Embed Size (px)
Citation preview

24
Leong et al.: Harmful algal blooms in Singapore waters
Three new records of dinoflagellates in Singapore’s coastal waters, with observations on environmental conditions associated with microalgal growth in the Johor Straits
Sandric Leong* Chee Yew, Lim Lay Peng, Chew Siew Moon, Jerome Kok Wai Kit & Serena Teo Lay Ming
Abstract. The coastal environment in Singapore is increasingly affected by algal blooms. In addition to some 20 species previously reported in the literature, we have recently detected three dinoflagellate species, namely Karenia mikimotoi and two species of Karlodinium, i.e., K. cf. australe and K. cf. veneficum, for the first time in Singapore waters. Our studies indicate that the recent dinoflagellate blooms in the Johor Strait were formed during neap tide under physical settings that encouraged eutrophic conditions. In such conditions, several nitrogen (N) sources are available at high concentrations in contrast to low silicate (Si) concentration. A high N:Si ratio appears to allow a dinoflagellate bloom to be maintained for a prolonged period. The dinoflagellates Takayama xiamenensis, Karlodinium cf. veneficum and K. australe identified during the recent bloom appear to be associated with the fish kills along Johor. Detailed studies are needed to understand the dynamics of these blooms and causal factors initiating such blooms. Implementation of a long term science-based harmful algal blooms (HAB) management programme will be useful to safeguard our health of our coastal waters, mariculture activities and human health.
Key words. Fish kills, algal blooms, Karenia sp., Karlodinium spp., Singapore, toxins
RAFFLES BULLETIN OF ZOOLOGY Supplement No. 31: 24–36Date of publication: 10 July 2015http://zoobank.org/urn:lsid:zoobank.org:pub:8039D2A1-A032-4E5F-882D-D69BA18711E6
© National University of SingaporeISSN 2345-7600 (electronic) | ISSN 0217-2445 (print)
Tropical Marine Science Institute, National University of Singapore, 18 Kent Ridge Road, Singapore 119227; Email: [email protected] (*corresponding author)
INTRODUCTION
To date, some 20 species of dinoflagellates have been reported from Singapore waters (Chua, 1973; Khoo & Wee, 1996; Holmes & Teo, 2002; Tang et al., 2007, 2010, 2012; also undetermined species of Karenia and Scrippsiella in the Provasoli- Guillard National Centre for Marine Algae and Microbiota type collection, Bigelow Laboratory for Ocean Sciences, USA). Some bloom- forming phytoplankton species such as Alexandrium spp. are capable of producing biotoxins, which can be very potent when ingested, resulting in mortalities of marine mammals and humans (Leong et al., 2004). Other compounds with ichthyotoxic properties are often associated with fish kills resulting in heavy losses of mariculture (Hoagland et al., 2002; Kim, 2010). Some toxic cells even cause death of other organisms by direct contact (Uchida et al., 1995; Kamiyama et al., 2001; Leong et al., 2006).
Algal toxins may bio-accumulate in shellfish resulting in severe seafood poisoning, a major concern for seafood industries. Accumulated algal toxins in shellfish and other filter feeders could be passed on to higher trophic levels resulting in human intoxication. Consumption of these
organisms can also lead to severe shellfish poisoning that is associated with high mortality rates (Sobel & Painter, 2005). There will be global threats to human health if affected organisms are exported worldwide. Therefore, it is important to examine the extent of HAB vulnerability of major seafood export regions.
In Southeast Asia (SEA), many coastal regions are affected by regular HABs. For instance, in Malaysia, six algal blooms over a ten-year period between 2000 and 2010 were reported including shellfish poisoning cases (Lim et al., 2012). In the Philippines, there were more than 40 cases of HAB events during the period of 1983 to 2002 (Aquino et al., 2010), and in Vietnam, nine cases of HABs were reported during the period of 1993–2002 (Tang et al., 2004). These HAB events demonstrate the vulnerability of SEA coastal environment to HAB occurrences. The coastal zone of many SEA countries is characterised by a high level of anthropogenic activity that can have an effect on the dynamics of HABs (e.g., Adam et al., 2011; Leong et al., 2012; Mohammad-Noor et al., 2012). Changes in nutrient dynamics can result in the eutrophication of coastal waters, which is one of the stimuli for algal blooms that encourage HAB development (e.g., Leong & Taguchi, 2004).
In December 2009, a toxic algal bloom in the East Johor Strait caused massive fish kills that resulted in substantial economic losses (e.g., Quek & Lim, 2010; Leong et al., 2012). Prior to this event, there were few public reports of fish kill events associated with algal blooms. Subsequently, during late January to early February 2014, another HAB

25
RAFFLES BULLETIN OF ZOOLOGY 2015
event occurred in both.the East and West Johor Straits. This particular bloom induced massive fish-kills, amounting to 160 tons of fish lost valued in millions of dollars (Lee, 2014). These incidents suggest an increasing vulnerability of the Johor Straits of Singapore to HAB occurrences. However, the species responsible for these blooms have not been positively identified. The environmental conditions associated with such blooms are also poorly understood. In this study, the phytoplankton community in the Johor Straits water column was profiled on two occasions: (1), a pre-monsoon study from 16–30 October 2012 which examined environment parameters in relation to phytoplankton community structure; and (2), during a fish kill event from 10–12 February 2014 which examined the biological diversity in a HAB.
MATERIAL AND METHODS
Environmental studies. Field observations for bloom formation were carried out daily at the Outward Bound School jetty (1°25.166’N, 103°55.674’E) on Ubin Island located in the East Johor Strait, Singapore from 16–30 October, 2012 (Fig. 1) during the CMBS workshop.
Environmental and biological parameters were measured using a YSI (Yellow Springs Instrument Co.) multi-parameter instrument (Model SONDE 6600 V2-4), equipped with conductivity, temperature, depth (CTD), optical chlorophyll, dissolved oxygen, and colored dissolved organic matter (CDOM) sensors. Quinine sulfate (QS), a strong fluorescent alkaloid suitable as an indicative index of CDOM, was used as a standard for calibrating the CDOM sensor.
For the analysis of nutrients, cells abundance and species of phytoplankton, water samples were collected from the sea surface using a clean bucket. Water samples were then transferred into 4 L dark bottles and stored in a cooler box until further analysis. Subsamples for nutrient (ammonium, nitrate, nitrite, urea, phosphate and silicate) analyses were filtered through 0.45 μm pore size filter unit (Millipore), stored in dark bottles and frozen immediately until further analysis. Ammonium was determined using a modified Berthelot reaction using salicylate similar to the method as described by Patton & Crouch (1977). Nitrate and nitrite concentrations were measured using a multi-channel nutrient analyser (EcoLAB 2; EnviroTech Instruments). Urea was assayed using the diacetylmonoxime reagent method (Mulvenna & Savidge, 1992). Phosphate and silicate concentrations were determined following the protocol of JGOFS Report 19 (Knap et al., 1996). Phytoplankton species in the natural samples were identified microscopically at 400× magnification (Olympus IX-51), and their relative composition estimated by direct microscopic counts using a Sedgwick-Rafter cell.
DNA sampling and analysis. Sampling during the February 2014 algal bloom was carried out in Johor Straits, Singapore from 10–12 February 2014. Surface water was collected along East Johor Straits on 10 February from Punggol Jetty (Fig. 2). Samples were collected along the West Johor Strait off Tuas on 11 February and from Tuas and Lim Chu Kang
Fig. 1. Sampling stations (grey inverted triangle) in and around Ubin Island, Singapore. Inset shows locations of sampling stations during the April 2014 bloom in the East Johor Strait, Singapore. Maps were extracted using Geophysical Survey Data from NOAA, National Geophysical Data Center website. Ubin Island is about 7.5 km in length from east to west.
Fig. 2. Sampling stations during the February 2014 bloom along Johor Straits, Singapore. Square symbol: Tuas area; circle: Lim Chu Kang Jetty; triangle: Punggol Jetty. Inset shows locations of sampling stations at Tuas area. Maps were extracted using Geophysical Survey Data from NOAA, National Geophysical Data Center website.
jetties on 12 February (Fig. 2). Samples were stored in 4L dark bottles and kept in a cooler box until further analysis of cell abundance, species identification and DNA in the laboratory. DNA extraction and PCR amplification were carried out using ethanol-preserved phytoplankton samples which were centrifuged at 4°C and 15,000 rpm for 3 min. The cell pellet was re-suspended in TE buffer (10 mM Tris HCl and 1 mM EDTA, pH 8.0) and Triton-X 10, and boiled at 70°C for about 5 min. Further extraction steps were conducted according to instructions provided by the manufacturer (DNeasy Plant Mini Kit).
26
Leong et al.: Harmful algal blooms in Singapore waters
PCR amplification was done following the methods outlined by Kok et al. (2012a). The first PCR amplification of the 18s rDNA was carried out using PP18S-408F [5´- TAC CAC ATC YAA GGA AGG CAG - 3´] and PP18S-1332R [5´- CTC GTT CGT TAA CGG AAT TAA C - 3´] primers. A 25 µl reaction mixture containing Premix Ex Taq (Takara), 0.5 µM each of the forward primer PP18S-408F and reverse primer PP18S-1332R, and 1 µl of the genomic DNA template was prepared. PCR amplification comprised of initial denaturation at 94°C for 3 min; followed by 35 cycles each consisting of a denaturation step at 94°C for 30 s, an annealing step at 62°C for 30 s, and an extension step at 72°C for 1 min 30 s; and one final extension step at 72°C for 5 min.
Nested PCR was then performed after the first PCR amplification by using 1 μ1 of first PCR reactant with PP18S-431F [5´- GGC GCG YAA ATT ACC CAA TM - 3´] and PP18S-1133R [5´- TCA GCC TTG CGA CCA TAC TC - 3´]. The ingredients used in the PCR master mix for this nested PCR step included Premix Ex Taq (Takara), 0.5 µM of each forward primer PP18S-431F and reverse primer PP18S-1133R, as well as 1 µl of the first PCR reactant. PCR amplification conditions comprised an initial denaturation at 94°C for 3 min; followed by 30 cycles each consisting of denaturation at 94°C for 30 s, annealing at 62°C for 30 s and extension at 72°C for 1 min; and finally 72°C for 5 min. The amplified DNA was then purified with GFX kit (GE Healthcare Life Science).
Fragments of 18S rDNA that were obtained by nested PCR were cloned into pT7Blue T-vector (Novagen), ligated with Ligation High Ver. 2 (Toyobo), and left overnight. Recombinant plasmids were used for transformation into DH5-alpha competent Escherichia coli cells. The transformants were spread on an L-amp plate, which was made using 40 µg mL-1 X-gal, 0.5 mM IPTG, 100 µg mL-1 ampicillin and agar. White colonies (cells with plasmid insert) were selected and picked at random and subcultured into 96-well plates (each well contained 100 µL LB medium with 100 µg mL-1 ampicillin). Colonies were then incubated overnight at 37°C. PCR amplification was then performed following procedures described earlier for the nested PCR step. The resultant DNA amplicons were purified with Exosap IT and put through a two-step amplification at 37°C for 15 min followed by 80°C for 15 min. Purified amplicons were then sent to AIT Biotech (Singapore) for sequencing. Sequencing was carried out using the ABI 3730XL DNA Analyzer (PE Biosystems, Vernon Hills, IL, USA).
Species identification and phylogenetic analysis. 18S rDNA sequences obtained in this study were compared and analysed with sequences retrieved from the National Center for Biotechnology Information (NCBI) nucleotide database (http://www.ncbi.nlm.nih.gov/). Rhodella violacea (Kornmann) Wehrmeyer (AF168624) was selected as an out-group in the neighbor-joining (NJ) tree construction. Sequences were aligned using ClustalX (Thompson et al., 1997) with further analysis carried out using GENETYX (version 12.1.0). Bootstrap values were estimated and set from 1,000 replicates. The nucleotide sequences obtained in
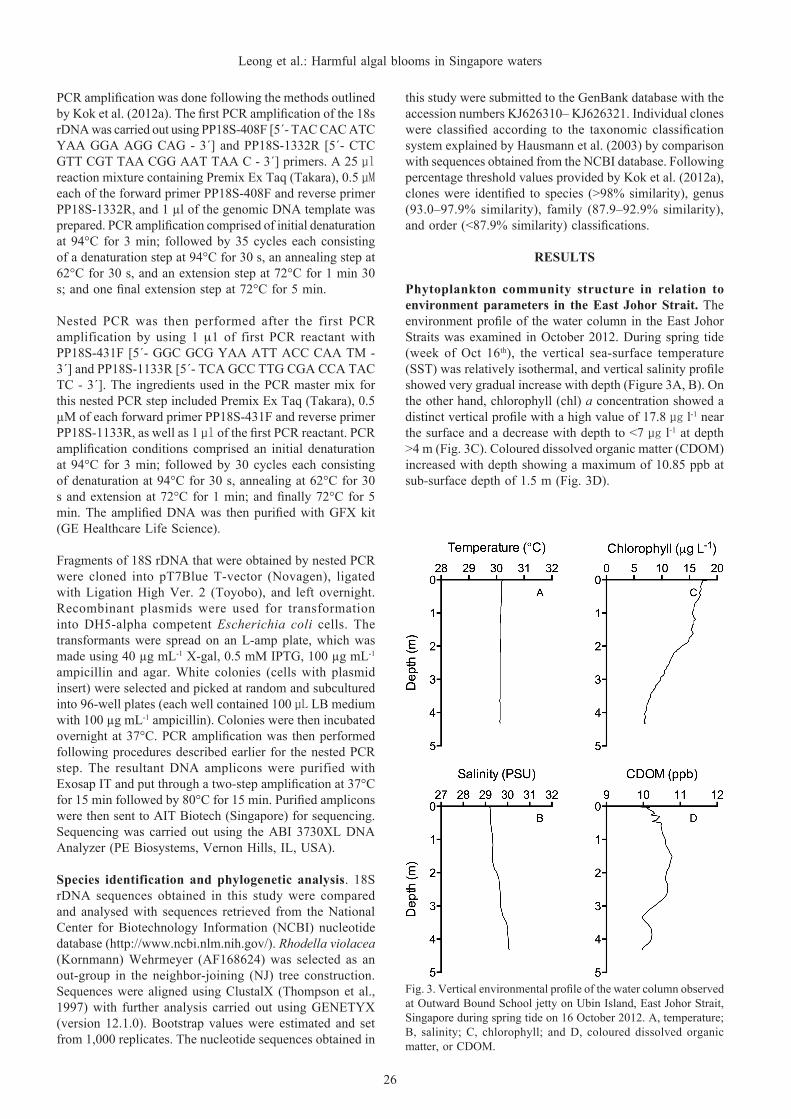
Fig. 3. Vertical environmental profile of the water column observed at Outward Bound School jetty on Ubin Island, East Johor Strait, Singapore during spring tide on 16 October 2012. A, temperature; B, salinity; C, chlorophyll; and D, coloured dissolved organic matter, or CDOM.
this study were submitted to the GenBank database with the accession numbers KJ626310– KJ626321. Individual clones were classified according to the taxonomic classification system explained by Hausmann et al. (2003) by comparison with sequences obtained from the NCBI database. Following percentage threshold values provided by Kok et al. (2012a), clones were identified to species (>98% similarity), genus (93.0–97.9% similarity), family (87.9–92.9% similarity), and order (<87.9% similarity) classifications.
RESULTS
Phytoplankton community structure in relation to environment parameters in the East Johor Strait. The environment profile of the water column in the East Johor Straits was examined in October 2012. During spring tide (week of Oct 16th), the vertical sea-surface temperature (SST) was relatively isothermal, and vertical salinity profile showed very gradual increase with depth (Figure 3A, B). On the other hand, chlorophyll (chl) a concentration showed a distinct vertical profile with a high value of 17.8 µg l-1 near the surface and a decrease with depth to <7 µg l-1 at depth >4 m (Fig. 3C). Coloured dissolved organic matter (CDOM) increased with depth showing a maximum of 10.85 ppb at sub-surface depth of 1.5 m (Fig. 3D).
27
RAFFLES BULLETIN OF ZOOLOGY 2015
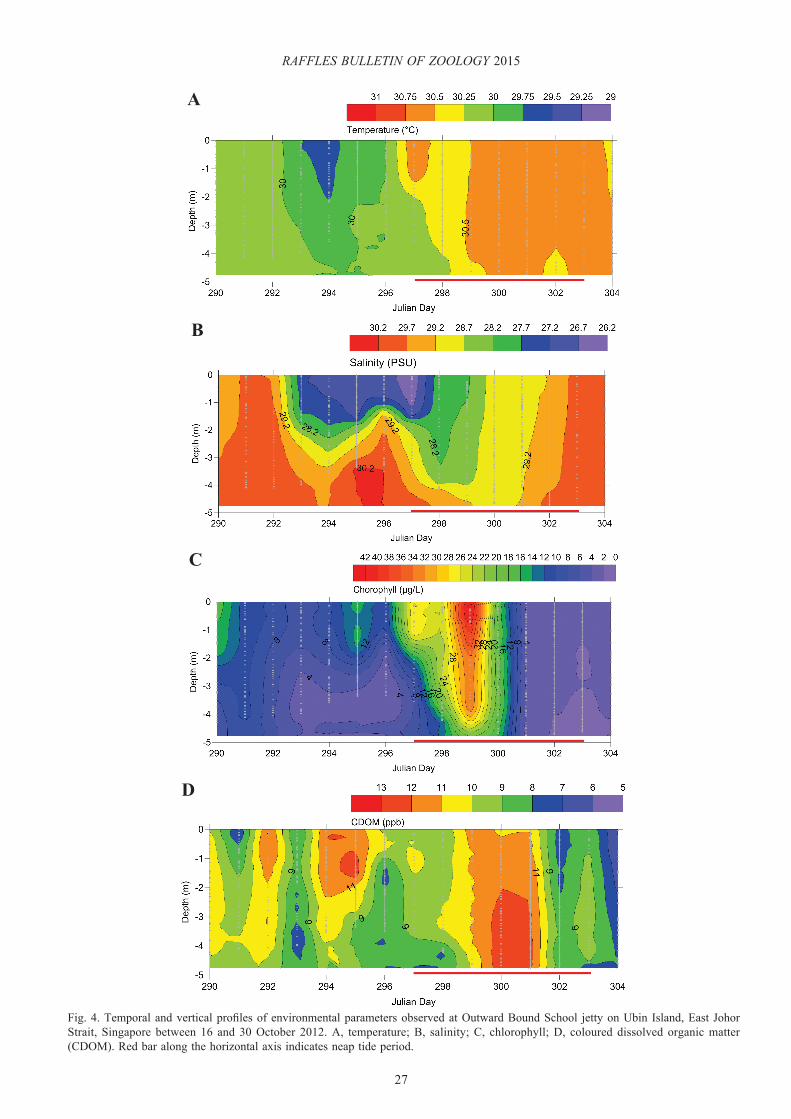
Fig. 4. Temporal and vertical profiles of environmental parameters observed at Outward Bound School jetty on Ubin Island, East Johor Strait, Singapore between 16 and 30 October 2012. A, temperature; B, salinity; C, chlorophyll; D, coloured dissolved organic matter (CDOM). Red bar along the horizontal axis indicates neap tide period.
A
C
D
B
28
Leong et al.: Harmful algal blooms in Singapore waters
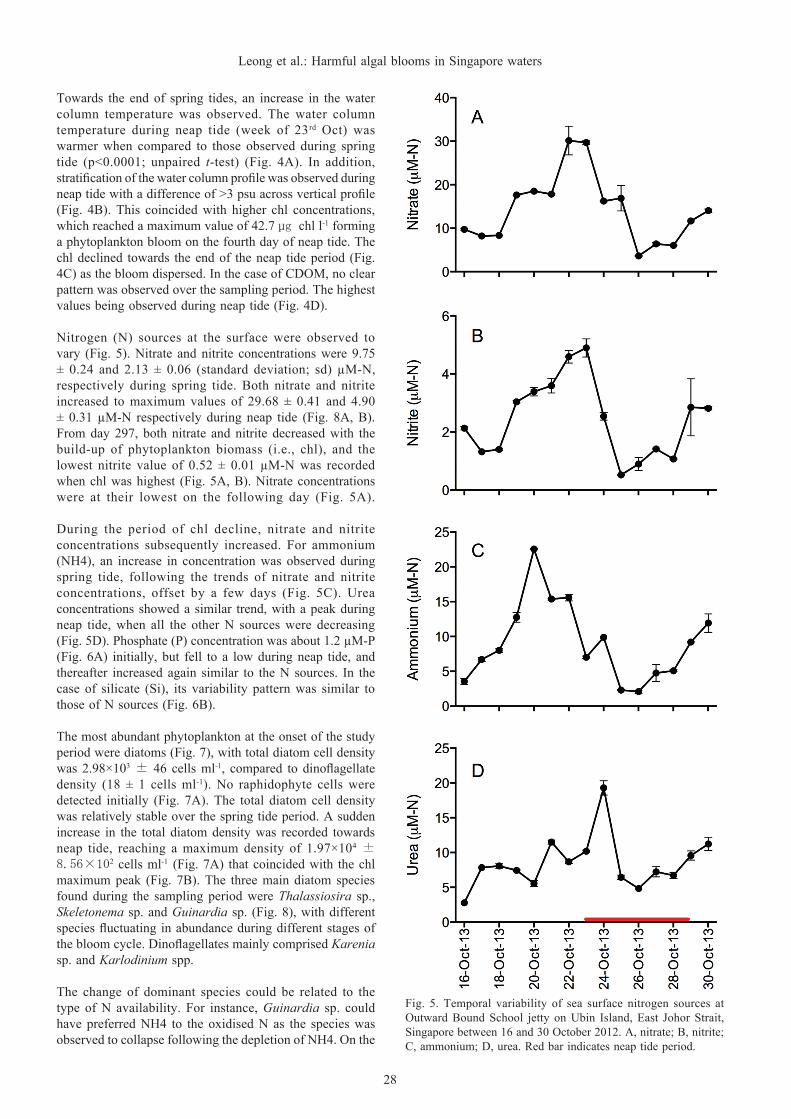
Fig. 5. Temporal variability of sea surface nitrogen sources at Outward Bound School jetty on Ubin Island, East Johor Strait, Singapore between 16 and 30 October 2012. A, nitrate; B, nitrite; C, ammonium; D, urea. Red bar indicates neap tide period.
Towards the end of spring tides, an increase in the water column temperature was observed. The water column temperature during neap tide (week of 23rd Oct) was warmer when compared to those observed during spring tide (p<0.0001; unpaired t-test) (Fig. 4A). In addition, stratification of the water column profile was observed during neap tide with a difference of >3 psu across vertical profile (Fig. 4B). This coincided with higher chl concentrations, which reached a maximum value of 42.7 µg chl l-1 forming a phytoplankton bloom on the fourth day of neap tide. The chl declined towards the end of the neap tide period (Fig. 4C) as the bloom dispersed. In the case of CDOM, no clear pattern was observed over the sampling period. The highest values being observed during neap tide (Fig. 4D).
Nitrogen (N) sources at the surface were observed to vary (Fig. 5). Nitrate and nitrite concentrations were 9.75 ± 0.24 and 2.13 ± 0.06 (standard deviation; sd) µM-N, respectively during spring tide. Both nitrate and nitrite increased to maximum values of 29.68 ± 0.41 and 4.90 ± 0.31 µM-N respectively during neap tide (Fig. 8A, B). From day 297, both nitrate and nitrite decreased with the build-up of phytoplankton biomass (i.e., chl), and the lowest nitrite value of 0.52 ± 0.01 µM-N was recorded when chl was highest (Fig. 5A, B). Nitrate concentrations were at their lowest on the following day (Fig. 5A).
During the period of chl decline, nitrate and nitrite concentrations subsequently increased. For ammonium (NH4), an increase in concentration was observed during spring tide, following the trends of nitrate and nitrite concentrations, offset by a few days (Fig. 5C). Urea concentrations showed a similar trend, with a peak during neap tide, when all the other N sources were decreasing (Fig. 5D). Phosphate (P) concentration was about 1.2 µM-P (Fig. 6A) initially, but fell to a low during neap tide, and thereafter increased again similar to the N sources. In the case of silicate (Si), its variability pattern was similar to those of N sources (Fig. 6B).
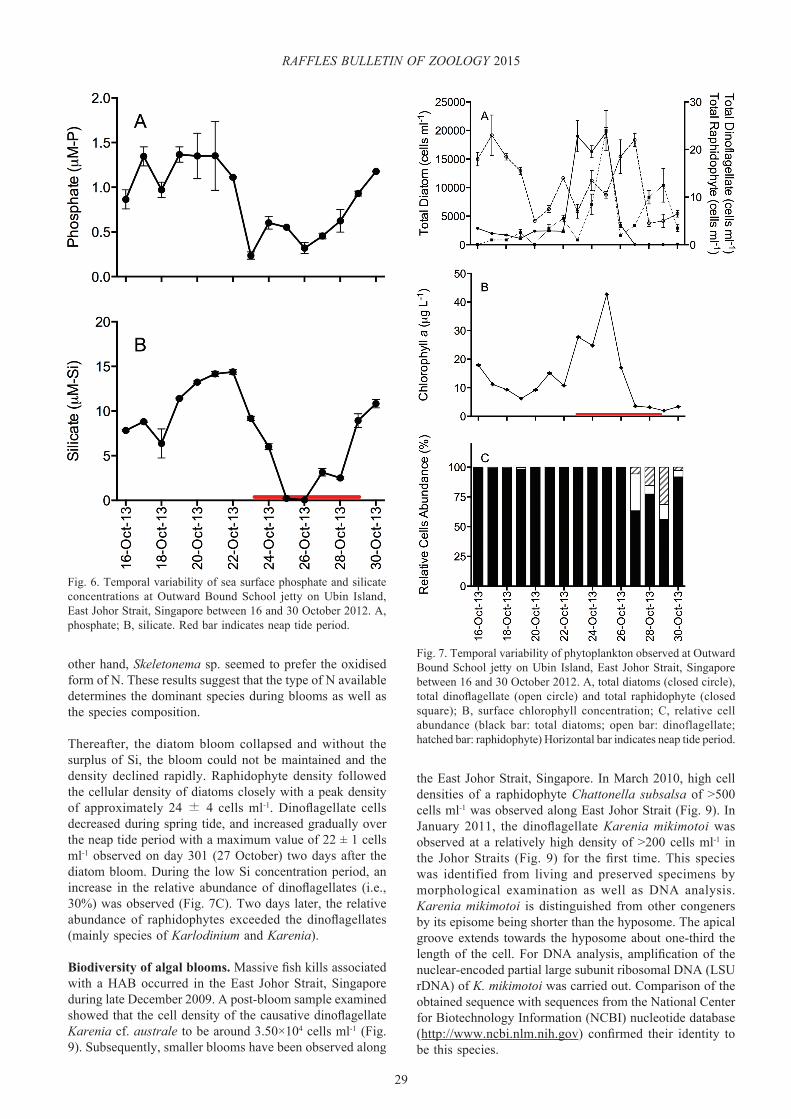
The most abundant phytoplankton at the onset of the study period were diatoms (Fig. 7), with total diatom cell density was 2.98×103 ± 46 cells ml-1, compared to dinoflagellate density (18 ± 1 cells ml-1). No raphidophyte cells were detected initially (Fig. 7A). The total diatom cell density was relatively stable over the spring tide period. A sudden increase in the total diatom density was recorded towards neap tide, reaching a maximum density of 1.97×104 ± 8.56×102 cells ml-1 (Fig. 7A) that coincided with the chl maximum peak (Fig. 7B). The three main diatom species found during the sampling period were Thalassiosira sp., Skeletonema sp. and Guinardia sp. (Fig. 8), with different species fluctuating in abundance during different stages of the bloom cycle. Dinoflagellates mainly comprised Karenia sp. and Karlodinium spp.
The change of dominant species could be related to the type of N availability. For instance, Guinardia sp. could have preferred NH4 to the oxidised N as the species was observed to collapse following the depletion of NH4. On the
29
RAFFLES BULLETIN OF ZOOLOGY 2015
Fig. 7. Temporal variability of phytoplankton observed at Outward Bound School jetty on Ubin Island, East Johor Strait, Singapore between 16 and 30 October 2012. A, total diatoms (closed circle), total dinoflagellate (open circle) and total raphidophyte (closed square); B, surface chlorophyll concentration; C, relative cell abundance (black bar: total diatoms; open bar: dinoflagellate; hatched bar: raphidophyte) Horizontal bar indicates neap tide period.
other hand, Skeletonema sp. seemed to prefer the oxidised form of N. These results suggest that the type of N available determines the dominant species during blooms as well as the species composition.
Thereafter, the diatom bloom collapsed and without the surplus of Si, the bloom could not be maintained and the density declined rapidly. Raphidophyte density followed the cellular density of diatoms closely with a peak density of approximately 24 ± 4 cells ml-1. Dinoflagellate cells decreased during spring tide, and increased gradually over the neap tide period with a maximum value of 22 ± 1 cells ml-1 observed on day 301 (27 October) two days after the diatom bloom. During the low Si concentration period, an increase in the relative abundance of dinoflagellates (i.e., 30%) was observed (Fig. 7C). Two days later, the relative abundance of raphidophytes exceeded the dinoflagellates (mainly species of Karlodinium and Karenia).
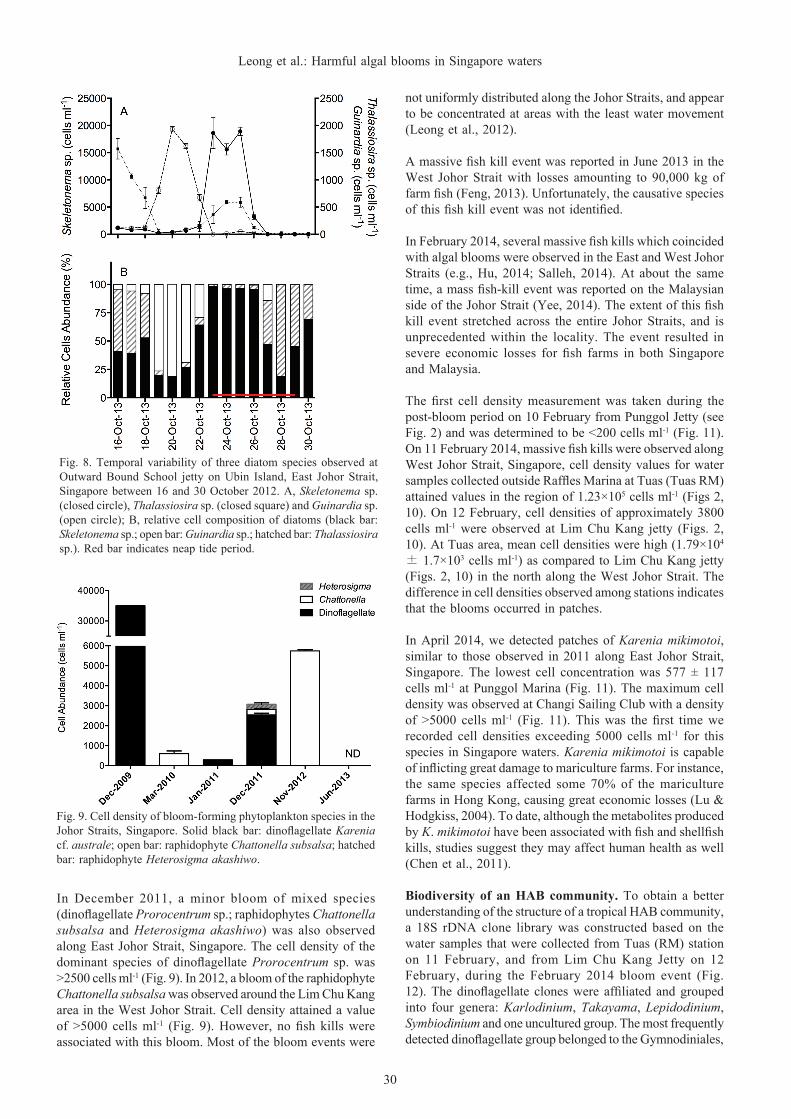
Biodiversity of algal blooms. Massive fish kills associated with a HAB occurred in the East Johor Strait, Singapore during late December 2009. A post-bloom sample examined showed that the cell density of the causative dinoflagellate Karenia cf. australe to be around 3.50×104 cells ml-1 (Fig. 9). Subsequently, smaller blooms have been observed along
the East Johor Strait, Singapore. In March 2010, high cell densities of a raphidophyte Chattonella subsalsa of >500 cells ml-1 was observed along East Johor Strait (Fig. 9). In January 2011, the dinoflagellate Karenia mikimotoi was observed at a relatively high density of >200 cells ml-1 in the Johor Straits (Fig. 9) for the first time. This species was identified from living and preserved specimens by morphological examination as well as DNA analysis. Karenia mikimotoi is distinguished from other congeners by its episome being shorter than the hyposome. The apical groove extends towards the hyposome about one-third the length of the cell. For DNA analysis, amplification of the nuclear-encoded partial large subunit ribosomal DNA (LSU rDNA) of K. mikimotoi was carried out. Comparison of the obtained sequence with sequences from the National Center for Biotechnology Information (NCBI) nucleotide database (http://www.ncbi.nlm.nih.gov) confirmed their identity to be this species.
Fig. 6. Temporal variability of sea surface phosphate and silicate concentrations at Outward Bound School jetty on Ubin Island, East Johor Strait, Singapore between 16 and 30 October 2012. A, phosphate; B, silicate. Red bar indicates neap tide period.
30
Leong et al.: Harmful algal blooms in Singapore waters
Fig. 9. Cell density of bloom-forming phytoplankton species in the Johor Straits, Singapore. Solid black bar: dinoflagellate Karenia cf. australe; open bar: raphidophyte Chattonella subsalsa; hatched bar: raphidophyte Heterosigma akashiwo.
Fig. 8. Temporal variability of three diatom species observed at Outward Bound School jetty on Ubin Island, East Johor Strait, Singapore between 16 and 30 October 2012. A, Skeletonema sp. (closed circle), Thalassiosira sp. (closed square) and Guinardia sp. (open circle); B, relative cell composition of diatoms (black bar: Skeletonema sp.; open bar: Guinardia sp.; hatched bar: Thalassiosira sp.). Red bar indicates neap tide period.
In December 2011, a minor bloom of mixed species (dinoflagellate Prorocentrum sp.; raphidophytes Chattonella subsalsa and Heterosigma akashiwo) was also observed along East Johor Strait, Singapore. The cell density of the dominant species of dinoflagellate Prorocentrum sp. was >2500 cells ml-1 (Fig. 9). In 2012, a bloom of the raphidophyte Chattonella subsalsa was observed around the Lim Chu Kang area in the West Johor Strait. Cell density attained a value of >5000 cells ml-1 (Fig. 9). However, no fish kills were associated with this bloom. Most of the bloom events were
not uniformly distributed along the Johor Straits, and appear to be concentrated at areas with the least water movement (Leong et al., 2012).
A massive fish kill event was reported in June 2013 in the West Johor Strait with losses amounting to 90,000 kg of farm fish (Feng, 2013). Unfortunately, the causative species of this fish kill event was not identified.
In February 2014, several massive fish kills which coincided with algal blooms were observed in the East and West Johor Straits (e.g., Hu, 2014; Salleh, 2014). At about the same time, a mass fish-kill event was reported on the Malaysian side of the Johor Strait (Yee, 2014). The extent of this fish kill event stretched across the entire Johor Straits, and is unprecedented within the locality. The event resulted in severe economic losses for fish farms in both Singapore and Malaysia.
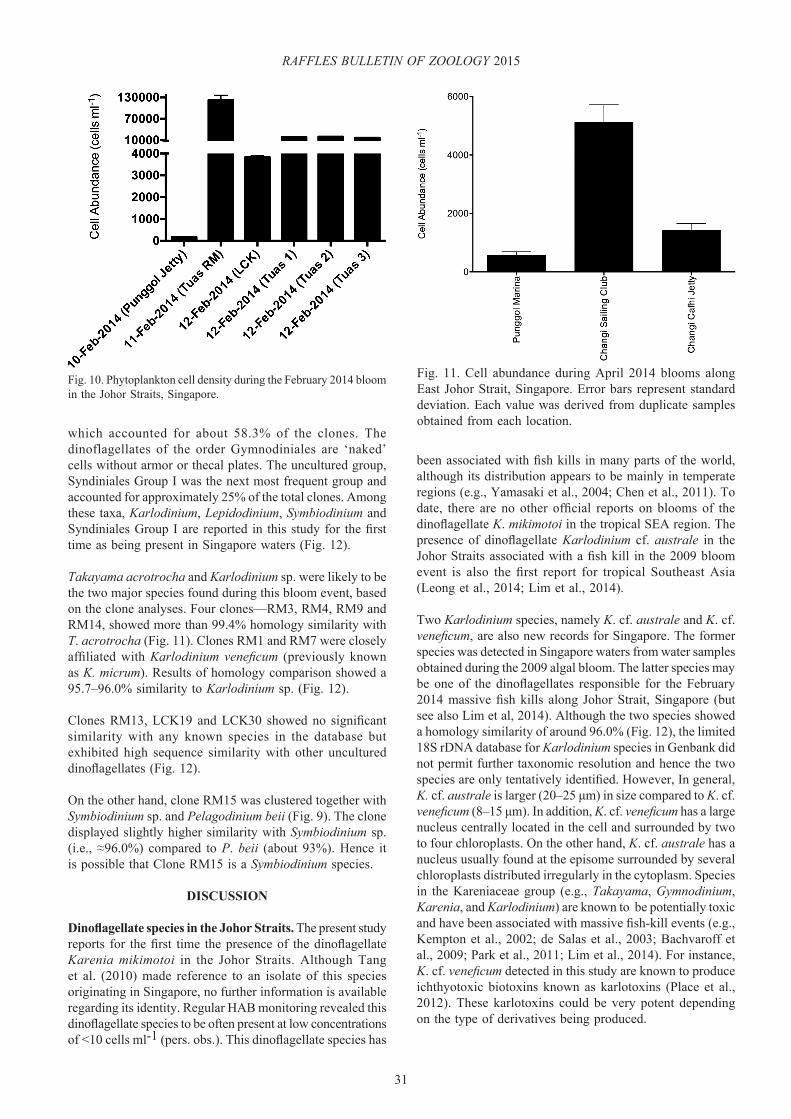
The first cell density measurement was taken during the post-bloom period on 10 February from Punggol Jetty (see Fig. 2) and was determined to be <200 cells ml-1 (Fig. 11). On 11 February 2014, massive fish kills were observed along West Johor Strait, Singapore, cell density values for water samples collected outside Raffles Marina at Tuas (Tuas RM) attained values in the region of 1.23×105 cells ml-1 (Figs 2, 10). On 12 February, cell densities of approximately 3800 cells ml-1 were observed at Lim Chu Kang jetty (Figs. 2, 10). At Tuas area, mean cell densities were high (1.79×104 ± 1.7×103 cells ml-1) as compared to Lim Chu Kang jetty (Figs. 2, 10) in the north along the West Johor Strait. The difference in cell densities observed among stations indicates that the blooms occurred in patches.
In April 2014, we detected patches of Karenia mikimotoi, similar to those observed in 2011 along East Johor Strait, Singapore. The lowest cell concentration was 577 ± 117 cells ml-1 at Punggol Marina (Fig. 11). The maximum cell density was observed at Changi Sailing Club with a density of >5000 cells ml-1 (Fig. 11). This was the first time we recorded cell densities exceeding 5000 cells ml-1 for this species in Singapore waters. Karenia mikimotoi is capable of inflicting great damage to mariculture farms. For instance, the same species affected some 70% of the mariculture farms in Hong Kong, causing great economic losses (Lu & Hodgkiss, 2004). To date, although the metabolites produced by K. mikimotoi have been associated with fish and shellfish kills, studies suggest they may affect human health as well (Chen et al., 2011).
Biodiversity of an HAB community. To obtain a better understanding of the structure of a tropical HAB community, a 18S rDNA clone library was constructed based on the water samples that were collected from Tuas (RM) station on 11 February, and from Lim Chu Kang Jetty on 12 February, during the February 2014 bloom event (Fig. 12). The dinoflagellate clones were affiliated and grouped into four genera: Karlodinium, Takayama, Lepidodinium, Symbiodinium and one uncultured group. The most frequently detected dinoflagellate group belonged to the Gymnodiniales,
31
RAFFLES BULLETIN OF ZOOLOGY 2015
Fig. 10. Phytoplankton cell density during the February 2014 bloom in the Johor Straits, Singapore.
Fig. 11. Cell abundance during April 2014 blooms along East Johor Strait, Singapore. Error bars represent standard deviation. Each value was derived from duplicate samples obtained from each location.
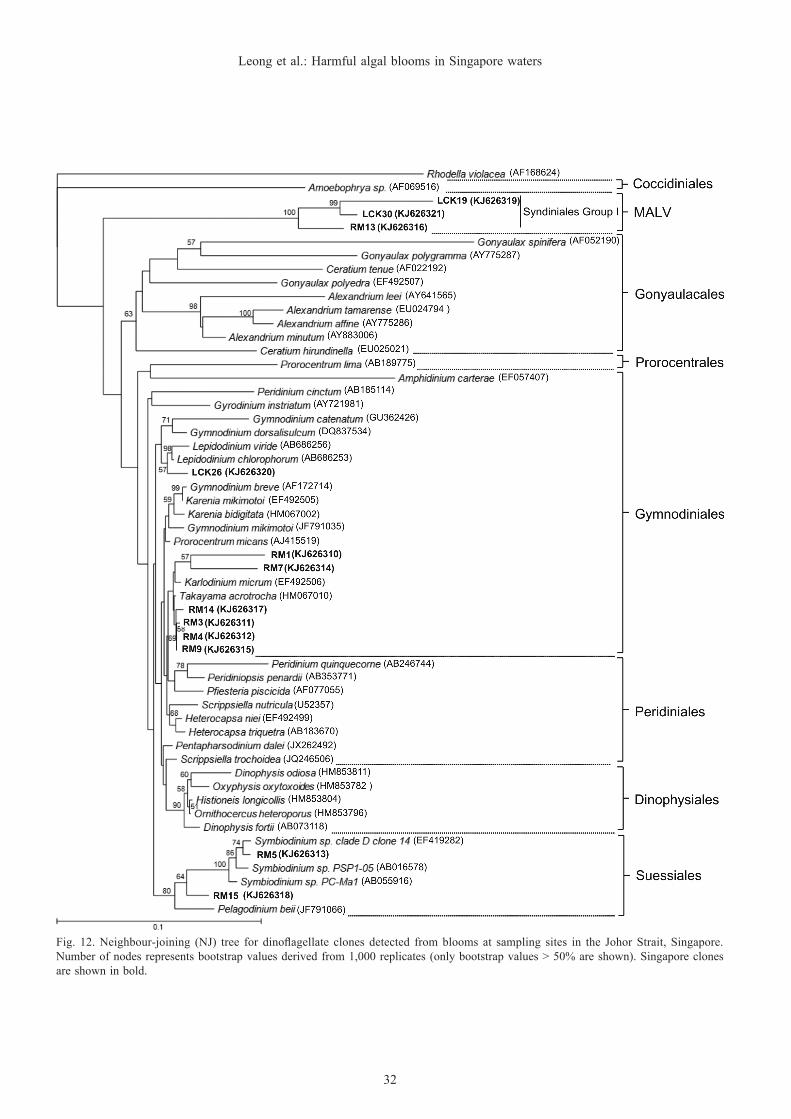
which accounted for about 58.3% of the clones. The dinoflagellates of the order Gymnodiniales are ‘naked’ cells without armor or thecal plates. The uncultured group, Syndiniales Group I was the next most frequent group and accounted for approximately 25% of the total clones. Among these taxa, Karlodinium, Lepidodinium, Symbiodinium and Syndiniales Group I are reported in this study for the first time as being present in Singapore waters (Fig. 12).
Takayama acrotrocha and Karlodinium sp. were likely to be the two major species found during this bloom event, based on the clone analyses. Four clones—RM3, RM4, RM9 and RM14, showed more than 99.4% homology similarity with T. acrotrocha (Fig. 11). Clones RM1 and RM7 were closely affiliated with Karlodinium veneficum (previously known as K. micrum). Results of homology comparison showed a 95.7–96.0% similarity to Karlodinium sp. (Fig. 12).
Clones RM13, LCK19 and LCK30 showed no significant similarity with any known species in the database but exhibited high sequence similarity with other uncultured dinoflagellates (Fig. 12).
On the other hand, clone RM15 was clustered together with Symbiodinium sp. and Pelagodinium beii (Fig. 9). The clone displayed slightly higher similarity with Symbiodinium sp. (i.e., ≈96.0%) compared to P. beii (about 93%). Hence it is possible that Clone RM15 is a Symbiodinium species.
DISCUSSION
Dinoflagellate species in the Johor Straits. The present study reports for the first time the presence of the dinoflagellate Karenia mikimotoi in the Johor Straits. Although Tang et al. (2010) made reference to an isolate of this species originating in Singapore, no further information is available regarding its identity. Regular HAB monitoring revealed this dinoflagellate species to be often present at low concentrations of <10 cells ml-1 (pers. obs.). This dinoflagellate species has
been associated with fish kills in many parts of the world, although its distribution appears to be mainly in temperate regions (e.g., Yamasaki et al., 2004; Chen et al., 2011). To date, there are no other official reports on blooms of the dinoflagellate K. mikimotoi in the tropical SEA region. The presence of dinoflagellate Karlodinium cf. australe in the Johor Straits associated with a fish kill in the 2009 bloom event is also the first report for tropical Southeast Asia (Leong et al., 2014; Lim et al., 2014).
Two Karlodinium species, namely K. cf. australe and K. cf. veneficum, are also new records for Singapore. The former species was detected in Singapore waters from water samples obtained during the 2009 algal bloom. The latter species may be one of the dinoflagellates responsible for the February 2014 massive fish kills along Johor Strait, Singapore (but see also Lim et al, 2014). Although the two species showed a homology similarity of around 96.0% (Fig. 12), the limited 18S rDNA database for Karlodinium species in Genbank did not permit further taxonomic resolution and hence the two species are only tentatively identified. However, In general, K. cf. australe is larger (20–25 μm) in size compared to K. cf. veneficum (8–15 μm). In addition, K. cf. veneficum has a large nucleus centrally located in the cell and surrounded by two to four chloroplasts. On the other hand, K. cf. australe has a nucleus usually found at the episome surrounded by several chloroplasts distributed irregularly in the cytoplasm. Species in the Kareniaceae group (e.g., Takayama, Gymnodinium, Karenia, and Karlodinium) are known to be potentially toxic and have been associated with massive fish-kill events (e.g., Kempton et al., 2002; de Salas et al., 2003; Bachvaroff et al., 2009; Park et al., 2011; Lim et al., 2014). For instance, K. cf. veneficum detected in this study are known to produce ichthyotoxic biotoxins known as karlotoxins (Place et al., 2012). These karlotoxins could be very potent depending on the type of derivatives being produced.
32
Leong et al.: Harmful algal blooms in Singapore waters
Fig. 12. Neighbour-joining (NJ) tree for dinoflagellate clones detected from blooms at sampling sites in the Johor Strait, Singapore. Number of nodes represents bootstrap values derived from 1,000 replicates (only bootstrap values > 50% are shown). Singapore clones are shown in bold.

33
RAFFLES BULLETIN OF ZOOLOGY 2015
Another unarmoured dinoflagellate species Takayama acrotrocha was previously recorded from Singapore waters in 2004 and 2007 (Tang et al., 2012). However, Gu et al. (2013) argued that T. acrotrocha identified by Tang et al. (2012) should be T. xiamenensis, based on the finger-like sulcal intrusion observed in ventral view of the latter species. The morphology of four Takayama clones from Tuas in this study concurs with the description of T. xiamenensis in Gu et al. (2013). Takayama acrotrocha do not cause fish kills (Tang et al., 2012). However, in the 2014 bloom event, Takayama species was associated with massive fish-kills, suggesting further that T. xiamensensis may be present in Singapore. More studies are necessary to determine if T. xiamenensis has bioactive metabolites.
Our observations showed that the bloom in February 2014 was a multi-species dinoflagellate bloom with different dominant dinoflagellates found at different locations. For instance T. xiamenensis was found to dominate at Tuas RM. As for the Lim Chu Kang samples, we did not find any of the dinoflagellates detected in Tuas area (i.e., Takayama sp. or Karlodinium spp.; K. veneficum and K. australe) from clone analysis.
Syndiniales Group I was the second main group identified in the present study. Group I belongs to Marine Alveolates Group (MALV) (Díez et al., 2001; López-García et al., 2001; Moon-van der Staay et al., 2001; Groisillier et al., 2006). According to Guillou et al. (2008), Syndiniales Group I are endosymbiont parasites that associate with the Alveolata host cells (Coats & Park, 2002; Guillou et al., 2008). Alveolata is a major eukaryotic group, which comprises the Ciliophora, Apicomplexa, Perkinsea, and dinoflagellates. In a separate study within Sagami Bay in Japan, Kok et al. (2012b) showed that the Syndiniales Group I accounted for about 53% of the total dinoflagellate clones, which was about twice the percentage obtained in this present study from the sample collected along the Johor Straits.
On the other hand, clones RM13, LCK19 and LCK30 showed no significant similarity with any known species in the database but exhibited high sequence similarity with other uncultured dinoflagellates (Fig. 14). Clone RM13 showed 100% similarity with “uncultured alveolate 18S rRNA gene, clone C5” that had been isolated from the Southern Baltic Sea (Piwosz & Pernthaler, 2010). Clones LCK19 and LCK30 exhibited 97% and 99% similarity with “uncultured dinoflagellate gene for 18S rRNA, partial sequence, clone: PJ11” and “uncultured dinoflagellate gene for 18S rRNA, partial sequence, clone: PJ22” respectively (Kok et al., 2012a). Both the PJ11 and PJ22 clones had been isolated from Sagami Bay, Japan.
Symbiodinium sp. was also identified in the present study apart from the uncultured dinoflagellates. This symbiotic dinoflagellate is usually associated with coral reefs, foraminifera and marine invertebrates (Carlos et al., 1999). This group of symbiotic dinoflagellates is not well understood in tropical waters. More studies are necessary to confirm their role and ecological function in tropical ecosystem.
There is a high diversity of bloom-forming species in the Singapore coastal environments, some of which are known to be toxic (Holmes & Teo 2002; Gin et al., 2006; Leong et al., 2014), and many remain to be properly documented and identified. Analyses of samples from routine sampling using normal molecular approaches or microscopic observations cannot possibly completely screen for every single species present. For instance, endosymbiotic parasites and symbiotic dinoflagellates cannot be observed through microscopic methods. However, by using the DNA cloning approach, we could detect such species, and could enhance our understanding about the community structure and biodiversity in Singapore waters and surrounding regions. Not all blooms were associated with fish kills, and most blooms rarely caused serious impacts to the coastal ecosystem. HAB events observed in Singapore waters have been sporadic and not associated with any one particular species. While most species have not previously caused local HAB events, they can potentially cause serious problems in the future.
Environment of the Johor Straits. The environmental conditions in the Johor Strait are affected by multiple sources of variation including the monsoons (i.e., Northeast and Southwest monsoons), terrestrial and atmospheric inputs, as well as tidal currents. In particular, the salinity along the length of the Johor Straits can differ significantly especially during the Northeast monsoon period and is an important factor affecting the growth of phytoplankton in this ecosystem. During times of regular heavy rainfall and terrestrial input, salinity values can fall below 20 psu. Under favorable environmental conditions, phytoplankton biomass could accumulate and form an algal bloom in the Johor Straits, such as the diatom bloom described in this study. These blooms may lead to eutrophication events.
The Johor Straits is a unique coastal ecosystem with high concentration of dissolved organic materials (Leong et al., 2012) as shown by the CDOM observed in the present study. The CDOM concentration in Johor Straits is high compared to the Singapore Strait. CDOM in the Johor Straits ecosystem has not been studied in detail and is poorly understood. In general, CDOM is known as an absorber of sunlight, in particular ultra-violet radiation (Belzile et al., 2002), and it is a major factor in determining the optical properties of coastal waters. CDOM can also serve as a source of nitrogen in marine waters (Stedmon et al., 2007).
Nutrients utilisation by the phytoplankton communities in coastal ecosystem is relatively complex. The nutrients were utilised differently by the different species during the study period (Figs. 7, 8). The reduced form of N source, namely NH4, was being utilised first, followed by the oxidised form of N (i.e., nitrate and nitrite) and then the reduced organic N, i.e., urea. Other nutrients such as P displayed a pattern similar to NH4, while Si followed the trend of nitrate. The nutrient concentrations in Johor Strait were relatively high, possibly influenced by significant terrestrial inputs which encourage high phytoplankton biomass (Gin et al., 2000; Leong et al., 2012). Depending on the composition of nutrients, these conditions increase the risk of HABs.

34
Leong et al.: Harmful algal blooms in Singapore waters
Studies have demonstrated that raphidophyte species such as Heterosigma akashiwo found in Johor Strait, exhibit high growth rates in response to nutrients (Kok & Leong, 2012).
The eutrophic conditions in Johor Strait could be an important driving force for bloom to develop under favorable physical settings. The present study indicates that blooms could be formed during a neap tide under favorable physical settings such as a stable water column, leading to high toxic cell density incidents. In particular, several N sources available at high concentration as compared to the Si concentration allow a bloom to be maintained over a prolonged period.
CONCLUSIONS
Over the past few years, we have detected new species of dinoflagellates in our local waters such as Karenia mikimotoi and its undetermined congeners. The present study identified the causative species responsible for the recent massive fish kills along Johor Strait as Takayama xiamenensis, Karlodinium cf. veneficum and K. cf. australe. This indicates that Singapore coastal waters are increasingly affected by HABs. The presence of harmful phytoplankton species in our waters is not well documented and identified. Further investigations of these species and their fish kill mechanisms are needed, as their toxin properties are poorly known. Our findings have also shown that multi-species dinoflagellate blooms can occur in different locations along the Johor Straits at different times. Again, detailed studies are required to understand the dynamics of these blooms and causal factors initiating such blooms. Such information and knowledge would be beneficial for the prevention and management of future occurrences of HABs in Singapore.
ACKNOWLEDGEMENTS
The Johor Straits marine biodiversity workshop on Pulau Ubin, Singapore was organised by the National Parks Board and National University of Singapore and held from 15 October to 2 November 2012 at Outward Bound School. The workshop, as part of the Comprehensive Marine Biodiversity Survey (CMBS) was supported by generous contributions from Asia Pacific Breweries Singapore, Care-for-Nature Trust Fund, Shell Companies in Singapore and The Air Liquide Group. We also wish to thank the management and staff of the Outward Bound School for kindly accommodating our special needs for a successful workshop. Staff and students from TMSI and Reef Ecology Laboratory assisted in collecting water samples. We are grateful to Kok S.P. from Soka University, Japan for assisting in the clone analysis.
LITERATURE CITED
Adam A, Mohammad-Noor N, Anton A, Saleh E, Saad S & Muhd Saleh SR (2011) Temporal and spatial distribution of harmful algal bloom (HAB) species in coastal waters of Kota Kinabalu, Sabah, Malaysia. Harmful Algae, 10: 495–502.
Aquino J, Flores B & Naguit M (2010) Harmful algal bloom occurrence in Murcielagos Bay amidst climate change. E-International Scientific Research Journal, 2: 358–365.
Bachvaroff TR, Adolf JE & Place AR (2009) Strain variation in Karlodinium veneficum (Dinophyceae): toxin profiles, pigments, and growth characteristics. Journal of Phycology, 45: 137–153.
Belzile C, Gibson JAE & Vincent WF (2002) Colored dissolved organic matter and dissolved organic carbon exclusion from lake ice: Implications for irradiance transmission and carbon cycling. Limnology and Oceanography, 47: 1283–1293
Carlos, AA, Baillie BK, Kawachi M & Maruyama T (1999) Phylogenetic position of Symbiodinium (Dinophyceae) isolates from tridacnids (Bivalvia), cardiids (Bivalvia), a sponge (Porifera), a soft coral (Anthozoa), and a free-living strain. Journal of Phycology, 35: 1054–1062.
Chen Y, Yan T, Yu R & Zhou M (2011) Toxic effects of Karenia mikimotoi extracts on mammalian cells. Chinese Journal of Oceanology and Limnology, 29: 860–868.
Chua TE (1973) An ecological study of the Ponggol estuary in Singapore. Hydrobiologia, 43: 505–533.
Coats DW & Park MG (2002) Parasitism of photosynthetic dinoflagellates by three strains of Amoebophrya (Dinophyta): parasite survival, infectivity, generation time, and host specificity. Journal of Phycology, 38: 520–528.
De Salas MF, Bolch CJS, Botes L, Nash G, Wright SW & Hallegraeff G M (2003) Takayama gen. nov. (Gymnodiniales, Dinophyceae), a new genus of unarmored dinoflagellates with sigmoid apical grooves, including the description of two new species. Journal of Phycology, 39: 1233–1246.
De Salas MF, Bolch CJS & Hallegraeff GM (2005) Karlodinium australe sp. nov. (Gymnodiniales, Dinophyceae), a new potentially ichthyotoxic unarmored dinoflagellate from lagoonal habitats of south-eastern Australia. Phycologia, 44: 640–650. Díez B, Pedrós-Alió C & Massana R (2001) Study of genetic diversity of eukaryotic picoplankton in different oceanic regions
by small-subunit rRNA gene cloning and sequencing. Applied and Environmental Microbiology, 67: 2932–2941.
Feng JL (2013) Jul 3. Thousands of fish dead as dry spell hits farms in Lim Chu Kang. Straits Times. http://www.straitstimes.com/breaking-news/singapore/story/thousands-fish-dead-dry-spell- hits-farms-lim-chu-kang-20130703 (Accessed 4 July 2013).
Gin KYH, Holmes MJ, Zhang S & Lin X (2006) Phytoplankton structure in the tropical port waters of Singapore. In: Wolanski E (ed.) The Environment in Asia Pacific Harbours. Springer, Netherlands. Pp 347–375.
Gin KYH, Lin X & Zhang S (2000) Dynamics and size structure of phytoplankton in the coastal waters of Singapore. Journal of Plankton Research, 22: 1465–1484.
Groisillier A, Massana R, Valentin K, Vaulot D & Guillou L (2006) Genetic diversity and habitats of two enigmatic marine alveolate lineages. Aquatic Microbial Ecology, 42: 277–291.
Gu H, Luo Z, Zhang X, Xu B & Fang Q (2013) Morphology, ultrastructure and phylogeny of Takayama xiamenensis sp. nov. (Gymnodiniales, Dinophyceae) from the East China Sea. Phycologia, 52: 256–265.
Guillou L, Viprey M, Chambouvet A, Welsh RM, Kirkham AR, Massana R, Scanlan DJ & Worden AZ (2008) Widespread occurrence and genetic diversity of marine parasitoids belonging to Syndiniales (Alveolata). Environmental Microbiology, 10: 3349–3365.
Hausmann K, Hülsmann N & Radek R (2003) Protistology. Third Revised Edition. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, 379 pp.
Hoagland P, Anderson, DM, Kaoru Y & White AW (2002) The economic effects of harmful algal blooms in the United States: estimates, assessment issues, and information needs. Estuaries, 25: 819–837.
Holmes MJ & Teo SLM (2002) Toxic marine dinoflagellates in Singapore waters that cause seafood poisonings. Clinical and Experimental Pharmacology and Physiology, 29: 829–836.

35
RAFFLES BULLETIN OF ZOOLOGY 2015
Hu JL (2014) Farms along East Johor Straits report fish mortality. Channel News Asia. http://www.channelnewsasia.com/news/singapore/farms-along-east-johor/988628.html (Accessed 10 February 2014).
Kamiyama T, Takayama H, Nishii Y & Uchida T (2001) Grazing impact of the field ciliate assemblage on a bloom of the toxic dinoflagellate Heterocapsa circularisquama. Plankton Biology and Ecology, 48: 10–18.
Kempton JW, Lewitus AJ, Deeds JR, Law JM & Place AR (2002) Toxicity of Karlodinium micrum (Dinophyceae) associated with a fish kill in a South Carolina brackish retention pond. Harmful Algae, 1: 233–241.
Khoo HW & Wee JY (1997) Phytoplankton monitoring and harmful algae occurrences in Singapore waters – historical perspective and status. In: Vigers G., Ong KS, McPherson C, Millson N, Watson I & Tang A (eds.), ASEAN Marine Environmental Management: Quality Criteria and Monitoring for Aquatic Life and Human Health Protection. Proceedings of the ASEAN-Canada Technical Conference on Marine Science (24–28 June 1996), Penang, Malaysia. EVS Environmental consultants, North Vancouver and Department of Fisheries, Malaysia. Pp. 58–67.
Kim GH (2010) An overview on the occurrences of harmful algal blooms (HABs) and mitigation strategies in Korean coastal waters. In: Ishimatsu A & Lie HJ (eds.) Coastal Environmental and Ecosystem Issues of the East China Sea. TERRAPUB, Nagasaki University. Pp. 121–131.
Knap A, Michaels A, Close A, Ducklow H & Dickson A (1996) Protocols for the Joint Global Ocean Flux Study (JGOFS) core measurements. JGOFS Report Nr. 19, vi+170 pp. Reprint of the IOC Manuals and Guides No. 29, UNESCO 1994.
Kok JWK & Leong SCY (2012) Growth and physiological responses of the Singapore strain of Heterosigma to various nitrogen sources and light conditions. Oceans 2012, IEEE, New York, p. 4.
Kok SP, Kikuchi T, Toda T & Kurosawa N (2012a) Diversity analysis of protistan microplankton in Sagami Bay by 18S rRNA gene clone analysis using newly designed PCR primers. Journal of Oceanography, 68: 599–613.
Kok SP, Kikuchi T, Toda T & Kurosawa N (2012b) Diversity and community dynamics of protistan microplankton in Sagami Bay revealed by 18S rRNA gene clone analysis. Plankton and Benthos Research, 7: 75–86.
Lee A (2014) 160 tonnes of dead fish found in farms along Johor Straits. Today Online. http://www.todayonline.com/ singapore/160-tonnes-dead-fish-found-farms-along-johor- straits/ (Accessed 25 March 2014).
Lim HC, Leaw CP, Tan TH, Kon NF, Yek LH, Hii KS, Teng ST, Razali, RM, Usup G, Iwataki M & Lim PT (2014) A bloom of Karlodinium australe (Gymodiniales, Dinophycease) associated with mass mortality of cage-cultured fishes in West Johor Strait, Malaysia. Harmful Algae, 40: 51–62.
Leong SCY, Chen PZX, Kok JWK, Chew SM & Lim LP (2014) Toxic bloom-forming phytoplankton species in Singapore coastal waters. IOC/WESTPAC 9th International Scientific Symposium (presentation), Nha Trang,Vietnam, 22–25 April 2014.
Leong SCY, Tkalich P & Patrikalakis NM (2012) Monitoring harmful algal blooms in Singapore: developing a HABs observing system. Oceans 2012, IEEE, New York, p. 5.
Leong SCY, Nakazawa M & Taguchi S (2006) Physiological and optical responses of the harmful dinoflagellate Heterocapsa circularisquama to a range of salinity. Hydrobiologia, 559: 149–159.
Leong SCY, Murata A, Nagashima Y & Taguchi S (2004) Variability in toxicity of the dinoflagellate Alexandrium tamarense in response to different nitrogen sources and concentrations. Toxicon, 43: 407–415.
Leong SCY & Taguchi S (2004) Response of the dinoflagellate Alexandrium tamarense to a range of nitrogen sources and concentrations: growth rate, chemical carbon and nitrogen, and pigments. Hydrobiologia, 515: 215–224.
Lim PT, Usup G & Leaw CP (2012) Harmful algal blooms in Malaysian waters. Sains Malaysiana, 41: 1509–1515.
López-García P, Rodríguez-Valera F, Pedrós-Alió C & Moreira D (2001) Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton. Nature, 409: 603–607.
Lu SH & Hodgkiss IJ (2004) Harmful algal bloom causative collected from Hong Kong waters. Hydrobiologia, 512: 231–238.
Moon-van der Staay SY, Watchter RD & Vaulot D (2001) Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature, 409: 607–610.
Mohammad-Noor N, Ong FS & Encik Weliyadi EA (2012) Seasonal distribution of harmful algal bloom species in East Coast of Sabah, Malaysia. Journal of Fisheries and Aquatic Science, 7: 431–438.
Mulvenna PF & Savidge G (1992) A modified manual method for the determination of urea in seawater using diacetylmonoxime reagent. Estuarine, Coastal and Shelf Science, 34: 429–438.
Park TG, Ok YR, ParkYT & Lee CY (2011) Temporal changes in the abundance of the fish-killing dinoflagellate Karlodinium veneficum (Dinophyceae) in Tongyeong, Korea. Algae, 26: 237–241.
Patton CJ & Crouch SR (1977) Spectrophotometric and kinetics investigation of the Berthelot reaction for the determination of ammonia. Analytical Chemistry, 49: 464–469.
Piwosz K & Pernthaler J (2010) Seasonal population dynamics and trophic role of planktonic nanoflagellates in coastal surface waters of the Southern Baltic Sea. Environmental Microbiology, 12: 364–377.
Place AR, Bowers HA, Bachvaroff TR, Adolf JE, Deeds JR & Sheng J (2012) Karlodinium veneficum—The little dinoflagellate with a big bite. Harmful Algae, 14: 179–195.
Quek C & Lim J (2010) Plankton bloom causes losses reaching hundreds of thousands of dollars. Wild Singapore. http:// wildsingaporenews.blogspot.com/2010/01/200000-fish-in- farmsoff-pasir-ris-dead.html (Accessed 25 January 2010).
Salleh NAM (2014) Pasir Ris mass fish deaths may lead to over $1 million loss for farmers. Straits Times. http://www.straitstimes. com/breaking-news/singapore/story/pasir-ris-mass-fish-deaths- may-lead-over-1-million-loss-farmers-201402 (Accessed 14
February 2014).Sobel J & Painter J (2005) Illnesses caused by marine toxins.
Clinical Infectious Diseases, 41: 1290–1296.Stedmon CA, Markager S, Tranvik L, Kronberg L, Slätis T &
Martinsen W (2007) Photochemical production of ammonium and transformation of dissolved organic matter in the Baltic Sea. Marine Chemistry, 104: 227–240.
Tang DL, Kawamura H, Doan-Nhu H & Takahashi W (2004) Remote sensing oceanography of a harmful algal bloom off the coast of southeastern Vietnam. Journal of Geophysical Research, 109: C03014
Tang YZ, Kong L & Holmes MJ (2007) Dinoflagellate Alexandrium leei (Dinophyceae) from Singapore coastal waters produces a water soluble ichthyotoxin. Marine Biology, 150: 541–549.
Tang YZ, Koch F & Gobler CJ (2010) Most harmful algal bloom species are vitamin B1 and B12 auxotrophs. Proceedings of the National Academy of Sciences of the United States of America, 107 (48): 20756–20761.
Tang YZ, Kong L, Morse RE, Gobler CJ & Holmes MJ (2012) Report of a bloom-forming dinoflagellate Takayama acrotrocha from tropical coastal waters of Singapore. Journal of Phycology, 48: 455–466.

36
Leong et al.: Harmful algal blooms in Singapore waters
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F & Higgins DG (1997) The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research, 25: 4876–4882.
Uchida T, Yamaguchi M, Matsuyama Y & Honjo T (1995) The red- tide dinoflagellate Heterocapsa sp. kills Gyrodinium instriatum by cell contact. Marine Ecology Progress Series, 118: 301–303.
Yamasaki Y, Kim DI, Matsuyama Y, Oda T & HonjoT (2004) Production of superoxide anion and hydrogen peroxide by the red tide dinoflagellate Karenia mikimotoi. Journal of Bioscience And Bioengineering, 97: 212–215.
Yee XY (2014) Mass death of fish hits Johor fishermen. The Star Online. http://www.thestar.com.my/News/Nation/2014/02/13/ Johor-Mass-Fish-Death (Accessed 13 February 2014).





























