Embed Size (px)
Citation preview

This article was originally published in the Encyclopedia of Animal Behavior published by Elsevier, and the attached copy is provided by Elsevier for the
author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in
instruction at your institution, sending it to specific colleagues who you know, and providing a copy to your institution�’s administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open
internet sites, your personal or institution�’s website or repository, are prohibited. For exceptions, permission may be sought for such use through
Elsevier's permissions site at:
http://www.elsevier.com/locate/permissionusematerial
Cocroft R.B. (2010) Vibrational Communication. In: Breed M.D. and Moore J., (eds.) Encyclopedia of Animal Behavior, volume 3, pp. 498-505 Oxford:
Academic Press.
© 2010 Elsevier Ltd. All rights reserved.

Author's personal copy
Vibrational CommunicationR. B. Cocroft, University of Missouri, Columbia, MO, USA
ã 2010 Elsevier Ltd. All rights reserved.
Introduction
Using Substrate Vibrations to Communicate
Of all forms of communication that make use of mechan-ical vibrations propagating through a medium, vibrationalcommunication is by far the most common. However,it is also the least familiar to biologists, and the leaststudied. One consequence of this lack of attention is thatmany of our inferences about this communication modal-ity are based on a relatively small number of studies.Another is that many opportunities remain for ground-breaking study of a complex and fascinating mode ofcommunication that occurs in taxa ranging from spidersto elephants.
How does the use of vibrations differ from that ofairborne sound? The two modalities are closely related.However, while organisms using sound are embeddedwithin the medium they use for communication, thoseusing vibration are at the boundary between air and adenser medium such as soil, water, or a solid structure likea plant stem. Communication takes place among organ-isms in contact with this dense medium, using vibrationsthat travel along its surface. Sound in air or water propa-gates as a longitudinal pressure wave, with motion of themedium occurring in the direction of travel of the wave.In contrast, substrate vibrations propagate as a variety ofwave types, with receivers generally detecting the com-ponent of medium motion perpendicular to the directionof travel of the wave. This difference in the nature ofsignal propagation leads to differences in the structuresused to produce and detect sound and vibration, and inthe properties of the signals themselves.
Taxonomic Distribution
Terrestrial animal species generally spend much or all oftheir time in contact with a substrate. Vibrations intro-duced into that substrate by the movement of other organ-isms provide a rich source of information about thebehavior and proximity of conspecifics, predators, orprey. Many species, from worms to spiders to elephants,have evolved means of detecting these vibrations, and thedetection of substrate vibrations underlies many social andecological interactions. A recurring theme in the study ofvibrational communication is the discovery of taxa inwhich individuals not only passively monitor substratevibrations but also introduce vibrations into the substratein order to influence the behavior of other individuals.
The use of substrate vibrations for communication iswidespread in arthropods, including insects, crabs, andspiders and other arachnids. It is estimated that 265 000species of insects communicate with substrate vibration.However, the true number is probably much larger,because this estimate includes only the species that havebeen formally named. For many groups of insects, thereare far more species yet to be described; for example, inaddition to the 20 000 described species of leafhoppers,there are an estimated 80 000 additional undescribed spe-cies, mostly in tropical forests. Whatever the real num-bers, it is probably safe to say that 98% of the diversity ofinsect vibrational signals remains unrecorded. A similarsituation likely exists in the other arthropod groups.
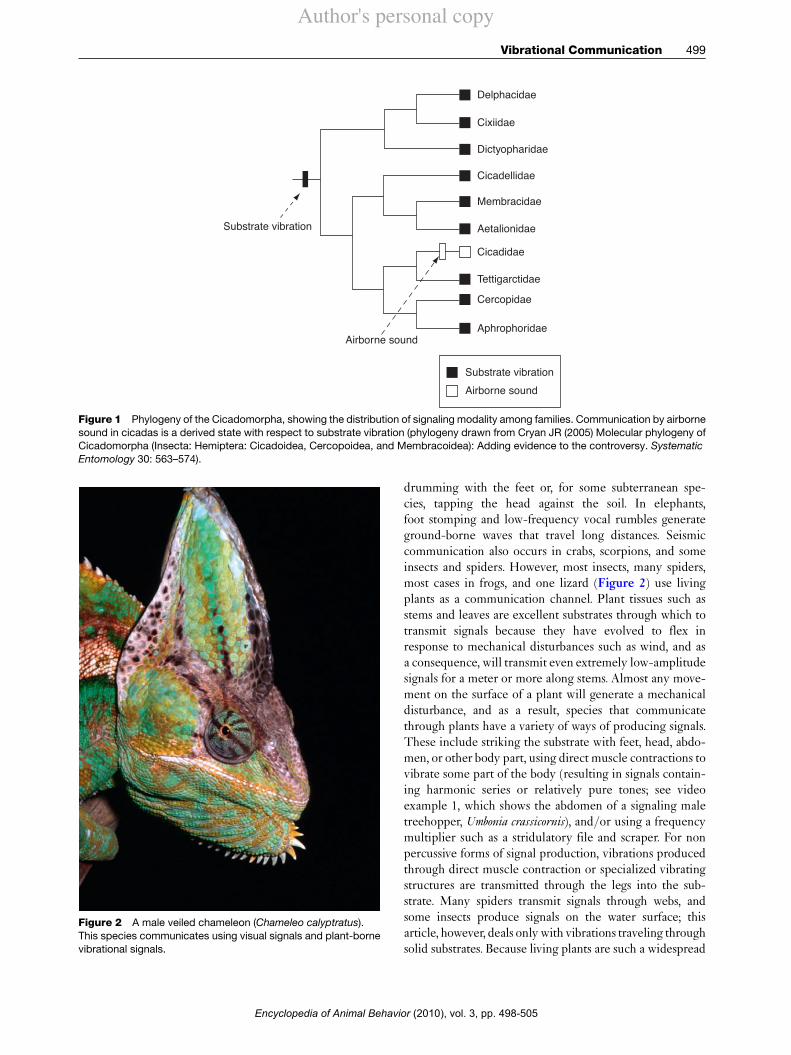
In insects, vibrational communication represents anancient communication modality, evolving before the ori-gin of airborne sound communication. The cicadas andtheir relatives provide a good illustration: vibrational com-munication is widespread, while airborne sound occursonly in the cicadas, where it is clearly derived with respectto vibrational communication (Figure 1). Even within thecicadas, vibrational communication occurs in basal spe-cies, and it is possible that it occurs in many others, used incombination with airborne sound. Recent discovery ofvibrational signaling in a Gondwanan relict species,along with comparative phylogenetic analysis, suggeststhat vibrational signaling may have evolved in the ances-tors of today’s Hemiptera, some 230 million years ago.
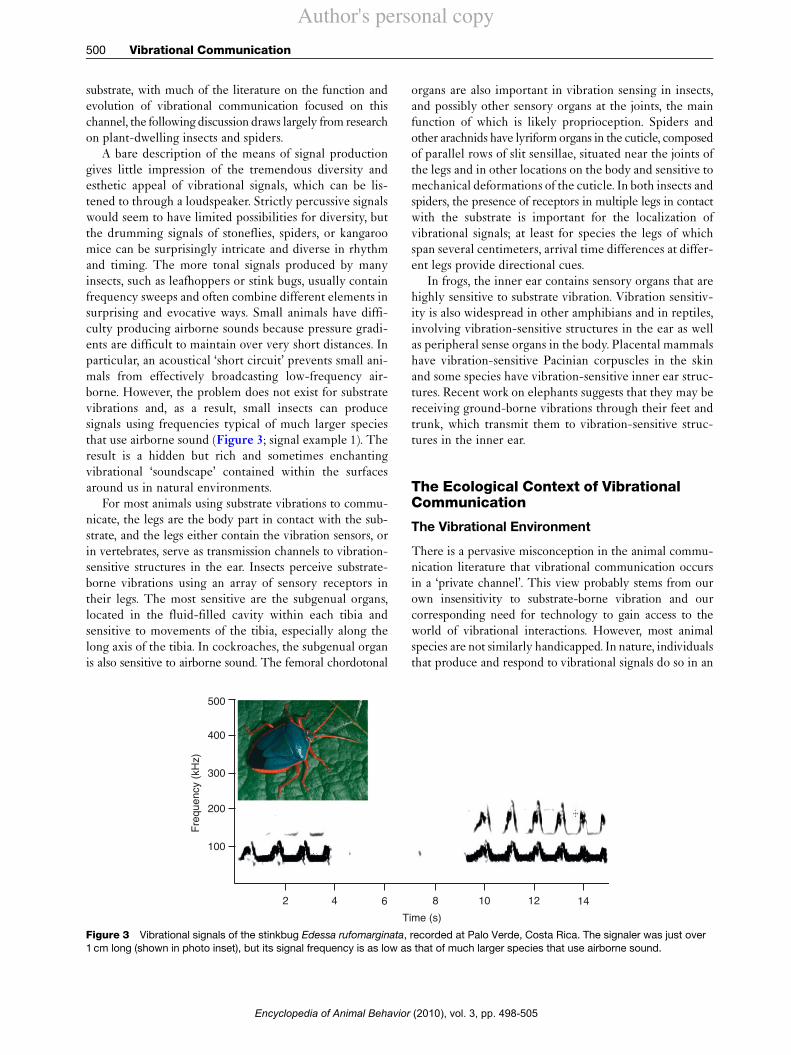
Vibrational communication is also widespread in ver-tebrates; again, the species known to communicate usingthis modality represent only the tip of the iceberg. Thereis increasing evidence for its importance in frogs and somereptiles, including the veiled chameleon (Figure 2).Vibrational communication is widespread in mammals,particularly in rodents, and more recently documentedin large species, including elephants.
Perception and Production ofVibrational Signals
The substrates used for vibrational communication varywith taxonomic group and, to some extent, with size. Inaddition, the nature of the substrate influences how signalspropagate and how they are perceived. Vibrationally signal-ing mammals typically use the ground as a communicationchannel (vibrations of the earth’s surface are referred to asseismic signals). In most cases, signals are produced by
498
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505
Author's personal copy
drumming with the feet or, for some subterranean spe-cies, tapping the head against the soil. In elephants,foot stomping and low-frequency vocal rumbles generateground-borne waves that travel long distances. Seismiccommunication also occurs in crabs, scorpions, and someinsects and spiders. However, most insects, many spiders,most cases in frogs, and one lizard (Figure 2) use livingplants as a communication channel. Plant tissues such asstems and leaves are excellent substrates through which totransmit signals because they have evolved to flex inresponse to mechanical disturbances such as wind, and asa consequence, will transmit even extremely low-amplitudesignals for a meter or more along stems. Almost any move-ment on the surface of a plant will generate a mechanicaldisturbance, and as a result, species that communicatethrough plants have a variety of ways of producing signals.These include striking the substrate with feet, head, abdo-men, or other body part, using direct muscle contractions tovibrate some part of the body (resulting in signals contain-ing harmonic series or relatively pure tones; see videoexample 1, which shows the abdomen of a signaling maletreehopper, Umbonia crassicornis), and/or using a frequencymultiplier such as a stridulatory file and scraper. For nonpercussive forms of signal production, vibrations producedthrough direct muscle contraction or specialized vibratingstructures are transmitted through the legs into the sub-strate. Many spiders transmit signals through webs, andsome insects produce signals on the water surface; thisarticle, however, deals onlywith vibrations traveling throughsolid substrates. Because living plants are such a widespread
Delphacidae
Substrate vibration
Airborne sound
Cixiidae
Dictyopharidae
Cicadellidae
Membracidae
Aetalionidae
Cicadidae
Tettigarctidae
Cercopidae
Aphrophoridae
Substrate vibrationAirborne sound
Figure 1 Phylogeny of the Cicadomorpha, showing the distribution of signaling modality among families. Communication by airbornesound in cicadas is a derived state with respect to substrate vibration (phylogeny drawn from Cryan JR (2005) Molecular phylogeny ofCicadomorpha (Insecta: Hemiptera: Cicadoidea, Cercopoidea, and Membracoidea): Adding evidence to the controversy. SystematicEntomology 30: 563–574).
Figure 2 A male veiled chameleon (Chameleo calyptratus).This species communicates using visual signals and plant-bornevibrational signals.
Vibrational Communication 499
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505
Author's personal copy
substrate, with much of the literature on the function andevolution of vibrational communication focused on thischannel, the following discussion draws largely from researchon plant-dwelling insects and spiders.
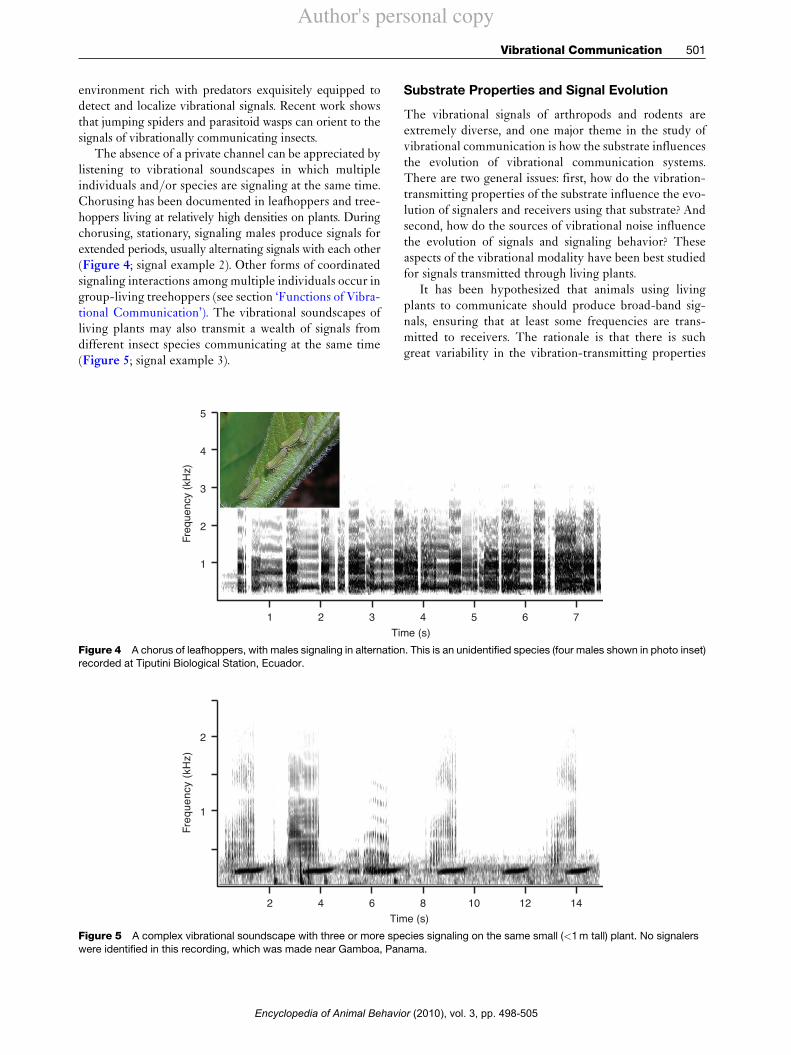
A bare description of the means of signal productiongives little impression of the tremendous diversity andesthetic appeal of vibrational signals, which can be lis-tened to through a loudspeaker. Strictly percussive signalswould seem to have limited possibilities for diversity, butthe drumming signals of stoneflies, spiders, or kangaroomice can be surprisingly intricate and diverse in rhythmand timing. The more tonal signals produced by manyinsects, such as leafhoppers or stink bugs, usually containfrequency sweeps and often combine different elements insurprising and evocative ways. Small animals have diffi-culty producing airborne sounds because pressure gradi-ents are difficult to maintain over very short distances. Inparticular, an acoustical ‘short circuit’ prevents small ani-mals from effectively broadcasting low-frequency air-borne. However, the problem does not exist for substratevibrations and, as a result, small insects can producesignals using frequencies typical of much larger speciesthat use airborne sound (Figure 3; signal example 1). Theresult is a hidden but rich and sometimes enchantingvibrational ‘soundscape’ contained within the surfacesaround us in natural environments.
For most animals using substrate vibrations to commu-nicate, the legs are the body part in contact with the sub-strate, and the legs either contain the vibration sensors, orin vertebrates, serve as transmission channels to vibration-sensitive structures in the ear. Insects perceive substrate-borne vibrations using an array of sensory receptors intheir legs. The most sensitive are the subgenual organs,located in the fluid-filled cavity within each tibia andsensitive to movements of the tibia, especially along thelong axis of the tibia. In cockroaches, the subgenual organis also sensitive to airborne sound. The femoral chordotonal
organs are also important in vibration sensing in insects,and possibly other sensory organs at the joints, the mainfunction of which is likely proprioception. Spiders andother arachnids have lyriform organs in the cuticle, composedof parallel rows of slit sensillae, situated near the joints ofthe legs and in other locations on the body and sensitive tomechanical deformations of the cuticle. In both insects andspiders, the presence of receptors in multiple legs in contactwith the substrate is important for the localization ofvibrational signals; at least for species the legs of whichspan several centimeters, arrival time differences at differ-ent legs provide directional cues.
In frogs, the inner ear contains sensory organs that arehighly sensitive to substrate vibration. Vibration sensitiv-ity is also widespread in other amphibians and in reptiles,involving vibration-sensitive structures in the ear as wellas peripheral sense organs in the body. Placental mammalshave vibration-sensitive Pacinian corpuscles in the skinand some species have vibration-sensitive inner ear struc-tures. Recent work on elephants suggests that they may bereceiving ground-borne vibrations through their feet andtrunk, which transmit them to vibration-sensitive struc-tures in the inner ear.
The Ecological Context of VibrationalCommunication
The Vibrational Environment
There is a pervasive misconception in the animal commu-nication literature that vibrational communication occursin a ‘private channel’. This view probably stems from ourown insensitivity to substrate-borne vibration and ourcorresponding need for technology to gain access to theworld of vibrational interactions. However, most animalspecies are not similarly handicapped. In nature, individualsthat produce and respond to vibrational signals do so in an
Time (s)
Freq
uenc
y (k
Hz)
100
2 4 6 8 10 12 14
200
300
400
500
Figure 3 Vibrational signals of the stinkbug Edessa rufomarginata, recorded at Palo Verde, Costa Rica. The signaler was just over1 cm long (shown in photo inset), but its signal frequency is as low as that of much larger species that use airborne sound.
500 Vibrational Communication
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505
Author's personal copy
environment rich with predators exquisitely equipped todetect and localize vibrational signals. Recent work showsthat jumping spiders and parasitoid wasps can orient to thesignals of vibrationally communicating insects.
The absence of a private channel can be appreciated bylistening to vibrational soundscapes in which multipleindividuals and/or species are signaling at the same time.Chorusing has been documented in leafhoppers and tree-hoppers living at relatively high densities on plants. Duringchorusing, stationary, signaling males produce signals forextended periods, usually alternating signals with each other(Figure 4; signal example 2). Other forms of coordinatedsignaling interactions among multiple individuals occur ingroup-living treehoppers (see section ‘Functions of Vibra-tional Communication’). The vibrational soundscapes ofliving plants may also transmit a wealth of signals fromdifferent insect species communicating at the same time(Figure 5; signal example 3).
Substrate Properties and Signal Evolution
The vibrational signals of arthropods and rodents areextremely diverse, and one major theme in the study ofvibrational communication is how the substrate influencesthe evolution of vibrational communication systems.There are two general issues: first, how do the vibration-transmitting properties of the substrate influence the evo-lution of signalers and receivers using that substrate? Andsecond, how do the sources of vibrational noise influencethe evolution of signals and signaling behavior? Theseaspects of the vibrational modality have been best studiedfor signals transmitted through living plants.
It has been hypothesized that animals using livingplants to communicate should produce broad-band sig-nals, ensuring that at least some frequencies are trans-mitted to receivers. The rationale is that there is suchgreat variability in the vibration-transmitting properties
Time (s)1 2 3 4 5 6 7
5
4
3
2
1
Freq
uenc
y (k
Hz)
Figure 4 A chorus of leafhoppers, with males signaling in alternation. This is an unidentified species (four males shown in photo inset)recorded at Tiputini Biological Station, Ecuador.
2
1
2 4 6Time (s)
8 10 12 14
Freq
uenc
y (k
Hz)
Figure 5 A complex vibrational soundscape with three or more species signaling on the same small (<1m tall) plant. No signalerswere identified in this recording, which was made near Gamboa, Panama.
Vibrational Communication 501
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505

Author's personal copy
of living plants – the frequency spectrum of a signal variessubstantially, and often unpredictably, with distance fromthe signaler – that specialization on a single, optimalfrequency is unlikely to function well. This hypothesishas been influential but has never been formally tested,although studies of wolf spiders show that their broad-band, percussive signals are successful in courtship onmultiple substrates. It is clear, however, that it does notapply to all species, because although many vibrationallycommunicating species do produce broad-band signals,many others produce tightly tuned signals, with mostof their energy concentrated in a very narrow range offrequencies.
There are relatively few studies of vibration trans-mission in plants, and fewer still that have characterizedmore than one or two examples of a given host plant.For most studies, there is little information about whichplant parts are used by signalers in nature, or about theproperties of more than one host species for organismsthat use many different host species for communication.Study of hosts used by the cosmopolitan green stink bugreveals that several of the plants it uses transmit vibra-tions in the 100-Hz range most effectively, suggesting thatsignals of pentatomid bugs are adapted to this ‘frequencywindow.’ In contrast, for the host plants used by somesignaling treehoppers with narrowly tuned signals, theoptimal frequency varies both between host species andbetween modules (such as stems vs. leaves) within theplant. For two closely related treehoppers using differentplant species and different modules (woody stems vs.leaf petioles), signal frequency has diverged to matchthe contrasting vibration-transmission properties of theirtypical substrate.
Sources of Noise in the Vibrational Channel
Wind is the major abiotic source of noise in living plants,and probably in other substrates such as leaf litter. Asanyone who has tried to record plant-borne vibrationsoutdoors can attest, the air is seldom completely still. Howdo organisms communicating with plant-borne vibrationsdeal with the pervasive noise generated by wind? Wind hasthe potential to be a major influence on the evolution ofvibrational communication systems, influencing the evolu-tion of signal form, signaling behavior, or both.
The most obvious way in which wind could influencethe evolution of vibrational signals is by selecting for theuse of frequencies that avoid masking by those present inthe wind. In living plants, most of the energy in wind-generated noise is in low frequencies, less than 100Hz.Lower wind velocities cause swaying and low-frequencyvibrations, while higher velocities cause leaves to flutterand stems to strike each other, exciting a broaderspectrum. There are at least two ways in which windmight exert an influence on signal evolution. First,
wind-induced noise might be important for some speciesand unimportant for others (e.g., species signaling onplants in sheltered locations). Second, even if wind isimportant for most species, the properties of wind-induced noise might differ from plant to plant, especiallyin the amount of energy present in different frequencies.Wind noise has only been characterized in a few plants; inone case, two structurally different plants were found tohave different frequency spectra of wind-induced vibra-tions, while in another, two structurally similar species ofwoody plants had very similar profiles. At present, it is notpossible to say whether wind imposes similar selection onvibrational signals (e.g., selecting for higher frequenciesthat avoid those present in wind noise) or whether it mayfavor the use of different signals for efficient communica-tion on different plants.
Whether or not the presence of wind influences theevolution of signal frequency, as a major source of noise inthe vibrational channel, it is very likely to influence thesignaling behavior of organisms. One way for organisms todeal with wind noise is spatial avoidance or communica-tion in relatively wind-free environments. Such environ-ments as the forest interior are characterized by lowerwind speeds; however, for plant-feeders, the most nutri-tionally valuable plant tissues are usually those in high-light environments, which are, of course, less sheltered. Asa strategy for avoiding wind noise, then, spatial avoidanceof wind may not be available to many species.
Temporal avoidance of wind can occur at two scales.First, often there are predictable daily patterns of rise andfall in wind velocity. Males of at least one treehopperspecies do signal more often during morning and eveninghours when wind speeds are lower in its forest edgehabitat, but whether this pattern has evolved specificallyin response to wind is unclear. Second, wind speed is oftenquite variable over a scale of minutes or seconds, andorganisms could use gap detection to signal during shortwind-free periods. For one treehopper, laboratory experi-ments show that males signal during brief wind-free gaps.This avoidance of signaling during wind gusts is likelyimportant for communication; females responded less tomale signals masked by wind, especially when the signalswere low in amplitude.
In addition to wind, rain is a significant source ofvibrations for living plants, leaf litter, and probably forany exposed substrate. In plant leaves, the impact ofa raindrop causes an initial high-amplitude, broad-band pop, with a decreasing tail of lower frequencies.A hard rainfall likely precludes communication, as doesheavy wind. But in environments subject to frequentlow-intensity rainfall, organisms likely have adaptationsthat enhance communication.
Airborne sounds are another source of noise inthe vibrational channel. Leaves vibrate in responseto airborne sound, and recordings made in natural
502 Vibrational Communication
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505

Author's personal copy
environments include not only the signals of vibrationallycommunicating species but also the signals of nearbyspecies signaling with airborne sound, such as cicadasand birds. The significance of these induced vibrationsfor communication is unknown, but in some cases, theymay provide useful information; for example, a wolf spi-der was shown to alter its behavior in response to inducedvibrations from the songs of potential avian predators.
Finally, a source of noise for one species may be animportant cue for another. The vibrations produced bywind blowing through mounds of grass are potential cuesfor foraging golden moles in their sand dune habitat,and the ground-borne rumbling of distant thunder maybe used by elephants to locate areas where rain hasrecently fallen.
Functions of Vibrational Communication
Sexual selection is the major engine of diversity in theevolution of animal signals. The vibrational signals ofinsects and spiders provide a remarkable panorama ofthe historical action of sexual selection, given the ancientnature of this communication channel, and the hundredsof thousands of living species that use it.
Evidence of the importance of sexual selection in thediversification of vibrational signals and in speciation comesprimarily from the study of insects, especially lacewings,stoneflies, and the Auchennorrhyncha (planthoppers, leaf-hoppers, and treehoppers). All of these groups containsibling species, close relatives that are phenotypically simi-lar except in mating signals and preferences. In the lacew-ings, for example, systematic study has revealed unexpectedacoustic diversity within what were thought to be singlespecies, and experimental research has implicated sexualselection as the evolutionary force driving both song diver-sification and species formation. Important work on sexualselection is also being conducted in spiders, where vibra-tional signals are often components of multimodal matingdisplays that include motion, color, and substrate vibra-tion. These multimodal signals are especially diverse injumping spiders and wolf spiders.
Vibrational signals are used not only in mating but alsoin communication among members of social groups. Incontrast to sexually selected signals, which are often com-plex and rapidly evolving, social signals are usually rathersimple, with their structure evolutionarily conservedamong closely related species. In the eusocial insects, vibra-tion is important in alarm communication in termites andsome ants, and in recruitment to profitable food resourcesin other ants. In noneusocial insects, such within-groupcommunication has been best studied in group-livingimmatures of sawflies and treehoppers. Vibrational signal-ing among group members is important for remaining in agroup, finding food, and, in species with maternal care,
alerting the mother to the arrival of a predator. Predatoralarm signaling also occurs in some rodents, although here,it is the parents that signal to alert the offspring.
Vibrational signaling is important for other social andecological relationships as well. Some caterpillars defendleaf territories using a repertoire of vibrational signals. Forlycaenid and riodinid caterpillars, and for some treehop-pers, vibrational signals facilitate the attraction of antmutualists or ant detection of potential predators.
Recording and Playback Methods forStudying Vibrational Communication
What follows is a discussion of methods for vibrationrecording and playback that have been used extensivelyfor vibrations transmitted through living plants, but muchwill be relevant to other substrates.
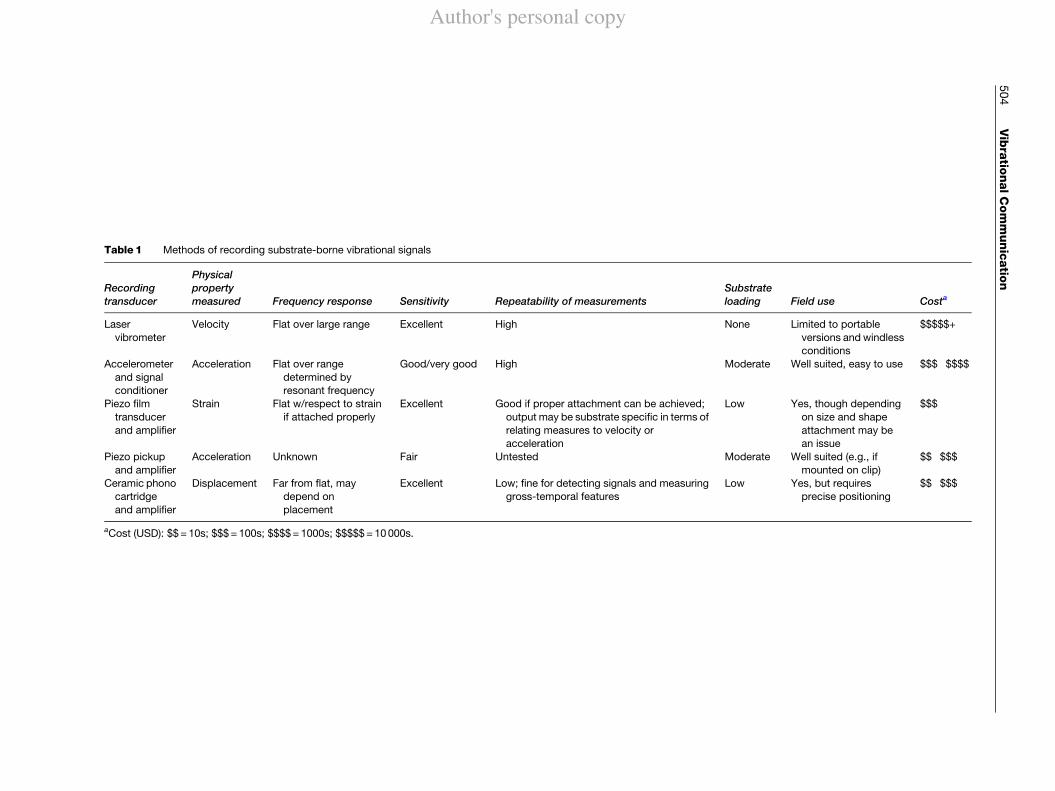
There are many applications for vibration measure-ment in engineering, manufacturing and testing, and themusic industry. As a result, there are many options avail-able for detecting and recording vibrational signals. Thesemethods range widely in cost and suitable applications.The gold standard is the laser vibrometer, which usesDoppler shifts in reflected laser light to measure thevelocity of a moving surface (analogous to the use ofradar for measuring vehicle speed). The day-to-day work-horse is the accelerometer, which is attached to the vibrat-ing substrate with wax or adhesive and has an outputproportional to the acceleration component of the signal.Newer technology includes piezoelectric film, which hashigh sensitivity when used with appropriate amplifiers,but has not been widely used. Other methods arise fromthe music industry, including phonograph cartridges andpiezo pickups; these are often convenient, of low-cost, andsensitive, but have disadvantages for some research pur-poses. Table 1 provides a comparison of some commonmethods of recording vibrational signals.
One general issue is the extent to which the measuringmethod alters the properties of the substrate, and as aresult of the transmitted signal. This ‘mass loading’ issuevaries in importance depending on the size of the sub-strate and the importance of making precise measure-ments of signal properties and/or of substrate effects onthose properties.
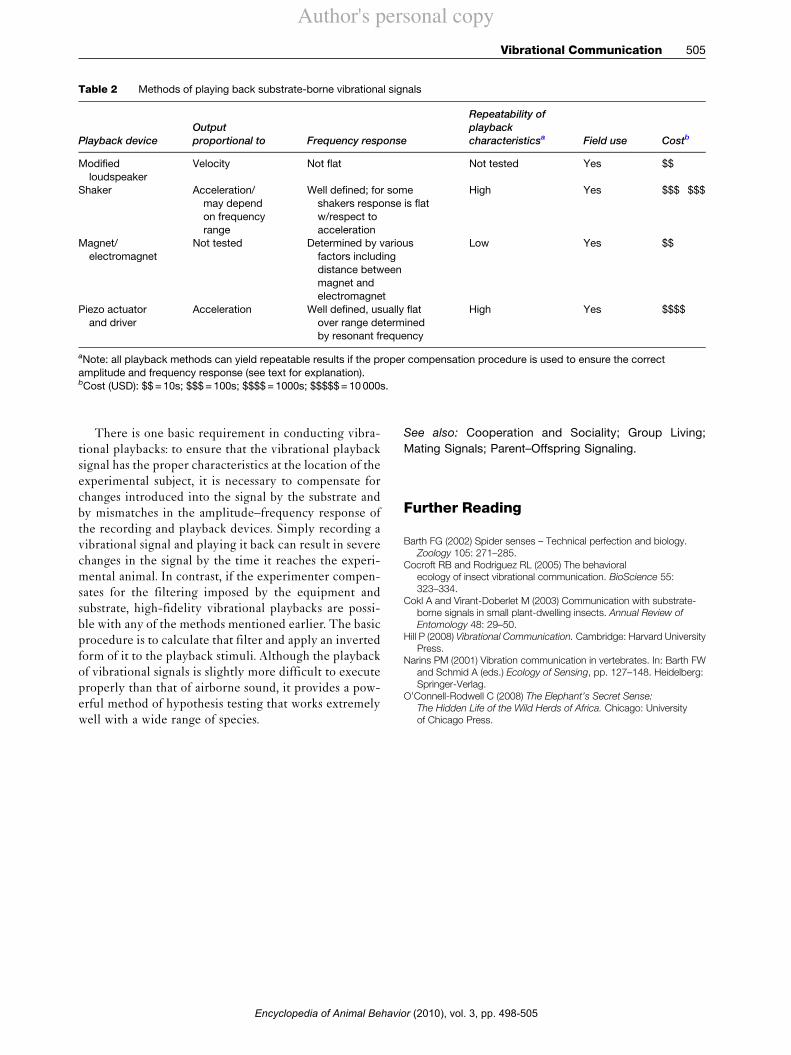
Vibrational playbacks are possible using a wide varietyof methods, compared in Table 2. The most commonplayback devices include a shaker, specifically designedfor coupling a vibration to a substrate; a magnet attachedto the substrate and driven by sending the signal throughan electromagnet placed in close proximity; and modifica-tions of other technology, such as removing the membranefrom a loudspeaker and coupling the movement of the coilto the substrate using a pin. Any of these methods canwork well, but their characteristics vary widely.
Vibrational Communication 503
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505
Author's personal copy
Table1 Methods of recording substrate-borne vibrational signals
Recordingtransducer
Physicalpropertymeasured Frequency response Sensitivity Repeatability of measurements
Substrateloading Field use Costa
Laservibrometer
Velocity Flat over large range Excellent High None Limited to portableversions and windlessconditions
$$$$$+
Accelerometerand signalconditioner
Acceleration Flat over rangedetermined byresonant frequency
Good/very good High Moderate Well suited, easy to use $$$!$$$$
Piezo filmtransducerand amplifier
Strain Flat w/respect to strainif attached properly
Excellent Good if proper attachment can be achieved;output may be substrate specific in terms ofrelating measures to velocity oracceleration
Low Yes, though dependingon size and shapeattachment may bean issue
$$$
Piezo pickupand amplifier
Acceleration Unknown Fair Untested Moderate Well suited (e.g., ifmounted on clip)
$$!$$$
Ceramic phonocartridgeand amplifier
Displacement Far from flat, maydepend onplacement
Excellent Low; fine for detecting signals and measuringgross-temporal features
Low Yes, but requiresprecise positioning
$$!$$$
aCost (USD): $$=10s; $$$=100s; $$$$=1000s; $$$$$=10000s.
504
Vibra
tionalCommunicatio
n
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505
Author's personal copy
There is one basic requirement in conducting vibra-tional playbacks: to ensure that the vibrational playbacksignal has the proper characteristics at the location of theexperimental subject, it is necessary to compensate forchanges introduced into the signal by the substrate andby mismatches in the amplitude–frequency response ofthe recording and playback devices. Simply recording avibrational signal and playing it back can result in severechanges in the signal by the time it reaches the experi-mental animal. In contrast, if the experimenter compen-sates for the filtering imposed by the equipment andsubstrate, high-fidelity vibrational playbacks are possi-ble with any of the methods mentioned earlier. The basicprocedure is to calculate that filter and apply an invertedform of it to the playback stimuli. Although the playbackof vibrational signals is slightly more difficult to executeproperly than that of airborne sound, it provides a pow-erful method of hypothesis testing that works extremelywell with a wide range of species.
See also: Cooperation and Sociality; Group Living;
Mating Signals; Parent–Offspring Signaling.
Further Reading
Barth FG (2002) Spider senses – Technical perfection and biology.Zoology 105: 271–285.
Cocroft RB and Rodriguez RL (2005) The behavioralecology of insect vibrational communication. BioScience 55:323–334.
Cokl A and Virant-Doberlet M (2003) Communication with substrate-borne signals in small plant-dwelling insects. Annual Review ofEntomology 48: 29–50.
Hill P (2008) Vibrational Communication. Cambridge: Harvard UniversityPress.
Narins PM (2001) Vibration communication in vertebrates. In: Barth FWand Schmid A (eds.) Ecology of Sensing, pp. 127–148. Heidelberg:Springer-Verlag.
O’Connell-Rodwell C (2008) The Elephant’s Secret Sense:The Hidden Life of the Wild Herds of Africa. Chicago: Universityof Chicago Press.
Table 2 Methods of playing back substrate-borne vibrational signals
Playback deviceOutputproportional to Frequency response
Repeatability ofplaybackcharacteristicsa Field use Costb
Modifiedloudspeaker
Velocity Not flat Not tested Yes $$
Shaker Acceleration/may dependon frequencyrange
Well defined; for someshakers response is flatw/respect toacceleration
High Yes $$$!$$$
Magnet/electromagnet
Not tested Determined by variousfactors includingdistance betweenmagnet andelectromagnet
Low Yes $$
Piezo actuatorand driver
Acceleration Well defined, usually flatover range determinedby resonant frequency
High Yes $$$$
aNote: all playback methods can yield repeatable results if the proper compensation procedure is used to ensure the correctamplitude and frequency response (see text for explanation).bCost (USD): $$=10s; $$$=100s; $$$$=1000s; $$$$$=10 000s.
Vibrational Communication 505
Encyclopedia of Animal Behavior (2010), vol. 3, pp. 498-505





























