 800microcomputer. After injections were completed, the roundwindow of the cochlea was sealed with a small disk ofgelfilm (Upjohn) or Saranwrap (Dow-Corning). Themuscleand skin were sutured to close the wound. Anesthesia wasmaintained and the animals were given fluid supplementsvia the intravenous catheter for 3–5 hours.
Preparation of cochlear and brainstemspecimens for NB cytochemistry
and light microscopy
Cochleas were preserved in vivo in heavily anesthetizedanimals by gentle perilymphatic perfusion of mixed alde-hyde fixative (0.5% paraformaldehyde and 2.5% glutaralde-hyde in 0.1 M phosphate buffer at pH 7.4) through theround and oval windows. Transcardiac perfusions wereconducted by using 5% lactated Ringer’s solution followedby a histological fixative (0.5% paraformaldehyde, 2.5%glutaraldehyde, and 4% sucrose in 0.1 M phosphate bufferat pH 7.4). Following transcardiac perfusion, the brain andtemporal bones were removed.The otic capsule bone of each cochlea was carefully
thinned with a diamond dental burr until the stria vascu-laris was clearly visible throughout the spiral. The roundwindow was enlarged anteriorly, and the bone of the
modiolus proximal to the spiral ganglion was thinned. Thecochlea was then decalcified for 12–18 hours in 0.1 MEDTA. Following decalcification, additional microdissec-tion was performed to isolate the hook and lower basalcochlear turn containing the injection sites and adjacentorgan of Corti. These specimens were then agitated on arotator for 2 hours in 2% dimethylsulfoxide (DMSO), acryoprotectant, and were then rapidly frozen in Freon 22cooled with liquid nitrogen. These additional steps arenecessary due to the relative impermeability of the tissuesurrounding the bony spiral lamina and Rosenthal’s canal,which appears to prevent adequate penetration of cyto-chemical reagents. After freezing, the isolated cochlearturns were rinsed in phosphate buffer and placed inVECTASTAINtABC reagent solution from Vector Labora-tories for 12 hours at room temperature.Horseradish peroxidase (HRP) reaction product was
demonstrated by using 3,38 diaminobenzidine tetrahydro-chloride (DAB, Sigma Chemical Co., St. Louis, MO) sub-strate with cobalt intensification as follows. Specimenswere placed in Tris-HCl buffer (Sigma) for 15 minutes,treated for 30 minutes in 0.5% CoCl2 in Tris, again rinsedin Tris for 10 minutes, and transferred to phosphate buffer(0.1 M at pH 7.2). Preincubation for 30 minutes in 0.1%DAB dissolved in phosphate buffer (pH 7.0) was followedby incubation in two changes of the same DAB solutionwith the addition of 0.1% hydrogen peroxide for a total of1.5 hours. All steps were carried out with continuousagitation on a rotator.After cytochemical processing, specimens were dehy-
drated, embedded in epoxy resin, and successive half turnsof the cochlear spiral were removed and mounted in epoxyon a glass slide with the basilar membrane parallel to theslide. The length of basilar membrane wasmeasured alongthe tops of the pillar cells, and the distance was marked inincrements of 0.5 mm. Blocks were removed at 0.5-mmintervals throughout the hook and lower basal turn of thespecimen containing the injection sites. Each block wasmounted on blank epoxy cylinders, and sections (1–2 µmthick) were collected every 25 µm. Alternate samples werestained with Toluidine blue.
Quantitative analysis of NB labelingin the cochlea
Unstained radial sections were viewed by using a Zeissphotomicroscope III, and a video image was captured byusing a Cohumonocrome video camera in conjunction witha video frame capture card (DT2255, Data Translation,Marlboro, MA) mounted in a MacIntosh Quadra 800microcomputer. Spiral ganglion injection sites were recon-structed by examination of sequential radial sections.Every 50 µm, a representative section was selected inwhich an IHC was clearly visible, and an image of theorgan of Corti was captured at a final magnification of34,000. The surface area of labeled fibers and terminalswas determined by using IMAGE (NIH, Bethesda, MD)and plotted as a function of distance from the cochlearbase. The frequency range represented at the injection sitewas defined by the distribution of labeled neurons in theosseous spiral lamina, and as recorded from 1-µm radialsections. The position of labeled fibers was first defined byabsolute location in millimeters from the basal end of thecochlea. By using a standard average total basilar mem-brane length of 24 mm (Liberman, 1982), the location ofthe injection site was calculated as proportional total
COCHLEAR TOPOGRAPHIC PROJECTIONS 295

basilar membrane length (% distance from base). Theaverage adult cochlear length measured by using thistechnique is 23.6 6 0.76 mm, and the average basilarmembrane length in P0–P3 kittens (n 5 13) is 23.5 6 1.06mm (unpublished data). Thus, average total adult cochlealength determined by using our methods is comparable toLiberman’s standard average basilar membrane length,and the average total cochlear length in neonatal kittens isnot significantly different from that in adults. Theseresults are in good agreement with the report of Romandand Romand (1982) that shows no difference between thelength of the adult spiral lamina and that of the neonatalcat. Therefore, we feel relatively secure in inferring acorrespondence between cochlear location in kittens andcochlear location (CF) in adults. The frequencies repre-sented at the location of the labeled fibers were thencalculated by using Greenwood’s frequency-position func-tion (Greenwood, 1974) with the revised constants sug-gested by Liberman (1982).
Preparation and analysis of CN
After vascular perfusion the brainstem (rostral mid-brain to the caudal medulla) was isolated, rinsed inRinger’s solution, and placed in 40% sucrose in 0.1 Mphosphate buffer at pH 7.4 for 12 to 72 hours at 3°C. Theright side of the brain was marked, and serial frozensections were cut at a thickness of 50 µm in the coronalplane. Sections were washed in 3 changes of 0.1 Mphosphate buffer, pH 7.6, followed by 3 changes of 0.05 MTris buffer. Sections were incubated in VECTASTAINtABC reagent solution from Vector Laboratories for 12hours at 3°C and rinsed in 3 changes of 0.05 M Tris buffer.All sections were then placed in the substrate solution forthe demonstration of HRP—20 mg DAB, 1 mg cobaltchloride, 1 mg nickel ammonium sulfate in 100 ml Trisbuffer. After 30 minutes incubation, sections were trans-ferred to a fresh solution of the DAB substrate, to whichone drop of 3% hydrogen peroxide had been added. Sec-tions were incubated for an additional 30–60 minutes,then washed in 3 changes of Tris buffer and mounted ongelatinized slides. The sections were dehydrated in gradedalcohol, cleared in xylenes, and coverslipped.Serial sections of the cochlear nucleus were examined
with the light microscope, and video images were capturedand analyzed by using the same methods as describedabove. The magnification of each nuclear series was cali-brated by capturing an image of an objective micrometerscale and superimposing that scale on the first sectionimage. All images were printed with a laser printer.Nomenclature for the cytoarchitectonic subdivisions andcell types of the cochlear nucleus used in this studygenerally follow those of Osen (1969), since smaller subdi-visions defined by others in Golgi material (Brawer et al.,1974; Cant and Morest, 1984) could not always be identi-fied in the present unstained experimental material. Fordescriptions of fiber and terminalmorphology the terminol-ogy of Rouiller et al. (1986) was adopted. The widths of thelamina of labeled fibers/terminals were determined byusing the measuring tool in Image and extended orthogo-nal to the long axis of the lamina. The center-to-centerdistance separating laminae in double-injection cases wasdetermined in all sections in which both laminae could beidentified. In each section, lamina thickness and separa-tion was measured at representative locations (at least 2in the VCN and one in the DCN) across each projection
lamina. Locations were chosen to avoid measurementsacross unusual extensions, indentations, section artifacts,etc., within each lamina. The edges of each lamina at eachlocation were determined subjectively. At each selectedlocation, wemeasured the thickness of the lamina by usingthe measuring tool in IMAGE, starting and stopping eachmeasurement where the density of labeled axons de-creased precipitously. All measurements were averagedseparately for each section, and then an overall averagewas calculated in each nucleus (AVCN, PVCN, and DCN).Although other,more objective,methods tomeasure laminathickness were evaluated, such methods resulted in singlemeasurements that were within 10% of these subjectivemeasurements, and they were much more time consum-ing. The area of each entire cochlear nucleus was deter-mined at a level just posterior to the entrance of theauditory nerve. This value was used to normalize themeasurements of lamina thickness for differences innuclear size. The square root of the area was used as anestimate of mean coronal CN width/height and to convertthe units of the normalizing measure to match those of thethickness measure. The experiments described in thisstudy were conducted under protocols approved by theanimal care committee at the University of California atSan Francisco.
RESULTS
Injections sites and projectionsin the cochlea
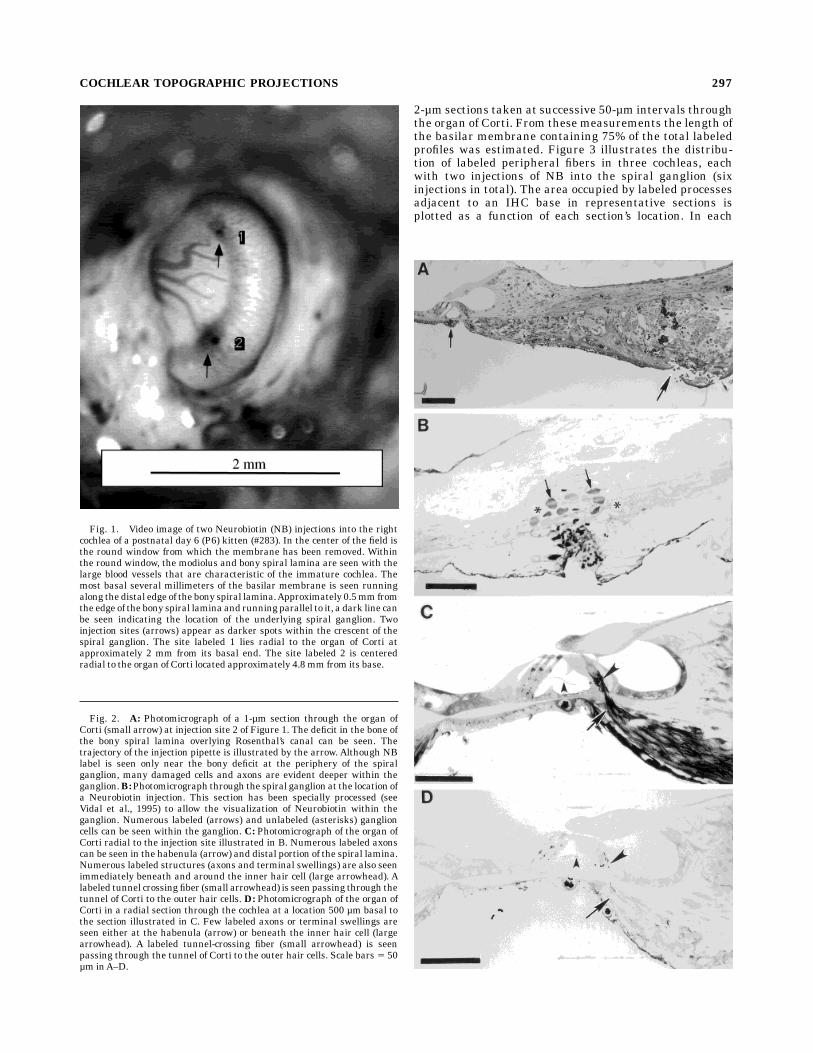
After removal of the round window membrane andaspiration of the perilymph, Rosenthal’s canal is identifi-able as a dark stripe of reduced reflectance in the bone ofthe osseous spiral lamina about 0.5 mm from the basilarmembrane. Spiral ganglion injections were made by inser-tion of a micropipette through the round window anddirectly into Rosenthal’s canal. With this approach, injec-tions can be made at locations up to 8 mm from theextreme base of the kitten cochlea, representing frequen-cies of 12 kHz to 60 kHz in adults. The location andrelative size of each injection were recorded as photo-graphic or video images taken through the operatingmicroscope (Fig. 1). When two injections were made in asingle cochlea the injection sites were always separated bya minimum of 1 mm. The depth and precise location ofeach micropipette penetration into Rosenthal’s canal wasdocumented by radial 1- to 2-µm sections of plastic embed-ded cochlear sectors (Fig. 2A,B).Peripheral to each injection site numerous labeled affer-
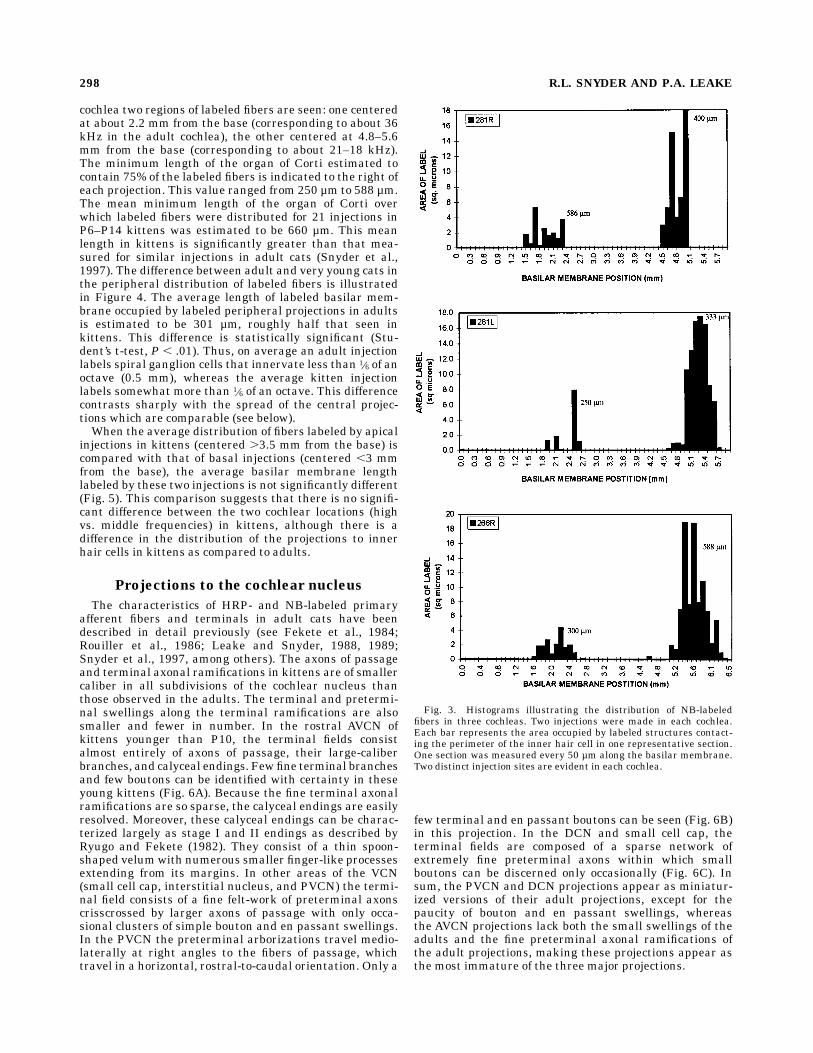
ent and efferent fibers were seen within the organ of Corti,habenula, and the peripheral portions of the bony spirallamina. Relatively few labeled processes were seen atmorecentral locations within the modiolus, presumably due topoor diffusion of the avidin–HRP complex into these areas.Within the organ of Corti, labeled fibers and terminalscould be seen beneath IHCs. Labeled fibers also were seencrossing the tunnel of Corti radially and running longitudi-nally along the cochlear spiral beneath the outer hair cells(Fig. 2C). The number of the labeled processes beneathIHCs was maximal in the regions nearest to the spiralganglion injection site and decreased in regions furtherapical and basal from the injection site (Fig. 2D).The peripheral distribution of labeled processes was
estimated by measuring the area occupied by all labeledprofiles immediately beneath IHCs in representative 1- to
296 R.L. SNYDER AND P.A. LEAKE
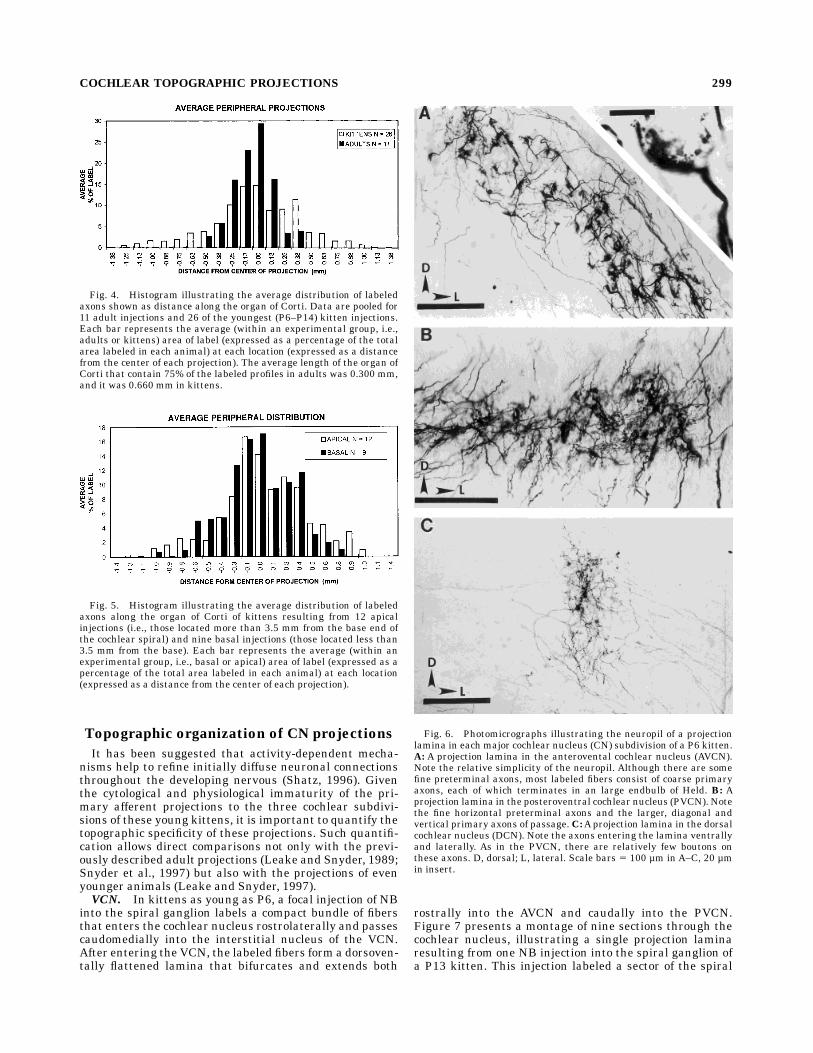
2-µm sections taken at successive 50-µm intervals throughthe organ of Corti. From these measurements the length ofthe basilar membrane containing 75% of the total labeledprofiles was estimated. Figure 3 illustrates the distribu-tion of labeled peripheral fibers in three cochleas, eachwith two injections of NB into the spiral ganglion (sixinjections in total). The area occupied by labeled processesadjacent to an IHC base in representative sections isplotted as a function of each section’s location. In each
Fig. 2. A: Photomicrograph of a 1-µm section through the organ ofCorti (small arrow) at injection site 2 of Figure 1. The deficit in the bone ofthe bony spiral lamina overlying Rosenthal’s canal can be seen. Thetrajectory of the injection pipette is illustrated by the arrow. Although NBlabel is seen only near the bony deficit at the periphery of the spiralganglion, many damaged cells and axons are evident deeper within theganglion.B:Photomicrograph through the spiral ganglionat the location ofa Neurobiotin injection. This section has been specially processed (seeVidal et al., 1995) to allow the visualization of Neurobiotin within theganglion. Numerous labeled (arrows) and unlabeled (asterisks) ganglioncells can be seen within the ganglion. C: Photomicrograph of the organ ofCorti radial to the injection site illustrated in B. Numerous labeled axonscan be seen in the habenula (arrow) and distal portion of the spiral lamina.Numerous labeled structures (axons and terminal swellings) are also seenimmediately beneath and around the inner hair cell (large arrowhead). Alabeled tunnel crossingfiber (small arrowhead) is seenpassing through thetunnel of Corti to the outer hair cells. D: Photomicrograph of the organ ofCorti in a radial section through the cochlea at a location 500 µm basal tothe section illustrated in C. Few labeled axons or terminal swellings areseen either at the habenula (arrow) or beneath the inner hair cell (largearrowhead). A labeled tunnel-crossing fiber (small arrowhead) is seenpassing through the tunnel of Corti to the outer hair cells. Scale bars 5 50µm inA–D.
Fig. 1. Video image of two Neurobiotin (NB) injections into the rightcochlea of a postnatal day 6 (P6) kitten (#283). In the center of the field isthe round window from which the membrane has been removed. Withinthe round window, the modiolus and bony spiral lamina are seen with thelarge blood vessels that are characteristic of the immature cochlea. Themost basal several millimeters of the basilar membrane is seen runningalong thedistal edge of thebony spiral lamina.Approximately 0.5mmfromthe edge of thebony spiral laminaand runningparallel to it, a dark line canbe seen indicating the location of the underlying spiral ganglion. Twoinjection sites (arrows) appear as darker spots within the crescent of thespiral ganglion. The site labeled 1 lies radial to the organ of Corti atapproximately 2 mm from its basal end. The site labeled 2 is centeredradial to the organ of Corti located approximately 4.8mm from its base.
COCHLEAR TOPOGRAPHIC PROJECTIONS 297
cochlea two regions of labeled fibers are seen: one centeredat about 2.2 mm from the base (corresponding to about 36kHz in the adult cochlea), the other centered at 4.8–5.6mm from the base (corresponding to about 21–18 kHz).The minimum length of the organ of Corti estimated tocontain 75% of the labeled fibers is indicated to the right ofeach projection. This value ranged from 250 µm to 588 µm.The mean minimum length of the organ of Corti overwhich labeled fibers were distributed for 21 injections inP6–P14 kittens was estimated to be 660 µm. This meanlength in kittens is significantly greater than that mea-sured for similar injections in adult cats (Snyder et al.,1997). The difference between adult and very young cats inthe peripheral distribution of labeled fibers is illustratedin Figure 4. The average length of labeled basilar mem-brane occupied by labeled peripheral projections in adultsis estimated to be 301 µm, roughly half that seen inkittens. This difference is statistically significant (Stu-dent’s t-test, P , .01). Thus, on average an adult injectionlabels spiral ganglion cells that innervate less than 1⁄6 of anoctave (0.5 mm), whereas the average kitten injectionlabels somewhat more than 1⁄6 of an octave. This differencecontrasts sharply with the spread of the central projec-tions which are comparable (see below).When the average distribution of fibers labeled by apical
injections in kittens (centered .3.5 mm from the base) iscompared with that of basal injections (centered ,3 mmfrom the base), the average basilar membrane lengthlabeled by these two injections is not significantly different(Fig. 5). This comparison suggests that there is no signifi-cant difference between the two cochlear locations (highvs. middle frequencies) in kittens, although there is adifference in the distribution of the projections to innerhair cells in kittens as compared to adults.
Projections to the cochlear nucleus
The characteristics of HRP- and NB-labeled primaryafferent fibers and terminals in adult cats have beendescribed in detail previously (see Fekete et al., 1984;Rouiller et al., 1986; Leake and Snyder, 1988, 1989;Snyder et al., 1997, among others). The axons of passageand terminal axonal ramifications in kittens are of smallercaliber in all subdivisions of the cochlear nucleus thanthose observed in the adults. The terminal and pretermi-nal swellings along the terminal ramifications are alsosmaller and fewer in number. In the rostral AVCN ofkittens younger than P10, the terminal fields consistalmost entirely of axons of passage, their large-caliberbranches, and calyceal endings. Fewfine terminal branchesand few boutons can be identified with certainty in theseyoung kittens (Fig. 6A). Because the fine terminal axonalramifications are so sparse, the calyceal endings are easilyresolved. Moreover, these calyceal endings can be charac-terized largely as stage I and II endings as described byRyugo and Fekete (1982). They consist of a thin spoon-shaped velumwith numerous smaller finger-like processesextending from its margins. In other areas of the VCN(small cell cap, interstitial nucleus, and PVCN) the termi-nal field consists of a fine felt-work of preterminal axonscrisscrossed by larger axons of passage with only occa-sional clusters of simple bouton and en passant swellings.In the PVCN the preterminal arborizations travel medio-laterally at right angles to the fibers of passage, whichtravel in a horizontal, rostral-to-caudal orientation. Only a
few terminal and en passant boutons can be seen (Fig. 6B)in this projection. In the DCN and small cell cap, theterminal fields are composed of a sparse network ofextremely fine preterminal axons within which smallboutons can be discerned only occasionally (Fig. 6C). Insum, the PVCN and DCN projections appear as miniatur-ized versions of their adult projections, except for thepaucity of bouton and en passant swellings, whereasthe AVCN projections lack both the small swellings of theadults and the fine preterminal axonal ramifications ofthe adult projections, making these projections appear asthe most immature of the three major projections.
Fig. 3. Histograms illustrating the distribution of NB-labeledfibers in three cochleas. Two injections were made in each cochlea.Each bar represents the area occupied by labeled structures contact-ing the perimeter of the inner hair cell in one representative section.One section was measured every 50 µm along the basilar membrane.Two distinct injection sites are evident in each cochlea.
298 R.L. SNYDER AND P.A. LEAKE
Topographic organization of CN projections
It has been suggested that activity-dependent mecha-nisms help to refine initially diffuse neuronal connectionsthroughout the developing nervous (Shatz, 1996). Giventhe cytological and physiological immaturity of the pri-mary afferent projections to the three cochlear subdivi-sions of these young kittens, it is important to quantify thetopographic specificity of these projections. Such quantifi-cation allows direct comparisons not only with the previ-ously described adult projections (Leake and Snyder, 1989;Snyder et al., 1997) but also with the projections of evenyounger animals (Leake and Snyder, 1997).VCN. In kittens as young as P6, a focal injection of NB
into the spiral ganglion labels a compact bundle of fibersthat enters the cochlear nucleus rostrolaterally and passescaudomedially into the interstitial nucleus of the VCN.After entering the VCN, the labeled fibers form a dorsoven-tally flattened lamina that bifurcates and extends both
rostrally into the AVCN and caudally into the PVCN.Figure 7 presents a montage of nine sections through thecochlear nucleus, illustrating a single projection laminaresulting from one NB injection into the spiral ganglion ofa P13 kitten. This injection labeled a sector of the spiral
Fig. 4. Histogram illustrating the average distribution of labeledaxons shown as distance along the organ of Corti. Data are pooled for11 adult injections and 26 of the youngest (P6–P14) kitten injections.Each bar represents the average (within an experimental group, i.e.,adults or kittens) area of label (expressed as a percentage of the totalarea labeled in each animal) at each location (expressed as a distancefrom the center of each projection). The average length of the organ ofCorti that contain 75% of the labeled profiles in adults was 0.300 mm,and it was 0.660 mm in kittens.
Fig. 5. Histogram illustrating the average distribution of labeledaxons along the organ of Corti of kittens resulting from 12 apicalinjections (i.e., those located more than 3.5 mm from the base end ofthe cochlear spiral) and nine basal injections (those located less than3.5 mm from the base). Each bar represents the average (within anexperimental group, i.e., basal or apical) area of label (expressed as apercentage of the total area labeled in each animal) at each location(expressed as a distance from the center of each projection).
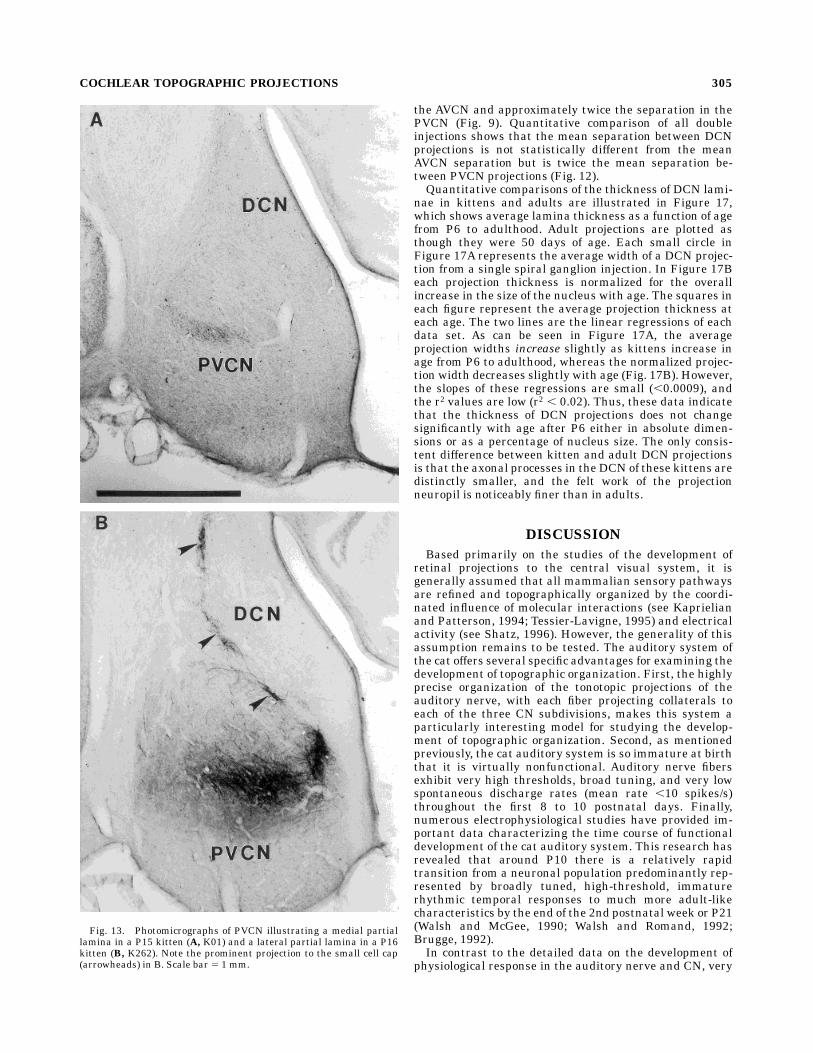
Fig. 6. Photomicrographs illustrating the neuropil of a projectionlamina in each major cochlear nucleus (CN) subdivision of a P6 kitten.A: A projection lamina in the anterovental cochlear nucleus (AVCN).Note the relative simplicity of the neuropil. Although there are somefine preterminal axons, most labeled fibers consist of coarse primaryaxons, each of which terminates in an large endbulb of Held. B: Aprojection lamina in the posteroventral cochlear nucleus (PVCN). Notethe fine horizontal preterminal axons and the larger, diagonal andvertical primary axons of passage.C:Aprojection lamina in the dorsalcochlear nucleus (DCN). Note the axons entering the lamina ventrallyand laterally. As in the PVCN, there are relatively few boutons onthese axons. D, dorsal; L, lateral. Scale bars 5 100 µm in A–C, 20 µmin insert.
COCHLEAR TOPOGRAPHIC PROJECTIONS 299
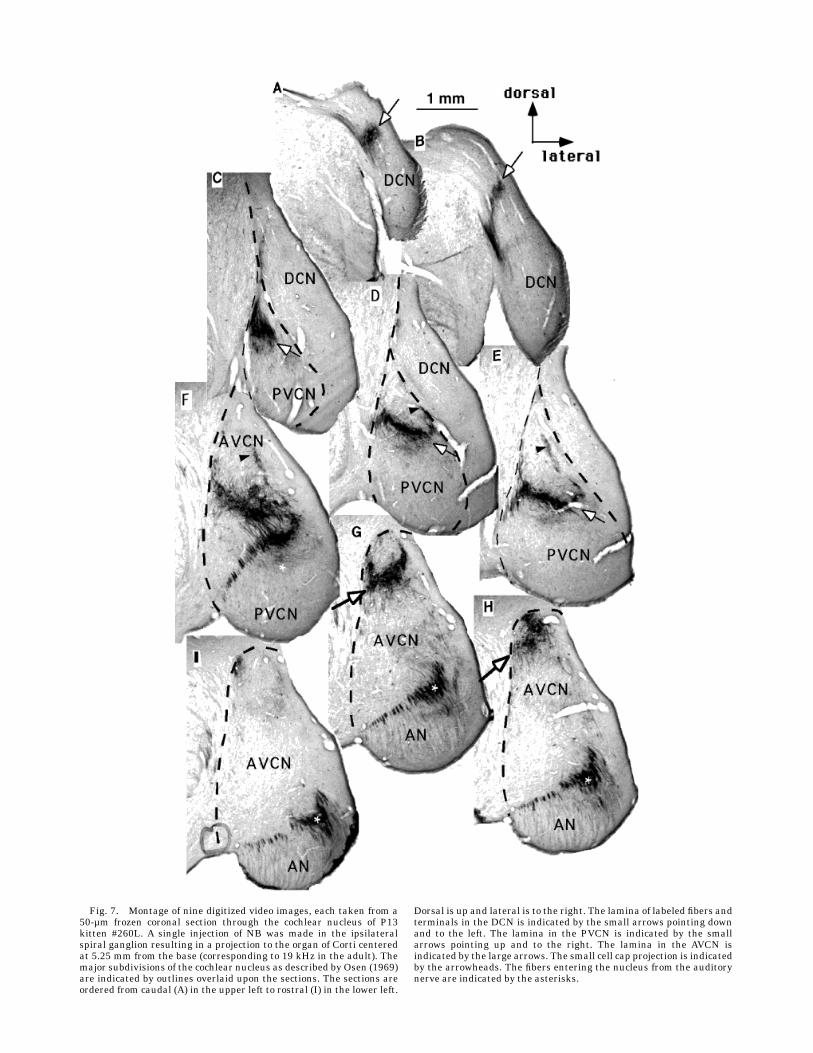
Fig. 7. Montage of nine digitized video images, each taken from a50-µm frozen coronal section through the cochlear nucleus of P13kitten #260L. A single injection of NB was made in the ipsilateralspiral ganglion resulting in a projection to the organ of Corti centeredat 5.25 mm from the base (corresponding to 19 kHz in the adult). Themajor subdivisions of the cochlear nucleus as described by Osen (1969)are indicated by outlines overlaid upon the sections. The sections areordered from caudal (A) in the upper left to rostral (I) in the lower left.
Dorsal is up and lateral is to the right. The lamina of labeled fibers andterminals in the DCN is indicated by the small arrows pointing downand to the left. The lamina in the PVCN is indicated by the smallarrows pointing up and to the right. The lamina in the AVCN isindicated by the large arrows. The small cell cap projection is indicatedby the arrowheads. The fibers entering the nucleus from the auditorynerve are indicated by the asterisks.
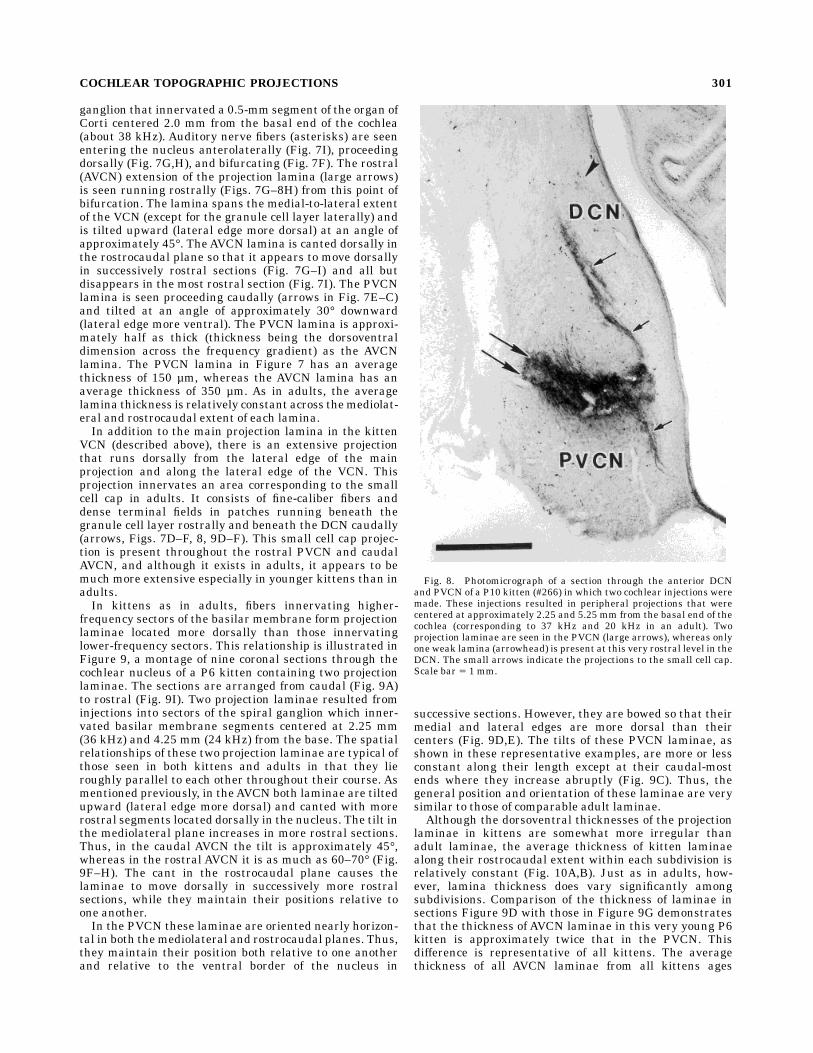
ganglion that innervated a 0.5-mm segment of the organ ofCorti centered 2.0 mm from the basal end of the cochlea(about 38 kHz). Auditory nerve fibers (asterisks) are seenentering the nucleus anterolaterally (Fig. 7I), proceedingdorsally (Fig. 7G,H), and bifurcating (Fig. 7F). The rostral(AVCN) extension of the projection lamina (large arrows)is seen running rostrally (Figs. 7G–8H) from this point ofbifurcation. The lamina spans the medial-to-lateral extentof the VCN (except for the granule cell layer laterally) andis tilted upward (lateral edge more dorsal) at an angle ofapproximately 45°. TheAVCN lamina is canted dorsally inthe rostrocaudal plane so that it appears to move dorsallyin successively rostral sections (Fig. 7G–I) and all butdisappears in the most rostral section (Fig. 7I). The PVCNlamina is seen proceeding caudally (arrows in Fig. 7E–C)and tilted at an angle of approximately 30° downward(lateral edge more ventral). The PVCN lamina is approxi-mately half as thick (thickness being the dorsoventraldimension across the frequency gradient) as the AVCNlamina. The PVCN lamina in Figure 7 has an averagethickness of 150 µm, whereas the AVCN lamina has anaverage thickness of 350 µm. As in adults, the averagelamina thickness is relatively constant across themediolat-eral and rostrocaudal extent of each lamina.In addition to the main projection lamina in the kitten
VCN (described above), there is an extensive projectionthat runs dorsally from the lateral edge of the mainprojection and along the lateral edge of the VCN. Thisprojection innervates an area corresponding to the smallcell cap in adults. It consists of fine-caliber fibers anddense terminal fields in patches running beneath thegranule cell layer rostrally and beneath the DCN caudally(arrows, Figs. 7D–F, 8, 9D–F). This small cell cap projec-tion is present throughout the rostral PVCN and caudalAVCN, and although it exists in adults, it appears to bemuch more extensive especially in younger kittens than inadults.In kittens as in adults, fibers innervating higher-
frequency sectors of the basilar membrane form projectionlaminae located more dorsally than those innervatinglower-frequency sectors. This relationship is illustrated inFigure 9, a montage of nine coronal sections through thecochlear nucleus of a P6 kitten containing two projectionlaminae. The sections are arranged from caudal (Fig. 9A)to rostral (Fig. 9I). Two projection laminae resulted frominjections into sectors of the spiral ganglion which inner-vated basilar membrane segments centered at 2.25 mm(36 kHz) and 4.25 mm (24 kHz) from the base. The spatialrelationships of these two projection laminae are typical ofthose seen in both kittens and adults in that they lieroughly parallel to each other throughout their course. Asmentioned previously, in theAVCN both laminae are tiltedupward (lateral edge more dorsal) and canted with morerostral segments located dorsally in the nucleus. The tilt inthe mediolateral plane increases in more rostral sections.Thus, in the caudal AVCN the tilt is approximately 45°,whereas in the rostral AVCN it is as much as 60–70° (Fig.9F–H). The cant in the rostrocaudal plane causes thelaminae to move dorsally in successively more rostralsections, while they maintain their positions relative toone another.In the PVCN these laminae are oriented nearly horizon-
tal in both themediolateral and rostrocaudal planes. Thus,they maintain their position both relative to one anotherand relative to the ventral border of the nucleus in
successive sections. However, they are bowed so that theirmedial and lateral edges are more dorsal than theircenters (Fig. 9D,E). The tilts of these PVCN laminae, asshown in these representative examples, are more or lessconstant along their length except at their caudal-mostends where they increase abruptly (Fig. 9C). Thus, thegeneral position and orientation of these laminae are verysimilar to those of comparable adult laminae.Although the dorsoventral thicknesses of the projection
laminae in kittens are somewhat more irregular thanadult laminae, the average thickness of kitten laminaealong their rostrocaudal extent within each subdivision isrelatively constant (Fig. 10A,B). Just as in adults, how-ever, lamina thickness does vary significantly amongsubdivisions. Comparison of the thickness of laminae insections Figure 9D with those in Figure 9G demonstratesthat the thickness of AVCN laminae in this very young P6kitten is approximately twice that in the PVCN. Thisdifference is representative of all kittens. The averagethickness of all AVCN laminae from all kittens ages
Fig. 8. Photomicrograph of a section through the anterior DCNand PVCN of a P10 kitten (#266) in which two cochlear injections weremade. These injections resulted in peripheral projections that werecentered at approximately 2.25 and 5.25 mm from the basal end of thecochlea (corresponding to 37 kHz and 20 kHz in an adult). Twoprojection laminae are seen in the PVCN (large arrows), whereas onlyone weak lamina (arrowhead) is present at this very rostral level in theDCN. The small arrows indicate the projections to the small cell cap.Scale bar 5 1 mm.
COCHLEAR TOPOGRAPHIC PROJECTIONS 301
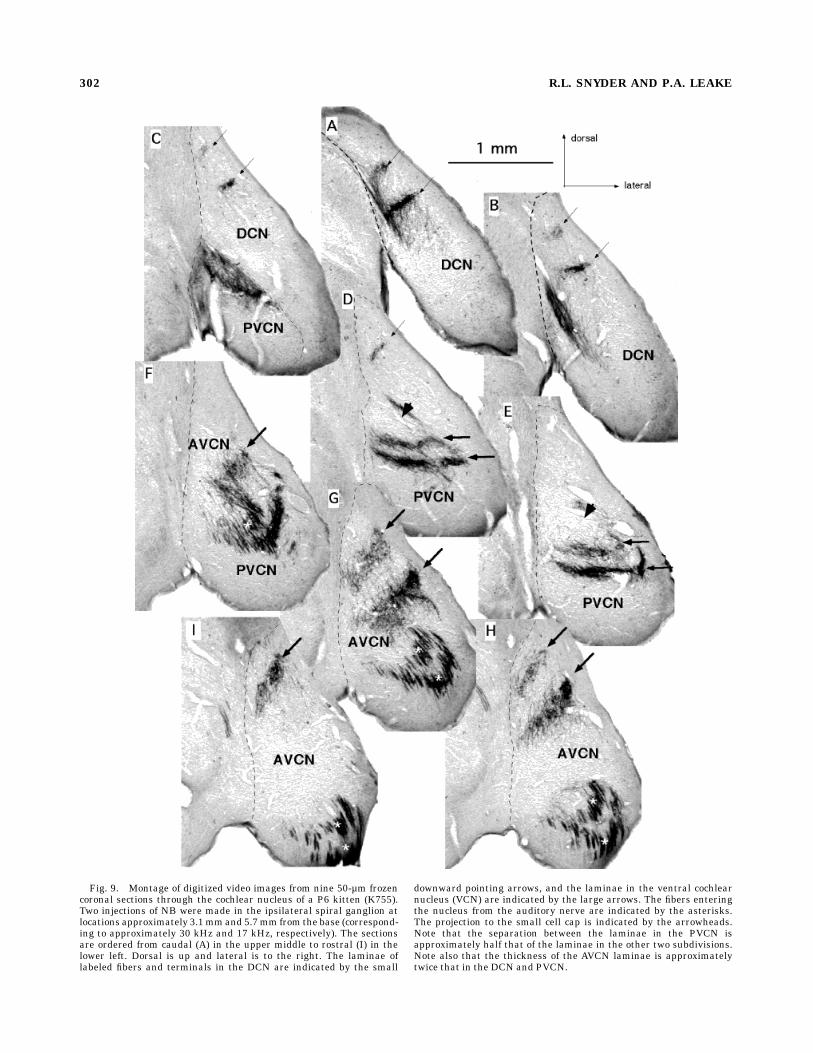
Fig. 9. Montage of digitized video images from nine 50-µm frozencoronal sections through the cochlear nucleus of a P6 kitten (K755).Two injections of NB were made in the ipsilateral spiral ganglion atlocations approximately 3.1mmand 5.7mm from the base (correspond-ing to approximately 30 kHz and 17 kHz, respectively). The sectionsare ordered from caudal (A) in the upper middle to rostral (I) in thelower left. Dorsal is up and lateral is to the right. The laminae oflabeled fibers and terminals in the DCN are indicated by the small
downward pointing arrows, and the laminae in the ventral cochlearnucleus (VCN) are indicated by the large arrows. The fibers enteringthe nucleus from the auditory nerve are indicated by the asterisks.The projection to the small cell cap is indicated by the arrowheads.Note that the separation between the laminae in the PVCN isapproximately half that of the laminae in the other two subdivisions.Note also that the thickness of the AVCN laminae is approximatelytwice that in the DCN and PVCN.
302 R.L. SNYDER AND P.A. LEAKE
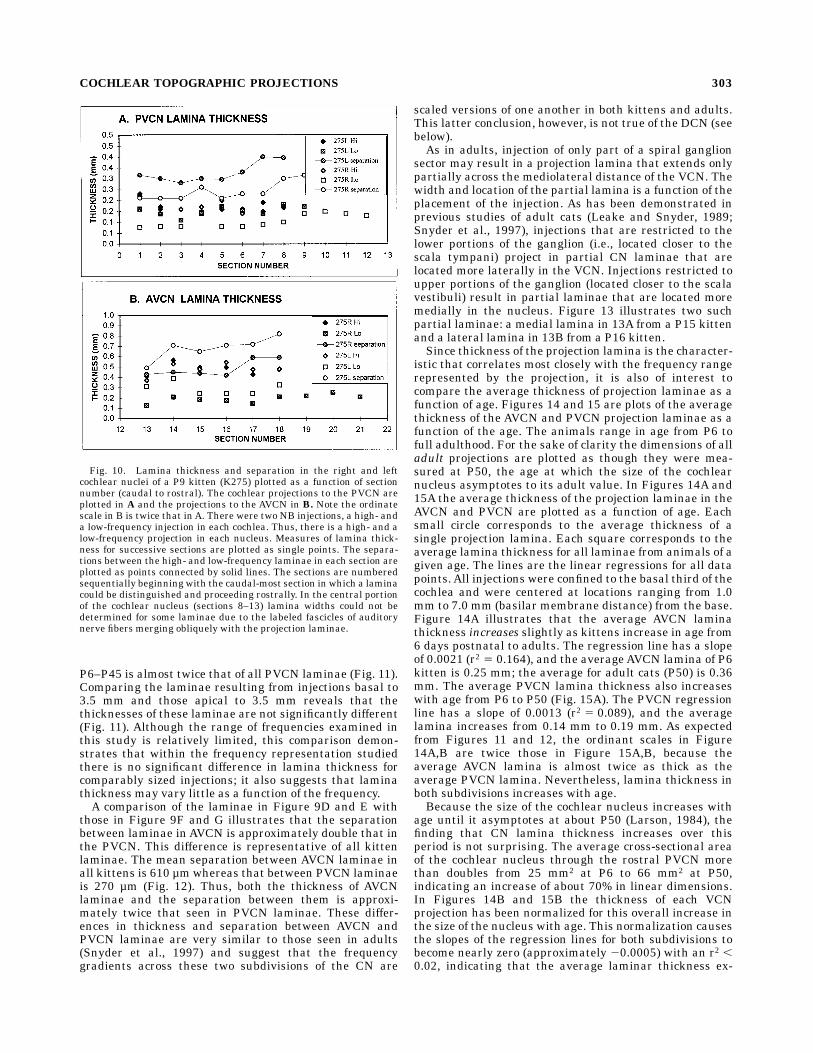
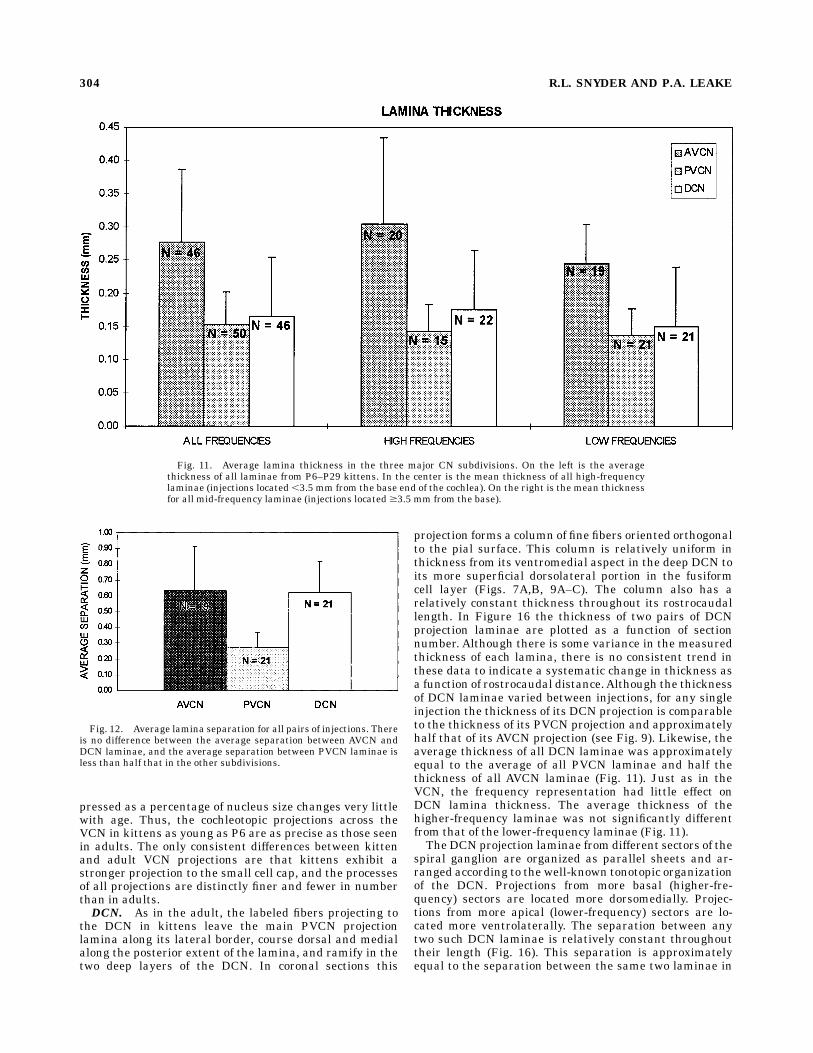
P6–P45 is almost twice that of all PVCN laminae (Fig. 11).Comparing the laminae resulting from injections basal to3.5 mm and those apical to 3.5 mm reveals that thethicknesses of these laminae are not significantly different(Fig. 11). Although the range of frequencies examined inthis study is relatively limited, this comparison demon-strates that within the frequency representation studiedthere is no significant difference in lamina thickness forcomparably sized injections; it also suggests that laminathickness may vary little as a function of the frequency.A comparison of the laminae in Figure 9D and E with
those in Figure 9F and G illustrates that the separationbetween laminae in AVCN is approximately double that inthe PVCN. This difference is representative of all kittenlaminae. The mean separation between AVCN laminae inall kittens is 610 µmwhereas that between PVCN laminaeis 270 µm (Fig. 12). Thus, both the thickness of AVCNlaminae and the separation between them is approxi-mately twice that seen in PVCN laminae. These differ-ences in thickness and separation between AVCN andPVCN laminae are very similar to those seen in adults(Snyder et al., 1997) and suggest that the frequencygradients across these two subdivisions of the CN are
scaled versions of one another in both kittens and adults.This latter conclusion, however, is not true of the DCN (seebelow).As in adults, injection of only part of a spiral ganglion
sector may result in a projection lamina that extends onlypartially across the mediolateral distance of the VCN. Thewidth and location of the partial lamina is a function of theplacement of the injection. As has been demonstrated inprevious studies of adult cats (Leake and Snyder, 1989;Snyder et al., 1997), injections that are restricted to thelower portions of the ganglion (i.e., located closer to thescala tympani) project in partial CN laminae that arelocated more laterally in the VCN. Injections restricted toupper portions of the ganglion (located closer to the scalavestibuli) result in partial laminae that are located moremedially in the nucleus. Figure 13 illustrates two suchpartial laminae: a medial lamina in 13A from a P15 kittenand a lateral lamina in 13B from a P16 kitten.Since thickness of the projection lamina is the character-
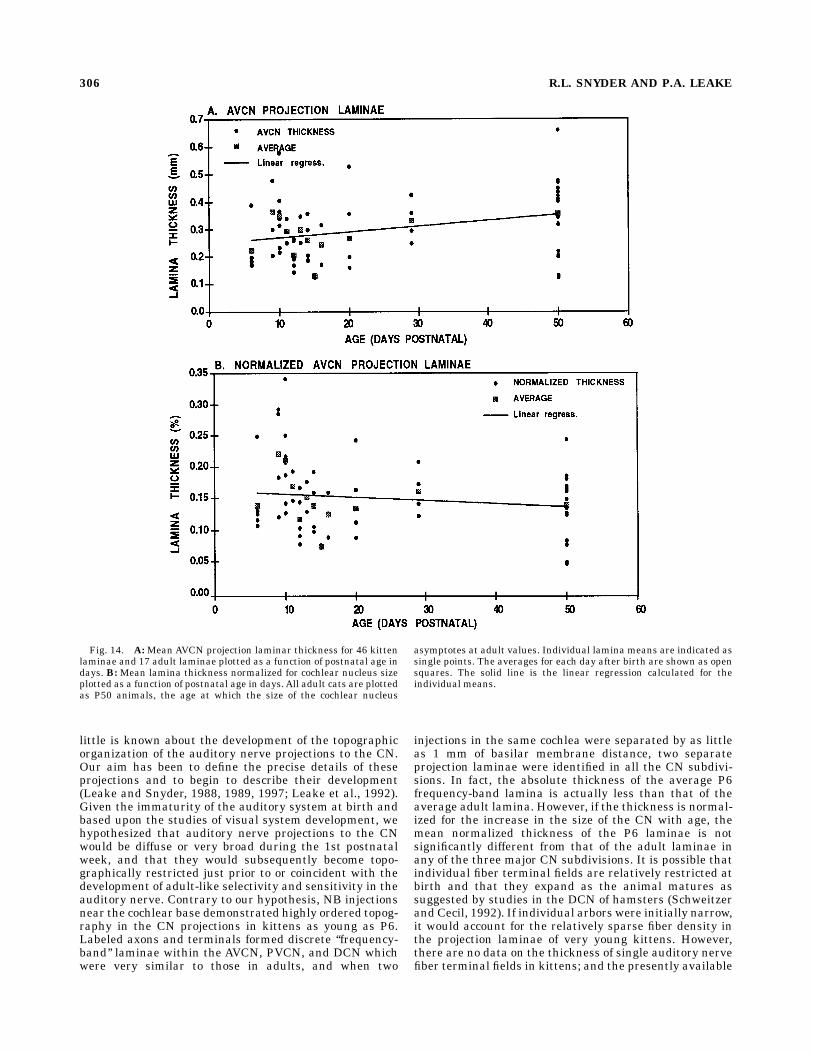
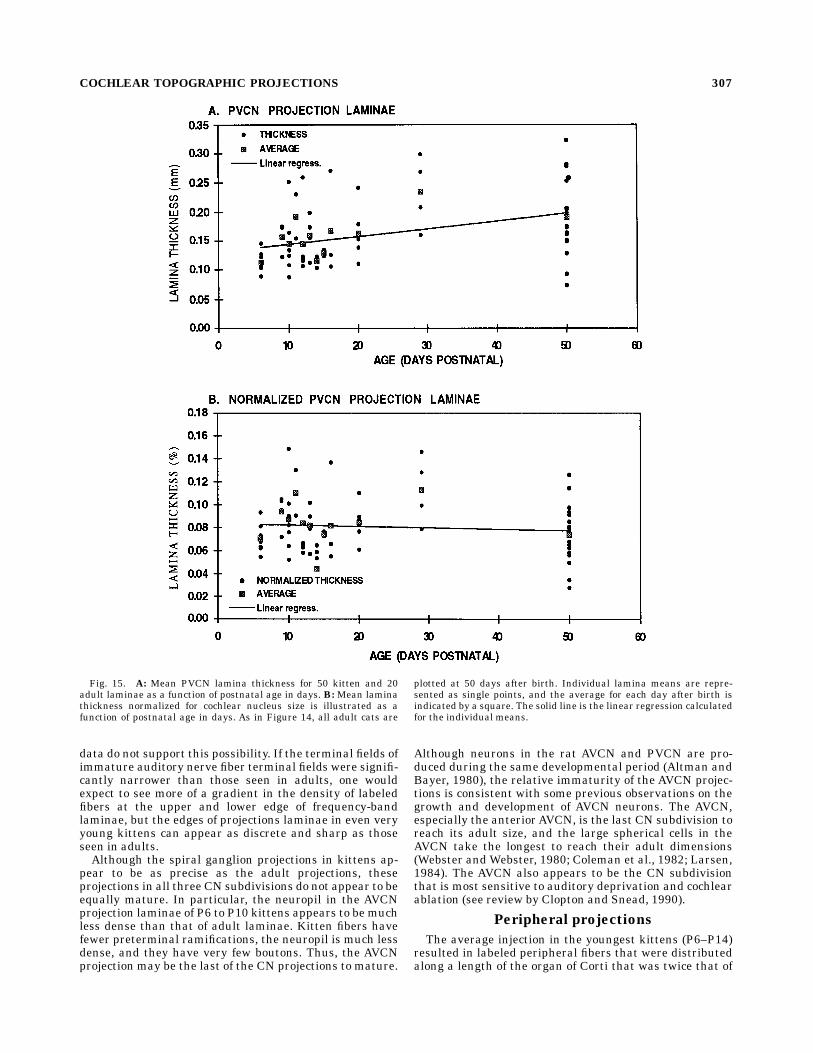
istic that correlates most closely with the frequency rangerepresented by the projection, it is also of interest tocompare the average thickness of projection laminae as afunction of age. Figures 14 and 15 are plots of the averagethickness of the AVCN and PVCN projection laminae as afunction of the age. The animals range in age from P6 tofull adulthood. For the sake of clarity the dimensions of alladult projections are plotted as though they were mea-sured at P50, the age at which the size of the cochlearnucleus asymptotes to its adult value. In Figures 14A and15A the average thickness of the projection laminae in theAVCN and PVCN are plotted as a function of age. Eachsmall circle corresponds to the average thickness of asingle projection lamina. Each square corresponds to theaverage lamina thickness for all laminae from animals of agiven age. The lines are the linear regressions for all datapoints. All injections were confined to the basal third of thecochlea and were centered at locations ranging from 1.0mm to 7.0 mm (basilar membrane distance) from the base.Figure 14A illustrates that the average AVCN laminathickness increases slightly as kittens increase in age from6 days postnatal to adults. The regression line has a slopeof 0.0021 (r2 5 0.164), and the averageAVCN lamina of P6kitten is 0.25 mm; the average for adult cats (P50) is 0.36mm. The average PVCN lamina thickness also increaseswith age from P6 to P50 (Fig. 15A). The PVCN regressionline has a slope of 0.0013 (r2 5 0.089), and the averagelamina increases from 0.14 mm to 0.19 mm. As expectedfrom Figures 11 and 12, the ordinant scales in Figure14A,B are twice those in Figure 15A,B, because theaverage AVCN lamina is almost twice as thick as theaverage PVCN lamina. Nevertheless, lamina thickness inboth subdivisions increases with age.Because the size of the cochlear nucleus increases with
age until it asymptotes at about P50 (Larson, 1984), thefinding that CN lamina thickness increases over thisperiod is not surprising. The average cross-sectional areaof the cochlear nucleus through the rostral PVCN morethan doubles from 25 mm2 at P6 to 66 mm2 at P50,indicating an increase of about 70% in linear dimensions.In Figures 14B and 15B the thickness of each VCNprojection has been normalized for this overall increase inthe size of the nucleus with age. This normalization causesthe slopes of the regression lines for both subdivisions tobecome nearly zero (approximately 20.0005) with an r2 ,0.02, indicating that the average laminar thickness ex-
Fig. 10. Lamina thickness and separation in the right and leftcochlear nuclei of a P9 kitten (K275) plotted as a function of sectionnumber (caudal to rostral). The cochlear projections to the PVCN areplotted in A and the projections to the AVCN in B. Note the ordinatescale in B is twice that inA. There were two NB injections, a high- anda low-frequency injection in each cochlea. Thus, there is a high- and alow-frequency projection in each nucleus. Measures of lamina thick-ness for successive sections are plotted as single points. The separa-tions between the high- and low-frequency laminae in each section areplotted as points connected by solid lines. The sections are numberedsequentially beginning with the caudal-most section in which a laminacould be distinguished and proceeding rostrally. In the central portionof the cochlear nucleus (sections 8–13) lamina widths could not bedetermined for some laminae due to the labeled fascicles of auditorynerve fibers merging obliquely with the projection laminae.
COCHLEAR TOPOGRAPHIC PROJECTIONS 303
pressed as a percentage of nucleus size changes very littlewith age. Thus, the cochleotopic projections across theVCN in kittens as young as P6 are as precise as those seenin adults. The only consistent differences between kittenand adult VCN projections are that kittens exhibit astronger projection to the small cell cap, and the processesof all projections are distinctly finer and fewer in numberthan in adults.DCN. As in the adult, the labeled fibers projecting to
the DCN in kittens leave the main PVCN projectionlamina along its lateral border, course dorsal and medialalong the posterior extent of the lamina, and ramify in thetwo deep layers of the DCN. In coronal sections this
projection forms a column of fine fibers oriented orthogonalto the pial surface. This column is relatively uniform inthickness from its ventromedial aspect in the deep DCN toits more superficial dorsolateral portion in the fusiformcell layer (Figs. 7A,B, 9A–C). The column also has arelatively constant thickness throughout its rostrocaudallength. In Figure 16 the thickness of two pairs of DCNprojection laminae are plotted as a function of sectionnumber. Although there is some variance in the measuredthickness of each lamina, there is no consistent trend inthese data to indicate a systematic change in thickness asa function of rostrocaudal distance.Although the thicknessof DCN laminae varied between injections, for any singleinjection the thickness of its DCN projection is comparableto the thickness of its PVCN projection and approximatelyhalf that of its AVCN projection (see Fig. 9). Likewise, theaverage thickness of all DCN laminae was approximatelyequal to the average of all PVCN laminae and half thethickness of all AVCN laminae (Fig. 11). Just as in theVCN, the frequency representation had little effect onDCN lamina thickness. The average thickness of thehigher-frequency laminae was not significantly differentfrom that of the lower-frequency laminae (Fig. 11).The DCN projection laminae from different sectors of the
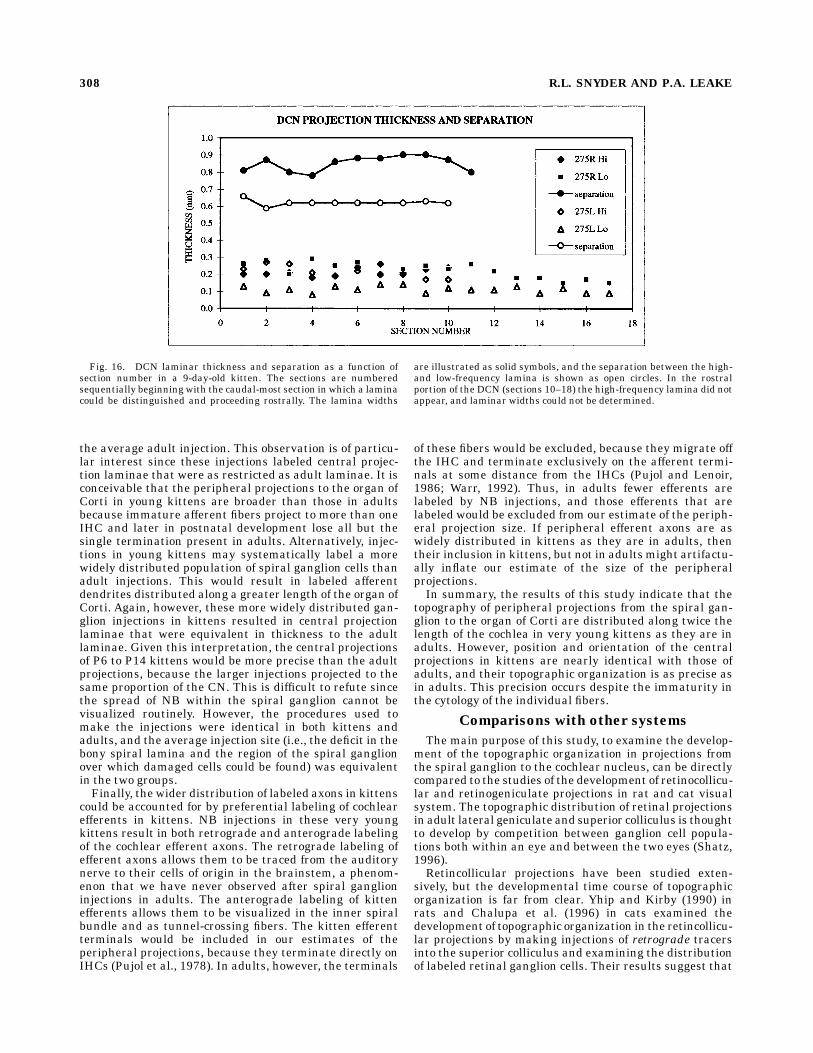
spiral ganglion are organized as parallel sheets and ar-ranged according to the well-known tonotopic organizationof the DCN. Projections from more basal (higher-fre-quency) sectors are located more dorsomedially. Projec-tions from more apical (lower-frequency) sectors are lo-cated more ventrolaterally. The separation between anytwo such DCN laminae is relatively constant throughouttheir length (Fig. 16). This separation is approximatelyequal to the separation between the same two laminae in
Fig. 11. Average lamina thickness in the three major CN subdivisions. On the left is the averagethickness of all laminae from P6–P29 kittens. In the center is the mean thickness of all high-frequencylaminae (injections located ,3.5 mm from the base end of the cochlea). On the right is the mean thicknessfor all mid-frequency laminae (injections located $3.5 mm from the base).
Fig. 12. Average lamina separation for all pairs of injections. Thereis no difference between the average separation between AVCN andDCN laminae, and the average separation between PVCN laminae isless than half that in the other subdivisions.
304 R.L. SNYDER AND P.A. LEAKE
the AVCN and approximately twice the separation in thePVCN (Fig. 9). Quantitative comparison of all doubleinjections shows that the mean separation between DCNprojections is not statistically different from the meanAVCN separation but is twice the mean separation be-tween PVCN projections (Fig. 12).Quantitative comparisons of the thickness of DCN lami-
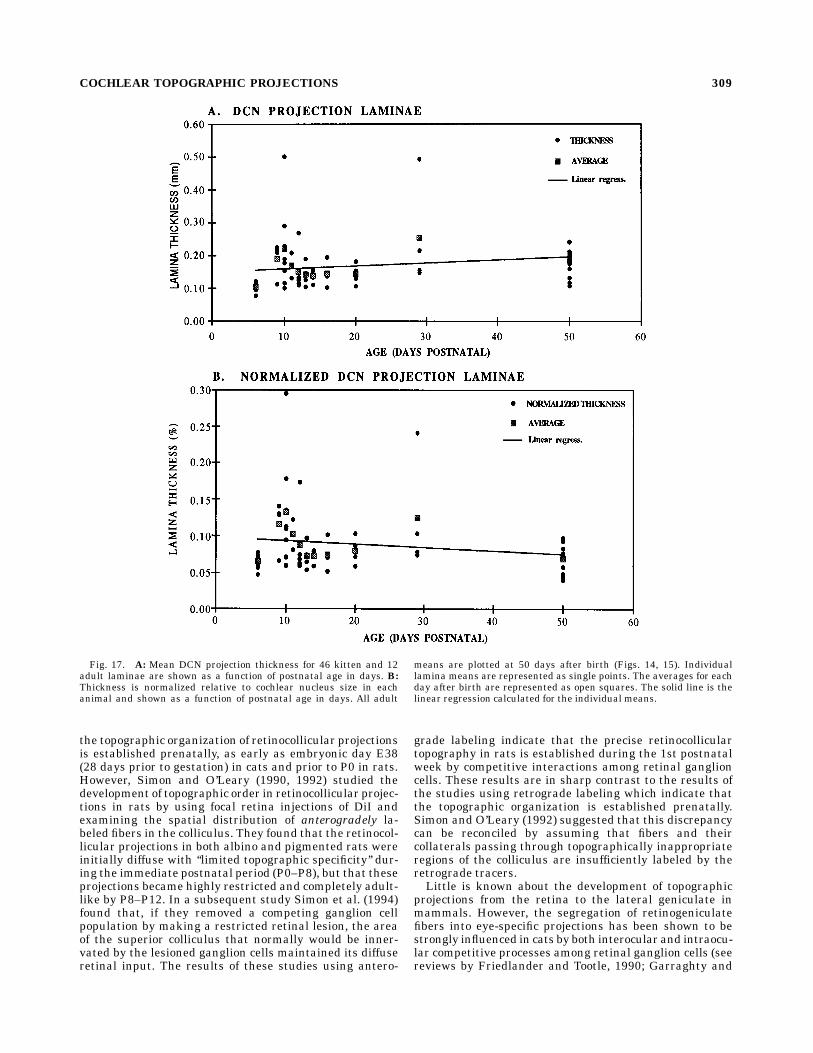
nae in kittens and adults are illustrated in Figure 17,which shows average lamina thickness as a function of agefrom P6 to adulthood. Adult projections are plotted asthough they were 50 days of age. Each small circle inFigure 17A represents the average width of a DCN projec-tion from a single spiral ganglion injection. In Figure 17Beach projection thickness is normalized for the overallincrease in the size of the nucleus with age. The squares ineach figure represent the average projection thickness ateach age. The two lines are the linear regressions of eachdata set. As can be seen in Figure 17A, the averageprojection widths increase slightly as kittens increase inage from P6 to adulthood, whereas the normalized projec-tion width decreases slightly with age (Fig. 17B). However,the slopes of these regressions are small (,0.0009), andthe r2 values are low (r2 , 0.02). Thus, these data indicatethat the thickness of DCN projections does not changesignificantly with age after P6 either in absolute dimen-sions or as a percentage of nucleus size. The only consis-tent difference between kitten and adult DCN projectionsis that the axonal processes in the DCN of these kittens aredistinctly smaller, and the felt work of the projectionneuropil is noticeably finer than in adults.
DISCUSSION
Based primarily on the studies of the development ofretinal projections to the central visual system, it isgenerally assumed that all mammalian sensory pathwaysare refined and topographically organized by the coordi-nated influence of molecular interactions (see Kaprielianand Patterson, 1994; Tessier-Lavigne, 1995) and electricalactivity (see Shatz, 1996). However, the generality of thisassumption remains to be tested. The auditory system ofthe cat offers several specific advantages for examining thedevelopment of topographic organization. First, the highlyprecise organization of the tonotopic projections of theauditory nerve, with each fiber projecting collaterals toeach of the three CN subdivisions, makes this system aparticularly interesting model for studying the develop-ment of topographic organization. Second, as mentionedpreviously, the cat auditory system is so immature at birththat it is virtually nonfunctional. Auditory nerve fibersexhibit very high thresholds, broad tuning, and very lowspontaneous discharge rates (mean rate ,10 spikes/s)throughout the first 8 to 10 postnatal days. Finally,numerous electrophysiological studies have provided im-portant data characterizing the time course of functionaldevelopment of the cat auditory system. This research hasrevealed that around P10 there is a relatively rapidtransition from a neuronal population predominantly rep-resented by broadly tuned, high-threshold, immaturerhythmic temporal responses to much more adult-likecharacteristics by the end of the 2nd postnatal week or P21(Walsh and McGee, 1990; Walsh and Romand, 1992;Brugge, 1992).In contrast to the detailed data on the development of
physiological response in the auditory nerve and CN, very
Fig. 13. Photomicrographs of PVCN illustrating a medial partiallamina in a P15 kitten (A, K01) and a lateral partial lamina in a P16kitten (B, K262). Note the prominent projection to the small cell cap(arrowheads) in B. Scale bar 5 1 mm.
COCHLEAR TOPOGRAPHIC PROJECTIONS 305
little is known about the development of the topographicorganization of the auditory nerve projections to the CN.Our aim has been to define the precise details of theseprojections and to begin to describe their development(Leake and Snyder, 1988, 1989, 1997; Leake et al., 1992).Given the immaturity of the auditory system at birth andbased upon the studies of visual system development, wehypothesized that auditory nerve projections to the CNwould be diffuse or very broad during the 1st postnatalweek, and that they would subsequently become topo-graphically restricted just prior to or coincident with thedevelopment of adult-like selectivity and sensitivity in theauditory nerve. Contrary to our hypothesis, NB injectionsnear the cochlear base demonstrated highly ordered topog-raphy in the CN projections in kittens as young as P6.Labeled axons and terminals formed discrete ‘‘frequency-band’’ laminae within the AVCN, PVCN, and DCN whichwere very similar to those in adults, and when two
injections in the same cochlea were separated by as littleas 1 mm of basilar membrane distance, two separateprojection laminae were identified in all the CN subdivi-sions. In fact, the absolute thickness of the average P6frequency-band lamina is actually less than that of theaverage adult lamina. However, if the thickness is normal-ized for the increase in the size of the CN with age, themean normalized thickness of the P6 laminae is notsignificantly different from that of the adult laminae inany of the three major CN subdivisions. It is possible thatindividual fiber terminal fields are relatively restricted atbirth and that they expand as the animal matures assuggested by studies in the DCN of hamsters (Schweitzerand Cecil, 1992). If individual arbors were initially narrow,it would account for the relatively sparse fiber density inthe projection laminae of very young kittens. However,there are no data on the thickness of single auditory nervefiber terminal fields in kittens; and the presently available
Fig. 14. A:Mean AVCN projection laminar thickness for 46 kittenlaminae and 17 adult laminae plotted as a function of postnatal age indays. B: Mean lamina thickness normalized for cochlear nucleus sizeplotted as a function of postnatal age in days. All adult cats are plottedas P50 animals, the age at which the size of the cochlear nucleus
asymptotes at adult values. Individual lamina means are indicated assingle points. The averages for each day after birth are shown as opensquares. The solid line is the linear regression calculated for theindividual means.
306 R.L. SNYDER AND P.A. LEAKE
data do not support this possibility. If the terminal fields ofimmature auditory nerve fiber terminal fields were signifi-cantly narrower than those seen in adults, one wouldexpect to see more of a gradient in the density of labeledfibers at the upper and lower edge of frequency-bandlaminae, but the edges of projections laminae in even veryyoung kittens can appear as discrete and sharp as thoseseen in adults.Although the spiral ganglion projections in kittens ap-
pear to be as precise as the adult projections, theseprojections in all three CN subdivisions do not appear to beequally mature. In particular, the neuropil in the AVCNprojection laminae of P6 to P10 kittens appears to be muchless dense than that of adult laminae. Kitten fibers havefewer preterminal ramifications, the neuropil is much lessdense, and they have very few boutons. Thus, the AVCNprojection may be the last of the CN projections to mature.
Although neurons in the rat AVCN and PVCN are pro-duced during the same developmental period (Altman andBayer, 1980), the relative immaturity of the AVCN projec-tions is consistent with some previous observations on thegrowth and development of AVCN neurons. The AVCN,especially the anterior AVCN, is the last CN subdivision toreach its adult size, and the large spherical cells in theAVCN take the longest to reach their adult dimensions(Webster and Webster, 1980; Coleman et al., 1982; Larsen,1984). The AVCN also appears to be the CN subdivisionthat is most sensitive to auditory deprivation and cochlearablation (see review by Clopton and Snead, 1990).
Peripheral projections
The average injection in the youngest kittens (P6–P14)resulted in labeled peripheral fibers that were distributedalong a length of the organ of Corti that was twice that of
Fig. 15. A: Mean PVCN lamina thickness for 50 kitten and 20adult laminae as a function of postnatal age in days. B: Mean laminathickness normalized for cochlear nucleus size is illustrated as afunction of postnatal age in days. As in Figure 14, all adult cats are
plotted at 50 days after birth. Individual lamina means are repre-sented as single points, and the average for each day after birth isindicated by a square. The solid line is the linear regression calculatedfor the individual means.
COCHLEAR TOPOGRAPHIC PROJECTIONS 307
the average adult injection. This observation is of particu-lar interest since these injections labeled central projec-tion laminae that were as restricted as adult laminae. It isconceivable that the peripheral projections to the organ ofCorti in young kittens are broader than those in adultsbecause immature afferent fibers project to more than oneIHC and later in postnatal development lose all but thesingle termination present in adults. Alternatively, injec-tions in young kittens may systematically label a morewidely distributed population of spiral ganglion cells thanadult injections. This would result in labeled afferentdendrites distributed along a greater length of the organ ofCorti. Again, however, these more widely distributed gan-glion injections in kittens resulted in central projectionlaminae that were equivalent in thickness to the adultlaminae. Given this interpretation, the central projectionsof P6 to P14 kittens would be more precise than the adultprojections, because the larger injections projected to thesame proportion of the CN. This is difficult to refute sincethe spread of NB within the spiral ganglion cannot bevisualized routinely. However, the procedures used tomake the injections were identical in both kittens andadults, and the average injection site (i.e., the deficit in thebony spiral lamina and the region of the spiral ganglionover which damaged cells could be found) was equivalentin the two groups.Finally, the wider distribution of labeled axons in kittens
could be accounted for by preferential labeling of cochlearefferents in kittens. NB injections in these very youngkittens result in both retrograde and anterograde labelingof the cochlear efferent axons. The retrograde labeling ofefferent axons allows them to be traced from the auditorynerve to their cells of origin in the brainstem, a phenom-enon that we have never observed after spiral ganglioninjections in adults. The anterograde labeling of kittenefferents allows them to be visualized in the inner spiralbundle and as tunnel-crossing fibers. The kitten efferentterminals would be included in our estimates of theperipheral projections, because they terminate directly onIHCs (Pujol et al., 1978). In adults, however, the terminals
of these fibers would be excluded, because they migrate offthe IHC and terminate exclusively on the afferent termi-nals at some distance from the IHCs (Pujol and Lenoir,1986; Warr, 1992). Thus, in adults fewer efferents arelabeled by NB injections, and those efferents that arelabeled would be excluded from our estimate of the periph-eral projection size. If peripheral efferent axons are aswidely distributed in kittens as they are in adults, thentheir inclusion in kittens, but not in adultsmight artifactu-ally inflate our estimate of the size of the peripheralprojections.In summary, the results of this study indicate that the
topography of peripheral projections from the spiral gan-glion to the organ of Corti are distributed along twice thelength of the cochlea in very young kittens as they are inadults. However, position and orientation of the centralprojections in kittens are nearly identical with those ofadults, and their topographic organization is as precise asin adults. This precision occurs despite the immaturity inthe cytology of the individual fibers.
Comparisons with other systems
The main purpose of this study, to examine the develop-ment of the topographic organization in projections fromthe spiral ganglion to the cochlear nucleus, can be directlycompared to the studies of the development of retinocollicu-lar and retinogeniculate projections in rat and cat visualsystem. The topographic distribution of retinal projectionsin adult lateral geniculate and superior colliculus is thoughtto develop by competition between ganglion cell popula-tions both within an eye and between the two eyes (Shatz,1996).Retincollicular projections have been studied exten-
sively, but the developmental time course of topographicorganization is far from clear. Yhip and Kirby (1990) inrats and Chalupa et al. (1996) in cats examined thedevelopment of topographic organization in the retincollicu-lar projections by making injections of retrograde tracersinto the superior colliculus and examining the distributionof labeled retinal ganglion cells. Their results suggest that
Fig. 16. DCN laminar thickness and separation as a function ofsection number in a 9-day-old kitten. The sections are numberedsequentially beginning with the caudal-most section in which a laminacould be distinguished and proceeding rostrally. The lamina widths
are illustrated as solid symbols, and the separation between the high-and low-frequency lamina is shown as open circles. In the rostralportion of the DCN (sections 10–18) the high-frequency lamina did notappear, and laminar widths could not be determined.
308 R.L. SNYDER AND P.A. LEAKE
the topographic organization of retinocollicular projectionsis established prenatally, as early as embryonic day E38(28 days prior to gestation) in cats and prior to P0 in rats.However, Simon and O’Leary (1990, 1992) studied thedevelopment of topographic order in retinocollicular projec-tions in rats by using focal retina injections of DiI andexamining the spatial distribution of anterogradely la-beled fibers in the colliculus. They found that the retinocol-licular projections in both albino and pigmented rats wereinitially diffuse with ‘‘limited topographic specificity’’ dur-ing the immediate postnatal period (P0–P8), but that theseprojections became highly restricted and completely adult-like by P8–P12. In a subsequent study Simon et al. (1994)found that, if they removed a competing ganglion cellpopulation by making a restricted retinal lesion, the areaof the superior colliculus that normally would be inner-vated by the lesioned ganglion cells maintained its diffuseretinal input. The results of these studies using antero-
grade labeling indicate that the precise retinocolliculartopography in rats is established during the 1st postnatalweek by competitive interactions among retinal ganglioncells. These results are in sharp contrast to the results ofthe studies using retrograde labeling which indicate thatthe topographic organization is established prenatally.Simon and O’Leary (1992) suggested that this discrepancycan be reconciled by assuming that fibers and theircollaterals passing through topographically inappropriateregions of the colliculus are insufficiently labeled by theretrograde tracers.Little is known about the development of topographic
projections from the retina to the lateral geniculate inmammals. However, the segregation of retinogeniculatefibers into eye-specific projections has been shown to bestrongly influenced in cats by both interocular and intraocu-lar competitive processes among retinal ganglion cells (seereviews by Friedlander and Tootle, 1990; Garraghty and
Fig. 17. A: Mean DCN projection thickness for 46 kitten and 12adult laminae are shown as a function of postnatal age in days. B:Thickness is normalized relative to cochlear nucleus size in eachanimal and shown as a function of postnatal age in days. All adult
means are plotted at 50 days after birth (Figs. 14, 15). Individuallamina means are represented as single points. The averages for eachday after birth are represented as open squares. The solid line is thelinear regression calculated for the individual means.
COCHLEAR TOPOGRAPHIC PROJECTIONS 309

Sur, 1993; Shatz, 1996). Initially, retinal ganglion cells ineach eye project to all layers of the lateral geniculate.During development interocular competition results inprojections from the two eyes that are segregated intoseparate and specific geniculate layers (Sretavan andShatz, 1986, 1987). By the 2nd postnatal day (P2) retinalganglion cells in the contralateral eye project almostexclusively to layers A, C, and C2 of the lateral geniculate,and the ipsilateral ganglion cells project primarily tolayers A1 and C1. The segregation of the projections is notcomplete until the beginning of the 2nd postnatal week(P7–8) when there is with no overlap between the projec-tions of the two eyes. Interocular competition is alsonecessary for maintenance of this segregation once it hasbeen established, as removal of one eye in the earlypostnatal period (P7–9), after segregation of retinogenicu-late projection into the adult-like pattern, results in spout-ing of some of the remaining retinogeniculate axons intodenervated areas of the geniculate (Guillery, 1972; Hickey,1975; Robson, 1981). Thus, interocular competition duringthe pre- and postnatal period culminates in and maintainssegregated eye-specific retinogeniculate terminal fields. Ina similar manner intraocular competition strongly influ-ences the size of terminal arbors of retinogeniculate axons.For example, competition during postnatal developmentbetween X andY axons results in the shrinkage of X arborsas the arbors of Y axons expand (Sur et al., 1984). Over thissame time interval (12–16 weeks postnatal) the spatialresolution of lateral geniculate neurons increases, anobservation that is consistent with a refinement of retino-geniculate terminal fields (Friedlander and Tootle, 1990).It is these final refined termnal fields of retinogeniculatearbors that most closely corresponds to the frequency-band laminae reported here.Therefore, based on the development of retinocollicular
projections in the rat, the development of eye-specificprojections in the cat retinogeniculate pathway, and therefinement of X cell terminal fields in the cat, we hypoth-esized that in the auditory system spiral ganglion cellsmight initially project diffusely across each cochlear nucleussubdivision and then become more selective and refinedduring pre- and postnatal development. Surprisingly, thisprocess of refinement in topography of cochlear projectionsappears to be already complete by P6, the age of theyoungest kittens reported in this study.
ACKNOWLEDGMENTS
The authors acknowledge the assistance of Beth Dwanand Gary Hradek in the conduct of these experiments andDrs. J.H. LaVail and S.B. Tieman for their suggestions onthe manuscript.
LITERATURE CITED
Altman, J., and S.A. Bayer (1980) Development of the brain stem in the rat.III. Thymidine-radiographic study of the time of origin of neurons of thevestibular and auditory nuclei of the upper medulla. J. Comp. Neurol.194:877–904.
Bourk, T.R., J.M.Mielcarz, and B.E. Norris (1981) Tonotopic organization ofthe anteroventral cochlear nucleus of the cat. Hearing Res. 4:215–241.
Brawer, J.R., D.K. Morest, and E.I. Kane (1974) The neuronal architectureof the cochlear nucleus of the cat. J. Comp. Neurol. 155:251–300.
Brugge, J.F. (1988) Stimulus coding in the developing auditory system. InG.M. Edelman, W.E. Gall, and W.M. Cowan (eds): Auditory Function.Neurobiological Bases of Hearing. New York: John Wiley & Sons, pp.113–136.
Brugge, J.F. (1992) Development of the lower auditory brainstem of the cat.In R. Romand (ed): Development of Auditory and Vestibular Systems 2.NewYork: Elsevier, pp. 273–296.
Brugge, J.F., E. Javel, and L.M. Kitzes (1978) Signs of functional matura-tion of peripheral auditory system in discharge patterns of neurons inthe anteroventral cochlear nucleus in kitten. J. Neurophysiol. 41:1557–1579.
Cant, N.B., and D.K.Morest (1984) The structural basis for stimulus codingin the cochlear nucleus of the cat. In C. Berlin (ed): Hearing Science:RecentAdvances. San Diego, CA: College-Hill, pp. 371–421.
Carlier, E., M. Abonnenc, and R. Pujol (1975) Maturation des responsesunitaires a la stimulation tonale dans le nerf cochleaire du chaton. J.Physiol. (Paris) 70:129–138.
Chalupa, L.M., C.J. Snider, and M.A. Kirby (1996) Topographic organiza-tion in the retinocollicular pathway of the fetal cat demonstrated byretrograde labeling of ganglion cells. J. Comp. Neurol. 368:295–303.
Clements, M., and J.B. Kelly (1978) Directional responses by kittens to anauditory stimulus. Dev. Psychobiol. 11:505–511.
Clopton, B.M., and C.R. Snead (1990) Experimental factors in auditorydevelopment. In R. Romand (ed): Development ofAuditory and Vestibu-lar Systems 2. NewYork: Elsevier, pp. 317–338.
Coleman, J., B.J. Blatchley, and J.E. Williams (1982) Development of thedorsal and ventral cochlear nucleus in rat and effects of acousticdeprivation. Dev. Brain Res. 4:119–123.
Ehret, G., and R. Romand (1981) Postnatal development of absoluteauditory thresholds in kittens. J. Comp. Physiol. Psychol. 95:304–311.
Fekete, D.M., E.M. Rouiller, M.C. Liberman, and D.K. Ryugo (1984) Thecentral projections of intracellularly labeled auditory nerve fibers incats. J. Comp. Neurol. 229:432–450.
Feng, A.S., and M. Vater (1985) Functional organization of the cochlearnucleus of rufous horseshoe bats (Rhinolophus rouxi): frequencies andinternal connections are arranged in slabs. J. Comp. Neurol. 235:529–553.
Foss, I., and G. Flottorp (1974) A comparative study of the development ofhearing and vision in various species commonly used in experiments.Acta Otolaryngol. 77:202–214.
Friedlander, M.J., and J.S. Tootle (1990) Postnatal anatomical and physi-ological development of the visual system. In J.R. Coleman (ed):Development of Sensory System in Mammals. New York: Wiley, pp.61–124.
Garraghty, P.E., andM. Sur (1993) Competitive interactions influencing thedevelopment of retinal axonal arbors in cat lateral geniculate nucleus.Physiol. Rev. 73:529–545.
Greenwood, D.D. (1974) Critical bandwidth in man and some other speciesin relation to the traveling wave envelope. In H.R. Moskowitz and J.C.Stevens (eds): Sensation and Measurement. Boston: Reidel, pp. 231–239.
Guillery, R.W. (1972) Experiments to determine whether retinogeniculateaxons can form translaminar collateral sprouts in the dorsal lateralgeniculate nucleus of the cat. J. Comp. Neurol. 146:407–420.
Hickey, T.L. (1975) Translaminar growth of axons in the kitten dorsallateral geniculate nucleus following removal of one eye. J. Comp.Neurol. 161:359–382.
Kaprielian, Z., and P.H. Patterson (1994) The molecular basis of retinotec-tal topography. Bioessays 16:1–11.
Keithley, E.M., and R. Cronin-Schreiber (1987) Frequency map of the spiralganglion in the cat. J. Acoust. Soc. Am. 81:1036–1042.
Kettener, R.E., J.Z. Feng, and J.F. Brugge (1985) Postnatal development ofthe phase-locked response to low frequency tones of auditory nervefibers in the cat. J. Neurosci. 5:275–283.
Kim, D.O., and C.E. Molnar (1972) A population study of cochlear nervefibers: comparison of spatial distributions of average-rate and phase-locking measures of responses to single tones. J. Neurophysiol. 42:16–30.
Larsen, S.A. (1984) Postnatal maturation of the cat cochlear nuclearcomplex. Acta Otolaryngol. [Suppl.] 417:1–43.
Leake, P.A., and R.L. Snyder (1988) An additional dimension in thetopographic organization of the spiral ganglion projection to the ventralcochlear nucleus. Soc. Neurosci. Abstr. 14:322.
Leake, P.A., and R.L. Snyder (1989) Topographic organization of the centralprojections of the spiral ganglion in cats. J. Comp. Neurol. 281:612–629.
Leake, P.A., and R.L. Snyder (1996) Topographic organization of theprojections of the spiral ganglion to the ventral cochlear nucleus. InW.A. Ainsworth (ed.): Advances in Speech, Hearing and LanguageProcessing, Vol. 3. London: JAI, pp. 3–17.
310 R.L. SNYDER AND P.A. LEAKE

Leake, P.A., and R.L. Snyder (1997) Postnatal refinement in the spiralganglion projections to the cochlear nucleus in cats. Assoc. Res.Otolaryngol. 20:53.
Leake, P.A., R.L. Snyder, andM.M.Merzenich (1992) Topographic organiza-tion of the cochlear spiral ganglion demonstrated by restricted lesions ofthe anteroventral cochlear nucleus. J. Comp. Neurol. 320:468–478.
Liberman, M.C. (1978) Auditory-nerve responses from cats raised in alow-noise chamber. J. Acoust. Soc. Am. 63:442–455.
Liberman, M.C. (1982) The cochlear frequency map for the cat: labelingauditory-nerve fibers of known characteristic frequency. J. Acoust. Soc.Am. 72:1441–1449.
Noda, Y., and W. Prisig (1974) Anatomical projections of the cochlea to thecochlear nucleus of the guinea pig. Arch. Otorhinolaryngol. 208:107–120.
Olmstead, C.E., and J.R. Villablanca (1980) Development of behavioralaudition in the kitten. Physiol. Behav. 24:705–712.
Osen, K.K. (1969) Cytoarchitecture of the cochlear nuclei in the cat. J.Comp. Neurol. 136:453–484.
Osen, K.K. (1970) Course and termination of the primary afferents in thecochlear nuclei of the cat: an experimental anatomical study. Arch. Ital.Biol. 108:21–51.
Pujol, R., and M. Lenoir (1986) The four types of synapses in the organ ofCorti. In R.A. Altschuler, R.P. Bobin, and D.W. Hoffman (eds): Neurobi-ology of Hearing: The Cochlea. NewYork: Raven, pp. 161–172.
Pujol, R., E. Carlier, and C. Devigne (1978) Different patterns of cochlearinnervation during the development in the kitten. J. Comp. Neurol.117:529–536.
Robson, J.A. (1981) Abnormal axonal growth in the dorsal lateral genicu-late nucleus of the cat. J. Comp. Neurol. 195:453–476.
Romand, R. (1983) Development in the frequency selectivity of auditorynerve fibers in the kitten. J. Neurosci. Lett. 35:271–276.
Romand, R., and R. Marty (1975) Postnatal maturation of the cochlearnuclei in the cat: a neurophysiological study. Brain Res. 83:225–233.
Rose, J.E., R. Galambos, and J. Hughes (1960) Organization of frequencysensitive neurons in the cochlear nuclear complex of the cat. In G.L.Rassmussen and W.F. Windle (eds): Neuronal Mechanisms of theAuditory and Vestibular Systems. Springfield: Thomas, pp. 116–136.
Rouiller, E.M., R. Cronin-Schreiber, D.M. Fekete, and D.K. Ryugo (1986)The central projections of intracellularly labeled auditory nerve fibersin cats: an analysis of terminal morphology. J. Comp. Neurol. 249:261–278.
Ryugo, D.K., and D.M. Fekete (1982) Morphology of primary axosomaticendings in the anteroventral cochlear nucleus of the cat: a study of theendbulbs of Held. J. Comp. Neurol. 210:239–257.
Ryugo, D.K., and S.K. May (1993) The projections of intracellularly labeledauditory nerve fibers to the dorsal cochlear nucleus of cats. J. Comp.Neurol. 329:20–35.
Sando, I. (1965) The anatomical interrelationships of the cochlear nervefibers. Acta Otolaryngol. 59:417–436.
Schweitzer, L., and T. Cecil (1992) Morphology of HRP-labeled cochlearnerve axons in the dorsal cochlear nucleus of the developing hamster.Hearing Res. 60:34–44.
Shatz, C. (1996) Emergence of order in visual system development. Proc.Natl. Acad. Sci. U.S.A. 93:602–608.
Simon, D.K., and D.D.M. O’Leary (1990) Limited topographic specificity inthe targeting and branching of mammalian retinal axons. Dev. Biol.137:125–134.
Simon, D.K., and D.D.M. O’Leary (1992) Development of topographic orderin the mammalian retinocollicular projection. J. Neuroscience 12:1212–1232.
Simon, D.K., A.L. Roskies, and D.D.M. O’Leary (1994) Plasticity in thedevelopment of topographic order in the mammalian retinocollicularprojection. Dev. Biol. 162:384–393.
Snyder, R.L., and P.A. Leake (1988) Intrinsic connections within andbetween cochlear nucleus subdivisions in the cat. J. Comp. Neurol.278:209–225.
Snyder, R.L., P.A. Leake, and G.T. Hradek (1997) Quantitative analysis ofspiral ganglion projections to the cat cochlear nucleus. J. Comp. Neurol.379:133–149.
Sretavan, D., and C. Shatz (1986) Prenatal development of retinal ganglioncell axons: segregation into eye-specific layers within the cat’s lateralgeniculate nucleus. J. Neurosci. 6:234–251.
Sretavan, D., and C. Shatz (1987) Axon trajectories and pattern of terminalarborization during prenatal development of the cat’s retinogeniculatepathway. J. Comp. Neurol. 255:356–400.
Sur, M., R.E. Weller, and S.M. Sherman (1984) Development of X- andY-cellretinogeniculate terminations in kittens. Nature 310:246–249.
Tessier-Lavigne, M. (1995) Eph receptor tyrosine kinases, axon repulsion,and the development of topographic maps. Cell 82:345–348.
Vidal, S., M. Lombard, P. Sanchez, A. Roman, and L. Moya (1995) An easymethod for the removal of Epon resin form semi-thin sections. Applica-tion of the avidin-biotin technique. Histochem. J. 27:204–209.
Walsh, E.J., and J. McGee (1986) The development of function in theauditory periphery. In R.A. Altschuler, D.W. Hoffman, and R.P. Bobbin(eds): Neurobiology of Hearing: The Cochlea. New York: Raven, pp.247–269.
Walsh, E.J., and J. McGee (1987) Postnatal development of auditory nerveand cochlear nucleus neuronal responses in kittens. Hearing Res.28:97–116.
Walsh, E.J., and J. McGee (1988) Rhythmic discharge properties of caudalcochlear nucleus neurons during postnatal development in cats. Hear-ing Res. 36:233–248.
Walsh, E.J., and J. McGee (1990) Frequency selectivity in the auditoryperiphery: similarities between damaged and developing ears. Am. J.Otolaryngol. 11:23–32.
Walsh, E.J., and R. Romand (1992) Functional development of the cochleaand the cochlear nerve. In R. Romand (ed): Development of Auditoryand Vestibular Systems 2. NewYork: Elsevier, pp. 161–219.
Walsh, E.J., J. McGee, and E. Javel (1986) Development of auditory-evokedpotentials in the cat. I. Onset of response and development of sensitivi-ties. J. Acoust. Soc. Am. 79:712–724.
Warr, W.B. (1992) Organization of olivocochlear efferent systems in mam-mals. In D.B. Webster, A.N. Popper, and R.R. Fay (eds): The Mamma-lian Auditory Pathway: Neuroanatomy. New York: Springer-Verlag, pp.410–448.
Webster, D., and M. Webster (1980) Mouse brainstem auditory nucleidevelopment. Ann. Otol. Rhinol. Laryngol. (Suppl.) 89:254–256.
Webster, W.B., J. Serviere, C. Batini, and S. Laplante (1978) Autoradio-graphic demonstration with 2-[14C] deoxyglucose of frequency selectiv-ity in the auditory system of cats under conditions of functional activity.Neurosci. Lett. 10:43–48.
Yhip, J.P., and M.A. Kirby (1990) Topographic organization of retinocollicu-lar projections in the neonatal rat. Vis. Neurosci. 4:313–329.
COCHLEAR TOPOGRAPHIC PROJECTIONS 311

























![Journal Wrist Ganglion[1]](https://img.pdfslide.us/doc/110x75/577cc6881a28aba7119e84ab/journal-wrist-ganglion1.jpg)



