Embed Size (px)
Citation preview

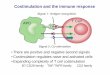
The Two-Signal Model of T-cell Activation
TCRMHC
CD4 or CD8
1
2
DC T cell
COSTIMULATION

• Experimental evidences for the two signal model of T cell activation• Experimental evidences that CD28 is a costimulatory receptor• Effects of CD28-mediated costimulation on T cells• Mechanism of action• The ligands for CD28: B7 molecules• The complexity of the B7/CD28 system: CD28 plays a role in promoting T cell tolerance!!
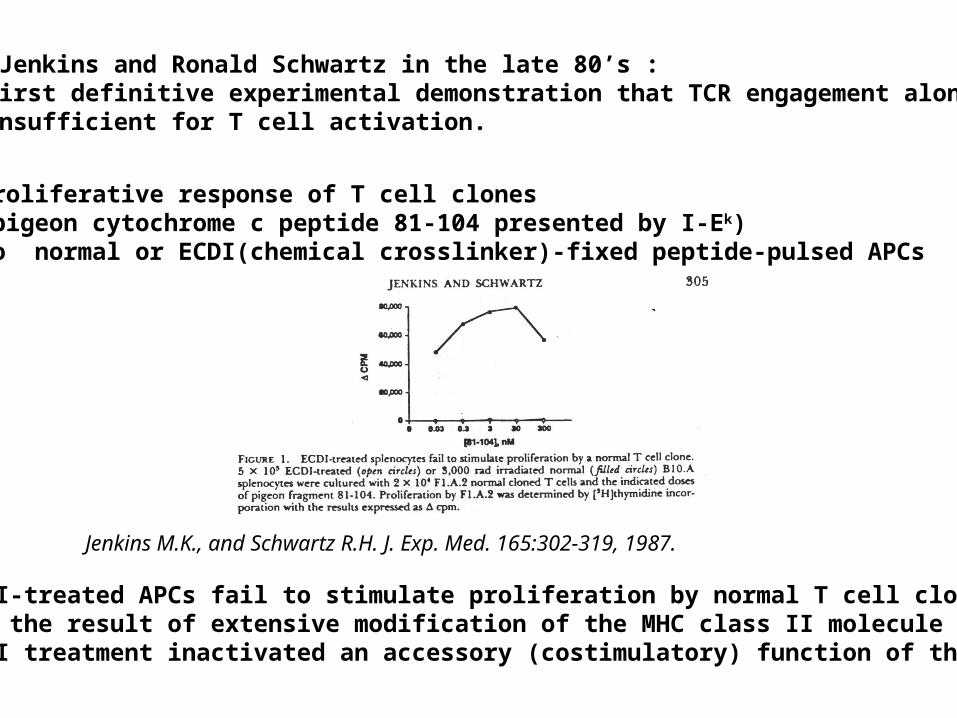
Marc Jenkins and Ronald Schwartz in the late 80’s :The first definitive experimental demonstration that TCR engagement alone was insufficient for T cell activation.
Proliferative response of T cell clones (pigeon cytochrome c peptide 81-104 presented by I-Ek)to normal or ECDI(chemical crosslinker)-fixed peptide-pulsed APCs
ECDI-treated APCs fail to stimulate proliferation by normal T cell clones :Not the result of extensive modification of the MHC class II moleculeECDI treatment inactivated an accessory (costimulatory) function of the APC
Jenkins M.K., and Schwartz R.H. J. Exp. Med. 165:302-319, 1987.

T cell clones stimulated in vitro by EDCI-treated APCswere induced into a state of unresponsiveness (Anergy).
Jenkins M.K., and Schwartz R.H. J. Exp. Med. 165:302-319, 1987.

Cells with this costimulatory function:
macrophages, activated B cells and dendritic cells (professional antigen presenting cells).
Subsequently,it was shown that:
Naïve T cells have similar requirement for these costimulatory signals in order to produce IL-2 and progress through the cell cycle.

TCR
peptide/MHC I
Parenchymal cell
T cell Self-reactive Anergy
This requirement for costimulation and the restricted pattern of expression of the costimulatory ligands to professional APCs,was proposed to be a mechanism for maintenance of peripheral T cell tolerance.
Costimulatory Ligands
Costimulatory Receptors

PROLIFERATION +PROLIFERATION +IL-2 PRODUCTIONIL-2 PRODUCTION
PREVENTION ANERGYPREVENTION ANERGY
T cell
Ag/MHC TCR
CD28CD28
PROLIFERATION +PROLIFERATION +IL-2 PRODUCTIONIL-2 PRODUCTION
INDUCTION OF ANERGYINDUCTION OF ANERGY
PROLIFERATION +PROLIFERATION +IL-2 PRODUCTIONIL-2 PRODUCTION
Ag/MHC TCR
CD28CD28
The concept of costimulation was consistent with the characteristicsof a newly identified molecule called CD28, which is expressed onnaive CD4+ and CD8+ T lymphocytes.
Artificial APC
Physiological APC
TCR
CD28CD28
Beads
Artificial APC
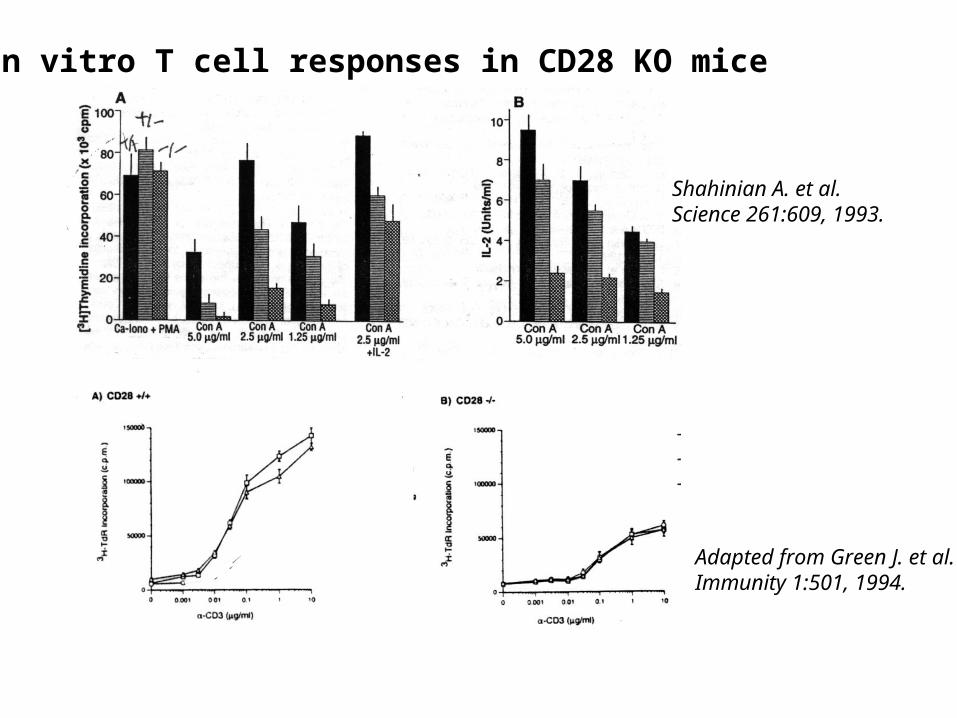
In vitro T cell responses in CD28 KO mice
Shahinian A. et al.Science 261:609, 1993.
Adapted from Green J. et al.Immunity 1:501, 1994.

The importance of CD28-mediated costimulation has also been demonstrated in vivo.
Blockade of CD28-mediated costimulation has been shown to:
• Prolong graft survival
• Block primary antibody response
• Prevent or reduce in intensity experimentally, antigen-induced models of
autoimmunity including arthritis, encephalomyelitis, myocarditis, thyroiditis and
myasthenia gravis

ProliferationIL-2 (transcription, mRNA stabilization)IL-2R up-regulationG1 cell cycle kinasesCell cycle inhibitor p27Kip
Survival Bcl-xL
Effector function CD40-L, OX-40, 41BB, ICOS cytokines expression cytotoxic molecules
The major effects of CD28-mediated costimulation are to augment and sustain T cellresponses initiated by antigen receptor signal by promoting T-cell survival andenabling cytokines to initiate T cell clonal expansion and differentiation.
Major effects of CD28-mediated costimulation in T cells

T cells stimulated in the presence of signal 1 alone can respond only when a certain number of TCR complexes are triggered (threshold ≈ 8000 TCR engaged molecules).
In the presence of costimulation through CD28, the number of TCR complexes that should be engaged for the induction of a response isdecreased to 1500 or less.
CD28 engagement lowers the activation threshold, thus making T cells more sensitiveto antigenic stimulation.
In the presence of CD28 stimulationIn the absence of CD28 stimulation
A + B = human T cellsC= T cell clones
Viola, A. and Lanzavecchia A. Science 273: 104, 1996
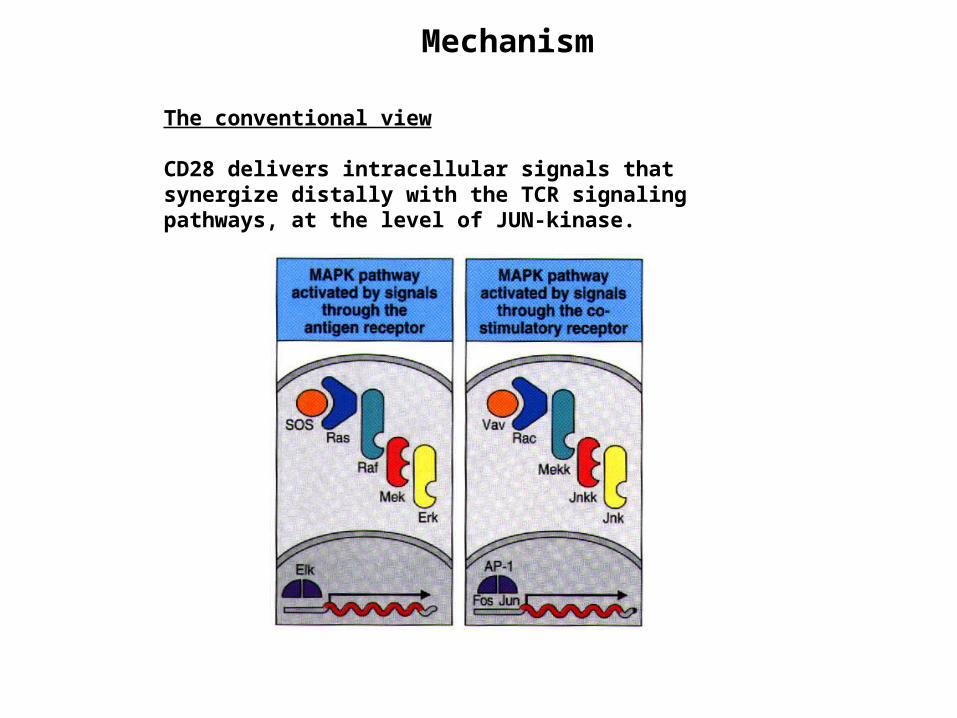
The conventional view
CD28 delivers intracellular signals that synergize distally with the TCR signaling pathways, at the level of JUN-kinase.
Mechanism

The new concept (based on the visualization of T cell activation)Kupfer et al. have shown that T cells polarize toward the APCs, thereby forming a highly structured synapse (the immune synapse).This process involves a redistribution of surface and intracellular molecules.Some molecules, such as TCR, CD4, src kinases are enriched in a central zone, whereas others, such as LFA-1 and CD45, remain outside.
• helps maintaining the interaction between the TCR and the peptide/MHC complexes (low concentration and low affinity for the TCR)
• helps in the local accumulation of intracellular signaling complexes

Figure 1. Bead movement toward the interface. Single frames of a video microscopy experiment (36) of the interaction of 5C.C7 T cells, loaded with 4.5-µm anti-ICAM-1 (YN1) beads, with peptide-loaded CH27 B cell lymphoma cells (35) are shown. The CH27 cell is substantially larger than the T cells. The T cell intracellular calcium concentration is overlaid in a false color scale from blue (low concentration) to red (high concentration) to mark the onset of T cell activation, set to time 0:00 min. Although the beads, one of which is marked with an arrow, are bound to the posterior end of the central T cell before and at the time of its activation, they can be seen to move toward the T cell-B cell interface in subsequent frames.
Wulfing, C. and and Davies, M.M. Science 282:2266, 1998.1. Actine cytoskeleton reorients toward the T cell-APC interface soon after the start of T cell activation
Beads are attached to the surface of T cells and monitor the movement of the cytoskeleton in the T cells.

2. CD28 engagement is necessary and sufficient to induce the movement of the T cell actine cytoskeleton toward the newly formed T cell-APC interface.
CD28 activates an actin-myosin-driven transport process that delivers receptors and signaling complexes to the TCR contact area.
Type of beads/treatment/APC Movement Movementtoward interface away interface No mov n
---------------------------------------------------------------------------------------------------------------Type of beadsAnti-ICAM 74% 0% 26% 81Streptavidin (surface biotin) 83% 7% 10% 29Streptavidin (lipid biotin) 64% 5% 32% 22Antibody blockinganti-B7 8% 0% 92% 24anti-VCAM 62% 0% 38% 21anti-CD48 75% 0% 25% 20
Adapted from Wulfing, C. and and Davies, M.M. Science 282:2266, 1998.

Viola, Schroeder, Sakakibara and Lanzavecchia, Science 1999:The plasma membrane is composed of discrete lipid microdomains in which membrane molecules are differentially partitioned (lipid rafts). These rafts concentrate glycophosphatidylinositol-linked proteins, glycosphingolipids, as well as several molecules involved in signal transduction such as Lck, LAT, Ras, and G proteins.CD28 participates in the organization of the immune synapse by recruiting rafts into it.The recruitment of rafts to the site of TCR engagement mayrepresent a general mechanism by which costimulation can increase the signaling process.
A new paradigm is emerging that is built around the central role of costimulation in contact formation necessary for T cell activation.
CD28-induced raft redistribution to the site of TCR engagement. Resting T cells were stained with FITC-CTx and analyzed by confocal microscopy.(A) Unstimulated resting T cells. (B) T cells stimulated for 20 min with beads coated with anti-CD3 alone (10 µg/ml) and (C) stimulated for 20 min with beads coated with anti-CD3 (10 µg/ml) plus anti-CD28. raft marker (FITC)-labeled cholera toxin (CTx) B subunit, which binds the GM1 glycosphingolipid

The ligands for CD28
1982, B7.1(CD80): Artificial APCs transfected with the cDNA coding for B7.1 induceT cell proliferation and IL-2 secretion.
However, when T cells are stimulated with professional APCsanti-B7.1mAb does not block completely T cell activationwhile anti-CD28 mAb does.
Yes, there is another ligand for CD28: B7.2 (CD86) (1993)
Artificial APCs transfected with the cDNA coding for B7.1 or B7.2induce similar T cell proliferation and IL-2 production.

B7.1 and B7.2: What’s the difference?
Expression: Primarily limited to APCs B7.1 is absent from unstimulated cells.B7.2 is constitutively expressed at low levels on unstimulated DCs and blood monocytes.Activation of APCs induces B7 expression.In general, the induction of B7.2 expression follows faster kinetics and usually reaches higher values than does B7.1 induction. In vivo, anti B7.2 mAb was shown to be more efficient than anti-B7.1 mAb to block the expansion of Ag-specific T cells.B7.1 KO vs B7.2 KO:When immunized intravenously without adjuvant, B7.2-deficient mice failed to form germinal centers, whereas B7.1-deficient mice gave Ab responses comparable with wild-type mice.
B7.2 is probably the major initial ligand for CD28 while B7.1 which is expressed later sustains T cell activation.

The differentiation of CD4+ T cells into Th1 or Th2 subsets has profound effectson the outcome of immune responses:Th1: INFg, eradication of intracellular pathogens, autoimune disease .Th2:IL-4, IL-5, IL-10 eradication of extracellular parasites, allergic reactions.
EAE: Th1-dependent autoimmune diseaseanti-B7.1 mAb treatment reduced the incidence of disease of murine EAE, anti B7.2 mAb made the disease worse (Kuchroo et al., 1995).
It has been suggested that this differential effect of the B7 mAbs reflected the abilitiy of B7.1 and B7.2 to direct T cell differentiation along the Th1 or Th2 pathway, respectively.
B7.1 Th1 differentiation B7.2 Th2 differentiation

Production of cytokines when T cells are primed in the absence of B7.1 or B7.2.
T cells = TCR Tg CD4+ specific for an OVA peptide presented by I-Ad
T cells + WT APC pulsed with OVA peptide
T cells + B7.1 KO APC pulsed with OVA peptide
T cells + B7.2 KO APC pulsed with OVA peptide
4 daysT cells + WT APC
2 days
T cells + WT APC
T cells + WT APC
IFNIL-4
IFNIL-4
IFNIL-4
Schweitzer N.A. et al. J.Immunol. 158:2713, 1997.

When T cells are primed under optimal signal 1, neither B7.1 or B7.2 plays an obligatory rolein priming for production of a particular cytokine. These data do not support a simple Th1/Th2-determining role for either B7.1 or B7.2.
When T cells are primed under suboptimal signal 1, both B7.1 and B7.2 are important forthe induction of Th1 or Th2 cytokines synthesis, with B7.2 having a greater influence compared with B7.1.
The influence of B7.1 or B7.2 on T cell differentiation is determined mainly by their distincttemporal kinetics and level of expression, rather than distinct signaling capacity of therespective B7.1 and B7.2 molecules.
IFN (pg/ml)
IL-4 (pg/ml)
12 x 106/ml APCs
1.3 x 106/ml APCs
Adapted from Schweitzer N.A. et al. J.Immunol. 158:2713, 1997.

The B7/CD28 pathway is much more complicated than expected
NOD mice develop diabetes at 10 weeks of age. This is the result of a T cell-dependent autoimmune process against the pancreatic beta cells.
B7-/- NOD:
Salomon, B., et al. Immunity 12:431-440, 2000
More severe diabetes!!
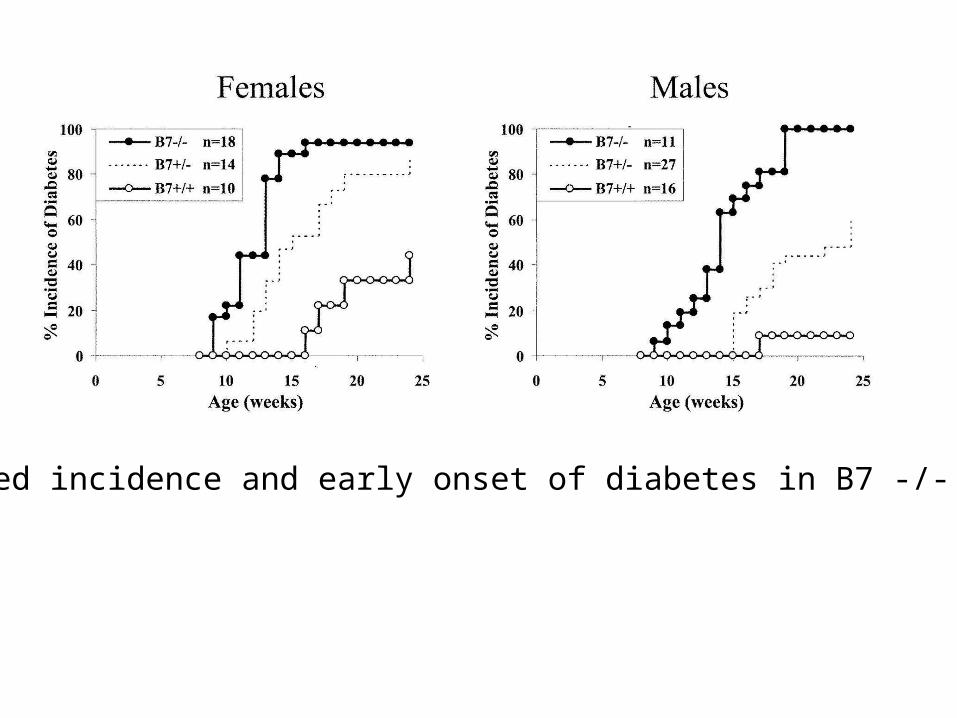
Increased incidence and early onset of diabetes in B7 -/- NOD mice

In the NOD model, Th1 cytokines such as IFNg are pathogenic while Th2 cytokines, like IL-4 could block disease development.
CD28 -/- NOD mice have impaired Th2 differentiation.
Hypothesis: Imbalance in Th1/Th2 differentiation might be a possible mechanism for exacerbation of diabetes in NOD B7-/- mice.
T cells from B7-deficient mice were stimulated with anti-CD3 mAbs, and supernatants were harvested to determine the levels of IL-4, IL-5, and IFNg production. The production of all cytokines was significantlyreduced as compared to cytokines produced by T cells from wild-type mice.
Isotype analysis of antibodies to glutamic acid decarboxylase (GAD), a potent autoantigen in autoimmunediabetes, showed that the levels of the Th1-dependent IgG2b isotype were severely decreased in these mice.The levels of GAD-specific Th2-dependent, IgG1 isotype gG1 antibodies were significantly reduced in five out of nine mice, but they were comparable to wild-type mice in the remaining four animals.
Thus, a selective decrease in Th2 cytokine production could not explain the increased disease severity in this B7-deficient NOD strain.

Virtually all islets isolated from 9-week-old prediabetic male and female B7-deficient mice were highlyinfiltrated and scored as severe insulitis. In contrast, in wild-type NOD mice, less than 10 % of the islets from female and only 3% of the islets from male mice displayed a severe insulitis.A kinetic analysis of islet infiltration in B7-deficient mice was carried out by scoring the intensity of insulitis at different time points in young prediabetic animalsA rapid increase in the levels of islet infiltration was observed by 9 weeks of age. The increased islet infiltration was not progressive as in control NOD mice butoccurred rapidly between 7 and 9 weeks of age. TThe accelerated kinetics of islet infiltration in B7-deficient mice suggested that a regulatory process that normally modulates the autoreactivity in the pancreas is lacking in B7 deficient NOD mice.

Previous studies have demonstrated the existence of regulatory CD4+CD25+ T cells that can prevent or limit autoimmunity. In the absence of this regulatory subset, normal mice develop several types of autoimmune disease spontaneously. In most mouse strains, this subset represents 7%–10% of the CD4+ T cell subset. In NOD mice, these cells are reduced in number, representing only 5%–6% of CD4+ T cells.The possibility that B7 expression might affect the regulatory CD4+CD25+ T cells in NOD mice was examined.A profound decrease of CD4+CD25+ T cells was observed in the B7-/- NOD mice. Less than 1% of CD4+ T cells expressed CD25 as compared to 5.4% in littermate B7+/+ controls.Thus, B7-/- mice are deficient in the immunoregulatory cells present in wild-type NOD mice, supporting a potential role for B7 engagement in CD4+CD25+ regulatory T cell development or homeostasis.
Profound absence of these cells also in CD28-deficient mice suggesting that the B7/CD28 interaction was critical for the homeostasis of these cells.
To determine if B7/CD28 costimulation is required continuously for the homeostasis of this T cell subset oris required for their development.Blockade of B7 stimulation in WT NOD miceresulted in decreased number of these cells and in exacerbated diabetes.

To determine whether the CD4+ CD25+ T cells control diabetes in CD28-deficient mice: Transfer of CD28-/- T cells into NOD.SCID. When CD4+ CD25+ are co-transferred, no diabetes. They also examined whether the transfer of a limited number of regulatory T cells directly into whole prediabetic CD28-deficient NOD mice could alter the progression of disease in these highly susceptible animals. All the CD28-deficient NOD mice either injected with PBS or with CD4+CD25- NOD T cells became diabetic by 11 weeks of age. In contrast, in four out of the six CD28-deficient mice, the transfer of CD4+CD25+ NOD T cells significantly delayed the development of diabetes. In two animals, the disease did not develop through 15 weeks of age.
Together, these studies suggest that the deficit of CD4+CD25+ regulatory T cells is responsible for the exacerbation of diabetes of the B7/CD28-deficient NOD mice.

Thus, in addition to a role in initial activation of T cells, B7/CD28 interactions are important in promoting T cell tolerance because they have a critical role in the homeostasis of CD4+ CD25+ regulatory T cells which play an essential role in regulating self-tolerance.