Embed Size (px)
Citation preview

lable at ScienceDirect
Biomaterials 32 (2011) 4773e4781
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
The role of bFGF in down-regulating a-SMA expression of chondrogenicallyinduced BMSCs and preventing the shrinkage of BMSC engineered cartilage
Qiong Li a,b, Tianyi Liu a,b, Lu Zhang a,b, Yu Liu a,b, Wenjie Zhang a,b, Wei Liu a,b, Yilin Cao a,b,*,Guangdong Zhou a,b,*
aDepartment of Plastic and Reconstructive Surgery, Shanghai 9th People’s Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai Key Laboratory of Tissue Engineering,Shanghai Stem Cell Institute, 639 Zhi Zao Ju Road, Shanghai 200011, PR ChinabNational Tissue Engineering Center of China, Shanghai, PR China
a r t i c l e i n f o
Article history:Received 20 February 2011Accepted 6 March 2011Available online 2 April 2011
Keywords:Bone marrow stromal cells (BMSCs)ChondrogenesisShrinkageSmooth muscle actin-alpha (a-SMA)Basic fibroblast growth factor (bFGF)
* Corresponding authors. Department of PlasticShanghai 9th People’s Hospital, Shanghai Jiao Tong UShanghai Key Laboratory of Tissue Engineering, ShanZhi Zao Ju Road, Shanghai 200011, PR China. Tel.: þ8621 53078128.
E-mail addresses: [email protected] (Y. Cao)(G. Zhou).
0142-9612/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.biomaterials.2011.03.020
a b s t r a c t
Bone marrow stromal cells (BMSCs) have proved to be an ideal cell source for cartilage regeneration. Ourprevious studies demonstrated that a three-dimensional (3D) cartilage could be constructed successfullyin vitro using BMSCs and biodegradable scaffolds. However, an obvious shrinkage and deformation wasobserved during in vitro chondrogenic induction. According to the literatures, it can be speculated thatthe up-regulation of smooth muscle actin-alpha (a-SMA) caused by transforming growth factor beta(TGFb) is one of the leading reasons and that basic fibroblast growth factor (bFGF) could antagonize therole of TGFb to down-regulate a-SMA expression and prevent the shrinkage of BMSC engineered carti-lage. This study testified these speculations by adding bFGF to chondrogenic media. According to thecurrent results, chondrogenic induction significantly up-regulated a-SMA expression of BMSCs at bothcell and tissue levels, and the engineered tissue only retained 12.4% of original size after 6 weeks ofchondrogenic induction. However, the supplement of bFGF in chondrogenic media efficiently down-regulated a-SMA expression and the engineered tissue still retained over 60% of original size after 6weeks of culture. Moreover, bFGF showed a beneficial influence on 3D cartilage formation of BMSCs interms of gene expression and deposition of cartilage specific matrices. All these results suggested thatbFGF could repress a-SMA expression caused by chondrogenic induction, efficiently prevent shrinkage ofBMSC engineered tissue, and have a positive influence on cartilage formation, which provides a clue forboth shape control and quality improvement of BMSC engineered 3D cartilage.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Because of strong proliferation ability, fine chondrogenic poten-tial, and easy to harvest withminimum injury, bone marrow stromalcells (BMSCs) have served as an ideal cell source for cartilage regen-eration [1e3]. Our previous studies have demonstrated that a three-dimensional (3D) cartilage could be constructed successfully usingBMSCs and biodegradable scaffolds by in vitro chondrogenic induc-tion [4]. However, BMSC engineered tissue with a large size tends toshrink and deform during chondrogenic induction and fails to
and Reconstructive Surgery,niversity School of Medicine,ghai Stem Cell Institute, 63921 23271699x5192; fax: þ86
All rights reserved.
accurately maintain the original shape and size (See SupplementalFigure 1), which obviously limits its application in repairing carti-lage defects with specific shapes, such as auricular and nasal defects.
Why BMSC engineered cartilage easily presents shrinkage anddeformation is still unclear. Smooth muscle actin-alpha (a-SMA)has been known as one of the main proteins that cause cellcontraction [5,6]. Some studies reported that the up-regulation ofa-SMA expression in skin and cornea fibroblasts was correlated toscar contraction [7,8]. However, no studies show whether expres-sion of a-SMA is correlated with the shrinkage and deformation ofBMSC engineered cartilage. It has been demonstrated that trans-forming growth factor beta (TGFb) could remarkably up-regulatea-SMA expression in both fibroblasts and BMSCs [9,10]. Unfortu-nately, TGFb is the essential growth factor for BMSC chondrogenesisin vitro [11]. Therefore, it can be speculated that the up-regulationof a-SMA expression caused by TGFb is probably one of theimportant reasons that cause the shrinkage of BMSC engineeredcartilage in vitro. In this case, down-regulating the expression of

Table 1Primer sequences of related genes.
Gene Size (bp) Primer sequences
SMA-a1 385 5-AGGAAGGACCTCTATGCTAACAAT�3,5-AACACATAGGTAACGAGTCAGAGC-3
Collagen II 293 5-GCGAGACTTGCGTCTACCC-3,5-ATTGGAGCCCTGGATGAGC-3
b-actin 208 5-CCACCGCAAATGCTTCTAG-35-GCTGTCACCTTCACCGTTCC-3
Q. Li et al. / Biomaterials 32 (2011) 4773e47814774
a-SMA would help to prevent the shrinkage of BMSC engineeredcartilage and thus maintain the original shape and size moreproperly.
Some studies showed that basic fibroblast growth factor (bFGF)could reduce the expression of a-SMA in cell level [12], but nostudies show whether bFGF can regulate a-SMA expression in 3Dtissue level. Especially, it is unclear whether bFGF can antagonizethe role of TGFb to down-regulate expression of a-SMA and preventthe shrinkage of BMSC engineered 3D cartilage. In the currentstudy, the chondrogenic media with or without bFGF were used forboth monolayer cell culture and 3D tissue construction. Expressionof a-SMA and size change of engineered tissue was analyzed toevaluate the feasibility of regulating a-SMA expression andshrinkage of engineered tissue by adding bFGF. In addition, carti-lage specific examinations were also performed to evaluate theeffect of bFGF on cartilage formation.
2. Materials and methods
2.1. BMSC culture and expansion
Bonemarrow aspirates were obtained from new born pigs provided by ShanghaiChuansha Breeding Factory (n ¼ 6). BMSCs were cultured and expanded in vitro inDulbecco’s modified Eagle medium (DMEM. Gibco BRL, Grand Island, NY) containing10% fetal bovine serum (FBS) according to previously established methods [13]. Cellsin passage two were used for the following experiments. An institutional reviewcommittee of Shanghai Jiao Tong University School of Medicine approved all animalstudy protocols.
2.2. Preparation of cell-scaffold constructs
Twenty five milligrams of unwoven polyglycolic acid (PGA) fibers (provided byDonghua University, Shanghai, China) were evenly pressed into a 13 � 13 � 2 mmrectangle. According to previously established methods [14], polylactic acid (PLA,Sigma, St. Louis, MO, USA) dissolved in dichloromethane was added to the scaffoldwith a final mass ratio of about 6% for maintaining the original shape.
BMSCs in passage two with a number of 1 � 107 cells in 200 mL were evenlyseeded onto each PLAePGA scaffold to form cell-scaffold constructs [15]. All theconstructs were cultured in DMEM plus 10% FBS for 2 days allowing sufficient cellattachment to PGA fibers and then used for grouping experiments.
2.3. Grouping design
In order to evaluate the role of bFGF in regulating a-SMA expression of chon-drogenically induced BMSCs and preventing shrinkage of BMSC engineered cartilagein vitro respectively, both BMSCs and the constructs were divided into four groupsand cultured in four different media: A. RM: Regular medium (DMEMþ10%FBS); B.CM: Chondrogenic medium (RMþ10 ng/ml TGFb1, 50 ng/ml insulin-like growthfactor I (IGF-I), 40 ng/ml dexamethasone) [16]; C. CMþ bFGF: CMþ50 ng/ml bFGF; D.bFGF: RMþ50 ng/ml bFGF. In monolayer culture system, cells were harvested at 12,24 and 48 h respectively for further analysis. In 3D culture system, the constructswere harvested at 2, 4 and 6 weeks respectively for further analysis.
2.4. Immunofluorescent staining
Cells at different time points (12, 24 or 48 h) in all groups were fixed in 10%formalin for 15 min and washed by PBS. Expression of a-SMA was detected usingmouse anti-human SMA monoclonal antibody (1:200 in PBS, Sigma), followed bygoat anti-mouse fluorescent second antibody (Invitrogen, A11029) [17]. Normalsmoothmuscle cells were used as positive control and articular cartilage as negativecontrol.
2.5. Quantitative PCR
Total RNA was extracted from both cell and tissue samples and cDNA wasobtained by reverse polymerase transcription (RT) according to previously describedmethods [18]. Quantitative polymerase chain reaction (QPCR) was performed toquantitatively analyze mRNA level of a-SMA and cartilage specific genes [18]. The b-actin mRNA level was quantified as an internal control. The forward and reverseprimer sequences were listed in Table 1.
2.6. Gross observation
The gross views of cell-scaffold constructs in different groups were recorded byphotos every week. The area ratio of each sample compared to 6-well culture plate
was calculated respectively according to the formula: the area of the constructs/thebottom area of 6-well culture plate in the same picture. The area change of eachspecimenwith culture timewas quantified using a percentage, whichwas calculatedaccording to the formula: the area ratio of the same specimen at different timepoint/its original area ratio � 100%. The curves of area change with culture timewere drawn according to the quantitative percentages in all groups.
2.7. ELISA
Enzyme linked immunosorbent assay (ELISA) was used to quantitatively analyzethe level of a-SMA expression in different groups. According to the literatures [19],specimens from both cells and engineered tissues were completely homogenized torelease total proteins. After centrifuging (to get rid of the residue of specimens), thesupernatant was harvested for ELISA.
2.8. Quantitative analysis of cartilage specific matrices
The contents of cartilage specific matrices in engineered tissues were quanti-tatively detected to evaluate the influence of bFGF on 3D chondrogenesis of BMSCs.After 6-week in vitro culture, wet weight of samples was measured by an electronicbalance. Total glycosaminoglycan (GAG) content was analyzed by spectrophoto-metric microdetermination with dimethylmethylene blue as previously described[19]. The quantitative amount of type II collagen was measured by ELISA asmentioned above.
2.9. Histological and immunohistochemical analysis
The specimens were fixed in 10% formalin for 24 h prior to processing andembedded in paraffin for sectioning. Specimens were sectioned to 6 mm, mountedon glass slides, and stained with hematoxylin and eosin (H&E) or Safranin-O usingstandard histological techniques.
Expression of type II collagen was detected using mouse anti-human type IIcollagen monoclonal antibody (1:200 in PBS, Santa Cruz, CA, USA), followed byhorseradish peroxidase (HRP)-conjugated anti-mouse antibody (1:200 in PBS, SantaCruz) and color development with diaminobenzidine tetrahydrochloride (DAB,Santa Cruz) [19].
2.10. Statistical analysis
The percentage of area change, wet weight, GAG content, collagen II content,ELISA as well as QPCR results were analyzed using Student’s t-test. A p-value lessthan 0.05 was considered as statistically significant.
3. Results
3.1. The role of bFGF in down-regulating a-SMA expression causedby chondrogenic induction
The role of bFGF in regulating a-SMA expression of BMSCs wasfirstly investigated in monolayer cell culture system. As shown inFig. 1, an obvious change in cell morphology was observed in CMand CM þ bFGF groups compared to RM and bFGF groups, sug-gesting a result of chondrogenic induction. According to the resultsof immunofluorescent staining (Fig. 1), no visible positive stainingof a-SMA was shown in RM and bFGF groups. However, strongpositive staining of a-SMA was observed in CM group at all timepoints, indicating that chondrogenic induction could obviously up-regulate a-SMA expression. Noticeably, a relatively weak positivestaining of a-SMA (compared to CM group) was observed inCM þ bFGF group, indicating that supplement of bFGF could
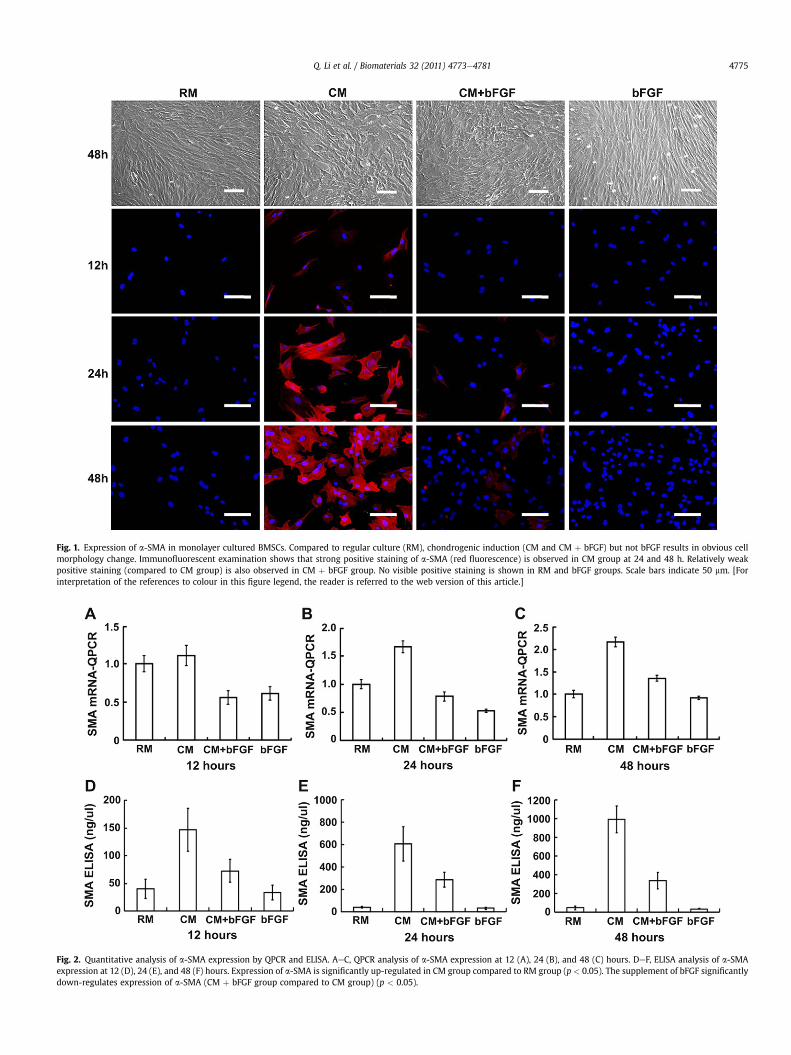
Fig. 1. Expression of a-SMA in monolayer cultured BMSCs. Compared to regular culture (RM), chondrogenic induction (CM and CM þ bFGF) but not bFGF results in obvious cellmorphology change. Immunofluorescent examination shows that strong positive staining of a-SMA (red fluorescence) is observed in CM group at 24 and 48 h. Relatively weakpositive staining (compared to CM group) is also observed in CM þ bFGF group. No visible positive staining is shown in RM and bFGF groups. Scale bars indicate 50 mm. [Forinterpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.]
Fig. 2. Quantitative analysis of a-SMA expression by QPCR and ELISA. AeC, QPCR analysis of a-SMA expression at 12 (A), 24 (B), and 48 (C) hours. DeF, ELISA analysis of a-SMAexpression at 12 (D), 24 (E), and 48 (F) hours. Expression of a-SMA is significantly up-regulated in CM group compared to RM group (p < 0.05). The supplement of bFGF significantlydown-regulates expression of a-SMA (CM þ bFGF group compared to CM group) (p < 0.05).
Q. Li et al. / Biomaterials 32 (2011) 4773e4781 4775
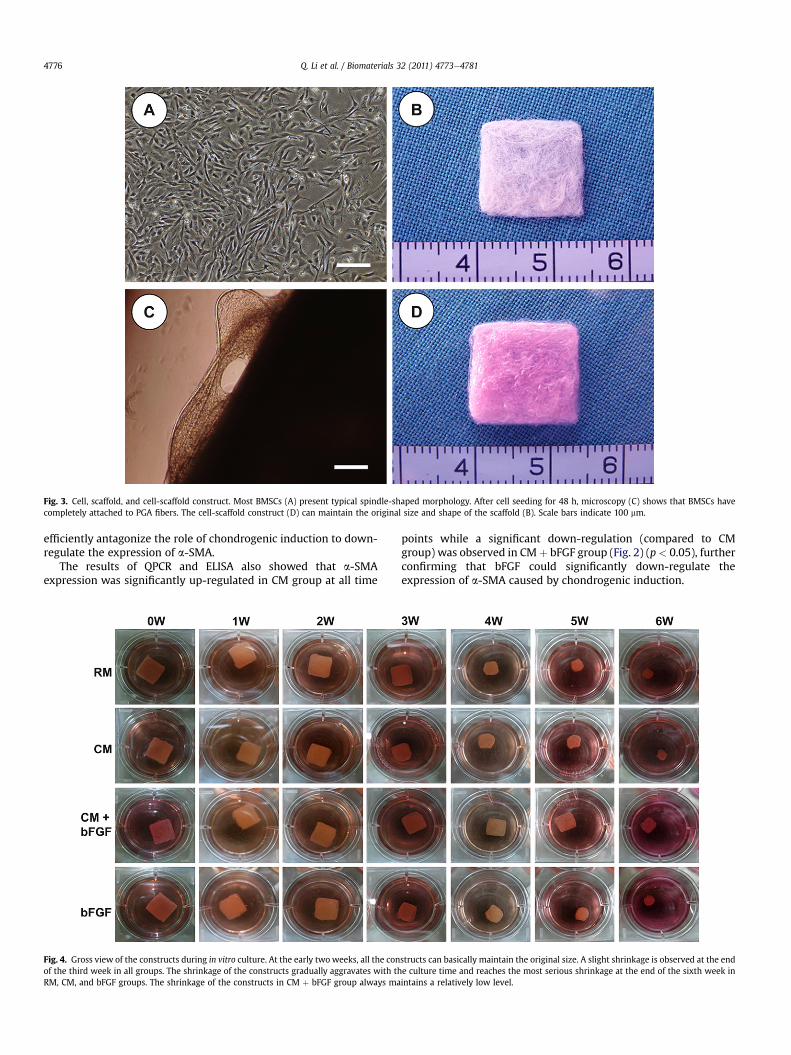
Fig. 3. Cell, scaffold, and cell-scaffold construct. Most BMSCs (A) present typical spindle-shaped morphology. After cell seeding for 48 h, microscopy (C) shows that BMSCs havecompletely attached to PGA fibers. The cell-scaffold construct (D) can maintain the original size and shape of the scaffold (B). Scale bars indicate 100 mm.
Q. Li et al. / Biomaterials 32 (2011) 4773e47814776
efficiently antagonize the role of chondrogenic induction to down-regulate the expression of a-SMA.
The results of QPCR and ELISA also showed that a-SMAexpression was significantly up-regulated in CM group at all time
Fig. 4. Gross view of the constructs during in vitro culture. At the early two weeks, all the conof the third week in all groups. The shrinkage of the constructs gradually aggravates with thRM, CM, and bFGF groups. The shrinkage of the constructs in CM þ bFGF group always ma
points while a significant down-regulation (compared to CMgroup) was observed in CMþ bFGF group (Fig. 2) (p< 0.05), furtherconfirming that bFGF could significantly down-regulate theexpression of a-SMA caused by chondrogenic induction.
structs can basically maintain the original size. A slight shrinkage is observed at the ende culture time and reaches the most serious shrinkage at the end of the sixth week inintains a relatively low level.
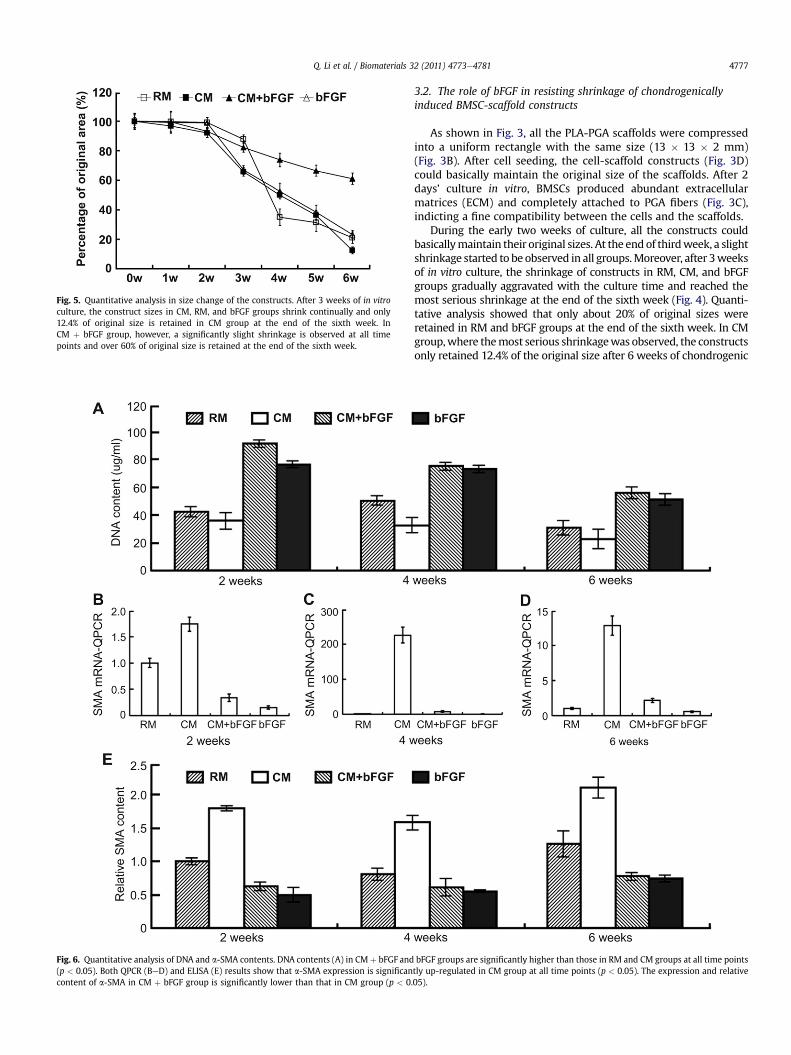
Fig. 5. Quantitative analysis in size change of the constructs. After 3 weeks of in vitroculture, the construct sizes in CM, RM, and bFGF groups shrink continually and only12.4% of original size is retained in CM group at the end of the sixth week. InCM þ bFGF group, however, a significantly slight shrinkage is observed at all timepoints and over 60% of original size is retained at the end of the sixth week.
Fig. 6. Quantitative analysis of DNA and a-SMA contents. DNA contents (A) in CM þ bFGF and(p < 0.05). Both QPCR (BeD) and ELISA (E) results show that a-SMA expression is significancontent of a-SMA in CM þ bFGF group is significantly lower than that in CM group (p < 0.
Q. Li et al. / Biomaterials 32 (2011) 4773e4781 4777
3.2. The role of bFGF in resisting shrinkage of chondrogenicallyinduced BMSC-scaffold constructs
As shown in Fig. 3, all the PLA-PGA scaffolds were compressedinto a uniform rectangle with the same size (13 � 13 � 2 mm)(Fig. 3B). After cell seeding, the cell-scaffold constructs (Fig. 3D)could basically maintain the original size of the scaffolds. After 2days’ culture in vitro, BMSCs produced abundant extracellularmatrices (ECM) and completely attached to PGA fibers (Fig. 3C),indicting a fine compatibility between the cells and the scaffolds.
During the early two weeks of culture, all the constructs couldbasicallymaintain their original sizes. At the end of thirdweek, a slightshrinkage started to be observed in all groups.Moreover, after 3weeksof in vitro culture, the shrinkage of constructs in RM, CM, and bFGFgroups gradually aggravated with the culture time and reached themost serious shrinkage at the end of the sixth week (Fig. 4). Quanti-tative analysis showed that only about 20% of original sizes wereretained in RM and bFGF groups at the end of the sixth week. In CMgroup,where themost serious shrinkagewas observed, the constructsonly retained 12.4% of the original size after 6 weeks of chondrogenic
bFGF groups are significantly higher than those in RM and CM groups at all time pointstly up-regulated in CM group at all time points (p < 0.05). The expression and relative05).
Q. Li et al. / Biomaterials 32 (2011) 4773e47814778
induction (Fig. 5). In CM þ bFGF group, however, an obviously slightshrinkagewas shownduring invitro culture (Fig. 4) andover60%of theoriginal sizewas retained at the end of the sixthweek, a percentage ofover four folds of that in CM group (Fig. 5) (p< 0.05). All these resultsindicated that bFGF could significantly resist the shrinkage of theconstructs caused by chondrogenic induction.
3.3. The role of bFGF in regulating cell proliferation and a-SMAexpression of chondrogenically induced BMSC-scaffold constructs
In consideration of the influence of cell amount on a-SMAcontent, total DNA contents in all groups were firstly detected so asto standardize relative content of a-SMA. As shown in Fig. 6, DNAcontents in all the bFGF-containing groups (CM þ bFGF and bFGFgroups) were significantly higher than those in bFGF-free groups(RM and CM groups) at all time points (Fig. 6A) (p < 0.05), indi-cating that bFGF could better retain cell viability on 3D scaffolds.
The QPCR results showed that a-SMA expressionwas significantlyhigher in CM group compared to RM group at all time points(Fig. 6BeD)(p < 0.05), indicating that chondrogenic induction up-regulated a-SMA expression of BMSCs on 3D scaffolds. Noticeably,a-SMA expression in CM þ bFGF group was significantly lower thanthat in CM group at all time points (Fig. 6BeD)(p < 0.05), indicatingthat supplement of bFGF could down-regulate a-SMA expressioncaused by chondrogenic induction. Consistent with the gene expres-sion level, the relative content of a-SMA (standardized by DNAcontent) in CM group was significantly higher than all other groups(Fig. 6E)(p < 0.05), further suggesting that chondrogenic inductioncould significantly enhance a-SMA content and this role could beantagonized by bFGF.
3.4. The influence of bFGF on 3D cartilage formation ofchondrogenically induced BMSC-scaffold constructs
The influence of bFGF on 3D cartilage formation was furtherevaluated by histology and quantitative analysis of cartilage specific
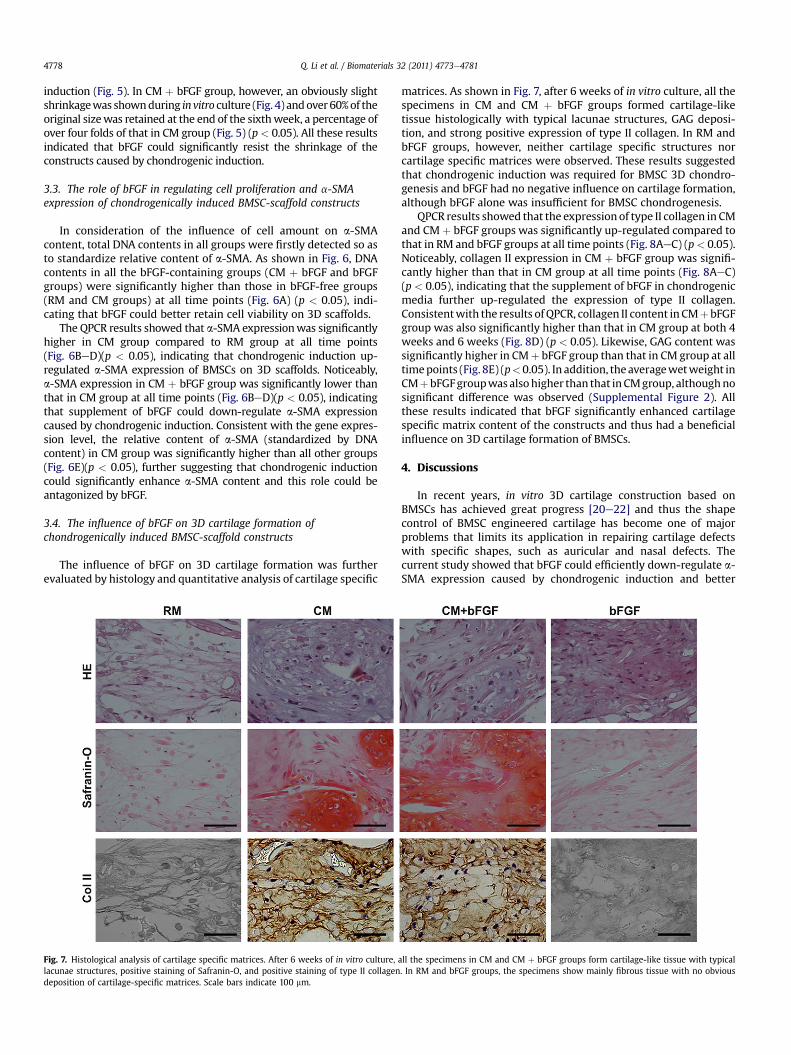
Fig. 7. Histological analysis of cartilage specific matrices. After 6 weeks of in vitro culture,lacunae structures, positive staining of Safranin-O, and positive staining of type II collagendeposition of cartilage-specific matrices. Scale bars indicate 100 mm.
matrices. As shown in Fig. 7, after 6 weeks of in vitro culture, all thespecimens in CM and CM þ bFGF groups formed cartilage-liketissue histologically with typical lacunae structures, GAG deposi-tion, and strong positive expression of type II collagen. In RM andbFGF groups, however, neither cartilage specific structures norcartilage specific matrices were observed. These results suggestedthat chondrogenic induction was required for BMSC 3D chondro-genesis and bFGF had no negative influence on cartilage formation,although bFGF alone was insufficient for BMSC chondrogenesis.
QPCR results showed that the expression of type II collagen in CMand CM þ bFGF groups was significantly up-regulated compared tothat in RM and bFGF groups at all time points (Fig. 8AeC) (p< 0.05).Noticeably, collagen II expression in CM þ bFGF group was signifi-cantly higher than that in CM group at all time points (Fig. 8AeC)(p < 0.05), indicating that the supplement of bFGF in chondrogenicmedia further up-regulated the expression of type II collagen.Consistentwith the results ofQPCR, collagen II content in CMþbFGFgroup was also significantly higher than that in CM group at both 4weeks and 6 weeks (Fig. 8D) (p < 0.05). Likewise, GAG content wassignificantly higher in CMþ bFGF group than that in CM group at alltimepoints (Fig. 8E) (p<0.05). In addition, the averagewetweight inCMþbFGFgroupwasalsohigher than that inCMgroup, althoughnosignificant difference was observed (Supplemental Figure 2). Allthese results indicated that bFGF significantly enhanced cartilagespecific matrix content of the constructs and thus had a beneficialinfluence on 3D cartilage formation of BMSCs.
4. Discussions
In recent years, in vitro 3D cartilage construction based onBMSCs has achieved great progress [20e22] and thus the shapecontrol of BMSC engineered cartilage has become one of majorproblems that limits its application in repairing cartilage defectswith specific shapes, such as auricular and nasal defects. Thecurrent study showed that bFGF could efficiently down-regulate a-SMA expression caused by chondrogenic induction and better
all the specimens in CM and CM þ bFGF groups form cartilage-like tissue with typical. In RM and bFGF groups, the specimens show mainly fibrous tissue with no obvious
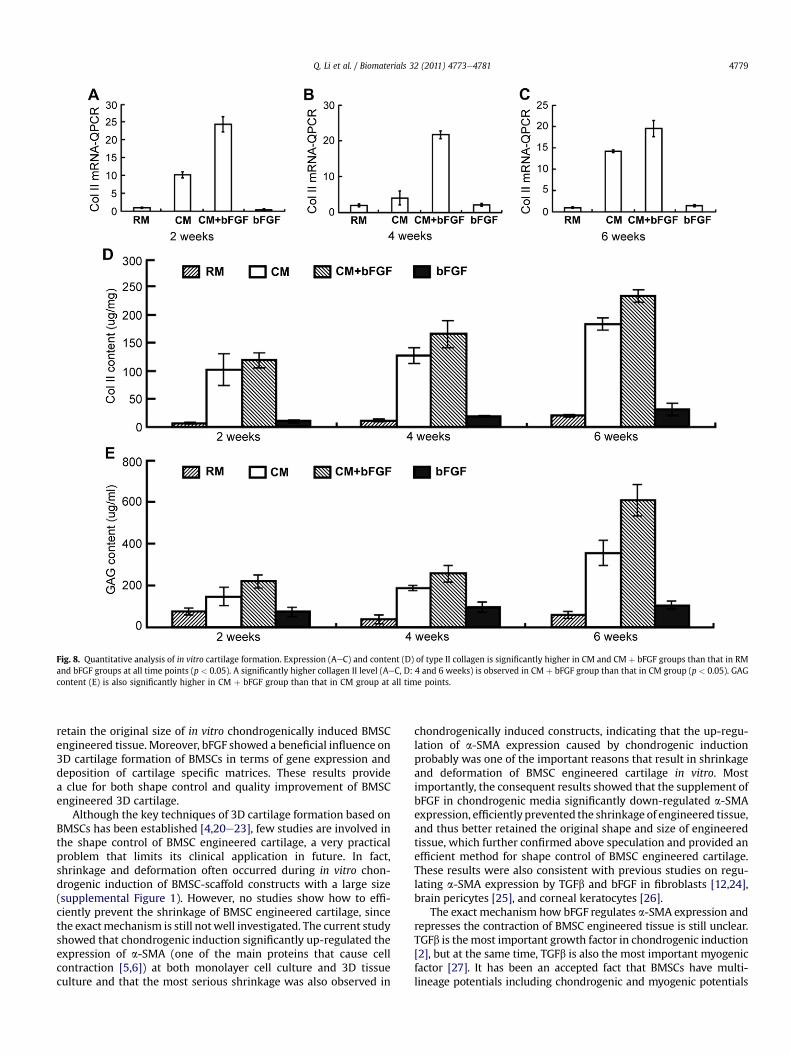
Fig. 8. Quantitative analysis of in vitro cartilage formation. Expression (AeC) and content (D) of type II collagen is significantly higher in CM and CM þ bFGF groups than that in RMand bFGF groups at all time points (p < 0.05). A significantly higher collagen II level (AeC, D: 4 and 6 weeks) is observed in CM þ bFGF group than that in CM group (p < 0.05). GAGcontent (E) is also significantly higher in CM þ bFGF group than that in CM group at all time points.
Q. Li et al. / Biomaterials 32 (2011) 4773e4781 4779
retain the original size of in vitro chondrogenically induced BMSCengineered tissue. Moreover, bFGF showed a beneficial influence on3D cartilage formation of BMSCs in terms of gene expression anddeposition of cartilage specific matrices. These results providea clue for both shape control and quality improvement of BMSCengineered 3D cartilage.
Although the key techniques of 3D cartilage formation based onBMSCs has been established [4,20e23], few studies are involved inthe shape control of BMSC engineered cartilage, a very practicalproblem that limits its clinical application in future. In fact,shrinkage and deformation often occurred during in vitro chon-drogenic induction of BMSC-scaffold constructs with a large size(supplemental Figure 1). However, no studies show how to effi-ciently prevent the shrinkage of BMSC engineered cartilage, sincethe exactmechanism is still notwell investigated. The current studyshowed that chondrogenic induction significantly up-regulated theexpression of a-SMA (one of the main proteins that cause cellcontraction [5,6]) at both monolayer cell culture and 3D tissueculture and that the most serious shrinkage was also observed in
chondrogenically induced constructs, indicating that the up-regu-lation of a-SMA expression caused by chondrogenic inductionprobably was one of the important reasons that result in shrinkageand deformation of BMSC engineered cartilage in vitro. Mostimportantly, the consequent results showed that the supplement ofbFGF in chondrogenic media significantly down-regulated a-SMAexpression, efficiently prevented the shrinkage of engineered tissue,and thus better retained the original shape and size of engineeredtissue, which further confirmed above speculation and provided anefficient method for shape control of BMSC engineered cartilage.These results were also consistent with previous studies on regu-lating a-SMA expression by TGFb and bFGF in fibroblasts [12,24],brain pericytes [25], and corneal keratocytes [26].
The exact mechanism how bFGF regulates a-SMA expression andrepresses the contraction of BMSC engineered tissue is still unclear.TGFb is themost important growth factor in chondrogenic induction[2], but at the same time, TGFb is also the most important myogenicfactor [27]. It has been an accepted fact that BMSCs have multi-lineage potentials including chondrogenic and myogenic potentials

Q. Li et al. / Biomaterials 32 (2011) 4773e47814780
[28]. In general, TGFb initiating differentiation of BMSCs towardswhether chondrogenic or myogenic mainly depends on thecombined factors. TGFb combining with insulin-like growth factor I(IGF-I) and dexamethasone mainly promotes chondrogenic differ-entiation [29,30], whereas TGFb combining with platelet-derivedgrowth factor (PDGF)-BB mainly promotes myogenic differentiation[31]. Nevertheless, TGFb in the current chondrogenic media appar-ently partially initiated BMSCs’ myogenic differentiation, sincea-SMA, as one of relatively specific markers of smooth muscle cells(SMCs) [32], was significantly up-regulated during chondrogenicinduction. The latest studies reported that TGFb could up-regulatethe expression of SMC marker genes (including a-SMA) throughactivating Src family of tyrosine kinase [33], whereas bFGF couldrepress this role of TGFb by activating MEK1 pathway [34]. In addi-tion, studies also demonstrated that bFGF could enhance BMSCchondrogenesis by regulating MAPK and Wnt signaling [35], whichwas consistent with the current results. All these evidences impliedthat the role of bFGF in regulating a-SMA expression and repressingcontraction of BMSC engineered tissue was mainly mediatedby antagonizing TGFb-driven SMC differentiation and enhancingchondrogenic differentiation. Nevertheless, the exact mechanismstill needs to be further investigated.
It needs to be proposed that the final effect of bFGF resistingcontraction of BMSC engineered tissue was a result of multifactorparticipation but not a-SMA alone. Firstly, themechanical property ofthe scaffold was one of the important factors. The PGA fibers used inthe current study usually started to degrade after 2 weeks of in vitroculture and thus themechanical strength of the constructs decreasedgradually, which provided a reasonable explanation that no visiblecontraction was observed in all groups within two weeks. Secondly,the deposition ratio of extracellular matrices (ECM) was anotherimportant factor. In RM and bFGF groups, due to lack of chondrogenicinduction, the deposition ratio of ECMwas much slower than degra-dation ratio of the scaffolds.With the degradation of the scaffolds, theconstructs inevitably formed small size tissue due to insufficient ECM,which had no relationship with a-SMA level. In CM þ bFGF group,bFGF combining with chondrogenic induction significantly enhancedthe deposition ratio of ECM,whichfilledwith the space caused by thedegradation of the scaffold and thus helped to retain the original sizeand shape especially in the case of low level of a-SMA. In CM group,however, the high level of a-SMA had become dominating factor thatcaused contraction, although chondrogenic induction ensured a highdeposition ratio of ECM. Finally, bFGF antagonizing the role of TGFbprobably did not limit the regulation of a-SMA. The role of bFGF inregulating cell mitosis [36] and other contraction-related proteins(such as smooth muscle protein 22-a [34], fibronection [37] etc.) wasalso opposite to TGFb, which also probably helped to represscontraction caused by chondrogenic induction.
5. Conclusion
In summary, this study demonstrated that the up-regulation ofa-SMA expression was an important reason that resulted in theshrinkage of BMSC engineered cartilage and that bFGF couldsignificantly repress a-SMA expression, efficiently prevent theshrinkage, and have a positive influence on 3D cartilage formation.Although the exact mechanism still needs to be further investi-gated, this study provides a clue for both shape control and qualityimprovement of BMSC engineered 3D cartilage.
Acknowledgements
This research was supported by the National Basic ResearchProgram of China (2005CB522702)Hi-Tech Research and Develop-ment Program of China (2006AA02A126)National Natural Science
Foundation of China (30973131, 30973130, 50830105, and81000677)Shanghai Shu Guang Foundation (08SG19)andShanghaiRising-star Program (09QH1401600). The authors appreciate thetechnical supports from Juanjuan Wu, Demin Yin, and Lijuan Zongof our laboratory.
Appendix. Supplementary material
Supplementary data related to this article can be found online atdoi:10.1016/j.biomaterials.2011.03.020.
References
[1] Mackay AM, Beck SC, Murphy JM, Barry FP, Chichester CO, Pittenger MF.Chondrogenic differentiation of cultured human mesenchymal stem cellsfrom marrow. Tissue Eng 1998;4(4):415e28.
[2] Johnstone B, Hering TM, Caplan AI, Goldberg VM, Yoo JU. In vitro chondro-genesis of bone marrow-derived mesenchymal progenitor cells. Exp Cell Res1998;238(1):265e72.
[3] Barry F, Boynton RE, Liu B, Murphy JM. Chondrogenic differentiation ofmesenchymal stem cells from bone marrow: differentiation-dependent geneexpression of matrix components. Exp Cell Res 2001;268(2):189e200.
[4] Liu K, Zhou GD, Liu W, Zhang WJ, Cui L, Liu X, et al. The dependence of in vivostable ectopic chondrogenesis by human mesenchymal stem cells on chon-drogenic differentiation in vitro. Biomaterials 2008;29(14):2183e92.
[5] Kinner B, Spector M. Smooth muscle actin expression by human articularchondrocytes and their contraction of a collagen-glycosaminoglycan matrix invitro. J Orthop Res 2001;19(2):233e41.
[6] Hinz B, Celetta G, Tomasek JJ, Gabbiani G, Chaponnier C. Alpha-smooth muscleactin expression upregulates fibroblast contractile activity. Mol Biol Cell2001;12(9):2730e41.
[7] Akasaka Y, Ono I, Tominaga A, Ishikawa Y, Ito K, Suzuki T, et al. Basic fibroblastgrowth factor in an artificial dermis promotes apoptosis and inhibitsexpression of alpha-smooth muscle actin, leading to reduction of woundcontraction. Wound Repair Regen 2007;15(3):378e89.
[8] Kurosaka H, Kurosaka D, Kato K, Mashima Y, Tanaka Y. Transforming growthfactor-beta 1 p.omotes contraction of collagen gel by bovine corneal fibro-blasts through differentiation of myofibroblasts. Invest Ophthalmol Vis Sci1998;39(5):699e704.
[9] Arora PD, Narani N, McCulloch CA. The compliance of collagen gels regulatestransforming growth factor-beta induction of alpha-smooth muscle actin infibroblasts. Am J Pathol 1999;154(3):871e82.
[10] Narita Y, Yamawaki A, Kagami H, Ueda M, Ueda Y. Effects of transforminggrowth factor-beta 1 and ascorbic acid on differentiation of human bone-marrow-derived mesenchymal stem cells into smooth muscle cell lineage.Cell Tissue Res 2008;333(3):449e59.
[11] Hellingman CA, Blaney Davidson E, Koevoet W, Vitters EL, van den Berg WB,van Osch G, et al. Smad signaling determines chondrogenic differentiation ofbone-marrow derived mesenchymal stem cells: inhibition of Smad 1/5/8Pprevents terminal differentiation and calcification. Tissue Eng Part A; 2011[Epub ahead of print].
[12] Khouw IM, van Wachem PB, Plantinga JA, Vujaskovic Z, Wissink MJ, de Leij LF,et al. TGF-beta and bFGF affect the differentiation of proliferating porcinefibroblasts into myofibroblasts in vitro. Biomaterials 1999;20(19):1815e22.
[13] Zhou G, Liu W, Cui L, Wang X, Liu T, Cao Y. Repair of porcine articularosteochondral defects in non-weightbearing areas with autologous bonemarrow stromal cells. Tissue Eng 2006;12(11):3209e21.
[14] Xu L, Cao D, Liu W, Zhou G, Zhang WJ, Cao Y. In vivo engineering of a efunc-tional tendon sheath in a hen model. Biomaterials 2010;31(14):3894e902.
[15] Liu Y, Zhang L, Zhou G, Li Q, Liu W, Yu Z, et al. In vitro engineering of humanear-shaped cartilage assisted with CAD/CAM technology. Biomaterials2010;31(8):2176e83.
[16] Liu TY, Zhou GD, Wei X, Wu XL, Chen FG, Cui L, et al. [Influence of trans-forming growth factor-beta1 inducing time on chondrogenesis of bonemarrow stromal cells (BMSCs): in vitro experiment with porcine BMSCs].Zhonghua Yi Xue Za Zhi 2007;87(31):2218e22.
[17] Wang C, Yin S, Cen L, Liu Q, Liu W, Cao Y, et al. Differentiation of adipose-derived stem cells into contractile smooth muscle cells induced by trans-forming growth factor-beta1 and bone morphogenetic protein-4. Tissue EngPart A 2010;16(4):1201e13.
[18] Zhu J, Li J, Wang B, Zhang WJ, Zhou G, Cao Y, et al. The regulation of phenotypeof cultured tenocytes by microgrooved surface structure. Biomaterials2010;31(27):6952e8.
[19] Yan D, Zhou G, Zhou X, Liu W, Zhang WJ, Luo X, et al. The impact of low levelsof collagen IX and pyridinoline on the mechanical properties of in vitroengineered cartilage. Biomaterials 2009;30(5):814e21.
[20] Alves da Silva ML, Martins A, Costa-Pinto AR, Correlo VM, Sol P,Bhattacharya M, et al. Chondrogenic differentiation of human bone marrowmesenchymal stem cells in chitosan-based scaffolds using a flow-perfusionbioreactor. J Tissue Eng Regen Med; 2010 [Epub ahead of print].

Q. Li et al. / Biomaterials 32 (2011) 4773e4781 4781
[21] Park JS, Yang HN, Woo DG, Chung HM, Park KH. In vitro and in vivo chon-drogenesis of rabbit bone marrow-derived stromal cells in fibrin matrixmixed with growth factor loaded in nanoparticles. Tissue Eng Part A2009;15(8):2163e75.
[22] Liu L, Wu W, Tuo X, Geng W, Zhao J, Wei J, et al. Novel strategy to engineertrachea cartilage graft with marrow mesenchymal stem cell macroaggregateand hydrolyzable scaffold. Artif Organs 2010;34(5):426e33.
[23] Worster AA, Brower-Toland BD, Fortier LA, Bent SJ, Williams J, Nixon AJ.Chondrocytic differentiation of mesenchymal stem cells sequentially exposedto transforming growth factor-beta1 in monolayer and insulin-like growthfactor-I in a three-dimensional matrix. J Orthop Res 2001;19(4):738e49.
[24] Narine K, DeWever O, Van Valckenborgh D, Francois K, BrackeM, DeSmet S, et al.Growth factor modulation of fibroblast proliferation, differentiation, and inva-sion: implications for tissue valve engineering. TissueEng2006;12(10):2707e16.
[25] Verbeek MM, Otte-Holler I, Wesseling P, Ruiter DJ, de Waal RM. Induction ofalpha-smooth muscle actin expression in cultured human brain pericytes bytransforming growth factor-beta 1. Am J Pathol 1994;144(2):372e82.
[26] Jester JV, Barry-Lane PA, Cavanagh HD, Petroll WM. Induction of alpha-smooth muscle actin expression and myofibroblast transformation in culturedcorneal keratocytes. Cornea 1996;15(5):505e16.
[27] Florini JR, Ewton DZ. Actions of transforming growth factor-beta on musclecells. J Cell Physiol 1988;135(2):301e8.
[28] Pansky A, Roitzheim B, Tobiasch E. Differentiation potential of adult humanmesenchymal stem cells. Clin Lab 2007;53(1e2):81e4.
[29] Longobardi L, O’Rear L, Aakula S, Johnstone B, Shimer K, Chytil A, et al. Effect ofIGF-I in the chondrogenesis of bone marrow mesenchymal stem cells in thepresence or absence of TGF-beta signaling. J Bone Miner Res2006;21(4):626e36.
[30] Hickok NJ, Haas AR, Tuan RS. Regulation of chondrocyte differentiation andmaturation. Microsc Res Tech 1998;43(2):174e90.
[31] Tian H, Bharadwaj S, Liu Y, Ma H, Ma PX, Atala A, et al. Myogenic differenti-ation of human bone marrow mesenchymal stem cells on a 3D nano fibrousscaffold for bladder tissue engineering. Biomaterials 2010;31(5):870e7.
[32] Skalli O, Ropraz P, Trzeciak A, Benzonana G, Gillessen D, Gabbiani G.A monoclonal antibody against alpha-smooth muscle actin: a new probe forsmooth muscle differentiation. J Cell Biol 1986;103(6 Pt 2):2787e96.
[33] Sato M, Kawai-Kowase K, Sato H, Oyama Y, Kanai H, Ohyama Y, et al. c-Src andhydrogen peroxide mediate transforming growth factor-beta1-inducedsmooth muscle cell-gene expression in 10T1/2 cells. Arterioscler Thromb VascBiol 2005;25(2):341e7.
[34] Kawai-Kowase K, Sato H, Oyama Y, Kanai H, Sato M, Doi H, et al. Basicfibroblast growth factor antagonizes transforming growth factor-beta1-induced smooth muscle gene expression through extracellular signal-regu-lated kinase 1/2 signaling pathway activation. Arterioscler Thromb Vasc Biol2004;24(8):1384e90.
[35] Solchaga LA, Penick K, Porter JD, Goldberg VM, Caplan AI, Welter JF. FGF-2enhances the mitotic and chondrogenic potentials of human adult bonemarrow-derivedmesenchymal stem cells. J Cell Physiol 2005;203(2):398e409.
[36] Carcamo-Orive I, Tejados N, Delgado J, Gaztelumendi A, Otaegui D, Lang V,et al. ERK2 p.otein regulates the proliferation of human mesenchymal stemcells without affecting their mobilization and differentiation potential. ExpCell Res 2008;314(8):1777e88.
[37] Frieri M, Capetandes A. The effect of enantiomers of beta-agonists on myofi-broblast-derived vascular endothelial growth factor and other matrixcomponents in the presence of dust-mite extract. Allergy Asthma Proc2008;29(2):182e8.





























