Embed Size (px)
Citation preview

J. Cell Sci. 38, 345-356 (1979) 3 4 5
Printed in Great Britain © Company of Biologists Limited 1979
THE TIME COURSE OF MALE MEIOSIS
IN THE RED-BACKED SALAMANDER,
PLETHODON CINEREUS
GARRY T. MORGAN*Department of Zoology, University of Leicester,Leicester, LEi 7RH, England
SUMMARY
Tritiated-thymidine autoradiography was used to follow the progress of cells throughmeiosis in male P. cinereus at 20 °C. Spermatocytes spend 7 days in leptotene, 5 days in zygoteneand 3 days in pachytene before entering the diffuse stage. Diffuse lasts for 8 days and is followedby a diplotene of 2 days. First and second meiotic metaphase occur a total of 26 days after theend of premeiotic S.
Considering the information for P. cinereus together with that for 3 other species, it appearsthat in amphibians the duration of meiosis does not show the linear relation with C-value thathas been described for other groups of organisms.
INTRODUCTION
The time course of male meiosis is known for only 3 amphibian species (Wimber& Prensky, 1963; Callan & Taylor, 1968; Kalt, 1976). I have attempted to time malemeiosis in another amphibian, Plethodon cinereus cinereus (Green), the red-backedsalamander. A study of the time course of meiosis in P. cinereus appeared especiallyuseful for 2 reasons:
(1) The total duration of meiosis and the times spent in particular meiotic stagesmay be very different in different organisms. Part of the diversity in total durationhas been explained as an effect of nuclear DNA content, or C-value (Bennett, 1971,1977). Comparison of widely unrelated diploid angiosperm species has shown thatthere exists a linear, positive relation between the duration of meiosis and nuclearDNA content (Bennett, 1971), just as the minimum cell cycle time of mitotically-dividing plant cells is linearly related to their nuclear DNA content (Van't Hof &Sparrow, 1963). Among animals also, it appears that the duration of meiosis may belinearly related to nuclear DNA content, although only if animals within the sameClass are compared (Bennett, 1977). Although Bennett (1977) has proposed that thisrelationship exists in mammals, insects and amphibians, data for just 2 amphibianspecies were available to him. Since the C-value of P. cinereus is accurately known(Mizuno & Macgregor, 1974), knowledge of the time course of its meiosis wouldallow further examination of the relation between C-value and meiotic duration inamphibian species.
* Present address: Department of Zoology, University of Washington, Seattle, Washington9819s. U.S.A.

346 G. T. Morgan
(2) In many organisms meiocytes pass through a period of chromosome deconden-sation after pachytene (Wilson, 1925; Klasterska, 1976). The decondensation may beslight and result merely in the appearance of ' fuzzy' bivalents or it may be extensiveand cause the virtual disappearance of bivalent axes from the nucleus. Stages ofchromosome decondensation are found during female meiosis (Darlington, 1937),including the best-known examples of the condition, the lampbrush chromosomesthat are found in the oocytes of many animals, especially those forming yolky eggs(Callan, 1957). Periods of well marked chromosome decondensation, or 'diffuse'stages, have been described in the developing asci of fungi (Barry, 1969), in pollenmother cells (Darlington, 1937; Klasterska, 1976) and during spermatogenesis inanimals (Darlington, 1937; Kezer & Macgregor, 1971; Fuge, 1976; Bogdanov, 1977).P. cinereus, like other plethodontid salamanders, possesses excellent oocyte lampbrushchromosomes (Vlad & Macgregor, 1975) and also provides a good example of thespermatocyte diffuse stage (Kezer, 1970; Kezer & Macgregor, 1971). The urodelelampbrush stage lasts for several months (Callan, 1963) and it seemed of interest tocompare the lengths of diffuse and lampbrush stages and to determine what effect thepresence of a diffuse stage has on the total duration of male meiosis.
There are several advantages in using P. cinereus as material for studying sperma-togenesis. In addition to the facts that the meiotic stages have been well studied and areeasily identifiable, the stages are serially arranged in the testis. The sequential arrange-ment of meiotic events results from the fact that meiotic divisions appear first at theposterior end of the testis and spread through the gonad as a caudocephalic 'wave'during a period of about 2 months. This feature made possible the unequivocaldemonstration of the diffuse stage in P. cinereus (Kezer & Macgregor, 1971) and washelpful in the interpretation of the meiotic timing study presented here. The timecourse of meiosis in P. cinereus was studied by an autoradiographic method that tracesthe fastest passage of spermatocytes through meiosis. Essentially, an injection of[3H]thymidine was given to a group of salamanders, and the progress of the label intomeiotic chromosomes of various stages was monitored in testes fixed at knownintervals after the injection.
MATERIALS AND METHODSSalamanders were collected during the spring in Shenandoah National Park, Va., U.S.A.
by Dr R. Highton of the University of Maryland, and in the Watchung Reservation, N.J.,U.S.A. by Dr J. Kezer of the University of Oregon. Meiotic divisions were present when theexperiment was begun in June. The animals were kept in a 20 ±0-5 °C incubator with a 16:8 hlight: dark cycle provided by a fluorescent tube, and were fed twice weekly with livingDrosophila.
[3H]thymidine was injected subcutaneously into the animals (average weight 05 g) on themid-ventral surface using a ioo-/tl Hamilton syringe. Each animal was given 25 /tCi of[Me-3H]thymidine (TRK 418, 46 Ci/mmol, from the Radiochemical Centre, Amersham,England). At daily intervals after injection of label, from +1 day to +37 days, animals werekilled by submersio'n in a solution of MS-222 (Sandoz). The testes were immediately removedand fixed in 3:1 absolute ethanol:glacial acetic acid. Testes were stored in fixative at about— 20 °C until the timing series had been completed.
Squash preparations were prepared as follows: each of the roughly cylindrical testes was

Time course of meiosis in P. cinereus 347
cut into 6-8 transverse slices with a razor blade and each slice placed in a drop of 45 % aceticacid on a siliconized coverslip. The testis fragments were carefully teased apart, and epitheliumand connective tissue removed. When a reasonably homogeneous cell suspension was obtained,the coverslip was inverted and dropped onto a subbed slide. The cells were squashed by theapplication of firm thumb pressure and the slides then placed on a bed of powdered dry-ice.After 5-10 min, the coverslips were flipped off with a razor blade, the preparations placed in95 % ethanol for 15-20 min and then allowed to dry in air.
For autoradiography, preparations were coated with Kodak NTB-2 liquid emulsion, (diluted1:1 with distilled water) at 45 °C. After exposing for 25 weeks, autoradiographs were developedin Kodak Dio,b for 2-5 min at 20 CC, rinsed in distilled water and fixed in Metafix (Kodak).Autoradiographs were stained with Giemsa (Improved R-66, Gurr) as follows: slides wereplaced in Coplin jars containing 6 ml of 0-07 M Na2HPO4, 4 ml of 0-07 M KH2PO4 and 90 mlof distilled water, final pH 68. About 3 ml of the Giemsa solution were added and the prepara-tions stained for 10-20 min; then they were air dried and left unmounted.
Tn order to determine whether [°H]thymidine was incorporated into cellular constituentsother than DNA, some air-dried preparations were treated before autoradiography with eitherDNase or RNase. DNA was digested in 100 /tg/ml DNase I E.P. (Sigma) in 0066 M phosphatebuffer (pH 6-2) with 0-005 M MgCl2 at 37 °C for 1 h. Preparations were washed after incubationin 2 changes each of 2 x SSC, 70 % and 95 % ethanol and then air dried (SSC = 015 M NaCl,0015 M Na3 citrate, pH 7-0). The RNase treatment involved incubating preparations in 50 fig/ml pancreatic RNase A (Sigma) plus 10 units/ml RNase Tj (Sigma) in 2 x SSC at 37 CC for1 h. The RNase had previously been boiled to inactivate contaminating DNase. After incuba-tion the slides were washed as above.
RESULTS
The sets of autoradiographs prepared from testes fixed at given intervals after labelinjection were each examined for the most advanced labelled meiotic stage. Thesequential arrangement of meiotic stages in the plethodontid testis means that in eachset of autoradiographs, the more posterior testis slices contain the more advancedstages. The most advanced labelled cells will be those that were completing premeioticS-phase when label became available, and they therefore possess less label thanless-advanced cells, which replicated for longer in the presence of [3H]thymidine.Accordingly, within each testis the most advanced labelled stage appeared as lightlylabelled nuclei that were posterior to more heavily labelled nuclei of the same orearlier stage, and anterior to unlabelled nuclei of the same or more advanced stage.
In autoradiographs from testes fixed 1 day after [3H]thymidine injection, labelledand unlabelled spermatogonia and spermatocytes were found. The amount of labellingover spermatogonial nuclei varied widely; some were completely black with silverwhile others showed a small number of scattered grains. Leptotene nuclei were themost advanced labelled meiotic stage and showed large differences in the amount anddistribution of label. Some possessed a light, uniform scatter of grains, and suchnuclei may indicate the spill-over of a small amount of DNA synthesis into earlyleptotene, as found in Triturus vulgaris (Callan & Taylor, 1968). In other lightlylabelled leptotene nuclei, silver grains were concentrated into patches that presumablyreflect the distribution of late-replicating regions of the genome (Fig. 1). This patternof patchy or 'late labelling' was usually found in the most advanced labelled cellsfrom +1 to +26 days. The stages reached by the most advanced labelled cells afterdifferent intervals between injection of label and testis fixation are given in Table 1

G. T. Morgan

Time course of meiosis in P. cinereus 349
(see Figs. 1-9). From this information the time course of male meiosis in P. cinereusat 20 °C can be deduced.
Leptotene lasts for 7 days and is followed by a zygotene stage that lasts for 5 days.Fully synapsed, pachytene bivalents are present for 3 days before they begin to lose
Table 1. Passage through meiosis of P. cinereus spermatocyteslabelled with [3H]thymidine during the premeiotic S-phase
No. of daysafter
injection
i
2
3456789
IO
II
1 2
1314
Stage reached by mostadvanced, labelled cells
\
Leptotene
1> Zygotene
J> Pachytene
No. of daysafter
injection
1516
i 718
i92O
2 1
2 2
23242526
2728
Stage reached by mostadvanced, labelled cells
Pachytene
Diffuse
} Diplotene
Metaphase I - Anaphase II
> Spermatid
their distinct axes at the onset of the diffuse stage. The transition to diffuse is notinstantaneous; intermediate stages are often seen in which chromosome decondensa-tion is not maximal or symmetrical over the whole nucleus. The extent of squashingand staining may affect the detailed appearance of diffuse nuclei, adding to the diffi-culty of assigning transitional nuclei to diffuse or pachytene. I have classified nucleias diffuse if some regions of their bivalents were markedly fuzzier than other regions.Spermatocytes spend 8 days in diffuse and then condensed bivalents appear at thebeginning of diplotene, which lasts for 2 days. In preparations made from testes fixed26 days after label injection, some first metaphase bivalents were patchily labelled
Figs, I - I I show [3H]thymidine incorporation by P. cinereus spermatocytes. Squashpreparations given autoradiographic exposures of 2'5 weeks. All scale bars represent10 fim.
Fig. 1. Leptotene nucleus with a small number of patches of label. The darklystained chromocentres are unlabelled. Testis fixed 1 day after injection of label.
Fig. 2. Leptotene nucleus. Chromocentres unlabelled, label confined to a smallnumber of patches. + 6 days.
Fig. 3. Leptotene nucleus with heavier labelling than that in Fig. 2. Chromocentreslabelled. +6 days.
Fig. 4. Early zygotene nucleus with patches of label over chromocentres. + 9 days.Fig. 5. Pachytene nucleus, patchily labelled. +14 days.
23 CEL 38
350 G. T. Morgan
6t • V •
" • & • • • • * • • , ; • . , .v- y;
• • ' • • • • • . . • • * • * • - . . * * • ,
7 *
0 V
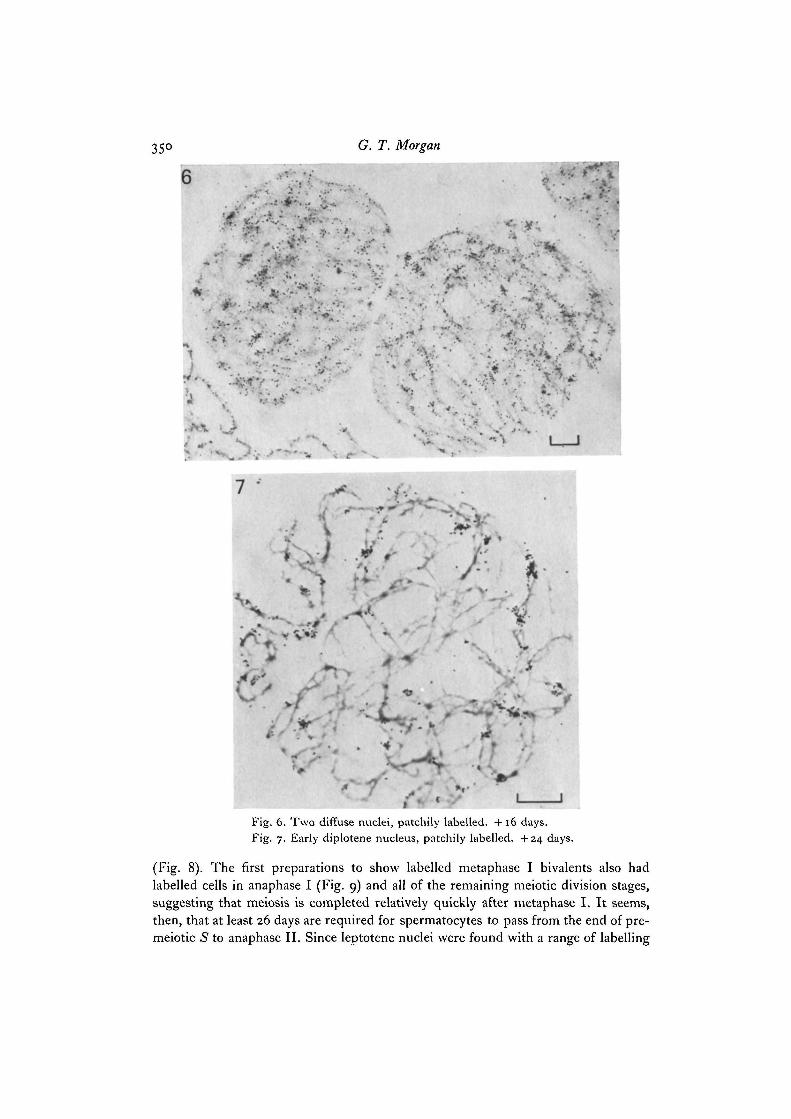
•4/-K-
Fig. 6. Two diffuse nuclei, patchily labelled. +16 days.Fig. 7. Early diplotene nucleus, patchily labelled. +24 days.
(Fig. 8). The first preparations to show labelled metaphase I bivalents also hadlabelled cells in anaphase I (Fig. 9) and all of the remaining meiotic division stages,suggesting that meiosis is completed relatively quickly after metaphase I. It seems,then, that at least 26 days are required for spermatocytes to pass from the end of pre-meiotic S to anaphase II. Since Ieptotene nuclei were found with a range of labelling

Time course of meiosis in P. cinereus 351
8
Fig. 8. First meiotic metaphase. Label confined to i region of i bivalent. + 26 days.Fig. 9. First meiotic anaphase and telophase (t) chromosomes with patchy labelling.
Asynchronous incorporation by a pair of half-bivalents is marked by arrows. + 26 days.Fig. 10. First meiotic metaphase bivalents labelled uniformly. +37 days.Fig. 11. Part of a second meiotic prophase with non-localized labelling of all chromo-
somes. Sister chromatids appear equally labelled (arrows). +37 days.23-2

352 G. T. Morgan
patterns just i day after label injection, it appears that premeiotic G2 is a short stageand that the total duration of meiosis in P. cinereus is about 26 days.
From + 27 days to + 37 days the most advanced labelled cells were early sperma-tids, although heavily and uniformly labelled metaphase and anaphase chromosomeswere also present (Fig. 10). A minimum estimate of the duration of the premeiotic5-phase can be made from the labelling patterns of sister chromatids (Callan &Taylor, 1968). Unfortunately, the +27 to +37 animals were well advanced intomeiosis when label was injected and yielded few labelled metaphases. However, thosesecond meiotic division chromosomes that were examined possessed equal amountsof labelling on both chromatids up to +37 days (Fig. 11). Even in the absence of a'chase' to limit carry-over of [3H]thymidine from one S-phase to another, one wouldexpect the amount of label incorporated by a given chromosome region during agonial 5-phase to be greater than that incorporated during a subsequent premeiotic5-phase. Therefore, the absence of label segregation in cells at + 37 days indicatesthat these cells were already in premeiotic 5-phase when label was injected. Sub-tracting the 26 days spent in meiosis, this suggests that premeiotic S-phase couldlast for a minimum of about 11 days. Another observation supports this suggestion.Diplotene and pachytene chromosomes that were late labelled were found in prepara-tions from testes fixed about 11 days after the first appearance of late-labelled cellsin these stages.
The chromocentres of prophase nuclei were not labelled in the most advancedlabelled cells of a given testis (Figs, i, 2) but were labelled in slightly less advancedcells (Fig. 3). In the diplotene, metaphase and anaphase chromosomes that firstappeared labelled, the patches of late label were usually found at non-centromericlocations. In slightly less advanced but labelled bivalents, centromeric regions werelabelled. The centromeric heterochromatin thus appears to complete its replicationlate in premeiotic S-phase, but not right at the end.
Two potential sources of error in this experiment should be recognized: (1)[3H]thymidine can become incorporated into cellular constituents other than DNAin some situations (Goldspink & Goldberg, 1973; Ito & Hotta, 1973). To check thatthe label incorporated during premeiotic 5-phase in this experiment was exclusivelylocated in DNA, some preparations were treated with either RNase or DNase. Thosetreated with RNase showed the same amount and distribution of label as untreatedpreparations, but all of the label was removed from preparations that were treated withDNase. (2) If the small amounts of DNA synthesis that occur during meiotic pro-phase (Stern & Hotta, 1977) incorporate autoradiographically detectable amounts of[3H]thymidine, then the duration of early meiotic stages will be underestimated.Three observations suggest that such incorporation does not occur in this experi-mental system. First, in the timing experiment labelled zygotene and pachytenenuclei were not found until +8 and +13 days, respectively. Secondly, labelledmetaphases were not found in preparations fixed from +13 to +23 days. Since ittakes only about 13 days for spermatocytes to progress from the end of pachytene tometaphase I, any label incorporated during zygotene or pachytene would be expectedto produce labelled metaphase I bivalents before the 5-phase label reached metaphase

Time course of meiosis in P. cinereus 353
at 4- 26 days. Lastly, a preliminary experiment was undertaken in which salamanderseach received 2 injections of 50 /tCi of [3H]thymidine (4 times the dose used in thetiming experiment) 18 h apart, and then were killed 9 h after the second injection.No labelling of meiotic stages later than early leptotene was found in autoradiographsexposed for up to a year. Gonial and premeiotic /S-phase nuclei were heavily labelledafter 2-day exposures.
DISCUSSION
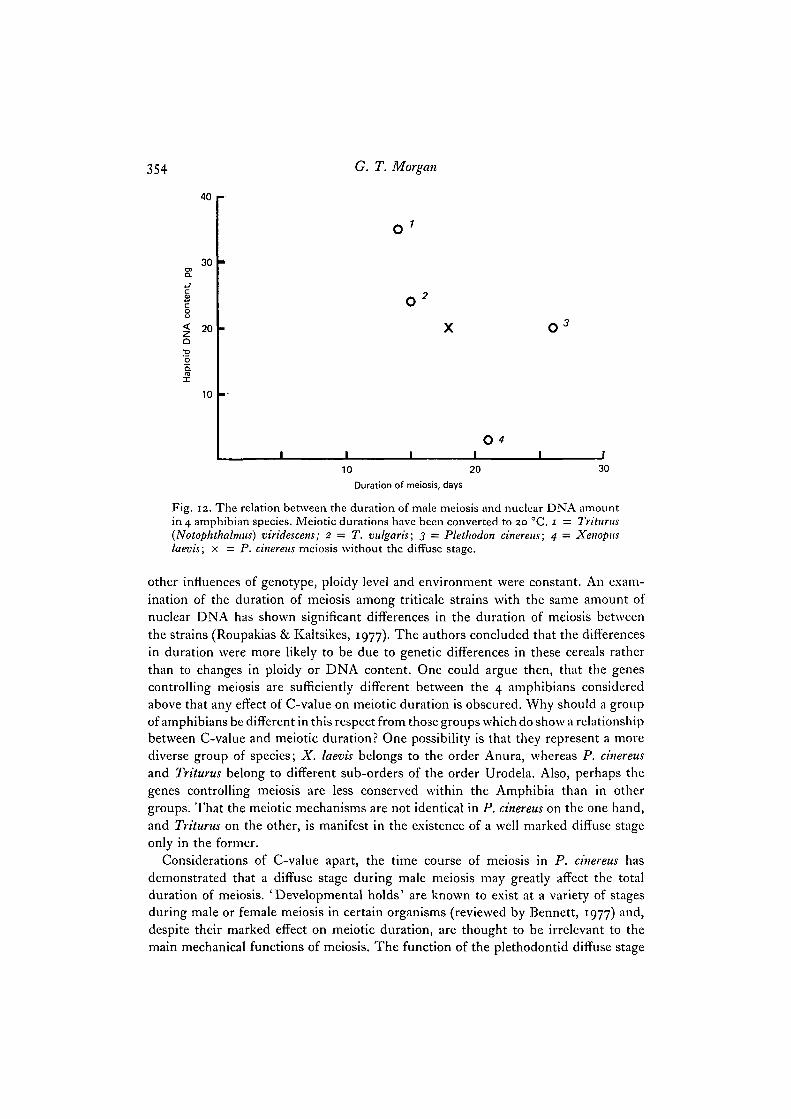
The results obtained have shown that male meiosis in P. cinereus at 20 °C takes atleast 26 days to complete. One of the reasons for my attempting this study was todetermine whether information obtained from amphibians supports the proposalthat C-value and meiotic duration are positively and linearly related in various groupsof organisms (Bennett, 1971, 1977)- The time required to complete male meiosis hasbeen measured in 3 other amphibian species. Meiosis takes 20-21 days in Triturusvulgaris at 16 °C (Callan & Taylor, 1968), 13 days in Triturus (= Notophthalmus)viridescens at 20-22 °C (Wimber & Prensky, 1963) and 24 days in Xenopus laevis at18 °C (Kalt, 1976). In order for these data to be strictly comparable they must beconverted to the same temperature since an increase in temperature decreases theduration of meiosis over the temperature range in which organisms can survive(Bennett, 1977). The Q10 values obtained for meiosis in animals are about 2-0-2-3(Bennett, 1977). Applying this as an approximate conversion factor, at a temperatureof 20 °C meiosis would take about 15 days in T. vulgaris, 14 days in T. viridescens and21 days in X. laevis. The haploid nuclear DNA contents of the 4 amphibians forwhich the duration of meiosis is known, are as follows: P. cinereus, 20 pg (Mizuno &Macgregor, 1974); T. vulgaris, 24 pg (Olmo, 1973, 1974); T. viridescens, 35 pg(Olmo, 1973, 1974); X. laevis, 3-2 pg (Thiebaud & Fischberg, 1977). Thus as far asthese amphibians are concerned, the duration of meiosis is not positively correlatedto C-value (Fig. 12). For instance, X. laevis has a genome one tenth the size of T. viri-descens but takes much longer to complete meiosis. Similarly, P. cinereus takes longerto complete meiosis than either Triturus species, despite its smaller C-value. Thelonger meiosis in P. cinereus may be attributed in part to the presence of an 8-daydiffuse stage, which is not found in the Triturus species. However, even if P. cinereusspermatocytes passed immediately from pachytene to diplotene, meiosis would lastfor about 18 days at 20 °C, still appreciably longer than in both Triturus species at thattemperature.
The precise relationship between nuclear DNA content and the duration of meiosisthat exists in diploid angiosperms has been interpreted as demonstrating that themass of DNA per nucleus plays a major role in determining the duration of meiosis(Bennett, 1971, 1977). However, Bennett (1971, 1977) has also pointed out thatgenotype, ploidy level and environmental conditions also influence the rate of meioticdevelopment, the duration of meiosis in a given species being determined by a complexinteraction of these factors. The comparison of meiotic durations and C-values fora group of organisms might then be expected to reveal a good correlation only if the
354 G. T. Morgan
40
30
20
10
10 20
Duration of meiosis, days
30
Fig. 12. The relation between the duration of male meiosis and nuclear DNA amountin 4 amphibian species. Meiotic durations have been converted to 20 °C. 1 = Triturus(Notophthalmus) viridescens; 2 — T. vulgaris; 3 = Plethodon cinereus; 4 — Xenopuslaevis; x = P. cinereus meiosis without the diffuse stage.
other influences of genotype, ploidy level and environment were constant. An exam-ination of the duration of meiosis among triticale strains with the same amount ofnuclear DNA has shown significant differences in the duration of meiosis betweenthe strains (Roupakias & Kaltsikes, 1977). The authors concluded that the differencesin duration were more likely to be due to genetic differences in these cereals ratherthan to changes in ploidy or DNA content. One could argue then, that the genescontrolling meiosis are sufficiently different between the 4 amphibians consideredabove that any effect of C-value on meiotic duration is obscured. Why should a groupof amphibians be different in this respect from those groups which do show a relationshipbetween C-value and meiotic duration? One possibility is that they represent a morediverse group of species; X. laevis belongs to the order Anura, whereas P. cinereusand Triturus belong to different sub-orders of the order Urodela. Also, perhaps thegenes controlling meiosis are less conserved within the Amphibia than in othergroups. That the meiotic mechanisms are not identical in P. cinereus on the one hand,and Triturus on the other, is manifest in the existence of a well marked diffuse stageonly in the former.
Considerations of C-value apart, the time course of meiosis in P. cinereus hasdemonstrated that a diffuse stage during male meiosis may greatly affect the totalduration of meiosis. 'Developmental holds' are known to exist at a variety of stagesduring male or female meiosis in certain organisms (reviewed by Bennett, 1977) and,despite their marked effect on meiotic duration, are thought to be irrelevant to themain mechanical functions of meiosis. The function of the plethodontid diffuse stage

Time course of meiosis in P. cinereus 355
is unknown. On the basis of light-microscope observations, Kezer & Macgregor (1971)have suggested that the spermatocyte diffuse stage represents a situation comparableto the lampbrush condition of amphibian oocytes. In support of this comparison,autoradiographic analysis of [3H]uridine incorporation during meiotic prophase inplethodontid spermatocytes has suggested that more RNA is synthesized during thediffuse stage than in other prophase stages (Owen, 1973; P.Leon, personal com-munication). However, the ultrastructure of transcription appears different in lamp-brush and diffuse nuclei (Morgan, unpublished observations), and the 2 stages clearlydiffer with respect to their duration. The urodele lampbrush stage has been estimatedto last for a minimum of about 200 days (Callan, 1963), whereas I have found thespermatocyte diffuse stage to have a duration of just 8 days. A detailed comparisonof chromosome organization during diffuse and lampbrush stages appears necessaryin determining to what extent the 2 stages may be considered equivalent.
I am grateful to Dr R. Highton and Dr J. Kezer for providing the animals used in this study.This work was carried out whilst in receipt of a Science Research Council studentship.
REFERENCES
BARRY, E. G. (1969). The diffuse diplotene stage of meiotic prophase in. Neurospora. Chromo-soma 26, 119-129.
BENNETT, M. D. (1971). The duration of meiosis. Proc. R. Soc. B 178, 277-299.BENNETT, M. D. (1977). The time and duration of meiosis. Phil. Trans. R. Soc. Ser. B 277,
201-226.BOGDANOV, Yv. F. (1977). Formation of cytoplasmic synaptonemal-like polycomplexes at
leptotene and normal synaptonemal complexes at zygotene in Ascaris suum male meiosis.Chromosoma 61 , 1—21.
CALLAN, H. G. (1957). The lampbrush chromosomes of Sepia, Anilocra and ScyIlium and ofAmphibia. Pubbl. Staz. zool. Napoli 29, 329-346.
CALLAN, H. G. (1963). The nature of lampbrush chromosomes. Int. Rev. Cytol. 15, 1-34.CALLAN, H. G. & TAYLOR, J. H. (1968). A radioautographic study of the time course of male
meiosis in the newt Tritimis vulgaris. J. Cell Sci. 3, 615-626.DARLINGTON, C. D. (1937). Recent Advances in Cytology, 2nd edn. London: Churchill.FUGE, H. (1976). Ultrastructure of cytoplasmic nucleolus-like bodies and nuclear RNP particles
in late prophase of tipulid spermatocytes. Chromosoma 56, 363-379.GOLDSPINK, D. F. & GOLDBERC, A. L. (1973). Problems in the use of [Me-3H]-thymidine for
the measurement of DNA synthesis. Biochim. biophys. Acta 299, 521-532.ITO, M. & HOTTA, Y. (1973). Radioautography of incorporated 3H-thymidine and its meta-
bolism during meiotic prophase in microsporocytes of Lilium. Chromosoma 43, 391-398.K.ALT, M. R. (1976). Morphology and kinetics of spermatogenesis in Xenopus laevis. J. exp.
Zool. 195, 393-408.KEZER, J. (1970). Observations on salamander spermatocyte chromosomes during the first
meiotic division. Drosoph. Inf. Serv. 45, 194-200.KEZER, J. & MACGREGOR, H. C. (1971). A fresh look at meiosis and centromeric heterochroma-
tin in the red-backed salamander, Plethodon cinereus cinereus (Green). Chromosoma 33,146-166.
KLASTERSKA, I. (1976). A new look on the role of the diffuse stage in problems of plant andanimal meiosis. Hereditas 82, 193-204.
MIZUNO, S. & MACGRECOR, H. C. (1974). Chromosomes, DNA sequences and evolution insalamanders of the genus Plethodon. Chromosoma 48, 239-296.
OLMO, E. (1973). Quantitative variations in the nuclear DNA and phylogenesis of the Amphibia.Caryologia 26, 43-68.

356 G.T. Morgan
OLMO, E. (1974). Further data on the genome size in the urodeles. Boll. Zool. 41, 29-33.OWEN, C. A. (1973). The Cytology of Meiosis in Male Salamanders. M. Phil. Thesis, University
of Leicester, England.ROUPAKIAS, D. G. & KALTSIKES, P. J. (1977). Genomic effects on the duration of meiosis in
triticale and its parental strains. Can. J. Genet. Cytol. 19, 331-343.STERN, H. & HOTTA, Y. (1977). Biochemistry of meiosis. Phil.Trans. R.Soc.Ser.B 277,277-294.THIEBAUD, C. H. & FISCHBERG, M. (1977). DNA content in the genus Xenopus. Chromosoma
59, 253-257.VAN'T HOF, J. & SPARROW, A. H. (1963). A relationship between DNA content, nuclear volume
and minimum mitotic cycle time. Proc. natn. Acad. Sci. U.S.A. 49, 897-902.VLAD, M. & MACGREGOR, H. C. (1975). Chromomere number and its genetic significance in
lampbrush chromosomes. Chromosoma 50, 327-347.WILSON, E. B. (1925). The Cell in Development and Heredity, 3rd edn. New York: Macmillan.WIMBER, D. E. & PRENSKY, W. (1963). Autoradiography with meiotic chromosomes of the
male newt (Triturus viridescens) using 3H-thymidine. Genetics, Princeton 48, 1731-1738.
{Received 13 February 1979)





























