Embed Size (px)
Citation preview

The roles of microorganisms in litter decomposition and soilformation
Satoru Hobara • Takashi Osono • Dai Hirose •
Kenta Noro • Mitsuru Hirota • Ronald Benner
Received: 1 August 2012 / Accepted: 15 September 2013 / Published online: 9 October 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Much has been learned about the micro-
bial decomposition of plant litter, but relatively little is
known about microbial contributions to litter and soil
chemistry. We conducted a 3-year litterbag experi-
ment and measured hydrolyzable amino acids (AA)
and amino sugars (AS) to gain insights about micro-
bial contributions to the chemical characteristics of
decomposing litter and soil. Microscopic observations
of hyphae were used to estimate fungal contributions
to litter. The carbon (C)-normalized yields of AA and
AS increased during decomposition along with nitro-
gen (N), indicating a shift in chemical characteristics
from C-rich plant-derived biopolymers to N-rich,
microbially-derived biochemicals. The contributions
of fungal biomass to C and N were minor, but
necromass of fungi as melanized and clamp-bearing
hyphae increased during litter decomposition. Yields
of glucosamine and galactosamine in litter approached
those in microorganisms, particularly bacteria, sug-
gesting major contributions of bacterial residues to
litter during decomposition. The microbial contribu-
tions to decomposing litter were consistent with those
observed in organic and mineral soils. Microorgan-
isms play important roles in the organization and
stabilization of soil organic matter as well as N
immobilization and organic C preservation.
Keywords Litter decomposition � Carbon and
nitrogen dynamics �Amino sugars �Amino acids �Fungi � Bacteria
Responsible Editor: Kathleen Lohse
Electronic supplementary material The online version ofthis article (doi:10.1007/s10533-013-9912-7) contains supple-mentary material, which is available to authorized users.
S. Hobara (&) � K. Noro
Department of Environmental and Symbiotic Science,
Rakuno Gakuen University, 582 Midorimachi,
Bunkyodai, Ebetsu 069-8501, Japan
e-mail: [email protected]
S. Hobara � R. Benner
Department of Biological Sciences, University of South
Carolina, Columbia, SC 29208, USA
T. Osono
Center for Ecological Research, Kyoto University,
Otsu 520-2113, Japan
D. Hirose
College of Pharmacy, Nihon University, Funabashi,
Chiba 274-8555, Japan
M. Hirota
Graduate School of Life and Environmental Sciences,
University of Tsukuba, Tsukuba 305-8572, Japan
123
Biogeochemistry (2014) 118:471–486
DOI 10.1007/s10533-013-9912-7

Abbreviations
AA Amino acid
AS Amino sugar
THAA Total hydrolyzable amino acid
THAS Total hydrolyzable amino sugar
GlcN Glucosamine
GalN Galactosamine
OC Organic carbon
Introduction
Litter decomposition in terrestrial ecosystems is an
important process for nutrient regeneration, maintaining
productivity and influencing ecosystem structure (Swift
et al. 1979). The process of litter decomposition is
closely associated with microbial activities, which alter
the chemical composition of litter and control carbon
(C) and nitrogen (N) dynamics in soil (Berg and
McClaugherty 2008). Hence, investigations of microbial
contributions to litter chemistry during decomposition
are essential for fully understanding the mechanisms of
litter decomposition and C and N dynamics in soil.
The enzymatic activities of microorganisms play a
major role in the decomposition of plant litter and soil
formation (Aber and Melillo 2001; Berg and McClaugh-
erty 2008), whereas much less is known about microbial
contributions to litter chemistry. Microbial growth and
the resulting biomass and necromass significantly alter
the chemical characteristics of soil organic matter, as
observed in nutrient immobilization (Hart et al. 1994;
Bengtsson et al. 2003; Tremblay and Benner 2006;
Simpson et al.2007; Wanek et al. 2010; Liang and Balser
2011; Cotrufo et al. 2013). In addition, the transforma-
tions and fates of microbial biomass and necromass
affect soil formation, stabilization, and fertility (Six et al.
2006; Fontaine et al. 2011; Miltner et al. 2011).
The effects of microbial growth and decomposition
processes can be traced through measurements of
amino acids (AA) and amino sugars (AS), biomole-
cules that are much more abundant in microbes than in
plant litter (Amelung et al. 2001; Tremblay and
Benner 2006). The composition and yields of AA and
AS in litter changes with sources and decomposition
processes, and some of these biomolecules, such as
muramic acid (MA), are uniquely found in specific
microorganisms (Parsons 1981; Benner and Kaiser
2003; Amelung et al. 2008). Furthermore, AA and AS
are important components of N immobilization
(Marumoto et al. 1982; Mengel 1996; Tremblay and
Benner 2006; He et al. 2011a, b) and C sequestration
(Glaser et al. 2006; Liang et al. 2007; Knicker 2011) in
soils. Therefore, the characterization of AA and AS in
decomposing litter provides important information
about microbial contributions to litter chemistry and
the biogeochemical processes leading to soil forma-
tion. However, the characterization of AA and AS has
been performed mostly for bulk litter and/or soil
samples (Solomon et al. 2002; Ding et al. 2010), and
few studies have examined relationships between litter
and microbial characteristics during decomposition
processes (Guggenberger et al. 1999).
We hypothesize that microorganisms play major
roles in both litter decomposition and soil formation.
In the present study, we measured total hydrolyzable
AA and AS in decomposing plant litter from three
plant species, and compared them with analyses of
soils (organic and mineral) and microorganisms (fungi
and bacteria) to investigate the origins and dynamics
of these major biochemicals during the decomposition
process. Fungi typically dominate soil microbial
biomass during the early stages of litter decomposition
(Berg and McClaugherty 2008). Thus, we also
estimated the biomass and necromass of fungi to
further elucidate their contributions to litter chemistry.
Materials and methods
Study sites, litterbag experiment, and sample
collections
Litterbag experiments were carried out in grasslands
and forests of Sugadaira Montane Research Center,
University of Tsukuba, Nagano, Japan (368300N,
1288200E). The soil was originally derived from
volcanic ash, classified as Typic Melanudand (Iimura
et al. 2010). Three sites that were adjacent to each
other but dominated by different plant species were
selected for the litterbag experiments: a grassland
dominated by Miscanthus sinensis, a coniferous
evergreen forest dominated by Pinus densiflora, and
a deciduous broad-leaf forest dominated by Quercus
crispula. We conducted the litterbag experiments at
these sites using litter of the dominant species. These
different plant species were chosen to gain a more
comprehensive understanding of biogeochemical pro-
cesses during litter decomposition and soil formation.
472 Biogeochemistry (2014) 118:471–486
123

Further detailed description of the study site and the
soils is provided in (Kato and Hayasi (2006) and
Iimura et al. (2010).
Freshly fallen leaves of the dominant plant species
(Miscanthus sinensis, Pinus densiflora, or Quercus
crispula) were collected from the soil surface at each
site during November 2003. The leaves were dried in
an oven at 40 �C for 1 week. Litter (*2 g) was placed
in litterbags (18 cm 9 18 cm) made of polypropylene
shade cloth with a mesh size of approximately 2 mm. A
total of 420 bags were prepared. Approximately 20 g
of each litter sample was stored for chemical analyses.
The decomposition experiment covered a 36-month
period from November 2003 to November 2006. A
transect was established at each site, and 10 subplots
(2 m 9 2 m) were located at 1 m intervals along the
transect, making a total of 30 subplots for the three
sites. Litterbags were place on the litter layer in
November 2003, with 14 bags per subplot. The
litterbags were fixed to the litter layer using metal
pins to prevent movement and to ensure good contact
between the bags and the litter layer. Sampling of the
bags took place 7 times: 6, 9, 12, 18, 21, 24, and
36 months after initiation of the experiment. On each
sampling occasion, 20 bags were collected for each
plant species from 10 subplots (2 bags each) and
transported them to the laboratory.
One bag from each subplot was used for estimation
of hyphal mass (see below), and another bag was used
for mass determination and chemical analyses. Sam-
ples were dried to a constant mass at 40 �C, and mean
values of mass loss were calculated for each sampling.
The subsamples for ash and chemical analyses were
ground in a laboratory mill to pass a 0.5-mm screen
and combined prior to analyses.
Soil and fungi samples were also analyzed to
compare their chemical compositions with those of
decomposing litter. Soils were sampled from the
organic (Oe?Oa) horizon and the mineral soil A
horizon in each site. After sieving with a 2 mm mesh
screen, soils were dried at 40 �C for 1 week, and
ground for analyses. One of the dominant fungal
species at all sites, Nigrospora sphaerica (Osono
2010), was grown in the laboratory for chemical
analyses. Nigrospora sphaerica is the dominant fungal
species colonizing decomposing leaves. We chose this
species because this was the only fungi observed in all
study sites throughout the experiment (T. Osono
unpublished data). The frequency of occurrence of
this species in litter was 75–90 % for Miscanthus
sinensis (Osono 2010), 20–25 % for Pinus densiflora,
and 25–45 % for Quercus crispula (T. Osono unpub-
lished data). Three isolates of N. sphaerica were
obtained from Miscanthus sinensis leaves, and grown
as a pure culture in Czapek medium (NaNO3 0.2 %,
K2HPO4 0.1 %, MgSO4�7H2O 0.05 %, KCl 0.05 %,
FeSO4 0.001 %, and sucrose 3 %). We used chemi-
cally defined, synthetic Czapek medium, instead of
natural medium, to incubate N. sphaerica isolates to
avoid the contamination of amino acids in the
medium. The cultured fungal biomass was harvested,
washed with sterile distilled water, dried at 40 �C, and
ground for analyses. We also analyzed a natural fungal
community, Cortinarius armillathus, and cultures of
Micrococcus lysodeikticus ATCC 4698 and cultures of
Aerobacter aerogenes and Azobacter vinelandii.
These bacterial cultures were obtained Sigma
Chemicals.
Hyphal length
Hyphal length in litter was measured by the agar-film
method in litter samples collected at 12, 24 and
36 months (Osono and Takeda 2001). Briefly, a
subsample (1 g) from each litterbag was homogenized
in a blender at 10,000 rev/min in 49 ml of deionized
water for 3 min. The suspension (20 ml) was diluted
with 20 ml of molten agar solution (final concentra-
tion 1.5 % (w/v)) and mixed at low speed on a
magnetic stirring plate. Three agar films were prepared
for each suspension in a haemocytometer (0.1 mm
depth), transferred to a glass slide, and dried for 24 h.
The films were dual-stained with fluorescent bright-
ener (FB) and acridine orange (AO) each for 1 h.
The stained films were mounted on slides with one
drop of immersion oil and examined with an epiflu-
orescent microscope (Nikon Microphot-SA, Nikon,
Tokyo). Darkly pigmented hyphae that were not
stained with FB were observed by bright-field micros-
copy. Microscope fields were selected randomly and
25 fields were observed for each slide. Total hyphal
length was calculated as the sum of FB-stained and
darkly pigmented hyphal lengths. We used following
definitions for fungal hyphae in this study: AO-stained
hyphae as living hyphae (Ono 1998); darkly-pig-
mented hyphae as melanized hyphae; clamp-bearing
hyphae as basidiomycetous hyphae. The Basidiomy-
cota includes such functional groups as lignin
Biogeochemistry (2014) 118:471–486 473
123

decomposing and mycorrhizal fungi. Hence, the
amount of clamp-bearing hyphae can represent
potential functional capabilities of fungal communi-
ties. It is recognized, however, that the fungal biomass
of the Basidiomycota may have been underestimated
because the frequency of clamp formation varies
between species.
Estimation of hyphal mass and AA, GlcN,
and GalN as hyphae
We used a conversion from hyphal length to hyphal
mass for estimating the AA, glucosamine (GlcN), and
galactosamine (GalN) contents of hyphae. We con-
verted hyphal length to hyphal mass according to the
equation in Paul and Clark (1989):
F ¼ pr2L D S� 102
where F is hyphal mass (g g-1 (w/w)), r is average
hyphal radius (cm) (0.00011 for Miscanthus sinensis
litter, 0.000131 for Pinus densiflora litter, 0.000135
for Quercus crispula litter), L is hyphal length
(m g-1), D is hyphal density (g cm-3) (1.1, from
Saito (1955)), and S is solid content (0.15, from (Baath
and Soderstrom (1977)). Total hydrolyzable AA,
GlcN, and GalN yields as hyphae in litter were
calculated multiplying the hyphal mass in litter by AA,
GlcN, and GalN yields in the Nigrospora culture,
respectively. In this study, we defined living hyphal
mass as fungal biomass and the remnant hyphae (total
minus living hyphae) as fungal necromass.
Ash and chemical analyses
Ash determinations were made by combusting subs-
amples from each litterbag at 800 �C for 4 h.
Elemental C and N contents were measured using a
CHN analyzer (Sumigraph NC-22, Sumitomo Chem-
ical Co., Osaka, Japan). Weight losses and elemental
compositions are reported on an ash-free dry weight
(AFDW) basis.
Total hydrolyzable amino acids (THAA) were
analyzed in dried samples (*5 mg; decomposing
litter, soils, fungal body, and references of other
microorganisms) that were placed in a sealed vacuum-
reaction-tube with 6 M HCl at 110 �C for 20 h.
Following hydrolysis, amino acids were analyzed
using the HPLC AccQ method (Reverter et al. 1997).
Separations were carried out on a reversed-phase
column (AccQ Tag C18, Waters Japan, Tokyo) heated
to 40 �C with a flow rate of 1.0 ml min-1 and eluted
with mobile phase A (0.04 M KH2PO4) and mobile
phase B (acetonitrile/water = 60/40 (v/v)) using a
linear gradient program. The following amino acids
were analyzed: alanine, arginine, aspartic acid, glu-
tamic acid, glycine, histidine, isoleucine, leucine,
lysine, methionine, phenylalanine, proline, serine,
threonine, tyrosine, and valine.
Total hydrolyzable amino sugars (THAS) were
analyzed in dried samples (*5 mg) placed in sealed
ampules with 3M HCl at 100 �C for 5 h (Kaiser and
Benner 2000). Acetyl groups are removed from amino
sugars during acid hydrolysis, so concentrations of the
deacetylated forms are presented. Hydrolyzed samples
were centrifuged, and the supernatant was neutralized
with the self-absorbed AG 11 A8 resin (Bio-Rad).
After desalting using the cation-exchanger AG50 X8
resin (Na?-form, Bio-Rad), concentrations of GalN,
GlcN, and mannosamine (ManN) were measured
using high-performance anion exchange chromatog-
raphy (HPAEC) with pulsed amperometric detection
(PAD). Although GalN and MA are assumed to
originate mostly from bacteria (Parsons 1981), we
used GalN as a tracer of bacteria in this study because
it is more abundant and persistant in litter and soil than
MA (Amelung 2003, Tremblay and Benner 2006,
Engelking et al. 2007). A Dionex ion chromatography
system with a CarboPac PA1 column with guard
column and PAD detector were used for all analyses.
Separation was performed under isocratic conditions
with a 12 mM NaOH mobile phase at a flow rate of
1 ml min-1. In the present paper, we do not report
ManN concentrations because they were low and often
below the level of quantification. Although GlcN/
GalN ratio has been used for evaluating the influence
of fungi, we used GlcN/GalN ratio as a diagenetic
indicator in this study because this ratio decreases with
decomposition in both soil (Amelung et al. 2001) and
water (Tremblay and Benner 2006), irrespective of the
presence of fungi.
The C- and N-normalized yields of AA and AS was
used in this study because they trace the fraction of
total C and N in litter and soil as these biochemicals. A
degradation index (DI) based on the composition of
protein amino acids was used to investigate the extent
of diagenetic alteration in decomposing litter and soils
(Dauwe et al. 1999).
474 Biogeochemistry (2014) 118:471–486
123

The percentage of detrital N in litter and soils
derived from microorganisms were quantified using
the following equation (Tremblay and Benner 2006):
%Nimmob ¼ 100 Nsample � Nplant
� �=Nsample;
where Nimmob is the fraction of N immobilized by
microbes, Nsample is the total N content of the sample,
and Nplant is the total N content in the plant litter. The
Nplant from the remaining at time t was determined
assuming plant N had the same reactivity as plant C
(indicated from litter C content at time t (Ct) and fresh
litter C content (C0)) during decomposition:
Nplant;t ¼ Nplant;0 Ct=C0ð Þ:
One-way analysis of variance (ANOVA) was used
to compare the yields of AA and AS (average of all
sites) between fresh (initial) and decomposed
(36 months) litter. Correlation and regression analyses
were performed using JMP 8.0.2 software (SAS
Institute Japan).
Results
Bulk parameters
Changes in bulk parameters during the experiments
were variable among plant species (Table 1). Rates of
mass loss (AFDW) from litterbags were lower during
winter, but seasonal effects on mass loss were
relatively minor. Quercus litter showed higher losses
(89 %) during 36 months than Miscanthus (71 %) and
Pinus litter (67 %). The carbon content of litter
decreased with time for Miscanthus (*7 % during
36 months) and Quercus (*2 % during 36 months)
litter, but remained fairly constant for Pinus litter. The
losses of C from decomposing litter ranged from 67 to
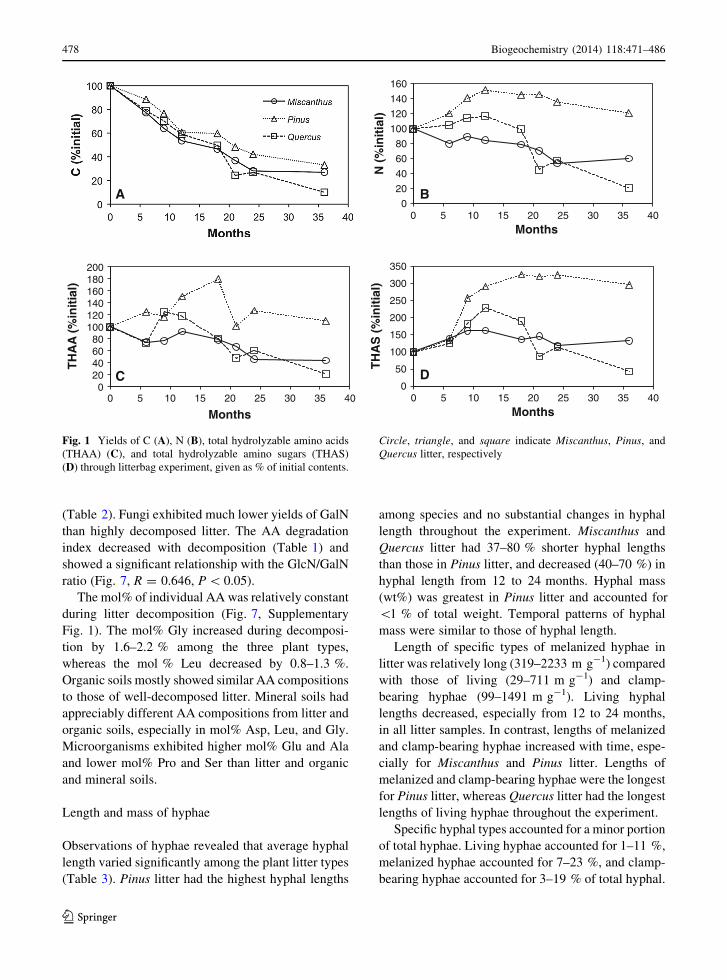
90 % (Fig. 1A).
The N content of litter increased up to four fold
during the experiments (Table 1), leading to net
increases in the % initial N, especially during the first
year (Fig. 1B). Pinus litter N increased to 152 % of the
initial N in the first year and was 121 % at the end of
the experiment. The % initial N for Quercus litter also
increased during the first 12 months of decomposition,
but decreased rapidly to 21 % at the end of the study.
The pattern of N dynamics in the Miscanthus litter was
quite different and steadily declined during the study
to 60 % of the initial N content after 36 months
(Table 1).
Molar ratios of C/N in the three litter types
decreased gradually with time (Table 1). Variability
among C/N ratios of the initial litter samples ranged
from 58.9 to 148.8 and decreased to 23.7–40.6 by the
end of the experiment. The C/N values for the highly
decomposed litter samples (14.6–32.8) were
approaching those in organic and mineral soils. Fungi
had C/N ratios of 14.2–21.0, which were similar to
those in litter and soil samples, whereas bacteria had
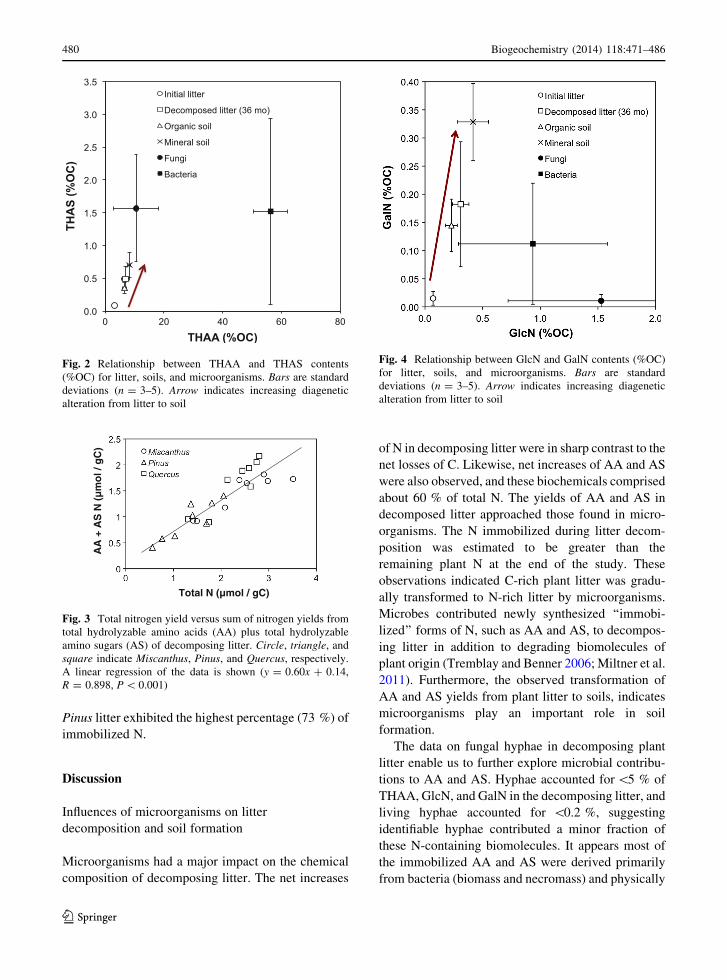
much lower C/N ratios (3.7–4.7) (Table 2).
Amino acids and amino sugars yields in litter, soil
and fungi
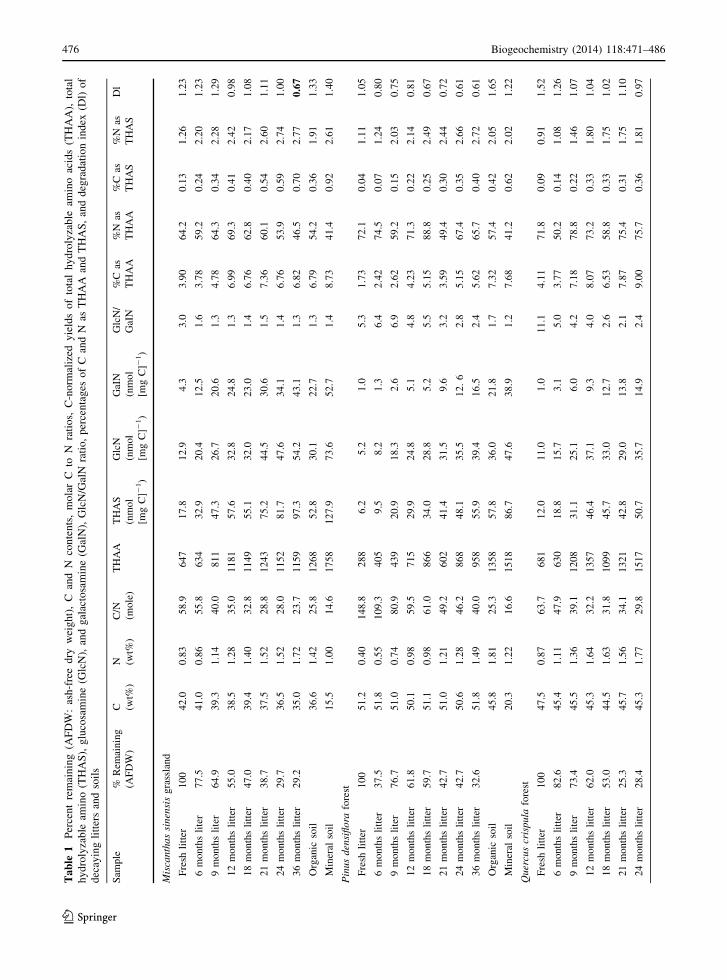
The C-normalized yields of THAA were 12- to 57-fold
higher than those of THAS for litter and soil samples
(Table 1). Pinus litter had lower THAA and THAS
yields than Miscanthus and Quercus. The percentage
of C as AA and AS in litter samples ranged from 1.73
to 9.00 % and from 0.04 to 0.70 %, respectively, and
the percentage of N as AA and AS ranged from 46.5 to
88.8 % and from 0.91 to 2.77 %, respectively. The
yields of THAA and THAS in litter increased during
the experiment, and appeared to approach those of
soils during decomposition. Significant increases
(P \ 0.05 by ANOVA) in yields of THAA and THAS
were observed during litter decomposition. The
increase was greater in THAS (4- to 9-fold) than
THAA (2- to 3-fold) and was particularly high in
Pinus litter. The yields of THAA and THAS in soils
were similar to those of more highly decomposed litter
and were higher in the mineral soil layer than the organic
layer. Nigrospore hyphae, the most abundant fungal
hyphae at the study site, exhibited 1.7- to 9.0-fold higher
THAS yields than those in litter (Tables 1 and 2). In
contrast, the THAA yield in Nigrospore hyphae
(970 nmol [mg C]-1) was in the middle of the range
of values in litter (288–1517 nmol [mg C]-1) (Table 2).
Bacteria had 8- to 43-fold higher THAA yields than
those in litter and were generally higher than THAS
yields in litter. The THAA yields of bacteria were also 2-
to 9-fold higher than those of fungi.
The percentages of C as AA and AS in litter
increased with decomposition, approaching those of
soils and microorganisms, especially fungi (Fig. 2).
The percentage of N as AA generally fluctuated
Biogeochemistry (2014) 118:471–486 475
123
Ta
ble
1P
erce
nt
rem
ain
ing
(AF
DW
:as
h-f
ree
dry
wei
gh
t),
Can
dN
con
ten
ts,
mo
lar
Cto
Nra
tio
s,C
-no
rmal
ized
yie
lds
of
tota
lh
yd
roly
zab
leam
ino
acid
s(T
HA
A),
tota
l
hy
dro
lyza
ble
amin
o(T
HA
S),
glu
cosa
min
e(G
lcN
),an
dg
alac
tosa
min
e(G
alN
),G
lcN
/Gal
Nra
tio
,p
erce
nta
ges
of
Can
dN
asT
HA
Aan
dT
HA
S,
and
deg
rad
atio
nin
dex
(Dl)
of
dec
ayin
gli
tter
san
dso
ils
Sam
ple
%R
emai
nin
g
(AF
DW
)
C (wt%
)
N (wt%
)
C/N
(mo
le)
TH
AA
TH
AS
(nm
ol
[mg
C]-
1)
Glc
N
(nm
ol
[mg
C]-
1)
GaI
N
(nm
ol
[mg
C]-
1)
Glc
N/
GaI
N
%C
as
TH
AA
%N
as
TH
AA
%C
as
TH
AS
%N
as
TH
AS
Dl
Mis
canth
as
sinen
sis
gra
ssla
nd
Fre
shli
tter
10
04
2.0
0.8
35
8.9
64
71
7.8
12
.94
.33
.03
.90
64
.20
.13
1.2
61
.23
6m
on
ths
lite
r7
7.5
41
.00
.86
55
.86
34
32
.92
0.4
12
.51
.63
.78
59
.20
.24
2.2
01
.23
9m
on
ths
lite
r6
4.9
39
.31
.14
40
.08
11
47
.32
6.7
20
.61
.34
.78
64
.30
.34
2.2
81
.29
12
mo
nth
sli
tter
55
.03
8.5
1.2
83
5.0
11
81
57
.63
2.8
24
.81
.36
.99
69
.30
.41
2.4
20
.98
18
mo
nth
sli
tter
47
.03
9.4
1.4
03
2.8
11
49
55
.13
2.0
23
.01
.46
.76
62
.80
.40
2.1
71
.08
21
mo
nth
sli
tter
38
.73
7.5
1.5
22
8.8
12
43
75
.24
4.5
30
.61
.57
.36
60
.10
.54
2.6
01
.11
24
mo
nth
sli
tter
29
.73
6.5
1.5
22
8.0
11
52
81
.74
7.6
34
.11
.46
.76
53
.90
.59
2.7
41
.00
36
mo
nth
sli
tter
29
.23
5.0
1.7
22
3.7
11
59
97
.35
4.2
43
.11
.36
.82
46
.50
.70
2.7
70
.67
Org
anic
soil
36.6
1.4
225.8
1268
52.8
30.1
22.7
1.3
6.7
954.2
0.3
61.9
11.3
3
Min
eral
soil
15
.51
.00
14
.61
75
81
27
.97
3.6
52
.71
.48
.73
41
.40
.92
2.6
11
.40
Pin
us
den
sifl
ora
fore
st
Fre
shli
tter
10
05
1.2
0.4
01
48
.82
88
6.2
5.2
1.0
5.3
1.7
37
2.1
0.0
41
.11
1.0
5
6m
on
ths
litt
er3
7.5
51
.80
.55
10
9.3
40
59
.58
.21
.36
.42
.42
74
.50
.07
1.2
40
.80
9m
on
ths
lite
r7
6.7
51
.00
.74
80
.94
39
20
.91
8.3
2.6
6.9
2.6
25
9.2
0.1
52
.03
0.7
5
12
mo
nth
sli
tter
61
.85
0.1
0.9
85
9.5
71
52
9.9
24
.85
.14
.84
.23
71
.30
.22
2.1
40
.81
18
mo
nth
sli
tter
59
.75
1.1
0.9
86
1.0
86
63
4.0
28
.85
.25
.55
.15
88
.80
.25
2.4
90
.67
21
mo
nth
sli
tter
42
.75
1.0
1.2
14
9.2
60
24
1.4
31
.59
.63
.23
.59
49
.40
.30
2.4
40
.72
24
mo
nth
sli
tter
42
.75
0.6
1.2
84
6.2
86
84
8.1
35
.51
2.
62
.85
.15
67
.40
.35
2.6
60
.61
36
mo
nth
sli
tter
32
.65
1.8
1.4
94
0.0
95
85
5.9
39
.41
6.5
2.4
5.6
26
5.7
0.4
02
.72
0.6
1
Org
anic
soil
45.8
1.8
125.3
1358
57.8
36.0
21.8
1.7
7.3
257.4
0.4
22.0
51.6
5
Min
eral
soil
20
.31
.22
16
.61
51
88
6.7
47
.63
8.9
1.2
7.6
84
1.2
0.6
22
.02
1.2
2
Qu
ercu
scr
ispu
lafo
rest
Fre
shli
tter
10
04
7.5
0.8
76
3.7
68
11
2.0
11
.01
.01
1.1
4.1
17
1.8
0.0
90
.91
1.5
2
6m
on
ths
litt
er8
2.6
45
.41
.11
47
.96
30
18
.81
5.7
3.1
5.0
3.7
75
0.2
0.1
41
.08
1.2
6
9m
on
ths
lite
r7
3.4
45
.51
.36
39
.11
20
83
1.1
25
.16
.04
.27
.18
78
.80
.22
1.4
61
.07
12
mo
nth
sli
tter
62
.04
5.3
1.6
43
2.2
13
57
46
.43
7.1
9.3
4.0
8.0
77
3.2
0.3
31
.80
1.0
4
18
mo
nth
sli
tter
53
.04
4.5
1.6
33
1.8
10
99
45
.73
3.0
12
.72
.66
.53
58
.80
.33
1.7
51
.02
21
mo
nth
sli
tter
25
.34
5.7
1.5
63
4.1
13
21
42
.82
9.0
13
.82
.17
.87
75
.40
.31
1.7
51
.10
24
mo
nth
sli
tter
28
.44
5.3
1.7
72
9.8
15
17
50
.73
5.7
14
.92
.49
.00
75
.70
.36
1.8
10
.97
476 Biogeochemistry (2014) 118:471–486
123

between 50 and 70 % during litter decomposition,
while the %N as AS constantly increased by 1–2 %.
Nitrogen yields as AA plus AS were correlated with
total N content in decomposing litter from the three
plant species (R = 0.898, P \ 0.001) (Fig. 3). This
strong relationship indicated changes in total N
content of litter reflected those in AA and AS
throughout decomposition. In addition, the slope of
the line indicated AA and AS accounted for about
60 % of total N in the decomposing litter.
The % initial yields of THAA and THAS typically
increased during the early stages of decomposition and
decreased during the latter stages (Figs. 1C, D). The %
initial yield of THAS increased to a greater extent than
did THAA yield. Net increases in % initial yields were
observed for THAA of Pinus and THAS of Miscan-
thus and Pinus. Changing patterns in % initial yields of
THAA and THAS tracked those of % initial N,
especially for THAS.
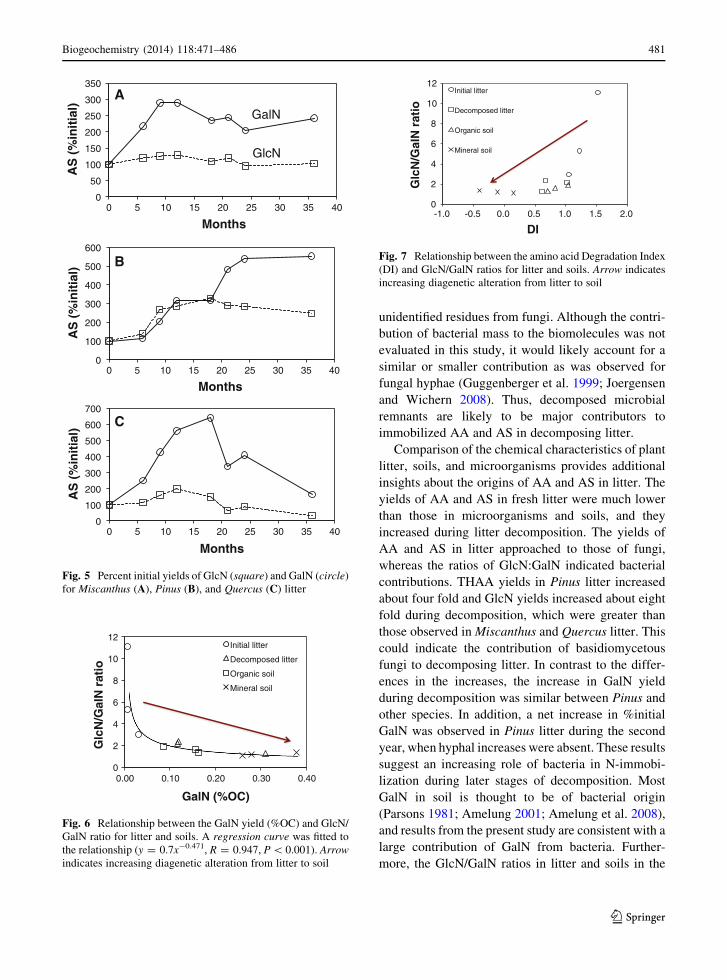
The yields of GlcN and GalN increased in all litter
samples during the experiment. Yields increased 3- to
8-fold for GlcN and 10- to 17-fold for GalN,
approaching those in soils (Table 1, Fig. 4). The
yields of GlcN and GalN in decomposing litter were
more similar to those of bacteria rather than fungi.
Glucosamine was the most abundant AS in fresh litter,
with 3- to 11-fold higher yields than GalN, whereas the
increases in yields were greater for GalN than GlcN
(Fig. 5). In the first year, the % initial of both GlcN and
GalN increased in litter for all plant species. Greater
increases in %initial GalN than GlcN were observed
for Miscanthus and Quercus litter in the first year,
while Pinus litter showed similar increases in %initial
GlcN and GalN. In contrast, in the second year, greater
increases in %initial GalN than GlcN was observed
for Pinus litter, but not for Miscanthus and Quercus
litter. The GlcN/GalN ratio of litter showed clear
patterns in all plant species, decreasing (3.0–11.1 to
1.3–2.4) to values similar to those in soils (Table 1).
The GlcN/GalN ratio decreased in Pinus and Quercus
litter throughout the study period, whereas the GlcN/
GalN ratio in Miscanthus litter decreased only during
the early stages of decomposition. Significant varia-
tion in the GlcN/GalN ratio was a function of the
C-normalized yield of GalN (Fig. 6; R = 0.636,
P \ 0.05). Soils exhibited only minor variations in
AS composition among sites. Although fungi and
bacteria both had higher yields of GlcN than GalN, the
yield of GalN was considerably high in some bacteriaTa
ble
1co
nti
nu
ed
Sam
ple
%R
emai
nin
g
(AF
DW
)
C (wt%
)
N (wt%
)
C/N
(mo
le)
TH
AA
TH
AS
(nm
ol
[mg
C]-
1)
Glc
N
(nm
ol
[mg
C]-
1)
GaI
N
(nm
ol
[mg
C]-
1)
Glc
N/
GaI
N
%C
as
TH
AA
%N
as
TH
AA
%C
as
TH
AS
%N
as
TH
AS
Dl
36
mo
nth
sli
tter
10
.54
5.5
1.7
53
0.4
14
32
51
.73
5.4
16
.32
.28
.50
72
.90
.37
1.8
91
.02
Org
anic
soil
46.0
1.4
032.8
1043
34.7
22.9
11.8
1.9
5.7
257.5
0.2
51.6
01.9
3
Min
eral
soil
23
.71
.35
17
.51
58
17
7.1
40
.93
6.2
1.1
8.1
34
5.4
0.5
61
.89
1.1
3
Av
erag
eo
fal
lsi
tes
Fre
shli
tter
46
.90
.70
90
.55
39
12
.09
.72
.16
.53
.25
69
.40
.09
1.0
91
.27
36
mo
nth
sli
tter
44
.11
.65
31
.61
18
36
8.3
43
.02
5.3
1.9
6.9
86
1.7
0.4
92
.46
0.7
7
Org
anic
soil
43.1
1.3
147.2
1055
40.9
25.3
15.5
3.2
5.7
960.3
0.2
91.6
81.4
2
Min
eral
soil
19
.81
.21
16
.21
61
99
7.2
54
.04
2.6
1.3
8.1
34
2.7
0.7
02
.17
1.2
5
Sta
nd
ard
dev
iati
on
of
all
site
Fre
shli
tter
4.6
0.2
65
0.6
21
85
.84
.01
.94
.21
.32
4.5
0.0
40
.17
0.2
4
36
mo
nth
sli
tter
8.5
0.1
48
.52
38
25
.29
.91
5.4
0.6
1.4
41
3.7
0.1
80
.50
0.2
2
Org
anic
soil
5.3
0.2
34.2
162
12.1
6.6
6.0
0.3
0.8
21.9
0.0
90.2
30.3
0
Min
eral
soil
4.1
0.1
41
.51
24
27
.01
7.2
8.9
0.1
0.5
32
.40
.19
0.3
80
.13
Biogeochemistry (2014) 118:471–486 477
123
(Table 2). Fungi exhibited much lower yields of GalN
than highly decomposed litter. The AA degradation
index decreased with decomposition (Table 1) and
showed a significant relationship with the GlcN/GalN
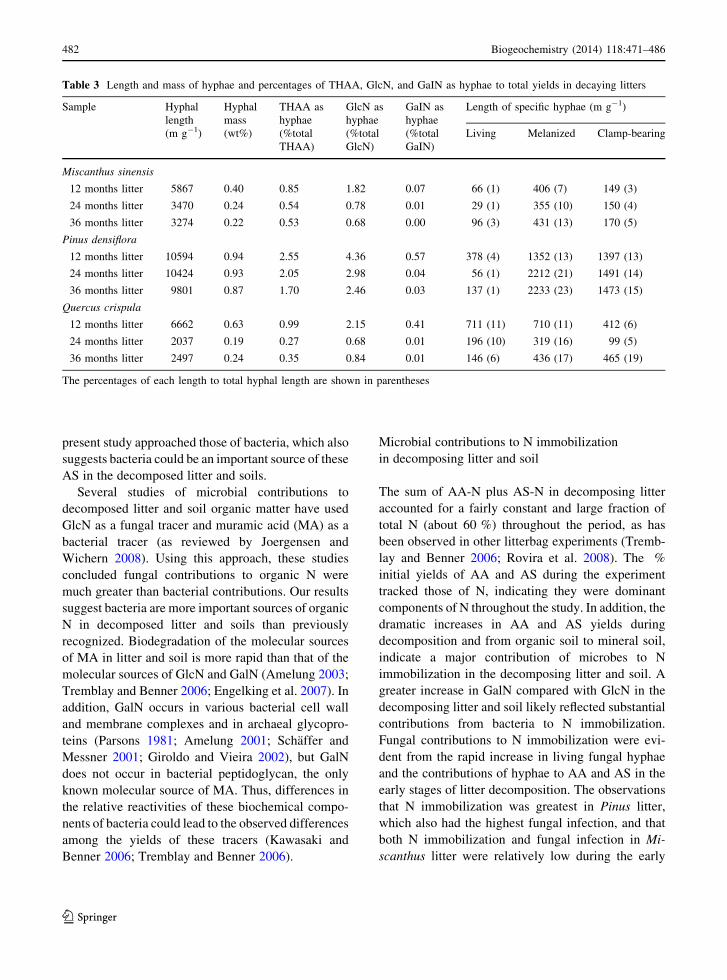
ratio (Fig. 7, R = 0.646, P \ 0.05).
The mol% of individual AA was relatively constant
during litter decomposition (Fig. 7, Supplementary
Fig. 1). The mol% Gly increased during decomposi-
tion by 1.6–2.2 % among the three plant types,
whereas the mol % Leu decreased by 0.8–1.3 %.
Organic soils mostly showed similar AA compositions
to those of well-decomposed litter. Mineral soils had
appreciably different AA compositions from litter and
organic soils, especially in mol% Asp, Leu, and Gly.
Microorganisms exhibited higher mol% Glu and Ala
and lower mol% Pro and Ser than litter and organic
and mineral soils.
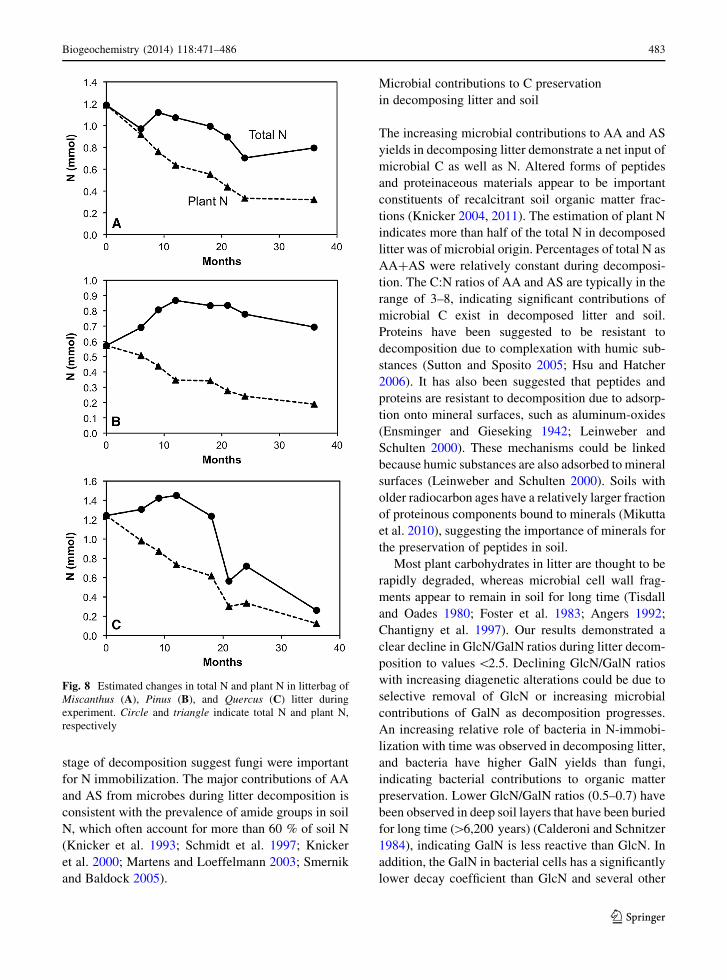
Length and mass of hyphae
Observations of hyphae revealed that average hyphal
length varied significantly among the plant litter types
(Table 3). Pinus litter had the highest hyphal lengths
among species and no substantial changes in hyphal
length throughout the experiment. Miscanthus and
Quercus litter had 37–80 % shorter hyphal lengths
than those in Pinus litter, and decreased (40–70 %) in
hyphal length from 12 to 24 months. Hyphal mass
(wt%) was greatest in Pinus litter and accounted for
\1 % of total weight. Temporal patterns of hyphal
mass were similar to those of hyphal length.
Length of specific types of melanized hyphae in
litter was relatively long (319–2233 m g-1) compared
with those of living (29–711 m g-1) and clamp-
bearing hyphae (99–1491 m g-1). Living hyphal
lengths decreased, especially from 12 to 24 months,
in all litter samples. In contrast, lengths of melanized
and clamp-bearing hyphae increased with time, espe-
cially for Miscanthus and Pinus litter. Lengths of
melanized and clamp-bearing hyphae were the longest
for Pinus litter, whereas Quercus litter had the longest
lengths of living hyphae throughout the experiment.
Specific hyphal types accounted for a minor portion
of total hyphae. Living hyphae accounted for 1–11 %,
melanized hyphae accounted for 7–23 %, and clamp-
bearing hyphae accounted for 3–19 % of total hyphal.
0
20
40
60
80
100
120
140
160
0 5 10 15 20 25 30 35 40
N(%
init
ial)
Months
0 20 40 60 80
100 120 140 160 180 200
0 5 10 15 20 25 30 35 40
TH
AA
(%in
itia
l)
Months
0
50
100
150
200
250
300
350
0 5 10 15 20 25 30 35 40
TH
AS
(%in
itia
l)
Months
A B
C D
Fig. 1 Yields of C (A), N (B), total hydrolyzable amino acids
(THAA) (C), and total hydrolyzable amino sugars (THAS)
(D) through litterbag experiment, given as % of initial contents.
Circle, triangle, and square indicate Miscanthus, Pinus, and
Quercus litter, respectively
478 Biogeochemistry (2014) 118:471–486
123
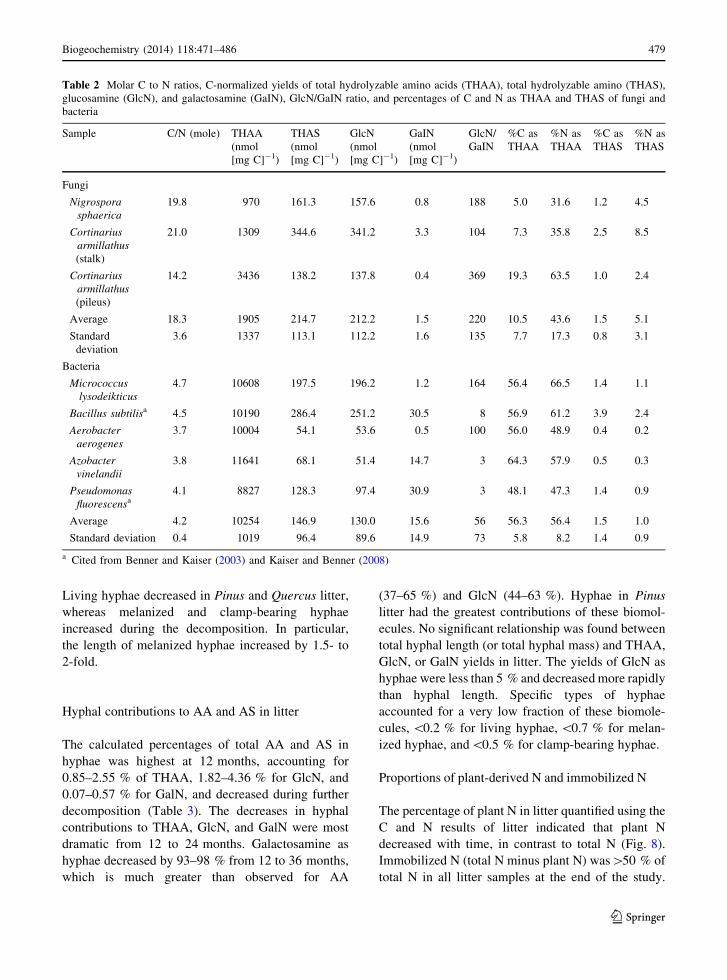
Living hyphae decreased in Pinus and Quercus litter,
whereas melanized and clamp-bearing hyphae
increased during the decomposition. In particular,
the length of melanized hyphae increased by 1.5- to
2-fold.
Hyphal contributions to AA and AS in litter
The calculated percentages of total AA and AS in
hyphae was highest at 12 months, accounting for
0.85–2.55 % of THAA, 1.82–4.36 % for GlcN, and
0.07–0.57 % for GalN, and decreased during further
decomposition (Table 3). The decreases in hyphal
contributions to THAA, GlcN, and GalN were most
dramatic from 12 to 24 months. Galactosamine as
hyphae decreased by 93–98 % from 12 to 36 months,
which is much greater than observed for AA
(37–65 %) and GlcN (44–63 %). Hyphae in Pinus
litter had the greatest contributions of these biomol-
ecules. No significant relationship was found between
total hyphal length (or total hyphal mass) and THAA,
GlcN, or GalN yields in litter. The yields of GlcN as
hyphae were less than 5 % and decreased more rapidly
than hyphal length. Specific types of hyphae
accounted for a very low fraction of these biomole-
cules, \0.2 % for living hyphae, \0.7 % for melan-
ized hyphae, and\0.5 % for clamp-bearing hyphae.
Proportions of plant-derived N and immobilized N
The percentage of plant N in litter quantified using the
C and N results of litter indicated that plant N
decreased with time, in contrast to total N (Fig. 8).
Immobilized N (total N minus plant N) was[50 % of
total N in all litter samples at the end of the study.
Table 2 Molar C to N ratios, C-normalized yields of total hydrolyzable amino acids (THAA), total hydrolyzable amino (THAS),
glucosamine (GlcN), and galactosamine (GaIN), GlcN/GaIN ratio, and percentages of C and N as THAA and THAS of fungi and
bacteria
Sample C/N (mole) THAA
(nmol
[mg C]-1)
THAS
(nmol
[mg C]-1)
GlcN
(nmol
[mg C]-1)
GaIN
(nmol
[mg C]-1)
GlcN/
GaIN
%C as
THAA
%N as
THAA
%C as
THAS
%N as
THAS
Fungi
Nigrospora
sphaerica
19.8 970 161.3 157.6 0.8 188 5.0 31.6 1.2 4.5
Cortinarius
armillathus
(stalk)
21.0 1309 344.6 341.2 3.3 104 7.3 35.8 2.5 8.5
Cortinarius
armillathus
(pileus)
14.2 3436 138.2 137.8 0.4 369 19.3 63.5 1.0 2.4
Average 18.3 1905 214.7 212.2 1.5 220 10.5 43.6 1.5 5.1
Standard
deviation
3.6 1337 113.1 112.2 1.6 135 7.7 17.3 0.8 3.1
Bacteria
Micrococcus
lysodeikticus
4.7 10608 197.5 196.2 1.2 164 56.4 66.5 1.4 1.1
Bacillus subtilisa 4.5 10190 286.4 251.2 30.5 8 56.9 61.2 3.9 2.4
Aerobacter
aerogenes
3.7 10004 54.1 53.6 0.5 100 56.0 48.9 0.4 0.2
Azobacter
vinelandii
3.8 11641 68.1 51.4 14.7 3 64.3 57.9 0.5 0.3
Pseudomonas
fluorescensa4.1 8827 128.3 97.4 30.9 3 48.1 47.3 1.4 0.9
Average 4.2 10254 146.9 130.0 15.6 56 56.3 56.4 1.5 1.0
Standard deviation 0.4 1019 96.4 89.6 14.9 73 5.8 8.2 1.4 0.9
a Cited from Benner and Kaiser (2003) and Kaiser and Benner (2008)
Biogeochemistry (2014) 118:471–486 479
123
Pinus litter exhibited the highest percentage (73 %) of
immobilized N.
Discussion
Influences of microorganisms on litter
decomposition and soil formation
Microorganisms had a major impact on the chemical
composition of decomposing litter. The net increases
of N in decomposing litter were in sharp contrast to the
net losses of C. Likewise, net increases of AA and AS
were also observed, and these biochemicals comprised
about 60 % of total N. The yields of AA and AS in
decomposed litter approached those found in micro-
organisms. The N immobilized during litter decom-
position was estimated to be greater than the
remaining plant N at the end of the study. These
observations indicated C-rich plant litter was gradu-
ally transformed to N-rich litter by microorganisms.
Microbes contributed newly synthesized ‘‘immobi-
lized’’ forms of N, such as AA and AS, to decompos-
ing litter in addition to degrading biomolecules of
plant origin (Tremblay and Benner 2006; Miltner et al.
2011). Furthermore, the observed transformation of
AA and AS yields from plant litter to soils, indicates
microorganisms play an important role in soil
formation.
The data on fungal hyphae in decomposing plant
litter enable us to further explore microbial contribu-
tions to AA and AS. Hyphae accounted for \5 % of
THAA, GlcN, and GalN in the decomposing litter, and
living hyphae accounted for \0.2 %, suggesting
identifiable hyphae contributed a minor fraction of
these N-containing biomolecules. It appears most of
the immobilized AA and AS were derived primarily
from bacteria (biomass and necromass) and physically
Fig. 2 Relationship between THAA and THAS contents
(%OC) for litter, soils, and microorganisms. Bars are standard
deviations (n = 3–5). Arrow indicates increasing diagenetic
alteration from litter to soil
Total N (µmol / gC)
AA
+ A
S N
(µ
mo
l / g
C)
Fig. 3 Total nitrogen yield versus sum of nitrogen yields from
total hydrolyzable amino acids (AA) plus total hydrolyzable
amino sugars (AS) of decomposing litter. Circle, triangle, and
square indicate Miscanthus, Pinus, and Quercus, respectively.
A linear regression of the data is shown (y = 0.60x ? 0.14,
R = 0.898, P \ 0.001)
Fig. 4 Relationship between GlcN and GalN contents (%OC)
for litter, soils, and microorganisms. Bars are standard
deviations (n = 3–5). Arrow indicates increasing diagenetic
alteration from litter to soil
480 Biogeochemistry (2014) 118:471–486
123
unidentified residues from fungi. Although the contri-
bution of bacterial mass to the biomolecules was not
evaluated in this study, it would likely account for a
similar or smaller contribution as was observed for
fungal hyphae (Guggenberger et al. 1999; Joergensen
and Wichern 2008). Thus, decomposed microbial
remnants are likely to be major contributors to
immobilized AA and AS in decomposing litter.
Comparison of the chemical characteristics of plant
litter, soils, and microorganisms provides additional
insights about the origins of AA and AS in litter. The
yields of AA and AS in fresh litter were much lower
than those in microorganisms and soils, and they
increased during litter decomposition. The yields of
AA and AS in litter approached to those of fungi,
whereas the ratios of GlcN:GalN indicated bacterial
contributions. THAA yields in Pinus litter increased
about four fold and GlcN yields increased about eight
fold during decomposition, which were greater than
those observed in Miscanthus and Quercus litter. This
could indicate the contribution of basidiomycetous
fungi to decomposing litter. In contrast to the differ-
ences in the increases, the increase in GalN yield
during decomposition was similar between Pinus and
other species. In addition, a net increase in %initial
GalN was observed in Pinus litter during the second
year, when hyphal increases were absent. These results
suggest an increasing role of bacteria in N-immobi-
lization during later stages of decomposition. Most
GalN in soil is thought to be of bacterial origin
(Parsons 1981; Amelung 2001; Amelung et al. 2008),
and results from the present study are consistent with a
large contribution of GalN from bacteria. Further-
more, the GlcN/GalN ratios in litter and soils in the
0
100
200
300
400
500
600
700
0 5 10 15 20 25 30 35 40
AS
(%in
itia
l)
0
50
100
150
200
250
300
350
0 5 10 15 20 25 30 35 40
AS
(%in
itia
l)
Months
0
100
200
300
400
500
600
0 5 10 15 20 25 30 35 40
AS
(%in
itia
l)
A
B
C
GalN
GlcN
Months
Months
Fig. 5 Percent initial yields of GlcN (square) and GalN (circle)
for Miscanthus (A), Pinus (B), and Quercus (C) litter
0
2
4
6
8
10
12
0.00 0.10 0.20 0.30 0.40
Glc
N/G
a lN
rati
o
GalN (%OC)
Initial litter
Decomposed litter
Organic soil
Mineral soil
Fig. 6 Relationship between the GalN yield (%OC) and GlcN/
GalN ratio for litter and soils. A regression curve was fitted to
the relationship (y = 0.7x-0.471, R = 0.947, P \ 0.001). Arrow
indicates increasing diagenetic alteration from litter to soil
0
2
4
6
8
10
12
-1.0 -0.5 0.0 0.5 1.0 1.5 2.0
Glc
N/G
alN
r ati
o
DI
Initial litter
Decomposed litter
Organic soil
Mineral soil
Fig. 7 Relationship between the amino acid Degradation Index
(DI) and GlcN/GalN ratios for litter and soils. Arrow indicates
increasing diagenetic alteration from litter to soil
Biogeochemistry (2014) 118:471–486 481
123
present study approached those of bacteria, which also
suggests bacteria could be an important source of these
AS in the decomposed litter and soils.
Several studies of microbial contributions to
decomposed litter and soil organic matter have used
GlcN as a fungal tracer and muramic acid (MA) as a
bacterial tracer (as reviewed by Joergensen and
Wichern 2008). Using this approach, these studies
concluded fungal contributions to organic N were
much greater than bacterial contributions. Our results
suggest bacteria are more important sources of organic
N in decomposed litter and soils than previously
recognized. Biodegradation of the molecular sources
of MA in litter and soil is more rapid than that of the
molecular sources of GlcN and GalN (Amelung 2003;
Tremblay and Benner 2006; Engelking et al. 2007). In
addition, GalN occurs in various bacterial cell wall
and membrane complexes and in archaeal glycopro-
teins (Parsons 1981; Amelung 2001; Schaffer and
Messner 2001; Giroldo and Vieira 2002), but GalN
does not occur in bacterial peptidoglycan, the only
known molecular source of MA. Thus, differences in
the relative reactivities of these biochemical compo-
nents of bacteria could lead to the observed differences
among the yields of these tracers (Kawasaki and
Benner 2006; Tremblay and Benner 2006).
Microbial contributions to N immobilization
in decomposing litter and soil
The sum of AA-N plus AS-N in decomposing litter
accounted for a fairly constant and large fraction of
total N (about 60 %) throughout the period, as has
been observed in other litterbag experiments (Tremb-
lay and Benner 2006; Rovira et al. 2008). The %
initial yields of AA and AS during the experiment
tracked those of N, indicating they were dominant
components of N throughout the study. In addition, the
dramatic increases in AA and AS yields during
decomposition and from organic soil to mineral soil,
indicate a major contribution of microbes to N
immobilization in the decomposing litter and soil. A
greater increase in GalN compared with GlcN in the
decomposing litter and soil likely reflected substantial
contributions from bacteria to N immobilization.
Fungal contributions to N immobilization were evi-
dent from the rapid increase in living fungal hyphae
and the contributions of hyphae to AA and AS in the
early stages of litter decomposition. The observations
that N immobilization was greatest in Pinus litter,
which also had the highest fungal infection, and that
both N immobilization and fungal infection in Mi-
scanthus litter were relatively low during the early
Table 3 Length and mass of hyphae and percentages of THAA, GlcN, and GaIN as hyphae to total yields in decaying litters
Sample Hyphal
length
Hyphal
mass
THAA as
hyphae
GlcN as
hyphae
GaIN as
hyphae
Length of specific hyphae (m g-1)
(m g-1) (wt%) (%total
THAA)
(%total
GlcN)
(%total
GaIN)
Living Melanized Clamp-bearing
Miscanthus sinensis
12 months litter 5867 0.40 0.85 1.82 0.07 66 (1) 406 (7) 149 (3)
24 months litter 3470 0.24 0.54 0.78 0.01 29 (1) 355 (10) 150 (4)
36 months litter 3274 0.22 0.53 0.68 0.00 96 (3) 431 (13) 170 (5)
Pinus densiflora
12 months litter 10594 0.94 2.55 4.36 0.57 378 (4) 1352 (13) 1397 (13)
24 months litter 10424 0.93 2.05 2.98 0.04 56 (1) 2212 (21) 1491 (14)
36 months litter 9801 0.87 1.70 2.46 0.03 137 (1) 2233 (23) 1473 (15)
Quercus crispula
12 months litter 6662 0.63 0.99 2.15 0.41 711 (11) 710 (11) 412 (6)
24 months litter 2037 0.19 0.27 0.68 0.01 196 (10) 319 (16) 99 (5)
36 months litter 2497 0.24 0.35 0.84 0.01 146 (6) 436 (17) 465 (19)
The percentages of each length to total hyphal length are shown in parentheses
482 Biogeochemistry (2014) 118:471–486
123
stage of decomposition suggest fungi were important
for N immobilization. The major contributions of AA
and AS from microbes during litter decomposition is
consistent with the prevalence of amide groups in soil
N, which often account for more than 60 % of soil N
(Knicker et al. 1993; Schmidt et al. 1997; Knicker
et al. 2000; Martens and Loeffelmann 2003; Smernik
and Baldock 2005).
Microbial contributions to C preservation
in decomposing litter and soil
The increasing microbial contributions to AA and AS
yields in decomposing litter demonstrate a net input of
microbial C as well as N. Altered forms of peptides
and proteinaceous materials appear to be important
constituents of recalcitrant soil organic matter frac-
tions (Knicker 2004, 2011). The estimation of plant N
indicates more than half of the total N in decomposed
litter was of microbial origin. Percentages of total N as
AA?AS were relatively constant during decomposi-
tion. The C:N ratios of AA and AS are typically in the
range of 3–8, indicating significant contributions of
microbial C exist in decomposed litter and soil.
Proteins have been suggested to be resistant to
decomposition due to complexation with humic sub-
stances (Sutton and Sposito 2005; Hsu and Hatcher
2006). It has also been suggested that peptides and
proteins are resistant to decomposition due to adsorp-
tion onto mineral surfaces, such as aluminum-oxides
(Ensminger and Gieseking 1942; Leinweber and
Schulten 2000). These mechanisms could be linked
because humic substances are also adsorbed to mineral
surfaces (Leinweber and Schulten 2000). Soils with
older radiocarbon ages have a relatively larger fraction
of proteinous components bound to minerals (Mikutta
et al. 2010), suggesting the importance of minerals for
the preservation of peptides in soil.
Most plant carbohydrates in litter are thought to be
rapidly degraded, whereas microbial cell wall frag-
ments appear to remain in soil for long time (Tisdall
and Oades 1980; Foster et al. 1983; Angers 1992;
Chantigny et al. 1997). Our results demonstrated a
clear decline in GlcN/GalN ratios during litter decom-
position to values \2.5. Declining GlcN/GalN ratios
with increasing diagenetic alterations could be due to
selective removal of GlcN or increasing microbial
contributions of GalN as decomposition progresses.
An increasing relative role of bacteria in N-immobi-
lization with time was observed in decomposing litter,
and bacteria have higher GalN yields than fungi,
indicating bacterial contributions to organic matter
preservation. Lower GlcN/GalN ratios (0.5–0.7) have
been observed in deep soil layers that have been buried
for long time ([6,200 years) (Calderoni and Schnitzer
1984), indicating GalN is less reactive than GlcN. In
addition, the GalN in bacterial cells has a significantly
lower decay coefficient than GlcN and several other
Fig. 8 Estimated changes in total N and plant N in litterbag of
Miscanthus (A), Pinus (B), and Quercus (C) litter during
experiment. Circle and triangle indicate total N and plant N,
respectively
Biogeochemistry (2014) 118:471–486 483
123

bacterial tracers, including D-amino acids and MA,
which are found in various cell wall components
(Kawasaki and Benner 2006).
Relatively high GlcN/GalN ratios in organisms and
its gradual decrease to values \2 during decomposi-
tion is consistently observed in litterbag studies
conducted in soil (Amelung et al. 2001) and water
(Tremblay and Benner 2006). In addition, highly
decomposed marine organic matter typically exhibits
low GlcN/GalN ratios (Ogawa et al. 2001; Benner and
Kaiser 2003; Kaiser and Benner 2009). The differ-
ences in reactivity between GlcN and GalN in
decomposing bacteria and the decreasing GlcN/GalN
ratio with increasing decomposition indicates GlcN/
GalN ratios are useful indicators of the extent of
diagenetic alteration of natural organic matter in a
wide ranges of systems. Likewise, yields of common
biochemicals, such as AA, are also useful indicators of
the diagenetic state of organic matter (Cowie and
Hedges 1994).
Although GlcN appears to be less resistant to
decomposition than GalN, the substantial (3- to 8-fold)
increases in GlcN yields indicate it occurs in resistant
forms of organic matter. Chitin, a polymer of acety-
lated GlcN subunits, is rapidly degraded when added
to soil, whereas GlcN yields in soil are relatively high,
indicating that GlcN in soil could be in molecular
complexes that are stabilized against microbial attack
(Bondietti et al. 1972; Parsons 1981). The higher
percentage of melanized and clamp-bearing hyphae to
total hyphae in the latter stages of the decomposition,
suggests the melanization process increases the resis-
tance of basidiomycetous hyphae to decomposition.
Melanins are assumed to be generated by the conden-
sation of phenolic precursors present in fungal cell
walls and to increase the resistance of hyphae to
decompose (Linhares and Martin 1978; Martin and
Haider 1986; Chenu and Cosentino 2011).
Organic matter derived from microorganisms plays
a critical role in the formation and preservation of soil
organic matter (Liang et al. 2011). The molecular
signatures of heterotrophic microbes are evident in
decomposing litter and soils, particularly in amino
acids and amino sugars. The yields of these microbial
biomolecules increase rapidly during litter decompo-
sition and appear to be important for shaping the
chemical composition and long-term stabilization of
soil organic matter.
Acknowledgments We thank Karl Kaiser and Mike Philben
for analyzing C, N, and AA in several microorganisms and for
assistance with AA and AS analyses. We also thank Yoko
Morimoto, Ruth Flerus, Tomohiro Kasuga, Seiya Shiratori,
Marin Otomichi, Hiroki Inoue, Hidetomo Iwano, Isao Kato,
Hiroshi Yokota, and Teruo Matsunaka for support in the
laboratory. This research was supported by grant from the
Japanese Society for the Promotion of Science (No. 21710014)
and by Grants-in-Aid to Cooperative Research from Rakuno
Gakuen University (2008). Ronald Benner acknowledges
support from NSF grant 0843417.
References
Aber JD, Melillo JM (2001) Terrestrial ecosystems, 2nd edn.
Academic Press, New York
Amelung W (2001) Methods using amino sugars as markers for
microbial residues in soil. In: Lal R, Kimble JM, Follett RF,
Stewart BA (eds) Assessment methods for soil carbon
pools. Advances in soil science. CRC/Lewis Publishers,
Boca Raton, pp 233–270
Amelung W (2003) Nitrogen biomarkers and their fate in soil.
J Plant Nutr Soil Sci 166:677–686
Amelung W, Miltner A, Zhang X, Zech W (2001) Fate of
microbial residues during litter decomposition as affected
by minerals. Soil Sci 166:598–606
Amelung W, Brodowski S, Sandhage-Hofmann A, Bol R (2008)
Combining biomarker with stable isotope analyses for
assessing the transformation and turnover of soil organic
matter. Adv Agron 100:155–250
Angers DA (1992) Changes in soil aggregation and organic
carbon under corn and alfalfa. Soil Sci Soc Am J
56:1244–1249
Baath E, Soderstrom B (1977) Mycelial lengths and fungal
biomasses in some Swedish coniferous forest soils, with
special reference to a pine forest in central Sweden.
Technical Report. Swedish Coniferous Forest Project,
Uppsala, pp 1–45
Bengtsson G, Bengtson P, Mansson KF (2003) Gross nitrogen
mineralization-, immobilization-, and nitrification rates as
a function of soil C/N ratio and microbial activity. Soil Biol
Biochem 35:143–154
Benner R, Kaiser K (2003) Abundance of amino sugars and
peptidoglycan in marine particulate and dissolved organic
matter. Limnol Oceanogr 48:118–128
Berg B, McClaugherty C (2008) Plant litter, decomposition,
humus formation, carbon sequestration, 2nd edn. Springer,
Berlin
Bondietti E, Martin JP, Haider K (1972) Stabilization of amino
sugar units in humic type polymers. Soil Sci Soc Am J
36:597–602
Calderoni G, Schnitzer M (1984) Nitrogen distribution as a
function of raiodcarbon age in Paleosol humic acids. Org
Geochem 5:203–209
Chantigny MH, Angers DA, Prevost D, Vezina L-P, Cahlifour
F-P (1997) Soil aggregation and fungal and bacterial bio-
mass under annual and perennial cropping systems. Soil
Sci Soc Am J 61:262–267
484 Biogeochemistry (2014) 118:471–486
123

Chenu C, Cosentino D (2011) Microbial regulation of soil
structural dynamics. In: Ritz K, Young I (eds) The archi-
tecture and biology and soils: life in inner space. CAB
International, London, pp 37–70
Cotrufo MF, Wallenstein MD, Boot CM, Denef K, Paul E
(2013) The Microbial Efficiency-Matrix Stabilization
(MEMS) framework integrates plant litter decomposition
with soil organic matter stabilization: Do labile plant inputs
form stable soil organic matter? Global Change Biol.
doi:10.1111/gcb.12113
Cowie GL, Hedges JI (1994) Biochemical indicators of diage-
netic alteration in natural organic-matter mixtures. Nature
369:304–307
Dauwe B, Middelburg JJ, Herman PMJ, Heip CHR (1999)
Linking diagenetic alteration of amino acids and bulk
organic matter reactivity. Limnol Oceanogr 44:1809–1814
Ding X, Zhang X, He H, Xie H (2010) Dynamics of soil amino
sugar pools during decomposition processes of corn resi-
dues as affected by inorganic N addition. J Soil Sediment
10:758–766
Engelking B, Flessa H, Joergensen RG (2007) Shifts in amino
sugar and ergosterol contents after addition of sucrose and
cellulose to soil. Soil Biol Biochem 39:2111–2118
Ensminger L, Gieseking JE (1942) Resistance of clay-adsorbed
proteins to proteolytic hydrolysis. Soil Sci 53:205–209
Fontaine S, Henault C, Aamor A, Bdioui N, Bloor JMG, Maire
V, Mary B, Revaillot S, Maron PA (2011) Fungi mediate
long term sequestration of carbon and nitrogen in soil
through their priming effect. Soil Biol Biochem 43:86–96
Foster RC, Rovira AD, Cock TW (1983) Ultrastructure of the
root-soil interface. American Phytopathological Society,
St. Paul
Giroldo D, Vieira AAH (2002) An extracellular sulfated fucose-
rich polysaccharide produced by a tropical strain of
Cryptomonas obovata (Cryptophyceae). J Appl Phycol
14:185–191
Glaser B, Millar N, Blum H (2006) Sequestration and turnover
of bacterial- and fungal- derived carbon in a temperate
grassland soil under long-term elevated atmospheric
pCO2. Global Change Biol 12:1521–1531
Guggenberger G, Frey SD, Six J, Paustian K, Elliott ET (1999)
Bacterial and fungal cell wall residues in conventional and
no-tillage agroecosystems. Soil Sci Soc Am J 63:1188–
1198
Hart SC, Nason GE, Myrold DD (1994) Dynamics of gross
nitrogen transformations in an old-growth forest: the car-
bon connection. Ecology 75:880–891
He HB, Lia XB, Zhang W, Zhang XD (2011a) Differentiating
the dynamics of native and newly immobilized amino
sugars in soil frequently amended with inorganic nitrogen
and glucose. Eur J Soil Sci 62:144–151
He H, Zhang W, Zhang X, Xie H, Zhuang J (2011b) Temporal
responses of soil microorganisms to substrate addition as
indicated by amino sugar differentiation. Soil Biol Bio-
chem 43:1155–1161
Hsu P-H, Hatcher PG (2005) New evidence for covalent cou-
pling of peptides to humic acids based on 2D NMR spec-
troscopy: a means for preservation. Geochim Cosmochim
Acta 69:4521–4533
Iimura Y, Fujimoto M, Hirota M, Tamura K, Higashi T,
Yonebayashi K, Fujitake N (2010) Effects of ecological
succession on surface mineral horizons in Japanese vol-
canic ash soil. Geoderma 159:122–130
Joergensen RG, Wichern F (2008) Quantitative assessment of
the fungal contribution to microbial tissue in soil. Soil Biol
Biochem 40:2977–2991
Kaiser K, Benner R (2000) Determination of amino sugars in
environmental samples with high salt content by high-
performance anion-exchange chromatography and pulsed
amperometric detection. Anal Chem 72:2566–2572
Kaiser K, Benner R (2008) Major bacterial contribution to the
ocean reservoir of detrital organic carbon and nitrogen.
Limnol Oceanogr 53:99–112
Kaiser K, Benner R (2009) Biochemical composition and size
distribution of organic matter at the Pacific and Atlantic
time-series stations. Mar Chem 113:63–77
Kato J, Hayasi I (2006) Quantitative analysis of a stand of Pinus
densiflora undergoing succession to Quercus mongolica
ssp. crispula: 1. A 31-year record of growth and population
dynamics of the canopy trees. Ecol Res 21:503–509
Kawasaki N, Benner R (2006) Bacterial release of dissolved
organic matter during cell growth and decline: molecular
origin and composition. Limnol Oceanogr 51:2170–2180
Knicker H (2004) Stabilization of N-compounds in soil and
organic matter rich sediments - What is the difference?
Marine Chem 92:167–195
Knicker H (2011) Soil organic N—An under-rated player for C
sequestration in soils? Soil Biol Biochem 43:1118–1129
Knicker H, Frund R, Ludemann H-D (1993) The chemical
nature of nitrogen in soil organic matter. Naturwissens-
chaften 80:219–221
Knicker H, Schmidt MWI, Kogel-Knabner I (2000) Nature of
organic nitrogen in fine particle size separates of sandy
soils of highly industrialized areas as revealed by NMR
spectroscopy. Soil Biol Biochem 32:241–252
Leinweber P, Schulten HR (2000) Nonhydrolyzable forms of
soil organic nitrogen: extractability and composition.
J Plant Nutr Soil Sci 163:433–439
Liang C, Balser TC (2011) Microbial production of recalcitrant
organic matter in global soils: implications for productivity
and climate policy. Nature Rev Microbiol 9:75
Liang C, Zhang X, Balser TC (2007) Net microbial amino
sugars accumulation process in soil as influenced by dif-
ferent plant material inputs. Biol Fert Soils 44:1–7
Liang C, Cheng G, Wixon DL, Balser TC (2011) An absorbing
Markov chain approach to understanding the microbial role
in soil carbon stabilization. Biogeochemistry 106:303–309
Linhares LF, Martin JP (1978) Decomposition in soil of the
humic acid-type polymers (melanins) of Eurotium echi-
nulatum, Aspergillus glaucus sp. and other fungi. Soil Sci
Soc Am J 42:738–743
Martens DA, Loeffelmann KL (2003) Soil amino acid composi-
tion quantified by acid hydrolysis and anion chromatogra-
phy-pulsed amperometry. J Agr Food Chem 51:6521–6529
Martin JP, Haider K (1986) Influence of mineral colloids on
turnover rates of soil organic carbon. In: Huang PM,
Shnitzer M (eds) Interactions of soil minerals with natural
organics and microbes, vol 17. SSSA Special Publication,
Madison, pp 283–304
Marumoto T, Anderson JPE, Domsch KH (1982) Decomposi-
tion of 14C- and 15N-labelled microbial cells in soil. Soil
Biol Biochem 14:461–467
Biogeochemistry (2014) 118:471–486 485
123

Mengel K (1996) Turnover of organic nitrogen in soils and its
availability to crops. Plant Soil 181:83–93
Mikutta R, Kaiser K, Dorr N, Vollmer A, Chadwick OA,
Chorover J, Kramer MG, Guggenberger G (2010) Miner-
alogical impact on organic nitrogen across a long-term soil
chronosequence (0.3–4100 kyr). Geochim Cosmochim
Acta 74:2142–2164
Miltner A, Bombach P, Schmidt-Brucken B, Kastner M (2011)
SOM genesis: microbial biomass as a significant source.
Biogeochemistry. doi:10.1007/s10533-011-9658-z
Ogawa H, Amagai Y, Koike I, Kaiser K, Benner R (2001)
Production of refractory dissolved organic matter by bac-
teria. Science 292:917–920
Ono Y (1998) A study on the initial decomposition process of
needle litter in a Chamaecyparis obtusa forest. Masters
Thesis, Kyoto University, Kyoto, Japan (in Japanese)
Osono T (2010) Decomposition of grass leaves by ligninolytic
litter-decomposing fungi. Grassland Sci 56:31–36
Osono T, Takeda H (2001) Organic chemical and nutrient
dynamics in decomposing beech leaf litter in relation to
fungal ingrowth and succession during 3-year decompo-
sition processes in a cool temperate deciduous forest in
Japan. Ecol Res 16:649–670
Parsons JW (1981) Chemistry and distribution of amino sugars
in soils and soil organisms. In: Paul EA, Ladd JN (eds)
Soil Biochemistry, vol 5. Marcel Dekker, New York,
pp 197–227
Paul EA, Clark FE (1989) Soil microbiology and biochemistry.
Academic Press, New York
Reverter M, Lundh T, Lindberg JE (1997) Determination of free
amino acids in pig plasma by precolumn derivatization
with 6-N- aminoquinolyl-N-hydroxysuccinimidyl carba-
mate and high performance liquid chromatography.
J Chromatogr B Biomed Sci Appl. 696((1):1–8
Rovira P, Kurz-Besson C, Hernandez P, Couteaux M–M, Val-
lejo VR (2008) Searching for an indicator of N evolution
during organic matter decomposition based on amino acids
behaviour: a study on litter layers of pine forests. Plant Soil
307:149–166
Saito T (1955) The significance of plate counts of soil fungi and
the detection of their mycelia. Ecol Rev 14:69–74
Schaffer C, Messner P (2001) Glycobiology of surface layer
proteins. Biochimie 83:591–599
Schmidt MWI, Knicker H, Hatcher PG, Kogel-Knabner I (1997)
Improvement of 13C and 15N CPMAS NMR spectra of bulk
soils, particle size fractions and organic material by treat-
ment with 10% hydrofluoric acid. Eur J Soil Sci 48:319–328
Simpson AJ, Simpson MJ, Smith E, Kelleher BP (2007) Micro-
bially derived inputs to soil organic matter: are current
estimates too low? Environ Sci and Techonol 41:8070–8076
Six J, Frey SD, Thiet RK, Batten KM (2006) Bacterial and
fungal contributions to carbon sequestration in agroeco-
systems. Soil Sci Soc Am J 70:555–569
Smernik RJ, Baldock JA (2005) Does solid-state 15N NMR
spectroscopy detect all soil organic nitrogen? Biogeo-
chemistry 75:507–528
Solomon D, Fritzsche F, Tekalign M, Lehmann J, Zech W
(2002) Soil organic matter composition in the Subhumid
Ethiopian Highlands as influenced by deforestation and
agricultural management. Soil Sci Soc Am J 66:68–82
Sutton R, Sposito G (2005) Molecular structure in soil humic
substances: the new view. Environ Sci Technol 39:
9009–9015
Swift MJ, Heal OW, Anderson JM (1979) Decomposition in
terrestrial ecosystems. Studies in ecology, vol 5. University
of California Press, Berkeley
Tisdall JM, Oades JM (1980) The management of ryegrass to
stabilize aggregates of a red-brown earth. Aust J Soil Res
18:415–422
Tremblay L, Benner R (2006) Microbial contributions to
N-immobilization and organic matter preservation in
decomposing plant detritus. Geochim Cosmochim Acta
70:133–146
Wanek W, Mooshammer M, Blochl A, Hanreich A, Richter A
(2010) Determination of gross rates of amino acid pro-
duction and immobilization in decomposing leaf litter by a
novel 15N isotope pool dilution technique. Soil Biol Bio-
chem 42:1293–1302
486 Biogeochemistry (2014) 118:471–486
123





























