Embed Size (px)
Citation preview

The Use of General Ecological Principles in ArchaeologyAuthor(s): Donald L. HardestySource: Advances in Archaeological Method and Theory, Vol. 3 (1980), pp. 157-187Published by: SpringerStable URL: http://www.jstor.org/stable/20170156 .
Accessed: 15/06/2014 10:53
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Springer is collaborating with JSTOR to digitize, preserve and extend access to Advances in ArchaeologicalMethod and Theory.
http://www.jstor.org
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

The Use of General
Ecological Principles in Archaeology
DONALD L HARDESTY
Ecology and archaeology have been acquaintances for a long time, and
the personality of archaeological methods and theory has been strongly in
fluenced by the contact. The relationship is, however, distant. Watson,
LeBlanc, and Redman (1971:107) summarize it best as a perspective, a way of looking at the world that may be useful in answering some questions; that is, ecology is used heuristically, not as a set of formal explanatory prin
ciples that can be brought to bear upon archaeological problems. Not
withstanding the large contribution that the heuristic approach has made to
archaeological research, it has its disadvantages. Explanatory models are ad
hoc, constructed for archaeological data from a particular site or region; because they are tied to specific environments and to specific human
groups, the models are unlikely to have any relevance elsewhere. The reason
is that predictions drawn from such models are most likely to be confirmed
under ecological and cultural settings that are identical to the model. That
is, of course, never the case; because of historical factors, no two settings are the same. The probable outcome is the failure to confirm most of the
predictions, giving the impression that there are no general ecological prin
ciples, that the human ecology of each site or region is unique. But the possibility of that impression being an illusion cannot be over
looked. Simon (1969) points out, for example, that a few simple princi
ples operating in complex environments can produce an almost infinite
variety of human behavior; the same may be true of the principles of
ecology. If so, a new research strategy is needed that recognizes a closer
4
ADVANCES IN ARCHAEOLOGICAL METHOD Copyright ? 1980 by Academic Press, Inc. AND THEORY, VOL. 3 All rights of reproduction in any form reserved.
ISBN 0-12-003103-5
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

158 DONALD L. HARDESTY
camaraderie between ecology and archaeology and that provides an alter
native to the self-fulfilling prophesy of the ad hoc approach. That is exactly the direction proposed by a few archaeologists. Thus, Ford (1977:183) com
ments that
Anthropology has produced few, if any, lawlike statements; but the physical and
biological sciences have. If we make our problems operational in terms appropriate to
analysis of a more general theory such as synthetic evolutionary biology, we can employ those principles and free ourselves from induction and tautology.
The purpose of this chapter is to look closely at the new research strategy, to
consider its strengths and weaknesses, and to make some suggestions about
how it may be used to further our understanding of the past. First is an
illustration of how general ecological principles have been used in ar
chaeological research. The second part of this contribution turns to a crit
icism of the strategy and what future direction is needed; that direction is
likely to demand much more interaction between ecology and archaeology than is suggested by the preceding quote. Finally, the chapter suggests the
way in which general ecological principles can be used to explain the pro cesses of cultural diversification, a key problem in archaeological inter
pretation (Binford 1962).
HOW GENERAL PRINCIPLES ARE USED: AN EXAMPLE
Yellen (1977) gives an interesting example of how general ecological prin
ciples can be applied to archaeological data by using the stability-time
hypothesis to explain and predict observations of the behavior of hunters
and gatherers living in deserts. Ethnographers and archaeologists alike have
been perplexed with the social organization of aboriginal Australia, the
Kalahari desert of southern Africa, and the American Great Basin. On the
one hand, there is abundant evidence that looseness and fluidity should be
considered as principles of social organization; thus Julian Steward as
signed these principles to the Great Basin Shoshoni (1938). But on the other
hand, there is equally abundant evidence for rather rigid organizing prin
ciples, and that has impressed another group of researchers. Service (1962)
reconstructs, for example, patrilineal, exogamous bands for the precontact
Shoshoni, and Jennings and Norbeck (1955) see resistance to rapid change, and a general conservatism, as the key to understanding Great Basin pre
history. Why does the contradiction exist? The easy way out would be to
dismiss the evidence of one side or the other as being erroneous, nonrep
resentative, or as having some other methodological or interpretative weakness. Yellen, however, accepts the existence of both kinds of evidence
and argues that both flexibility and persistence are organizing principles consistent with the most advantageous strategies of desert life. His argu
ment follows from the predictions of the stability-time hypothesis.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 159
The most general use of the hypothesis is as an explanation for bio
geographical differences in species diversity and behavior; it gives an
answer to the question of why so many kinds of species live in some places and so few in others. Sanders (1969) and others who have developed the
hypothesis argue that the question is best answered by the ecological
strategies that are made advantageous, or not, by environmental stability, on the one hand, and stress, on the other. These conditions are observed as
endpoints on the continua of physical severity, such as extreme tempera tures and the magnitude of fluctuations in resources or other life oppor tunities. Yellen makes a good case for replacing the latter continuum with
the predictability of resources; that is, a stressful place to live is one in
which it is very difficult to predict when, where, and what resources will be
available, and the opposite is true for stability. Deserts are considered by Yellen to be severe and unpredictable to hunters and gatherers, and he ac
cordingly turns to that part of the stability-time hypothesis that explains and predicts response to stress.
What kind of strategy is most advantageous in an unpredictable environ
ment? Resources (or other life opportunities) change suddenly and unex
pectedly, acting against the exclusive use of one or a few of them. The use of
many resources is, however, advantageous. Such a strategy demands be
havior sufficiently flexible to rapidly shift from one to another, and
Yellen cites studies of bird communities in various parts of the world to sup
port the general proposition that behavioral plasticity and flexibility of
organisms is greater in stressful than in stable environments. The behavior
of hunters and gatherers in deserts is, then, expected to be flexible, a predic tion that is confirmed by ethnographic and archaeological data on the social
organization of such groups. But what about the contradiction? Here Yellen introduces us to Holling's
(1973) argument that resiliency is advantageous in unpredictable envi
ronments; that is, the organism is expected to roll with the punch, chang
ing enough to cope with perturbations, such as shifting behavior to make up for a suddenly failed resource but still keeping the same fundamental struc
ture and behavior. In other words, persistence is predicted, and that is
precisely how the social organization of desert-dwelling hunters and gath erers has been interpreted by some. Thus, Yellen is able to use the stability time hypothesis to shed new light on an old archaeological and anthropo
logical problem. And what more can be asked of a research strategy?
SOME LOGICAL PROBLEMS
Attempts such as Yellen's have not always met with sweetness and light, however. Schiffer (1978) argues, for example, that the use of general eco
logical principles in archaeology and anthropology is plagued with log ical weaknesses. Analogy is one use whereby parallels are drawn between a
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

160 DONALD L. HARDESTY
principle and human behavior; Gall and Saxe's (1977) application of the
principles of succession to explain the evolution of complex states is given as
an example. Here, parallels are drawn between the increasing diversity of
ecological communities undergoing succession, and their correspondingly
greater ability to "buffer" environmental stress, and the increasing diver
sity of human societies undergoing evolutionary change. Gall and Saxe con
tend that if a social system becomes better buffered against environmental
stress as it becomes more complex and diversified, then complex societies
have a selective advantage over simple societies and are bound to replace them over time. The weakness of the succession analogy, and one that can
be extended to similar analogies according to Schiffer, is that "superficially
appealing" correspondences are opposed by numerous areas of complete
dissimilarity that are either not recognized or are ignored. Thus, Gall and
Saxe pay no attention to other predictions deduced from the principles of
succession that seemingly do not apply to the evolution of the state: these
include the greater stability of climax communities, although states are
usually less stable, and the greater energy efficiency of climax communities, in apparent contrast to states.
Another use is homology. Here, general ecological principles and human
behavior are assumed to be identical in important ways, as illustrated by Osborne's (1977) use of r-selection and k-selection to explain the shift from
land-based to marine resources in Peru. The homology comes from the
assumption that the growth curves of all animals, including Homo sapiens, have the same sigmoid shape and are controlled by the same processes.
Rapid growth in the beginning selects for ecological strategies that, among other things, maximize resource use. The process is called r-selection, and
Osborne argues that the rapid growth stage of hunting and gathering
populations will be associated with r-selection strategies, typically the
specialized exploitation of a few highly productive resources such as big
game. After growth-inhibiting processes level off the growth curve, the ad
vantage shifts to ecological strategies that are more intensive and energy ef
ficient. Such k-selection strategies allow individuals to cope more effectively with a limited resource base. Osborne proposes that hunters and gatherers
began to use marine resources as part of a shift to k-selection strategies tied
to the stabilization of population size at carrying capacity. Schiffer sees Osborne's approach as a "born-again population pressure
model, with all of its theoretical and operational weaknesses, that no
amount of ecological baptism can conceal" (1978:14). In other words, the
homology, if it does exist, is trivial (we already know about population
pressure), and it obscures the real problem of making such models workable
for archaeological data. Homologues may exist, according to Schiffer, but
they must be nontrivial and workable before they will advance archaeolog ical knowledge.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 161
If analogy and homology are poor logical arguments, then, what is the
proper role of general ecological principles in archaeological explanation? One possibility that Schiffer conceives is reduction, "effected when one
discipline's principles can be deduced from the theoretical premises of
another" (1978:15-16). That implies two sets of principles that are indepen dent: one making up general ecological theory, the other making up a
theory of human behavior. Unfortunately, neither one exists, at least in
Kuhn's (1970) sense of a paradigm. Schiffer argues that logical reduction is
not yet possible because of the absence of agreed upon principles of cultural
processes, and it can also be argued that general ecological principles are in
the same state. Despite its rigorous methodology, mathematical precision, and experimentation, ecology is not much further along, if at all, than
the social sciences in discovering general laws, and competing schools of
thought vie with each other for general acceptability. The hazards of crossing disciplinary boundaries on search and seizure
missions are clearly stated in Schiffer's critique. General ecological prin
ciples are unlikely to become an important part of explanation in ar
chaeology unless more attention is^iven to the problems of making them
workable for archaeological data and to logical relationships between ecol
ogy and archaeology. The latter problem arises from two characteristics
of ecological theory. First, the theory has developed largely independent of
data on human ecology; that is, the explanatory principles and hypotheses
making up the theory have been mostly tested with data on the ecology of
species other than Homo sapiens. The reason for this is mostly historical, but the implications are crucial to the use of ecological principles in the
human sciences: it is likely that modifications in the theory will have to be
made, including the discovery of new principles, to accommodate people. That would also be true for the theory to take into account new data on the
ecology of any species; it has nothing to do with human "exceptionalism."
Accordingly, there is no reason to assume that the theory, as it now stands, is sufficiently general to cover human ecology.
Second, and already mentioned, ecological theory is in a state of flux, with many competing principles and hypotheses. Only the most foolhardy, therefore, would use one principle or another, without showing why it is a
better explanation than the others. The transfer of general ecological prin
ciples across disciplinary boundaries is, then, dependent upon the results of
multiple-hypothesis testing. Those principles that are the most general, and
probably that have been tested with the greatest variety of species, have the
best chance of being applicable. All of this suggests that it is wrong to relate ecology and archaeology as
the donor and recipient of explanatory principles; rather, the most produc tive relationship is one of interaction between a general theory and a new
data base. The relationship is new to human ecology, but it is not new to
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

162 DONALD L HARDESTY
the palaeosciences. Palaeontology, our sibling discipline, is using its dia
chronic, evolutionary data on species ecology to test and modify general
ecological models based upon synchronie and experimental data (e.g., Gould 1977; Raup 1977; Raup et al 1973; Van Valen 1965, 1973). The ad
vantage of diachronic data is also held by archaeology. Notwithstanding the
incompleteness of their data, it appears that the palaeosciences have the best
potential for serving as the "evolutionary" arm of general ecology. Such a
role is consistent with Plog's (1974) argument that the goals of archaeology should be limited to evolutionary problems.
The purpose of the last part of this paper is to illustrate what is involved
in using general ecological principles and archaeological data interactively.
Competing models of species diversity are tested with archaeological data as
possible explanations for evolutionary patterns of cultural diversity. The
first section discusses how such models can be made workable for ar
chaeological data; the second outlines the test implications of the various
models considered and discusses how the test results can be used to modify
general ecological theory.
THE ECOLOGICAL EXPLANATION OF CULTURAL DIVERSITY
The explication of the processes of cultural diversification is one of the
most important general problems in archaeology and anthropology (Bin ford 1962). Why are there more kinds of cultures in some places and in
some periods of time than in others? The same question is asked by
ecologists but is aimed more generally?thus the title of Hutchinson's
(1959) classic paper "Homage to Santa Rosalia, or Why are There so Many Kinds of Animals?" Models of species diversity have been devised to
answer Hutchinson's question, and here we are interested in whether these
models can shed light on cultural diversity and in whether data on cultural
diversity can be used to improve the generality of general ecological theory. The approach is opposite, and is an alternative, to the argument that
cultural "diversity and (the) processes of diversification are not explicable in terms of biological process" (Binford 1962:218).
Why should explanations of species diversity have anything at all to do
with the principles of cultural diversity? Such a question follows from the
larger problem of why explanations of variability among species should
have any meaning for our understanding of variability within a single
species. At least part of the answer lies in what ecological theory seeks to ex
plain: differences among organisms that have to do with how they cope with
environmental problems. Biological differences, such as those defining
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 163
species, are important only in that they provide a way of keeping genetic in
structions for environmental problem-solving reasonably intact. But if
problem solving of that kind mostly originates from learned behavior, and
only incidentally from genetic instructions, differences in cultural and other
rules of behavior gain priority in ecological explanation; the biological
species concept is no longer appropriate. Cultural rules are propagated in
social groups, and social groups thus become equivalent to the species for
purposes of ecological analysis. Because of the flexibility of learned
behavior, social groups with essentially the same genetic instructions can
devise very different rules for solving environmental problems. For that
reason, the ecological contrasts among neighboring social groups of the same species are often as great, if not greater, than the contrasts among
coexisting species. That is true not only for Homo sapiens but also for other
higher mammals (e.g., Kummer 1971). Therefore, because it is the ecolog ical differences that are important to ecological theory, there is no reason
why models of species diversity should not be applicable to intraspecific
diversity, provided that significant differences do exist in the ecology of
subspecific social groups.
Making the Models Workable for Archaeological Data
Before turning to the models of species diversity, we must consider how
they can be tested with archaeological data. What kind of information is
needed? First of all, the intraspecific equivalent of the species must be de
fined, and the means by which it can be recognized must be identified;
otherwise, there is no way of measuring "diversity." The ecological mean
ing of the species concept is a significant difference in lifestyle, as discussed
in the preceding paragraph. That implies that intraspecific differences in
lifestyle about the same as that separating species can be used to define the
species equivalent. How much difference is that? The answer lies, perhaps, in the same methods that some evolutionary taxonomists have used to
decide whether a local population belongs to one subspecies or another
(namely Mayr 1963); that is, rules have been devised to do the separation, such as two populations belong to the same subspecies if 95% of their
members hold 95% of their biological features in common. For our pur
poses, the separation rule can be stated as the allowable difference in the
behavior of the social groups being compared. Archaeological manifesta
tions of that behavior will be in the form of artifacts and other material re
mains; therefore, the differences among human groups that define species
equivalents are measured by the similarity in artifact assemblages. That is, of course, a method that is familiar to most archaeologists. The question
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

164 DONALD L HARDESTY
that has to be answered is whether the 95% rule should be used or whether some other level of similarity is best. Once that question has been answered, the recognition of "cultural species" with archaeological data is straightfor
ward.
The measurement of cultural diversity can then be made comparable to
that of species diversity. Ecologists use the concept of diversity in three
ways (Pielou 1975; Whittaker 1977):
1. Species richness?the number of species present 2. Species evenness or equitability?the relative importance of species
present 3. Total species diversity?measures that combine species richness and
evenness
Species richness is the most simple measure, requiring only that the number
of species in a standard sample be counted, and seems to be the most effec
tive measure of diversity (Whittaker 1977:2-4). "Standard sample" is, of
course, a key variable. The number of cultural species is partly a function of
the size of the geographical area and the length of the time period in which
the count was made; accordingly, scale must be considered. But what is the
appropriate scale to study cultural diversity? The answer to that question is
tied to the kind of explanatory model being used, among other things. MacArthur's (1972) theory of island biogeography, for example, argues
that scale in itself is an important cause of differences in species diversity. The other kind of information needed to test most models of species
diversity is suggested by their similar underlying logic, using the following
key assumptions (Hutchinson 1965; Diamond 1978; Schoener 1974; Whit
taker and Levin 1975):
1. Resources or other life opportunities in the environment are finite. 2. These finite resources subpartitioned among species sharing the en
vironment; the ecological niche defines what share a species re
ceives.
3. The ecological niche is occupied more or less exclusively.
Therefore, if the environment is partitioned into large shares, and the share
of each species is exclusive, it is reasonable to argue that species diversity is
less than if smaller shares were available. Questions such as how wide
(large) are the niches of single species and how far apart are the niches of
two species, or how much do they overlap, are, for that reason, especially
important to the understanding of species diversity. That is agreed upon. The models disagree, however, over the cause of partitioning and niche dif
ferentiation.
Because the niche concept is an important part of most models of species
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions
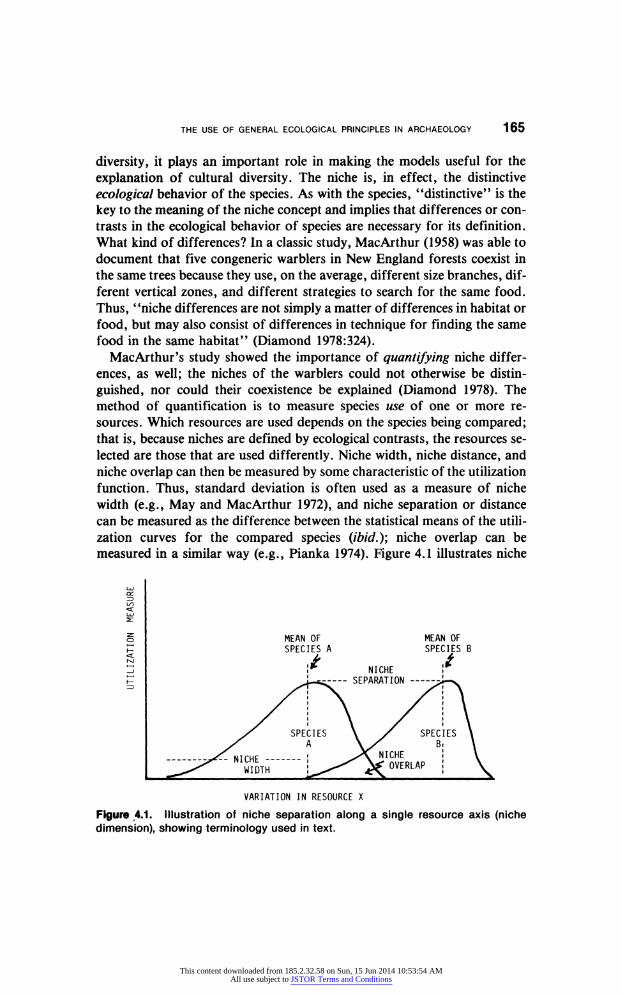
THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 165
diversity, it plays an important role in making the models useful for the
explanation of cultural diversity. The niche is, in effect, the distinctive
ecological behavior of the species. As with the species, "distinctive" is the
key to the meaning of the niche concept and implies that differences or con
trasts in the ecological behavior of species are necessary for its definition.
What kind of differences? In a classic study, MacArthur (1958) was able to
document that five congeneric warblers in New England forests coexist in
the same trees because they use, on the average, different size branches, dif
ferent vertical zones, and different strategies to search for the same food.
Thus, "niche differences are not simply a matter of differences in habitat or
food, but may also consist of differences in technique for finding the same
food in the same habitat" (Diamond 1978:324). MacArthur's study showed the importance of quantifying niche differ
ences, as well; the niches of the warblers could not otherwise be distin
guished, nor could their coexistence be explained (Diamond 1978). The
method of quantification is to measure species use of one or more re
sources. Which resources are used depends on the species being compared; that is, because niches are defined by ecological contrasts, the resources se
lected are those that are used differently. Niche width, niche distance, and
niche overlap can then be measured by some characteristic of the utilization
function. Thus, standard deviation is often used as a measure of niche
width (e.g., May and MacArthur 1972), and niche separation or distance
can be measured as the difference between the statistical means of the utili
zation curves for the compared species (ibid.); niche overlap can be
measured in a similar way (e.g., Pianka 1974). Figure 4.1 illustrates niche
MEAN OF SPECIES B
i
VARIATION IN RESOURCE X
Figure 4.1. Illustration of niche separation along a single resource axis (niche
dimension), showing terminology used in text.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

166 DONALD L HARDESTY
definition with resource utilization curves. In practice, of course, a single en
vironmental variable (niche dimension or axis) is unlikely to give complete information about the niche differences that exist among species; several
niche dimensions are better indicators. Together, they define what Hutchin
son (1965) calls a niche hypervolume or hyperspace, the total ecological life
style. The measurement of the niches of cultural species can, then, be based
upon two kinds of data:
1. Data that show differences in the way that resources or other life op
portunities limited by the environment are utilized.
2. Data that show differences in how resources are acquired. Hardesty
(1975) illustrates how the niche width of cultural species can be measured with data on resource use, taking advantage of a method devised by Levins
(1968). The method uses an index of resource diversity n
niche width = 1 /L? (Pi)2 i where p? is the proportion of the total subsistence of a species contributed
by resource /, and n is the total number of resources used for subsistence.
Table 4.1 illustrates the method with subsistence data on the Mistassini Cree
TABLE 4.1
Niche Width Calculations from Total Resource Variety for Mistassini Cree, Canada3
Resource Biomass(lb) p? (p?)2
Moose 4000 .448 .202 Caribou 1500 .168 .029
Bear 210 .024 .000 Beaver 2120 .237 .058
Hare 114 .013 .000 Muskrat 240 .027 .001
Porcupine 60 .007 .000 Mink 33 .004 .000
Squirrel 8 .000 .000 Marten 5 .000 .000
Otter 110 .012 .000 Loon 44 .005 .000
Geese 67 .008 .000 Ducks 231 .026 .001
Ptarmigan 150 .017 .000 Spruce grouse 38 .004 .000 Ruffled grouse 1 .000 .000
Owl 1 .000 .000
?(p,)2 = .291
Niche width = 1/0.291 = 3.436
Data from Rogers (1972:104, Table 3.1). Source: Donald L. Hardesty, "The Niche Concept: Some Suggestions for Its Use in Human
Ecology." Human Ecology 3, 1975. Copyright ? 1975 by Plenum Publishing Corporation.
Reprinted by permission of the publisher.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions
THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 167
of Canada given by Rogers (1972:104, Table 3.1). The method assumes that
resource diversity is a fair measure of how the environment of neighboring
groups is being partitioned. Niche overlap among cultural species can be
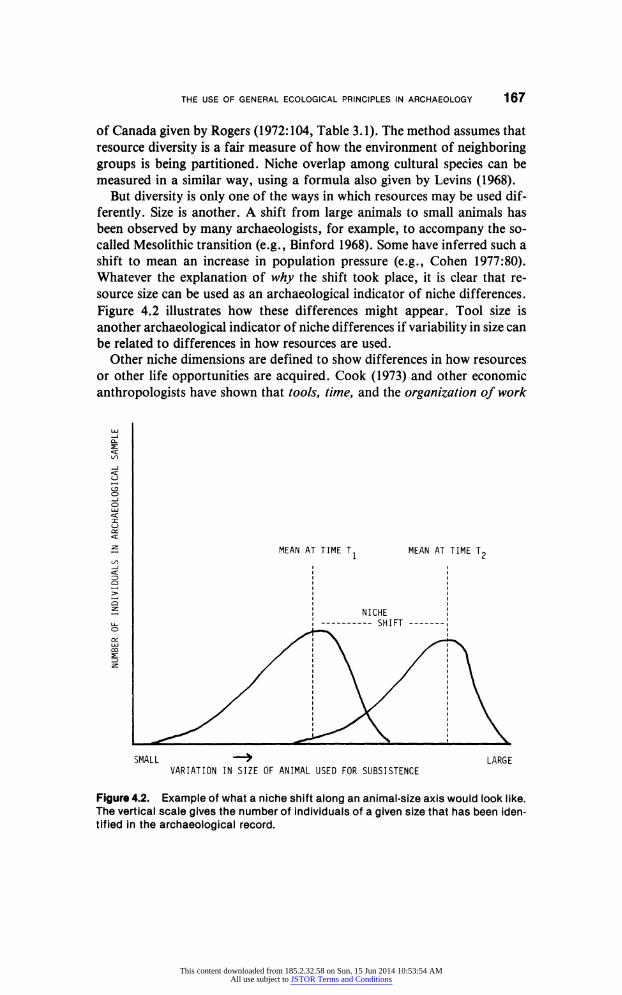
measured in a similar way, using a formula also given by Levins (1968). But diversity is only one of the ways in which resources may be used dif
ferently. Size is another. A shift from large animals to small animals has
been observed by many archaeologists, for example, to accompany the so
called Mesolithic transition (e.g., Binford 1968). Some have inferred such a
shift to mean an increase in population pressure (e.g., Cohen 1977:80). Whatever the explanation of why the shift took place, it is clear that re
source size can be used as an archaeological indicator of niche differences.
Figure 4.2 illustrates how these differences might appear. Tool size is
another archaeological indicator of niche differences if variability in size can
be related to differences in how resources are used.
Other niche dimensions are defined to show differences in how resources
or other life opportunities are acquired. Cook (1973) and other economic
anthropologists have shown that tools, time, and the organization of work
MEAN AT TIME T0
SMALL VARIATION IN SIZE OF ANIMAL USED FOR SUBSISTENCE
LARGE
Figure 4.2. Example of what a niche shift along an animal-size axis would look like.
The vertical scale gives the number of individuals of a given size that has been iden
tified in the archaeological record.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions
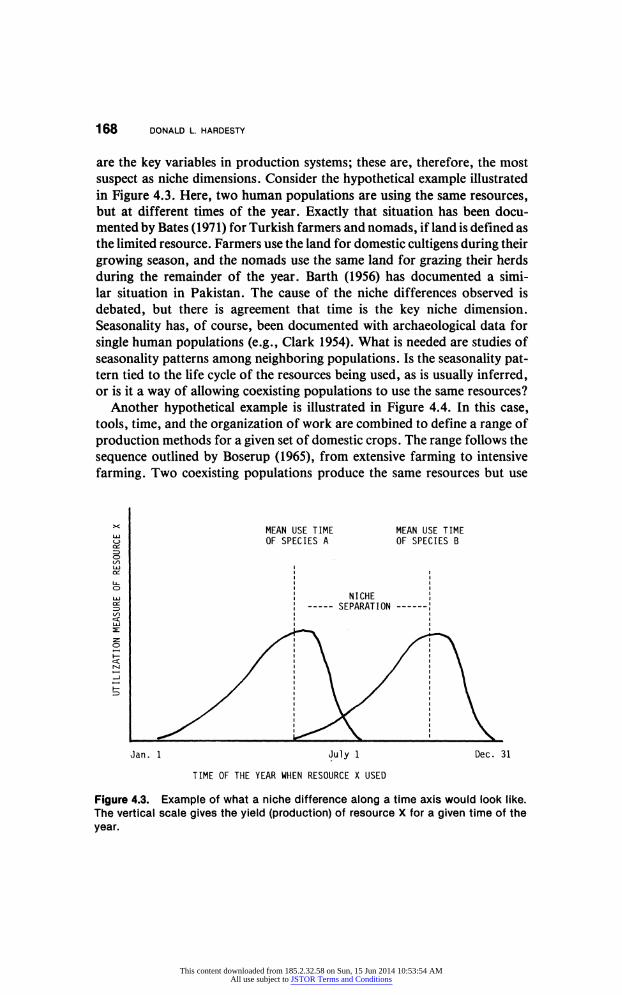
168 DONALD L. HARDESTY
are the key variables in production systems; these are, therefore, the most
suspect as niche dimensions. Consider the hypothetical example illustrated
in Figure 4.3. Here, two human populations are using the same resources, but at different times of the year. Exactly that situation has been docu
mented by Bates (1971) for Turkish farmers and nomads, if land is defined as
the limited resource. Farmers use the land for domestic cultigens during their
growing season, and the nomads use the same land for grazing their herds
during the remainder of the year. Barth (1956) has documented a simi
lar situation in Pakistan. The cause of the niche differences observed is
debated, but there is agreement that time is the key niche dimension.
Seasonality has, of course, been documented with archaeological data for
single human populations (e.g., Clark 1954). What is needed are studies of
seasonality patterns among neighboring populations. Is the seasonality pat tern tied to the life cycle of the resources being used, as is usually inferred, or is it a way of allowing coexisting populations to use the same resources?
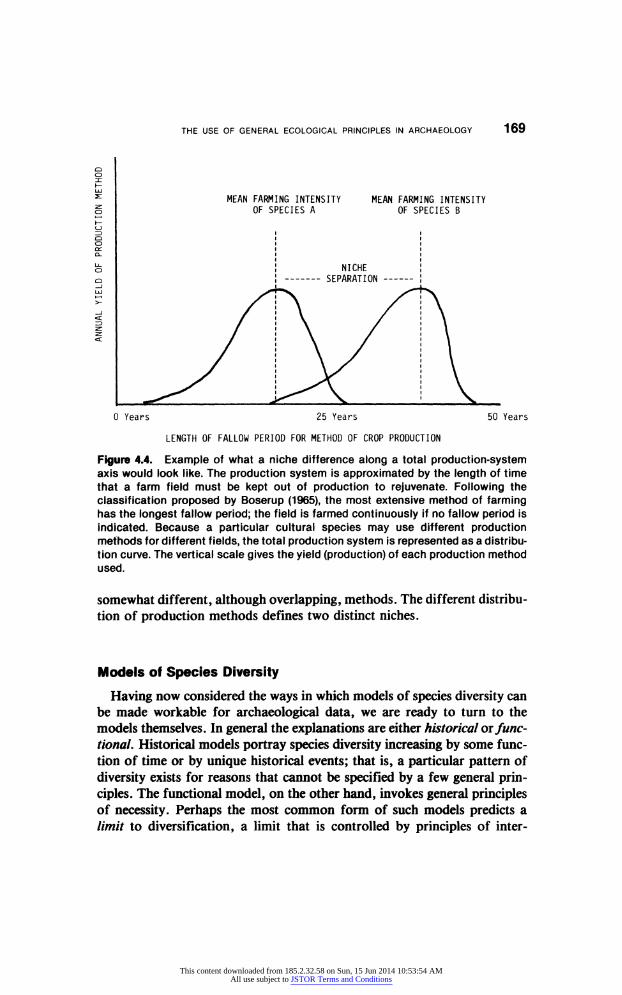
Another hypothetical example is illustrated in Figure 4.4. In this case,
tools, time, and the organization of work are combined to define a range of
production methods for a given set of domestic crops. The range follows the
sequence outlined by Boserup (1965), from extensive farming to intensive
farming. Two coexisting populations produce the same resources but use
MEAN USE TIME OF SPECIES A
MEAN USE TIME OF SPECIES B
Jan. 1 July 1
TIME OF THE YEAR WHEN RESOURCE X USED
Dec. 31
Figure 4.3. Example of what a niche difference along a time axis would look like.
The vertical scale gives the yield (production) of resource X for a given time of the
year.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions
THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 169
MEAN FARMING INTENSITY OF SPECIES A
MEAN FARMING INTENSITY OF SPECIES B
0 Years 25 Years
LENGTH OF FALLOW PERIOD FOR METHOD OF CROP PRODUCTION
50 Years
Figure 4.4. Example of what a niche difference along a total production-system axis would look like. The production system is approximated by the length of time that a farm field must be kept out of production to rejuvenate. Following the classification proposed by Boserup (1965), the most extensive method of farming has the longest fallow period; the field is farmed continuously if no fallow period is indicated. Because a particular cultural species may use different production
methods for different fields, the total production system is represented as a distribu tion curve. The vertical scale gives the yield (production) of each production method used.
somewhat different, although overlapping, methods. The different distribu
tion of production methods defines two distinct niches.
Models of Species Diversity
Having now considered the ways in which models of species diversity can
be made workable for archaeological data, we are ready to turn to the
models themselves. In general the explanations are either historical or func tional. Historical models portray species diversity increasing by some func
tion of time or by unique historical events; that is, a particular pattern of
diversity exists for reasons that cannot be specified by a few general prin
ciples. The functional model, on the other hand, invokes general principles of necessity. Perhaps the most common form of such models predicts a
limit to diversification, a limit that is controlled by principles of inter
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

170 DONALD L HARDESTY
specific competition, pr?dation, or physical stress. Some models have, of
course, elements of both historical and functional explanation; the stability time hypothesis is an example.
The Stability-Time Hypothesis Yellen's paper (1977), discussed in an earlier section, clearly shows how
the stability-time hypothesis can be used to explain and predict some aspects of the behavior of hunters and gatherers; it is also an explanation for species
diversity. The hypothesis states that species diversity increases continuously over time but that environmental stress counteracts the trend (Bretsky and
Lorenz 1970; Sanders 1969). There are at least two test implications. The
first is that older ecosystems and communities should be more diverse than
younger ones, given the same amount of stress; the second is that, given
ecosystems and communities of the same age, those under the least stress
will be the most diverse. Unfortunately, neither of the implications is easily observed: the age of ecosystems and communities is difficult to ascertain, and environmental stress cannot be easily measured. Indeed, Sanders uses
physiological stress as a measure because its effects can be observed on the
organism; but, because cause and effect are thereby not separated, a cir
cular argument ensues. These problems with testing the stability-time hy
pothesis have led to the accusation that it is a tautology (Peters 1976:9-10).
Why, then, should it be used as an explanation of cultural diversity? The reasons are twofold. In the first place, the archaeological evidence of hu
man populations making up an ecological community is often dateable,
making it possible to compare communities of more or less the same age.
Second, if environmental stress is equated with the predictability of re
sources or other life opportunities, as Yellen does, then circular reasoning is avoided; that is, stress, as environmental predictability, can be observed
and measured independently of its supposed effects. Here is a good example of how archaeological data on human ecology can be used to test a model of
species diversity that is otherwise plagued with logical weaknesses.
A few additional comments about the predictability of the environment
are in order before going further; it cannot be measured separately from the
organism being studied. The same environment may be very predictable for
some and completely unpredictable for others, depending upon how re
sources or other life opportunities in the environment are being used.
Technology and social organization are, therefore, essential parts of any at
tempt to measure environmental predictability for human populations. Both have an impact upon the meaning of an environment for a population, not only because they define the means of production but also because
they can modify the predictability of the environment. Technology makes
changes by controlling and redistributing water (irrigation technology), by
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 171
weather modification, by artificial fertilization, and the like. Social or
ganization makes changes through economic networks that involve the
transportation of resources from one place to another and from one time to
another. Thus, the desert habitat that Yellen found to be so unpredictable for hunters and gatherers is considerably more predictable for modern
irrigation farmers. There is, therefore, reason to doubt that "external"
measures of environmental predictability, such as rainfall variability (e.g.,
Baerreis, Bryson, and Kutzbach 1976; Jorder 1977; Reher 1977), will be
of much value to general explanations. The best measure is probably
something that considers the simultaneous effects of climate, technology, social organization, and the like. Perhaps something equivalent to the an
nual yield of production systems would work: a highly variable yield sug
gests an unpredictable environment, and a sustained yield suggests a pre dictable one. That measure is, of course, difficult to make workable for
archaeological data, but suitable material indicators of variability in yield are possible.
Now let us return to the test implications of the stability-time hypothesis for cultural diversity. There are two kinds of predictions: comparative
predictions and evolutionary predictions. Comparative predictions are the
same as those stated in the first paragraph of this section; that is, they hold
constant the age and the stress of ecological communities made up of
cultural species and compare cultural diversity. But it is an evolutionary
prediction that may be of most interest. The stability-time hypothesis leads to the prediction that evolutionary patterns of cultural diversity in
an ecological community will reflect environmental predictability and its
changes over time. Consider an environment that is initially highly predict able to a cultural species. Diversification continues over time and produces a curve of continually increasing cultural diversity. But if episodes of en
vironmental stress shock the evolving community of species, diversity would either fall drastically or level off, depending upon the severity of the
shock. Holling (1973) argues, for example, that the diverse, equilibrium communities usually found in stable environments are most susceptible to an unexpected perturbation; the result is often a catastrophic change in the
community, causing a drastic reduction in species diversity. Because of this, Gould (1977:222) suggests that the "ultimate control (of diversity in equilib rium communities) is exerted by the very rare major fluctuation (of the envi
ronment)." If so, the expected shape of the diversification curve shows
rapid growth interspersed with periods of sharp declines or plateaus. The stability-time hypothesis predicts, on the other hand, that in un
predictable environments, cultural diversity is kept at or below a limit set by the necessity of having wide niches. That is, a reasonably large number of resources or other life opportunities must be available to each species to
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

172 DONALD L. HARDESTY
minimize the risk of an unexpected loss of one resource or another; there
fore, wide niches are demanded and, because of that, few species are possi ble. The limiting similarity models, to be discussed later, give some explana tions of exactly what the functional limits on diversity are. Notwithstanding the theoretical limit, however, the evolutionary pattern of cultural diversifi
cation may show peaks or episodes of rapid proliferation corresponding to
periods of increased predictability of the environment; such peaks will, of
course, be squashed by a return to normal conditions.
Limiting Similarity Models
Perhaps the most provocative of the functional explanations propose that
species diversity is limited by niche similarity; that is, the ecological lifestyle of coexisting species can become only so much alike before the relationship is disrupted. Why? Two explanations are currently popular. One argues that niche similarity is partly limited by environmental variability but that a
limit exists independently of moderate to small fluctuations; the other sees
niche similarity limited by competition among species.
Variability in the Environment The first kind of explanation is il
lustrated by a model proposed by Robert May and Robert MacArthur (May and MacArthur 1972; May 1974, 1976; see also Nisbet et al. 1978). Based
upon mathematical studies, the May-Mac Arthur model predicts that under
the conditions of a constant, deterministic environment, there is no limit to
how closely together neighboring niches may be packed; therefore, there is
no theoretical limit on species diversity. But if the environment is allowed to
fluctuate randomly, a different situation prevails. May and MacArthur
(1972:1109) initially conjectured that "in the real world, environmental
fluctuations will put a limit to the closeness of species packing compatible with an enduring community, and that species will be packed closer or wider
as the environmental variations are smaller or larger." That was found to
be true if the fluctuations were severe. Species packing is about propor tional to the amount of variance, as would also be expected by the stability time hypothesis. But if the random fluctuations are kept within the range from 30 to .01% of the mean, then "the closest species packing consistent
with stability falls only from 2 to 1 times the utilization function variance"
(ibid.); that is, the model predicts a definite limit on species diversity. May and MacArthur interpret such a limit to mean that environmental variabil
ity is not an important cause of species packing unless it is severe. If true, the model contradicts the stability-time hypothesis.
Species packing in the May-MacArthur model is measured by niche
separation. Using a single niche dimension, food size, the model portrays the distance between neighboring niches as the difference in the way that
food size is used. The actual measure is d/w, where d is the difference be
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 173
tween the means of the compared utilization curves and w is 1 standard
deviation. Thus, the predicted limit, given above, is d/w = 1 or 2.
The test implications of the May-MacArthur model for patterns of
cultural diversity are similar to those of the stability-time hypothesis, but
more precise predictions are possible. Once again environmental fluctu
ations are to be equated with the variability in an index of production yield or something similar; that is, predictability in effective environment is
measured. In very unpredictable environments, cultural diversity is ex
pected to be more or less proportional to the amount of uncertainty. And in
very predictable environments, cultural diversity is expected to have no
limit; that implies that it will increase continuously over time. Both of these
predictions are consistent with those of the stability-time hypothesis. But in
most environments, those with only moderate stochasticism, cultural diver
sity is expected to reach and maintain a limit. That suggests that any com
munity of cultural species with moderate to small fluctuations in production
yield, whether foragers, farmers, or industrialists, should have approxi
mately the same diversity if niche dimensions are equivalent. And in the
same way, the evolution of a community of cultural species in the same kind
of environment should increase to a limit and then level off; that is, diver
sification should not be continuous over time, as implied by the stability time hypothesis.
The May-MacArthur model further predicts that the limit is approx
imately d/w = 1 or 2, and archaeological data can be used to test this more
specific expectation. Niche dimensions that best separate cultural species must first be defined; then, utilization curves can be plotted and niche widths and distances measured. Again, recall that the best niche dimensions to use are limited resources or other life opportunities that are used or pro duced differently. The dimensions need not be natural resources: indeed, such "created" resources as art objects could probably be used to define the niches of social classes in complex societies; certainly there is a difference in use (see Hardesty 1978, 1979 for a further discussion of niche dimensions in human ecology).
Competition. The May-MacArthur model is supported extensively,
although not exclusively, by mathematical evidence, but by little observable
evidence (May 1976; Nisbet etal 1978). Furthermore, several of the assump tions used in the model, such as a saturated environment, resources equally available to interacting species, and no significant difference in species
abundance, may not be realistic in many situations (Whittaker and Levin
1975:176-177). Thus, Pianka (1974:2141) notes that the May-MacArthur model "assumes an equilibrium community in a fully saturated environ
ment with all resources being used fully; as such, variation in the intensity of competition is not modelled." What effect does competition have upon
diversity? Pianka argues that if environment is not limiting, for whatever
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

174 DONALD L HARDESTY
reason, there is absolutely no reason why the same niche should not be oc
cupied by different species. Thus, Connell (1975) documents that competi
tion, forcing niche exclusion, is virtually absent under heavy pr?dation or
environmental stress; population size is kept low so that the supply of resources doesn't exceed demand. Nor is the environment limiting under
conditions of superabundant resources, and Weins (1977:591-592) sees
competition as unimportant under such conditions. In general, if the supply of resources or other life opportunities exceeds demand, it can be assumed
that competition has little impact upon niche differentiation and, therefore,
species diversity.
But, as environment becomes more limiting, that is, as demand exceeds
supply, competition intensifies. What are the conditions under which that is
likely to happen? Environmental predictability seems to be important, for
two reasons: (1) competition models assume an equilibrium community, which are found in predictable environments, and (2) continuous competi tion is assumed by competition models, and that is most likely in predictable environments (Weins 1977:591). Whatever the cause, however, intensified
competition is likely to make the principle of competitive exclusion (Hardin
1960) more effective and, consequently, to limit niche similarity. A limit is
thereby placed upon species diversity.
Testing Pianka's "niche overlap hypothesis," as he calls it, depends upon some way of recognizing and measuring competition. Diamond (1978: 325-328) considers niche shifts to be the best evidence of competition, in
cluding the following:
1. Niche differences among neighboring species that are very similar in
other ways 2. Niche differences in otherwise identical species that occupy environ
ments with and without other similar species 3. Diachronie changes in the niche of the same species after it has
moved from an environment with potential competitors to one with
out, or vice versa
Notwithstanding arguments that such niche shifts could be explained by yet unknown limiting factors and the like (e.g., Connell 1975; Weins 1977), the
evidence seems to favor competition as the best explanation. But the obser
vation of niche shifts per se cannot be used to measure competition without
getting into the same kind of circular reasoning that weakened the stability time hypothesis; that is, the cause is not empirically separated from the ef
fect. One way of separating the two is to follow MacArthur's (1972) con
cept of diffuse competition, also used in Pianka's model. MacArthur and
Pianka propose that weak competition from many sources is additive and
has the same effect as more intense competition from a few sources; for that
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 175
reason, the number of competitors can be viewed as a measure of the inten
sity of competition. For our purposes, then, the intensity of competition is
measured by population size or population density, and that provides a
source of empirical data that is independent of data used to measure niche
shifts.
Now we can consider the implications of the niche overlap model for pat terns of cultural diversity. The limit on diversity is set by the intensity of
competition; therefore, a test of the model requires that population size or
density or another measure of competition be plotted against the average niche overlap of cultural species, either using comparative or evolutionary data. If the model is correct, the ensuing curve should have a definite upper
plateau. Care must be taken, however, to be sure that environmental vari
ability is controlled for the test; otherwise, the effects of competition and
variability cannot be separated. Probably the best method is to compare the
competition curves in environments with different degrees of predictability. The curves should be similar if the Pianka model is correct.
Equilibrium Theory of Island Biogeography
Perhaps the most influential functional model of species diversity to
emerge in recent years is the equilibrium theory of island biogeography pro
posed by Robert MacArthur (MacArthur and Wilson 1967; MacArthur
1972; Simberloff 1974). The model portrays islands or other isolated
patches being packed with species up to a predictable limit determined by the size of the island and its distance from a colonizing source?a mainland
in the case of islands. That limit is reached at the intersection of two
mathematical functions: the rate of immigration of new species from the
colonizing source, and the rate of extinction of species already on the
island. When a vacant island is first colonized, the rate of immigration is
relatively high. Almost all colonists are representatives of new species. With
continual migration, however, the chance that a new arrival is a new species decreases; the rate of immigration correspondingly decreases. The immigra tion rate reaches zero when all species from the colonizing source have ar
rived. Mathematically, then, the immigration rate is some inverse function
of species diversity. By contrast, the island extinction rate is an increasing function of species diversity because of the following: (1) there are more
species to become extinct as diversity increases, and (2) the intensity of com
petition, pr?dation, and the like increase with diversification. The falling
immigration rate curve eventually intersects with the rising extinction rate
curve: the intersection point is the predicted level of species diversity on the
island. Intersection points, the equilibria of species diversity, are not the same from one island to another because of the effect of distance and island size. Immigration rates are highest when an island is close to its colonizing
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

176 DONALD L HARDESTY
source and lowest when it is far away. Extinction rates are highest on a
small island, principally because random population fluctuations are more
pronounced, and lowest on a large island; accordingly, diversity levels, or
equilibria, are highest on large islands close to a source of colonization.
Conversely, the equilibria are lowest on small islands that are far away. Some promising applications of MacArthur's theory of island biogeog
raphy to archaeological data have been made in Oceania (see Clark and Ter
rell 1978). Kaplan (1976) has, for example, nicely illustrated its usefulness
for understanding cultural and biological processes on the small "stepping stone" islands of Nissan and Pinipel in Papua, New Guinea. Both are
situated, along with the neighboring Feni Islands, between New Ireland, a
large island in the Bismarck Archipelago, and Buka, in the northern Solo
mons. Without the stepping stones, New Ireland and Buka would be sepa rated by more than 100 miles of open water. Did Nissan and its neighbors
play a role in creating the ethnographic similarities and striking differences
in race, language, and culture between the two? MacArthur and Wilson
(1967) suggest that stepping stones are important to species dispersal in two
ways: (1) they shorten distances that must be traveled at any one time and,
therefore, are the most likely routes of dispersal, and (2) they regulate the
frequency and the rate of species movement between larger land masses.
Both of these hypotheses share a fundamental assumption with "gravity" models in geography (Chorley and Haggett 1967:559), namely that the like
lihood of attraction between two social groups or geographical areas is a
direct function of size or mass and an inverse function of distance. For
species dispersal, the attraction of a small stepping stone is greatly reduced
because of intensified competition among colonists. The result is that only a
few species from a source area successfully make the leap, making the step
ping stone an effective filter.
Using this perspective, Kaplan (p.78) defines two research goals, as
follows:
1. To what extent has the presence of Nissan as an island intermediate between New
Ireland and Buka served to increase the likelihood that trade goods, ideas, and people
might travel between these two regions?
2. In what ways has Nissan acted as a "filter," or regulator, influencing the transmis
sion, or "dispersal," of trade goods, ideas, and people between the Solomons and
New Ireland?
Theoretical, ethnographic, and archaeological approaches are used to
answer the questions. Terrell (1974) built an "interaction" model of the
region showing the most likely trade routes among the islands if the
distance-mass assumptions of the biogeography and gravity models are
realistic. Kaplan then compares that theoretical model with actual trade
routes reconstructed from ethnographic data; the fit is quite close. The
stepping-stone model is, then, a good predictor of human interaction in this
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 177
part of the Pacific, suggesting that short hops rather than long-distance movement best explains trading and communication routes.
Directly to the point of Kaplan's question, both the stepping-stone model
of island biogeography and ethnographic data support trade between Buka
and New Ireland via Nissan, Pinipel, and the Feni Islands. Furthermore, an
analysis of ceramics from archaeological sites on Nissan shows an unbroken
stylistic sequence from ad 500 to historic times, styles that are virtually the
same as those from Buka. One sherd is stylistically foreign and suggests trade with other islands. Kaplan's first question can, then, be answered:
"There is some indication that known historic patterns of interaction be
tween New Ireland and the Solomons via Nissan developed during the
prehistoric past over a period of more than a thousand years" (Kaplan
1976:85). The second question is partly answered with ethnographic data showing
that Nissan did indeed act as a filter. Kaplan (p. 82) notes, for example, that
Buka cookware, a breakable commodity, was not usually traded beyond
Nissan; most of it was consumed on that island. Archaeological data have
not yet been brought to bear upon the question, but the implications for ar
chaeological research in the future are clear.
That theories of island biogeography have been used to explain human
ecology on real islands is not surprising. We should not, however, overlook
the possibility of their usefulness for understanding the human ecology of
conceptual islands: land-bound geographical regions that are isolated from
other regions in much the same way that real islands are. Perhaps the
American Great Basin and the Mexican highlands, both topographically and otherwise divided into small basins, would be ideal. Each basin can be
modeled as an island. The entire region would then consist of a cluster of
islands of different sizes and distances from one another; race, language, and cultural diversity could then be predicted from the calculated equilibria for each island. For the purposes of this chapter, the diversity of cultural
species could also be predicted from the equilibria. How can archaeological data be used to test the accuracy of the equilib
rium model of island biogeography? May (1978: 163-164) lists three kinds
of data that are needed, and they can be made workable for data on the di
versity of cultural species. The first is data on changes in cultural diversity that take place over time on islands where an equilibrium is suspected to ex
ist; if the model is correct, the changes should be best explained as random
fluctuations. Second, the model can be tested with data on changes in
cultural diversity on islands that were vacant or depopulated and that subse
quently refilled. The evolutionary pattern expected over time is rapid diver
sification in the beginning, followed by a leveling off as the equilibrium is
reached. Archaeological data are, of course, most appropriate for this kind
of test. Finally, data on evolutionary changes in the diversity of cultural
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

178 DONALD L. HARDESTY
species are needed on islands that are suspected to have diversities well
above the equilibrium because of unique invasions, forced colonization, and the like; the diversity level should drop back to the predicted
equilibrium.
Stochastic Models
The models that have been discussed so far are deterministic; that is, the
underlying cause of species diversity, cultural or otherwise, is mostly com
petition, environmental stress, or some other controlling process. That
assumption is not, however, shared by all. Some see causation as the result
of a very large number of interacting processes, no one of which has con
trol; the cause of diversity is, therefore, best understood as the sum total of
the unique histories of particular species (Simberloff 1978; see also Raup et
al. 1973:526). And because of its complexity, the cause of diversity from
one situation to another is unlikely to be the same.
Such an interpretation does not mean that the study of patterns of species
diversity is fruitless. The cause of each coin tossed landing heads or tails, for example, is too complex to specify, depending upon a multitude of fac
tors: wind velocity, wind direction, balance of the coin, direction of the
toss, and so forth (this example is given in Raup et al. 1973:526). Yet we
know that the pattern in the long run can be predicted to be 50% heads and
50% tails. Why? Events with complex, multivariate causes behave as if they were random variables, and random variables have statistically predictable
patterns of behavior. For example, we know from the "central limit theo
rem" of probability that the sum of many independent random variables
will have a very nearly normal distribution (MacArthur 1972:42). The ex
pected pattern of events with very complex causes can, then, be deduced
from a probabilistic model.
That is an approach presently being taken by some paleontologists and
biogeographers to see if observed patterns of species diversity can be ex
plained by the null hypothesis of randomness (e.g., Boucot 1976, 1978; Gould 1977; Kitts 1975; Levandowsky and White 1977; Osman and Whit
latch 1978; Raup 1977; Raup et al. 1973, 1975, 1976; Simberloff 1978). In a
classic study of the effects of randomness on diversity, for example, Raup et
al. (1973) set out to see if patterns of "phyletic diversification in the fossil
record" (p. 525) could be explained by a model based upon the following
assumptions: (1) every geographical region has a limit on the number of
taxa that can coexist, and (2) after that limit, or equilibrium, has been
reached, changes in diversity over time can be predicted by stochastic pro cesses. That is, the causes of the extinction or origins of each taxon are com
plex, in effect unknowable, but the overall pattern of extinctions and
originations can be simulated with a random walk (ibid., p. 526). And that
simulation is exactly what the authors do.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 179
A computer program was written to see what would happen if a single
lineage were subjected to stochastic processes over time. At each successive
time period, the simulated lineage met one of three fates: it became extinct, it branched to produce a second lineage, or it was allowed to persist without
branching. Random numbers automatically generated by the computer chose the fate. The simulation was then compared to the actual fossil record
of several regions. Many similarities are apparent, so that stochasticism
could not be eliminated as a cause of evolutionary patterns of phyletic
diversity, but neither could the possibility of deterministic causes be dis
counted; some periods of rapid extinction are, for example, definitely not
best explained by random walk and suggest that some deterministic process was responsible.
The probabilistic model is not really an explanation for species diversity because it does not specify a cause; rather, it is a null hypothesis that must
be rejected before deterministic explanations can be retained. For that
reason, simulations of what evolutionary patterns of cultural diversity look
like under stochastic conditions are badly needed. The computer program
logic could be very similar to that used by Raup et al. Start with a single cultural species colonizing an uninhabited area, and assign to the species a
set of cultural rules for making an artifact in a particular style. Begin the
species on a journey through time, divided into periods of, say, 100 years, each of which commences with a fate that is randomly chosen by computer.
(Recall that the fate of the cultural species is assumed by the model to have
complex, specific causes; the actual distribution of fates over time can be,
therefore, best described as a random walk.) What fates are possible? The
species can become extinct without disseminating its unique artifact style; the species can persist without disseminating its artifact style; the species can fission into two, each of which keeps the same artifact style; the species can fission into two, the offspring creating a new artifact style; the spe cies can persist and adopt a new artifact style; and so forth. Several simula
tion runs will produce the evolutionary patterns of artifact style diversity to
be expected if the causes are stochastic. The predicted patterns can be then
compared to actual observations to ascertain whether the null hypothesis can be rejected. If so, more deterministic models of diversity, such as
limiting similarity, can be tested; if not, ad hoc explanations may be the
best.
Discussion
Models of species diversity can, then, be made workable as possible ex
planations of cultural diversity and can be tested with archaeological data, at least in theory. The models that have been discussed are not intended to
be exhaustive; other competing models of species diversity can be gleaned
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

180 DONALD L HARDESTY
from the ecological literature. Rather, they illustrate something of the range of explanations that exist and the kind of problems that must be overcome
to render them useful for archaeological interpretation. More importantly, the discussion shows the need for multiple hypothesis testing if general
ecological models are to be used effectively in archaeology and anthro
pology. What about the generality of now-existing models of species diversity?
Are they capable of accommodating human ecology data and principles? Or
must the models be modified? Human behavior is unique in its capacity to
change the amplitude and predictability of environmental fluctuations, and
it is here that many of the species diversity models are likely to be too nar
row or specialized. Those that use responses to environmental variability as
explanatory principles, such as the stability-time hypothesis and the May MacArthur model, assume that environment is controlled by external pro
cesses; there is no provision for a species that responds by changing its en
vironment, for a process of ert?ronmental change that is internal to the
model. What must be added to make the models more general is an interac
tive process that connects the community of cultural or biological species and its environment.
To illustrate what that means in practice, consider two models that use
environmental variation to explain the evolution of complex human so
cieties. The first is Gall and Saxe's (1977) argument, already discussed, that the evolutionary process is analogous to that of ecological succession.
In their model, increasing cultural diversity is assumed to have a selective
advantage because diversity buffers the impact of environmental change and makes the ecological community more stable. But diversification may not be the cause of community stability; rather, increased diversity and
community stability may be the consequence of environmental stability
(Goodman 1975; May 1975; Pielou 1975; Whittaker 1977). If so, it is the
cause of environmental stability that is critical to the evolution of complex human societies, and that is not considered by Gall and Saxe.
Isbell (1978), by contrast, develops a model to explain the emergence of
the state in the central Andes that does take environmental variability, and
its modification by human behavior, into account; Jorde (1977) suggests a
similar model for the American Southwest. The model gives the selective
advantage to behavior that reduces the amplitude and increases the predict
ability of environmental fluctuations. Economic exchange networks con
necting geographically separated human populations are examples of this
kind of behavior, as is the establishment of a storage complex to "even
out" fluctuations over time. Both of these are expected to be advantageous
and, therefore, to become more common over time. Isbell gives archae
ological evidence to show that the culture history of the central Andes is
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 181
marked by just that kind of behavior: increasingly elaborate storage and
distribution facilities, increasingly centralized decision-making apparatus, and political entities with increasingly larger geographical territories. The
last is added here because recent mathematical models have shown that
greater spatial heterogeneity can increase the stability of an ecological com
munity (May 1974; Murdoch and Oaten 1975). An interesting implication of Isbell's model is that ideology may play an
important role in making the environments of human ecological com
munities more stable. Willey (1962) argues, for example, that widespread
religious horizons, such as the Olmec and Chavin, served to integrate
geographically separated populations. The ecological significance of closer
ties is that it encourages and makes possible economic exchange networks, behavior that dampens local fluctuations in the environment. If so, it rein
forces Vayda and Rappaport's (1968) argument that ideology is important to ecological explanation and does not justify the separation of "spiritual" and ecological models, as does, for example, Adams (1977:94).
The "internalization" of the cause of environmental variability is, as far
as we know, absent in general ecological models. Yet Homo sapiens is not
the only species that uses behavior to change its effective environment in a
habitat, although Homo has capitalized upon that ability. Burrowing animals do exactly the same thing, for example, by using biological struc
tures and mostly genetically controlled behavior to create microclimates
that are considerably different from the surrounding habitat. What that im
plies, of course, is that animals have two kinds of responses to environmen
tal fluctuations: (1) they can change their behavior or biological structure, as general ecological models assume, and (2) they can change their environ
ment by using biological structures, technology, social organization, and
the like. The addition of the latter response to general ecological models
would render them much more capable of accommodating human be
havior.
If environmental change is the response chosen, an interactive process
connecting human behavior, or that of another species, and the environ
ment can be conceptualized. Environmental uncertainty is countered by some kind of behavior, such as intergroup exchange of uncertain resources, that lowers the level of the uncertainty. But a less risky environment is
suitable for finer niche partitioning and a finely tuned adaptation. And that
is associated with the diversification of biological or cultural species. Fine
tuning, however, makes the species, now occupying more specialized niches, susceptible to smaller, more localized fluctuations in the environ
ment. The expected consequence is even more elaborate "buffering" be
havior to make the environment even more predictable. And so the pro cess continues toward 100% predictability. The evolutionary pattern of
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

182 DONALD L. HARDESTY
species diversification to be expected from the positive feedback process
depends upon which model is the best explanation. May and MacArthur's
limiting similarity model, for example, predicts that diversification will con
tinue as environmental fluctuations are reduced down to about 30% of the mean. At that point, no further change in diversity should take place until
the predictability of the environment has been increased to nearly 100%.
After that, diversification can continue without limit.
But as a community of biological or cultural species diversifies in an in
creasingly stable environment, the dilemma of all equilibrium communities
must be faced: unexpected major disturbances in the environment are likely to be disastrous (Bretsky and Lorenz 1970; Gould 1977; Holling 1973). The
disturbance may be external, such as a major climatic shift, or it may be in
ternal, such as the collapse of behavior that buffers unexpected fluctua
tions; the overthrow of a strong central government is an example if it
destroys exchange and storage systems. Whatever the reason for the shock, the finely tuned, narrow-niched species making up the community are likely to become extinct, either as a biological entity or as a behavioral entity. A
period of revolutionary cultural change is, for example, the likely archaeo
logical manifestation of the breakdown. The interactive process then begins anew.
CONCLUSION
The use of general ecological principles is a viable alternative to ad hoc
ecological explanation in archaeology. Changes must be made in their ap
plication to archaeological data, however, if logical problems are to be overcome. First of all, and perhaps most important, the principles cannot
be obtained by search and seizure and used without consideration of their
place in ecological theory. Few, if any, of the principles in general ecology are so generally accepted that they are paradigmatic; consequently, com
peting principles are likely to exist. And, furthermore, the principles may not be sufficiently general to accommodate human ecology. That suggests that the best use of general ecological principles is interactively with ar
chaeological data, testing competitors and making changes to improve their
generality. The second major change that must be made if models derived
from general ecology are to be useful for archaeological explanation is to
make them workable. Few applications have considered this problem; yet if
such models cannot be tested, the explanation is inadequate. More research
in these two directions may bring about a revolution in ecological explana tion.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 183
REFERENCES
Adams, R. E. W.
1977 Prehistoric Mesoamerica. Boston: Little, Brown.
Baerreis, D., R. A. Bryson, and J. E. Kutzbach.
1976 Climate and culture in the western Great Lakes region. Midcontinental Journal of
Archaeology 1:39-57.
Barth, F.
1956 Ecologie relationships of ethnic groups in Swat, North Pakistan. American An
thropologist 58:1079-1089.
Bates, D.
1971 The role of the state in peasant-nomad mutualism. Anthropological Quarterly 44:109-131.
Binford, L. R.
1962 Archaeology as anthropology. American Antiquity 28:217-225.
1968 Post-Pleistocene adaptations. In New perspectives in archaeology, edited by S. R. and L. R. Binford. Chicago: Aldine. Pp. 313-341.
Boserup, E.
1965 The conditions of agricultural growth. Chicago: Aldine.
Boucot, A. J.
1976 Stochastic models of phylogeny and the evolution of diversity: a discussion. The
Journal of Geology 84:731-733.
1978 Community evolution and rates of cladogenesis. In Evolutionary biology (Vol.
11), edited by M. Hecht, W. Steere, and B. Wallace. New York: Plenum. Pp.
545-655.
Bretsky, P. W., and D. Lorenz
1970 Adaptive response to environmental stability: a unifying concept in paleontology.
Proceedings of North American Paleontology (Part E Vol. 1). Pp. 522-550.
Chorley, R. J., and P. Haggett 1967 Integrated models in geography. In Models in geography (Vol. 4). London:
Methuen.
Clark, J. G. D.
1954 Excavations at Star Can. Cambridge, Massachusetts: Harvard University Press.
Clark, J. T., and J. Terrell
1978 Archaeology in Oceania. Annual Review of Anthropology 7:292-319.
Cohen, M.
1977 The food crisis in prehistory. New Haven, Connecticut: Yale University Press.
Connell, J. H.
1975 Some mechanisms producing structure in natural communities: a model and
evidence from field experiments. In Ecology and evolution of communities, edited
by M. L. Cody and J. M. Diamond. Cambridge, Massachusetts: Belknap, Har
vard University Press. Pp. 460-490.
Cook, S.
1973 Production, ecology, and economic anthropology: notes toward an integrated frame of reference. Social Science Information 12:25-52.
Diamond, J. M.
1978 Niche shifts and the rediscovery of interspecific competition. American Scientist
66:322-331.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

184 DONALD L. HARDESTY
Ford, R.
1977 Evolutionary ecology and the evolution of human ecosystems: a case study from
the midwestern U.S.A. In Explanation of prehistoric change, edited by J. N. Hill.
Albuquerque: University of New Mexico Press. Pp. 153-184.
Gall, P. L., and A. A. Saxe
1977 The ecological evolution of culture: the state as predator in prehistory. In Ex
change systems in prehistory, edited by T. Earle and J. E. Ericson. New York:
Academic Press. Pp. 255-268.
Goodman, D.
1975 The theory of diversity-stability relationships in ecology. The Quarterly Review of
Biology 50:237-266.
Gould, S. J.
1977 Palaeontology plus ecology as palaeobiology. In Theoretical ecology, edited by R.
May. Philadelphia: Saunders. Pp. 218-236.
Hardesty, D. L.
1975 The niche concept: suggestions for its use in human ecology. In Human Ecology
3:71-85.
1978 Human evolutionary ecology. In Human evolution (4th edition), edited by N.
Korn. New York: Holt. Pp. 234-244.
1979 Measurement in human ecology. In Population, adaptation, and evolution, edited
by J. C. Calhoun. New York: Praeger.
Hardin, G.
1960 The competitive exclusion principle. Science 131:1291-1297.
Holling, C. S.
1973 Resilience and stability of ecological systems. Annual Review of ecology and
Systematics 4:1-23.
Hutchinson, G. E.
1959 Homage to Santa Rosalia, or why are there so many kinds of animals? American
Naturalist 93:145-159.
1965 The ecological theater and the evolutionary play. New Haven, Connecticut: Yale
University Press.
Isbell, W. H.
1978 Environmental perturbations and the origins of the Andean state. In Social ar
cheology, beyond subsistence and dating, edited by C. Redman, W. T. Langhorne
Jr., M. J. Berman, N. M. Versaggi, E. V. Curtin, and J. C. Wanser. New York:
Academic Press. Pp. 303-313.
Jennings, J. D., and E. Norbeck
1955 Great Basin prehistory: a review. American Antiquity 21:1-11.
Jorde, L. B.
1977 Precipitation cycles and cultural buffering in the prehistoric Southwest. In For
theory building in archaeology, edited by L. Binford. New York: Academic Press.
Pp. 385-396.
Kaplan, S. A.
1976 Ethnographic and biogeographical significance of pottery sherds from Nissan
Island, Papua New Guinea. Fieldiana: Anthropology 66:35-89.
Kitts, D. B.
1975 Stochastic models of phylogeny and the evolution of diversity: a discussion. The
Journal of Geology 83:125-126.
Kuhn, T. S.
1970 The structure of scientific inquiry (2nd edition). Chicago: University of Chicago
Press.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 185
Kummer, H.
1971 Primate societies. Chicago: Aldine.
Levandowsky, W., and B. S. White
1977 Randomness, time scale, and the evolution of biological communities. In Evolu
tionary biology (Vol. 10), edited by M. Hecht, W. C. Steere, and B. Wallace. New
York: Plenum. Pp. 69-161.
Levins, R.
1968 Evolution in changing environments. Princeton, New Jersey: Princeton Univer
sity Press.
MacArthur, R. H.
1958 Population ecology of some warblers of northeastern coniferous forests. Ecology
39:599-619.
1972 Geographical ecology. New York: Harper.
MacArthur, R. H., and E. O. Wilson
1967 The theory of island biogeography. Princeton, New Jersey: Princeton University Press.
May, R.
1974 On the theory of niche overlap. Theoretical Population Biology 5:297-332.
1976 Patterns in multispecies communities. In Theoretical ecology, edited by R. May.
Philadelphia: Saunders. Pp. 142-162.
1978 The evolution of ecological systems. Scientific American (September 1978). Pp.
161-173.
May, R. (editor) 1975 Stability and complexity in model ecosystems (2nd edition). Princeton, New
Jersey: Princeton University Press.
May, R., and R. H. MacArthur
1972 Niche overlap as a function of environmental variability. Proceedings of the Na
tional Academy of Sciences 69:1109-1113.
Mayr, E.
1963 Animal species and evolution. Cambridge, Massachusetts: Belknap, Harvard
University Press.
Murdoch, W. W., and A. Oaten
1975 Pr?dation and population stability. In Advances in ecological research (Vol. 9),
edited by A. Macfadyen. New York: Academic Press. Pp. 1-31.
Nisbet, R. M., W. S. C. Gurney, and M. A. Pettipher 1978 Environmental fluctuations and the theory of the ecological niche. Journal of
Theoretical Biology 75:223-237.
Osborne, A.
1977 Strandloopers, mermaids, and other fairy tales: ecological determinants of marine
resource utilization?the Peruvian case. In For theory building in archaeology, edited by L. R. Binford. New York: Academic Press. Pp. 157-205.
Osman, R. W., and R. B. Whitlatch
1978 Patterns of species diversity: fact or artifact? Paleobiology 4:41-54.
Peters, R. H.
1976 Tautology in evolution and ecology. American Naturalist 110:1-12.
Pianka, E. R.
1974 Niche overlap and diffuse competition. Proceedings of the National Academy of
Science 71:2141-2145.
Pielou, E. C.
1975 Ecological diversity. New York: Wiley.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

186 DONALD L HARDESTY
Plog, F.
1974 The study of prehistoric change. New York: Academic Press.
Raup, D. M.
1977 Probabilistic models in evolutionary paleobiology. American Scientist 65:50-57.
Raup, D. M., S. J. Gould, T. J. M. Schopf, and D. S. Simberloff
1973 Stochastic models of phylogeny and the evolution of diversity. The Journal of
Geology 81:525-542.
1975 Reply to Kitts. The Journal of Geology 83:126-127. 1976 Reply to Boucot. The Journal of Geology 84:734-735.
Reher, C. A.
1977 Adaptive process on the shortgrass plains. In For theory building in archaeology, edited by L. R. Binford. New York: Academic Press. Pp. 13-40.
Rogers, E. S.
1972 The Mistassini Cree. In Hunters and gatherers today, edited by M. G. Bicchieri.
New York: Holt. Pp. 90-137.
Sanders, H. L.
1969 Benthic marine diversity and the stability-time hypothesis. In Diversity and
stability in ecological systems, edited by G. M. Wood well and H. H. Smith.
Brookhaven Symposium in Biology No. 22.
Schiffer, M. B.
1978 Some issues in the philosophy of science and archaeology. Unpublished paper
delivered at the State University of New York, Binghamton, November 1978.
Schoener, T.
1974 Resource partitioning in ecological communities. Science 185:27-39.
Service, E.
1962 Primitive social organization. New York: Random House.
Simberloff, D. S.
1974 Equilibrium theory of island biogeography and ecology. Annual Review of
Ecology and Systematics 5:161-182.
1978 Using island biogeographic distributions to determine if colonization is stochastic.
American Naturalist 112:713-726.
Simon, H. A.
1969 The sciences of the artificial. Cambridge, Massachusetts: MIT Press.
Steward, J. H.
1938 Basin-plateau aboriginal sociopolitical groups. Washington, D.C.: Smithsonian
Institution, Bureau of American Ethnology, Bulletin 120.
Terrell, J.
1974 Comparative human and lower animal biogeography in the Solomon Islands.
Solomon Island studies in human biogeography 3. Department of Anthropology, Field Museum of Natural History.
Van Valen, L.
1965 Morphological variation and width of ecological niche. American Naturalist
99:377-390.
1973 A new evolutionary law. Evolutionary Theory 1:1-30.
Vayda, A. P., and R. A. Rappaport 1968 Ecology, cultural and noncultural. In Introduction to cultural anthropology,
edited by J. Clifton. Boston: Houghton. Pp. 477-497.
Watson, P. J., S. A. LeBlanc, and C. Redman
1971 Explanation in archaeology: an explicitly scientific approach. New York: Colum
bia University Press.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions

THE USE OF GENERAL ECOLOGICAL PRINCIPLES IN ARCHAEOLOGY 187
Weins, J. A.
1977 On competition in variable environments. American Scientist 65:590-597.
Whittaker, R. H.
1977 Evolution of species diversity in land communities. In Evolutionary biology (Vol.
10), edited by M. Hecht, W. C. Steere, and B. Wallace. New York: Plenum.
Pp. 1-67.
Whittaker, R. H., and S. A. Levin (editors) 1975 Niche, theory and application, Pennsylvania: Stroudsburg, Dowden, Hutchinson
and Ross.
Willey, G. R.
1962 The early great styles and the rise of the pre-Columbian civilizations. American
Anthropologist 64:1-14.
Yellen, J.
1977 Long term hunter-gatherer adaptation to desert environments: a biogeographical
perspective. World Archaeology 8:262-274.
This content downloaded from 185.2.32.58 on Sun, 15 Jun 2014 10:53:54 AMAll use subject to JSTOR Terms and Conditions





























