Embed Size (px)
DESCRIPTION
research article
Citation preview

Pa
TEa
b
c
a
ARA
KBFDM
1
v
tg1aThhlsspSs
FP
0h
Postharvest Biology and Technology 70 (2012) 51–58
Contents lists available at SciVerse ScienceDirect
Postharvest Biology and Technology
jou rna l h omepa g e: www.elsev ier .com/ locate /postharvbio
roteomic analysis of banana fruit reveals proteins that are differentiallyccumulated during ripening
atiana Torres Toledoa , Silvia Beserra Nogueiraa , Beatriz Rosana Cordenunsia,b , Fábio César Gozzoc ,duardo Jorge Pilauc, Franco Maria Lajoloa,b, João Roberto Oliveira do Nascimentoa,b,∗
University of São Paulo, Department of Food Science and Experimental Nutrition, FCF, São Paulo, BrazilUniversity of São Paulo – NAPAN – Food and Nutrition Research Center, BrazilDepartment of Organic Chemistry, Institute of Chemistry, PO Box 6154, University of Campinas, UNICAMP, 13084-862 Campinas, SP, Brazil
r t i c l e i n f o
rticle history:eceived 31 August 2011ccepted 9 April 2012
eywords:ananaruit ripeningifferential proteomeusa spp.
a b s t r a c t
Bananas (Musa spp.) are highly perishable fruit of notable economic and nutritional relevance. Becausethe identification of proteins involved in metabolic pathways could help to extend green-life and improvethe quality of the fruit, this study aimed to compare the proteins of banana pulp at the pre-climactericand climacteric stages. The use of two-dimensional fluorescence difference gel electrophoresis (2D-DIGE)revealed 50 differentially expressed proteins, and comparing those proteins to the Mass SpectrometryProtein Sequence Database (MSDB) identified 26 known proteins. Chitinases were the most abundanttypes of proteins in unripe bananas, and two isoforms in the ripe fruit have been implicated in thestress/defense response. In this regard, three heat shock proteins and isoflavone reductase were alsoabundant at the climacteric stage. Concerning fruit quality, pectate lyase, malate dehydrogenase, andstarch phosphorylase accumulated during ripening. In addition to the ethylene formation enzyme amino
cyclo carboxylic acid oxidase, the accumulation of S-adenosyl-l-homocysteine hydrolase was neededbecause of the increased ethylene synthesis and DNA methylation that occurred in ripening bananas.Differential analysis provided information on the ripening-associated changes that occurred in proteinsinvolved in banana flavor, texture, defense, synthesis of ethylene, regulation of expression, and proteinfolding, and this analysis validated previous data on the transcripts during ripening. In this regard, thedifferential proteomics of fruit pulp enlarged our understanding of the process of banana ripening.. Introduction
Banana is a fruit crop of strong economic and nutritional rele-ance that is cultivated in tropical and subtropical regions around
Abbreviations: 2D-DIGE, two-dimensional fluorescence difference gel elec-rophoresis; ACC, amino cyclo carboxylic acid; ACN, acetonitrile; AGPase, ADP-lucose pyrophosphorylase; CHAPS, 3-[(3-cholamidopropyl) dimethylammonio]--propanesulfonate; Cy3, carbocyanine 3; Cy5, carbocyanine 5; DAA, days afternthesis; DTT, dithiothreitol; EDTA, ethylenediaminetetraacetic acid; ESI-QUAD-OF, electrospray ionization-quadrupole-time of flight; FA, formic acid; HPAE-PAD,igh-pressure liquid chromatography with pulse amperometric detection; HSPs,eat shock proteins; IFR, isoflavone reductase; IAA, iodoacetamide; IPG, immobi-
ized pH gradient; LC–MS/MS, liquid chromatography coupled with tandem masspectrometry; LED, light-emitting diode; MDH, malate dehydrogenase; MS, masspectrometry; MSDB, Mass Spectrometry Protein Sequence Database; Pi, inorganichosphate; PL, pectate lyase; SAHH, S-adenosyl-l-homocysteine hydrolase; SAM,-adenosylmethionine; SDS, sodium dodecyl sulfate; SDS-PAGE, sodium dodecylulfate polyacrylamide gel electrophoresis.∗ Corresponding author at: Departamento de Alimentos e Nutric ão Experimental,CF, Universidade de São Paulo, Av. Lineu Prestes 580, Bloco 14, CEP 05508-900, Sãoaulo, SP, Brazil. Tel.: +55 11 30913647; fax: +55 11 38154410.
E-mail address: [email protected] (J.R.O. do Nascimento).
925-5214/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.postharvbio.2012.04.005
© 2012 Elsevier B.V. All rights reserved.
the world. In developing countries, the consumption of bananas is21 kg per person per year (Arias et al., 2003), and this fruit providesnot only calories but also other important components for humannutrition, such as vitamins, minerals, antioxidants, and prebiotics(Der Agopian et al., 2008; Prasanna et al., 2007; White, 2002).
As a climacteric fruit, the physico-chemical changes that occurduring banana ripening are fast, resulting in a notably perish-able fruit. Ripening is a genetically programmed, coordinated andirreversible phenomenon, which leads to the desirable qualityattributes of edible fruit (Prasanna et al., 2007). However, thesechanges also increase the susceptibility of the fruit to physicalinjury and decrease the resistance of the fruit to microbial infec-tion (Giovannoni, 2001). Therefore, a better understanding of theprocess of fruit ripening may help to improve the nutritional andsensorial quality of fruit and reduce the postharvest fruit losses.In the case of banana this knowledge would be of primary rele-vance.
Investigation of gene expression during fruit ripening at thetranscriptional level provides important information, but the abun-dance of mRNA is not always correlated to the abundance ofexpressed proteins (Chen and Harmon, 2006). Because proteins

5 logy a
mctfdttc
t22ehpppo(oitairAmu2
ibctcbd2Smt
2
2
oki(afitirfa
2
tce
2 T.T. Toledo et al. / Postharvest Bio
ay be considered the effectors of gene expression, and they playentral roles in the regulation of metabolism, an investigation ofhe proteins that are functional during ripening may provide cluesor the activation/deactivation of metabolic pathways and for theevelopment of the phenotypes of edible fruits. In this context, pro-eomics can contribute to the understanding of ripening throughhe identification of the proteins that are affected during this pro-ess.
Recently, studies concerning the proteomics of the ripening ofhe following fruits have been reported: tomato (Faurobert et al.,007; Rocco et al., 2006), grape (Giribaldi et al., 2007; Lücker et al.,009; Negri et al., 2008; Zhang et al., 2008), and strawberry (Biancot al., 2009). However, in the case of banana, only two studiesave been performed on the proteome of the meristem of thelant (Carpentier et al., 2007; Samyn et al., 2006). The analysis ofroteins from banana fruit pulp is limited to the separations ofroteins that are extracted from the fruit at four different stagesf maturation. Submitting these samples to 2D electrophoresisDominguez-Puigjaner et al., 1992) resulted in the identificationf only five proteins of the same molecular weight, which weredentified by immunoblotting to be polygalacturonase-related pro-eins. The differential analysis of banana fruit proteome wouldllow the identification of the proteins changing in abundance dur-ng ripening, which would help to discriminate those putativelyesponsible for the desirable quality attributes of the ripe fruit.t the same time, it may allow access to the post-translationalodification of proteins, which constitutes another level of reg-
lation of many enzymes or regulatory proteins (Mann and Jensen,003).
To get information on the ripening process of banana, whichs a commercially relevant but non-model fruit, the proteomes ofanana pulp at the pre-climacteric and climacteric stages wereompared using two-dimensional fluorescence difference gel elec-rophoresis (2D-DIGE), and the proteins were sequenced usingoupled mass spectrometry (MS). New data on the proteomics ofanana fruit can complement the few transcriptional and genomicata that are available (Clendennen and May, 1997; Godoy et al.,010; Gupta et al., 2006; Manrique-Trujillo et al., 2007; Medina-uárez et al., 1997) and can enhance our understanding of theolecular basis of banana quality through the identification of pro-
eins that are associated with ripening.
. Materials and methods
.1. Plant material
Mature green bananas (Musa acuminata AAA, cv. Nanicão) werebtained approximately 110 d after anthesis (DAA) at a local mar-et on the day of harvest and were allowed to ripen spontaneouslyn an incubator with controlled temperature (20 ◦C) and humidity90%). Respiration, ethylene production, and the amount of starchnd soluble sugars were monitored on a daily basis for at leastve fruit. Based on physiological and physico-chemical parame-ers, pre-climacteric and climacteric bananas were identified. Threendependent ripening experiments were performed (I, II and III),esulting in three biological replicates of each sample group. Theruit that were sampled were peeled, sliced, frozen in liquid N2,nd pooled before storage at −80 ◦C.
.2. Ethylene and CO2 measurements
Fruit were placed in closed jars for 1 h, and air samples wereaken from their headspaces to measure ethylene and CO2 con-entrations by gas chromatography as described previously (Godoyt al., 2010).
nd Technology 70 (2012) 51–58
2.3. Carbohydrate determinations
Starch determination was performed enzymatically asdescribed by Cordenunsi and Lajolo (1995), and soluble sug-ars were extracted with 80% ethanol at 80 ◦C. The sugars weresubsequently analyzed by high-pressure liquid chromatographywith pulse amperometric detection (HPAE-PAD) using a PA1column (Dionex, Sunnyvale, CA, USA). Total soluble sugars werecalculated to be the sum of glucose, fructose and sucrose values.
2.4. Protein extraction from banana pulp
One gram of ground, frozen pulp from each of the fruitsamples was resuspended in 5 mL of ice-cold extraction buffer(50 mmol L−1 Tris–HCl, pH 8.5; 5 mmol L−1 EDTA; 100 mmol L−1
KCl; 1% (w/v) DTT; and 30% (w/v) sucrose) with protease (P9599)and phosphatase I (P2850) and II (P5726) inhibitor cocktails(Sigma–Aldrich). These samples were subsequently extracted usinga phenol–methanol/ammonium acetate precipitation method(Carpentier et al., 2005). The resulting protein pellet was resus-pended in 1 mL lysis buffer (7 mol L−1 urea; 2 mol L−1 thiourea; 4%CHAPS; and 1% DTT) for 1 h at room temperature, and the contam-inants were removed using the 2-D Clean-Up Kit (GE Healthcare).The purified protein pellet was resuspended in DIGE lysis buffer(8 mol L−1 urea; 2 mol L−1 thiourea; 4% CHAPS; and 10 mmol L−1
Tris–HCl, pH 8.0), and the concentrations of the proteins weredetermined using the 2D-Quant Kit (GE Healthcare).
2.5. Protein separation on 2D-DIGE
Proteins were labeled using the Amersham CyDye DIGE Fluorsminimal dyes kit according to the manufacturer’s instructions (GEHealthcare). Two fluorescent markers, Cy3 and Cy5, were used tolabel the internal standard and the samples, respectively.
To separate proteins in the first dimension, each 24 cm IPGstrip (pH 4–7, GE Healthcare) was rehydrated using a mixture of50 �g of Cy3-labeled internal standard and 50 �g of a specific Cy5-labeled sample in a maximum of 450 �L of rehydration solution(7 mol L−1 urea; 2 mol L−1 thiourea; 1% DTT; 4% CHAPS, containinga 0.5% mixture of ampholytic carriers, pH 4–7; and a trace of bro-mophenol blue). Rehydration was performed passively, and IEF wasperformed using an Ettan IPGphor II (GE Healthcare) at 20 ◦C and atotal current of 2 mA with the following program: 500 V for 1 min,3500 V for 1 h 30 min and 3500 V for 16 h 20 min.
After being run, the IPG strips were incubated for 15 min inequilibration buffer (75 mmol L−1 Tris–HCl, pH 8.8; 6 mol L−1 urea;29.3% glycerol; 2% SDS; and a trace of bromophenol blue) with1% DTT added for protein reduction followed by another 15 minin equilibration buffer with 2.5% iodoacetamide (IAA) added forprotein alkylation. SDS-PAGE separations were performed on large12.5% polyacrylamide gels using 25 mmol L−1 Tris–glycine (pH 8.3)running buffer containing 0.1% SDS. The electrophoretic runs werecarried out at 150 V and 12 mA per gel until the bromophenol blueline reached the bottom of the gels. One DIGE gel was run for eachgroup of fruit of the three ripening experiments (I, II and III). Thepreparative 2DE gels for spot picking were prepared essentially inthe same way as described above except that IPG strips were loadedwith 1 mg of non-labeled protein and extracted, and the proteinspots were revealed by colloidal Coomassie Blue (G-250) stainingaccording to Neuhoff et al. (1985).
2.6. Image analysis
The gels were scanned using a Versa Doc 4000 MP (Bio-Rad) apparatus, and images from the samples that had been

logy a
flgMu6Aotoptoitccoo0
2
taA5mswvrbat(wl
2
WHwuCcdFt1f
uMtcMia(fitwu
T.T. Toledo et al. / Postharvest Bio
uorescently labeled with Cy3 were acquired by excitation withreen LED (553 ± 3 nm) and an emission filter of 605 nm ± 50 BP.eanwhile, the samples labeled with Cy5 were visualized
sing red LED excitation (648 ± 3 nm) and an emission filter of95 nm ± 55 BP. The images were analyzed using the PDQuestdvanced software (v.8.0.1) and edited as follows. First, the imagesf each gel were cropped, similarly positioned, and grouped withheir respective internal standards in order to create a matched setf the two groups of fruit. To detect spots, automatic matching waserformed, and the spots were confirmed manually. Spot intensi-ies were normalized in relation to the respective internal standardf each gel and were presented as parts per million of the totalntegrated optical density of the spots. Student’s t-test was usedo evaluate significant variations, and the spots above the 1.5-fold-hange threshold were determined to be significant. Therefore thelimacteric/pre-climacteric intensity ratio above 1.5 was indicativef a protein more abundant at the climacteric, while the proteinsf decreased abundance were those with intensity ratio less than.66.
.7. In-gel protein digestion
The differential protein spots were excised from the prepara-ive gels and destained in 50% acetonitrile (ACN) and 25 mmol L−1
mmonium bicarbonate. The gel plugs were dehydrated with 100%CN and rehydrated with 20 mmol L−1 DTT. After incubation at6 ◦C for 40 min, the suspension was centrifuged, and the gel frag-ents were placed in a 55 mmol L−1 iodoacetamide solution. The
uspensions were kept in the dark for 30 min, and the gel piecesere dehydrated again with 100% ACN. After removing the sol-
ent, the gel pieces were allowed to air dry and were subsequentlyehydrated with a 10 �g L−1 of trypsin in 25 mmol L−1 ammoniumicarbonate solution. The tubes were maintained at 37 ◦C for 12 h,nd the peptides were extracted from the gel pieces by incubatinghem twice with 50 �L of a 60% (v/v) ACN and 1% (v/v) formic acidFA) solution and once with 50 �L of ACN. The three supernatantsere combined and vacuum-dried, and the peptide-containing pel-
ets were re-suspended in 0.1% (v/v) FA for MS analysis.
.8. Protein identification by LC–MS/MS
Peptide mixtures were sequenced using a LC–MS/MS in aaters nanoAcquity UPLC that was coupled to a Waters SYNAPT
DMS mass spectrometer (Q-TOF geometry). The UPLC systemas equipped with a Waters Symmetry C18 desalination col-mn (20 mm × 180 �m; 5 �m particle size) followed by a Waters18 BEH130 analytical column (100 mm × 100 �m; 1.7 �m parti-le size). The samples were injected into the column trap and wereesalted for 3 min with a solution of 97:3 H2O/ACN and 0.1% (v/v)A at a flow rate of 83 nL s−1. The proteins were eluted from therap column and directed to the analytical column at a flow rate of7 nL s−1 with a gradient of 97:3–30:70 H2O/ACN and 0.1% (v/v) FAor a total analysis time of 60 min.
After LC–MS/MS, RAW files containing the data were processedsing the ProteinLynx Global Server (version 2.2, Waters) and theascot MS/MS Ion Search (www.matrixscience.com). To obtain
he possible identity of the proteins, the Mascot search engineompared the obtained sequences with those deposited into theSDB. The search of the database was performed using the follow-
ng parameters: a maximum of one missed trypsin cleavage wasllowed, a taxonomy of Viridiplantae was used, carbamidomethylC) was a fixed modification, oxidation (M) was a variable modi-
cation, 2+ ion charge, values of monoisotopic masses, a peptideolerance of 0.1 Da, a fragment tolerance of 0.1 Da, the instrumentas a ESI-QUAD-TOF, and the micromass data format (.pkl) wassed.nd Technology 70 (2012) 51–58 53
3. Results
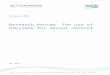
The typical phenotype change that occurred in ripening bananascould be observed in all three experiments, which enabled the def-inition of two well-characterized groups for differential proteomeprofiling. The group of pre-climacteric and unripe bananas wascomposed of fruit that had been sampled one day after harvest inall three experiments when respiration and endogenous ethylenewere low and the starch concentration was high. In contrast, theclimacteric bananas were sampled at the respiration burst whensoluble sugars accumulated, which corresponded to 21 d for exper-iment I and 15 d after harvest for experiments II and III. As can beobserved in Fig. 1, the samples that were classified in the samephysiological group had similar levels of respiration, starch andsugars. When considering the commercial peel color index, the fruitthat were compared in this study were between indices 1 and 5.
The 2DE maps revealed that the protein spots resolved on thegels were well distributed between 97 kDa and 14.4 kDa and werealong the range of pH 4–7 (Fig. 2). Except for a group of proteinsat approximately 30 kDa that were more intense in pre-climactericfruit, the protein maps obtained from the two groups of fruit werequite similar. Overall, the 2DE separations of banana extracts werecomparable to the differential analysis because no vertical or hori-zontal streaks were apparent.
The DIGE approach allowed for the detection of 168 validatedspots of banana proteins, and 50 of those proteins were selectedby Student’s t-test to be significantly above the 1.5-fold threshold(Fig. 2). Among those spots, 5 were present only in pre-climactericfruit, and 9 spots were exclusive to the climacteric group. Aftersequencing the 50 spots, only 26 were identified by comparingtheir sequences with the MSDB using the Mascot search engine asis presented in Table 1. No significant similarity was found to theremaining 24 proteins.
Nineteen spots corresponded to sequences from M. acuminata,and 6 were more similar to the sequences from other plants.The majority of the Musa proteins (13 spots) were related to twochitinases, while the other proteins were involved in metabolismof carbohydrate and ethylene, secondary products or heat-shockproteins. Among the Musa sequences that corresponded to the pro-tein spots, there were also three ripening-associated proteins ofunknown function detected in the pulp of ripe fruit.
Only one class III chitinase was found in the down-regulatedspots or those detected only in the pre-climacteric fruit. In contrast,the other putative chitinase and proteins involved in cell wall disas-sembly, carbohydrate metabolism, ethylene production, synthesisof secondary metabolites or the defense/stress response accumu-lated during ripening. Overall, the protein changes revealed by DIGEwere in good agreement with the previous reports on the profilesof transcripts for the corresponding genes in ripening bananas.
4. Discussion
Considering the importance of gene expression for the regula-tion of metabolic and physico-chemical changes in ripening fruitsand the pivotal role of proteins in this process, this study aimedto provide a differential map of the proteins associated with theripening of bananas. The comparison of two contrasting physiolog-ical stages by 2DE-DIGE revealed the most significant differencesin protein abundance in banana pulp.
Chitinases were the most abundant proteins in the pulp ofunripe bananas, and 11 30-kDa isoforms of the class III acidic
chitinase were identified. These proteins are present in variousorganisms, and their main function is in defense against pathogens,although they have also been implicated in growth and develop-ment processes (Kasprzewska, 2003). Differential accumulation of
54
T.T. Toledo
et al.
/ Postharvest
Biology and
Technology 70
(2012) 51–58
Table 1List of proteins identified.
Spot no. Protein name Organism Scorea Accession code Peptides number Th/Expb pI Th/Expb Mr Ratio (C/PC)c
1 Ripening-associated protein (fragment) Musa acuminata (banana) 194 O22322 MUSAC 7 6.70/6.94 21155/21341 2.54204 Putative chitinase Musa acuminata (banana) 163 Q8VXF0 MUSAC 5 6.84/6.70 34301/33934 3.04
1101 Class III acidic chitinase Musa acuminata (banana) 522 Q6QUK8 MUSAC 18 7.59/6.48 35275/30439 0.081304 Pectate lyase (fragment) Musa acuminata (banana) 94 Q43783 MUSAC 2 6.51/6.47 44312/43506 N.d.2004 Ripening-associated protein (fragment) Musa acuminata (banana) 255 O22322 MUSAC 5 6.67/6.35 21155/20968 2.992103 Class III acidic chitinase Musa acuminata (banana) 506 Q6QUK8 MUSAC 14 7.59/6.29 35732/30439 0.072202 Isoflavone reductase (fragment) Musa acuminata (banana) 133 Q6DQ90 MUSAC 2 5.38/6.24 20165/35553 5.082203 Putative chitinase Musa acuminata (banana) 121 Q8VXF0 MUSAC 4 6.84/6.23 34301/33934 4.062504 S-Adenosyl-l-homocysteine hydrolase Hordeum vulgare (barley) 496 Q4LB20 HORVU 11 5.81/6.29 49960/63157 2.343101 Class III acidic chitinase Musa acuminata (banana) 416 Q6QUK8 MUSAC 9 7.59/6.13 35732/29508 0.053102 Class III acidic chitinase Musa acuminata (banana) 256 Q6QUK8 MUSAC 5 7.59/6.08 35732/30438 N.d.3301 Malate dehydrogenase Nicotiana tabacum (tobacco) 153 Q9FSF0 TOBAC 3 5.91/6.07 35727/44879 1.524101 Class III acidic chitinase Musa acuminata (banana) 546 Q6QUK8 MUSAC 11 7.59/5.91 35732/29970 0.114102 Class III acidic chitinase Musa acuminata (banana) 238 Q6QUK8 MUSAC 9 7.59/5.85 35732/29970 0.054104 Class III acidic chitinase Musa acuminata (banana) 562 Q6QUK8 MUSAC 16 7.59/5.67 35732/29970 0.044505 ADP-glucose pyrophosphorylase small subunit Solanum tuberosum (potato) 71 Q2PXI9 SOLTU 2 6.73/5.75 57545/60282 1.865101 Class III acidic chitinase Musa acuminata (banana) 103 Q6QUK8 MUSAC 3 7.59/5.63 35732/29970 0.055911 Starch phosphorylase Ipomoea batatas (sweet potato) 90 T10947 2 5.26/5.60 109079/105438 N.d.6101 Class III acidic chitinase Musa acuminata (banana) 158 Q6QUK8 MUSAC 5 7.59/5.44 35732/30439 N.d.6103 Class III acidic chitinase Musa acuminata (banana) 93 Q6QUK8 MUSAC 3 7.59/5.40 35732/29970 0.057003 Ripening-associated protein Musa acuminata (banana) 199 O22322 MUSAC 3 6.67/5.24 21155/21296 2.157202 Class III acidic chitinase Musa acuminata (banana) 61 Q6QUK8 MUSAC 3 7.59/5.26 35732/30439 N.d.7302 1-Aminocyclopropane-1-carboxylate oxidase (ACC oxidase) Musa acuminata (banana) 63 Q6LC46 MUSAC 3 5.33/5.18 36256/42836 N.d.8801 High molecular weight heat shock protein Malus domestica (apple) 147 Q9M6R1 MALDO 5 5.17/5.06 71570/79725 12.498802 Heat shock protein 70 Cucumis sativus (cucumber) 201 Q9M4E6 CUCSA 6 5.29/5.01 71183/79725 N.d.8827 Heat shock protein 90 Nicotiana tabacum (tobacco) 171 Q6R0J1 TOBAC 3 4.95/4.99 80358/87511 12.00
a Mascot score for the most significant hits.b Theoretical (Th) and experimental (Exp) isoelectric point (pI) and molecular weight (Mr) of identified proteins.c Ratio of spot intensity at the climacteric (C) and pre-climacteric (PC). Only the spots with ratio values above 1.5 (increased abundance) or less than 0.66 (decreased abundance) were considered. The intensity ratio was not
determined (N.d.) for spots selected by qualitative analysis.

T.T. Toledo et al. / Postharvest Biology and Technology 70 (2012) 51–58 55
0
15
30
45
0,0
0,5
1,0
1,5
0
50
100
150
200
0
1
2
0
10
20
0
50
100
150
200
0
50
100
150
200
0,0
0,5
1,0
1,5
0
10
20
30
5 10 15 200
50
100
150
200
3 6 9 12 15 180
50
100
150
200
3 6 9 12 150
50
100
150
200
B
g k
g-1
�g k
g-1
s-1
A A
B
C
D
A
B
C
D
ng k
g-1
s-1
C
Experiment II
DaysDaysDays
Experiment I Experiment III
g k
g-1
D
F , �), rf D of t
tbntatiriegGir
aitcai
ig. 1. Characterization of ripening bananas. From top to bottom: ethylene (Part Arom experiments I, II, and III during post-harvest ripening. Error bars indicate the S
ranscripts in bananas has been reported (Clendennen et al., 1997),ut Western blotting revealed that the very abundant, 31 kDa,on-catalytic, class III acidic chitinase decreased during ripening,hereby suggesting that this chitinase serves as a source of aminocids for protein synthesis (Peumans et al., 2002). In contrast, thewo isoforms (spots 204 and 2203) that accumulated during ripen-ng were chitinases that have been implicated in the stress/defenseesponse. In fact, the antifungal activity of chitinases has been foundn fruits, such as fig (Li et al., 2005), grape (Fernandez-Caballerot al., 2009), and papaya (Chen et al., 2007), and two homolo-ous chitinases that inhibit Fusarium oxysporum were isolated fromold bananas (Ho and Ng, 2007). Therefore, the two up-regulated
soforms revealed in this study could be important in pathogenesistance or in adaptation to postharvest conditions.
Pectin is the main component of cell walls and middle lamella,nd its degradation is usually noted as the cause of fruit soften-ng. Since pectate lyase (PL, EC 4.2.2.2) catalyzes the breaking of
he glucosidic bonds of the pectin chain (Yadav et al., 2009), itould contribute to polymer solubilization and fruit softening. Theccumulation of protein (i.e., spot 1304) is in agreement with thencrease in transcript levels and activity in the peel and pulp ofespiration (Part B, �), starch (Part C, ©), and soluble sugars (Part D, �) of bananashe mean (n = 5).
climacteric bananas (Gupta et al., 2006; Kesari et al., 2007; Payasiand Sanwal, 2003), but the relevance of pectate lyase to fruit soft-ening remains to be established. According to Shiga et al. (2011)the softening of Nanicão and Mysore bananas seems to be moredependent upon starch levels than to cell wall changes but in acultivar-dependent manner. If this possibility were true, pectatelyase expression could play a minor role in the process of bananapulp softening.
Besides contributing to fruit softening, starch degradation isimportant for the sweetness of ripe fruit, which occurs becauseof the concerted action of amylolytic and phosphorolytic enzymes(Da Mota et al., 2002; Mainardi et al., 2006). Starch phosphory-lase (EC 2.4.1.1) catalyzes the reversible conversion of �-1,4-glucanand Pi into glucose-1-phosphate, and the presence of spot 5911in climacteric fruit is in agreement with the proposed role of thisprotein in starch degradation and with the regulation of its activ-ity level by protein synthesis. However, this protein spot showed
identity to the sweet potato (T10947), maize (CAB69360) and riceenzyme (Q9ATK9 ORYSA), among others, but not to the banana(AY463025) enzyme that had been previously studied (Mainardiet al., 2006). This finding could indicate that a new isoform has been
56 T.T. Toledo et al. / Postharvest Biology and Technology 70 (2012) 51–58
Fig. 2. A 2DE map of banana fruit. (A) Representative proteome revealed by the image of a Cy3-labeled, internal standard that was pooled from the protein extracts frompre-climacteric and climacteric fruit focused from pH 4 to 7 and separated with 12.5% SDS-PAGE. The fifty protein spots that showed abundance changes during ripeningare indicated by the arrows and identified by numbers attributed by PDQuest. The pH gradient is indicated at the top, and the positions of the protein markers are indicatedby numbers on the left (97–14.4 kDa). (B) Highlights of the 26 differential spots identified by comparing the sequences of those proteins with the MSDB using the Mascotsearch engine. The images from the internal standard (Cy3, in blue), and pre-climacteric (PC) or climacteric (C) extracts (Cy5, in red) are superimposed. The more abundantt ear. Tb e boxr
fi
brsgpbrdA
he protein is in relation to the internal standard, the more reddish the spot will appy an arrow ( ) or letters (a or b) when other unrelated spots are also visible in theferred to the web version of the article.)
ound, which could contribute to the ethylene-responsive increasen enzyme activity at the beginning of starch degradation.
Because degradation of starch is a major occurrence duringanana ripening, the accumulation of ADP-glucose pyrophospho-ylase (AGPase, EC 2.7.7.27, spot 4505), which participates in theynthesis of starch through the formation of ADP-glucose fromlucose-1-P and ATP (Martin and Smith, 1995), was an unex-ected result. However, although the presence of AGPase would
e more consistent with an early developmental stage, the occur-ence of concomitant starch degradation and synthesis has beenescribed in ripening banana (Hill and Rees, 1994). Variations ofGPase expression have also been reported in watermelon (Kimhe differential spots are designated by their identification numbers and pointed toes. (For interpretation of the references to color in this figure legend, the reader is
et al., 1998), decreasing from the small-green to the medium-greenstage and increasing during ripening. The accumulation of bananaAGPase could reflect an adjustment of the biosynthetic capabilityof the fruit through the levels of some enzymes in order to sustainthe occurrence of concomitant synthesis and degradation of starch.
The amounts of organic acids influence banana flavor, andmalic acid is the main organic acid in the fruit pulp (Sweetmanet al., 2009). Although malic acid can be a respiratory substrate
(Tucker, 1993), climacteric fruits, such as bananas and mangoes,seem to not use it during their respiratory peaks (Sweetman et al.,2009). A malate dehydrogenase (MDH, EC 1.1.1.37, spot 3301) wasmore abundant at the climacteric stage, suggesting that it was
logy a
dcc
e7itdtSeciSmgis(e
wet22Bsin
trsrfiwaHbbeetitii
5
riWptsbopifr
T.T. Toledo et al. / Postharvest Bio
ifferentially expressed. Since this protein catalyzes the reversibleonversion of oxaloacetate to malate, MDH accumulation couldontribute to the development of the ripe flavor of banana.
Ethylene is an important regulator of banana ripening, and thethylene biosynthetic enzyme ACC oxidase was detected in spot302. The substrate of ACC oxidase is produced by the lyase activ-
ty of ACC synthase on S-adenosylmethionine (SAM), which is alsohe limiting step of the pathway. This compound is a major methylonor group in several reactions, but a byproduct of methylation ishe S-adenosyl-l-homocysteine, which is a competitive inhibitor ofAM-dependent methyltransferases (Brzezinski et al., 2008). How-ver, S-adenosyl-l-homocysteine hydrolase (SAHH, spot 2504)atalyzes the reversible hydrolysis of S-adenosyl-l-homocysteinento adenosine and l-homocysteine, which can be regenerated toAM through l-methionine and sustains the ACC synthesis andethylation processes. The accumulation of SAHH in bananas sug-
ests that it may be in demand because of the increased use of SAMn ethylene synthesis and DNA methylation during the climacterictage because this process is a mechanism for suppressing genesGibney and Nolan, 2010) and may be important in regulating genexpression during ripening.
Three heat shock proteins (HSPs) of different molecular weightsere abundant in ripe fruit (spots 8801, 8802, and 8827). The differ-
nt classes of HSPs are involved in responses to stress in plants, buthey have also been observed during ripening of fruits (Bianco et al.,009; Deytieux et al., 2007; Faurobert et al., 2007; Giribaldi et al.,007; Guarino et al., 2007; Muccilli et al., 2009; Negri et al., 2008).ecause these proteins are usually involved in stabilizing proteintructures during plant development in non-stressful conditions,t is likely that they are needed for the stabilization of ancient andewly synthesized proteins during ripening.
Isoflavone reductase (IFR) is a secondary metabolism enzymehat is also involved in the stress response that occurs duringipening (spot 2202). IFR catalyzes the key step in the biosynthe-is of isoflavonoids, which are compounds produced by plants inesponse to biotic and abiotic stresses. Differential proteomics con-rmed the induction of IFR as was suggested by Gupta et al. (2006),ho found increased transcription of the IFR gene during ripening
nd suggested that this gene was highly responsive to ethylene.owever, accumulation of IFR is not limited to the climacteric stageecause IFR from non-climacteric fruits, such as grapes and straw-erries, also increased during ripening (Bianco et al., 2009; Giribaldit al., 2007; Negri et al., 2008; Sarry et al., 2004). Due to the ben-fits of isoflavonoids to human health, detection of the enzymehat participates in the biosynthesis of these compounds in fruits ismportant not only for improving the stress response but becausehis enzyme may also have biotechnological applications, such asn the genetic improvement of fruits to have higher contents ofsoflavonoids.
. Concluding remarks
This study reports the first proteomic analysis of banana fruitipening and has identified some proteins that may be involvedn the biochemical processes that are important for fruit quality.
e have identified changes in storage, defense and biosyntheticroteins that could be involved in development of banana flavor,exture, defense, synthesis of ethylene, regulation of gene expres-ion and protein folding. Differential analyses of protein maps fromanana pulp using 2D-DIGE technology provided new information
n the ripening-associated changes in protein levels and validatedrevious data concerning the amounts of transcripts produced dur-ng banana ripening. In this regard, the differential proteomics ofruit pulp has enlarged our understanding of the process of bananaipening.
nd Technology 70 (2012) 51–58 57
Acknowledgments
The authors thank Fundac ão de Amparo à Pesquisa do Estadode São Paulo (FAPESP) for providing financial support (Project2008/52447-0) and Conselho Nacional de Desenvolvimento Cien-tífico e Tecnológico (CNPq) for providing scholarships to T.T.T. andS.B.N. The authors also thank Dr. Luciano Paulino da Silva and Dr.Marcelo Porto Bemquerer from “Embrapa Recursos Genéticos eBiotecnologia” for training T.T.T. and S.B.N. in mass spectrometryanalysis.
References
Arias, P., Dankers, C., Liu, P., Pilkauskas, P., 2003. The World Banana Economy.Food and Agriculture Organization of the United Nations, Rome, 1985–2002.http://www.fao.org/kids/en/index.html (accessed 06.07.10).
Bianco, L., Lopez, L., Scalone, A.G., Di Carli, M., Desiderio, A., Benvenuto, E., Perrotta,G., 2009. Strawberry proteome characterization and its regulation during fruitripening and in different genotypes. Journal of Proteomics 72, 586–607.
Brzezinski, K., Bujacz, G., Jaskolski, M., 2008. Purification, crystallization and pre-liminary crystallographic studies of plant S-adenosyl-l-homocysteine hydrolase(Lupinus luteus). Acta Crystallographica 64, 671–673.
Carpentier, S.C., Witters, E., Laukens, K., Deckers, P., Swennen, R., Panis, B., 2005.Preparation of protein extracts from recalcitrant plant tissues: an evaluation ofdifferent methods for two-dimensional gel electrophoresis analysis. Proteomics5, 2497–2507.
Carpentier, S.C., Witters, E., Laukens, K., Onckelen, H.V., Swennen, R., Panis, B., 2007.Banana (Musa spp.) as a model to study the meristem proteome: acclimation toosmotic stress. Proteomics 7, 92–105.
Chen, S., Harmon, A.C., 2006. Advances in plant proteomics. Proteomics 6,5504–5516.
Chen, Y., Hsu, L., Huang, I., Tsai, T., Lee, G., Shaw, J., 2007. Gene cloning and char-acterization of a novel recombinant antifungal chitinase from papaya (Caricapapaya). Journal of Agricultural and Food Chemistry 55, 714–722.
Clendennen, S.K., May, C.D., 1997. Differential gene expression in ripening bananafruit. Plant Physiology 115, 463–469.
Clendennen, S.K., Lopez-Gomez, R., Gomez-Lim, M., Arntzen, C.J., May, G.D., 1997.The abundant 31-kilodalton banana pulp protein is homologous to class-IIIacidic chitinases. Phytochemistry 47, 613–619.
Cordenunsi, B.R., Lajolo, F.M., 1995. Starch breakdown during banana ripening:sucrose synthase and sucrose phosphate synthase. Journal of Agricultural andFood Chemistry 43, 347–351.
Da Mota, R.V., Cordenunsi, B.R., Nascimento, J.R.O., Purgatto, E., Rossetto, M.R.M.,Lajolo, F.M., 2002. Activity and expression of banana starch-phosphorylasesduring fruit development and ripening. Planta 216, 325–333.
Der Agopian, R.G., Soares, C.A., Purgatto, E., Cordenunsi, B.R., Lajolo, F.M., 2008.Identification of fructooligosaccharides in different banana cultivars. Journal ofAgricultural and Food Chemistry 56, 3305–3310.
Deytieux, C., Geny, L., Lapaillerie, D., Claverol, S., Bonneu, M., Donèche, B., 2007. Pro-teome analysis of grape skins during ripening. Journal of Experimental Botany58, 1851–1862.
Dominguez-Puigjaner, E., Vendrell, M., Ludevid, M.D., 1992. Differential proteinaccumulation in banana fruit during ripening. Plant Physiology 98, 157–162.
Faurobert, M., Mihr, C., Bertin, N., Pawlowski, T., Negroni, L., Sommerer, N., Causse,M., 2007. Major proteome variations associated with cherry tomato pericarpdevelopment and ripening. Plant Physiology 143, 1327–1346.
Fernandez-Caballero, C., Romero, I., Gon, O., Escribano, M.I., Merodio, C., Sanchez-Ballesta, M.T., 2009. Characterization of an antifungal and cryoprotective classI chitinase from table grape berries (Vitis vinifera Cv. Cardinal). Journal of Agri-cultural and Food Chemistry 57, 8893–8900.
Gibney, E.R., Nolan, C.M., 2010. Epigenetics and gene expression. Heredity 105, 4–13.Giovannoni, J., 2001. Molecular biology of fruit maturation and ripening. Annual
Review of Plant Physiology and Plant Molecular Biology 52, 725–749.Giribaldi, M., Perugini, I., Sauvage, F., Schubert, A., 2007. Analysis of protein changes
during grape berry ripening by 2-DE and MALDI-TOF. Proteomics 7, 3154–3170.Godoy, A., Cordenunsi, B.R., Lajolo, F.M., Nascimento, J.R.O., 2010. Differential dis-
play and suppression subtractive hybridization analysis of the pulp of ripeningbanana. Scientia Horticulturae 124, 51–56.
Gupta, S.M., Srivastava, S., Sane, A.P., Nath, P., 2006. Differential expression of genesduring banana fruit development, ripening and 1-MCP treatment: presenceof distinct fruit specific, ethylene induced and ethylene repressed expression.Postharvest Biology and Technology 42, 16–22.
Guarino, C., Arena, S., De Simone, L., D’Ambrosio, C., Santoro, S., Rocco, M., Scaloni, A.,Marra, M., 2007. Proteomic analysis of the major soluble components in Annurcaapple flesh. Molecular Nutrition & Food Research 51, 255–262.
Hill, S.A., Rees, T.ap., 1994. Fluxes of carbohydrate metabolism in ripening bananas.
Planta 192, 52–60.Ho, V.S.M., Ng, T.B., 2007. Chitinase-like proteins with antifungal activity fromemperor banana fruits. Protein and Peptide Letters 14, 828–831.
Kasprzewska, A., 2003. Plant chitinases – regulation and function. Cellular andMolecular Biology Letters 8, 809–824.

5 logy a
K
K
L
L
M
M
M
MM
M
N
N
Yadav, S., Yadav, P.K., Yadav, D., Yadav, K.D.S., 2009. Pectin lyase: a review. Process
8 T.T. Toledo et al. / Postharvest Bio
esari, R., Trivedi, P.K., Nath, P., 2007. Ethylene-induced ripening in banana evokesexpression of defense and stress related genes in fruit tissue. Postharvest Biologyand Technology 46, 136–143.
im, I.J., Kahng, H.Y., Chung, W.I., 1998. Characterization of cDNAs encoding smalland large subunits of ADP-glucose pyrophosphorylases from watermelon (Cit-rullus vulgaris S.). Bioscience, Biotechnology and Biochemistry 62, 550–555.
i, Y., Yang, Y., Hsu, J.S.F., Wu, D., Wu, H., Tzen, J.T.C., 2005. Cloning and immunolo-calization of an antifungal chitinase in jelly fig (Ficus awkeotsang) achenes.Phytochemistry 66, 879–886.
ücker, J., Laszczak, M., Smith, D., Lund, S.T., 2009. Generation of a predicted proteindatabase from EST data and application to iTRAQ analyses in grape (Vitis Viniferacv. Cabernet Sauvignon) berries at ripening initiation. BMC Genomics 10, 50–67.
ainardi, J.A., Purgatto, E., Vieira Jr., A., Bastos, W.A., Cordenunsi, B.R., Nascimento,J.R.O., Lajolo, F.M., 2006. Effects of ethylene and 1-methylcyclopropene (1-MCP)on gene expression and activity profile of r-1,4-glucan-phosphorylase duringbanana ripening. Journal of Agricultural and Food Chemistry 54, 7294–7299.
ann, M., Jensen, O.N., 2003. Proteomic analysis of post-translational modifications.Nature Biotechnology 21, 255–261.
anrique-Trujillo, S.M., Ramírez-López, A.C., Ibarra-Laclete, E., Gómez-Lim, M.A.,2007. Identification of genes differentially expressed during ripening of banana.Journal of Plant Physiology 164, 1037–1050.
artin, C., Smith, A.M., 1995. Starch biosynthesis. The Plant Cell 7, 971–985.edina-Suárez, R., Manning, K., Fletcher, J., Aked, J., Bird, C.R., Seymour, C.B., 1997.
Gene expression in the pulp of ripening bananas’ two-dimensional sodiumdodecyl sulfate-polyacrylamide gel electrophoresis of in vitro translation prod-ucts and cDNA cloning of 25 different ripening-related mRNAs. Plant Physiology115, 453–461.
uccilli, V., Licciardello, C., Fontanini, D., Russo, M.P., Cunsolo, V., Saletti, R., Recu-pero, G.R., Foti, S., 2009. Proteome analysis of Citrus sinensis L. (Osbeck) flesh atripening time. Journal of Proteomics 73, 134–152.
egri, A.S., Prinsi, B., Rossoni, M., Failla, O., Scienza, A., Cocucci, M., Espen, L., 2008.
Proteome changes in the skin of the grape cultivar Barbera among differentstages of ripening. BMC Genomics 9, 378.euhoff, V., Stamm, R., Eibl, H., 1985. Clear background and highly sensitive proteinstaining with Coomassie Blue dyes in polyacrylamide gels: a systematic analysis.Electrophoresis 6, 427–448.
nd Technology 70 (2012) 51–58
Prasanna, V., Prabha, T.N., Tharanathan, R.N., 2007. Fruit ripening phenomena – anoverview. Critical Reviews in Food Science and Nutrition 47, 1–19.
Payasi, A., Sanwal, G.G., 2003. Pectate lyase activity during ripening of banana fruit.Phytochemistry 63, 243–248.
Peumans, W.J., Proost, P., Swennen, R.L., Van Damme, E.J.M., 2002. The abundant classIII chitinase homolog in young developing banana fruits behaves as a transientvegetative storage protein and most probably serves as an important supply ofamino acids for the synthesis of ripening-associated proteins. Plant Physiology130, 1063–1072.
Rocco, M., D’ambrosio, C., Arena, S., Faurobert, M., Scaloni, A., Marra, M., 2006. Pro-teomic analysis of tomato fruits from two ecotypes during ripening. Proteomics6, 3781–3791.
Samyn, B., Sergeant, K., Carpentier, S., Debyser, G., Panis, B., Swennen, R., Beeumen,J.V., 2006. Functional proteome analysis of the banana plant (Musa spp.) using deNovo sequence analysis of derivatized peptides. Journal of Proteome Research6, 70–80.
Sarry, J.-E., Sommerer, N., Sauvage, F.-X., Bergoin, A., Rossignol, M., Albagnac, G.,Romieu, C., 2004. Grape berry biochemistry revisited upon proteomic analysisof the mesocarp. Proteomics 4, 201–215.
Shiga, T.M., Soares, C.A., Nascimento, J.R., Purgatto, E., Lajolo, F.M., Cordenunsi, B.R.,2011. Ripening-associated changes in the amounts of starch and non-starchpolysaccharides and their contributions to fruit softening in three banana culti-vars. Journal of the Science of Food and Agriculture 91, 1511–1516.
Sweetman, C., Deluc, L.G., Cramer, G.R., Ford, C.M., Soole, K.L., 2009. Regulation ofmalate metabolism in grape berry and other developing fruits. Phytochemistry70, 1329–1344.
Tucker, G.A., 1993. Introduction. In: Seymour, G.B., Taylor, J.E., Tucker, G.A. (Eds.),Biochemistry of Fruit Ripening. Chapman & Hall, London, pp. 1–43.
White, P.J., 2002. Recent advances in fruit development and ripening: an overview.Journal of Experimental Botany 53, 1995–2000.
Biochemistry 44, 1–10.Zhang, J., Ma, H., FenG., J., Zeng, L., Wang, Z., Chen, S., 2008. Grape berry plasma
membrane proteome analysis and its differential expression during ripening.Journal of Experimental Botany 59, 2979–2990.