Embed Size (px)
Citation preview

The Value of Ontogenetic Data in Reconstructing Protozoan PhylogeniesAuthor(s): John O. CorlissSource: Transactions of the American Microscopical Society, Vol. 87, No. 1 (Jan., 1968), pp. 1-20Published by: Wiley on behalf of American Microscopical SocietyStable URL: http://www.jstor.org/stable/3224333 .
Accessed: 28/06/2014 11:57
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Wiley and American Microscopical Society are collaborating with JSTOR to digitize, preserve and extendaccess to Transactions of the American Microscopical Society.
http://www.jstor.org
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

TRANSACTIONS
of the
American Microscopical Society VOL. 87 JANUARY 1968 NO. 1
THE VALUE OF ONTOGENETIC DATA IN RECONSTRUCTING PROTOZOAN PHYLOGENIES'
JOHN O. CORLISS
Department of Biological Sciences, University of Illinois at Chicago Circle, Chicago
ABSTRACT: Ontogenetic information, properly considered, can play a role in recon- struction of metazoan phylogenies; and it is proposed that such data can also be of value in postulating phylogenetic interrelationships among certain groups within the phylum Protozoa. One must agree at the outset that protozoa have ontogenies. Several other assumptions are employed: that certain present-day forms have gone essentially unchanged for many aeons; that there is positive correspondence between degree of similarity and recency of common ancestry; that the "simple" is commonly ancestral to the "complex." As a source of examples of possible ontogenetic recapitula- tion of phylogeny, the ciliate protozoa are chosen because of the convenience of the infraciliature in comparative studies of morphogenetic phenomena occurring during ciliate ontogenies (life cycles). Selected specific illustrations involve several groups of the Ciliophora at the subclass or ordinal level: the astomes, the trichostomes, the chonotrichs, the suctorians, the peritrichs, the holotrichs (and spirotrichs), and the hymenostomes (and scuticociliates). Patterns of stomatogenesis may often supply valuable clues in determination of possible phylogenetic interrelationships. Dangers and pitfalls to the use of ontogeny in reconstructing protozoan phylogenies are many (e.g., recognition of homology, selection, polyphyly, parallel and convergent evolution, adaptation to similar environment) and thus extreme caution must be employed; but it is suggested that protozoologists, so warned, might well make worthwhile progress in this field of research.
Although Haeckel's (1866, 1875) now century-old aphorism, "ontogeny briefly recapitulates phylogeny," represents a theory replete with inadequacies and mis- leading implications which have been thoroughly and repeatedly pointed out by many authors during the past quarter of a century (and even much earlier: see de Beer, 1951, for historical review), the fact remains that evidences from embryology, considered with proper caution, can play an important, legitimate role in reconstruction of metazoan phylogenies. Von Baer's (1828) pre-evolu- tionary "fourth law," which was postulated some 40 years before the time of Haeckel's hypothesis and which stated that the young stages in the development of an animal resemble the young stages, only, of other animals lower on the scale, has long met with acceptance as a more appropriate interpretation of "recapitula- tion" in its broadest sense. Nevertheless, some students of evolutionary or phylo- genetic systematics today seem wary of using such a postulate in construction, for example, of schemes of classification (e.g., see remarks in Mayr, 1965, and Simpson, 1961). Yet, as Libbie Hyman (19,40) has pointed out, with character-
Past-President's address, delivered at the 83rd Annual Meeting of the American Micro- scopical Society, held at the University of Arizona, Tucson, August 1967. Support of National Science Foundation grants GB-2800 and GB-7013X is gratefully acknowledged. TRANS. AMER. MICROSC. Soc. 87(1): 1-20. 1968.
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

2 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
istic straightforward judiciousness, "ancestral resemblance during ontogeny is a general biological principle." And a well-known student of certain vertebrate groups has recently stressed the value of ontogenetical studies in systematics and evolution (Orton, 1955).
Hadzi (1952), in Yugoslavia, and von Gelei (1950), in Hungary, have sug- gested, in a general way, that the principles of "phylembryogenesis"-Sewertzoff's (1931) term for the relation between ontogeny and phylogeny-might be ap- plied to the "protista"; and, in very recent times, protozoologists in Poland and Russia have supported usage of the "biogenetic law" in studies on ciliate phylog- eny (e.g., see Cheissin & Poljansky, 1963; Poljansky & Cheissin, 1965; Raabe, 1958, 1964a; and references therein). In this country, Furgason (1940), over 25 years ago, described the presence of a simple, tripartite adoral zone of membranelles in the oral cavity of a species of holotrich ciliate (Tetrahymena) and promulgated the important postulate that such an inconspicuous set of structures might well represent the forerunner of (and thus, phylogenetically, the primitive state of) the well-known, prominent multipartite AZM characteristic of most spirotrich ciliates.
It is, however, to the French schools of Chatton and Faure-Fremiet, partic- ularly the latter's, that we must give credit for heuristic attempts to broaden our recognition of the potential significance of morphological and morphogenetic similarities among organellar systems in protozoa (especially ciliates) and to focus fresh attention on the possibility that recapitulation of ancestral stages occurs in the ontogeny or morphogenetic life cycle of various species (e.g., see Chatton, 1942; Chatton & Lwoff, 1935, 1936, 1949, 1950; Chatton & Perard, 1921; Chatton & Seguela, 1940; Dragesco, 1960, 1963; Faure-Fremiet, 1950, 1953, 1959, 1961a, b, 1962, 1963, 1965, 1968; Guilcher, 1951; Mugard, 1949; Noirot-Timothee, 1960; Noirot-Timothee & Lom, 1965; Roque, 1961; Tuffrau, 1952, 1960; Villeneuve- Brachon, 1940). It is unnecessary to point out that the organisms must be sub- mitted to appropriately refined cytological study.
In part from the proposed phylogenetic interrelationships resulting from many studies, it has, in recent years, become possible to construct revised systems of classification at the higher taxonomic levels within the phylum Protozoa which allegedly represent considerable improvement over older schemes (e.g., see Corliss, 1963a, 1967b, 1968; Honigberg et al., 1964; Levine, 1966; and the many references cited in those publications; also see Appendix II).
My own interests in the overall subject developed during my graduate student days at New York University, 20 years ago. I was particularly stimulated, at that time, by the works of such experimental embryologists as Ross Granville Harrison, Hans Spemann, Paul Weiss, and J. Runnstrom; of such evolutionary biologists as Gavin de Beer, W. Garstang, Julian Huxley, George Gaylord Simpson, and Ernst Mayr; and of such incomparable ciliate protozoologists as Chatton and Lwoff, Faure-Fremiet, and the latter's very active corps of doctoral students. A
year's postdoctoral fellowship in Professor Faure-Fremiet's laboratory in Paris and, subsequently, two years on the staff at Yale University offered environments which served to intensify my desire to study ciliate morphogenesis as a kind of "embryology at the protozoan level" (Corliss, 1953). And I continue, today, to believe that such investigation is profitable from the point of view of an approach to an understanding of phylogenetic interrelationships among protozoan orga- nisms, though we have a long, long way to go.
PRELIMINARY CONSIDERATIONS
Before discussing selected cases, some old and others very new, in illustration, or defense, of my theme-that ontogenetic data, critically collected and analyzed and cautiously interpreted, can be of genuine value in reconstruction of protozoan
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 3
phylogenies-I should like to dispose of some preliminary matters as briefly as possible.
1. Definition of terms.-I am in agreement with the generally accepted and oft-quoted modern concepts surrounding such terminology as "ontogeny," "phylog- eny," "morphogenesis," "differentiation," "homology" (the amount of literature on this word is fantastic!), "convergence," "parallel evolution," "monophyly," "polyphyly," and "natural selection." Thus no definitions need to be supplied here.2 I do not agree with Hadzi (1952), incidentally, that protozoa exhibit no ontogeny, although I understand his point of view and must admit that the multiple phases or generations in the life cycles of many protozoan species, par- ticularly non-ciliates, complicate the picture.
2. Differences between protozoa and metazoa.-I decry the generally con- tinued emphasis on differences, principally because of the single-celledness of the former, between protozoa and metazoa, primarily because I do not accept the implication that the protozoa are clearly "hopeless" organisms with respect to proper study of their phylogeny and systematics. (Geneticists of the protozoa have run into the same problem.) Ignoring unicellular organisms when con- sidering basic tenets of phylogeny and systematics (or genetics) is not only unfair but may be unfortunate, for it is possible that solutions to some such problems being studied by students of the protozoa could profitably be applied to parallel situations existing for certain metazoan (or even metaphytan) groups of orga- nisms. Indeed, in spite of the protozoologists's (necessary) preoccupation, gen- erally speaking, with the alpha stage of taxonomy, he actually may be far ahead of-or may be able to apply far more sophisticated approaches to some problems than-biologists working with many animal and plant assemblages which remain outside the pale of the few "ideal," well-known, much-studied groups of organisms.
3. Necessary assumptions.-In view of the admittedly discouraging and nearly insurmountable disadvantages to the successful study of protozoan phylog- enies (widespread lack of a fossil record, typically microscopic size, subcellular organization, cosmopolitan nature of many species, frequent absence of sexuality: see Corliss, 1959a, 1960), certain assumptions-often also applied to problems with various metazoan groups-must be allowed the student of protozoan evolu- tion. The primary assumption, since, with rare exception, our whole system must be based on similarities and differences found only in extant forms, is that certain present-day forms have gone essentially unchanged for many aeons. Thus the protozoologist may consider that some of the forms with which he is dealing (continue to) resemble primitive species more closely than do others. Therefore the factor of time is, indirectly, not neglected. But note that no present-day members of a species are claimed to have descended from present-day members of any other species or group, for this would be a most unlikely evolutionary origin for the majority of species comprising any diverse group of extant organisms.
The basic evolutionary principle that there is positive correspondence between degree of similarity of organisms and their recency of common ancestry is also invoked. Even the admittedly highly dangerous assumption that the simple is ancestral to the elaborate or complex is openly employed, but only in a general and cautious way and keeping clearly in mind the many actual or potential exceptions to justifiable use of the generalization. One must be well aware, for example, of the possible effects of selection, adaptations to particular environ- ments, etc., etc.
2 A single excellent source of definitions, explanations, and discussion of many of these terms is Simpson's (1961) book on principles of animal classification. There is no reason to become entangled here-with respect to some of this terminology-in the disputations, often highly semantic in nature, of the purists.
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

4 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
Finally, the "when," "where," "why," "how," and "how fast" of the evolutionary changes which must have taken place to give us our present diversity of forms need not, in my opinion, be considered of primary importance in our initial ap- proaches to protozoan phylogeny, evolution, and systematics. Such questions, as Wagner (1968) has pointed out in his stimulating discussion of constructing classifications for plant material, are subsidiary to the questions of how much change has occurred, what characters are involved, in what directions the changes have proceeded, and in what order they have occurred.
SOURCES OF EXAMPLES
Groups within the several major taxa of protozoa could provide examples of alleged phylogenetic sequences, the elucidation of which might be aided by knowledge of ontogenetic data for the various forms involved. Even fossil evi- dence might be invoked in such cases as certain lines of the foraminiferidans, coccolithophorids, radiolarians, or testaceous amoebae. A striking example of "ontogenetic recapitulation of phylogeny" can be assumed with respect to the transformation undergone by the dimorphic amoeboflagellates, though the phe- nomenon is too general to help in reconstruction of any specific phylogenetic series.
Protozoa with elaborate cysts or other pellicular or external differentiations showing considerable diversity among groups of species lend themselves to the kind of study which I am suggesting: examples could range from myxosporideans, with their sculptured spore-valves, to the gregarine sporozoa, with their diversi- fied gametocyst membranes. The newly discovered ultrastructural similarities in forms comprising the several stages found in both malarial and coccidian life cycles also could be cited. The thecae of many phytoflagellates, particularly the dinoflagellates, in which the arrangements of the thecal plates are superficially reminiscent of the patterns of bones in the developing vertebrate skull, might be similarly investigated, too.
The mastigont system of the flagellate protozoa, particularly as found in the so-called "higher" zooflagellates, represents an ideal system for comparative investigation which might cast light on phylogenetic interrelationships at various taxonomic levels within the whole assemblage. The (primarily) intracytoplasmic structures or organelles of the system can be studied with regard to variation in size, shape, number, and position, and even chemical composition, as well as presence or absence. Kirby (e.g., 1944, 1949, 1950) and, more recently, Honigberg (e.g., 1963a,b, 1967b) have undertaken investigation of such fascinating and challenging problems presented by these zooflagellates; much remains to be done in this fertile area of inquiry, however.
But, in many ways, the ciliate protozoa, with their high degree of pellicular or cortical development and their diversity in external ciliature and in patterns of ciliature, would seem to offer the best source of examples for study of possible phylogenetic interrelationships. I would choose the ciliates, too, from a personal point of view, because I feel much more at home with them than with the non- ciliate protozoan groups. Also, with rare exception, the Ciliophora have already been so studied more extensively than any other protozoa to date, although, admittedly, few of the investigations have represented the indepth studies which would be most desirable to have carried out.
Before citing, very briefly and with deliberate omission of the associated de- tailed data, examples from among the Ciliophora, it would be pertinent to treat, also most briefly, the underlying problem in all morphological-phylogenetic con- siderations; namely, what features can be thought of as truly "basic," "fundamen- tal," "unspecialized," or "most primitive"-and with what justification? Are there certain characteristics of a conservative enough nature among ciliates to warrant
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 5
their treatment as properties of genuine evolutionary value? Can a priori weight- ing of a few arbitrarily chosen characteristics be avoided? That is, do we cur- rently know enough in the field of ciliate systematics to be able to invoke a posteriori reasoning (see Mayr, 1964) with respect to taxonomic characters?
I think that the problem is, in the last analysis, no more difficult for the protozoa than for the metazoa or metaphyta. Subjectivity and intuition may indeed be involved in making evolutionary judgments in general; or in reaching conclusions with regard to choice or weighting of characteristics; but I do not consider such action, for any kinds of organisms, as illegitimate or unscientific.
The infraciliature has not been established and recognized beyond doubt as the most stable feature of the ciliate protozoa (Corliss, 1956, 1959b, 1961), thanks primarily to the development of techniques for demonstrating it unequivocally (including, today, electron microscopy, as well as silver and other cytological methods at the light microscopic level: Corliss, 1953c, 1963b; Corliss et al., 1968; Dragesco, 1962; Grimstone, 1961, 1966; Honigberg, 1967a; Pitelka, 1963; Tuffrau, 1967; Uhlig, 1967). The patterns of the infraciliary systems (kinetosomes and associated fibers) and of the external ciliature often arising from the infraciliature lend themselves beautifully to comparative study, as has been known for some time.
What is even more important, however, are the changes in such patterns which occur during the course of the morphogenetic phenomena which are normal parts of the full life cycle of any ciliate. In other words, tracing out the dynamics of the pattern during ontogeny can provide data potentially of great significance in determination of the organism's phylogenetic affinities. Revelation of what happens to the somatic infraciliature during fission or encystment or to the buccal infraciliature during new mouth formation (stomatogenesis) is thus of great importance to the conscientious comparative phylogeneticist.
Fundamental biological properties or characteristics of all or many cells and organisms-such as polarity, axes of symmetry, modes of fission, types of stomato- genesis, regeneration, cyst formation, polymorphism, allometric growth, and de- and redifferentiation-come under scrutiny in the kinds of comparative investigations undertaken by the thorough phylogeneticist referred to above. At all times, of course, the eager investigator must temper his enthusiasm and excitement-and control his possibly natural inclination or desire to promulgate new hypotheses-by awareness of the age-old vexatious problems of parallel evolution, convergence, polyphyly, independent origin of similar processes or characteristics, false homologies, irretrievable loss of intermediate forms or "missing links," numerous inexplicable adaptations, differential selection factors, variation in rates of evolution, etc., etc. Above all, perhaps, one must keep in mind that all currently recognized groups of the ciliates have surely enjoyed a very long evolutionary history of their own (Corliss, 1956, 1961)-thus, strictly speaking, present-day members of no extant taxon are probably directly ancestral to any other taxon (as stressed earlier: see p. 3).
SPECIFIC EXAMPLES
To return to the main topic of this paper, seven examples, though presented very briefly and with little comment, may suffice to serve as illustration of the value of knowledge of ontogeny in attempts to reconstruct protozoan phylogenies. Nearly all of these are concerned with evolutionary affinities at the ordinal level within the subphylum Ciliophora: recall that currently some fifteen orders are generally recognized in this major subdivision of the Protozoa embracing some 6,000 or so known species (Corliss, 1956, 1959b, 1961; and see Appendices I and II). I am not going to use as examples evolutionary sequences within sub- orders or families. Allow me, nonetheless, to point out that beautiful cases do
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

6 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
exist at such levels; the literature contains many appropriate illustrations: for example, see such papers as Chatton & Lwoff (1935, 1949, 1950), Faure-Fremiet (1959, 1961a,b, 1963), Noirot-Timothee (1960), and de Puytorac (1954, 1963), to cite selected outstanding works.3
1. The astomes were often formerly considered as "primitive" ciliates because of the single fact that they are mouthless. Some erstwhile authorities (e.g., Calkins, 1933) even lumped them together with the flagellate opalinids, protozoa strikingly different in every other way, for the same single reason of mouthless- ness. Modern studies of the whole life cycle of astomes, however, particularly as carried out by de Puytorac (e.g., 1954, 1959, 1963), have revealed basic infraciliary similarities to other groups, especially to certain members of the order of thigmotrichs. The absence (= loss?) of a mouth is rightly held to be a secondary character related to adaptation to a specialized endoparasitic en- vironment and thus of limited evolutionary or systematic value (Corliss, 1956, 1961). The pattern of the infraciliature during ontogeny is considered a much more reliable source of data for postulating possible affinities.
2. The taxonomic position or evolutionary status of the trichostomes has long baffled ciliate protozoologists.4 Although many problems still remain unsolved, already classical work such as Tuffrau's (1952) study of the family Colpodidae (see also Stout, 1960) reveals that the rather complex form and cortical morphology of the mature organism result from allometric growth in the individuals produced by binary fission and that the mouthless, radially sym- metrical pre-division form is strikingly reminiscent of rhabdophorine gymnostome species, which comprise a group many of which are generally considered as the most "primitive" representatives of the extant Ciliophora (Corliss, 1956, 1961). Thus we may well have here a kind of recapitulation of ancestral conditions, un- recognized before the work of Tuffrau and its interpretation by Faure-Fremiet (e.g., 1950, 1963). Recent studies by Grain (1966) and Prelle (1961, 1963) have supplied us with many additional examples.
3. The chonotrichs have for a long time represented an enigmatic group of organisms. Stalked, sedentary, with unusually-shaped mature stage and occur- ring in a specialized ecological habitat (ectocommensal on crustaceans, generally marine), they have not only been set apart taxonomically but have been thought to hold little degree of affinity with other ciliates. Study of their infraciliature- and certain other characters, such as the macronucleus and the peduncle-during ontogeny, however, carried out principally by Guilcher (1951) and Faure-Fremiet (1950, 1957; and see also Faure-Fremiet et al., 1956), threw entirely new light on the problem. Such striking resemblances to certain of the cyrtophorine gymno- stomes were discovered that the evolutionary hypothesis of the origin of the former from the latter appears to be a sound one, in spite of persistent objections from the chonotrich specialist Mohr (1959, 1966) and scattered general proto- zoologists (e.g., Poljansky & Cheissin, 1965).
4. The suctorians have traditionally been considered unique protozoa, hardly even belonging within the subphylum Ciliophora. Work by French protozool- ogists in particular, however (e.g., Chatton et al., 19,29; Faure-Fremiet, 1950;
3 Citations need not be restricted to French papers; e.g., Dogiel (1927, 1946), many years ago in Russia, described "orthogenetic lines" among the ophryoscolecids (a family of symbiotic spirotrich ciliates). Numerous other appropriate citations to the older literature may be found in Corliss (1961); very recently, Jankowski (1964a,b) has described some interesting sequences and interrelationships in the groups of ciliates which he has been studying. 4 And it still does! Obviously more data, especially concerning the full life histories of various and sundry curious forms routinely tossed into the order Trichostomatida, need to be collected and analyzed from a comparative point of view. One of the taxonomically most enigmatic genera, Cyathodinium, has recently been subjected to careful restudy (Paulin & Corliss, 1964, 1967)-it may turn out to be a (most aberrant) suctorian!
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 7
Guilcher, 1951; and see other citations in Canella, 1957; Corliss, 1961), empha- sized not only that the larval or migratory forms are ciliated but also that the infraciliary kinetosomes responsible for production of this ciliation appear to have their origin in a field of barren (i.e., without cilia) basal bodies ever-present in the cortex of the mature, sedentary form in the life cycle. With the additional realization that certain other formerly "unique" characteristics of suctorians, such as their stalks and scopulae, may well have their homologues in structures known in species belonging to other ordinal groups of ciliates, the hypothesis was born (Faure-Fremiet, 1950) that the suctorians are very closely allied to the holotrich ciliates, if indeed not just one of the eight or nine included orders within that great subclass (Corliss, 1956, 1959b, 1960, 1961). This phylogenetic-systematic conclusion is widely accepted today (e.g., see Hawes, 1961, Honigberg et al., 1964, and Hyman, 1959; but also note the continued more conservative view, as exemplified by Jankowski, 1967, Kudo, 1966, and Poljansky & Cheissin, 1965).
5. The peritrichs, whose "uniqueness" is particularly fiercely defended by specialists on the group (e.g., Clark & Finley, 1967; Finley et al., 1959; see also Hyman, 1959; Raabe, 1964b), have long been known to possess striking charac- teristics, especially in the mature forms, not often found among species of other ciliates. Though the battle continues to rage, homologies suggested some years ago (Corliss, 1956; Faure-Fremiet, 1950; see also Faure-Fremiet, 1965; Lon, 1964) on the basis of comparative studies of silver-impregnated specimens have now gained confirmative support from work with electron microscopy (Faure- Fremiet et al., 1962; Lomn et al., 1966, 1968; Noirot-Timothee & Lom, 1965). There appears to be a common structural plan for organelles comprising the buccal apparatus in organisms belonging to the separate taxonomic groups of hymenostomes, thigmotrichs, and peritrichs (see especially Lomn et al., 1968). Such data, including studies on stomatogenesis of selected forms, may not con- stitute unequivocal evidence of a direct phylogenetic line linking the three orders involved (Hymenostomatida, Thigmotrichida, Peritrichida), since parallel de- velopment may well be the most appropriate interpretation. Of importance, however, is the indication that the groups at least very likely had their origin in a common ancestral taxon-and this could never have even been postulated without study of ontogenies of species comprising the three orders.
6. Traditionally, and often still today, the great group of holotrichs (now considered as a subclass within the class Ciliatea) has been set apart from that of the spirotrichs on the basis of the possession by the latter only of an adoral zone of membranelles (AZM) in the oral area. Furgason (1940), as mentioned above, recognized that the relatively minute set of ciliary organelles in the buccal cavity of the small hymenostome Tetrahymena could be interpreted as represent- ing a "primitive AZM." (Studies on the ontogeny of the ciliate aided in verifi- cation of their presence.) And he proceeded further to postulate that the AZM would be found in additional holotrichs, that it represented an homologous structure, and that from the simple picture found in such forms as Tetrahymena one could imagine the evolution of the complex AZM long known for the spirotrich ciliates. This heuristic hypothesis stimulated a series of expansions and refinements by Faure-Fremiet and the writer and their associates (for selected examples, see Berger, 1963; Corliss, 1956, 1958, 1961; Corliss & Roque, 1967; Evans & Corliss, 1964; Faure-Fremiet, 1950, 1954, 1961b, 1965, 1968; Faure- Fremiet & Andre, 1967; Mugard, 1949; Thompson & Corliss, 1958; Tuffrau, 1963; Yusa, 1957) which involved a large number of specific orders within both the holotrich and spirotrich subclasses. Proof of the homology implied by Furgason -or, indeed, even precise definition of what one means by "membranelle" (see
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

8 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
Corliss, 1967a; Evans & Corliss, 1964; Lom et al., 1968)-is, admittedly, still mostly lacking today, but the value of the initial idea cannot be over-estimated.
7. Within the large holotrich order of hymenostomes, separation into sub- orders has been aided greatly by studies of stomatogenesis. Two generalizations have been proposed very recently (Corliss & Roque, 1967): (1) that the mode of new mouth formation and the types of oral ciliature involved allow recognition of groups at the level of suborder; and (2) that details of the process of stoma- togenesis permit the setting of boundaries at the level of family. Ontogenetic studies are, once again, proving indispensable.
In an important work very recently published, Small (1967), in agreement with Furgason (e.g., see Furgason et al., 1960), has emphasized the importance of patterns in the whole stomatogenic sequence. As a result of his comprehensive and detailed studies of stomatogenesis in genera from among the Hymenostoma- tida sensu lato and related groups, Small proposes a new order within the subclass Holotrichia, the Scuticociliatida, an order which will contain the (former) hymenostome suborder Pleuronematina (enlarged by the addition of certain species currently assigned to the thigmotrichs) and two other suborders com- prised of certain genera presently considered members of the hymenostome suborder Tetrahymenina. This research (the necessity for which had been fore- seen and the avenue of approach predicted: see especially Corliss, 1961, 1967a; Evans & Corliss, 1964) represents an important step forward in elucidation of the many mysteries enshrouding both the systematics and evolution of the generally small, relatively undifferentiated holotrich ciliates which I (Corliss, 1961) have labelled as "the pivotal group" in the phylogeny of most of the other members of the entire subphylum Ciliophora.
USE OF STOMATOCENESIS
A further comment should be inserted concerning stomatogenesis throughout all groups of mouthed ciliates, although I (Corliss, 1967a) have very recently discussed the phenomenon elsewhere in some detail: that paper is rich in references5 to pertinent literature which, therefore, need not be repeated here.
Although a dynamic morphogenetic process, there is-happily-growing evidence that stomatogenesis represents a character of considerable conservative- ness. Much greater diversity exists in patterns of buccal structures in the mature forms of ciliates than in basic patterns of new mouth-formation. I have proposed recognition of five categories of stomatogenesis (Corliss, 1964, 1967a; Evans & Corliss, 1964), but it should be emphasized that these do not relate directly to only five taxonomic groups of ciliates. Recognition of similar patterns in similar or dissimilar ciliates, however, especially if carried out with the precision seen, for example, in the work of Small (1967) cited above, might aid greatly in determining possible or even probable phylogenetic affinities among (and thus classificational positions for) species or higher groups which otherwise might remain taxonomically misplaced or unidentified.
Once again, extreme caution must be exercised in such comparative studies. One of the greatest potential stumbling blocks in examining patterns of stoma- togenesis in different taxonomic series of species would be-and, unfortunately, has been-assignment of the term "homologus" to similar-appearing structures which may, in reality, be entirely without homology. And forms that "don't fit" must not be shoved aside as "aberrant"! Many controversies in the recent ciliate
5Among several inadvertently omitted, however, were citations to the recent thorough investigations by Canella & Canella (1964) on stomatogenesis in Ophryoglena, a paper includ- ing discussion of the possible phylogenetic implications of their findings (also see Canella, 1964), and by Dobrzanska-Kacznowska (1963) on comparative morphogenesis of several important holotrich genera.
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 9
literature will be resolvable as more and better data become available, assuming judicious treatment of such new information and wholesome respect of alternative hypotheses by all of the research workers involved.
DISCUSSION AND SUMMIARY
Long discussion of the examples just given, or of the overall hypothesis that studies of ontogeny of protozoa are of solid value in phylogenetic considerations, seems inappropriate. A number of the major weaknesses, criticisms, dangers, or pitfalls have been mentioned along the way.
Although mild reproaches have emanated from time to time from American laboratories,6 especially from specialists on particular ordinal groups (e.g., Mohr, 1966, on chonotrichs; Finley et al., 1959, on peritrichs), the most comprehensive criticism of the phylogenetic ideas of Faure-Fremiet, Furgason, and myself is embodied in a lengthy monographic review recently published by Canella (1964) of Ferrara, Italy.7 Canella questions the actual homology of some structures (e.g., the AZM of Tetrahymena and of spirotrich ciliates); denies the broad usefulness of the tetrahymenal buccal apparatus concept; assails assumptions that certain groups are necessarily "ancestral" to others; decries concern over polyphyly; believes that facts are too often forced to fit hypotheses and that prejudicial criteria are employed; suggests that ecological and other factors are too little studied; has no faith at all in the "biogenetic law" and not much even in natural selection; considers the ciliate literature to be growing in confusion and contradiction and the complex of (new) hypotheses to be upsetting; chides workers for their lack of objectivity and accuses them of too much positiveness (they should show more doubt); and, finally, strongly advises a moratorium on all phylogenetic speculation.
The thought-provoking discussions of Canella are of genuine value, even if for no other reason than to make those of us who are prime objects of his criticism stop and re-think. Although he is guilty of occasional inconsistency, many of his points are exceedingly well made, legitimate, justifiable, and deserv- ing of far more comment than is possible here.
Though extensive consideration, or even rebuttal, of Canella's sobering argu- ments cannot be presented here, his paper must not be neglected by any worker contemplating entry into the realm of ciliate phylogeny. And several points are made below, even though limited to simple statements, since they may serve both to (at least partially) allay Canella's major objections and to serve as a summary of the present paper.
1. We must be careful not to apply the word "homology" too loosely with reference to structures which appear to be very similar morphologically or even morphogenetically. Other workers besides Canella have also recently stressed this all-important point (e.g., Grimstone, 1959).
2. We must select with extreme care characteristics considered to be of greatest value, avoiding, too, a priori weighting in such selection.
3. We must not consider as unquestionable proof of evolutionary relationship the occurrence in the ontogeny of a given ciliate of stages which are (merely) reminiscent of some stage(s) in the life cycle of a ciliate belonging to another taxonomic group.
6 And from certain European centers of ciliate research; e.g., see Raabe (1964a) of Poland, Poljansky & Cheissin (1965) and Jankowski (1967) of Russia, Grell (1956) of Germany. But the disagreements are usually only partial; indeed, differences among the critics themselves may sometimes be greater than between any one of their schemes and the one endorsed by the writer (see Appendix II).
7 I am indebted to Dr. Waldo H. Furgason, University of California at Los Angeles, for an English translation of this Italian paper.
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

10 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
4. We must recognize the widespread existence of polyphyly among pro- tozoan groups. And we must keep constantly in mind the problems posed by parallel evolution, adaptations to similar environments, and similar potential pitfalls.
5. We should retain objectivity as a major goal in this as well as other fields of protozoological research. Hypotheses not reasonably supportable by facts should be abandoned or viewed with heavy skepticism. Rampant speculation must be discouraged.
In short, the same criteria employed in the best work on problems of metazoan phylogeny must be applied to the area of research discussed in the present paper. (The reader is referred to the following convenient set of major authoritative books or monographs on the subjects of metazoan evolution, phylogeny, and systematics: Dougherty et al., 1963; Hennig, 1950; Mayr, 1963; Mayr et al., 1968; Remane, 1956; Rensch, 1959; Simpson, 1953, 1961).
If the above admonitions are kept in mind, as well as the dangers and pitfalls alluded to in earlier pages (and in previous publications: e.g., Corliss, 1960, 1962, 1963a, 1967b), I believe that there is no need to join Canella (1964) in pessimism about the value of protozoan phylogenetic investigations or the role of ontogenetic studies in such work. I do not share his dismay over the profusion and confusion of writings on the subject during the past two decades. Out of such seeming chaos may well come order.
Promulgation of phylogenetic hypotheses need not "degenerate into a kind of scholarly indoor past-time," as a British experimental biologist and recent Nobel-prize laureate has worriedly warned the world of systematists at large. Cautious speculation, it seems to me, is healthy-and it has proven to be heu- ristic.8 The recent and what appear to be very sound proposals of Small (1967), for example, would probably never have come into being if he had not had his curiosity aroused by the profound pronouncements of Furgason and Faure- Fremiet, hypotheses with which he is by no means in full agreement. The similarly recent monograph by Jankowski (1967), in which a strikingly new taxonomic hierarchy is proposed for the entire subphylum Ciliophora, represents a major overhauling of ciliate systematics which would have been quite impos- sible without a modern scheme of classification (much of which is based on phylogenetic considerations) to attack.
In closing these remarks, I should like to stress once again that the principal value of ontogenetic investigations in reconstruction of protozoan phylogenies rests on our giving critical attention to the pattern of developing structures or organellar systems in the full life cycles of the organisms under comparative study. Similarities which can be identified in key stages of the dynamic mor- phogenetic processes which comprise ontogeny generally stand a reasonable chance of involving truly homologous structures. Then-but only then-it would seem that we are justified in drawing tentative conclusions about the possible affinities of the specific forms or groups in question. Such research opportunities may exist in much greater numbers throughout the phylum than has been com- monly realized to date; so I am optimistic about our future progress in the field of protozoan phylogeny, slow though advances may come at first.
LITERATURE CITED
BAER, K. E., VON. 1828. Ueber Entwicklungsgeschichte der Thiere. Beobachtungen und Reflexion. Wilhelm Koch, Konigsberg.
8 Pertinent here, perhaps, is the optimistic principle endorsed by the late E. C. Dougherty (in Dougherty & Allen, 1960): "(it is) better to have a working hypothesis, even if based on fragile evidence, than to shrug aside a question of phylogeny as prematurely posed."
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 11
BERGER, J. 1963. The comparative buccal morphology of certain hymenostome ciliates entocommensal in echinoids. In Ludvik, J., Lom, J. & Vavra, J., eds., Progress in Pro-
tozoology, Proc. 1st Int. Cong. Protozool., Prague, Aug. 1961, pp. 86-88. CALKINS, G. N. 1933. The Biology of the Protozoa. 2nd ed. Lea and Febiger, Philadelphia. CANELLA, M. F. 1957. Studi e ricerche sui tentaculiferi nel quadro della biologia generale.
Ann. Univ. Ferrara (N. S., Sect. III), 1: 259-716. 1964. Strutture buccali, infraciliatura, filogenesi e sistematica dei ciliofori. Fatti, ipotesi,
speculazioni. Ann. Univ. Ferrara (N. S., Sect. III), 2: 119-188. CANELLA, M. F. & CANELLA, I. R. 1964. Morfo-fisiologia e morfogenesi dell'apparato orale
di Ophryoglena (Ciliata, Holotricha, Hymenostomatida). Ann. Univ. Ferrara (N. S., Sect. III), 2: 190-292.
CHATTON, E. 1942. Le probleme de la continuite du cinetome chez les hypotriches. Nou- velles recherches sur l'Euplotes crassus. Bull. biol. France Belg., 76: 314-335.
CHATTON, R. & LWOFF, A. 1935. Les cilies apostomes. I. Apercu historique et general; etude monographique des generes et des especes. Arch. Zool. exptl. gen., 77: 1-453.
1936. Les Pilisuctoridae Ch. et Lw. Cilies parasites des poils secreteurs des crustaces
edriophthalmes. Polarite, orientation, et desmodexie chez les infusoires. Bull. biol. France Belg., 70: 86-144.
1949. Recherches sur les cilies thigmotriches. I. Arch. Zool. exptl. gen., 86: 169-253. 1950. Recherches sur les cilies thigmotriches. II. Arch. Zool. exptl. gen., 86: 393-485.
CHATTON, R., LWOFF, A., LWOFF, M. & TELLIER, L. 1929. L'infraciliature et la continuite genetique des blepharoplastes chez l'acinetien Podophrya fixa 0. F. Muller. Compt. rend. Soc. Biol., Paris, 100: 1191-1196.
CHATTON, 1. & PERARD, C. 1921. Les Nicollellidae, infusoires intestinaux des gondis et des damans et le "cycle evolutif" des cilies. Bull. biol. France Belg., 55: 86-151.
CHATTON, 1. & SEGUELA, J. 1940. La continuite genetique des formations ciliaires chez les cilies hypotriches. Le cinetome et l'argyrome au cours de la division. Bull. biol. France
Belg., 74: 349-442. CHATTON, R. & VILLENEUVE, S. 1937. La division de la bouche et la formation du peristome
chez les peritriches (Cyclochaeta astropectinis, n. sp.). Leur continuite genetique im- mediate. Compt. rend. Acad. Sci., Paris, 204: 538-541.
CHEISSIN, E. & POLJANSKY, G. I. 1963. On the taxonomic system of Protozoa. Acta Pro- tozool., 1: 327-352.
CLARK, L. G. & FINLEY, H. E. 1967. Biological observations on the colonial peritrich Campanella tincta. (Abstr.) J. Protozool., 14(Suppl.): 32.
CORLISS, J. 0. 1953a. Protozoa and systematics. Yale Sci. Mag., 28: 14-17, 36, 38, 40. 1953b. Silver impregnation of ciliated protozoa by the Chatton-Lwoff technic. Stain Tech-
nol., 28: 97-100. 1956. On the evolution and systematics of ciliated protozoa. Syst. Zool., 5: 68-91, 121-
140. 1958. The phylogenetic significance of the genus Pseudomicrothorax in the evolution of
holotrichous ciliates. Acta biol. Acad. sci. hung., 8: 367-388. 1959a. Some basic problems in the study of evolution of the ciliated protozoa. Proc. XV
Int. Cong. Zool., London, 1958, pp. 179-182. 1959b. An illustrated key to the higher groups of the ciliated protozoa, with definition of
terms. J. Protozool., 6: 265-281. 1960. Comments on the systematics and phylogeny of the protozoa. Syst. Zool., 8(yr.
1959): 169-190. 1961. The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature.
Pergamon Press, London and New York. 310 pp. 1962. Taxonomic procedures in classification of protozoa. Symp. Soc. gen. Microbiol.,
12: 36-67. 1963a. Application of phylogenetic considerations to protozoan systematics. In Ludvik, J.,
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

12 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
Lom, J. & Vavra, J., eds., Progress in Protozoology, Proc. 1st Int. Cong. Protozool., Prague, Aug. 1961, pp. 47-56.
1963b. Application of modern techniques to problems in the systematics of the protozoa. Proc. XVI Int. Cong. Zool., Washington, D. C., Aug. 1963, 4: 97-102.
1964. The value of stomatogenesis in studies of ciliate phylogeny and systematics. (Abstr.) J. Protozool., 11(Suppl.): 4.
1967a. An aspect of morphogenesis in the ciliate protozoa. J. Protozool., 14: 1-8. 1967b. Newer trends in the systematics of the Protozoa. Bull. Nat. Inst. Sci. India, No.
34: 26-33. 1968. Systematics of the phylum Protozoa. In Florkin, M. & Scheer, B. T., eds., Chemical
Zoology, Vol. I, pp. 1-20. CORLISS, J. O., LOM, J. & SMALL, E. B. 1968. Use of silver impregnation techniques in
ciliate protozoology. (MS in preparation) CORLISS, J. O. & ROQUE, M. 1967. A direct use of stomatogenesis in supra-generic classifica-
tion of ciliate protozoa. (Abstr.) J. Protozool., 14(Suppl.): 33. DE BEER, G. R. 1951. Embryos and Ancestors. Clarendon Press, Oxford. DOBRZANSKA-KACZANOWSKA, J. 1963. Comparison de la morphogenese des cilies: Chilo-
donella uncinata (Ehrbg.), Allosphaerium paraconvexa et Heliochona scheuteni (Stein). Acta Protozool., 1: 353-394.
DOGIEL, V. A. 1927. Monographie der Familie Ophryoscolecidae. I. Arch. Protistenk., 59: 1-288.
1946. Phylogeny of the family Ophryoscolecidae from a palaeontological and parasito- logical standpoint. Zool. Zh., 25: 395-402.
DOUGHERTY, E. C. & ALLEN, M. B. 1960. Is pigmentation a clue to protistan phylogeny? In Allen, M. B., ed., Comparative Biochemistry of Photoreactive Systems. Symp. Comp. Biol., Vol. I, Academic Press, New York, pp. 129-144.
DOUGHERTY, E. C., BROWN, Z. N., HANSON, E. D. & HARTMAN, W. D., eds. 1963. The Lower Metazoa, Comparative Biology and Phylogeny. Univ. of Calif. Press, Berkeley.
DRAGESCO, J. 1960. Cilies mesopsammiques littoraux. Systematique, morphologie, ecologie. Trav. Sta. biol. Roscoff (N. S.), 12: 1-356.
1962. L'orientation actuelle de la systematique des cilies et la technique d'impregnation au proteinate d'argent. Bull. Microsc. appl., 11: 49-58.
1963. Complements a la connaissance des cilies mesopsammiques de Roscoff. I. Holo- triches. II. Heterotriches. III. Hypotriches. Cah. Biol. mar., 4: 91-119, 251-275.
EVANS, F. R. & CORLISS, J. 0. 1964. Morphogenesis in the hymenostome ciliate Pseudo- cohnilembus persalinus and its taxonomic and phylogenetic implications. J. Protozool., 11: 353-370.
FAURE-FREMIET, E. 1950. Morphologie comparee et systematique des cilies. Bull. Soc. zool.
France, 75: 109-122. 1953. La bipartition enantiotrope chez les cilies obligotriches. Arch. Anat. microsc.
Morphol. exptl., 42: 209-225. 1954. Morphogenese de bipartition chez Urocentrum turbo (cilie holotriche). J. Embryol.
exptl. Morph., 2: 227-238. 1957. Le macronucleus heteromere de quelques cilies. J. Protozool., 4: 7-17. 1959. La famille des Nassulidae (Ciliata Gymnostomatida) et le genre Nassulopsis n. gen.
Compt. rend. Acad. Sci., Paris, 249: 1429-1433. 1961a. Remarques sur la morphologie comparee et la systematique des Ciliata Hypotrichida.
Compt. rend. Acad. Sci., Paris, 252: 3515-3519. 1961b. Les cilies Cyrtophorina et leur diversification morphologique. Compt. rend. Acad.
Sci., Paris, 252: 3912-3916. 1962. Le genre Paranassula Kahl (Ciliata Cyrtophorina). Cah. Biol. mar., 3: 61-77. 1963. Remarques sur la morphologie comparee des cilies Trichostomatida. Compt. rend.
Acad. Sci., Paris, 257: 3089-3094. 1965. Morphologie comparee des cilies Peritrichida. Excerpta Medica, Int. Cong. Ser.
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS--VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 13
No. 91, 2nd Int. Conf. Protozool., London, Aug. 1965, Progress in Protozoology, pp. 13-14.
1968. La differenciation des structures buccales au cours de l'evolution des Ciliata.
(Unpublished MS) FAURE-FREMIET, E. & ANDRE, J. 1967. Etude au microscope electronique du cilie Pseudo-
microthorax dubius Maupas. J. Protozool., 14: 464-473. FAURE-FREMIET, E., FAVARD, P. & CARASSO, N. 1962. Etude au microscope electronique
des ultrastructures d'Epistylis anastatica (cilie peritriche). J. Microsc., 1: 287-312.
FAURE-FREMIET, E., ROUILLER, C. & GAUCHERY, M. 1956. Structure et origine du peduncule chez Chilodochona. J. Protozool., 3: 188-193.
FINLEY, H. E., MCLOUGHLIN, D. & HARRISON, D. M. 1959. Non-axenic and axenic growth of Vorticella microstoma. J. Protozool., 6: 201-205.
FURGASON, W. H. 1940. The significant cytostomal pattern of the "Glaucoma-Colpidium group," and a proposed new genus and species, Tetrahymena geleii. Arch. Protistenk., 94: 224-266.
FURGASON, W. H., SMALL, E. B. & LOEFER, J. B. 1960. New ideas regarding the mor- phogenesis of the buccal apparatus in hymenostome ciliates. Compt. rend. X Cong. Int. Biol. Cell., Paris, p. 230.
GELEI, J., VON. 1950. Die Morphogenese der Einzeller mit Rucksicht auf die morpho- genetischen Prinzipien von Sewertzoff. Acta biol. Acad. sci. hung., 1: 69-134.
GRAIN, J. 1966. Etude cytologique de quelques cilies holotriches endocommensaux des ruminants et des equides. Protistologica, 2(1): 59-141; 2(2): 5-51.
GRELL, K. G. 1956. Protozoologie. Springer, Berlin. (Revised edition expected in 1968) GRIMSTONE, A. V. 1959. Cytology, homology and phylogeny-a note on "organic design."
Amer. Nat., 93: 273-282. 1961. Fine structure and morphogenesis in protozoa. Biol. Rev., 36: 97-150. 1966. Structure and function in protozoa. Ann. Rev. Microbiol., 20: 131-150.
GUILCHER, Y. 1951. Contribution a l'etude des cilies gemmipares, chonotriches et tenta- culiferes. Ann. Sci. nat. Zool. (ser. 11), 13: 33-132.
HADZI, J. 1952. Application of the principles of phylembryogenesis to the protista. Nature, 169: 1019.
HAECKEL, E. 1866. Generelle Morphologie der Organismen. II. Allgemeine Entwicklungs- geschichte der Organismen. G. Reimer, Berlin.
1875. Ziel und Wege der heutigen Entwicklungsgeschichte. Hermann Dufft, Jena. HAWES, R. S. J. 1961. The Ciliata. In Mackinnon, D. L. & Hawes, R. S. J., An Introduction
to the Study of Protozoa. Clarendon Press, Oxford. HENNIG, W. 1950. Grundzuge einer Theorie der phylogenetischen Systematik. Deutscher
Zentralverlag, Berlin.
HONIGBERG, B. M. 1963a. A contribution to systematics of the non-pigmented flagellates. In Ludvik, J., Lorn, J. & Vavra, J., eds., Progress in Protozoology, Proc. 1st Int. Cong.
Protozool., Prague, Aug. 1961, pp. 68-69. 1963b. Evolutionary and systematic relationships in the flagellate order Trichomonadida
Kirby. J. Protozool., 10: 20-63. 1967a. Development in microscopy in relation to our understanding of protozoa. Trans.
Amer. Microsc. Soc., 86: 101-112. 1967b. Problems in classification of protozoa. Bull. Nat. Inst. Sci. India, No. 34: 13-19.
HONIGBERG, B. M., BALAMUTH, W., BOVEE, E. C., CORLISS, J. O., GOJDICS, M., HALL, R. P., KUDO, R. R., LEVINE, N. D., LOEBLICH, A. R., JR., WEISER, J. & WENRICH, D. H. 1964. A revised classification of the phylum Protozoa. J. Protozool., 11: 7-20.
JANKOWSKI, A. W. 1964a. Morphology and evolution of Ciliophora III. Diagnoses and
phylogenesis of 53 sapropelebionts, mainly of the order Heterotrichida. Arch. Pro-
tistenk., 107: 185-294. 1964b. Morphology and evolution of Ciliophora. IV. Sapropelebionts of the family Loxo-
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

14 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
cephalidae fam. nova, their taxonomy and evolutionary history. Acta Protozool., 2: 33-58.
1967. Principles of construction of a new system of ciliate protozoa (Protozoa: Cili-
ophora). Zool. Inst. Acad. Sci. USSR, Sci. sess., Leningrad, 63: 3-52. (In Russian) KIRBY, H. 1944. Some observations on cytology and morphogenesis in flagellate protozoa.
J. Morph., 75: 361-421. 1949. Morphology, morphogeny and kinship in some nonpigmented Flagellata. Compt.
rend. XIII Cong. Int. Zool., Paris, 1948, pp. 191-194. 1950. Systematic differentiation and evolution of flagellates in termites. Rev. Soc. Mex.
Hist. Nat., 10(yr 1949): 57-79. KUDO, R. R. 1966. Protozoology. 5th ed. Thomas, Springfield, Illinois. LEVINE, N. D., ed. 1966. Discussion of the classification of Protozoa at the 2nd Int. Conf.
on Protozool., London. J. Protozool., 13: 189-195. Lo\M, J. 1964. The morphology and morphogenesis of the buccal ciliary organelles in some
peritrichous ciliates. Arch. Protistenk., 107: 131-162. LoM, J., CORLISS, J. O. & NOIROT-TIMOTHEE, C. 1966. Comparison of the buccal apparatus
in thigmotrich and peritrich ciliates. (Abstr.) J. Protozool., 13(Suppl.): 8. 1968. Observations on the ultrastructure of the buccal apparatus in thigmotrich ciliates
and their bearing on thigmotrich-peritrich affinities. (MS in preparation) MAYR, E. 1963. Animal Species and Evolution. Harvard University Press, Cambridge.
1964. The new systematics. In Leone, C. A., ed., Taxonomic Biochemistry and Serology. Ronald Press, New York.
1965. Classification and phylogeny. Amer. Zool., 5: 165-174. MAYR, E., LINSLEY, E. G. & USINGER, R. L. 1968. Methods and Principles of Systematic
Zoology. 2nd ed. McGraw-Hill, New York. MOHR, J. L. 1959. Ciliates on crustaceans; the evolution of the order Chonotrichida. Proc.
XV Int. Cong. Zool., London, 1958, pp. 651-653. 1966. On the age of the ciliate group Chonotricha. In Barnes, H., ed., Some Contemporary
Studies in Marine Science, George Allen and Unwin Ltd., London, pp. 535-543. MUGARD, H. 1949. Contribution a l'etude des infusoires hymenostomes histiophages. Ann.
Sci. nat. Zool. (ser. 11), 10: 171-268. NOIROT-TIMOTHEE, C. 1960. Rtude d'une famille de cilies: les Ophryoscolecidae. Structures
et ultrastructures. Ann. Sci. nat. Zool. (ser. 12), 2: 527-718. NOIROT-TIMOTHEE, C. & LoNf, J. 1965. L'ultrastructure de l"'haplocinetie" des cilies peri-
triches comparison avec la membrane ondulante des hymenostomes. Protistologica, 1: 33-41.
ORTON, G. L. 1955. The role of ontogeny in systematics and evolution. Evolution, 9: 75-83.
PAULIN, J. J. & CORLISS, J. O. 1964. The somatic and oral infraciliature of the enigmatic ciliate Cyathodinium piriforme. J. Protozool., 11: 438-444.
1967. Ultrastructure of endosprits (= nemadesmata?) in species of Cyathodinium. (Abstr.) J. Protozool., 14(Suppl.): 26.
PITELKA, D. R. 1963. Electron-Microscopic Structure of Protozoa. Pergamon Press, London and New York.
POLJANSKY, G. I. & CHEISSIN, E. 1965. Dogiel's General Protozoology. Rev. ed., Oxford Univ. Press, London and New York. (English translation of the 1962 Russian revision of Dogiel's original 1951 publication)
PRELLE, A. 1961. Contribution a l'etude de Leptopharynx costatus (Mermod) (infusoire cilie). Bull. biol. France Belg., 95: 731-752.
1963. Bipartition et morphogenese chez le cilie holotriche Woodruffia metabolica (John- son et Larson, 1938). Compt. rend. Acad. Sci., Paris, 257: 2148-2151.
PUYTORAC, P. DE. 1954. Contribution a l'etude cytologique et taxonomique des infusoires astomes. Ann. Sci. Nat. Zool. (ser. 11), 16: 85-270.
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 15
1959. Quelques observations sur l'evolution et les origines des cilies Astomatida. Proc. XV Int. Cong. Zool., London, 1958, pp. 649-651.
1963. Contribution a l'etude des cilies astomes Haptophryidae Cepede, 1903 (cytologie, ultrastructure, taxinomie). Ann. Sci. Nat. Zool. (ser. 12), 5: 173-210.
RAABE, Z. 1958. An attempt of a revision of the system of Protozoa. Ann. Univ. M. Curie- Sklodowska, Sect. C, 3: 259-276. (In Polish with English summary)
1964a. Remarks on the principles and outline of the system of protozoa. Acta Protozool., 2: 1-18.
1964b. The taxonomic position and rank of Peritricha. Acta Protozool., 2: 19-32. REMANE, A. 1956. Die Grundlagen des Natiirlichen Systems der Vergleichenden Anatomie
und der Phylogenetik. Akademische Verlagsgesellschaft Geest and Portig, Leipzig. RENSCH, B. 1959. Evolution Above the Species Level. Columbia University Press, New
York. ROQUE, M. 1961. Recherches sur les infusoires cilies: les hymenostomes peniculiens. Bull.
biol. France Belg., 95: 432-519. SEWERTZWOFF, A. N. 1931. Die Morphologische Gesetzmiissigkeiten der Evolution. Fischer,
Jena. SIMPSON, G. G. 1953. The Major Features of Evolution. Columbia University Press, New
York. 1961. Principles of Animal Taxonomy. Columbia University Press, New York.
SMALL, E. B. 1967. The Scuticociliatida, a new order of the class Ciliatea (phylum Protozoa, subphylum Ciliophora). Trans. Amer. Microsc. Soc., 86: 345-370.
STOUT, J. D. 1960. Morphogenesis in the ciliate Bresslaua vorax Kahl and the phylogeny of the Colpodidae. J. Protozool., 7: 26-35.
THOmPSON, J. C., JR. & CORLIss, J. 0. 1958. A redescription of the holotrichous ciliate Pseudomicrothorax dubius with particular attention to its morphogenesis. J. Protozool., 15: 175-184.
TUFFRAU, M. 1952. La morphogenese de division chez les Colpodidae. Bull. biol. France Belg., 86: 309-320.
1960. Revision du genre Euplotes, fondee sur la comparaison des structures superficielles. Hydrobiologia, 15: 1-77.
1963. L'infraciliature, la morphogenese et la position systematique du genre Lembadion Perty, 1852 (cili6 hymenostome). Ann. Sci. Nat. Zool. (ser. 12), 5: 479-490.
1967. Perfectionnements et pratique de la technique d'impregnation au protargol des infusoires cilies. Protistologica, 3: 91-98.
UHLIG, G. 1967. Protargol-impregnation for taxonomic studies in ciliated protozoa. In Schlieper, C. Methoden der Meeresbiologischen Forschung. Fischer, Jena-Berlin.
VILLENEUVE-BRACHON, S. 1940. Recherches sur les cilies heterotriches. Arch. Zool. exptl. gen., 82: 1-180.
WAGNER, W. H., JR. 1968. The construction of a classification. (In a publication of the Proceedings of the "International Conference on Systematic Biology" held in Ann Arbor, Michigan, June 1967, in press)
YUSA, A. 1957. The morphology and morphogenesis of the buccal organelles in Paramecium with particular reference to their systematic significance. J. Protozool., 4: 128-142.
APPENDIX I
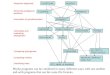
Although certain changes are already indicated, by recent work of the writer and of others, a "phylogenetic tree" or dendrogram showing alleged affinities among orders and suborders comprising the protozoan subphylum Ciliophora is here appended (see Fig. 1, p. 17) as an example of the theoretical results made possible by comparative study of extant ciliates, with the emphasis in such
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

16 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
investigations on morphogenetic patterns seen in selected ontogenies. The recon- structed "tree," altered only slightly from that published by the writer several years ago (see Corliss, 1962), represents an effort to indicate postulated principal evolutionary lines among ciliates, at the ordinal level. Most of the research leading to such a dendrogram has been carried out by French workers (see text of this paper); indeed, the tree presented here is based essentially on the stim- ulating one first published by Faure-Fremiet (1950), though later refinements of some importance have been incorporated.
Within the past two or three years a number of publications have supplied data which will undoubtedly necessitate further modification in the dendrogram shown here (see text). Especially to be cited are the significant works of very recent date by Jankowski (1967) and Small (1967). Although such papers underline the transitory nature of my ciliate genealogical tree, and also emphasize its obvious oversimplifications, they do not, in my opinion, change the value of our having even a temporary "frame of reference" on which to hang our tentative hypotheses. The tree gives its critics something definite to chop down! Without question, new data and fresh ideas will require new schemes of clas- sification and new proposals with respect to phylogenetic interrelationships among the major taxa of the ciliates.
APPENDIX II
For the convenience, especially, of the reader not too familiar with the recent literature on the systematics of the phylum Protozoa, the classification adopted by the internationally composed "Honigberg Committee" (see Honigberg et al., 1964) is given below (pp. 18-20). The writer is well aware of some of its deficiencies, yet the scheme-based, wherever possible, on phylogenetic as well as other considerations-represents: (a) a considerable improvement over the majority of classical or conventional-and generally more conservative-classifi- cations of the Protozoa; (b) an attempt to include significant data obtained in very recent years through the use, often, of sophisticated techniques (e.g., electron microscopy); (c) a compromise between extreme views of certain specialists on some groups; (d) an internally consistent, workable outline amenable to emendation as new data of pertinence accrue.
The classification is not expected to remain immutable nor to gain universal acceptance through "weight of authority." Already various criticisms have appeared in print (see text or glance down through the titles of papers in LITERATURE CITED) and more may be anticipated. Overall revisions will have to be undertaken from time to time. Even for the ciliates, which perhaps represent the least controversial section, changes will eventually have to be made; one may note, in passing, that the dendrogram discussed briefly in Appendix I, in which only two subclasses are shown, is at some variance with the distribution of orders (into four subclasses) given below (p. 20). As always-and as in all fields of biological research-"more work needs to be done." But one cannot, in my opinion, divorce classification from phylogenetic considerations-nor vice versa.
Important note: the original paper by Honigberg et al. (1964) should be seen by the interested reader-for not only the important characterizations included therein but also for its score of explanatory footnotes. In some of these carefully prepared footnotes, for example, the very criticisms which have sub- sequently (since 1964) arisen were foreseen.
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 17
HYPOTRICHIDA / (SPORADOTRICHINA) t
ENTODINIOMORPHIDA
V^/ \ 1Jilll/ HYPOTRICHIDA /
T
C
\. IS ^(STICHOT ICHINv
ODONTOSTOMATIDA
TINTINNIDA \
'' ^v OLIGOTRICHIDA / ORIA
7 1 HETEROTRCD HETEROTRICHIDA ,'"'<^\1^^~~~~ Ns.<~~ ^i.Jl'(LICNOPHORINA)
HETEROTRICHIDA (HETEROTRICHINA)
SPIROTRICI
HOLOTRICHA PERITRICHIDA /i'
4 (MOBILINA)
PERITRICHIDA (SESSILINA)
THIGMOTRICHIDA (ARHYNCHODINA)
/i/) : -/ THIGMOTRICHIDA ( ?/Xs i ^ 0''t;- / (RHYNCHODINA)
HYMENOSTOMATIDA ' . (PENICULINA) < HYMENOSTOMATIDA
\ (PLEURONEMATINA)
\ V S X 1f 1,,,,/
\ HYMENOSTOMATIDA ~ (TETRAHYMENINA) ASTOMATIDA
APOSTOMATIDA
? ? X \0 W "/<'
~
UNKNOWN ZOOFLAGELLATE ANCESTRY
FIG. 1. Dendrogram indicating possible affinities among the ciliate Protozoa.
HA
i;-- I
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

18 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
Phylum PROTOZOA Subphylum I. SARCOMASTIGOPHORA
Superclass I. Mastigophora Class 1. Phytomastigophorea
Order 1. Chrysomonadida Suborder (1) Euchrysomonadina Suborder (2) Rhizochrysidina Suborder (3) Chrysocapsina
Order 2. Silicoflagellida Order 3. Coccolithophorida Order 4. Heterochlorida Order 5. Cryptomonadida Order 6. Dinoflagellida
Suborder (1) Adinina Suborder (2) Diniferina
Order 7. Ebriida Order 8. Euglenida
Suborder (1) Euglenina Suborder (2) Peranematina Suborder (3) Petalomonadina
Order 9. Chloromonadida Order 10. Volvocida
Class 2. Zoomastigophorea Order 1. Choanoflagellida Order 2. Bicosoecida Order 3. Rhizomastigida Order 4. Kinetoplastida
Suborder (1) Bodonina Suborder (2) Trypanosomatina
Order 5. Retortamonadida Order 6. Diplomonadida Order 7. Oxymonadida Order 8. Trichomonadida Order 9. Hypermastigida
Suborder (1) Lophomonadina Suborder (2) Trichonymphina
Superclass II. Opalinata Order 1. Opalinida
Superclass III. Sarcodina Class 1. Rhizopodea
Subclass (1) Lobosia Order 1. Amoebida Order 2. Arcellinida
Subclass (2) Filosia Order 1. Aconchulinida Order 2. Gromiida
Subclass (3) Granuloreticulosia Order 1. Athalamida Order 2. Foraminiferida
Suborder (1) Allogromiina Suborder (2) Textulariina Suborder (3) Fusulinina Suborder (4) Miliolina Suborder (5) Rotaliina
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

CORLISS-VALUE OF ONTOGENY IN RECONSTRUCTING PHYLOGENY 19
Order 3. Xenophyophorida Subclass (4) Mycetozoia
Order 1. Acrasida Order 2. Eumycetozoida Order 3. Plasmodiophorida
Subclass (5) Labyrinthulia Order 1. Labyrinthulida
Class 2. Piroplasmea Order 1. Piroplasmida
Class 3. Actinopodea Subclass (1) Radiolaria
Order 1. Porulosida Order 2. Oculosida
Suborder (1) Nassellarina Suborder (2) Phaeodarina
Subclass (2) Acantharia Order 1. Acanthometrida Order 2. Acanthophractida
Subclass (3) Heliozoia Order 1. Actinophryida Order 2. Centrohelida Order 3. Desmothoracida
Subclass (4) Proteomyxidia Order 1. Proteomyxida
Subphylum II. SPOROZOA Class 1. Telosporea
Subclass (1) Gregarinia Order 1. Archigregarinida Order 2. Eugregarinida
Suborder (1) Acephalina Suborder (2) Cephalina
Order 3. Neogregarinida Subclass (2) Coccidia
Order 1. Protococcida Order 2. Eucoccida
Suborder (1) Adeleina Suborder (2) Eimeriina Suborder (3) Haemosporina
Class 2. Toxoplasmea Order 1. Toxoplasmida
Class 3. Haplosporea Order 1. Haplosporida
Subphylum III. CNIDOSPORA Class 1. Myxosporidea
Order 1. Myxosporida Suborder (1) Unipolarina Suborder (2) Bipolarina
Order 2. Actinomyxida Order 3. Helicosporida
Class 2. Microsporidea Order 1. Microsporida
Suborder (1) Monocnidina Suborder (2) Dicnidina
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions

20 TRANS. AMER. MICROSC. SOC., VOL. 87, NO. 1, JANUARY 1968
Subphylum IV. CILIOPHORA Class 1. Ciliatea
Subclass (1) Holotrichia Order 1 Gymnostomatida
Suborder (1) Rhabdophorina Suborder (2) Cyrtophorina
Order 2. Trichostomatida Order 3. Chonotrichida Order 4. Apostomatida Order 5. Astomatida Order 6. Hymenostomatida
Suborder (1) Tetrahymenina Suborder (2) Peniculina Suborder (3) Pleuronematina
Order 7. Thigmotrichida Suborder (1) Arhynchodina Suborder (2) Rhynchodina
Subclass (2) Peritrichia Order 1. Peritrichida
Suborder (1) Sessilina Suborder (2) Mobilina
Subclass (3) Suctoria Order 1. Suctorida
Subclass (4) Spirotrichia Order 1. Heterotrichida
Suborder (1) Heterotrichina Suborder (2) Licnophorina
Order 2. Oligotrichida Order 3. Tintinnida Order 4. Entodiniomorphida Order 5. Odontostomatida Order 6. Hypotrichida
Suborder (1) Stichotrichina Suborder (2) Sporadotrichina
II
This content downloaded from 141.101.201.103 on Sat, 28 Jun 2014 11:57:03 AMAll use subject to JSTOR Terms and Conditions