Embed Size (px)
Citation preview

Three-dimensional localisation of DNA in the nucieolus of Spirogyra by
correlated optical tomography and serial ultra-thin sectioning
E. GWYN JORDAN*
Biophysics Section, Division of Biomolecular Sciences, King's College London, 26-29 Dniry Lane, London WC2B 5RL, UK
and DAVID J. RAWLINS
Institute of Plant Science Research, John Innes Institute, Colney Lane, Nonoich NR4 7UH, UK
* Author for correspondence
Summary
Spirogyra nucleoli were shown by three-dimen-sional optical microscopy of DAPI fluorescence tocontain DNA with a pattern and distribution match-ing those of the fibrillar centres. This was con-firmed using different species with nucleoli show-ing different sizes of fibrillar centre. Much lowerlevels of fluorescence •were seen corresponding to
the dense fibrillar component. Nearly all the DAPIfluorescence arises from the fibrillar centres orfrom regions very close to their surface, indicatingthat this is the site of nucleolar transcription.
Key words: nucieolus, DNA, fibrillar centres.
Introduction
The nucieolus is the cellular site of ribosome synthesis.When active it contains the transcribing genes for themajor ribosomal RNAs. The nascent RNA rapidly be-comes RNP through the binding of ribosomal proteins(Chooi and Leiby, 1981). There are many rRNA genes ineukaryotic cells and their combined activity results in alarge accumulation of pre-ribosomal RNP particles in allstages of maturation. The resultant mass of protein,RNA and chromatin is the nucieolus (Goessens, 1984;Jordan et al. 1980; Stahl, 1982). It contains structurallydistinct components and the elucidation of the molecularbasis for the structural differences, especially the dis-covery of the location in situ of the transcribing rRNAgenes, is one of the first steps towards a full understand-ing of the nucieolus.
Nucleolar rRNA genes in the process of transcriptionhave been shown most clearly by the Miller spreadingtechnique (Miller and Beatty, 1969; Trendelenburg,1983). Notwithstanding the great insights this techniquehas afforded, we still do not have a clear idea of preciselywhere rRNA transcription units reside in the intactnucieolus.
The majority of reports conclude that the densefibrillar component is the site of active genes withinnucleoli (terms describing nucleolar ultrastructure areused according to the agreed nomenclature resulting fromthe discussion at the 8th Nucleolar Workshop; seeJordan, 1984; and see Fig. 3 of this paper). Wachtler etal. (1989) reviewed the literature for this view in a recent
Journal of Cell Science 95, 343-352 (1990)Printed in Great Britain © The Company of Biologists Limited 1990
paper and presented some good new evidence from in situhybridisation with fluorescence, in its favour.
Some authors, however, have identified structures thatthey believe to be the transcribing genes in intactnucleoli, which seem to correspond to the granularcomponent (Ghosh and Paweletez, 1987; Troster et al.1985; Hameister, 1977). These results parallel the obser-vation of transcription units in another product of massedgene activity - the Balbiani ring of Chironomus - wherethey are found in a peripheral granular cloud (Lamb andDaneholt, 1979; Olins and Olins, 1980). However, theidea that the granular component of nucleoli is thelocation of transcribing genes is not supported by anyother evidence.
In contrast, the first EM immunogold labelling studiessuggested that active rRNA genes are to be found neitherin the granular nor even in the dense fibrillar component,but solely within the fibrillar centres (Scheer and Rose,1984; Thiry et al. 1988a,b). These studies demonstratethat antibodies to polymerase I and DNA do not bind inthe granular component or the dense fibrillar componentbut only in the fibrillar centres. A further study by Thiry(1988) using bromodeoxyuridine (BrdUrd) incorpor-ation and monoclonal anti-BrdUrd antibodies supportsthis view. More-recent work using gold-labelled anti-bodies has confirmed that there is a high concentration ofpolymerase I and DNA in fibrillar centres but has shownin addition a low level of these in the dense fibrillarcomponents of very active cells (Raska et al. 1989). Theseworkers also showed that DNA topoisomerase I, whichmight be expected to coincide with the transcription sites
343

(Sternglanz, 1989), is found in the dense fibrillar com-ponent. However, another recent paper using rDNA insitu hybridisation at the electron microscope failed toreveal rDNA in the dense fibrillar component (Thiry andThiry-Blaise, 1989).
Thus, although there have been many attempts todescribe the structure of the nucleolus in relation to itsmolecular functions, in particular the location of theactive transcripts, the situation remains unresolved (Del-tour and Mosen, 1987; Jordan, 1987a,b; Goessens,1984). To this end we have studied the distribution ofDNA in the large well-differentiated nucleolus of Spiro-gyra.
Materials and methods
MaterialsTwo species of Spirogyra were used. The first was Spirogyragrevilleana (Griffen & George, Gerard Biological Centre, EastPreston, Sussex; catalogue no. ZDC-29SK), which wasassigned the number CCAP 678/1 by the Cambridge CultureCollection of Algae and Protozoa. This had filaments about40,um in diameter. The second, which had 25 ftm filaments,resulted from a collection made in a small river called 'The Rife'at Ferring, Sussex and was classified ZEC-210-030H (Griffen &George). Here we refer to it as 'species 210'. It is no longer inculture.
Algae were maintained in Bold's basal medium (Bischoff andBold, 1963) at 15°C, with a 18-6h light-dark cycle.
DArA fluorescenceSpirogyra filaments were fixed in 4% formaldehyde (freshlyprepared from paraformaldehyde) in buffer containing 50mMPipes, S mM EGTA and S mM MgSO4. Without further pre-treatment, they were then stained in ljUgml"1 DAPI for30-60 min, rinsed and mounted in anti-fade mountant (Citi-fluor; Citifluor Ltd, City University, London).
KaryotypesKaryotypes of Spirogyra grevilleana were prepared either withDAPI or by the iron alum acetocarmine technique of Godvvard(1948).
Light microscopySlides were examined with a Zeiss Universal microscopeequipped for epifluorescence and using a Leitz x63/NA 1.4objective. Images were obtained using a low light level ISITvideo camera linked to a framestore and VAX host computer asdescribed previously (Rawlins and Shaw, 1988). Optical sec-tions were collected at 0.5 j.lm z spacing under computercontrol, the fine focus of the microscope being stepped auto-matically using a microstepping motor. 512 video frames wereaveraged to reduce electronic noise from the camera beforestorage of the final image on disk. Optical sections weredeblurred as described previously (Rawlins and Shaw, 1988)using a simple, nearest-neighbour, deblurring algorithm (Wein-stein and Castleman, 1971; Castleman, 1979; Agard, 1984).Stacks of optical sections were projected at several differentangles about the z axis (Agard et al. 1988) and the projectionsviewed as stereo-pairs or in quick succession to give the effect ofrotation. Alternatively, tracings of micrographs of the opticalsections were made onto spaced acetate sheets and photo-graphed to give stereo-pairs as described previously for EM(Jordan and Saunders, 1976).
Electron miavscopyFilaments of Spirogyra were fixed in 1.5% glutaraldehyde in0.035 M sodium cacodylate with 5 % sucrose at room tempera-ture for 1 h. They were then washed in buffer, dehydratedthrough an ethanol series, washed in propylene oxide andembedded in Araldite or in Spurr's resin. Serial sections(0.1 fim) were cut and prepared as described by Bennett et al.(1979). Electron micrographs with magnifications of X 18000 orX15 000 were used to trace the section profiles of the fibrillarcentres. Three consecutive sections were traced onto oneacetate sheet, giving 20-30 sheets in the complete stack for asingle nucleolus. Stereo-photographs of the electron micro-graphs were prepared from submerged tracings on acetatesheets as described previously (Jordan and Saunders, 1976).For accurate registration of the serial sections in the stacks weused the outlined position of the nucleus, the nucleolus andother organelles.
Results
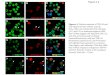
In DAPI-stained Spirogyra, the nucleoli appeared asdark spheres occupying a large proportion of the brightlyfluorescing nuclei (Fig. 1). Areas of fluorescence werealso visible inside the nucleolus. After deblurring of theoptical sections, in Spirogyra grevilleana, these areas hadthe appearance of many small spots or strings of spotswith diameters between about 0.1 and 0.3 /im (Fig. 1). Inaddition, a few characteristically larger spots measuringbetween 0.6 and 1.1 jUm were present and occupied amore peripheral position or appeared on the boundarybetween the nucleolus and the nucleus.
Examination of the optical sections and stereo-pairs ofprojections (Fig. 2A) or acetate profile stacks(Fig. 2B-D) indicated that this arrangement occurredthroughout the whole nucleolus in Spirogyra grevilleana.In addition, there appeared a very low level of fluor-escence around and between some of the spots.
Because precise measurements of fluorescence spots atthe limits in size of the resolution of the light microscopeare not very accurate, we examined the pattern of thedistribution of the fibrillar centres in electron micro-graphs of nucleoli of the same species. Nine nuclei wereserially sectioned and photographed. Fig. 3A shows asingle electron micrograph of a nucleolus from Spirogyragrevilleana. There was a marked similarity between thearrangement of the fibrillar centres in the electronmicrographs and that of the DAPI-labelled DNA. Inserial EM sections (Fig. 3B-G) the fibrillar centres hadcircular profiles between 0.13 and 0.3 /im in diameter andoccasional more-peripheral larger ones between 0.4 and0.8/im in diameter; these sizes correspond well to thoseobserved in the DAPI-stained nucleoli. In places, pro-files indicative of a longer structure were seen (Fig. 3E),but the general picture was of a bumpy or beaded threadcomposed of small fibrillar centres arranged as folded andtwisted strings of small spheres. Examination of stereo-pair reconstructions of EM serial sections (Fig. 4A-B)reinforced this conclusion. In some nucleoli, the fibrillarcentres were smaller and more scattered (Fig. 4A),indicating the variability encountered in Spirogyra grevil-leana when grown at 15°C.
It was not possible unambiguously to define a certain
344 E. G. Jordan and D. J. Rawlins

Fig. 1. A-0. De-blurred optical sections of DAPI-stained Spirogyra grevilleana nucleus. The nucleolus appears as a darksphere and contains small fluorescent spots arranged in folded rows. Between and around the small fluorescent spots is a lowerlevel of fluorescence. X3600.
number of strings of fibrillar centres in these nucleoli,due to their close association and the difficulty ofresolving pathways through regions of close entangle-ment. However, serial sections (Fig. 3B-G) show that
there is continuity between adjacent sections of thecomplexes resulting from the ensheathing of fibrillarcentres in dense fibrillar material. We did not observebranching of the strings.
DNA in Spirogyra nucleolus 345

Fig. 2. A. Stereo-pair of two projections throughthe images in Fig. 1. X3200. B. Stereo-pairreconstruction of profiles traced from the sectionsin Fig. 1 at the same magnification as A. C,D. Stereo-pair reconstructions as in B, but of twoother nm leoli from the same species to show thevariability in DNA distribution. X3200.
Although we were unable to determine the number ofloops directly we could count their ends. We recorded thenumber of points at which the fibrillar centres broke thesurface of the nucleolus to join the DNA of the nucleo-plasm (Fig. 3E, F). This was eight times for Spirogyragrevilleana, which is consistent with the hypothesis thatthey are the ends of four intranucleolar organizers.
From examination of the karyotypes, there seemed tobe 16 chromosomes, although we could also interpret thekaryotype as 14 with two satellites. We have not deter-
mined the number of organising chromosomes for thisspecies, but four seems most probable from the nucleolarultrastructure.
Species 210 showed a rather different nucleolar mor-phology. In the DNA-stained images, two discrete intra-nucleolar DNA structures could be resolved, one largerthan the other (Fig. 5A,B). In electron micrographs ofthis species, larger fibrillar centres were seen than thosein Spirogyra grevilleana (Fig. 5C-D, Fig. 6). Also, thepattern of fibrillar centres showed Vip as thick folded
346 E. G. Jordan and D. J. Rawlins

3A
2*
*Wi
.•'.'•/M^aiffiL
Fig. 3. Electron micrographs of a nucleolus of Spirogyra grevilleana. A. The spherical nucleolus shows the three structuralcomponents: fc, fibrillar centre; dfc, dense fibrillar component; and gc, granular component. X12000. B-G. Adjacent 0.1 /imsections showing small and larger fibrillar centres and an entry point (arrow) of a chain of fibrillar centres. X5900.
DNA in Spirogyra nucleolus 347

r*. "44A «•*>•?*•
Fig. 4. Stereo-pair reconstructions ofprofiles from serial electronmicrographs through a nucleolus ofSpirogyra grevilleana. A. Thisnucleolus has small fibrillar centresarranged in folded coils. X3200.B. Larger fibrillar centres are presentin this nucleolus as well as the smallerones. An invagination of the nuclearenvelope (arrow) abuts the nucleolusat the start point of one of theorganizers adjacent to a largeperipheral fibrillar centre. X3200.
bands rather than beads or strings of spots (Fig. 5D).Thus, the DNA pattern closely resembled that of thefibrillar centres.
Discussion
The intranucleolar DNA of Spirogyra can be visualisedby three-dimensional fluorescence microscopy of DAPI-stained cells. In Spirogyra grevilleana this DNA isdistributed throughout the nucleolus as small sphericalmasses between 0.1 jttm and 1 fim in diameter arranged asstrings of beads, which by electron microscopy have beenshown to correspond to the fibrillar centres of nucleoli.The pattern of the nucleolar DNA is variable and weinterpret this as reflecting patterns of nucleolar structurerelating to different points in the cell cycle or variablerates of cell metabolism and rRNA transcription. It hasbeen shown previously that the fibrillar centres are largerand less well distributed throughout the nucleolus inearly Gi (Jordan and Godward, 1969) and active cellsdisplay smaller more scattered fibrillar centres (Jordanand Chapman, 1971, 1973; Jordan and McGovern,1981). The species with the very large fibrillar centres(species 210) may have responded to a metabolic shockreceived during transfer of the cells from the wild habitatto laboratory conditions, and were perhaps alsoinfluenced by the higher density of cells in the laboratoryculture. We also saw small scattered rows of fibrillarcentres in this species by electron microscopy. Weinterpret the nucleoli with the smallest most-scatteredfibrillar centres as being the most active in rRNA
synthesis (Jordan and McGovern, 1981). The appearanceof a few larger fibrillar centres at the periphery of thenucleolus indicates either a slowing of transcription or astage before the development of full transcriptionalpotential (Jordan and Luck, 1976). But all patterns ofDNA distribution encountered in this study and theircorrespondence with nucleolar ultrastructure are in har-mony with previous work on Spirogyra identifying thefibrillar centres as looped-out nucleolar-organizing re-gions (Godward, 1950; O'Donnell, 1961; Godward andJordan, 1965; Ashraf and Godward, 1980). But this workextends those previous studies by showing the bumpy orbeaded nature of the loop and its correspondence to thefibrillar centres seen by electron microscopy. The findingthat the DNA within the fibrillar centres is rDNA issupported by in situ hybridisation studies at both thelight and the electron-microscope levels (Arroua et al.1982; Thiry and Thiry-Blaise, 1989). Although thepattern of fibrillar centre distribution for higher plants isreported to show branching (Motte et al. 1988), we haveno evidence for this in our work on Spirogyra.
When we turn to the problems of identifying the DNAof the nucleolus that corresponds to the axis of transcrip-tion units we must consider the weaker diffuse fluor-escence that occurs around and between the smallerDNA accumulations of the fibrillar centres. Is thereenough DNA outside the fibrillar centres to conform withthe traditional interpretations that point to the densefibrillar components and not the fibrillar centres as thesites of transcription (Hadjiolov, 1985; Jordan et al.1980; Stahl, 1982; Wachtler et al. 1989) and the zonefrom which the characteristic transcription units seen in
348 E. G. Jordan and D. J. Rawlins

B
Fig. 5. Species 210. A. Projection of deblurredoptical sections showing the two largeintranucleolar DNA structures. X3200.B. Reconstruction of DNA profiles as is Fig. 2B.Two regions of DNA, one a third the size of theother, can be seen. X3200. C-D. Reconstructionof EM profiles of two other nucleoli of species 210.C. Two large fibrillar centres comparable to theDNA distribution in A and B. X5000. D. Longercoils of more scattered fibrillar centres giving theorganizers a 'fluffy' loop appearance. XS000.
spread preparations have been said to have been derived(Jordan, 1987a)?
The most important point to establish in this connec-tion is the extent to which DNA occurs 'outside' fibrillarcentres and whether there is enough of it there to supportnucleolar transcription. Two considerations arise.
First, it is necessary to define how closely the distri-bution of fluorescent spots coincides with the fibrillarcentres. For instance, does some of the brighter fluor-escence arise from the dense fibrillar component that isaround the fibrillar centres or is it precisely confined
within their boundaries? It is difficult within the limits ofthe optical microscope and when using nucleoli fromdifferent phases of the cell cycle with variations in theirsizes of fibrillar centres to be as accurate as we wouldwish. This is especially true since the small fibrillarcentres and the thickness of the dense fibrillar componentthat ensheathes all fibrillar centres are just below thelimits of resolution of the light microscope (around0.1 /im). Nevertheless, we have made preliminarymeasurements that indicate that fluorescent spots areslightly larger than the fibrillar centres. Work on cells
DNA in Spirogyra nucleolus 349

gc
i
Fig. 6. A single section from a nucleus like that in Fig. 5C. The fibrillar centre forms a thick strand ensheathed in a layer ofdense fibrillar component with a very even thickness of around 0.17;Um surrounded by other layers of dense fibrillar componentdisplaying reticulate or nucleolonemal structure with the same regular thickness. The granular component (gc) is segregated intotwo discrete peripheral regions on either side of the other components. X23 000.
with uniformly sized fibrillar centres will be needed toconfirm this. If the interpretation is substantiated itwould not conflict with interpretations that place mostnucleolar transcription at the surface of the fibrillarcentres in the closely associated dense fibrillar com-ponent. It is not possible with small fibrillar centres todistinguish surface fluorescence from total fluorescence.
The second point, which requires careful analysis, isthe extent to which a low level of fluorescence indicatesDNA around and between the brighter spots in a positioncoinciding with the dense fibrillar component. Ourresults show very low levels of fluorescence in thesepositions and would conform with the most recent workon Poll and topoisomerase I localisation at the EM level(Raska et al. 1989) and the Feulgen-like osmium-ammine work of Derenzini et al. (1987). The very lowlevels of fluorescence, however, may be insufficient tojustify the conclusion that all transcription occurs here.Further, we observe that the amount of DNA in thefibrillar centres is very substantial and the number offibrillar centres very high, and in those nucleoli with thesmallest and most scattered fibrillar centres the idea thatthey alone can account for all nucleolar transcription isnot unreasonable and agrees closely with Scheer andRoses's interpretation that all transcription within nu-cleoli can be accounted for by the fibrillar centres. Itmight be argued that the bright fluorescence of thefibrillar centres indicates a level of chromatin conden-
sation incompatible with transcription. This argumentcan be met by noting that the DNA of the chloroplastgenomes, which is very dispersed, appears as small brightspots after DAPI staining, similar to the fibrillar centresof nucleoli. The chromatin of fibrillar centres is nottherefore necessarily condensed. The structure of fibril-lar centres as seen by electron microscopy also shows thatthe chromatin of fibrillar centres is dispersed and evenlacks nucleosomes (Derenzini et al. 1987). The low levelsof DNA fluorescence that we see in the dense fibrillarcomponent do not necessarily contradict the conclusionsbut may simply correspond to that DNA that linkstranscription sites together. We concede that the work ofWachtler et al. (1989), Raska et al. (1989), Derenzini etal. (1987) and the early conclusions of a large body ofresearch (see Stahl, 1982, for references) are opposed tothis interpretation and show the need for more rigorouswork before a new consensus is likely to be reached.
A problem in relating other organisms to Spirogyra isthe failure to reveal a similar distribution of DAPI-stained chromatin in their nucleoli. Spring et al. (1988)have seen small isolated DNA spheres by Feulgenfluorescence using confocal laser scanning microscopy, anobservation that we have confirmed (unpublished obser-vations) using conventional epi-fluorescence and DAPIon MRC5 fibroblasts, but in no case was the pattern offluorescence related to the distribution of fibrillar centres.In fact it seems that only occasional clumps of hetero-
350 E. G. Jordan and D. jf. Rawlins

chromatin were responsible for the patterns observed.The only conclusions that seems possible are either thatchromatin has a lower concentration in the fibrillarcentres of other organisms or that we have yet to establishthe right conditions to reveal it.
If DAPI staining is not revealing all the DNA innucleoli of other organisms we must be prepared todiscover that the distribution of DNA may yet be widerthan we have so far found in Spirogyra nucleoli. Answersto some of these questions should come from fullquantitative analysis of selected cell types at definedpositions in the cell cycle with uniform transcriptionalstatus.
The observations of what seem to be transcriptionunits in the granular component (Ghosh and Paweletz,1987; Hameister, 1977; Troster et al. 1985) and the axialstructures with attached fibres and granules shown thereby Olins and Olins (1980) in Chironomus salivary glandnucleoli warn us against too complacent an acceptance ofour current views.
The new power of image processing demonstratingthat single DNA molecules can be visualised using thelight microscope point the way ahead (Schwartz andKoval, 1989).
We thank Drs D. A. Agard and J. W. Sedat (University ofCalifornia, San Francisco) and P. J. Shaw (J.I.I.) for gener-ously providing us with their computer programs for deblurringand projection of optical sections, and Mrs Jane Penny ofGerard Biological Centre for directing us to the sources for thespecies of Spirogyra used. D.R. was supported by the Agricul-tural and Food Research Council via a grant-in-aid to the JohnInnes Institute. Support was also received from the GatsbyFoundation.
References
AGARD, D. A. (1984). Optical sectioning microscopy: Cellulararchitecture in three dimensions. A. Rev. Biophys. Bioengng 13,191-219.
AGARD, D. A., HIROAKA, Y. AND SEDAT, J. W. (1988). Three-dimensional light microscopy of diploid Drosophila chromosomes.CellMotil. Cyloskel. 10, 18-27.
ARROUA, M., HARTUNG, M., DEVICTOR, M., BERGE-LEFRANC, J. ANDSTAHL, A. (1982). Localisation of ribosomal genes by in situhybridisation in the fibrillar centre of the nucleolus in the humanspermatocyte. Biol. Cell 44, 337-340.
ASHRAF, M. AND GODWARD, M. B. E. (1980). The nucleolus intelophase, interphase and prophase.,7. Cell Sci. 41, 321-329.
BENNETT, M. D., SMITH, J. B., SIMPSON, S. AND WELLS, B. (1979).
Intranuclear fibrillar material in cereal pollen mother cells.Chromosoma 71, 289-332.
BISCHOFF, H. AND BOLD, H. C. (1963). Some soil algae fromEnchanted Rock and related algal species. Phycological Studies.IV. University of Texas Publ., no. 6318, pp. 1-95.
CASTLEMAN, K. R. (1979). Optical sectioning. In Digital ImageProcessing, pp. 351-360. Prentice Hall, New Jersey, USA.
CHOOI, W. AND LEIBY, K. (1981). An electron microscopic methodfor the localization of ribosomal proteins during transcription ofribosomal DNA: a method for studying protein assembly. Proc.natn. Acad. Sci. U.S.A. 78, 4823-4827.
DELTOUR, R. AND MOSEN, H. (1987). Proposals for themacromolecular organization of the higher plant nucleolonema.Biol. Cell 60, 75-86.
DERENZINI, M., HERNANDEZ-VERDUN, D., FARABEGOLI, F. ANDNOVELLO, F. (1987). Structure of ribosomal genes of mammaliancells in situ. Chromosoma 95, 63-70.
GHOSH, S. AND PAWELETZ, N. (1987). Active ribosomal cistrons andtheir primary transcripts in the nucleolus. Cell Biol. hit. Rep. 11,205-210.
GODWARD, M. B. E. (1948). The iron alum acetocarmine method foralgae. Nature, Land. 161, 203.
GODWARD, M. B. E. (1950). On the nucleolus and nucleolarorganising chromosomes of Spirogyra. Ann. Bot. 14, 39-53.
GODWARD, M. B. E. AND JORDAN, E. G. (1965). Electronmicroscopy of the nucleolus of Spirogyra britannica and Spirogyraellipsospora.J. R. Microsc. Soc. 84, 347-360.
GOESSENS, G. (1984). Nucleolar structure. Int. Rev. Cvtol. 87,107-158.
HADJIOLOV, A. A. (1985). The Nucleolus and Ribosome Biogenesis.Cell Biol. Monographs, 12. Springer-Verlag, Wein.
HAMEISTER, H. (1977). RNA synthesis in isolated polytene nucleifrom Chironomus teutons. Chromosoma 62, 217-242.
JORDAN, E. G. (1984). Nucleolar nomenclature. J. Cell Sci. 67,217-220.
JORDAN, E. G. (1987a). At the heart of the nucleolus. Nature, Loud.329, 489-490.
JORDAN, E. G. (19876). Nucleolar organizers in plants. InChromosomes Today, vol. 9 (ed. Stahl. A., Luciani, J. M. andVagner-Capadano, A. M.), pp. 272-283. Allen and Unwin,London.
JORDAN, E. G. AND CHAPMAN, J. M. (1971). Ultrastructural changesin the nucleoli of Jerusalem artichoke (Helianthus tubemstts) tuberdiscs. J. exp. Bot. 22, 627-634.
JORDAN, E. G. AND CHAPMAN, J. M. (1973). Nucleolar and nuclearenvelope ultrastructure in relation to cell activity in discs of carrotroot {Daucus carota L.). J. exp. Bot. 24, 197-209.
JORDAN, E. G. AND GODWARD, M. B. E. (1969). Some observationson the nucleolus of Spirogyra. J. Cell Sci. 4, 3-15.
JORDAN, E. G. AND LUCK, B. T. (1976). The nucleolus organizerand the synaptonemal complex in Endymion non-scriptus (L.).J.Cell Sci. 22, 75-86.
JORDAN, E. G. AND MCGOVERN, J. M. (1981). The quantitativerelationship of the fibrillar centres and other nucleolar componentsto changes in growth conditions, serum deprivation and low dosesof actinomycin D in cultured diploid human fibroblasts (strainMRC-5).J. Cell Sci. 52, 373-389.
JORDAN, E. G. AND SAUNDERS, A. M. (1976). The presentation ofthree-dimensional reconstructions from serial sections. J. Microsc.107, 205-206.
JORDAN, E. G., TIMMIS, J. N. AND TREWAVAS, A. J. (1980). The
Plant Nucleus. In The Biochemistry of Plants, vol. 1 (ed. E. Connand P. Stumpf), pp. 489-588. Academic Press, New York,London.
LAMB, M. M. AND DANEHOLT, B. (1979). Characterization of activetranscription units in Balbiani rings of Chironomus tentans. Cell17, 835-848.
MILLER, P. L. AND BEATTY, B. R. (1969). Visualization of nucleolargenes. Science 164, 955-997.
MOTTE, P., DELTOUR, R., MOSEN, H. AND BRONCHART, R. (1988).
Three-dimensional electron microscopy of the nucleolus andnucleolus-associated chromatin (NAC) during early germination ofZea mays L. Biol. Cell 62, 65-81.
O'DONNELL, E. H. J. (1961). DNA structures in the nucleolus.Nature, bond. 191, 1325-1326.
OLINS, A. L. AND OLINS, D. E. (1980). Stereo-electron microscopyof nucleoli, Balbiani rings and endoplasmic reticulum inChironomus salivary gland cells. Eur.J. Cell Biol. 22, 714-723.
RASKA, I., REIMER, G., JARNIK, M., KOSTROUCH, Z. AND RASKA, K.
(1989). Does the synthesis of ribosomal RNA take place withinnucleolar fibrillar centres or the dense fibrillar components? Biol.Cell 65, 79-82.
RAWLINS, D. J. AND SHAW, P. J. (1988). Three-dimensionalorganization of chromosomes of Crepis capillaris by opticaltomography. J . Cell Sci. 91, 401-414.
SCHEER, U. AND ROSE, K. (1984). Localization of RNA polymerase Iin interphase cells and mitotic chromosomes by light and electronmicroscopic immunocytochemistry. Proc. natn. Acad. Sci. U.S.A.81, 1431-1435.
SCHWARTZ, D. C. AND KOVAL, M. (1989). Conformational dynamics
DNA in Spirogyra nucleolus 351

of individual DNA molecules during gel electrophoresis. Nature,Land. 338, 520-522.
SPRING, H., TRENDELENBURG, M. F. AND MONTAG, M. (1988).
DNA-fluorescence of mammalian intra-nucleolar chromatindetected by confocal laser scanning microscopy (CLSM). Biol. Cell64, 371-374.
STAHL, A. (1982). The nucleolus and nucleolar chromosomes. In Themicleolus (ed. Jordan, E. G. and Cullis, C. A.), pp. 1-24.Cambridge University Press.
STERNGLANZ, R. (1989). DNA topoisomerases. Curr. Opinion CellBiol. 1, 533-535.
THIRY, M. (1988). Immunoeleetron microscope localization ofbromodeoxyuridine incorporated into DNA of Ehrlich tumour cellnucleoli. Expl Cell Res. 179, 204-213.
THIRY, M., SCHEER, U. AND GOESSENS, G. (1988«). Localization of
DNA within Ehrlich tumour cell nucleoli by immunoeleetronmicroscopy. Biol. Cell 63, 27-34.
THIRY, M., SCHEER, U. AND GOESSENS, G. (19886).
Immunoeleetron microscopic study of DNA during mitosis inEhrlich tumour cells. Eur.J. Cell Biol. 47, 346-357.
THIRY, M. AND THIRY-BLAISE, L. (1989). In situ hybridisation at the
electron microscope level: an improved method for preciselocalization of ribosomal DNA and RNA. Eur.J. Cell Biol. 50,235-243.
TRENDELENBURG, M. F. (1983). Progress in visualization ofeukaryotic transcription. Hum. Genet. 63, 197-215.
TROSTER, H., SPRING, H., MEISSNER, B., SCHULTZ, P., OUDET, P.
AND TRENDELENBURG, M. F. (1985). Structural organization of anactive, chromosomal nucleolar organizer region (NOR) identifiedby light microscopy, and subsequent TEM and STEM electronmicroscopy. Chromosoina 91, 151-163.
WACHTLER, F., HARTUNG, M., DEVICTOR, M., WIEGANT, J., STAHL,
A. AND SCHWARZACHER, H. G. (1989). Ribosomal DNA is locatedand transcribed in the dense fibrillar component of human sertolicell nucleoli. Expl Cell Res. 184, 61-71.
WEINSTEIN, M. AND CASTLEMAN, K. R. (1971). Reconstructing 3Dspecimens from 2D section images. Proc. Soc. Photo. Opt. Instr.Eng. 26, 131-138.
{Received 25 October 1989-Accepted 14 December 1989)
352 E. G. Jordan and D. J. Rawlins