Embed Size (px)
Citation preview

Immunopharmacology, 21 (1991) 121-128 Elsevier
IMPHAR 00541
121
The role of monocytes and T cells in 1,25-dihydroxyvitamin D3 mediated inhibition of B cell function in vitro
Klaus MOiler, Carsten Heilmann, Lars K. Poulsen, Torben Barington and Klaus Bendtzen Laboratory of Medical Immunology, Medical Department TTA, Rigshospitalet State University Hospital, Copenhagen, Denmark
(Received 6 November 1990; accepted 21 January 1991)
Abstract: 1,25-Dihydroxyvitamin D 3 (1,25-(OH)2D 3) inhibits immunoglobulin production by human mononuclear cells (MNC) in vitro. The present study was undertaken to evaluate the role of T cells and monocytes in 1,25-(OH)2D 3 induced suppression of B cell functions. The synthetic vitamin D 3 analogue MC 903 was examined in parallel. 1,25-(OH)2D 3 and MC 903 showed a dose-related inhibition of IgM, IgG and IgA plaque-forming cells in poke-weed mitogen (PWM) activated cultures of MNC. This effect was most likely mediated through impairment o fT cell and monocyte functions. First, the inhibitory effect was seen after PWM stimulation, but not after Epstein-Barr virus stimulation which activates B cells independently of T cells and monocytes. Second, 1,25-(OH)2D 3 was not effective in T cell and monocyte-depleted cultures. Third, the effect of 1,25-(OH)2D 3 on PWM driven MNC was reversed by addition of the recombinant monokines: interleukin (IL)-I/~, tumour necrosis factor c~ (rTNFc~), rlL-6, as well as the lymphokines: lymphotoxin (rLT) and rlL-2. This is consistent with the finding that 1,25-(OH)2D 3 also inhibited IL-I~, TNFc~ and LT production in these cultures. The assumption that B cells are not directly affected by 1,25-(OH)2D 3 was further supported by the fact that 24 h of culture with 10- 8 M 1,25-(OH)2D 3 failed to reduce immunoglobulin production by in vivo activated B cells.
Key words: 1,25-Dihydroxyvitamin D3; MC 903; Monocyte; B cell; T cell; Cytokine
Introduction
The active metabolite of vitamin D3, 1,25-Dihy- d r o x y v i t a m i n D 3 (1,25-(OH)2D3) is a major physiological regulator of calcium metabolism. Increasing evidence indicates that 1,25-(OH)2D 3 also modulates lymphocyte growth and function. Awareness of this immunoregulatory activity was prompted by the demonstration of specific recep- tors for vitamin D 3 in monocytes and in activated lymphocytes (Provvedini et al., 1983).
Correspondence: Klaus Mtiller, Laboratory of Medical Immunology, Medical Department TTA, 7544, Rigshospita- let, Tagensvej 20, Copenhagen, DK-2200, Denmark.
1,25-(OH)2D 3 inhibits production of interleu- kin (IL)-2 (Tsoukas et al., 1984) and lymphocyte proliferation (Rigby et al., 1984; Tsoukas et al., 1984). It activates secretion of hydrogen peroxide by human monocytes (Cohen et al., 1986). It also induces cytotoxicity and increases the number of Fc receptors and the fusion of alveolar macro- phages (Abe et al., 1984). The production of IL-1 has been reported to be stimulated by 1,25- (OH)2D 3 (Hodler et al., 1985; Bhalla et al., 1986), but recent studies contradict these results, show- ing an inhibitory effect on the release of IL-I~//~ (Tsoukas et al., 1989) and on IL-I~, IL-6, and tumour necrosis factor c~ (TNF~) (Mtlller et al., 1990).
0162-3109/91/$03.50 © 1991 Elsevier Science Publishers B.V.

122
It is well documented that 1,25-(OH)2D 3 inhi- bits immunoglobulin production by human mono- nuclear cells (MNC) in vitro (Lemire et al., 1984; Iho et al., 1986). However, it is unclear whether this suppressive effect is exerted directly on the B cell, or indirectly via an effect on T cells or mono- cytes (Rigby, 1988). Contradicting results have been reported concerning the role of T cells, and little is known about the role of monocytes in 1,25-(OH)zD3-mediated immune regulation.
The present study was undertaken to evaluate the role ofT cells and monocytes in 1,25-(OH)2D 3 induced suppression of B cell functions. The syn- thetic vitamin D 3 analogue MC 903 (Binderup and Bramm, 1988; Fig. 1) was examined in parallel.
Materials and Methods
D 3- vitamins 1,25-(OH)2D3 and MC 903 were gifts from Leo Pharmaceutical Products (Ballerup, Denmark) (Fig. 1). They were dissolved in 98 ~o ethanol and stored at 4°C. Before each experiment fresh dilutions were made in sterile, isotonic saline. The vitamins were tested at final concentrations between 10- 8 and 10 - 12 M. The final concentra- tions of ethanol were always less than 0.01 ~o v/v. These concentrations of ethanol were without effect on immunoglobulin production or pro- liferation of MNC in vitro.
OH
OH
OH OH
l ~ , 2 5 - d i h y d r o x y - cholecalciferol
MC 903
Fig. 1. Chemical structures of 1,25-(OH)2D 3 and the syn- thetic analogue MC 903.
Cytokines Human rlL-l~, 108U/mg, was provided by Immunex Corp. (Seattle, WA). Human rlL-1/~, 2 x 106 U/mg, was a gift from Novo-Nordisk (Bagsv~erd, Denmark). Their biological activities were confirmed by the mouse thymocyte co-sti- mulatory assay (Svenson and Bendtzen, 1989). Human rTNF~, 4 x 10 v U/mg and recombinant lymphotoxin (rLT), 108 U/mg, were provided by Boehringer (Vienna, Austria). Their biological activities were tested by the L-M fibroblast cyto- toxicity assay (Fomsgaard et al., 1988). Human rIL-6, 5 × 10 7 U/mg, was a gift from Drs. Hirano and Kishimoto (Osaka University, Osaka, Japan). Its bioactivity was ascertained by use of the B9 hybridoma assay (Aarden et al., 1987). Human rIL-2, 5 × 10 6 U/mg, was a gift from Dr. Smith (Dartmouth Medical School, Hanover, NH). Its activity was tested by the CTLL-2 proli- feration assay (Gillis et al., 1978).
Cell cultures In vitro activated cells. MNC were obtained from healthy adult volunteers by gradient centri- fugation of heparinized blood on Ficoll-Hypaque (Lymphoprep, Nyegaard, Oslo, Norway). In some experiments, MNC were fractionated by the sheep erythrocyte rosette technique (Saxon et al., 1976). Rosette-negative cells were separated from the rosette-forming cells and free erythrocytes by gradient centrifugation on Ficoll-Hypaque (<4~oLeu 1 + T cells, by FACS analysis (FACStar, Becton Dickinson, Mountain View, CA). In other studies, MNC were depleted of monocytes by incubation for 1 h at 37 °C in plastic Petri dishes (Nunc, Roskilde, Denmark) followed by aspiration of the non-adherent cells. Less than 2~o were Leu M3 positive by FACS analysis.
The cells were washed and resuspended in RPMI 1640 (Gibco, Paisley, U.K.) supplemented with 10~o fetal calf serum (FCS) (Flow, Irvine, U.K.), glutamine (0.8 mM), penicillin and strep- tomycin. The cells, 106/ml, were cultured at 37 °C in flat-bottomed microtiter plates (Nunc) in a humidified atmosphere containing 5 Yo CO2. The

cultures were stimulated with 0.8 #g/ml of poke- weed mitogen (PWM) (Gibco), with or without recombinant cytokines. Other cultures were sti- mulated by preincubation for 2 h in a supernatant containing Epstein-Barr virus (EBV) from a mar- moset cell line (B 95-8).
Vitamin D 3 analogues were added at initiation of the cultures, since time course studies showed that optimal effects of 1,25-(OH)2D 3 were ob- tained when the vitamin was added at this time. After 6 days of culture, the supernatants were harvested and the contents of IgG were measured. The contents of cytokines were measured after 24 h of culture.
In vivo activated cells. Three healthy donors were vaccinated subcutaneously with 0.5 ml of Haemophilus influenzae type b capsular polysac- charide, containing 25/~g polyribosylribitol phos- phate (PRP) coupled to 18 #g diphtheria toxoid (Proibit, Connaught Laboratories, Swiftwater, PA). MNC, including anti-PRP antibody-se- creting cells (AbSC), were obtained 8-9 days after immunization and cultured for 24 h at 4 x 10 6
MNC/ml in the presence of 10 -SM 1,25- (OH)zD 3 or the corresponding concentration of ethanol. All cultures were run in triplicate.
Cell viability in PWM-stimulated cultures of MNC after 6 days was examined by Trypan blue exclusion. There was no difference between cell viability in control cultures (18~o dead cells and cultures with 10 8 M 1,25-(OH)2D 3 (19~o dead cells; n = 3). The recombinant cytokines did not affect cell viability.
Quantitation of lgM-, IgG- and lgA-secreting lym- phocytes After 6 days of culture, the cells were harvested, washed in minimum essential medium (Flow), and resuspended to a concentration of 0.5-1 × 10 6 cells/ml before the plaque-forming cell (PFC) assay. The reverse, indirect PFC assay for detection of antibody-secreting cells (AbSC), including IgM-, IgG-, and IgA-secreting cells
123
(IgM-, IgG-, IgASC) was carried out as described previously (Agger et al., 1982). Briefly, Staphylo- coccus aureus protein A (SPA) (Pharmacia, Uppsala, Sweden) was conjugated to SRBC by chromium chloride. MNC were mixed with SPA- SRBC, rabbit anti-human IgM, IgG or IgA (Dakopatts, Glostrup, Denmark), guinea pig serum and warm agar with DEAE-dextran. This suspension, 200 #1, was placed under a coverslip and allowed to harden before overnight incu- bation at 37 ° C. PFC were counted using a dark- field stereo microscope and expressed per 10 6
cultured MNC. Antigen-specific AbSC were detected by a mo-
dification of this assay by exchanging the target cells with SRBC to which PRP was coupled by means of chromium chloride, as described (Barington et al., 1990). The anti-IgG- and anti- IgA-immunoglobulins were used at final concen- trations of 1 : 2400 and 1 : 1200, respectively.
Radioimmunoassay for IgG Maxisorp test tubes (Nunc) were coated over- night with F(ab)2-fragments of rabbit anti-human IgG antibody (Code No. A407, Dakopatts) diluted to 10 - 4 in phosphate buffered saline. 500 #1 of standards and samples, diluted in RPMI-1640 with 7.4 g/1 of EDTA and 5~o FCS, were added to the tubes and incubated for 2 h at room temperature. After wash, 500/~1 of mouse- anti-human IgG monoclonal antibody (Code No. Mo 6014, Oxoid, Basingstoke, Hampshire, U.K.) were added and incubated for 2h at 37°C, followed by wash and incubation with 500/~1 of ~eSiodine-labelled rabbit-anti-mouse IgG, as de- scribed (Djurup et al., 1984). The tubes were washed and counted in a V-scintillation counter. A human serum pool containing 11.6 + 0.2 mg/ml of IgG by a standard nephelometric method was used as standard. For routine use, a measurement range of 1.5 to 500 ng IgG/ml was employed. The intra- and interassay variations were 8 and 23 ~o, respectively. There was no cross-reaction with IgA, IgM or IgE. (Poulsen et al., submitted).

124
ELISA for IL-1 ~ and TNF7 IL-10¢ and TNFc¢ were detected by a slightly modi- fied, previously described ELISA (Fomsgaard et al., 1988). Briefly, high-titered specific antisera to human rlL-lc¢ and rTNFc~ were generated by repeated injections of purified human recombi- nant IL-1 :¢ or -TNF~ into high-responder rabbits. The immunoglobulins were precipitated by am- monium sulphate and the ELISAs were establish- ed using biotinylated antibodies for detection and unconjugated antibodies for catching. These ELI SAs do not cross-react with rlL-6, rLT, rlL-2, rlL-lc¢ or rlFNT, and there is no cross-reaction between rlL-le and rTNFe. The detection limits were 10 pg/ml (IL-le) and 30 pg/ml (TNFc¢). The intra- and interassay variations were below 10 %.
FI
125
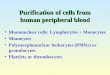
J 0 100 tv" I-- Z 0 75 (..)
LL 0 50 ,'X"
25
[3
1 2 5
d 0 100 r--- z 0 75 cJ
LL O 50 . \ "
25
IgM IgG
i i i i i 18
- 1 2 - 1 0 - 8 - 1 2 - 1 0 -
IgR
i i
-12 -10 -8
1,25-(OH)2D 3 (log M )
IgM IgG IgR
' 18 ; ' 18 ' ' ,'8 - 1 2 - 1 0 - - 2 - 1 0 - - 1 2 - 1 0 -
MC 903 (Log M)
Fig. 2. Effect of 1,25-(OH)2D 3 (A) and MC 903 (B) on num- bers of IgM, IgG and IgASC in cultures of PWM-stimulated MNC as measured by indirect plaque-forming cell assays.
* Significant effect of vitamin 9 3.
TABLE I
Effect of 1,25-(OH)2D3 and MC 903 on IgM-, IgG- and IgASC in Epstein-Barr virus-stimulated mononuclear cells (MNC) and T-cell-depleted MNC
Concentration of vitamin D 3
10 -SM 1 0 - r a M 10 12M
MNC 1,25-(OH)2D3:
IgM 119 (84/148) 145 (105/160) 97 (85/177) IgG 91 (59/209) 120 (36/244) 141 (106/205) IgA 141 (66/182) 102 (53/204) 132 (103/189)
MC 903: IgM 90 (76/143) 128 (87/176) 96 (70/155) IgG 118 (75/360) 91 (62/252) 110 (71,/177) IgA 127 (98/164) 114 (99/201) 100 (78/209)
T-cell-depleted MNC 1,25-(OH)2D 3
IgM 125 (62/172) ND a 100 (63/133) IgG 112 (84/182) ND 122 (80/145) IgA 118 (56/150) ND 92 (75/106)
MC 903: IgM 101 (82/143) ND 109 (98/192) IgG 130 (99/145) ND 145 (77/147) IgA 64 (25/114) ND 109 (91/131)
Results are given as percent of vitamin D 3 free controls (medians and quartiles; n = 5-7. The antibody-secreting cells (SC) were measured after 6 days of culture. a ND, not determined.
Statistics The Wilcoxon matched-pairs test was used for statistical evaluation of differences. P < 0.05 was considered significant.
Results
1 , 2 5 - ( O H ) 2 D 3 and MC 903 showed a dose- related inhibition of IgM, IgG, and IgASC in PWM activated cultures of MNC (Fig. 2). In this regard, the two compounds were almost equally potent. The contents of IgG in the culture super- natants were also reduced by 10 8M 1,25- ( O H ) E D 3 (36%, 15-52 (median, quartiles) of con- trois), while 25-OH D 3 w a s without effect (90%,

57-139 o f controls). Both 1,25-(OH)2D 3 and M C 903 failed to affect the P F C response by EBV stimulated M N C and by similarly activated T cell depleted M N C (Table I). These da ta suggested that the pr imary site o f ho rmone action is on T cells and/or monocytes .
Fur ther studies indicated that the inhibitory effect o f 1,25-(OH)2D 3 on the PWM-dr iven P F C response was partially overcome by addition of a condi t ioned med ium f rom a PWM-dr iven M N C culture (data not shown). Therefore, experiments were carried out in order to evaluate the ability of different r ecombinan t cytokines to overcome the inhibitory effect o f 1,25-(OH)zD 3 in these cul- tures. The monokines , r IL-6 (2000 U/ml) and rIL-1/? (100 U/ml) , as well as the lymphokines , r L T (4000 U/ml) and r IL-2 (20 U/ml) , all signifi- cantly counterac ted the inhibitory effect o f 1,25- ( O H ) z D 3 on I g G product ion (Table II). Consis- tent with this finding, Table I I I shows that 1 0 - S M 1,25-(OH)zD3 and M C 903, but not 2 5 - O H D3 (not shown), reduced the contents o f I L - l e and T N F e in the supernatants of the PWM-s t imu la t ed M N C cultures.
To evaluate further the role of lymphocytes and monocytes , the effect of 1,25-(OH)2D 3 in T cell- and monocy te depleted M N C cultures was studied. P W M alone failed to stimulate these cul- tures. However , the combinat ion of P W M and
125
TABLE II
Effect of 1,25-(OH)2D 3 on IgG production by mononuclear cells (MNC) stimulated by poke-weed mitogen, alone or with cytokine
Treatment Poke-weedmitogen +Cytokine U/ml
IgG in 6-day cultures
1 , 2 5 - ( O H ) 2 D 3- + 10- s M free control 1,25-(OH)2D 3
pg/ml % of 1,25-(OH)2D 3- free control
- 0.3 (0.2-4.7) 36 (15-52) a rTNFe 2000 3.8 (1.8-18.8) 74 (53-115) rTNFc~ 4000 6.5 (4.8-42.0) 65 (41-127) rIL-l/3 50 1.4 (0.5-6.7) 49 (16-94) rIL-1/~ 100 1.3 (0.9-40.3) 97 (44-123) b rIL-6 1000 2.5 (I.I-35.4) 67 (42-108) rIL-6 2000 4.4 (2.8-30.2) 90 (59-108) u rLT 2000 3.0 (1.0-12.4) 34 (5-81)" rLT 4000 1.6 (1.0-6.9) 90 (51-125) 8 rIL-2 10 1.1 (0.2-2.7) 60 (41-101) rlL-2 20 0.9 (0.2-4.4) 91 ( 4 4 - 3 7 1 ) b
Results are shown as medians and quartiles; n = 6. a Significant inhibition of IgG production compared with
1,25-(OH)2D 3 free control. Significantly less inhibitory effect of 1,25-(OH)2D 3 com- pared with the inhibitory effect obtained in cultures stimu- lated by poke-weed mitogen alone (A).
2000 U/ml r L T significantly stimulated I g G pro-
duction (Table IV). 1,25-(OH)2D 3 significantly
TABLE III
Effect of 1 , 2 5 - ( O H ) 2 D 3 on I L l e and TNFe production by mononuclear cells (MNC)
Poke-weed 1,25-(OH)2D 3 mitogen
nil 10 -8 M 10-12 M
IL-le (pg/ml) - 186 (36-400) 32 (25-10|) a + 275 (192-873) 195 (75-298) a
TNFe (pg/ml) - 205 (141-811) 128 (80-280) + 380 (123-1040) 148 (50-633)"
ND b 230 (141-365)
ND 221 (72-815)
Results are given as pg/ml (medians and quartiles; n = 6). The cytokines were measured after 24 h of culture. a Significant inhibition compared with 1,25-(OH)2D 3 free control. b Not determined.

126
T A B L E IV
Effect of 1,25-(OH)2D 3 on IgG product ion in cultures of monocyte- and T-cell-depleted mononuclear cells (MNC) stimulated with poke-weed mitogen without or with lymphotoxin
Poke-weed Lymphotoxin IgG in 6 day cultures mitogen 2000 U/ml
1,25-(OH)zD3-free control (#g/ml)
+ 10- 8 M 1,25-(OH)2D 3
~o of 1,25-(OH)2D3-free control
Monocyte-deple ted M N C a
- - 0.034 (0.0-0.09) + - 0.045 (0.0-0.26) + + 0.3 (0.012-34) 157 (72-235)
T cell-depleted M N C b
- - 0.082 (0.015-0.23) + - 0.096 (0.0085-0.25) + + 0.35 (0.05-3.6) 101 (43-400)
Results are shown as medians and range. a n = 4 . b n = 6 .
inhibited IgG production by MNC stimulated in this way (Table 1), but it failed to reduce IgG production both in T cell and in monocyte-de- pleted cultures (Table 4).
The assumption that B cells are not directly affected by 1,25-(OH)zD 3 was further supported by the fact that 24h of culture with 10 8M 1,25-(OH)2D 3 did not reduce anti-PRP-antibody secretion by in vivo activated B cells as measured by antigen-specific AbSC (Table V).
Discussion
A number of studies have demonstrated an inhibi- tory effect of 1,25-(OH)2D 3 on immunoglobulin production in vitro (Lemire et al., 1984; Iho et al., 1986). It is, however, uncertain whether this effect is exerted directly on the B cells or through impaired T cell or monocyte functions. Receptors for 1,25(OH)2D 3 have been demonstrated in mo- nocytes, in mitogen-activated MNC, in lines of
T A B L E V
Effect o f 1,25-(OH)2D 3 on numbers of in vivo-activated ant i -PRP secreting cells
Exp. no. 10 8 M 1,25-(OH)2D 3 IgG IgA
1 - 387 (31.7) 953 (26.4) + 376 (17.3) 1131 (28.8)
2 - 436 (20.3) 655 (47.6) + 434 (16.7) 751 (17.3)
3 - 255 (26.6) 401 (22.3) + 242 (12.0) 475 (81.4)
The results are given as the number of ant i -PRP secreting cells per 106 cultured cells after 24 h (means and SEM; n = 3). Each exper iment was run in triplicate. There were too few IgM P F C for evaluation.

malignant B- and T lymphocytes, and in EBV- transformed B cells after 2-3 weeks of culture (Provvedini et al., 1983, 1986). These findings suggest that receptors may also be present in mi- togen- and EBV-stimulated B cells in short-term cultures like the ones studied here. Since T cells, B cells and monocytes may carry these receptors, the site of hormone action may be on any of these cells.
Proliferation of human B cells to lectin stimula- tion has been reported to be both sensitive (Iho et al., 1986) and insensitive (Lemire et al., 1984) to 1,25-(OH)2D3. Staphylococcus Cowan-I (B lym- phocytotrophic)-driven PFC formation has been reported to be sensitive to 1,25-(OH)2D3(Iho et al., 1986). Later studies were, however, unable to demonstrate any notable reduction of the IgG contents in the supernatants of similarly stimu- lated, partially purified B cell cultures (Shiozawa etal., 1987; Chen etal., 1987). Furthermore, Komoriya et al. (1985) found that immunoglo- bulin production by murine splenocytes driven by a T cell independent antigen failed to respond to 1,25-(OH)2D 3. Finally, Provvedini and co- workers (1986) found that 1,25-(OH)zD 3 inhi- bited IgG production by a lymphoblast cell line (EBV transformed B cells in 3-week cultures), but short-term cultures were not investigated.
The data presented here demonstrate an inhibi- tory effect on immunoglobulin production by PWM stimulated MNC measured by PFC assays and IgG RIA. This inhibitory effect of 1,25- (OH)aD 3 was most likely mediated through impairment ofT cell and monocyte functions, and not by a direct effect on B cells. First, the inhi- bitory effect was seen after PWM stimulation, which is dependent upon the presence of mono- cytes and T cells (Heegaard and MOiler, 1988), but not after EBV stimulation which directly acti- vates B cells. Second, in cultures stimulated by PWM plus 2000 U/ml rLT, 1,25-(OH)zD 3 was effective only in the presence of both T cells and monocytes. Third, the reversal of the effect of 1,25-(OH)2D3 by monokines as well as lympho- kines suggests that the inhibitory effect of 1,25- ( O H ) 2 D 3 may be mediated through impairment of
127
monocyte and T cell functions. 1,25-(OH)2D3 has been shown to inhibit the in vitro production of IL-2 (Tsoukas et al., 1984) and LT (MOiler et al., unpublished data). 1,25-(OH)2D 3 also inhibited IL-I~ and TNFct production in the PWM stimu- lated cultures studied here. The above conclusion is further supported by the fact that 24 h of incu- bation with 1,25-(OH)zD 3 failed to reduce immu- noglobulin production by in vivo activated B cells, whereas production of cytokines by T cells and monocytes was inhibited by this treat- ment.
MC 903 is a vitamin D 3 analogue with limited calcium metabolic activity (Binderup and Bramm, 1988), possibly due to rapid degradation in the circulation. We have previously shown that 1,25- (OH)2D 3 and MC903 both suppress pro- liferation of rlL-1 driven murine thymocytes (Mtiller etai., 1988). In the present study, MC 903 and 1,25-(OH)zD 3 were equally potent with respect to inhibition of the PFC response.
In conclusion, this report confirms the previous observation that 1,25-(OH)2D3 inhibits B cell functions in vitro. It also provides evidence that this effect is caused by impairment of both T cell and monocyte functions, most likely mediated via reduced release of cytokines by these cells.
Acknowledgements
Financial support was obtained from Leo Phar- maceutical Company, The Danish Biotechnology Programme, and the Danish Medical Research Council. We thank Anne Ambjomsen for expert technical assistance.
References
Aarden LA, De Groot ER, Schaap OL, Lansdorp PM. Pro- duction of hybridoma growth factor by human monocytes. Eur J Immunol 1987; 17: 1411-1416.
Abe E, Shiina Y, Miyaura C, Tanaka H, Hayashi T, Kanegasaki S, Saito M, Nishii Y, DeLuca HF, Suda T. Activation and fusion induced by 1,25-dihydroxyvitamin D3 and their relation in alveolar macrophages. Proc Natl Acad Sci USA 1984; 81: 7112-7116.

128
Agger R, Petersen J, Dinesen B, Wiik A, Andersen V. Pro- duction and secretion of immunoglobulins by in vitro-acti- vated human B lymphocytes. Allergy 1982; 37: 179-185.
B arington T, Heilmann C, Andersen V. Quantitation of anti- body-secreting cells in the blood after vaccination with Haemophilus influenzae type b conjugate vaccine. Scand J Immunol 1990; 31: 515-522.
Bhalla AK, Amento EP, Krane SM. Differential effects of 1,25-dihydroxyvitamin D3 on human lymphocytes and monocyte/macrophages: inhibition of interleukin-2 and augmentation of interteukin-1 production. Cell Immunol 1986; 98: 311-322.
Binderup L, Bramm E. Effects of a novel vitamin D analogue MC 903 on cell proliferation and differentiation in vitro and on calcium metabolism in vivo. Biochem Pharmacol 1988; 37: 889-895.
Chen W-C, Vayuvegula B, Gupta S. 1,25-dihydroxyvitamin D3-mediated inhibition of human B cell differentiation. Clin Exp Immunol 1987; 69: 639-646.
Cohen MS, Mesler DE, Snipes RG, Gray TK. 1,25-dihy- droxyvitamin D3 activates secretion of hydrogen peroxide by human monocytes. J Immunol 1986; 136: 1049-1053.
Djurup R, Sondergaard I, Magnusson CGM, Weeke B. A three layer immunoradiometric assay for determination of IgG-subclass antibodies in human sera ('IgG-subclass RAST'). Allergy 1984; 39: 51-63.
Fomsgaard A, Worsaae H, Bendtzen K. Detection oftumour necrosis factor from lipopolysaccharide-stimulated human mononuclear cells by enzyme-linked immunosorbent assay and cytotoxicity bioassay. Scand J Immunol 1988; 27: 143-157.
Gillis S, Ferm MM, Ou W, Smith KA. T cell growth factor: parameters of production and a quantitative microassay for activity. J Immunol 1978: 120: 2027-2031.
Heegaard PHH, Mailer K. Lectins and the immune system. J Immunol Immunopharmacol 1988; 8: 239-247.
Hodler B, Evequoz V, Trechsel U, Fleisch H, Stadler B. Influence of vitamin D3, metabolites on the production of interleukin 1,2 and 3. Immunobiology 1985; 170: 256-269.
Iho S, Takahashi T, Kura F, Sigiyara H, Hoshini T. The effect of 1,25 Dihydroxyvitamin D3 on in vitro immunoglo- bulin production in human B-cells. J Immunol 1986; 136: 4427-4431.
Komoriya K, Nagata I, Tsuchimoto M, Kunisawa K, Takeshita T, Naruchi T. 1,25 dihydroxyvitamin D3 and 1,24-dihydroxy-vitamin D3 suppress in vitro antibody re- sponse to T-cell-dependent antigen. Biochem Biophys Res Comm 1985; 127: 753-758.
Lemire JM, Adams JS, Sakai R, Jordan SC. 1-alpha,25-dihy- droxyvitamin D3 suppresses proliferation and immunoglo- bulin production by normal human peripheral blood mononuclear cells. J Clin Invest 1984; 74: 657-661.
Mfiller K, Diamant M, Bendtzen K. Down-regulation of interleukin-l~, tumor-necrosis factor and interleukin-6 production by 1,25-Dihydroxyvitamin D3 and an analog, MC 903. Clin Rheumatol 1990; 9: 137-137.
Mtiller K, Svenson M, Bendtzen K. l~,25-dihydroxyvitamin D3 and a novel vitamin D analogue MC 903 are potent inhibitors of human interleukin 1 in vitro. Immunol Lett 1988; 17: 361-366.
Provvedini DM, Tsoukas CD, Deftos LJ, Manolagas SC. 1,25-dihydroxyvitamin D3 receptors in human leukocytes. Science 1983: 221: 1181-1183.
Provvedini DM, Tsoukas CD, Deftos LJ, Manolagas SC. 1 alfa,25-dihydroxyvitamin D3-binding macromolecules in human B lymphocytes: effects on immunoglobulin produc- tion. J Immunol 1986; 136: 2734-2740.
Rigby WFC, Denome S, Fanger MW. Regulation of lym- phokine production and human T lymphocyte activation by 1,25-dihydroxyvitamin D3. J Clin Invest. 1987; 79: 1659-1664.
Rigby WFC, Stacy T, Fanger MW. Inhibition of T lympho- cyte mitogenesis by 1,25-dihydroxyvitamin D3 (Calcitriol). J Clin Invest 1984; 74: 1451-1455.
Rigby WFC. The immunobiology of vitamin D. Immunol Today 1988; 9: 54-58.
Saxon A, Feldhaus J, Robins A. Single separation of human T and B cells using AET treated SRBC rosettes. J Immunol Meth 1976; 12: 285-288.
Shiosawa K, Tanaka Y, Fujita T. 1,25 Dihydroxyvitamin D3 inhibits proliferative response ofT- and B-lymphocytes in a serum free culture. Int J Immunopharmacol 1987; 9: 719-723.
Svenson M, Bendtzen K. Human urine deoxyribonuclease increases endogenous thymidine in the mouse thymocyte interleukin 1 assay: An artificial interleukin-1 inhibitor. Cytokine 1989; 1: 52-55.
Tsoukas CD, Provvedini DM, Manolagas SC. 1,25-dihy- droxy-vitamin D3: a novel immunoregulatory hormone. Science 1984; 224: 1438-1440.
Tsoukas CD, Watry D, Escobar SS, Provvedini DM, Dinarello CA, Hustmyer FG, Manolagas SC. Inhibition of interleukin-1 production by 1,25-dihydroxyvitamin D3. J Clin Endocrinol Metab 1989; 69: 127-133.