Embed Size (px)
Citation preview

Human Movement Science 31 (2012) 1014–1036
Contents lists available at SciVerse ScienceDirect
Human Movement Science
journal homepage: www.elsevier .com/locate/humov
The role of haptic information in shaping coordinationdynamics: Inertial frame of reference hypothesis
Paula L. Silva ⇑, Michael T. Turvey 1
University of Connecticut, Department of Psychology, Center for the Ecological Study of Perception and Action,406 Babbidge Road, Unit 1020, Storrs, CT 06269-1020, USA
a r t i c l e i n f o a b s t r a c t
Article history:Available online 26 December 2011
PsycINFO classification:2300232023302300
Keywords:Haptic proprioceptionAction coordination
0167-9457/$ - see front matter � 2011 Elsevier B.Vdoi:10.1016/j.humov.2011.10.002
⇑ Corresponding author. Present address: UniveTerapia Ocupacional, Departamento de FisioterapiMG, Brazil. Tel.: +55 31 88445349.
E-mail addresses: [email protected], paulala1 Tel.: +1 860 4863906.
Current research suggests that non-visual perception of the spatialorientation of body segments is tied to vectors representative oftheir mass moment distribution (vmm). Our question was whetherthe relative orientation of vmm of right and left hands (Dvmm = vmm
left � vmm right) constitutes haptic information supporting bimanualcoordination and, if so, how it contributes to coordination dynamics.Blindfolded participants coordinated the motions of a pair of cross-shaped, hand-held pendulums that were either symmetricallyloaded (Dvmm = 0) or asymmetrically loaded (Dvmm – 0). The signand magnitude of Dvmm, in particular of the first moment vector, sys-tematically affected the pattern of coordination (indexed by meanrelative phase /), but not its stability. These results suggest that(1) Dvmm specifies a frame of reference about which coordinationis organized; and (2) that the changes in pattern were a function ofthe experimentally induced biases in this perceptual frame of refer-ence and not a function of a functional asymmetry akin to detuning.The implications of the findings to the understanding of perceptualregulation of interlimb coordination were discussed.
� 2011 Elsevier B.V. All rights reserved.
1. Introduction
The ability to coordinate the motions of limbs and limb segments underlies the performance ofmany everyday activities of which walking and clapping are representative examples. The highly
. All rights reserved.
rsidade Federal de Minas Gerais, Escola de Educação Física, Fisioterapia ea, Av. Antonio Carlos 6627, Campus Pampulha, 31270-010 Belo Horizonte,
[email protected] (P.L. Silva), [email protected] (M.T. Turvey).

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1015
regular coordination patterns that typify these functional activities are not a matter of two or morelimbs (or limb segments) executing similar pre-programmed trajectories in relative independenceof each other. Rather, in the face of contextual change, the intended pattern is preserved through re-ciprocal adjustments to such trajectories (Black, Riley, & McCord, 2007). Pointedly, any changes in thetrajectories of one limb that could potentially deviate the system from its stable coordinative state arecompensated by complimentary changes in the trajectories of the other limb, such that the stablecoordinative state is preserved. This ability of biological movement systems to maintain functionalintegrity under varying context conditions necessarily implicates perception, that is, the use of infor-mation about the states of body segments to modulate their interactions according to specific taskgoals. Hence, central to the understanding of movement coordination supporting functional behavioris an understanding of its informational basis.
Bernstein (1967, 1996) strongly suggested that the fluidity and context sensitivity of large scalemovement patterns are only made possible by information revealed in the patterned activity ofmechanoreceptors in response to distortions of body tissues due to movements and environmentallyimposed contact forces. Awareness of the states of body segments (and of the environment adjacent tothe body) supported by this information is referred to as haptic perception (Gibson, 1966). Empiricaldemonstrations of the central contribution of haptic perception to interlimb coordination perfor-mance have been mainly obtained from studies with deafferented patients (Spencer, Ivry, Cattaerti,& Semjen, 2005) and from studies in which vibratory stimulation was applied to muscles or tendonsduring interlimb coordination (Steyvers, Verschueren, Levin, Ouamer, & Swinnen, 2001; Verschueren,Swinnen, Cordo, & Dounskaia, 1999). These studies have shown that changes in the afferent flow frommechanoreceptors induced by pathological conditions or atypical mechanical perturbations result in(1) deviations of the observed pattern of coordination from the intended one, and (2) reduced coordi-nation stability.
The changes in coordination performance observed as a result of altered mechanoreceptor activityare indicative of reduced coupling strength between limbs. There is little doubt, therefore, that themovement system exploits haptic information about the states of body segments (or haptic proprio-ception) to couple their motion and preserve the intended coordination pattern. However, the specificinformational parameters realized in the patterned activity of mechanoreceptors during performanceof coordination tasks are yet to be identified. Arguably, the identification of such parameters wouldallow a more systematic examination of the role of haptic proprioceptive information in shaping coor-dination dynamics.
1.1. Candidate informational parameters for haptic proprioception
A traditional assumption that persists to this date is that the temporally and spatially distributedpattern of mechanoreceptor activity that supports haptic proprioception refers to geometric parame-ters (Cordo, Flores-Vieira, Verschueren, Inglis, & Gurfinkel, 2002; Ribot-Ciscar, Bergenheim, Albert, &Roll, 2003). Cordo et al. (2002) argued that among the geometric variables attributed to haptic propri-oception, joint angle is the most fundamental one because more global spatial variables, such as theorientation of the whole limb, can be computed from it. The assumption that joint angle is the primaryreferent of haptic proprioception is rooted in early studies whose results suggested that individualswere able to reproduce previously experienced joint position (Cohen, 1958) and to match the joint po-sition of two different limbs without the benefit of vision (Horch, Clark, & Burgess, 1975).
An alternative possibility, first suggested by Pagano and Turvey (1995), is that the inertial axes (ormass moment vectors vmm) of body segments might constitute spatially oriented parameters support-ing haptic proprioception (Pagano, Garrett, & Turvey, 1996; Pagano & Turvey, 1995; van de Langen-berg, Kingma, & Beek, 2007, 2008). In particular, two candidate variables have been identified inthe literature for perception of limb orientation: ve3 (the inertial eigenvector defining the symmetryaxis of a limb or object which offers least resistance to rotational acceleration) and vCM (the centerof mass vector, defining an axis about which the static torque is zero). Given the cylindrical symmetryof body segments, ve3 and vCM typically coincide with their longitudinal axes. Hence, these variables(jointly referred to in the present paper as vmm) can be seen as informational parameters that directlyspecify limb orientation.

1016 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
Current research in dynamic touch favors the latter possibility and strongly suggests that sensitiv-ity to joint angle (hereafter referred to as a) might not be primary. Specifically, when a person, withoutbenefit of vision, attempts to align one arm with respect to the other, he or she does so by aligning themass moment vectors of right and left limbs and not by matching their joint angles as it has been tra-ditionally assumed (Pagano et al., 1996; van de Langenberg et al., 2007). This behavior has been ver-ified by (1) breaking the coincidence between the limbs’ geometric and inertial axes via theattachment of objects of the kind displayed in Fig. 2, and (2) measuring the difference between theangular positions of the participants’ right and left arms (Da = a right � a left).
Under conditions in which unilateral shifts in vmm away from the limb’s geometrical axis wereintroduced, a Da – 0 was observed (see Fig. 2, right panel). The magnitude of Da was proportionalto the experimentally induced difference between the direction of the mass moment vectors of rightand left limbs (hereafter referred to as Dvmm). The implication is that haptic perception of relative limbposition seems to be constrained by inertial and not geometrical parameters. The question of the pres-ent research is whether Dvmm is also exploited to monitor and regulate the spatial relations of thelimbs during interlimb rhythmic coordination, and if so, how it affects coordination performance. Adetailed characterization of coordination performance as a function of selective manipulations ofDvmm might provide an entry point into these questions.
1.2. A characterization of coordination performance rooted in dynamics
Bimanual rhythmic coordination offers a potentially rich set of dynamical measures for character-izing performance changes induced by haptic perceptual manipulations of the kind introduced by Pag-ano and Turvey (1995). The essential features of the cooperative behavior of two hands oscillating atthe same tempo can be fully characterized in relative ignorance of the dynamics of the individualhands (and related neuromuscular and physiological components). As developed by Kelso (1995)and colleagues (Fuchs, Jirsa, Haken, & Kelso, 1995; Haken, Kelso, & Bunz, 1985; Kelso, Del Colle, &Schöner, 1990; Schöner, Haken, & Kelso, 1986), bimanual 1:1 rhythmic coordination can be usefullymodeled by a motion equation in the collective variable of relative phase /—the difference in thephase angles (hleft � hright) of the left and right hands.
_/ ¼ � dVd/þ
ffiffiffiffiQ
pft ; ð1Þ
where the over dot signifies /’s rate of change and V is the potential function with respect to /:
Vð/Þ ¼ �d/� a cos /� b cos 2/: ð2Þ
Specifically, V(/) describes a potential landscape composed of two valleys. Intuitively, the currentcoordinative state of the system (or /) may be understood as a particle that eventually reaches thebottom of one of the potential wells, achieving then a steady state.
The positions of the valleys at / = 0� (representing in-phase coordination) and / = ±180� (repre-senting anti-phase coordination) correspond to stable solutions of Eq. (1). Coordinated rhythmicmovements are constrained to a range of / at or near these stable solutions, called attractors (/⁄), withdiffering attractive strengths. Intuitively, the current coordinative state of the system (or /) may beunderstood as a particle that eventually reaches the bottom of one of the potential wells, achievingthen a steady state (Fig. 1).
The cosine functions in / and 2/ determine the relative attractive strength or stability of the attrac-tors at / = 0� (more stable) and / = ±180� (less stable). As illustrated in Fig. 1, the stability of these twoattractive coordinative states (or k) is reflected in the relative depths and slopes of the potential wells,which determine how fast the system returns to its stable state after perturbations. Task constraints,such as increases in movement frequency, results in a progressive decrease in attractor stability, spe-cially of the one at or close to 180�. Such changes in coordination performance are captured by param-eters a and b. Finally, d is an imperfection parameter that accounts for the effects of differences in thecomponent hands giving rise to the coordination patterns (Fuchs et al., 1995; Kelso et al., 1990; Park &Turvey, 2008).
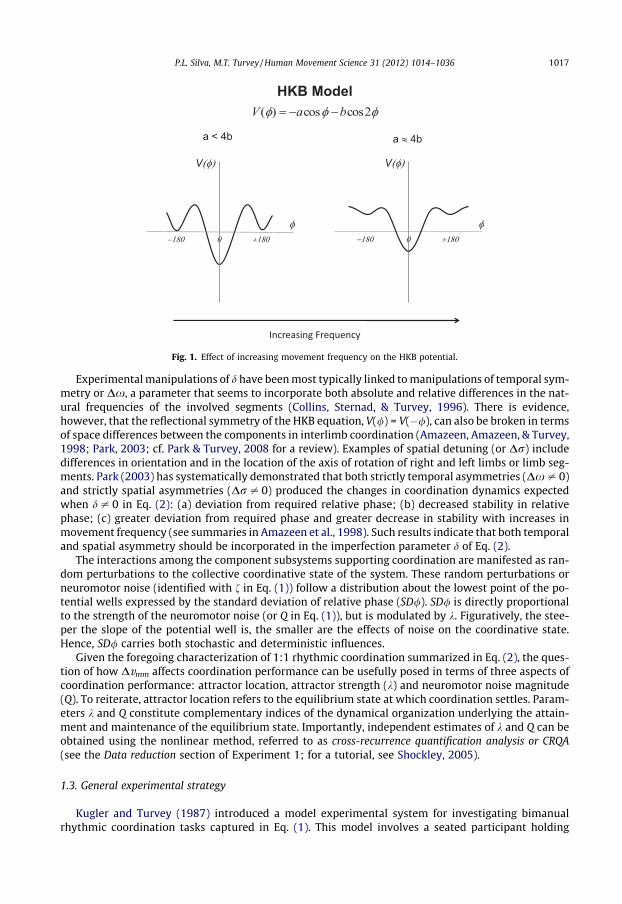
Fig. 1. Effect of increasing movement frequency on the HKB potential.
P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1017
Experimental manipulations of d have been most typically linked to manipulations of temporal sym-metry or Dx, a parameter that seems to incorporate both absolute and relative differences in the nat-ural frequencies of the involved segments (Collins, Sternad, & Turvey, 1996). There is evidence,however, that the reflectional symmetry of the HKB equation, V(/) = V(�/), can also be broken in termsof space differences between the components in interlimb coordination (Amazeen, Amazeen, & Turvey,1998; Park, 2003; cf. Park & Turvey, 2008 for a review). Examples of spatial detuning (or Dr) includedifferences in orientation and in the location of the axis of rotation of right and left limbs or limb seg-ments. Park (2003) has systematically demonstrated that both strictly temporal asymmetries (Dx – 0)and strictly spatial asymmetries (Dr – 0) produced the changes in coordination dynamics expectedwhen d – 0 in Eq. (2): (a) deviation from required relative phase; (b) decreased stability in relativephase; (c) greater deviation from required phase and greater decrease in stability with increases inmovement frequency (see summaries in Amazeen et al., 1998). Such results indicate that both temporaland spatial asymmetry should be incorporated in the imperfection parameter d of Eq. (2).
The interactions among the component subsystems supporting coordination are manifested as ran-dom perturbations to the collective coordinative state of the system. These random perturbations orneuromotor noise (identified with f in Eq. (1)) follow a distribution about the lowest point of the po-tential wells expressed by the standard deviation of relative phase (SD/). SD/ is directly proportionalto the strength of the neuromotor noise (or Q in Eq. (1)), but is modulated by k. Figuratively, the stee-per the slope of the potential well is, the smaller are the effects of noise on the coordinative state.Hence, SD/ carries both stochastic and deterministic influences.
Given the foregoing characterization of 1:1 rhythmic coordination summarized in Eq. (2), the ques-tion of how Dvmm affects coordination performance can be usefully posed in terms of three aspects ofcoordination performance: attractor location, attractor strength (k) and neuromotor noise magnitude(Q). To reiterate, attractor location refers to the equilibrium state at which coordination settles. Param-eters k and Q constitute complementary indices of the dynamical organization underlying the attain-ment and maintenance of the equilibrium state. Importantly, independent estimates of k and Q can beobtained using the nonlinear method, referred to as cross-recurrence quantification analysis or CRQA(see the Data reduction section of Experiment 1; for a tutorial, see Shockley, 2005).
1.3. General experimental strategy
Kugler and Turvey (1987) introduced a model experimental system for investigating bimanualrhythmic coordination tasks captured in Eq. (1). This model involves a seated participant holding

1018 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
one pendulum (typically a cylindrical rod) in each hand. Typically, each of the two pendulums isswung parallel to the sagittal plane of the body (i.e., in parasagittal planes) about an axis throughthe wrist. The pendulums can vary physically in shaft length, the weight attached to the bob, or both.
The pendulum paradigm has been traditionally used to introduce frequency competition betweenthe component rhythmic synergies (the right and left hands) by introducing differences in the rightand left hand-pendulum systems’ equivalent length (LE). LE can be calculated using the length andmass of the pendulum, hand, and added weight following standard methods for modeling any arbi-trary rigid body oscillating about a fixed point as a simple pendulum (see Kugler & Turvey, 1987 fordetails). The eigenfrequency (x) of each hand-pendulum system can be approximated by x = (g/LE)1/2. Hence, Kugler and Turvey’s system of hand-held pendulums allows one to experimentally con-trol the functional asymmetries between the hands performing the oscillations and to evaluate theireffects on coordination performance. Accordingly, the right and left hand-held pendulum systems willbe referred to in the present study simply as the right and left hand.
The basic experimental task in the present study combines Kugler and Turvey’s pendulum para-digm with the procedure introduced by Pagano and Turvey (1995) to break the coincidence betweenthe geometric and inertial axes of body segments (see Fig. 2). Participants were asked to coordinatetheir hand motions under selective manipulations of Dvmm (affecting the perceived relative orienta-tion of the hands). This experimental strategy is based on the premise that if Dvmm is exploited duringthis task, it will interfere with performance in a manner revealing of its role in shaping coordinationdynamics. The experimental protocol was approved by the Institutional Review Board for ethics in re-search of the University of Connecticut.
1.4. How might Dvmm shape coordination dynamics?
It is hypothesized that Dvmm constitutes an informational parameter exploited by the haptic per-ceptual system to couple the motions of body segments during rhythmic coordination tasks. Morespecifically, it is hypothesized that spatial–temporal patterns of interlimb coordination are organizedand maintained, at least in part, through regulation of interlimb spacing with respect to an inertial andnot to a geometrical frame of reference.
Fig. 2. Mass moment vector manipulations. Upper panel: Illustration of the experimental paradigm introduced by Pagano et al.(1996). The mass distribution on each object (alias splint) was manipulated so as to either maintain the coincidence betweenthe direction of the mass moment vector and the longitudinal axis of the forearm (left panel) or break the coincidence betweenthe two (right panel). The latter condition reveals that when attempting to align the right and left arms, participants aresensitive to Dvmm and not Da. Lower panel: Illustration of the present experimental paradigm. Coordination will be comparedbetween conditions where Dvmm = 0 (left panel) and conditions in which Dvmm – 0 (right panel).

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1019
If the synchronization of the hands relies on an inertial frame of reference, mean / should vary as afunction of both the sign and magnitude of Dvmm. For example, for the conditions in which Dvmm < 0(vmm right > vmm left), perfect in-phase coordination should feel as if the right hand is leading the leftdue to a bias in the proprioceptive frame of reference introduced by the unilateral shift in vmm. In thatcase, participants should compensate by adjusting the relative orientation (Da) between the handsaccordingly (i.e., advancing the left hand with respect to the right as illustrated in Fig. 2, bottom rightpanel).
The hypothesized adjustments in Da and shifts in mean / alone, however, are not sufficient to pro-vide support for a perceptual role of Dvmm given that the same pattern of results would be expected ifDvmm worked as an imperfection parameter (d in Eq. (2)). The effects of Dvmm on k and Q, however, doprovide a means to distinguish these hypotheses. Specifically, if Dvmm is akin to Dr, a spatial instan-tiation of d (Spatial Detuning hypothesis), any shifts in / from the required coordinative state (/ = 0� inthe present experiment) when Dvmm – 0 should be associated with a decrease in k. Figuratively, thespatial detuning hypothesis predicts shifts in the position and a decrease in the slope of the potentialwell constraining in-phase coordination (see Fig. 3, left panel).
If, in contrast, Dvmm provides a frame of reference for interlimb coupling, changes in the value ofDvmm should lead to the organization of different relations between the trajectories of the hands with-out sacrificing the stability of coordination. More precisely, the hypothesized changes in / as a func-tion of Dvmm – 0 are not expected to be related to decreases in k (Inertial Frame of Reference or IFRhypothesis). In dynamical terms, manipulations of the magnitude and sign of Dvmm should be relatedto corresponding shifts in attractor location without an associated change in the shape of the potentialwell (Fig. 3, right panel).
The noise parameter Q is related to activity evolving at time scales faster than the intersegmentalcoordination. Increases in Q have been interpreted as indexing control processes that are harnessed todeal with ongoing constraints on the dynamics of coordination (Schmidt, Shaw, & Turvey, 1993).Schmidt et al. (1993) further suggested that control processes subsidiary to the main dynamical re-gime (captured by Eq. (2)) are processes of perceptual tuning. Considering this argument, changesin the magnitude of Q may be observed if additional processes of perceptual tuning are needed whenDvmm – 0.
1.5. IFR hypothesis redux: An inventory of expected findings
If the IFR hypothesis holds, what changes in coordination performance would be expected? First,changes in Dvmm sign and magnitude should result in corresponding changes in attractor location. Sec-ond, the spatial relation between the hands (that is, Da) giving rise to / should change so as to align
Fig. 3. Haptic information and coordination dynamics. Competing hypothesis regarding the effects of Dvmm on the performanceof in-phase interlimb coordination.

1020 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
the vmm directions of the right and left hands. Third, the shifts in attractor location should not beaccompanied by changes in the underlying dynamics of coordination (that is, no changes in SD/, sta-bility k or noise Q).
2. Experiment 1: Effect of Dvmm magnitude on coordination dynamics
In Experiment 1, participants were required to produce in-phase coordination of a pair of hand-held pendulums under different magnitudes of unilateral vmm shift. Specifically, vmm direction of theright hand (with respect to its geometrical direction) was either equal to vmm direction of the left hand(Dvmm = 0) or shifted forward by two different amounts (Dvmm < 0). The effect of this manipulation onthe performance of the coordination task provided an initial examination of whether and how Dvmm
constrains coordination dynamics.
2.1. Method
2.1.1. ParticipantsEleven undergraduate students from the University of Connecticut participated in the present
study in partial fulfillment of course requirements. Eight participants classified themselves as right-handed and three as left-handed. None of them had previous experience with the task. The informa-tion given to participants prior to the experiment focused on its general purpose and procedure. Spe-cific description of the experimental manipulation and experimental hypotheses were only providedat the end.
2.1.2. MaterialsA 6D-Research (Skill Technologies, Inc., Phoenix, Arizona) motion analysis system was used to re-
cord the movements of the right and left pendulums. One sensor was attached 20 cm from the top ofeach pendulum. A digital signal processor computed each sensor’s position and orientation relative tothe transmitter. Signal sampling rate was 60 Hz per sensor.
The cross-shaped hand-held pendulums were constructed from wooden dowels. The stem of thependulums was 60 cm long. A crosspiece (22 cm in length) was attached to the stem of each pendulumat a 90� angle, 20 cm above its distal end. The crosspiece extended approximately 10 cm to each side ofthe stem. Cylindrical weights of different magnitudes were attached to the crosspieces so as to (a)maintain the alignment between the pendulum’s vmm and its longitudinal axis, or (b) break the coin-cidence between the two by shifting (rotating) vmm. Note that vmm orientation is a function of the ori-entation of both ve3 and vCM. These latter vectors were manipulated concurrently in the presentexperiment. Appendix A describes in detail the computations performed to estimate the orientationof ve3 and vCM.
The four experimental conditions illustrated in Fig. 4 were defined according to the configuration ofthe hand-held pendulums used to manipulate Dvmm. For Conditions 1A (2A), a 4� (10�) shift to theright pendulum’s vmm was introduced (Fig. 4, right panel) by asymmetrically distributing the weights
Condition 1A Condition 2A Condition 2B
ΔVmm= -4°
Condition 1B
ΔVmm= 0° ΔVmm= -10° ΔVmm= 0°
Fig. 4. Conditions of Experiment 1. Performance on the coordination task on Conditions 1B and 2B served as control forConditions 1A and 2A, respectively.

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1021
on the crosspiece. Specifically, for Condition 1A a 150 g cylindrical weight was attached to the cross-piece section in front of the hand and a 50 g cylindrical weight was attached to the crosspiece sectionbehind the hand, 8.5 cm from the pendulum’s stem. For Condition 2A, a 200 g weight was attached tothe crosspiece section in front of the hand (9.5 cm from the stem) and no weight was attached to thecrosspiece section behind the hand. For the left pendulum of Condition 1A (2A) cylindrical weights of100 g (200 g) were attached on each side of the crosspiece, 8.5 cm (9.5 cm) from the pendulums’ stem.Note that the masses were symmetrically distributed in this case (no vmm shift). Thus, as illustrated inFig. 4 (left panel), Dvmm of different magnitudes were introduced for Condition 1A (Dvmm = �4�:ve3 = 4.47�, vCM = 3.79�) and for Condition 2A (Dvmm = �10�, ve3 = 4.47�, vCM = 3.79�). The magnitudeof vmm shift was obtained by averaging the orientation of ve3 and vCM computed as detailed in theAppendix A.
The pendulums on the right and left hand in Conditions 1A and 2A only differed in vmm direction.However, the pendulums used in Condition 1A differed from the pendulums used in Condition 2A inequivalent length (LE: 1A = 36.28 cm; 2A = 40.61 cm), and hence, in their coupled eigenfrequencies(xc: 1A = 0.82 Hz; 2A = 0.79 Hz). This difference was a by-product of the Dvmm magnitude manipula-tion. Because xc may affect some of the dependent measures (e.g., SD/), two control conditions(Dvmm = 0) were designed (Fig. 4, right panel). For Condition 1B (2B), the right and left hand-held pen-dulums had the same configuration as the left hand-held pendulum in Condition 1A (2A).
2.1.3. ProcedureFor all trials participants sat in a custom-made, straight-back wooden chair with arm rests to sup-
port the forearms. The armrests were designed to restrict motions of the wrist joint while allowing thependulums to move without making contact with the chair. Participants were told to keep the twoforearms supported throughout the experimental trials with the hands extending approximately4 cm over the front of the armrest.
Once properly positioned, participants were handed a pair of pendulums and instructed to graspthem firmly so that the top of each pendulum’s handle was flush with the top of the respective hand.The experimenter used another pair of pendulums to demonstrate to the participants how to producesmooth and continuous oscillations of the two pendulums in the sagittal plane, using only radial andulnar deviations the wrist joint.
Participants had approximately 10 min of practice in order to allow familiarization with the task.Initially, participants were allowed to look at the pendulums. They were asked to note what it feelslike to restrict the motions to the sagittal plane and to contrast it with circular motions (which wereto be avoided). After that, they were blindfolded and told to experiment with moving the pendulumsin phase, at different frequencies and amplitudes, and to find the pattern of motion that was mostcomfortable and one that they would be able to maintain for a long period of time. As soon as partic-ipants indicated that they were confident with their performance, the sensors were attached to thependulum and a complete 3-min practice trial was performed. Participants only practiced withDvmm = 0�.
The experiment proper consisted of 12 trials of 3-min duration each (three trials for each of the 4different conditions illustrated in Fig. 4 and introduced in the Materials section). The order of trials wasfully randomized. On each trial, pendulums, with the sensors attached, were placed into the hands ofthe blindfolded participants. The hand-held pendulums were then positioned in the vertical directionwith the aid of vertically aligned supports and the orientation of the sensors was set to (0,0,0) withrespect to the three orthogonal coordinates of the transmitter: X (anterior-posterior), Y (side to side),and Z (up and down). Any changes in pendulum angular orientation during the coordination task wererelative to this static, space-fixed reference frame.
After the calibration procedure, the supports were removed and participants were blindfolded andtold to oscillate the pendulums. Data collection began at each trial as soon as participants indicated tothe experimenter that they reached a comfortable pattern of movement. A minimal 3-min rest wasprovided in between trials. Longer breaks were allowed at the discretion of participants. The experi-mental session lasted approximately 90 min. After all trials had been completed, participants were de-briefed and thanked for participation.

1022 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
2.1.4. Data reductionThe 6D-Research software generated data characterizing the three-dimensional positions of three
orthogonal vectors (x, y, z) centered in each sensor for each experimental trial. For all trials, the time-series measuring the angular excursions of the pendulums in the sagittal plane were obtained by com-puting, at each time step, the signed angle between the z-vectors (vertical vectors) of the sensors andthe basis vector (0,0,1) defining the z coordinate of the transmitter. The computed angular displace-ments were low-pass filtered with a cut off frequency of 10 Hz before further analyses. The dependentmeasures computed from the angular excursion time-series will be described in turn.
2.1.4.1. Relative angular orientation. The relative angular orientation (Da) of the hand-held pendulumswas computed for each sample of the displacement time-series by subtracting the angular position ofthe right sensor from the angular position of left sensor (Da = aleft � aright). Considering this conven-tion, a positive value indicates that the left pendulum is ahead of the right and a negative value indi-cates that the left pendulum is behind the right. Mean relative orientation or mean Da was computedfrom the resulting relative orientation time-series.
2.1.4.2. Mean / and SD/. Relative phase / is the difference in the phase angle of right and left pendu-lums. The phase angle, hi (�), of each pendulum was computed for each sample of the angular displace-ment time-series as:
hij ¼ arctan_aij
ðaij � �aiÞ
� �; ð3Þ
where ðaij � �aiÞ is the angular displacement of pendulum i at sample j minus the average displacementof that trial. _aij is the angular velocity at ith sample, normalized in terms of the mean angular fre-quency for the trial. In order to obtain the two velocity time series, the two angular displacementtime-series were differentiated. The relative-phase time-series was computed as the difference be-tween the left and right instantaneous phase angles (/ = hleft � hright). Mean / and SD/ were then com-puted from the resulting relative-phase time-series.
2.1.4.3. Attractor strength (k) and noise magnitude (Q). Cross recurrence quantification analysis (CRQA)was used in order to obtain independent indices of k and Q. This nonlinear analysis involves embed-ding a pair of time series in a multidimensional, time-delayed embedding space to ensure their suffi-cient unfolding (cf. Abarbanel, 1996). This procedure allows for the measurement of subtle time-correlated activity between the two signals by tracking when one or more values in one time seriescoincide with the value(s) in the second time series (i.e., when the values are cross-recurrent) inembedding space (Shockley, 2005; Webber & Zbilut, 1996a, 1996b).
Among the available measures from CRQA are the percentage of data points that are recurrent(%REC) and the longest trajectory of consecutive recurrent points (MAXLINE). MAXLINE is a measureof convergence between two phase-space trajectories and has been shown to be a reliable index of thespatial–temporal stability of interlimb coordination (i.e., stability of the dynamics of /) (Richardson,Schmidt, & Kay, 2007). Pointedly, the magnitude of MAXLINE is directly proportional to the magnitudeof k. %REC is a measure of the probability of finding a similarity of state between the two phase spacetrajectories and has been shown to be negatively related to the magnitude of noise in the system or Q(Richardson et al., 2007).
In order to quantify %REC and LMAX, the 180 s displacement time-series of the left and right handswere converted to z-scores to eliminate any difference in scale without influencing the distribution ofthe values within the time-series (Shockley, 2005). Next, each pair of time series (aright and aleft) wasembedded in a seven-dimensional space using its time-delayed copies as dimensions (see Shockley,2005). In the present experiment, the time-delay, selected individually for each movement time-ser-ies, was the time taken for the average mutual information function to reach its first minimum (Abar-banel, 1996).
The threshold radius used to determine which points in phase space trajectories constitute cross-recurrent points was set at 10% of the maximum distance separating points in the embedding space.

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1023
This definition of inclusion radius allowed recurrence to remain between 0.5% and 5% in all conditions,which is considered sufficient to make sense of a system’s dynamical structure while still allowing en-ough variation in other recurrence parameters, such as MAXLINE (Shockley, 2005).
2.2. Results and discussion
All subjects achieved the required 1:1 frequency locking in the different experimental conditions.Mean frequency ratios for Conditions 1A, 1B, 2A, 2B were 1 ± 0.002, 1 ± 0.004, 1 ± 0.0003, 1 ± 0.0002,respectively. As expected, self-selected coupled frequencies (xc) were inversely related to the pendu-lums’ LE, F(1,10) = 12.04, p = .06, g2 = .55, and were unaffected by Dvmm (F < 1).
For each dependent measure, the results were averaged across the three trials of each conditionand a 2 � 2 repeated measures ANOVA was conducted. The independent variables were the pendu-lums’ LE (pendulum type 1: LE = 36 cm; pendulum type 2: LE = 41 cm) and Dvmm (A: Dvmm < 0 and B:Dvmm = 0). As previously noted, differences in LE were introduced as a by-product of the Dvmm magni-tude manipulation (1A: Dvmm � �4�; 2A: Dvmm � �10�), which involved the attachment of weights ofdifferent magnitudes to the pendulums used in Conditions 1A and 2A (refer to Materials section fordetails). Hence, the effect of Dvmm magnitude on the study variables was assessed by the LE � Dvmm
interaction effect. To confirm whether significant interaction effects were driven by Dvmm magnitude,t-tests were conducted to verify if the difference observed between Conditions 2A and 2B in the meansof each dependent variable was greater than the difference in the same variable observed betweenConditions 1A and 1B.
Table 1 contains the means and standard errors for the measures of coordination performancedemonstrated under the different experimental conditions.
2.2.1. Did Dvmm affect Da?An ANOVA conducted with the dependent variable mean Da showed a significant LE � Dvmm inter-
action effect, F(1,10) = 14.83, p = .003, g2p = .60. Main effects of LE, F(1,10) = 14.4, p = .004, g2
p = .59, andDvmm, F(1,10) = 71.91, p = .001, g2
p = .88 were also observed. The significant interaction effect can beexplained by a greater difference in mean Da between Conditions 2A and 2B than between Conditions1A and 1B, t(10) = 3.85, p = .003. These results indicate the presence of an effect of Dvmm magnitude onDa. As can be seen in Table 1, the main effect of Dvmm is associated with the greater mean angle ob-served between the right and left pendulums in conditions in which Dvmm < 0 (Conditions 1A and 2A)when compared to those in which Dvmm = 0 (Conditions 1B and 2B). Contrast analysis did not showany significant difference in mean Da between the control Conditions 1B and 2B, t(10) = .39,p = .70). Hence, the main effect of LE was mainly driven by the relatively large changes in Da observedfor the longer pendulum when Dvmm = �10�.
2.2.2. Did Dvmm affect mean /?The repeated measures ANOVA demonstrated a significant LE � Dvmm interaction, F(1,10) = 8.11,
p = .02, g2p = .45, and a significant main effect of Dvmm, F(1,10) = 10.87, p = .01, g2
p = .52, on mean /.No main effect of pendulum LE was observed (F < 1). The main effect of Dvmm was consistent with
Table 1Means ± standard error of the dependent measures Da, /, SD/, MAXLINE, and %REC observed in Experiment 1.
Coordination measures Conditions
1A 1B 2A 2B
Da 1.88 ± 0.34 �0.53 ± 0.37 6.02 ± 0.28 �0.19 ± 0.36/ 2.49 ± 0.98 1.73 ± 1.38 3.73 ± 1.14 1.20 ± 1.32SD/ 8.22 ± 0.59 8.39 ± 0.53 8.22 ± 0.45 8.39 ± 0.49MAXLINE 1912.24 ± 300.4 1938.89 ± 328.9 1976.32 ± 261.3 1802.99 ± 291.7%REC 2.78 ± 0.20 2.99 ± 0.21 3.24 ± 0.19 3.03 ± 0.16
Note. Da, /, SD/ are reported in degrees.

1024 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
the experimental manipulation. Mean / obtained for Conditions 1A and 2A was greater than in Con-ditions 1B and 2B (see Table 1). Further inspection of the means in Table 1 suggests that the differencein mean / between Conditions 2A and 2B was significantly larger than the difference observed be-tween Conditions 1A and 1B. This observation was confirmed by a t-test, t(10) = 2.85, p = .02. Addition-ally, no significant difference in mean / between Conditions 1B and 2B, T(10) = 1.06, p = .31, wasobserved. Hence, the significant interaction indicates an effect of Dvmm magnitude on mean /. Moreprecisely, the larger the absolute value of Dvmm, the larger the shift in mean / from its canonical value(the canonical value meaning the value of mean / observed when Dvmm = 0).
2.2.3. Did Dvmm affect SD/, MAXLINE and %REC?As can be observed in Table 1, SD/ and MAXLINE were essentially constant in all conditions. This
was confirmed by the ANOVAs performed with these measures. There were no main effects of LE,Dvmm, or a LE � Dvmm interaction on either SD/ or MAXLINE, all Fs < 1. These results indicate thatthe shifts in attractor location (indexed by the changes in mean /) were not associated with changesin the stability of coordination.
Similarly, no main effect of Dvmm (F < 1) on mean %REC was observed. There was, however, a maineffect of LE, F(1,10) = 5.30, p = .04, g2
p = .35, and a significant LE � Dvmm interaction, F(1,10) = 5.14,p = .05, g2
p = .34 on %REC. Inspection of the means displayed in Table 1 indicates that the magnitudeof %REC is greater for the longer pendulum. The significant interaction suggests opposite effects ofDvmm for the heavier and lighter pendulums. However, contrast analysis does not indicate any signif-icant change in %REC when Conditions 1A and 2A were compared to their respective control condi-tions. The only difference observed in the pairwise comparisons was between Conditions 1A and2A. No explanation can be provided at this point for the latter observation. Nonetheless, the overallresults of the contrast analysis suggest that the contribution of stochastic processes was not modifiedby the asymmetry between the orientation of the right and left vmm.
Overall, the results of Experiment 1 suggest that Dvmm affects interlimb coordination. More pre-cisely, manipulations of Dvmm resulted in changes in the mean relative orientation of the hands andin their mean relative phase proportional to the magnitude Dvmm. No changes in k or Q accompaniedthe changes in pattern, which is consistent with the IFR hypothesis depicted in Fig. 3 (right panel).These results indicate that the angular displacements and velocities of the right and left pendulumswere equally constrained to stabilize / under the two Dvmm conditions. The only difference was thata different value of / was stabilized, which was specified by Dvmm magnitude. The invariance of theunderlying dynamics over the observed changes in pattern suggest that no additional processes of per-ceptual tuning were involved when Dvmm – 0 was compared to when Dvmm = 0.
Riley and Black (2003) reported a similar pattern of results when in-phase coordination patternswere performed after a unilateral change in the felt arm position was induced by prism adaptation.The authors suggested that the same spatial frames of reference altered by prism adaptation sup-ported the production of interlimb coordination patterns. However, a subsequent study did not sup-port the latter claim (Black & Riley, 2004). Specifically, interlimb coordination did not prove to bespecific to the direction of proprioceptive shift produced through optical displacement. This findingled the authors to conclude that the effects of prism adaptation worked as a general perturbation tothe coordination task. Hence, a thorough investigation of proprioceptive influences on coordinationdynamics should include not only manipulations of Dvmm magnitude but also of its sign.
3. Experiment 2: Effect of Dvmm sign on coordination dynamics
A strong prediction of both the detuning and IFR hypotheses concerns the sign of Dvmm. The changesin coordination pattern should occur in opposite directions if the sign of Dvmm goes from positive tonegative. However, as opposed to the detuning hypothesis, the IFR hypothesis expects no change in theunderlying dynamics (indexed by changes in SD/, MAXLINE or %REC) associated with the changes inpattern. A third hypothesis is that the effects of Dvmm are simply a result of a general perturbation tothe system (akin to the effects of proprioceptive shifts induced by prism adaptation). In that case, no

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1025
directional effects are expected. In addition, more variability would be expected as a result of a bilat-eral shift, even if Dvmm = 0. Experiment 2 was designed to distinguish these three hypotheses.
3.1. Method
3.1.1. ParticipantsTwelve undergraduate students from the University of Connecticut participated in the present
study in partial fulfillment of course requirements. Nine participants classified themselves as right-handed, two as left-handed and one as ambidextrous. None of them have had prior experience withthe task. Information regarding the experimental procedure and its purposes was provided as inExperiment 1.
3.1.2. Materials and procedureThe materials, procedure and experimental task were the same as in Experiment 1, except that only
pendulum type 2 (LE = 41 cm) was used. The design included four experimental conditions illustratedin Fig. 5. Conditions A (Dvmm = 10�) and B (Dvmm = 0�, control condition) in the present experimentwere identical to Conditions 2A and 2B in Experiment 1.
For Condition C, vmm of both pendulums were shifted forward by the same amount, maintainingDvmm = 0. This manipulation was accomplished by attaching a 400 g weight to the crosspiece of theright and left pendulum extending in front of the hand. No mass was added to the crosspiece extend-ing behind the hand. Finally, for Condition D, cylindrical weights of 200 g were attached on each sideof the crosspiece of the left hand pendulum. On the right pendulum, a 400 g weight was attached tothe crosspiece extending behind the hand and no weight was attached to the crosspiece extending infront of the hand. As illustrated in Fig. 5, Dvmm in Conditions A and D differed only in sign and not mag-nitude. No DLE (and consequently no Dx) were introduced between the right and left hands. Addition-ally, LE and xc magnitudes were the same for the four conditions.
3.1.3. Data reductionThe same procedures for data reduction described for Experiment 1 were used in the present
experiment.
3.2. Results and discussion
All subjects achieved the required 1:1 frequency locking in the different experimental conditions.Mean frequency ratios for Conditions A, B, C, and D were 1 ± 0.0007, 1 ± 0.0004, 1 ± 0.002, 1 ± 0.001,respectively. Self-selected frequency was unaffected by Dvmm (F < 1).
For each dependent measure, the results were averaged across the three trials of each condition.The overall design allowed for a single factor ANOVA to evaluate the effect of Dvmm on the perfor-mance of the coordination task. The means and standard errors for the dependent measures a, /,SD/, MAXLINE and %REC are summarized in Table 2.
Fig. 5. Conditions of Experiment 2. Performance on Condition B served as control.

Table 2Means ± standard error of the dependent measures Da, /, SD/, MAXLINE, and %REC observed in Experiment 2.
Coordination measures Conditions
A B C D
Da 6.18 ± 0.37 �0.66 ± 0.22 �0.53 ± 0.23 �8.00 ± 0.17/ 0.39 ± 1.35 �0.89 ± 1.47 �1.16 ± 0.98 �3.22 ± 1.15SD/ 9.70 ± 0.97 9.36 ± 0.92 9.55 ± 1.03 10.39 ± 0.98MAXLINE 1924.23 ± 460.84 2428.94 ± 640.37 2462.61 ± 448.56 2208.92 ± 695.59%REC 2.73 ± 0.35 2.62 ± 0.36 2.69 ± 0.38 2.62 ± 0.40
Note. Da, /, SD/ are reported in degrees.
1026 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
3.2.1. Did Dvmm affect Da?The repeated measures ANOVA demonstrated a significant effect of Dvmm, F(3,30) = 83.61, p < .001,
g2p = .89. Contrast analyses further demonstrated that (a) mean Da for Conditions A and D differed sig-
nificantly from mean Da for the control condition (Condition B), p < .001; and (b) mean Da for Condi-tion C was not significantly different from mean a for the control condition, p = .78.
Inspection of Table 2 reveals an effect of Dvmm sign on the spatial relation between limbs: while thedifference in mean Da between Conditions A and B have a positive sign, the difference in mean Dabetween Conditions D and B have a negative sign.
3.2.2. Did Dvmm affect /?The effects of Dvmm on / follow the same pattern of the effects described for Da. First, a significant
main effect of Dvmm was observed, F(3,30) = 7.91, p < .001, g2p = .42. Second, contrast analyses demon-
strated that (a) mean / for Conditions A and D differed significantly from mean / for the control con-dition (Condition B), p < .001; and (b) mean / for Condition C was not significantly different frommean / for the control condition, p = .81. Again, inspection of the means in Table 2 reveals that thespatiotemporal relations between the limbs are constrained not only by the magnitude but also bythe sign of Dvmm.
3.2.3. Did Dvmm affect SD/, MAXLINE and %REC?No significant main effect of Dvmm was observed on SD/, F(3,30) = 2.218, p = .11, g2
p = .18; on %REC,F < 1; or MAXLINE, F(3,30) = 1.22, p = .32, g2
p = .11.The results of Experiments 1 and 2 indicate that Dvmm does not play the role of an imperfection
parameter during interlimb coordination. Specifically, differences between the orientation of vmm rightand vmm left did not affect the main dynamical regime constraining the observed interlimb patterns.However, it does seem to provide a proprioceptive frame of reference for their organization. Thishypothesis, termed the IFR hypothesis, was supported by the observed scalar shift in the in-phaseattractor when Dvmm – 0, without changes in its specific shape. The implication is that haptic infor-mation about the relative orientation of the limbs (i.e., the relative orientation of vmm right and vmm
left) supports the organization of the synergy of 1:1 rhythmic coordination. The final question to beaddressed is which specific mass moment vector, ve3 or vCM, underlies the changes in coordination per-formance observed in Experiments 1 and 2.
4. Experiment 3: Which vmm vector shapes coordination dynamics?
As noted in the introduction, both ve3 and vCM typically coincide with the longitudinal axes of bodysegments, and, hence, both may be informative of their orientation in space. The manipulations of thependulums performed in Experiments 1 and 2, however, resulted in equivalent shifts of vCM and ve3,such that their individual effects could not be assessed. Fortunately, research by van de Langenberget al. (2007, 2008), has identified a methodology by which these two vectors can be manipulated inde-pendently so that their specific roles in shaping coordination dynamics can be evaluated. In applyingthe methodology, these authors showed that when individuals are asked to point to a target or to align

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1027
the position of their arms (without the benefit of vision) they do so guided by the limbs’ vCM and not bythe limb’s ve3. The present experiment exploited the methodology introduced by van de Langenberget al. (2007, 2008) to determine which vector was responsible for the effects observed in Experiments1 and 2.
4.1. Method
4.1.1. ParticipantsThirteen undergraduate students from the University of Connecticut participated in Experiment 3
in partial fulfillment of course requirements. Twelve participants reported a right hand preference andone participant reported a left hand preference. None of them had prior experience with the experi-mental task.
4.1.2. MaterialsIn order to employ the methodology introduced by van de Langenberg et al. (2007, 2008) in the
context of coordination dynamics, pendulums with two crossbars were required. Key to this method-ology is the fact that vCM is only influenced by the distribution of mass about the pendulum’s shaftwhile ve3 additionally depends on how the masses are distributed with respect to an axis of rotation(see Fig. 6, top panel). vCM is shifted out of the geometrical axis whenever the masses to the left and tothe right have different magnitudes or are positioned at different distances from the longitudinal axisof the pendulum (top panel, P2 and P3). The direction of ve3 is determined by the magnitude m of themasses to the right and to the left and their distances d to the center of rotation (e.g., an axis throughthe top of the pendulum). ve3 will be shifted out of the longitudinal geometrical axis if the products of
Fig. 6. Conditions of Experiment 3. Top panel: Hand-held pendulums with two cross bars allows independent manipulations ofthe hand’s vmm. vCM is the vector from the center of rotation (defined for the sake of illustration at the top of the pendulum) tothe pendulum’s center of mass. ve3 is the vector through the center of rotation about which the pendulum’s rotational inertia isminimal. Bottom panel: Experimental conditions of Experiment 3 configured with the two-bar pendulums. Performance onConditions 1B and 2B served as control for Conditions 1A and 2A, respectively. Dotted arrows refer to ve3 orientation; solidarrow refer to vCM orientation. Dotted line illustrates the direction of the gravity plumb line.

1028 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
inertia (or centrifugal moments) related to the rotational motion of the rightward mass distributionare not balanced (cancelled out) by the products of inertia related to the rotational motion of the left-ward mass distribution (as in panels P2 and P4 of Fig. 6 relative to panels P1 and P3). In respect to P3,for example, one can intuit the balance of centrifugal moments by noting that, relative to the leftwardattached mass, the rightward attached mass’s larger distance from the axis of rotation (dright = 3 � dleft)is countered by its smaller magnitude (mright = 1/3 �mleft).
The pendulums used in Experiment 3 were constructed following the principles described above inorder to design experimental conditions in which only differences in the orientation of one of the massmoment vectors (vmm) were introduced between hands (see Fig. 6, bottom panel). The hand-held pen-dulums were similar to the ones used in the previous experiments, except two crosspieces were at-tached to the stem. The upper and lower crosspieces were attached 60 cm and 20 cm, respectively,from the proximal end of the pendulum.
For Condition 1A, a 6� forward shift of the right hand’s ve3 (ve3 right) with respect to the left hand’sve3 (ve3 left) was introduced. With respect to the right pendulum, a 200 g cylindrical weight was posi-tioned on the upper crosspiece, 8.5 cm behind the hand, and a 200 g cylindrical weight was positionedon the lower crosspiece, 8.5 cm in front of the hand. With respect to the left pendulum, two cylindricalweights of 100 g were attached to the upper crosspiece and two cylindrical weights were attached tothe lower crosspiece, one 8.5 cm in front of the hand and one 8.5 cm behind the hand. Note that for theright pendulum, the torque produced by the mass on the lower crosspiece is higher than the torqueproduced by the mass on the upper crosspiece. However, the mass distribution with respect to the lon-gitudinal axis is symmetric on both pendulums. Hence, for Condition 1A, Dve3 = �7�, while DvCM = 0�(see Fig. 6, bottom panel).
For Condition 2A, a 6� forward shift of the right hand’s vCM (vCM right) with respect to the left hand’svCM (vCM left) was introduced also illustrated in Fig. 6 (bottom panel). Considering the right pendulum, a300 g cylindrical weight was positioned on the upper crosspiece, 8.5 cm in front of the hand, and a 100 gcylindrical weight was positioned on the lower crosspiece, 8.5 cm behind the hand. Note that the masson the upper bar of the right pendulum is three times larger than the mass on the lower bar, but it is alsothree times closer to the axis of rotation. Consequently, while vCM was shifted, ve3 was not. Consideringthe left pendulum, two cylindrical weights of 150 g (50 g) were attached to the upper (lower) crosspiece,one 8.5 cm in front of the hand and one 8.5 cm behind the hand. Since the masses were symmetricallydistributed both with respect to the longitudinal axis and with respect to the center of rotation, no shiftin vCM or ve3 was introduced. Hence, for Condition 2A, DvCM = �6�, while Dve3 = 0�.
As in Experiments 1 and 2, the right and left pendulums in Conditions 1A and 2A were configuredsuch that they only differed in the direction of vCM and ve3, respectively. Note also that the pendulumsused in Condition 1A differed from the pendulums used in Condition 2A in equivalent length (LE:1A = 39 cm; 2A = 47 cm), and hence, in their coupled eigenfrequencies (xc: 1A = 0.8 Hz;2A = 0.7 Hz). This difference was a by-product of the mass distribution used to obtain the independentshifts in vCM and ve3 of similar magnitudes on Conditions 1A and 2A, respectively. Because xc may af-fect some of the dependent measures (e.g., SD/), two control conditions (DvCM = 0� and Dve3 = 0�) weredesigned (Fig. 6, bottom panel). For control Condition 1B (2B), the right and left hand-held pendulumshad the same configuration as the left hand-held pendulum on experimental Condition 1A (2A).
The motions of the pendulums were recorded at a 60 Hz sampling rate in all conditions using thesame equipment described for Experiment 1.
4.1.3. ProcedureThe procedure was the same as in Experiments 1 and 2. Participants performed in-phase coordina-
tion of a pair of hand-held pendulums under the four different experimental conditions detailed in theMaterials section. There were 12 two-minute trials (three trials per condition). The order of trials wasfully randomized.
4.1.4. Data reductionIn order to characterize the effects of DvCM and Dve3 on coordination performance, the same mea-
sures Da, /, SD/, MAXLINE and %REC were obtained following the steps detailed in the Data reductionsection of Experiment 1.

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1029
4.2. Results and discussion
All subjects achieved the required 1:1 frequency locking in the different experimental conditions.Mean frequency ratios for Conditions 1A, 1B, 2A, 2B were 1 ± 0.002, 1 ± 0.004, 1 ± 0.0003, 1 ± 0.0002,respectively. A LE � Hand � Dvmm repeated measures ANOVA was performed to evaluate the effect ofthe study variables on mean period. As expected, self-selected coupled frequencies (xc) were inverselyrelated to pendulum LE, F(1,12) = 18.28, p = .001, g2
p = .60, and were unaffected by hand (F < 1).There was a small, but significant increase in frequency when Dvmm – 0 (regardless of which vector
was shifted), F(1,12) = 6.45, p = .03, g2p = .35, an effect that was not observed in Experiments 1 and 2.
Importantly, the frequencies were not different to the first decimal place (Dvmm = 0�: 0.96 ± 0.02 Hz;Dvmm – 0�: 0.99 ± 0.03 Hz). This small difference suggests that the change in frequency as a functionof Dvmm observed in the present experiment (though significant) was most likely not behaviorallymeaningful.
Effects of Dve3 and DvCM on the measures characterizing coordination performance were evaluatedby t-tests. The descriptive statistics are reported in Table 3.
4.2.1. Did Dve3 affect coordination performance?When Conditions 1A and 1B were compared, no differences in Da, /, SD/, MAXLINE or %REC were
observed (all ts < 1, all ps < .39). Hence, coordination performance was not affected by Dve3 – 0� whenDvCM = 0�.
4.2.2. Did DvCM affect coordination performance?There were significant differences in Da, T(12) = �7.71, p < .001, and in mean /, t(12) = 3.68,
p = .003, when Conditions 2A and 2B were compared. As documented in Table 3, mean / and meanDa were greater when for Condition 2B (DvCM – 0). Hence, both attractor location and the patternof interlimb spacing changed as a function of DvCM but not as a function of Dve3. No significant differ-ences in MAXLINE, t(12) = 1.09, p = .29, or %REC, t(12) = 0.21, p = .84, were observed.
The results of Experiment 3 indicate that the present coordination task was primarily supported byDvCM. This result might be surprising if we consider the physical meanings of both ve3 and vCM. Whileve3 defines an axis of dynamic balance of a limb or object, vCM defines an axis of static balance. The roleof vCM in the perception of limb orientation must, therefore, be grounded in the patterning of statictorques involved in balancing the limb in a gravitational field. In contrast, the perceptual role of ve3
must be grounded in its consequences to the patterning of dynamic torques generated to accelerate(or decelerate) limb rotations in space. Given the dynamic nature of the pendulum task, we expectedto see a contribution of ve3. Pointedly, participants were asked to coordinate the motions of theirhands in parasagittal planes. In this context, ve3 would be expected to specify the direction of muscularforces required to counteract reactive forces that could potentially accelerate the hands out of theparasagittal planes. The fact that participants were successful in complying with the task’s require-ment suggests that the movement system was sensitive to ve3. However, the absence of Dve3 effectson coordination performance indicates that ve3 exclusively constrained the local patterning of forces
Table 3Means ± standard error of the dependent measures Da, /, SD/, MAXLINE, and %REC observed in Experiment 4.
Pendulum properties Conditions
1A (ve3 < 0) 1B (ve3 = 0) 2A (vCM < 0) 2B (vCM = 0)
Da �1.66 ± 0.17 �0.75 ± 0.20 2.34 ± 0.21 �0.91 ± 0.27/ �0.55 ± 1.45 �1.09 ± 1.48 3.10 ± 1.10 1.51 ± 1.06SD/ 9.24 ± 0.76 9.65 ± 0.73 10.53 ± 0.74 10.36 ± 0.66MAXLINE 1668.33 ± 184.21 1539.23 ± 220.32 953.79 ± 172.11 1081.59 ± 122.81%REC 2.67 ± 0.26 2.56 ± 0.24 2.08 ± 0.22 2.11 ± 0.20
Note. Da, /, SD/ are reported in degrees.

1030 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
and was not exploited to synchronize the limbs. What might explain the dominance of DvCM over Dve3
in shaping coordination performance?The answer to this question may lie in the context of forces supporting task performance. The coor-
dination task performed in the present study, though dynamic, is organized so as to take full advan-tage of the gravitational potential (Kugler & Turvey, 1987). For example, there is evidence that duringthe performance of such task the neuromuscular machinery simply complements the passive dynam-ics of the hand + pendulum system in the gravitational field in order to sustain the oscillations(Goodman, Riley, Mitra, & Turvey, 2000). An expression of this strategy is the fact that self-selectedfrequency of oscillation roughly coincides with the natural frequency of the pendulums—the fre-quency that would be observed if the pendulums were free to oscillate in the gravitational field(Kugler & Turvey, 1987). The implication is that even though inertial (or dynamic) torques certainlycontribute to the total joint torque supporting the pendulum task, the gravitational (or static) compo-nent, to which DvCM is linked, is likely to be dominant. In support of that, Hollerbach and Flash (1982)showed that during the performance of arm movements, the effect of gravity completely dominatesany other term contributing to joint torque if the movement is sufficiently slow. Hence, the (likely)dominance of the gravitational component in our experimental task may explain the dominant effectof DvCM in guiding perception and regulation of relative limb motions in the present experiment. It ispossible that during the performance of high-speed tasks, the role of ve3 in orienting the body seg-ments with respect to each other and the environment might be revealed. This could be the topicof future studies.
5. General discussion
In the present paper, three experiments were presented in which bimanual coordination perfor-mance was evaluated under selective manipulations of mechanical parameters (generally referredto as Dvmm) potentially informative of the relative orientation of the limbs. The fact that Dvmm affectedcoordination performance reinforces the role of the haptic perceptual system in the organization andmaintenance of movement patterns appropriate for fulfilling task requirements. This result is in linewith findings of previous research that showed changes in spatial and temporal parameters of coor-dination performance as a function of pathology or mechanical perturbations that alter the afferentinflow from mechanoreceptors (e.g., Steyvers et al., 2001; Verschueren et al., 1999). However, becausethe latter studies involved only general perturbations to mechanoreceptor inflow, the specific natureof the information by action coupling underlying coordination could not be examined directly. Thepresent paradigm, in contrast, provided a window into the interface between coordination dynamicsand the haptic information that modulates its deployment, without interfering with the general func-tioning of the proprioceptive machinery. Consequently, this paradigm allowed the identification of ahaptic informational parameter that supports interlimb synchronization (specifically, vCM), and, con-sequently, allowed the examination of its specific role in shaping coordination dynamics.
The general hypothesis (termed the Inertial Frame of Reference hypothesis or IFR) was that monitor-ing and regulation of the spatial relations between hands during bimanual coordination is based on aninertial frame of reference defined by Dvmm. We drew an inventory of expected findings (captured inFig. 3) that would provide support to the IFR hypothesis in detriment to the detuning hypothesis (thestrongest competing hypothesis in our view). In all experiments, the predicted changes in coordina-tion performance were observed. First, Dvmm sign and magnitude resulted in proportional shifts inattractor location. Second, the shifts in / were associated with corresponding changes in the relativeorientation of the limbs. Third, the shifts in attractor location were not accompanied by changes in theunderlying dynamics of coordination, that is, no changes in SD/, stability k (indexed by LMAX) or noiseQ (indexed by %REC) were observed as a function of Dvmm, in particular DvCM. Therefore, participants’performance suggests that a spatial, proprioceptive frame of reference defined by the inertial axes(and not the geometrical axes) of the body segments is exploited in order to synchronize theirmotions.
How might vCM orientation be revealed to the haptic perceptual system in such tasks? As noted byvan de Langenberg et al. (2007, 2008) the tissue deformation consequences of changes in gravitational
P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1031
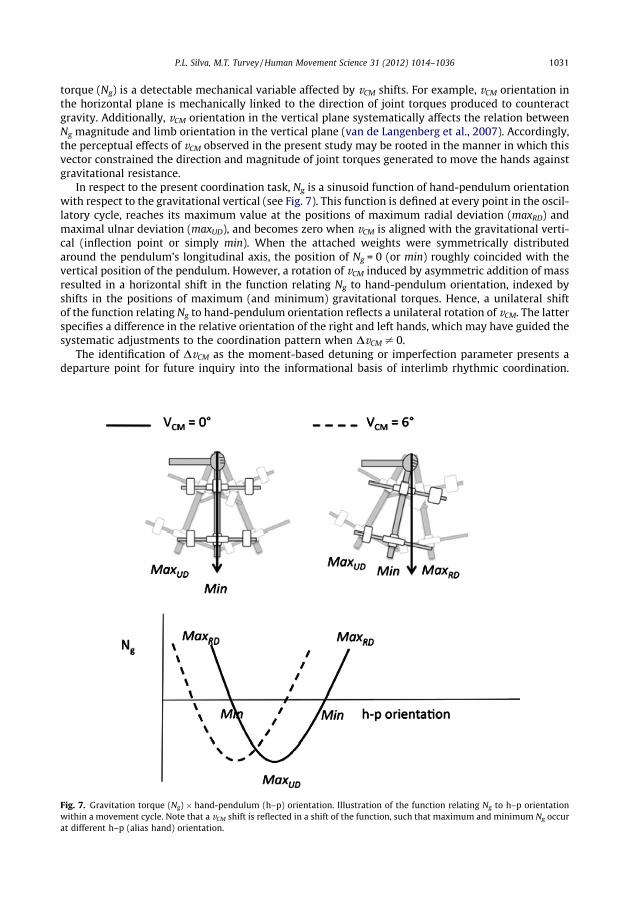
torque (Ng) is a detectable mechanical variable affected by vCM shifts. For example, vCM orientation inthe horizontal plane is mechanically linked to the direction of joint torques produced to counteractgravity. Additionally, vCM orientation in the vertical plane systematically affects the relation betweenNg magnitude and limb orientation in the vertical plane (van de Langenberg et al., 2007). Accordingly,the perceptual effects of vCM observed in the present study may be rooted in the manner in which thisvector constrained the direction and magnitude of joint torques generated to move the hands againstgravitational resistance.
In respect to the present coordination task, Ng is a sinusoid function of hand-pendulum orientationwith respect to the gravitational vertical (see Fig. 7). This function is defined at every point in the oscil-latory cycle, reaches its maximum value at the positions of maximum radial deviation (maxRD) andmaximal ulnar deviation (maxUD), and becomes zero when vCM is aligned with the gravitational verti-cal (inflection point or simply min). When the attached weights were symmetrically distributedaround the pendulum’s longitudinal axis, the position of Ng = 0 (or min) roughly coincided with thevertical position of the pendulum. However, a rotation of vCM induced by asymmetric addition of massresulted in a horizontal shift in the function relating Ng to hand-pendulum orientation, indexed byshifts in the positions of maximum (and minimum) gravitational torques. Hence, a unilateral shiftof the function relating Ng to hand-pendulum orientation reflects a unilateral rotation of vCM. The latterspecifies a difference in the relative orientation of the right and left hands, which may have guided thesystematic adjustments to the coordination pattern when DvCM – 0.
The identification of DvCM as the moment-based detuning or imperfection parameter presents adeparture point for future inquiry into the informational basis of interlimb rhythmic coordination.
Fig. 7. Gravitation torque (Ng) � hand-pendulum (h–p) orientation. Illustration of the function relating Ng to h–p orientationwithin a movement cycle. Note that a vCM shift is reflected in a shift of the function, such that maximum and minimum Ng occurat different h–p (alias hand) orientation.

1032 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
Our argument above was that vCM is realized in the tissue deformation consequences of the pattern ofstatic torques supporting limb oscillations. A prediction stemming from this argument is that informa-tion about the orientation of body segments would be primarily picked-up (or detected) at positions inthe cycle specified by easily differentiable patterns of tissue deformation about the wrist joint. In thecontext of the pendulum task, these would be the position where Ng = 0 (Min in Fig. 7) or the positionsat which Ng is maximal (MaxRD and MaxUD in Fig. 7). Min is expected to realize the most symmetricdeformation field about the wrist joint since forces are balanced at this point. In contrast, the reversalpoints (MaxRD and MaxUD) are expected to realize the least symmetric deformation fields (i.e., tissuesare mostly stretched on one side of the joint and mostly shortened on the other). Future investigationscan assess whether information for coordination is in fact primarily picked up at Min, MaxRD and Max-UD by measuring interlimb coupling strength at these positions and comparing it to interlimb couplingstrength at arbitrary positions in the oscillatory cycle. Of the three identified points, Min is expected tobe the least effective position for information pick up, particularly for higher frequencies, since it oc-curs at the high-speed portion of the cycle. In support of this hypothesis, Black et al. (2007) showedthat when frequency of oscillation increases, the strength of interlimb coupling at the middle portionof the cycle decreases more significantly than the strength of interlimb coupling at the reversal points.
Empirical demonstrations of the pattern of interlimb described above would have implicationsfor an information-based account of the phenomena predicted by the HKB dynamic. First, it wouldbe expected that a bimanual coordination pattern that requires arriving at MaxRD, MaxUD, and orMin at the same time would be easier to achieve. This might explain the relatively greater stabilityof in-phase coordination (hands reach all three positions at the same time) than anti-phase coor-dination (hands reach only Min at the same time). The more pronounced change in stability ofanti-phase coordination with increases in movement frequency might in turn be explained bythe detrimental effects of speed on information pick up predicted to be more significant at Min.Finally, the requirement for special perceptual tuning to achieve interlimb phase relations otherthan in-phase and anti-phase also follow naturally from this account given that none of the threeidentified positions is reached by both limbs or limb segments at the same time. In sum, the iden-tification of DvCM as the moment-based parameter specifying the frame of reference for interlimbcoupling points to the nature and challenges of detecting haptic information about the patterningand stability of interlimb coordination.
The fact that the magnitude of Q did not vary significantly across conditions suggests that infor-mation pick up did not differ between Dvmm – 0 and Dvmm = 0. This finding is consistent with theview of on-line haptic proprioception (Pagano & Turvey, 1995). The online perception of the rela-tive orientation of body segments is in principle possible because (a) vmm typically coincides withthe geometrical axis of body segments and, hence, constitutes information about its spatial orien-tation; and (b) vmm is a mechanical parameter that indexes invariant properties of tissue deforma-tion revealed during motion. Thus, one may know about relative orientation of the limbs onlinethrough detection of global invariants of the spatially and temporally distributed deformation pat-tern-specific to Dvmm. If this argument holds, then it can be assumed that the relative orientationof the hands was monitored in every condition of Experiments 1–3 in the same way: through on-line detection of the relative orientation of vmm right and vmm left. The implication is that, in allexperimental conditions, the trajectories of the two hands were organized so as to arrive at MaxRD,Min, and MaxUD at the same time, regardless of the actual geometrical configuration of the hands.Therefore, no additional or different kinds of haptic perception would be involved when Dvmm – 0,and hence, no changes in Q would be expected. This would explain the invariance in the dynamicalorganization over changes in pattern.
The effects of Dvmm on rhythmic coordination seem to differ from the effects of shifts in perceptual-motor frames of reference induced by prism adaptation. Under certain conditions, adaptation to opti-cally displacing prisms produces a change in the felt limb position called proprioceptive shift (PS).Black and Riley (2004) showed that the effects of prism-induced PS on coordination performanceare not direction specific and resulted in increases in SD/. This contrasts sharply with the effects ofDvmm observed in the present study. While PS seem to be generally disruptive to coordination, Dvmm
seems to shape the relations between limbs giving rise to the observed coordination pattern. Thesecontrasting findings indicate that the proprioceptive frame of reference affected by prism adaptation

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1033
plays a different role in interlimb coordination than the proprioceptive frame of reference defined byDvmm.
Despite the differences between the specific effects of Dvmm and PS described above, both studiespoint to a common conclusion: manipulations of proprioceptive parameters with a purely spatialcharacter may affect interlimb rhythmic coordination. This is not a trivial finding because the lattertask has a strong temporal component. Pointedly, perfect in-phase coordination can, in principle, beattained under different spatial configurations between limbs. However, the observed shifts in / whenDvmm – 0 suggest that the phase relations observed during interlimb synchronization are (at least inpart) a product of perceptual regulation of interlimb spacing.
What might be the implication of a vmm-based model of proprioception to the perceptual regulationof interlimb coordination? Of relevance to this question is the fact that even though vmm has a spatialcharacter, it is determined by the way in which forces act on the limb. Specifically, the torques re-quired to accelerate the limb about a fixed point by a certain amount are related to ve3, and the torquesrequired to sustain or to slowly move the limb against gravity are related to vCM. The implication isthat the proprioceptive frame of reference exploited to meet the kinematic (spatial) requirementsof interlimb rhythmic coordination can potentially also be exploited to meet its dynamical (force)requirements. This stands in sharp contrast with models of perceptually guided action that proposeseparate representations for sensory and motor parameters, with coordinate transformations to rec-oncile the differences between the two (cf. Flanders, Helms-Tillery, & Soechting, 1992). To reiterate,if spatial frames of references tied to the mass distribution of body segments were exploited in theproprioceptive control of action, the need for such explicit sensory-motor transformations would beobviated.
The fact that no change in attractor strength was observed in the present study supports the argu-ment that DvCM not only provided the spatial constraints required to synchronize the motions of thehands (which determines attractor location), but also thoroughly co-specified the magnitude anddirection of forces required to achieve and maintain the intended coordination pattern (which deter-mine attractor stability). Notably, spatial constraints may also be provided by the visual system(Amazeen, DaSilva, & Amazeen, 2008). It is not likely, however, that spatial information about limbrelations available visually could specify all aspects of limb dynamics. This is evidenced by the factthat movement coordination is profoundly affected when the contribution of the haptic system areremoved due to pathology even when vision is available (Cole, 1995). Perhaps, what makes the hapticproprioceptive system so special in the regulation of action is the fact that it more intimately connectsthe spatial and dynamical requirements of action.
Acknowledgment
This research was supported by NSF Grant SBR 04-23036.
Appendix A. Computation of ve3 and vCM orientations
The orientation of ve3 and vCM of the right and left wrist-pendulum systems were computed foreach experimental condition. For these computations, a coordinate system was defined, as illustratedin Fig. A1. In this appendix, a description is provided of the steps taken to compute the angle (also indi-cated in the figure) that indicates vmm orientation with respect to the longitudinal axis of thependulum.
The computation of ve3 direction was performed as follows. First, the inertia tensor of the hand (Ih)was estimated. Following Pagano and Turvey (1995), the closed hand was approximated as a solid,homogenous sphere, with its mass estimated at 443 g and its diameter estimated at 8 cm (Kugler &Turvey, 1987). Ih reflects the inertia of the hand about its center of mass (CM). The parallel axis the-orem was employed to obtain the inertia tensor of the hand with respect to the wrist (Ih,w). Specifi-cally, Ih was translated to the wrist along a single axis, the y-axis in the figure above. Subsequently,the inertia tensor of the stem (s), crosspiece (c) and attached loads (L1 and L2) were calculated alsowith respect to the wrist. The computations followed the same general steps described for the

Fig. A1. Frame of reference for computation of the mass moment vectors of hand-pendulum systems.
1034 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
computation of Ih,w. First, the tensor of each component of the pendulum (Is, Ic, IL1, IL2) was computedrelative to their respective CMs. Second, the parallel axis theorem was used to translate the computedtensors to the wrist along the y-axis through the forearm to obtain the different tensors with respectto the wrist (Is,w; Ic,w; IL1,w; IL2,w). The inertia tensor of the wrist-pendulum system (Itot) was obtainedby summing all the computed tensors:
Itot ¼ Ih;w þ Is;w þ Ic;w þ IL1;w þ IL2;w: ð4Þ
Note that Itot contains the off-diagonal terms of L1 and L2 that do not cancel each other out. In thatcase, e3 will not coincide with the z-axis through the pendulum, as occurs in the figure above. By diag-onalizing Itot, however, the set of eigenvectors (vei) can be obtained. The angle (or orientation) of ve3
with respect to the z-axis through the pendulum is:
ve3 ¼ arctanðYe3=Ze3Þ; ð5Þ
where Ye3 is the y-coordinate of ve3 and Ze3 is the z-coordinate of ve3.In order to compute vCM, the center of mass of each segment was first obtained (CMh, CMs, CMc,
CML1, CML2). Subsequently, the x, y, and z coordinates of the wrist-pendulum system’s CM was com-puted as:
CMtot ¼CMh �mh þ CMS �ms þ CMc �mc þ CML1 �mL1 þ CML2 �mL2
mh þms þmc þmL1 þmL2: ð6Þ
The angle of CMtot with the pendulum’s longitudinal axis is:
vCM ¼ arctanðYCM=ZCMÞ; ð7Þ
where YCM is the y-coordinate of vCM and ZCM is the z-coordinate of vCM.For Experiments 1 and 2, ve3 and vCM were shifted concurrently. In that case, their direction was
summarized in the variable vmm:
vmm ¼ve3 þ vCM
2: ð8Þ
In Experiment 3, the pendulums were designed with two bars and four masses such that ve3 andvCM would be independently rotated. The calculations performed for the latter experiment followedthe same steps described above, expect that Itot and CMtot would include the contributions of the sec-ond bar and additional masses.

P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036 1035
References
Abarbanel, H. D. I. (1996). Analysis of observed chaotic data. New York: Springer.Amazeen, P., Amazeen, E. L., & Turvey, M. T. (1998). Dynamics of human inter-segmental coordination: Theory and research. In
C. Collyer & D. Rosen-baum (Eds.), Sequencing and timing of movement: Neural, computational, and psychological perspectives.Cambridge, MA: MIT Press.
Amazeen, E. L., DaSilva, F., & Amazeen, P. G. (2008). Visual–spatial and anatomical constraints interact in a bimanualcoordination task with transformed visual feedback. Experimental Brain Research, 191, 13–24.
Bernstein, N. (1967). The coordination and regulation of movements. Oxford: Pergamon Press.Bernstein, N. (1996). On dexterity and its developments. In M. Latash & M. T. Turvey (Eds.), Dexterity and its development
(pp. 2–244). Mahwah, NJ: Erlbaum.Black, D. P., & Riley, M. A. (2004). Prism aftereffects disrupt interlimb rhythmic coordination. Journal of Motor Behavior, 36(2),
131–136.Black, D. P., Riley, M. A., & McCord, C. K. (2007). Synergies in intra- and interpersonal interlimb rhythmic coordination. Motor
Control, 11, 348–373.Cohen, L. A. (1958). Analysis of position sense in human shoulder. Journal of Neurophysiology, 21, 550–562.Cole, J. (1995). Pride and a daily marathon. Cambridge, MA: MIT Press.Collins, D., Sternad, D., & Turvey, M. T. (1996). An experimental note on designing frequency competition in intersegmental
coordination dynamics. Journal of Motor Behavior, 28, 299–304.Cordo, P. J., Flores-Vieira, C., Verschueren, S. M. P., Inglis, J. T., & Gurfinkel, V. (2002). Position sensitivity of human muscle
spindles: Single afferent and population representations. Journal of Neurophysiology, 87, 1186–1195.Flanders, M., Helms-Tillery, S. I., & Soechting, J. F. (1992). Early stages in a sensorimotor transformation. Behavioral and Brain
Sciences, 15, 309–362.Fuchs, A., Jirsa, V. K., Haken, H., & Kelso, J. A. S. (1995). Extending the HKB model of coordinated movement to oscillators with
different eigenfrequencies. Biological Cybernetics, 74(1), 1995.Gibson, J. J. (1966). The senses considered as perceptual systems. Boston: Houghton Mifflin.Goodman, L., Riley, M., Mitra, S., & Turvey, M. T. (2000). Advantages of rhythmic movements at resonance: Minimal active
degrees of freedom, minimal noise, and maximal predictability. Journal of Motor Behavior, 32, 3–8.Haken, H., Kelso, J. A. S., & Bunz, H. (1985). A theoretical model of phase transitions in human hand movements. Biological
Cybernetics, 51, 347–356.Hollerbach, J. M., & Flash, T. (1982). Dynamic interactions between limb segments during planar arm movement. Biological
Cybernetics, 44(1), 67–77.Horch, K. W., Clark, F. J., & Burgess, P. R. (1975). Awareness of knee joint angle under static conditions. Journal of Neurophysiology,
38, 1436–1447.Kelso, J. A. S. (1995). Dynamic patterns. Cambridge, MA: MIT Press.Kelso, J. A. S., Del Colle, J., & Schöner, G. (1990). Action–perception as a pattern formation process. In M. Jeanrrod (Ed.), Attention
and performance 13: Motor representation and control (pp. 139–169). Hillsdale, NJ: Lawrence Erlbaum Associates.Kugler, P. N., & Turvey, M. T. (1987). Information, natural law and self-assembly of rhythmic movements. New Jersey: Lawrence
Erlbaum Associates.Pagano, C., Garrett, S., & Turvey, M. T. (1996). Is limb proprioception a function of the limbs’ inertial eigenvectors? Ecological
Psychology, 8, 43–69.Pagano, C., & Turvey, M. T. (1995). The inertia tensor as a basis for the perception of limb orientation. Journal of Experimental
Psychology: Human Perception and Performance, 21, 1070–1087.Park, H. (2003). Induced spatial and temporal symmetry breaking in coordination dynamics. Ph.D. thesis, University of
Connecticut.Park, H., & Turvey, M. T. (2008). Imperfect symmetry and the elementary coordination law. In A. Fuchs & V. K. Jirsa (Eds.),
Coordination: Neural, behavioral and social dynamics (pp. 3–25). Berlin: Springer.Ribot-Ciscar, E., Bergenheim, M., Albert, F., & Roll, J.-P. (2003). Proprioceptive population coding of limb position in humans.
Experimental Brain Research, 149(4), 512–519.Richardson, M. J., Schmidt, R. C., & Kay, B. C. (2007). Distinguishing the noise and attractor strength of coordinated limb
movements using recurrence analysis. Biological Cybernetics, 96(1), 59–78.Riley, M. A., & Black, D. P. (2003). Prism exposure affects the proprioceptive frames of reference for interlimb rhythmic
coordination. Motor Control, 7, 57–70.Schmidt, R. C., Shaw, B. K., & Turvey, M. T. (1993). Coupling dynamics in interlimb coordination. Journal of Experimental
Psychology: Human Perception and Performance, 19(2), 397–415.Schöner, G., Haken, H., & Kelso, J. A. S. (1986). A stochastic theory of phase transitions in human movement. Biological
Cybernetics, 53, 247–257.Shockley, K. (2005). Cross recurrence quantification of interpersonal postural activity. In M. A. Riley & G. Van Orden (Eds.),
Tutorials in contemporary nonlinear methods for the behavioral sciences (pp. 142–177). Retrieved from <http://www.nsf.gov/sbe/bcs/pac/nmbs/nmbs.jsp/>.
Spencer, R. M. C., Ivry, R. B., Cattaerti, D., & Semjen, A. (2005). Bimanual coordination during rhythmic movements in theabsence of somatosensory feedback. Journal of Neurophysiology, 94, 2901–2910.
Steyvers, M., Verschueren, S. M., Levin, O., Ouamer, M., & Swinnen, S. P. (2001). Proprioceptive control of cyclical bimanualforearm movements across different movement frequencies as revealed by means of tendon vibration. Experimental BrainResearch, 140(3). 1432–1106.
van de Langenberg, R., Kingma, I., & Beek, P. J. (2007). Perception of limb orientation in the vertical plane depends on center ofmass rather than inertial eigenvectors. Experimental Brain Research, 180(4), 595–607.
van de Langenberg, R., Kingma, I., & Beek, P. J. (2008). The perception of limb orientation depends on the center of mass. Journalof Experimental Psychology: Human Perception and Performance, 34, 624–639.

1036 P.L. Silva, M.T. Turvey / Human Movement Science 31 (2012) 1014–1036
Verschueren, S. M. P., Swinnen, S. P., Cordo, P. J., & Dounskaia, N. V. (1999). Proprioceptive control of multijoint movement:Bimanual circle drawing. Experimental Brain Research, 127, 182–192.
Webber, C. L., Jr., & Zbilut, J. P. (1996a). Assessing deterministic structures in physiological systems using recurrence plotstrategies. In M. C. K. Khoo (Ed.), Bioengineering approaches to pulmonary physiology and medicine (pp. 137–148). New York:Plenum Press.
Webber, C. L., Jr., & Zbilut, J. P. (1996b). Dynamical assessment of physiological systems and states using recurrent plotstrategies. Journal of Applied Physiology, 76, 965–973.


























