Embed Size (px)
Citation preview

Thrombin peptide, TP508, stimulates angiogenic responses in animal
models of dermal wound healing, in chick chorioallantoic membranes,
and in cultured human aortic and microvascular endothelial cells
Andrea M. Norfleet, John S. Bergmann, Darrell H. Carney*
Chrysalis BioTechnology, Inc., 2200 Market Street, Suite 600, Galveston, TX, USA
Department of Human Biological Chemistry and Genetics, University of Texas Medical Branch, Galveston, TX, USA
Received 1 September 2001; accepted 1 October 2001
Abstract
The a-thrombin peptide, TP508, accelerates the healing of full-thickness wounds in both normal and ischemic skin. In wounds treated
with TP508, a pattern of increased vascularization is consistently observed both grossly and microscopically when compared to wounds
treated with saline. One possible mechanism by which the peptide accelerates wound healing is by promoting revascularization of granulation
tissue at the injured site. To evaluate the angiogenic potential of TP508, the peptide was tested in the chick embryo chorioallantoic membrane
(CAM), where it increased the density and size of CAM blood vessels relative to controls. Additionally, TP508 stimulated chemokinesis and
chemotaxis in a dose-dependent fashion in cultured human aortic and human microvascular endothelial cells. Taken together, these in vivo
and in vitro data support an angiogenic role for TP508 in wound healing. A working model is presented to explain how this 23-amino-acid
peptide, which lacks proteolytic activity, is generated during wound healing and contributes to the nonproteolytic functions associated with
a-thrombin during tissue repair. D 2002 Elsevier Science Inc. All rights reserved.
Keywords: Growth factors; Tissue repair; Synthetic peptides; Hemostasis
1. Cellular responses to A-thrombin: proteolytic and
nonproteolytic mechanisms
The functions of A-thrombin in response to an injury are
complex, evolving in a dynamic—and sometimes contra-
dictory—fashion as the microenvironment of the wound
changes over the course of healing (Grand et al., 1996;
Strukova, 2001). Initially, for example,A-thrombin is pivotal
in formation of the fibrin clot, aggregation of platelets, and
stimulation of proinflammatory responses. Later,A-thrombin
triggers events that limit its own formation, lead to dissolu-
tion of the clot, and contribute to cessation of inflammatory
cell activity. We are just beginning to gain an understanding
of howA-thrombin achieves this type of functional versatility
and how the diverse effects of thrombin contribute to the
intricate network of responses required for tissue repair.
One facet of A-thrombin’s multifunctionality is related to
its well-characterized activity as a serine protease. In this ca-
pacity, A-thrombin utilizes its proteolytic activity to catalyze
the hydrolysis of specific substrate molecules to yield unique
products, as exemplified by the cleavage of fibrinogen
to fibrin. In a less classical role, A-thrombin triggers intra-
cellular signaling pathways by hydrolytically removing
a portion of the amino-terminal exodomain of specific
G-protein-coupled receptors, called proteolytically activated
receptors (PARs). Three PAR subtypes (PAR-1, -3, -4) are
activated by A-thrombin via this unique mechanism, which
requires enzymatic cleavage of the PAR by the serine
protease (Vergnolle et al., 2001).
Another dimension to the complex biology ofA-thrombin
is a group of functions that are not dependent upon protease
activity. Compelling evidence for such interactions is based
on experiments in which cells respond to A-thrombin even
when the active site of the enzyme is rendered inert by
chemical blockade or genetic mutation (Bar-Shavit et al.,
1983, 1986; Carney et al., 1992a,b; Jenkins et al., 1995).
Moreover, this type of observation has led to the discovery
0306-3623/02/$ – see front matter D 2002 Elsevier Science Inc. All rights reserved.
PII: S0306 -3623 (01 )00118 -5
* Corresponding author. Chrysalis BioTechnology, 2200 Market Street,
Suite 600, Galveston, TX 77550, USA. Tel.: +1-409-750-9251; fax: +1-
409-750-9253.
E-mail address: [email protected] (D.H. Carney).
General Pharmacology 35 (2002) 249–254

and characterization of peptides corresponding to unique
regions of A-thrombin that lie outside the active site, yet
exhibit biological activity in vitro and in vivo (Bar-Shavit
and Wilner, 1986; Glenn et al., 1988).
In our working model of A-thrombin activity, events eli-
cited both by proteolytic cleavage and by nonproteolytic
binding comprise a dual signaling mechanism that is required
to achieve the full physiological response to A-thrombin.
Early studies showed that thrombin stimulation of cell
proliferation required two sets of signals: one generated by
proteolytic cleavage and one generated by high-affinity
interaction of A-thrombin (or a thrombin receptor monoclo-
nal antibody) with specific receptor sites (Carney et al.,
1986). We postulated that the signaling events initiated by
nonproteolytic or high-affinity interactions of thrombin are
mediated by separate receptor proteins, termed nonproteo-
lytically activated receptors or N-PARs. It is notable that
Factor Xa, another serine protease involved in the coagu-
lation cascade, has been found to enzymatically cleave and
activate PAR-2 (Cirino et al., 2000), as well as to specifically
bind and activate the EPR-1 receptor without proteolytically
altering it (Altieri, 1995). Thus, it may be that a number of
coagulation cascade proteases have evolved the ability to
initiate distinct physiological responses by cleavage of one
type of receptor and binding of another, thereby triggering
two completely different sets of intracellular signals and/or
allowing cross-talk between the two types of receptors.
To identify the functional binding domains of the A-
thrombin molecule involved in N-PAR activation and dual
signaling, we synthesized and tested a number of molecules
representing different regions of A-thrombin. We became
interested in a specific 23-amino-acid peptide corresponding
to a highly conserved region of the B-chain of the human
prothrombin sequence that extends from amino acids 508 to
530 that became known as thrombin peptide 508, or TP508.
Although the peptide has no proteolytic activity, we initially
found that the 23-mer competes for high-affinityA-thrombin
binding sites on fibroblasts and enhances proliferation of
fibroblasts co-stimulated with proteolytically active A-
thrombin or molecules that activate downstream signal events
normally generated by PAR-1 activation (Carney et al., 1984,
1986, 1992a,b). TP508 has also proved to be chemotactic for
neutrophils (Moller et al., in press), keratinocytes (Sower et
al., 1999), and, as discussed below, endothelial cells. In an
effort to understand how these effects relate to A-thrombin’s
role in tissue repair following injury, we began to test TP508
in animal models of dermal wound healing.
2. A-Thrombin peptide, TP508, as a possible angiogenic
factor in wound healing
In early studies in our laboratory (Carney et al., 1992a,b),
full-thickness incisions were created in the dorsal skin of
normal rats and immediately treated with a single topical
application of TP508 in saline. By Postwounding Day 7,
TP508-treated incisions exhibited up to 80% greater tensile
strength than saline controls, corresponding to a forward
shift in the time course of healing of over 4 days. Radio-
angiography and histology of the Day 7 wounds revealed
that TP508 treatment augmented revascularization of the
incisional wounds, which we speculated could have con-
tributed to the acceleration of healing.
Further studies using a full-thickness excisional model of
dermal wounding confirmed the ability of TP508 to accel-
erate the healing process (Stiernberg et al., 2000): Open
excisions that had been treated with a single topical applica-
tion of TP508 on Day 0 were 35% smaller than controls by
Day 7. Visual inspection of the underside of the dermis
showed an enhanced network of vessels directed towards
the TP508-treated wounds, compared to the saline-treated
wounds. This gross finding was consistent with microscopic
observations of Day 7 wound histology, which indicated
that the granulation tissue of TP508-treated wounds con-
tained larger blood vessels than corresponding control
wounds. In this study, we also found that between 12- and
24-h postwounding, a greater number of leukocytes had
infiltrated the TP508-treated wounds compared to controls.
In an experimental model of ischemic wound healing
(Norfleet et al., 2000), TP508 was again shown to accelerate
wound closure, to augment neovascularization of the granu-
lating wound tissue, and to enhance early leukocyte recruit-
ment. To produce ischemic wounds, full-thickness excisions
were made within a cranially based pedicle flap, a three-
sided skin flap to which all blood vessels are severed except
along the cranial (attached) edge. This procedure results in a
gradient of blood flow that decreases from the cranial to the
caudal end of the flap—and, therefore, from the cranial to
the caudal margin of the excisional wounds within the skin
flap. The decreased perfusion and concomitant increased
hypoxia at the caudal wound edge, relative to the cranial
edge of a given wound, were associated with the presence of
a greater number of large neovessels in the granulation
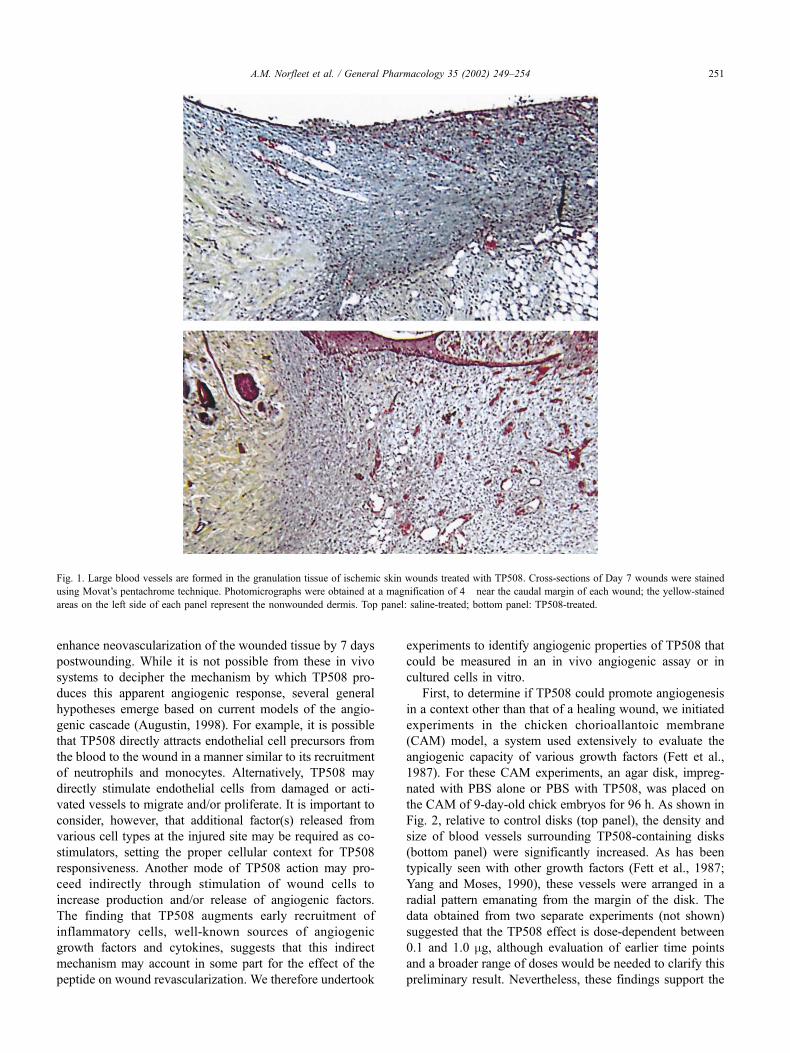
tissue at Postwounding Day 7. As shown in Fig. 1, TP508
treatment (lower panel) produced a further increase in the
number of large vessels in the granulation tissue at the
caudal margin of ischemic wounds as compared to those
treated with saline (upper panel). By Day 10 postwounding,
the numbers and sizes of the blood vessels did not differ
significantly in control and treated wounds. In fact, some
involution of the neovessels at the margins of the wounds
has occurred by this point, demonstrating that TP508 does
not induce an abnormal, persistent angiogenic response. In
this same study, we found that TP508 enhanced the infiltra-
tion of inflammatory cells into the ischemic wounds by 24 h.
Moreover, the granulation tissue of saline-treated ischemic
wounds was punctuated with high numbers of activated
neutrophils 14 days after wounds, while TP508-treated
wounds contained few of these cells, similar to Day 14
wounds in normal (nonischemic) skin.
Taken together, the observations from dermal wound
healing studies indicated that TP508 treatment results in
A.M. Norfleet et al. / General Pharmacology 35 (2002) 249–254250
enhance neovascularization of the wounded tissue by 7 days
postwounding. While it is not possible from these in vivo
systems to decipher the mechanism by which TP508 pro-
duces this apparent angiogenic response, several general
hypotheses emerge based on current models of the angio-
genic cascade (Augustin, 1998). For example, it is possible
that TP508 directly attracts endothelial cell precursors from
the blood to the wound in a manner similar to its recruitment
of neutrophils and monocytes. Alternatively, TP508 may
directly stimulate endothelial cells from damaged or acti-
vated vessels to migrate and/or proliferate. It is important to
consider, however, that additional factor(s) released from
various cell types at the injured site may be required as co-
stimulators, setting the proper cellular context for TP508
responsiveness. Another mode of TP508 action may pro-
ceed indirectly through stimulation of wound cells to
increase production and/or release of angiogenic factors.
The finding that TP508 augments early recruitment of
inflammatory cells, well-known sources of angiogenic
growth factors and cytokines, suggests that this indirect
mechanism may account in some part for the effect of the
peptide on wound revascularization. We therefore undertook
experiments to identify angiogenic properties of TP508 that
could be measured in an in vivo angiogenic assay or in
cultured cells in vitro.
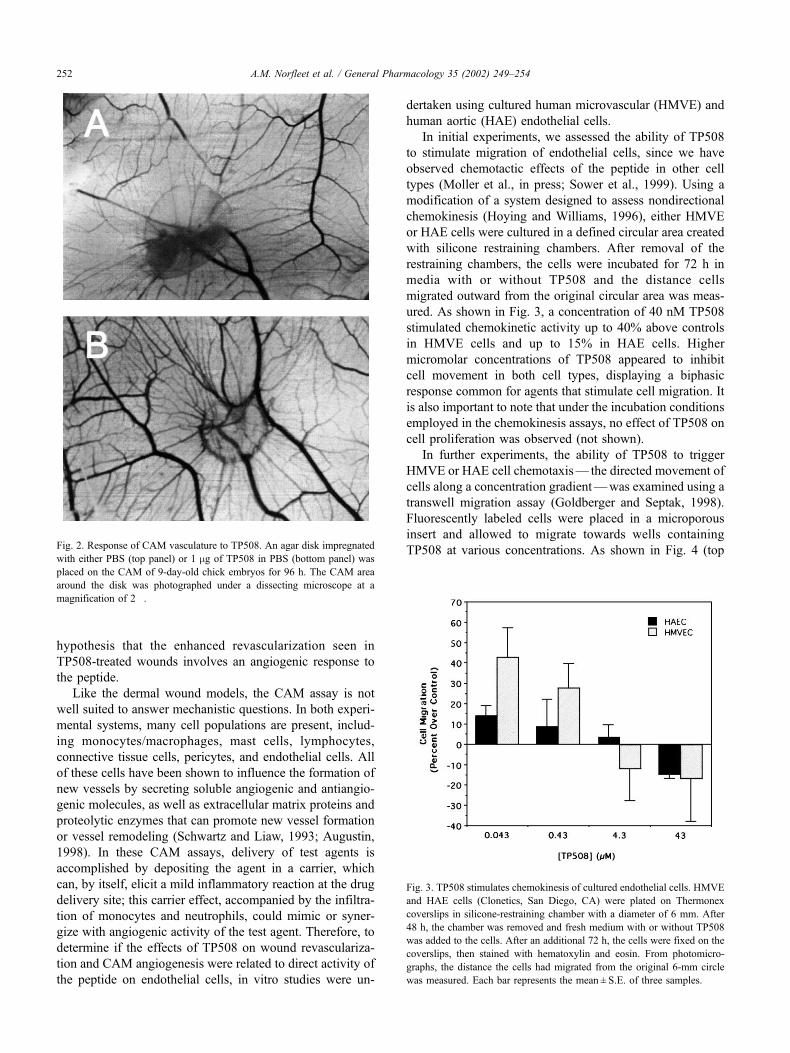
First, to determine if TP508 could promote angiogenesis
in a context other than that of a healing wound, we initiated
experiments in the chicken chorioallantoic membrane
(CAM) model, a system used extensively to evaluate the
angiogenic capacity of various growth factors (Fett et al.,
1987). For these CAM experiments, an agar disk, impreg-
nated with PBS alone or PBS with TP508, was placed on
the CAM of 9-day-old chick embryos for 96 h. As shown in
Fig. 2, relative to control disks (top panel), the density and
size of blood vessels surrounding TP508-containing disks
(bottom panel) were significantly increased. As has been
typically seen with other growth factors (Fett et al., 1987;
Yang and Moses, 1990), these vessels were arranged in a
radial pattern emanating from the margin of the disk. The
data obtained from two separate experiments (not shown)
suggested that the TP508 effect is dose-dependent between
0.1 and 1.0 mg, although evaluation of earlier time points
and a broader range of doses would be needed to clarify this
preliminary result. Nevertheless, these findings support the
Fig. 1. Large blood vessels are formed in the granulation tissue of ischemic skin wounds treated with TP508. Cross-sections of Day 7 wounds were stained
using Movat’s pentachrome technique. Photomicrographs were obtained at a magnification of 4� near the caudal margin of each wound; the yellow-stained
areas on the left side of each panel represent the nonwounded dermis. Top panel: saline-treated; bottom panel: TP508-treated.
A.M. Norfleet et al. / General Pharmacology 35 (2002) 249–254 251
hypothesis that the enhanced revascularization seen in
TP508-treated wounds involves an angiogenic response to
the peptide.
Like the dermal wound models, the CAM assay is not
well suited to answer mechanistic questions. In both experi-
mental systems, many cell populations are present, includ-
ing monocytes/macrophages, mast cells, lymphocytes,
connective tissue cells, pericytes, and endothelial cells. All
of these cells have been shown to influence the formation of
new vessels by secreting soluble angiogenic and antiangio-
genic molecules, as well as extracellular matrix proteins and
proteolytic enzymes that can promote new vessel formation
or vessel remodeling (Schwartz and Liaw, 1993; Augustin,
1998). In these CAM assays, delivery of test agents is
accomplished by depositing the agent in a carrier, which
can, by itself, elicit a mild inflammatory reaction at the drug
delivery site; this carrier effect, accompanied by the infiltra-
tion of monocytes and neutrophils, could mimic or syner-
gize with angiogenic activity of the test agent. Therefore, to
determine if the effects of TP508 on wound revasculariza-
tion and CAM angiogenesis were related to direct activity of
the peptide on endothelial cells, in vitro studies were un-
dertaken using cultured human microvascular (HMVE) and
human aortic (HAE) endothelial cells.
In initial experiments, we assessed the ability of TP508
to stimulate migration of endothelial cells, since we have
observed chemotactic effects of the peptide in other cell
types (Moller et al., in press; Sower et al., 1999). Using a
modification of a system designed to assess nondirectional
chemokinesis (Hoying and Williams, 1996), either HMVE
or HAE cells were cultured in a defined circular area created
with silicone restraining chambers. After removal of the
restraining chambers, the cells were incubated for 72 h in
media with or without TP508 and the distance cells
migrated outward from the original circular area was meas-
ured. As shown in Fig. 3, a concentration of 40 nM TP508
stimulated chemokinetic activity up to 40% above controls
in HMVE cells and up to 15% in HAE cells. Higher
micromolar concentrations of TP508 appeared to inhibit
cell movement in both cell types, displaying a biphasic
response common for agents that stimulate cell migration. It
is also important to note that under the incubation conditions
employed in the chemokinesis assays, no effect of TP508 on
cell proliferation was observed (not shown).
In further experiments, the ability of TP508 to trigger
HMVE or HAE cell chemotaxis— the directed movement of
cells along a concentration gradient—was examined using a
transwell migration assay (Goldberger and Septak, 1998).
Fluorescently labeled cells were placed in a microporous
insert and allowed to migrate towards wells containing
TP508 at various concentrations. As shown in Fig. 4 (topFig. 2. Response of CAM vasculature to TP508. An agar disk impregnated
with either PBS (top panel) or 1 mg of TP508 in PBS (bottom panel) was
placed on the CAM of 9-day-old chick embryos for 96 h. The CAM area
around the disk was photographed under a dissecting microscope at a
magnification of 2�.
Fig. 3. TP508 stimulates chemokinesis of cultured endothelial cells. HMVE
and HAE cells (Clonetics, San Diego, CA) were plated on Thermonex
coverslips in silicone-restraining chamber with a diameter of 6 mm. After
48 h, the chamber was removed and fresh medium with or without TP508
was added to the cells. After an additional 72 h, the cells were fixed on the
coverslips, then stained with hematoxylin and eosin. From photomicro-
graphs, the distance the cells had migrated from the original 6-mm circle
was measured. Each bar represents the mean ± S.E. of three samples.
A.M. Norfleet et al. / General Pharmacology 35 (2002) 249–254252
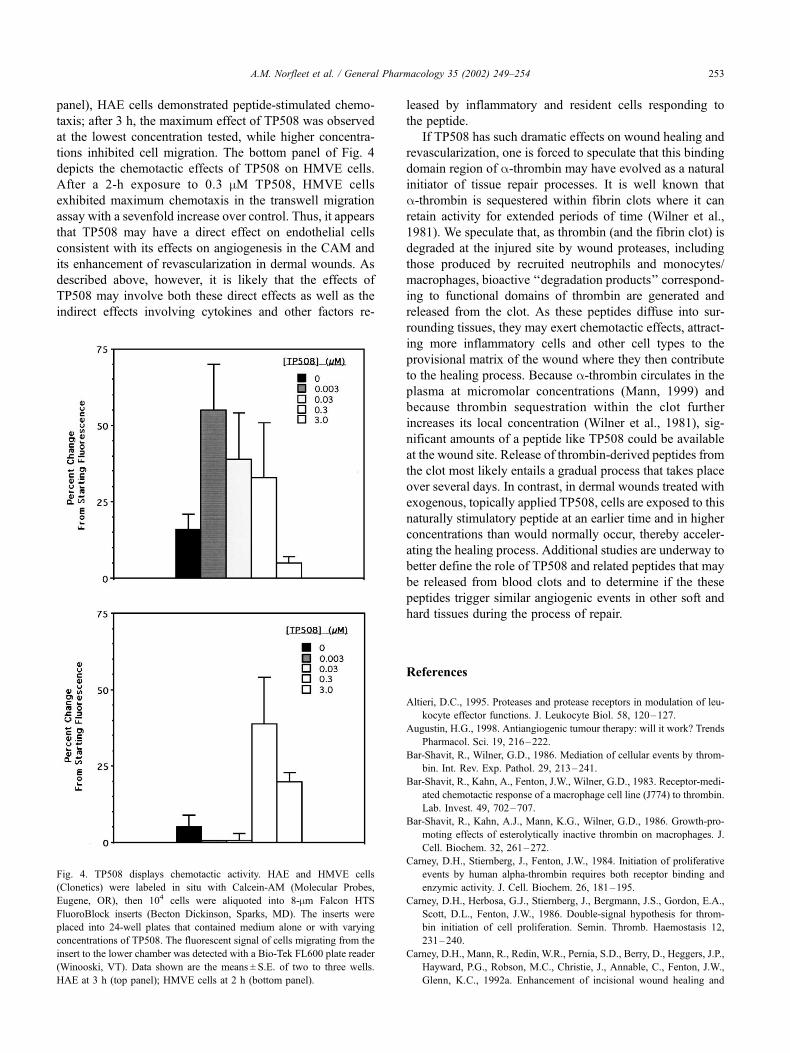
panel), HAE cells demonstrated peptide-stimulated chemo-
taxis; after 3 h, the maximum effect of TP508 was observed
at the lowest concentration tested, while higher concentra-
tions inhibited cell migration. The bottom panel of Fig. 4
depicts the chemotactic effects of TP508 on HMVE cells.
After a 2-h exposure to 0.3 mM TP508, HMVE cells
exhibited maximum chemotaxis in the transwell migration
assay with a sevenfold increase over control. Thus, it appears
that TP508 may have a direct effect on endothelial cells
consistent with its effects on angiogenesis in the CAM and
its enhancement of revascularization in dermal wounds. As
described above, however, it is likely that the effects of
TP508 may involve both these direct effects as well as the
indirect effects involving cytokines and other factors re-
leased by inflammatory and resident cells responding to
the peptide.
If TP508 has such dramatic effects on wound healing and
revascularization, one is forced to speculate that this binding
domain region of a-thrombin may have evolved as a natural
initiator of tissue repair processes. It is well known that
a-thrombin is sequestered within fibrin clots where it can
retain activity for extended periods of time (Wilner et al.,
1981). We speculate that, as thrombin (and the fibrin clot) is
degraded at the injured site by wound proteases, including
those produced by recruited neutrophils and monocytes/
macrophages, bioactive ‘‘degradation products’’ correspond-
ing to functional domains of thrombin are generated and
released from the clot. As these peptides diffuse into sur-
rounding tissues, they may exert chemotactic effects, attract-
ing more inflammatory cells and other cell types to the
provisional matrix of the wound where they then contribute
to the healing process. Because a-thrombin circulates in the
plasma at micromolar concentrations (Mann, 1999) and
because thrombin sequestration within the clot further
increases its local concentration (Wilner et al., 1981), sig-
nificant amounts of a peptide like TP508 could be available
at the wound site. Release of thrombin-derived peptides from
the clot most likely entails a gradual process that takes place
over several days. In contrast, in dermal wounds treated with
exogenous, topically applied TP508, cells are exposed to this
naturally stimulatory peptide at an earlier time and in higher
concentrations than would normally occur, thereby acceler-
ating the healing process. Additional studies are underway to
better define the role of TP508 and related peptides that may
be released from blood clots and to determine if the these
peptides trigger similar angiogenic events in other soft and
hard tissues during the process of repair.
References
Altieri, D.C., 1995. Proteases and protease receptors in modulation of leu-
kocyte effector functions. J. Leukocyte Biol. 58, 120–127.
Augustin, H.G., 1998. Antiangiogenic tumour therapy: will it work? Trends
Pharmacol. Sci. 19, 216–222.
Bar-Shavit, R., Wilner, G.D., 1986. Mediation of cellular events by throm-
bin. Int. Rev. Exp. Pathol. 29, 213–241.
Bar-Shavit, R., Kahn, A., Fenton, J.W., Wilner, G.D., 1983. Receptor-medi-
ated chemotactic response of a macrophage cell line (J774) to thrombin.
Lab. Invest. 49, 702–707.
Bar-Shavit, R., Kahn, A.J., Mann, K.G., Wilner, G.D., 1986. Growth-pro-
moting effects of esterolytically inactive thrombin on macrophages. J.
Cell. Biochem. 32, 261–272.
Carney, D.H., Stiernberg, J., Fenton, J.W., 1984. Initiation of proliferative
events by human alpha-thrombin requires both receptor binding and
enzymic activity. J. Cell. Biochem. 26, 181–195.
Carney, D.H., Herbosa, G.J., Stiernberg, J., Bergmann, J.S., Gordon, E.A.,
Scott, D.L., Fenton, J.W., 1986. Double-signal hypothesis for throm-
bin initiation of cell proliferation. Semin. Thromb. Haemostasis 12,
231–240.
Carney, D.H., Mann, R., Redin, W.R., Pernia, S.D., Berry, D., Heggers, J.P.,
Hayward, P.G., Robson, M.C., Christie, J., Annable, C., Fenton, J.W.,
Glenn, K.C., 1992a. Enhancement of incisional wound healing and
Fig. 4. TP508 displays chemotactic activity. HAE and HMVE cells
(Clonetics) were labeled in situ with Calcein-AM (Molecular Probes,
Eugene, OR), then 104 cells were aliquoted into 8-mm Falcon HTS
FluoroBlock inserts (Becton Dickinson, Sparks, MD). The inserts were
placed into 24-well plates that contained medium alone or with varying
concentrations of TP508. The fluorescent signal of cells migrating from the
insert to the lower chamber was detected with a Bio-Tek FL600 plate reader
(Winooski, VT). Data shown are the means ± S.E. of two to three wells.
HAE at 3 h (top panel); HMVE cells at 2 h (bottom panel).
A.M. Norfleet et al. / General Pharmacology 35 (2002) 249–254 253

neovascularization in normal rats by thrombin and synthetic thrombin
receptor-activating peptides. J. Clin. Invest. 89, 1469–1477.
Carney, D.H., Redin, W., McCroskey, L., 1992b. Role of high-affinity
thrombin receptors in postclotting cellular effects of thrombin. Semin.
Thromb. Haemostasis 18, 91–103.
Cirino, G., Napoli, C., Bucci, M., Cicala, C., 2000. Inflammation-coagu-
lation network: are serine protease receptors the knot? Trends Pharma-
col. Sci. 21, 170–172.
Fett, J.W., Bethune, J.L., Vallee, B.L., 1987. Induction of angiogenesis by
mixtures of two angiogenic proteins, angiogenin and acidic fibroblast
growth factor, in the chick chorioallantoic membrane. Biochem. Bio-
phys. Res. Commun. 146, 1122–1131.
Glenn, K.C., Frost, G.H., Bergmann, J.S., Carney, D.H., 1988. Synthetic
peptides bind to high-affinity thrombin receptors and modulate throm-
bin mitogenesis. Pept. Res. 1, 65–73.
Goldberger, A.S., Septak, M., 1998. A fluorescence blocking membrane
insert enhances analysis of cell motility assays. Becton Dickinson Tech.
Bull. 428, 1–5.
Grand, R.J., Turnell, A.S., Grabham, P.W., 1996. Cellular consequences of
thrombin-receptor activation. Biochem. J. 313, 353–368.
Hoying, J.B., Williams, S.K., 1996. Measurement of endothelial cell migra-
tion using an improved linear migration assay. Microcirculation 3,
167–174.
Jenkins, A.L., Howells, G.L., Scott, E., LeBonniec, B.F., Curtis, M.A.,
Stone, S.R., 1995. The response to thrombin of human neutrophils:
evidence for two novel receptors. J. Cell. Sci. 108, 3059–3066.
Mann, K.G., 1999. Biochemistry and physiology of blood coagulation.
Thromb. Haemostasis 82, 165–174.
Moller, M.L., Keherly, M.J., Carney, D.H., 2001. The thrombin peptide
TP508 is a potent chemotactic agent for human neutrophils (PMNs).
Mol. Biol. Cell 12, 257a.
Norfleet, A.M., Huang, Y., Sower, L.E., Redin, W.R., Fritz, R.R., Carney,
D.H., 2000. Thrombin peptide TP508 accelerates closure of dermal
excisions in animal tissue with surgically induced ischemia. Wound
Repair Regener. 8, 517–529.
Schwartz, S.M., Liaw, L., 1993. Growth control and morphogenesis in the
development and pathology of arteries. J. Cardiovasc. Pharmacol. 21,
S31–S49.
Sower, L.E., Huang, Y., Norfleet, A.M., Carney, D.H., 1999. Thrombin
peptide, TP508, induces proliferation and migration of keratinocytes.
Mol. Biol. Cell 10, 186a.
Stiernberg, J., Norfleet, A.M., Redin, W.R., Warner, W.S., Fritz, R.R.,
Carney, D.H., 2000. Acceleration of full-thickness wound healing in
normal rats by the synthetic thrombin peptide, TP508. Wound Repair
Regener. 8, 204–215.
Strukova, S.M., 2001. Thrombin as a regulator of inflammation and repar-
ative processes in tissues. Biochemistry (Moscow) 66, 8–18.
Vergnolle, N., Wallace, J.L., Bunnett, N.W., Hollenberg, M.D., 2001. Pro-
tease-activated receptors in inflammation, neuronal signaling and pain.
Trends Pharmacol. Sci. 22, 146–152.
Wilner, G.D., Danitz, M.P., Mudd, M.S., Hsieh, K.H., Fenton, J.W., 1981.
Selective immobilization of alpha-thrombin by surface-bound fibrin. J.
Lab. Clin. Med. 97, 403–411.
Yang, E.Y., Moses, H.L., 1990. Transforming growth factor beta 1-induced
changes in cell migration, proliferation, and angiogenesis in the chicken
chorioallantoic membrane. J. Cell. Biol. 111, 731–741.
A.M. Norfleet et al. / General Pharmacology 35 (2002) 249–254254





























