Embed Size (px)
Citation preview

Mutation Research, 104 (1982) 79-86 79 Elsevier Biomedical Press
The use of a yeast strain with a temperature-sensitive D N A ligase to estimate D N A repair after exposure
to mutagens
Richard S. Tippins , R a y m o n d Waters and James M. P a r r y
Department of Genetics, University College of Swansea, Singleton Park, Swansea SA2 8PP (Great Britain)
(Accepted 24 November 1981)
Summary
The yeast strain cdc9 which possesses a temperature-sensitive DNA ligase, was used to estimate DNA repair after mutagen exposure. Following low UV fluences, single-strand breaks in DNA were detected after an incubation at the restrictive temperature but were absent at the permissive temperature. These DNA breaks were shown to be equal to the number of pyrimidine dimers induced in DNA as measured by the presence of UV-endonuclease-sensitive sites. Similarly, after exposure to the chemical mutagen 4-chloromethyl-biphenyl (4CMB) single-strand breaks accumu- lated at the restrictive temperature. Hence the technique described should be appli- cable for the estimation of the early steps of repair of a wide range of different types of DNA damage induced in yeast by exposure to either physical or chemical mutagens.
The estimation of DNA repair in the yeast Saccharomyces cerevisiae has so far been hampered by several factors. One major problem is the absence of thymidine kinase in this organism, hence the inability of yeast workers to use many of the conventional techniques previously developed for the measurement of DNA repair in bacterial and mammalian systems which involve the uptake of radiolabelled thymidine or its analogue bromodeoxyuridine. The labelling of yeast DNA with a general nucleic acid precursor such as uracil has however enabled studies to be performed on the repair of damage induced after exposure to ethylating or methylating agents which are capable of inducing alkaline-labile lesions in DNA (Jachymezyk et al., 1977) and the repair of UV-induced pyrimidine dimers by the use of either chromatographic techniques (Unrau et al., 1971) or the measurement of incidence of UV-endonuclease-sensitive sites (Prakash, 1975; Reynolds, 1978).
0165-7992/82/0000-0000/$02.75 ~ Elsevier Biomedical Press

80
Despite these advances no technique had yet been developed which either accurately measures the rate of lesions repaired as opposed to those left in DNA or has a widespread application in being nonspecific as to the type of DNA lesion induced.
In this paper we describe a novel approach which provides for the direct monitoring of the repair of lesions induced by both chemical or physical mutagens in yeast DNA. Essentially the technique involves using the cell cycle mutant cdc9 (HartweU et al., 1973) which possesses a temperature-sensitive DNA ligase (Johnston and Nasmyth, 1978). In this strain the 'repair ' of lesions at the restrictive temperature of 37 ° results in the production of single-strand breaks due to the failure to ligate the repaired portion of DNA. The frequency of such unrepaired lesions may then be estimated by alkaline sucrose sedimentation of the DNA.
Materials and methods
The cdc9-1 (mutant 244) was obtained from the Yeast Genetic Stock Centre, Berkeley. This haploid strain is capable of growth at a permissible temperature of 24 ° but shows no growth at the restrictive temperature of 37 ° (Hartwell et al., 1973).
Growth and labelling conditions The strain was labelled overnight at 24 ° with 5 #Ci/ml [6-SH]uracil (Amersham)
in YEPG (yeast extract, peptone and glucose) until a density of 2 - 4 × l07 cells/ml was reached. They were then washed and resuspended in fresh unlabelled media for 1 h followed by a further 1-h incubation in nitrogen- and glucose-free starvation media to chase the radiolabel.
UV irradiation 10 ml of cells were irradiated with constant agitation on ice at a dose rate of 0.7
J. m -2 sec -1 in open petri dishes. The source of UV light was a Hanovia l l a low pressure mercury discharge tube. The dose was monitored by a calibrated photocell and voltmeter. All manipulations were carried out in red light to avoid photoreac- tivation.
4-CMB treatment The 4-chloromethylbiphenyl (4CMB) was supplied by J. Ashby at ICI Ltd.,
Macclesfield. The cells were exposed to a concentration of 10/~g/ml 4CMB for a period of 8 h at 24 ° in minimal medium.
Post-mutagen incubations Following mutagen treatment the cells were resuspended in minimal media to
which hydroxyurea (0.075 M) had been added. The cells were then incubated either

81
at the restrictive temperature (37 ° ) or at the permissible temperature (24 ° ) for
varying periods of time for up to 8 h.
Alkaline sucrose sedimentation The cells were washed 2 times with ice cold 0.05 M EDTA prior to being exposed
to 2°70/3-mercaptoethanol for 30 min on ice. The cells were resuspended in 1 ml of 1 M sorbitol, 0.5 M EDTA at pH 7.5 to which 100/d of zymolyase (Kirin Breweries, Japan) was added to give a final concentration of 5 mg/ml. The cells were incubated
for 15-20 min at 24 ° prior to being resuspended in ice cold 0.15 M NaC1, 0.1 M
EDTA and 100/~g/ml proteinase K (Boehringer). 100-#l samples of the spheroplasts produced by the above treatment were lysed directly onto the top of a 5-20o70 (w/v)
alkaline sucrose gradient (2 M NaC1, 0.33 M NaOH, 0.015 M EDTA). The lysis layer consisted of 100 #I of 7°7o sarkosyl, 0.15 M NaC1 and 0.1 M EDTA. Layered
samples were incubated at 20 o for 1 h prior to sedimentation in a Beckman model
L8-55 ultracentrifuge with an SW-60 rotor for 150 min at 28500 rpm at 20 ° After sedimentation the gradients were pumped from the bottom of the tubes
onto Whatman 17 paper strips marked into fractions every 2 cm. The fractions were collected at fixed time intervals to give 21-23 fractions per gradient. After fractio-
nation the strips were washed once in 5070 TCA and twice in 95°7o ethanol. The strips
were then dried overnight at 28 ° before being cut up and placed into scintillation vials with a toluene-based scintillation fluid. The samples were counted using a LKB 1216 Rackbeta scintillation counter. The resulting data were analysed with a
computer program calibrated with phage DNAs of known molecular weights, and which generated the weight-average molecular weights of the DNA samples. The
mutagen induced strand breakage was thus calculated as:
Number of breaks/108 dalton = 2 2 Mw treated DNA - Mw control DNA
The repair of mutagen produced lesions was calculated by:
Lesions repaired/10 ~ dalton/time = 2 2 Mw treated DNA - Mw treated DNA
at 37 ° at 24 °
Detection of pyrimidine dimers The protocol for detecting pyrimidine dimers using UV endonuclease has been
described elsewhere (Prakash, 1975; Reynolds, 1978).
Results
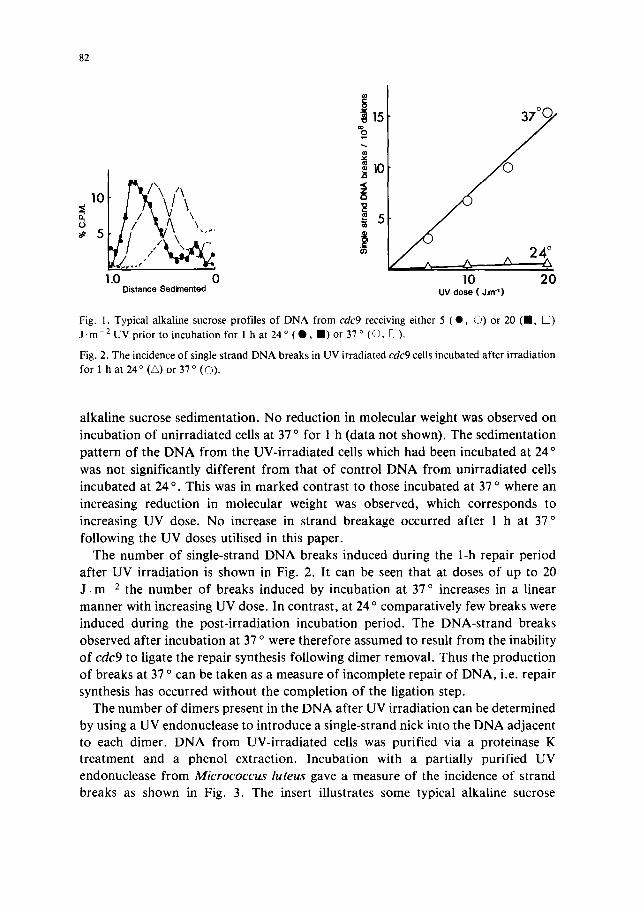
Fig. 1 shows the DNA profiles of UV-irradiated cdc9 cells that were irradiated with 5 or 20 J . m -2 of UV and incubated at either 24 ° or 37 ° for 1 h prior to
82
10
O 5
1.0
/ /
0 Distance Sedimented
:t= 15
%
z Q
10 UV dose (d.m-:)
2O
Fig. 1. Typical alkaline sucrose profiles of DNA from cdc9 receiving either 5 ( 0 , O) or 20 (11, [~) J.m 2 UV prior to incubation for 1 h at 24 ° (O , II) or 37 ° (O, I~).
Fig. 2. The incidence of single strand DNA breaks in UV irradiated cdc9 cells incubated after irradiation for 1 h at 24 ° (•) or 37 ° (O).
a lka l ine sucrose sed imen ta t ion . N o reduc t ion in molecu la r weight was observed on
incuba t ion o f un i r r ad i a t ed cells at 37 ° for 1 h (da ta no t shown). The sed imen ta t ion
pa t t e rn o f the D N A f rom the UV- i r r ad i a t ed cells which had been incuba ted at 24 °
was not s igni f icant ly d i f fe ren t f rom tha t o f con t ro l D N A f rom u n i r r a d i a t e d cells
i ncuba ted at 24 °. This was in m a r k e d con t ras t to those incuba ted at 37 ° where an
increas ing r educ t ion in mo lecu la r weight was observed, which co r r e sponds to
increas ing UV dose . N o increase in s t r and b reakage occur red af ter 1 h at 37 o
fo l lowing the UV doses ut i l ised in this paper .
The n u m b e r o f s ingle-s t rand D N A breaks induced dur ing the 1-h repai r pe r iod
af te r UV i r r ad i a t i on is shown in Fig. 2. I t can be seen tha t at doses o f up to 20
J . m -2 the n u m b e r o f b reaks induced by incuba t ion at 37 ° increases in a l inear
manne r with increas ing UV dose. In con t ras t , at 24 ° compara t i ve ly few breaks were
induced dur ing the pos t - i r r ad i a t i on incuba t ion per iod . The D N A - s t r a n d b reaks
observed af te r i ncuba t ion at 37 o were the re fo re assumed to result f rom the inabi l i ty
o f cdc9 to l igate the repa i r synthesis fo l lowing d imer removal . Thus the p roduc t i on
o f b reaks at 37 ° can be t aken as a measure o f incomple te repai r o f D N A , i.e. repa i r
synthesis has occur red wi thou t the comple t i on o f the l iga t ion step.
The n u m b e r o f d imers present in the D N A af ter UV i r r ad ia t ion can be de te rmined
by using a UV endonuc lease to in t roduce a s ingle-s t rand nick into the D N A ad jacen t
to each d imer . D N A f rom UV- i r r ad i a t ed cells was pur i f ied via a p ro te inase K
t r ea tmen t and a pheno l ex t rac t ion . Incuba t ion with a par t i a l ly pur i f ied UV
endonuc lease f rom Micrococcus luteus gave a measure o f the incidence o f s t rand
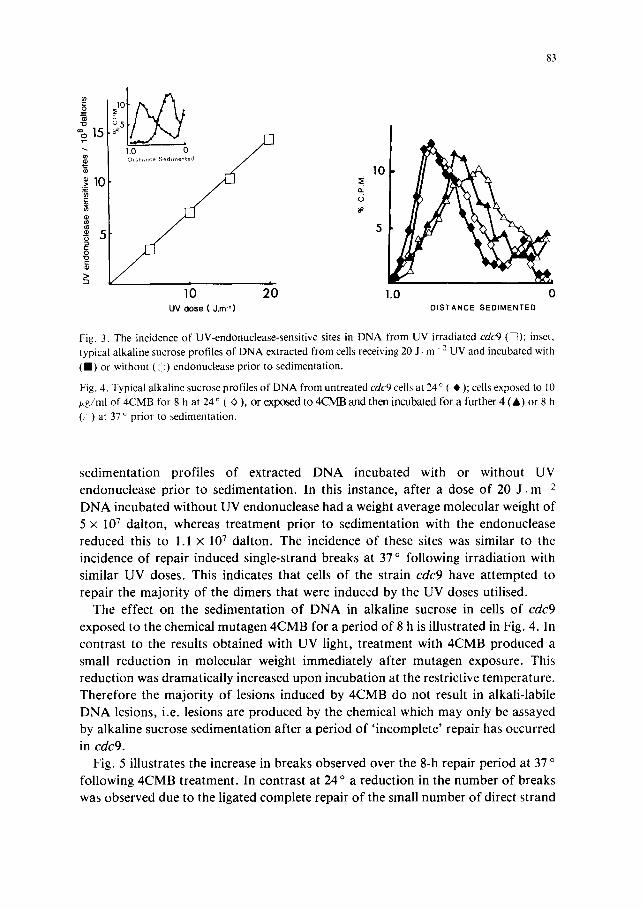
breaks as shown in Fig. 3. The insert i l lustrates some typica l a lka l ine sucrose
10
o
% 15
~ 10
~ 5 g g
10 20 UV dose ( J.m -~)
1.0 0 DISTANCE SEDIMENTED
83
Fig. 3. The incidence of UV-endonuclease-sensitive sites in DNA from UV irradiated cdc9 ([Z); inset, typical alkaline sucrose profiles of DNA extracted from cells receiving 20 J. m " 2 UV and incubated with (11) or without (<i) endonuclease prior to sedimentation.
Fig. 4. Typical alkaline sucrose profiles of DNA from untreated cdc9 cells at 24 ° ( ~, ); cells exposed to 10 p.g/ml of 4CMB for 8 h at 24 ° ( O ), or exposed to 4CMB and then incubated for a further 4 (A) or 8 h ( ) at 37 o prior to sedimentation.
s ed imen ta t i on prof i les o f ex t rac ted D N A incuba ted with or wi thout UV
endonuc lease p r io r to sed imenta t ion . In this instance, af ter a dose o f 20 J . m 2
D N A incuba ted wi thout UV endonuc lease had a weight average molecu la r weight o f
5 x 10 7 da l ton , whereas t r ea tmen t p r io r to sed imen ta t ion with the endonuc lease
reduced this to 1.1 × 107 da l ton . The incidence o f these sites was s imilar to the
incidence o f repai r induced s ingle-s t rand b reaks at 37 ° fol lowing i r r ad ia t ion with
s imi lar UV doses . This indicates that cells o f the s t ra in cdc9 have a t t e m p t e d to
repai r the m a j o r i t y o f the d imers tha t were induced by the UV doses uti l ised.
The effect on the sed imen ta t ion o f D N A in a lka l ine sucrose in cells o f cdc9
exposed to the chemical mutagen 4 C M B for a pe r iod o f 8 h is i l lus t ra ted in Fig. 4. In
con t ras t to the results ob ta ined with UV light, t r ea tment with 4 C M B produced a
small r educ t ion in molecu la r weight immed ia t e ly af te r mu tagen exposure . This
r educ t ion was d r ama t i ca l l y increased upon incuba t ion at the restr ict ive t empera tu re .
The re fo re the m a j o r i t y o f lesions induced by 4 C M B do not result in a lka l i - lab i le
D N A lesions, i .e. lesions are p r o d u c e d by the chemical which may only be assayed
by a lka l ine sucrose sed imen ta t ion af ter a pe r iod o f ' i n c omple t e ' repa i r has occurred
in cdc9.
Fig. 5 i l lustrates the increase in b reaks observed over the 8-h repai r per iod at 37 o
fo l lowing 4CMB t r ea tmen t . In con t ras t at 24 o a r educ t ion in the number o f b reaks
was observed due to the l igated comple te repai r o f the small number o f direct s t rand

84
].0t ~ ~ " c
a
~- o5 (/)
== 0 2 ~, 6 8 HOURS AT 37°C OR 24 "C
Fig. 5. The incidence of single strand DNA breaks in cdc9 exposed to 10 ~g/ml of 4CMB at 24 ° for 8 h prior to further post-treatment incubations at 24 ° or 37 ° for further periods up to 8 h.
breaks induced by this mutagen. The repair of 4CMB induced lesions was seen to be slower than that observed after UV-light exposure. Furthermore, as previously stated, no significant increase in breaks occurred after a l-h incubation at 37 ° in UV-irradiated samples. However, an increase in single-strand breaks occurred over an 8-h post-treatment period after exposure to 4CMB.
Discussion
The results presented in this paper indicate that the kinetics of DNA repair in yeast can be accurately determined at early times after UV irradiation. This may be achieved by utilizing cells o f the cdc9 strain that has a temperature-sensitive DNA ligase and by the measurement of the incidence of single-strand breaks that occur during incomplete repair at the restrictive temperature following UV irradiation.
The advantages of this technique are that: (a) it directly measures the repair of lesions induced rather than detecting those lesions which remain unrepaired in DNA, (b) DNA extraction and purification are not necessary which therefore circumvents the problems produced by extra manipulation of the DNA, (c) because the DNA-strand breaks observed using this technique appear to be due to non- ligated repair regions, it should not be specific in detecting the repair of only certain types of lesions induced in DNA.
Since the experiments reported here were undertaken similar experiments concerning UV irradiation have been reported by Wilcox and Prakash (1981). Essentially these authors have obtained similar data concerning the response of UV- irradiated cdc9 cells at the restrictive and permissive temperatures. In addition, the induction of single-strand breaks at 37 o was blocked in a rad3 cdc9 double mutant. As rad3 is defective in excision repair this observation indicates that the strand breakage observed in cdc9 cells was due to aborted DNA repair.
The observation that the number of strand breaks induced in cdc9 at 37 ° equals the number of UV-induced pyrimidine dimers induced at low UV doses indicates

85
that excision repair in yeast cells is uncoupled. That is, despite the non-l igat ion o f
lesions undergoing repair the cells still cont inue to under take the repair o f subsequent damage. This is in contrast to what has been observed in human cells
following UV irradiat ion and its subsequent repair in the presence o f the analogue cytosine arabinoside (Hiss and Preston, 1977; Collins et al., 1977).
The chemical mutagen 4CMB (Parry, 1982) may cause 2 forms o f D N A damage in yeast. However , the major i ty o f the lesions induced do not result in alkali-labile
sites and the breaks detected at the permissive temperature may be alkaline-labile
lesions or alkaline-stable lesions in the process of repair. Alkaline-stable lesions can be detected after subsequent incubat ion at the restrictive temperature. The kinetics
o f repair o f these chemically induced lesions appear to be slower than those observed with UV light. The technique o f utilising c d c 9 cells to moni tor the repair o f
D N A damage could thus be useful in detecting the repair o f damage induced by
environmental mutagens and carcinogens. The work presented here is currently being extended by incorporat ing a range o f
radiation-sensitive muta t ions into this strain. It is hoped that this will allow
calculations o f the relative contr ibut ion o f each locus to the D N A repair o f various
lesions induced in D N A by mutagens.
Acknowledgements
We would like to acknowledge the SRC and EEC Environmenta l Research
P rog ram for support o f this project . The technical assistance o f Mrs. Mary Lewis is
also gratefully acknowledged.
References
Collins, A.R.S., S.L. Schlor and R.T. Johnson (1977) The inhibition of reprit in UV-irradiated human cells, Mutation Res., 42, 413-432.
Hartwell, L.H., R.K. Mortimer, J. Culotti and M. Culotti (1973) Genetic control of the cell division cycle in yeast V, Genetic analysis of cdc mutants, Genetics, 74, 267-286.
Hiss, E.A., and R.J. Preston (1977) The effect of cytosine arabinoside on the frequency of single-strand breaks in DNA of mammalian cells following irradiation or chemical treatment, Biochim. Biophys. Acta, 478, 1-8.
Jachymezyk, W.J., E. Chlebowicz, Z. Swietlinska and J. Zuk (1977) Alkaline sucrose sedimentation studies of MMS-induced DNA single-strand breakage and rejoining in the wild-type and in UV-sensi- tive mutants of Saccharomyces cerevisiae, Mutation Res., 43, 1-10.
Johnston, L.H., and K.A. Nasmyth (1978) Saccharomyces cerevisiae cell cycle mutant cdc9 is defective in DNA ligase, Nature (London), 274, 891-893.
Parry, J.M. (1982) The genetic toxicology of 4-chloromethyl biphenyl (4CMB), 4-hydroxymethyl biphenyl (4HMB) and benzyl chloride (BC) as assessed by the UKEMS Genotoxicity Trial 1981, Muta- tion Res., 100, 411-416.

86
Prakash, L. (1975) Repair of pyrimidine dimers in nuclear and mitochondrial DNA of yeast irradiated with low doses of ultraviolet light, J. Mol. Biol., 98, 781-795.
Reynolds, R.J. (1978) Removal of pyrimidine dimers from Saccharomyces cerevisiae nuclear DNA under nongrowth conditions as detected by a sensitive enzymatic assay, Mutation Res., 50, 43-56.
Unrau, P., R. Wheatcroft and B.S. Cox (1971) The excision of pyrimidine dimers from DNA of ultra- violet irradiated yeast, Mol. Gen. Genet., 113, 359-362.
Wilcox, D.R., and L. Prakash (1981) Incision and post-incision steps of pyrimidine dimer removal in ex- cision defective mutants of Saccharomyces cerevisiae, J. Bacteriol., in press.





























