Embed Size (px)
Citation preview

1. Introduction
2. The relevance of AOXs in drug
metabolism and drug discovery
3. The number of active AOX
genes varies
in different animal species:
implications
for drug metabolism studies
4. Factors influencing the activity
of AOXs
5. Conclusion
6. Expert opinion
Review
The role of aldehyde oxidasein drug metabolismEnrico Garattini† & Mineko TeraoLaboratory of Molecular Biology, Istituto di Ricerche Farmacologiche “Mario Negri” , Milano, Italy
Introduction: Aldehyde oxidases (AOXs) are molybdo-flavoenzymes with com-
plex evolutionary profiles, as the number and types of active AOX genes vary
according to the animal species considered. Humans and higher primates
have a single functional AOX1 gene, while rodents are endowed with four
AOXs. Along with the endoplasmic cytochrome P450 system (CYP450), cyto-
plasmic AOX1 is the major enzyme involved in the hepatic phase I metabolism
of numerous xenobiotics.
Areas covered: The authors review literature to highlight the fact that alde-
hydes are not the only AOX substrates, as aza- and oxo-heterocycles, that rep-
resent the scaffold of many drugs, are also oxidized efficiently by these
enzymes. Additionally, the ndefine the different complements of AOX isoen-
zymes expressed in humans and animal models used in drug metabolism stud-
ies and discuss the implications. Furthermore, the authors report on human
AOX1 allelic variants that alter the activity of this enzyme. Finally, they discuss
the factors of potential importance in controlling the functional activity
of AOX1.
Expert opinion: There is evidence for an increasing relevance of AOX1 in the
metabolism and clearance of new drugs, as measures aiming at controlling
CYP450-dependent metabolism of prospective therapeutic agents are becom-
ing routine. This calls for investigations into the biology, catalytic properties
and substrate specificity of human AOX1.
Keywords: aldehyde oxidase, drug discovery, drug metabolism, molybdo-enzymes
Expert Opin. Drug Metab. Toxicol. (2012) 8(4):487-503
1. Introduction
Aldehyde oxidases (AOXs) are a group of enzymes belonging to the family ofmolybdo-flavoprotein (MOFE), requiring flavin adenine dinucleotide (FAD) anda molybdo-pterin cofactor (MoCo) for their catalytic activity [1-3]. AOXs are struc-turally conserved proteins and are represented throughout the animal kingdom,from insects to man [2,4,5]. AOXs are characterized by a high level of amino acididentity with another MOFE, xanthine oxidoreductase (XOR), which is the keyenzyme in the catabolism of purines, oxidizing hypoxanthine into xanthine and xan-thine into uric acid [6-11]. The gene coding for mammalian AOXs and XORs con-sists of 35/36 exons, whose structure and intron junctions are also highlyconserved. Until a few years ago it was believed that a single AOX enzyme was pres-ent in mammals, including primates and humans. In humans, this enzyme wasknown to be encoded by a single gene mapping to chromosome 2 where theXOR gene was also located [12,13]. It is now clear that the number of AOX genesvaries from one to four according to the vertebrate species considered [2,3]. Withrespect to this, the two extremes are represented by humans, whose genome containsa single functional gene, AOX1, whereas mice and rats are endowed with foursuch genes, AOX1, AOX3, AOX4 and AOX3l1, coding for an equivalent numberof structurally very conserved proteins.
10.1517/17425255.2012.663352 © 2012 Informa UK, Ltd. ISSN 1742-5255 487All rights reserved: reproduction in whole or in part not permitted
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

AOXs and XORs are characterized by similar primary, sec-ondary and tertiary structures. The catalytically activeenzymes are homodimers consisting of two identical subunitsendowed with a tripartite structure (Figure 1A). In fact, thetwo subunits contain three functionally distinct and conserveddomains separated by short hinge regions whose amino acidsequences vary in AOXs and XORs. The amino-terminal domain (25 kDa) contains two non-identical iron/sulfur centers, the central domain (45 kDa) is characterizedby a FAD binding site, and the carboxy-terminal domain con-sists of the MoCo binding site which is located in close prox-imity to the substrate pocket. AOXs are devoid of thenicotinamide adenine dinucleotide (NAD) binding site,which is present in XORs [4]. In fact, XORs exist in aNAD-dependent dehydrogenase and a NAD-independent
oxidase form [14], while AOXs are pure oxidases utilizingboth H2O and molecular oxygen to carry out their enzymaticfunction. A typical catalytic cycle for the oxidation of a gen-eral substrate, R-H, by AOXs is illustrated in Figure 1B. TheR-H substrate is oxidized into the corresponding R-OH prod-uct at the MoCo center. The electron equivalents are trans-ferred to FAD, which is reoxidized by molecular oxygenwith the formal production of hydrogen peroxide. The twoiron/sulfur centers mediate the transfer of the reducing equiv-alents from MoCo to FAD and act as electron sinks duringthe catalytic process [15].
The endogenous substrate(s) of AOXs have not yet beenidentified with certainty, although various molecules havebeen proposed [2]. At present, the only AOX for which thereis in vivo experimental evidence of an endogenous substrateis AOX4, which has been shown to oxidize retinaldehydeinto retinoic acid, the active metabolite of vitamin A [16].The physiological role of AOX1 and other mammalian mem-bers of the family is equally unknown. Nevertheless, recentevidence indicates that human AOX1 expression in theadipose tissue may be a determinant of fat deposition possiblyvia mechanisms that may be unrelated to the enzymaticactivity of the protein [17,18].
2. The relevance of AOXs in drug metabolismand drug discovery
Unlike XORs which are characterized by a significant level ofsubstrate specificity, AOXs act on a wide range of substrateswith different chemical structures and functionalities. Broadsubstrate specificity along with the presence of this enzymaticactivity in the hepatic tissue of humans and many other mam-mals are at the basis of the role which has been ascribed toAOXs in metabolizing foreign compounds of medicinaland toxicological importance [1,19-32]. Although it is wellestablished that the cytochrome P450 enzymatic system(CYP450) is the primary determinant of phase I metabolicconversion of xenobiotics, the significance of AOXs in drugmetabolism has been growing in the last few years [24].
AOXs derive their name from the well-known ability tooxidize aldehyde groups into the corresponding carboxylicacids (Figure 2A). Aldehyde functional groups are generallynot represented in common drugs; however, they are oftenintermediary products generated by alcohol dehydrogenasesor other enzymatic activities with which AOXs can act in con-cert. From a general point of view, it must be emphasized thatthe term “aldehyde oxidases” used to define the class ofenzymes representing the object of this article is misleading,as aromatic and aliphatic aldehydes are certainly not the mostprominent substrates of AOXs. In fact, a much larger set ofsubstrates that can be oxidized by AOXs very efficiently is rep-resented by aza-heterocycles and oxo-heterocycles (Figure 2B).This is the substrate subgroup of major significance in termsof drug metabolism, as aza- and oxo-heterocycles are oftenpresent in the backbone of registered drugs and drug candidates
Article highlights.
. Aldehyde oxidases are an evolutionary conserved familyof enzymes catalyzing the oxidation of a wide variety ofsubstrates. These enzymes require a molybdo-pterin cofactor (MoCo) and flavin adenine dinucleotidefor their catalytic activity and are structurally very similarto xanthine oxidoreductase, the key enzyme in thecatabolism of purines
. Along with the CYP450 system, aldehyde oxidases areprimary enzymes in the metabolism of xenobiotics ofmedical and toxicological interest. The substratesaldehyde oxidases act on are not limited to organiccompounds characterized by an aldehydic functionalgroup, which is oxidized into the correspondingcarboxylic acid. Aza- and oxo-heterocycles are a muchmore numerous group of compounds that can beefficiently oxidized by these enzymes
. The complement of aldehyde oxidase varies in differentanimal species. At the two extremes of the spectrum,humans are characterized by a single and functionallyactive aldehyde oxidase, AOX1, while mice and rats areendowed with four such enzymes. This is a majorproblem for the selection of the best experimentalmodel to be used as a proxy of the human situation indrug metabolism studies. On the basis of variousconsiderations, we propose that guinea pigs and Rhesusmonkeys may be the models of choice
. In view of the role played by aldehyde oxidase in drugmetabolism and in line with the emerging interest inpersonalized and stratified therapy, it is important totake into account the factors controlling the levels andthe functional activity of these enzymes in the liver.With respect to this, genetic polymorphisms affectingthe coding region of the AOX1 gene, sexual hormones,age and exposure to foreign compounds must beconsidered
. The emerging relevance of AOX1 in drug discovery callsfor the development of novel assays to be used for thescreening of compounds acting as substrates of thisenzyme. One of the priorities is represented by thedevelopment of effective systems for expression ofrecombinant and catalytically active human AOX1
This box summarizes key points contained in the article.
E. Garattini & M. Terao
488 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

under development. Further substrates of AOXs derived fromthe intermediary metabolism of drugs containing amine func-tional groups are iminium ions that are transformed into lac-tams (Figure 2C). Finally, there is in vitro evidence suggestingthat AOXs are endowed not only with oxidase activity,as they can act also as reductases, catalyzing the reduction ofN-oxides, sulfoxides, nitro groups and heterocycles in certainconditions.
Vanillin is an example of a primary xenobiotic containingan aldehyde group that is rapidly transformed into vanillic
acid by the AOX contained in liver cytosolic fractions [33].Vitamin B6 or pyridoxal is another example of primary andAOX-dependent drug metabolism [34]. The popular anti-estrogen, tamoxifen [35], used in the treatment of breastcancer; the antidepressant, citalopram [36], and vitamin A (ret-inol) [16,37] are biotransformed into aldehydic intermediatesthat act as specific substrates of AOXs in the liver. Prior toAOX oxidation, the two tertiary amines of tamoxifen andcytalopram are metabolized to aldehydes by monoaminooxidase and by CYP450, respectively. In contrast, retinol is
20 kDa 40 kDa
FAD
FAD
O
HN
H3C
O
HN
H3C
N
R
O
N
H3C
H3C N
O
N
R
O
N
H3C
H3C N
O
H2O2
Mo (VI)I
FAD
Mo (IV)
I
FADH2
Mo (VI).RH
I
FAD
Mo (VI)
I
FADH2.O2
ROH
RH + H2O
O2
A.
B.
I
II
Fe
Fe
SS
Fe
Fe
SS
I
II
Fe
Fe
SS
Fe
Fe
SS
85 kDa
Moco
Moco
Substrate
SubstrateO
OH
OP
Oe
OeOe
OS
S Mo
SN
N
H
H
O
OH
OP
Oe
OeOe
OS
S Mo
SN
N
H
H
Figure 1. Structure and catalytic activity of aldehyde oxidases. A. The panel illustrates a simplified model recapitulating the
homodimeric structure of aldehyde oxidase holoenzymes. Each monomeric subunit is characterized by a tripartite structure
consisting of conserved domains separated by two hinge regions. The structure of the two subunits is illustrated from the N-
to the C-terminus going from left to right. B. The panel shows the catalytic cycle of aldehyde oxidases. A generic substrate RH
is oxidized to the corresponding ROH product by the enzymes with a concomitant reduction of the molybdenum center (Mo).FAD: Flavin adenine dinucleotide; Mo: Molybdenum; Moco: Molybdenum cofactor.
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 489
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

CH
O
CO
2H
N
HO
CH
O
OH
N
HO
CO
2H
OH
ON
Me 2
O
CH
O
O
NC
FC
HO
CH
O
OH
OC
H3
O
O
NC
FC
O2H
CH
O
OH
OC
H3
AO
X1
AO
X1
AO
X1
AO
X1
AO
X1
A.
Ret
inal
Pyr
idox
alT
amox
ifen
MA
OC
YP
450
O
FN
Me 2
Cita
lopr
amV
anill
in
NC
CO
2H
Figure
2.Drugsandxenobiotics
metabolize
dbyaldehydeoxidases.
A.ThepanelillustratesthestructuresofselectedAOX
substratesch
aracterize
dbyanaldehyd
ic
function,whichisoxidizedinto
theco
rrespondingcarboxy
licacid.B.Thepanelshowsaselectednumberofaza
-heterocyclesofmedicalinterest
thatcanbeoxidizedby
AOXs.
C.Thepanelshowsselectedco
mpoundswhose
iminium
ioninterm
ediatesare
oxidizedbyAOXs.
E. Garattini & M. Terao
490 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

NN
NN
N
N NO
N
N HO
NM
e
Me
NN
O
OC
O2H
Me
NN
HN
H N
Br
N
N HO
O
NM
e
Me
NH N
O
OC
O2H
Me
O
NN
HN
H NN
Br
ONN
HN
H NN
Br
O
NO
CY
P45
0
AO
X1
AO
X1
AO
X1
AO
X1
AO
X1
AO
X1
NN
O
Nic
otin
e
DA
CA
XK
-469
Brim
onid
ine
Pro
linta
neA
zape
ptin
eB
.
N
Figure
2.Drugsandxenobiotics
metabolize
dbyaldehydeoxidases.
A(continued).ThepanelillustratesthestructuresofselectedAOXsubstratesch
aracterize
dbyan
aldehyd
icfunction,whichisoxidizedinto
theco
rrespondingcarboxy
licacid.B.Thepanelshowsaselectednumberofaza
-heterocyclesofmedicalinterest
thatcanbe
oxidizedbyAOXs.
C.Thepanelshowsselectedco
mpoundswhose
iminium
ioninterm
ediatesare
oxidizedbyAOXs.
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 491
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.
N
N
NNH
2N
H2N
NMe
H N
OC
O2H
CO
2H
N
N
NH N
NMe
H N
O
O
N
NN HN
SH
N
NN HN
SH
O
N
NN H
H
H N
SH
O
N N
O
OH
OH
HO
O
N
NN HN
O
O
Me
Me
Me
O
N
NN HN
O
O
Me
Me
Me
O
Met
hotr
exat
e6-
Mer
capt
opur
ine
Caf
fein
eZ
ebul
arin
e
AO
X1
AO
X1
AO
X1
AO
X1
XO
C.
NH
2
NH
2
CO
2H
CO
2H
HN N
O
OH
OH
HO
O
O
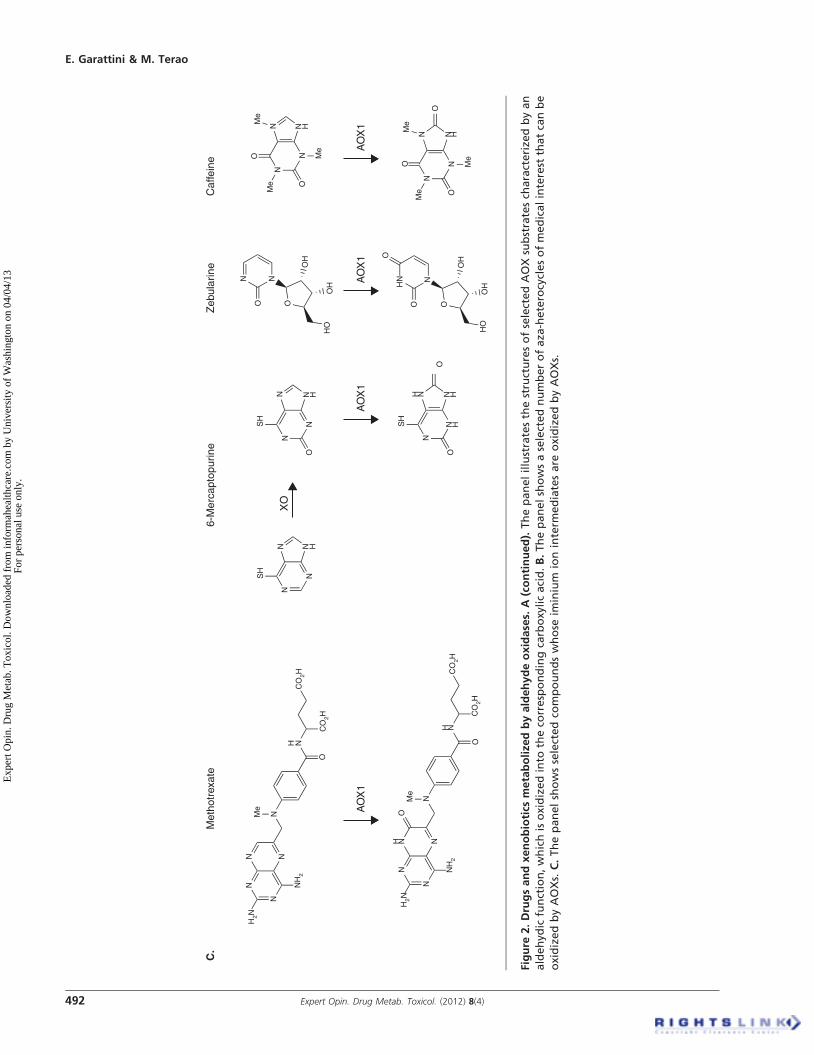
Figure
2.Drugsandxenobiotics
metabolize
dbyaldehydeoxidases.
A(continued).ThepanelillustratesthestructuresofselectedAOXsubstratesch
aracterize
dbyan
aldehyd
icfunction,whichisoxidizedinto
theco
rrespondingcarboxy
licacid.B.Thepanelshowsaselectednumberofaza
-heterocyclesofmedicalinterest
thatcanbe
oxidizedbyAOXs.
C.Thepanelshowsselectedco
mpoundswhose
iminium
ioninterm
ediatesare
oxidizedbyAOXs.
E. Garattini & M. Terao
492 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

biotransformed into retinal by an alcohol dehydrogenasebefore oxidation into retinoic acid by AOXs.
AOX drug substrates characterized by aza- or oxo-heterocyclic structures vastly outnumber those endowed withaldehyde functionalities. Phthalazines, quinazolines and dia-minopteridines are among some of the structures that can beoxidized by AOXs [38,39]. Methotrexate is a well-knownimmunosuppressant and antitumor agent belonging to thelast group of compounds. Methotrexate is oxidized to a7-hydroxylated derivative in humans and primates, but to amuch lesser extent in rodents [40]. As we will see, this is inline with the different complement of AOXs present inhumans and certain primates relative to mice and rats.AOXs have been documented to oxidize purine and pyrimi-dine analogs. Some of these are specific substrates for AOXswhile others are common to both AOXs and XORs. The anti-leukemia agent 6-mercaptopurine is metabolized by XORinto 6-thioxanthine. The resulting compound is further trans-formed by either AOX or XOR into 6-thiouric acid [31]. Bythe same token, caffeine is oxidized by AOX and XOR [41].These data indicate that AOX and XOR can compete forthe same type of substrates, although the cross-talk betweenthe two enzymes may be more complex and give rise to unpre-dicted types of interactions. A relevant case is represented byallopurinol, a well-known and selective inhibitor of XOR,which can be oxidized to oxypurinol by AOX, potentiallyaltering its ability to inhibit XOR in vivo [42]. Other examplesof aza-heterocycles of pharmacological interest that can beoxidized and inactivated by AOXs are zebularine, a pyrimidi-none nucleoside modulating DNA methylation [43], theacridine carboxamide, N-[2-(dimethylamino)-ethyl]acridine-4-carboxamide [44], and XK-469 [45], two experimental anti-neoplastic agents as well as brimonidine, an alpha-2 adrenergicagonist used in the treatment of open-angle glaucoma [46]. Allthe drugs or experimental agents with an aza-heterocyclic struc-ture mentioned above are representative of active compoundsmetabolized to inactive derivatives by AOX. However, the sci-entific literature also contains a few examples of pro-drugsselectively activated by AOX. AOX-dependent metabolism ofappropriate pro-drugs is a strategy that has been proposed tosolve the problems of bioavailability associated with the use of5-ethynyluracil and 5-fluorouracil [47,48].
Iminium ions are metabolic intermediates generated duringthe oxidation of cyclic amines, such as pyrrolidines and piper-idines, by MAO (monoamine oxidase) and CYP450. Theseintermediates can be good substrates of AOXs and further oxi-dized to the corresponding cyclic lactames [24]. Nicotine istransformed into a nicotine iminium ion by CYP450 that isfurther metabolized to cotinine by AOX [49]. Prolinatane, anappetite-stimulating agent and azapetine, an antihypertensivedrug, are biotransformed into inactive oxoprolintane [50] andoxazapetine [51], respectively, via similar coupled reactionsinvolving CYP450 and AOX.
Evidence present in the literature indicates that AOXs canact as reductases in certain conditions, catalyzing the
reduction of N-oxides, sulfoxides, nitro-compounds and het-erocycles. AOXs from different sources have been reportedto reduce nicotinamide N-oxide and imipramine N-oxideinto nicotinamide and imipramine [52,53]. The nonsteroidalanti-inflammatory sulfoxide, sulindac, can be reduced byAOX in the presence of different small molecules as electrondonors [53]. A series of nitroguanidine and nitromethyleneinsecticides are reduced by AOX to the corresponding nitrosoor amino derivatives [54]. Reduction of the antipsychotic drug,ziprasidone, is an example of AOX-dependent reduction of aheterocycle [21]. However the relevance of AOX reductaseactivity in vivo is questionable, as it occurs in vitro in the pres-ence of small molecules acting as electron donors or underanaerobic conditions with the drug substrate taking the placeof oxygen.
In addition to its relevance in the biotransformation of theregistered or developed drugs mentioned, there is increasingevidence for the role of AOXs in the metabolic inactivationand clearance of new drug candidates, with important impli-cations in the fields of drug discovery and development [1,24].The emerging significance of AOXs in drug discovery is likelyto be the consequence of the successful strategies put in placein the design and synthesis of new drug candidates that are poorsubstrates for the CYP450 system, as discussed extensively byPryde et al. in a recent review article [24]. In this article, theauthors report very interesting data on the prediction of AOXsubstrates present in the available collections of chemical com-pounds. The proportion of potential AOX substrates amongcompounds which have already progressed to the market is cal-culated to be 0.13. This proportion increases dramatically to0.45 for compounds currently under development. All this hasstirred interest in the development of drug design and synthesisstrategies aimed at limiting the chances that new molecules aremetabolized and inactivated by human AOX1. An example ofthis trend is the publication of an article [55] addressing the prob-lem with derivatives of N-[63-tetramethylcyclobutyl]imidazo[1,2-a]pyrimidine-3-carboxamide, a recently identified antago-nist of the androgen receptor. The approach is guided by anAOX1 structure-based model and reveals that either alteringthe heterocycle or blocking the reactive site is the more effectivestrategy for reducing AOX-dependent metabolism.
3. The number of active AOX genes variesin different animal species: implicationsfor drug metabolism studies
The available evidence indicates that AOXs evolved from anancestral XOR precursor gene via a primordial dupli-cation event. This was followed by a series of other duplica-tions, resulting in a number of AOX genes which variesaccording to the animal species considered. Fishes (such asTakifugu rubripes or Tetraodon nigroviridis) and amphibians(Xenopus laevis) are characterized by a single AOX gene, whilebirds (Gallus gallus) are endowed with two such genes. The
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 493
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

genome of rodents contains four AOXs, which is the largestnumber of active AOX genes so far described. The four mousegenes, Aox1, Aox3, Aox4 and Aox3l1 cluster, in this order, on ashort region of chromosome 1c1 with a head to tailconfiguration [56-58]. The available evolutionary data indicatethat the extant complement of rodent AOXs is the result ofasynchronous and independent gene duplication eventsdriven by the necessity to acquire an increasing number offunctional AOX isoenzymes characterized by different profilesof tissue- and cell-specific expression. Indeed, AOX1 andAOX3 are expressed predominantly in liver and lungs [56,59].The largest amounts of AOX4 are present in the Harderiangland, a major exocrine gland located in the orbital cavity,though detectable levels of the protein are also present in thekeratinized epithelia of the skin, oral cavity and esopha-gus [16,56]. Finally, AOX3l1 expression is restricted to the nasalmucosa predominantly in Bowman glands [58].The acquisition of new isoenzymatic forms of AOXs must
have stopped with the evolution of rodents, as the develop-ment of other mammals, including primates and humans ischaracterized by a process of progressive and species-specific gene suppression via deletion events or the generationof pseudogenes [2,3]. In humans, this process of gene suppres-sion has resulted in the maintenance of a single active gene,which is clearly the orthologue of mouse AOX1 [13]. HumanAOX1 is expressed in a rather ubiquitous fashion, althoughthe highest levels of the enzyme are measurable in liver,adrenal glands, lungs and pancreas [1,2].The variable complement of AOX isoenzymatic forms
observed in different animal species has important implica-tions for drug metabolism studies. In fact, it raises the ques-tion as to which is the best in vivo preclinical model to beused as a proxy of the human situation for studies involvingxenobiotics which are known or suspected to be AOX sub-strates. On the basis of the data present in the literature [2]
and an analysis of the genomes for which complete or almostcomplete sequencing data are available (EG and MT unpub-lished results), we can draw the following conclusions(Table 1). Some of the most common strains of mice andrats used for drug metabolism studies do not recapitulatethe human situation. The livers of inbred C57BL/6J or out-bred CD1 mice and Sprague-Dawley rats are characterizedby the expression of both AOX1 and AOX3 [58,60]. In addi-tion, AOX3 is the prevalent enzyme expressed in this organ.Hence, the metabolic profile of human AOX1 substrates isbound to be different in rodent liver both from a qualitativeand a quantitative standpoint. A pertinent and recent examplein support of this contention is represented by the large differen-ces in the metabolism and pharmacokinetics of the experimentalcompound and adenosine inhibitor, 6-(2-amino-4-phenylpyri-midine-5-yl)-2-isopropylpyridazin-3(2H)-one (FK3453), in ratsand humans [61]. Since rabbits are characterized by the presenceof the same complement of AOX genes as mice and rats (Table 1)and are predicted to synthesize AOX1 and AOX3 in their livers,they are also unlikely to be good proxies of the human situation.
Other experimental animals of potential interest in the field ofdrug metabolism, like dogs, cats and pigs do not recapitulatethe human complement of AOX genes either (Table 1). In fact,the cytosolic fractions of dogs and cats [20,62-66] are devoid ofhepatic AOX activity due to inactivation of the AOX1 geneand deletion/suppression of the AOX3 counterpart [66]. The sit-uation in pigs is more complicated, as our genome analysis pre-dicts the presence of inactive AOX1 and AOX3 pseudogenes inthis animal species, suggesting an absence of hepatic AOXactivity. However, a single report indicates that porcine livercytosolic fractions contain detectable levels of AOX activity,which was shown to be the main determinant of 3-methylindolemetabolism [67].
On the basis of the available data, we propose that guineapigs and Rhesus monkeys are likely to represent the experi-mental models of choice to perform in vivo pharmacokineticstudies of drugs or compounds known to be AOX substrates.Indeed, we cloned a single AOX cDNA from the liver of bothorganisms (EG and MT, unpublished results). Sequencing ofthe guinea pig and Rhesus monkey cDNAs demonstrated thatthey are the products of the human and mouse AOX1 orthol-ogous gene. Thus, the widespread use of guinea pig cytosolicextracts to study AOX-dependent drug metabolism is likely toprovide results predictive of the human situation [27-31,68-79].The same predictive value is associated with studies involvingRhesus monkey liver preparations [65,80-84]. These two modelsrepresent the best proxy of the human situation when systemicadministration of the compound under study and liver metab-olism are the objects of interest. We propose that a more cau-tious attitude in transferring the results from guinea pigs orRhesus monkeys to humans should be exercised when dealingwith AOX substrates to be administered topically via theintra-ocular (e.g., brimonidine) or the intra-nasal route. Infact, guinea pigs are characterized by the presence of Harderianglands, containing large amounts of AOX4. These exocrineglands are not present in primates or humans, have direct accessto the conjunctiva and are likely to be exposed to drugs instilledto the eye, resulting in metabolic transformation via AOX4activity. By the same token, drugs administered intra-nasallymay be metabolized by AOX3l1 in the nasal mucosa of guineapigs or Rhesus monkey but not in the same structures of thehuman nasal cavities. Finally, we surmise that an alternativeto the use of guinea pigs and Rhesus monkeys is representedby certain strains of mice, like DBA2 and CBA. These arenot popular in drug metabolism studies, but are characterizedby epigenetic suppression of Aox3 in the liver [68] and Aox4gene expression in the Harderian glands and skin [18]. Compar-ative studies using human and DBA2/CBA mouse cytosolicextracts or isolated hepatocytes with bona fide AOX1 substratesare necessary to support our contention.
4. Factors influencing the activity of AOXs
Given their role in the metabolism of xenobiotics, there isinterest in gaining information on the factors influencing
E. Garattini & M. Terao
494 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

the activity of AOX1 in humans, as this is of practical impor-tance in predicting individual responses to drugs that areknown to be metabolized by this enzyme. With respect tothis, it has long been known that the levels of AOX1 activityin the human population control inter-individual variabilityin the hydroxylation and inactivation of methotrexate [81].
A major source of inter-individual variability in the humanpopulation is represented by the presence of allelic variants ofthe AOX1 gene coding for proteins with increased or decreasedcatalytic activity. Missense single nucleotide polymorphisms(SNPs) in the coding region of the AOX1 gene may affectthe catalytic function of the enzyme both in a negative andpositive fashion. Non-sense nucleotide mutations in the het-erozygous state will decrease AOX1 activity by 50%, whilesuch mutations in the homozygous state delete it. As illus-trated in Table 2, numerous SNPs of the human AOX1 geneare available in the NCBI dbSNP database although for themajority of them allelic frequency data are not available. Theonly evidence available on the frequency of a selected numberof SNPs is present in a study performed on the ChurchillCounty of Nevada [85]. In addition, functional data on theinfluence that SNPs exert on the enzymatic activity of humanAOX1 have not yet been published. Indeed, the only resultsavailable on the effects exerted by amino acid mutations onthe catalytic activity of AOX1 were obtained in rats [86-89].To overcome these gaps in knowledge, we recently identifiedrelatively frequent SNPs in the coding region of the AOX1gene using a cohort of 180 volunteers, representative of theItalian population (Hartmann T, Terao M, Garattini E,Teutloff C, Alfaro JF, Jones JP and Leimkuhler S. The impactof Single Nucleotide Polymorphisms on human AldehydeOxidase. Drug Metabolism and Disposition, in press). Inthis population, we identified one non-sense mutation; onesynonymous and five non-synonymous SNPs. The non-sensemutation is located in exon 5 and predicts a short and inactiveprotein. This mutation has a relatively high incidence (calcu-lated allelic frequency, 2.6%) in the Italian population and isat the basis of a haplo-insufficient phenotype, being observed
only as a heterozygous trait. The most frequent missensemutation identified is a G/A substitution resulting in thepresence of an Arg instead of a Lys residue at position1297 (R1297K). The SNP maps to exon 34 and the trait ishomozygous in 6 out of the 13 positive individuals (calculatedallelic frequency, 5.3%). Exon 34 is likely to represent a muta-tional hotspot, as another relatively frequent SNP (T/C)resulting in the substitution of Leu for a Ser at position1271 (L1271S), and a synonymous SNP are located in thisexon. All the individuals carrying this last SNP are hetero-zygous for the trait and the calculated allelic frequency is3.9%. A relatively frequent missense SNP corresponding toan A to G transition, resulting in the substitution of Asnwith a Ser residue at position 1135 is observed in exon 30.Two further and rare non-synonymous SNPs map to exons22 and 25. They are the result of C-T (R802C) (calculatedallelic frequency, 0.6%) and G-A (R921H) (calculated allelicfrequency, 0.3%) transitions, respectively. Using a recentlydeveloped system for the expression of recombinant AOXsin E. coli, we concluded that individuals characterized by theR802C and the R921H variants may be considered poormetabolizers, since both variants are characterized by reducedcatalytic activity. In contrast, individuals with an N1135S andan H1297R genotype can be classified as fast metabolizers,since the two variants show high catalytic activity with all thetested substrates. Clearly further studies aimed at describingthe frequency and geographical distribution of all the SNPsso far identified and their significance for the catalytic activityof AOX1 are necessary. This last type of information will becrucial to classify human individuals into fast and slow drugmetabolizers not only according to the CYP450 but alsoaccording to the AOX1 genotype.
A second determinant of variability in terms of AOX1-de-pendent drug metabolism is likely to be represented by gen-der, although studies aimed at determining the influence ofsex and particularly sexual steroid hormones on humanAOX1 expression and functional activity are not available.This contention is supported by the data available in rodents.
Table 1. The table lists the complement of functionally active aldehyde oxidases expressed in the liver and other
tissues of humans, chimpanzees and popular animal experimental models.
Animal species Liver isoenzyme(s) Other isoenzyme(s) Pseudogenes
Human AOX1 - AOX3, AOX3L1Chimpanzee AOX1 - AOX3, AOX3L1Rhesus monkey AOX1 AOX3l1 AOX3, AOX4Guinea pig AOX1 AOX4, AOX3l1 -Dog - AOX4, AOX3l1 AOX1, AOX3Cat - AOX3l1 AOX1, AOX4Pig - AOX3L1 AOX1, AOX3, AOX4Mouse AOX3, AOX1 AOX4, AOX3l1 -Rat AOX3, AOX1 AOX4, AOX3l1 -Rabbit AOX3, AOX1 AOX4, AOX3l1 -
The number and type of inactive aldehyde oxidase pseudogenes are also indicated. Please note that the data in mice and rats refer to the C57BL/6J and Wistar
strains, since variability of AOX isoenzyme expression has been observed in other strains, as discussed in the article.
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 495
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.
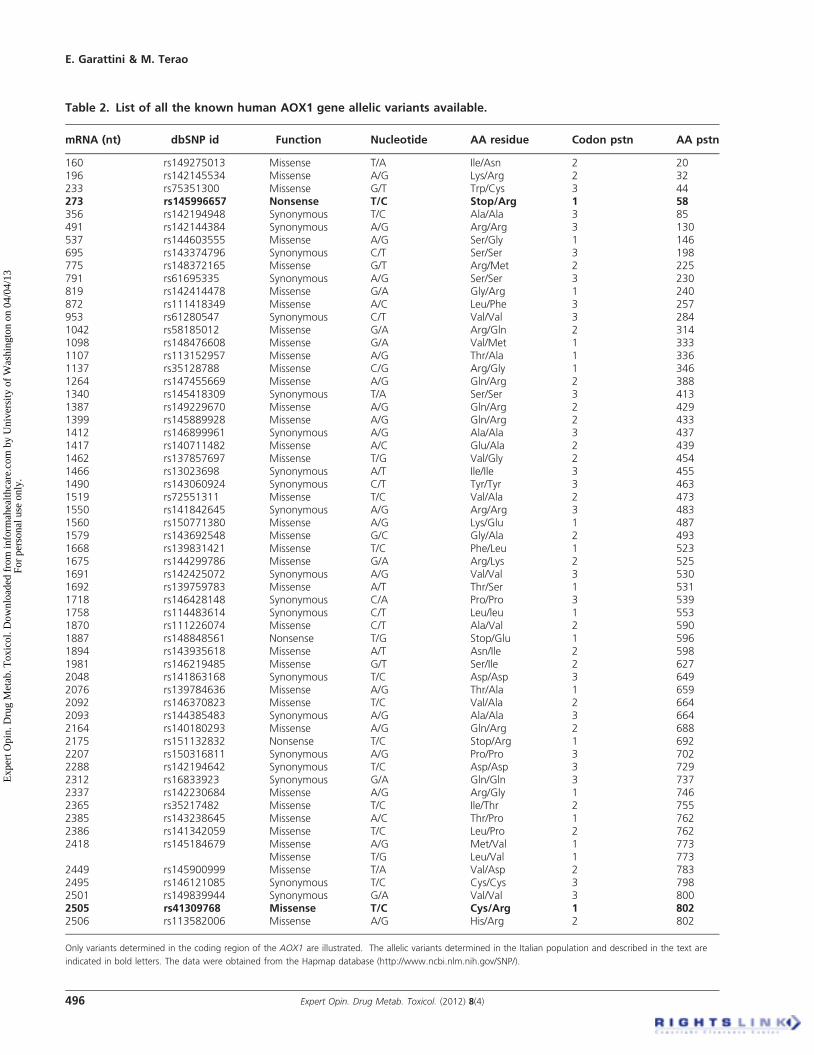
Table 2. List of all the known human AOX1 gene allelic variants available.
mRNA (nt) dbSNP id Function Nucleotide AA residue Codon pstn AA pstn
160 rs149275013 Missense T/A Ile/Asn 2 20196 rs142145534 Missense A/G Lys/Arg 2 32233 rs75351300 Missense G/T Trp/Cys 3 44273 rs145996657 Nonsense T/C Stop/Arg 1 58356 rs142194948 Synonymous T/C Ala/Ala 3 85491 rs142144384 Synonymous A/G Arg/Arg 3 130537 rs144603555 Missense A/G Ser/Gly 1 146695 rs143374796 Synonymous C/T Ser/Ser 3 198775 rs148372165 Missense G/T Arg/Met 2 225791 rs61695335 Synonymous A/G Ser/Ser 3 230819 rs142414478 Missense G/A Gly/Arg 1 240872 rs111418349 Missense A/C Leu/Phe 3 257953 rs61280547 Synonymous C/T Val/Val 3 2841042 rs58185012 Missense G/A Arg/Gln 2 3141098 rs148476608 Missense G/A Val/Met 1 3331107 rs113152957 Missense A/G Thr/Ala 1 3361137 rs35128788 Missense C/G Arg/Gly 1 3461264 rs147455669 Missense A/G Gln/Arg 2 3881340 rs145418309 Synonymous T/A Ser/Ser 3 4131387 rs149229670 Missense A/G Gln/Arg 2 4291399 rs145889928 Missense A/G Gln/Arg 2 4331412 rs146899961 Synonymous A/G Ala/Ala 3 4371417 rs140711482 Missense A/C Glu/Ala 2 4391462 rs137857697 Missense T/G Val/Gly 2 4541466 rs13023698 Synonymous A/T Ile/Ile 3 4551490 rs143060924 Synonymous C/T Tyr/Tyr 3 4631519 rs72551311 Missense T/C Val/Ala 2 4731550 rs141842645 Synonymous A/G Arg/Arg 3 4831560 rs150771380 Missense A/G Lys/Glu 1 4871579 rs143692548 Missense G/C Gly/Ala 2 4931668 rs139831421 Missense T/C Phe/Leu 1 5231675 rs144299786 Missense G/A Arg/Lys 2 5251691 rs142425072 Synonymous A/G Val/Val 3 5301692 rs139759783 Missense A/T Thr/Ser 1 5311718 rs146428148 Synonymous C/A Pro/Pro 3 5391758 rs114483614 Synonymous C/T Leu/leu 1 5531870 rs111226074 Missense C/T Ala/Val 2 5901887 rs148848561 Nonsense T/G Stop/Glu 1 5961894 rs143935618 Missense A/T Asn/Ile 2 5981981 rs146219485 Missense G/T Ser/Ile 2 6272048 rs141863168 Synonymous T/C Asp/Asp 3 6492076 rs139784636 Missense A/G Thr/Ala 1 6592092 rs146370823 Missense T/C Val/Ala 2 6642093 rs144385483 Synonymous A/G Ala/Ala 3 6642164 rs140180293 Missense A/G Gln/Arg 2 6882175 rs151132832 Nonsense T/C Stop/Arg 1 6922207 rs150316811 Synonymous A/G Pro/Pro 3 7022288 rs142194642 Synonymous T/C Asp/Asp 3 7292312 rs16833923 Synonymous G/A Gln/Gln 3 7372337 rs142230684 Missense A/G Arg/Gly 1 7462365 rs35217482 Missense T/C Ile/Thr 2 7552385 rs143238645 Missense A/C Thr/Pro 1 7622386 rs141342059 Missense T/C Leu/Pro 2 7622418 rs145184679 Missense A/G Met/Val 1 773
Missense T/G Leu/Val 1 7732449 rs145900999 Missense T/A Val/Asp 2 7832495 rs146121085 Synonymous T/C Cys/Cys 3 7982501 rs149839944 Synonymous G/A Val/Val 3 8002505 rs41309768 Missense T/C Cys/Arg 1 8022506 rs113582006 Missense A/G His/Arg 2 802
Only variants determined in the coding region of the AOX1 are illustrated. The allelic variants determined in the Italian population and described in the text are
indicated in bold letters. The data were obtained from the Hapmap database (http://www.ncbi.nlm.nih.gov/SNP/).
E. Garattini & M. Terao
496 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.
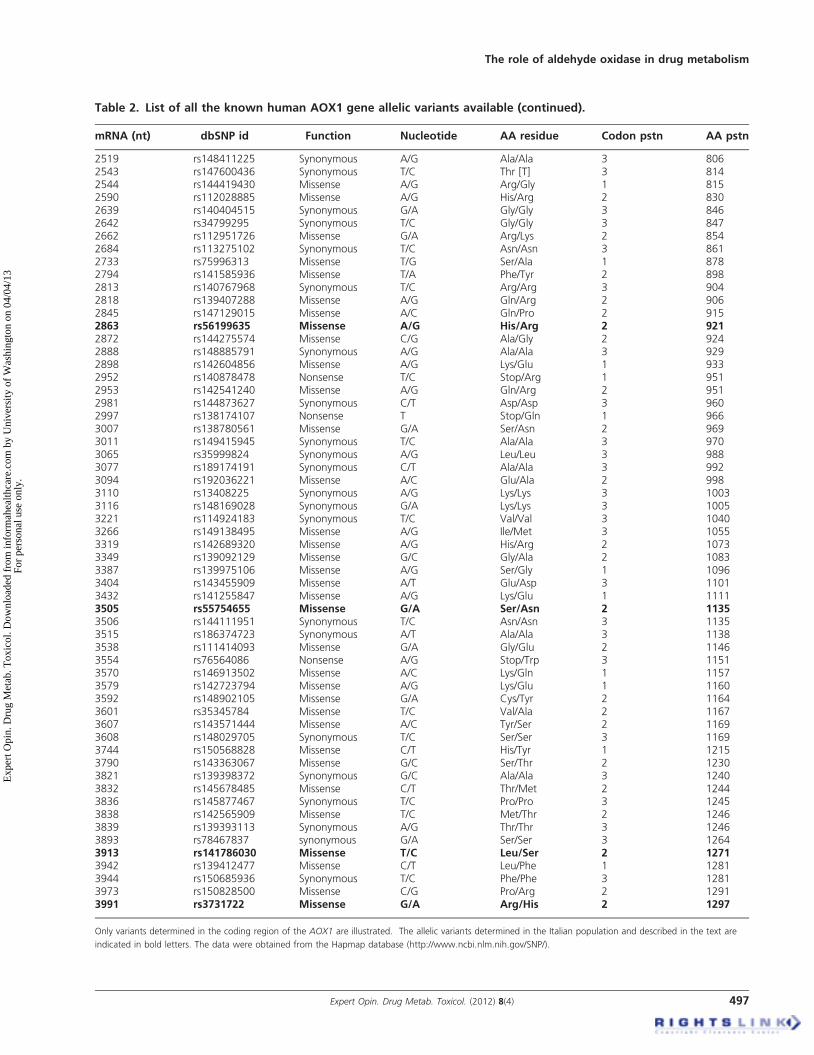
Table 2. List of all the known human AOX1 gene allelic variants available (continued).
mRNA (nt) dbSNP id Function Nucleotide AA residue Codon pstn AA pstn
2519 rs148411225 Synonymous A/G Ala/Ala 3 8062543 rs147600436 Synonymous T/C Thr [T] 3 8142544 rs144419430 Missense A/G Arg/Gly 1 8152590 rs112028885 Missense A/G His/Arg 2 8302639 rs140404515 Synonymous G/A Gly/Gly 3 8462642 rs34799295 Synonymous T/C Gly/Gly 3 8472662 rs112951726 Missense G/A Arg/Lys 2 8542684 rs113275102 Synonymous T/C Asn/Asn 3 8612733 rs75996313 Missense T/G Ser/Ala 1 8782794 rs141585936 Missense T/A Phe/Tyr 2 8982813 rs140767968 Synonymous T/C Arg/Arg 3 9042818 rs139407288 Missense A/G Gln/Arg 2 9062845 rs147129015 Missense A/C Gln/Pro 2 9152863 rs56199635 Missense A/G His/Arg 2 9212872 rs144275574 Missense C/G Ala/Gly 2 9242888 rs148885791 Synonymous A/G Ala/Ala 3 9292898 rs142604856 Missense A/G Lys/Glu 1 9332952 rs140878478 Nonsense T/C Stop/Arg 1 9512953 rs142541240 Missense A/G Gln/Arg 2 9512981 rs144873627 Synonymous C/T Asp/Asp 3 9602997 rs138174107 Nonsense T Stop/Gln 1 9663007 rs138780561 Missense G/A Ser/Asn 2 9693011 rs149415945 Synonymous T/C Ala/Ala 3 9703065 rs35999824 Synonymous A/G Leu/Leu 3 9883077 rs189174191 Synonymous C/T Ala/Ala 3 9923094 rs192036221 Missense A/C Glu/Ala 2 9983110 rs13408225 Synonymous A/G Lys/Lys 3 10033116 rs148169028 Synonymous G/A Lys/Lys 3 10053221 rs114924183 Synonymous T/C Val/Val 3 10403266 rs149138495 Missense A/G Ile/Met 3 10553319 rs142689320 Missense A/G His/Arg 2 10733349 rs139092129 Missense G/C Gly/Ala 2 10833387 rs139975106 Missense A/G Ser/Gly 1 10963404 rs143455909 Missense A/T Glu/Asp 3 11013432 rs141255847 Missense A/G Lys/Glu 1 11113505 rs55754655 Missense G/A Ser/Asn 2 11353506 rs144111951 Synonymous T/C Asn/Asn 3 11353515 rs186374723 Synonymous A/T Ala/Ala 3 11383538 rs111414093 Missense G/A Gly/Glu 2 11463554 rs76564086 Nonsense A/G Stop/Trp 3 11513570 rs146913502 Missense A/C Lys/Gln 1 11573579 rs142723794 Missense A/G Lys/Glu 1 11603592 rs148902105 Missense G/A Cys/Tyr 2 11643601 rs35345784 Missense T/C Val/Ala 2 11673607 rs143571444 Missense A/C Tyr/Ser 2 11693608 rs148029705 Synonymous T/C Ser/Ser 3 11693744 rs150568828 Missense C/T His/Tyr 1 12153790 rs143363067 Missense G/C Ser/Thr 2 12303821 rs139398372 Synonymous G/C Ala/Ala 3 12403832 rs145678485 Missense C/T Thr/Met 2 12443836 rs145877467 Synonymous T/C Pro/Pro 3 12453838 rs142565909 Missense T/C Met/Thr 2 12463839 rs139393113 Synonymous A/G Thr/Thr 3 12463893 rs78467837 synonymous G/A Ser/Ser 3 12643913 rs141786030 Missense T/C Leu/Ser 2 12713942 rs139412477 Missense C/T Leu/Phe 1 12813944 rs150685936 Synonymous T/C Phe/Phe 3 12813973 rs150828500 Missense C/G Pro/Arg 2 12913991 rs3731722 Missense G/A Arg/His 2 1297
Only variants determined in the coding region of the AOX1 are illustrated. The allelic variants determined in the Italian population and described in the text are
indicated in bold letters. The data were obtained from the Hapmap database (http://www.ncbi.nlm.nih.gov/SNP/).
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 497
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

In fact, male mice exhibit a 2 -- 4 fold higher liver AOX activ-ity than female mice [2]. This is likely to be due to the fact thatandrogens induce the expression of the mouse Aox1 and Aox3genes and corresponding proteins, while estrogens exert theopposite effect. An example of the potential relevance of sex-dependent differences in AOX1 expression is represented bythe results obtained with zebularine. In fact, the chemo-preventive activity of zebularine in experimental models ofintestinal polyposis is higher in female than maleanimals [43,90].Age, disease states, such as cancer, and drug usage may also
account for the inter-individual variability of humanAOX1 activity. The only information available on the influ-ence exerted by aging on AOXs relates to the expression ofthese enzymatic activities during development of youngorganisms. The level of hepatic AOX is undetectable in thedeveloping rat fetus, as assessed by measuring the oxidationof benzaldehyde to benzoic acid, N-1-methylnicotinamideto N-1-methyl-2-pyridone-5-carboxamide or N-1-methyl-4-pyridone-3-carboxamide, and methotrexate (MTX) to7-hydroxymethotrexate [91]. These oxidase activities aremarkedly increased soon after birth and maintained up to6 weeks. The results obtained in rats seem to recapitulatethe developmental profile of expression determined in acohort of Japanese children with the use of a non-invasive method measuring AOX-dependent metabolitesexcreted in the urine [92]. In this study, AOX activity wasassessed as the ratio of pyridone formation from N(1)-methyl-nicotinamide in 101 children. The findings indicate thatAOX activity begins to increase after birth and is relativelylow in children below 1 year of age as compared with adultindividuals. This has important implications for the adminis-tration of drugs with an AOX1-dependent metabolism to thepediatric population and it emphasizes the importance of doseadjustment in babies and young children to avoid unpredictedtoxicity and side effects.An extensive review of the human AOX1 expression profile
in different types of cancers and the potential relevance ofthese data for the activity of known antitumor agents is avail-able in a recent review article [1]. Suffice it to say that there arevery few human tumors, such as glioblastoma and breast can-cer, where high expression of the AOX1 gene has beenreported. AOX1 overexpression in these types of cancer couldbe exploited for the in situ activation of antineoplastic pro-drugs, like the aforementioned 5-fluoro-2-pyrimidone, toobtain an increased therapeutic index via organ selectivity inthe anticancer action. Cancers characterized by low expressionof AOX1 vastly outnumber those showing overexpression ofthe enzyme. The observation suggests that these tumors mayrepresent good targets for antitumor agents whose primarymetabolic pattern is inactivation by AOX1, as enrichment ofthe active drug in the neoplastic cell is expected. This is partic-ularly true in the case of hepatocellular carcinomas in whichthe predicted levels of AOX1 enzymatic activity are wellbelow those observed in normal hepatocytes. To the best of
our knowledge, there are no studies evaluating the effects ofcirculating agents produced by tumors on the activity of liverAOX1 and this should be an area of interest in thenear future.
Exposure to other drugs or environmental pollutants is thefinal determinant of AOX1 activity to be considered whenadministering drugs extensively metabolized by this enzymaticsystem. There is indeed a wide variety of compounds inhibit-ing AOX1 activity. Such diverse compounds as amsacrine [93],chlorpromazine [22], cimetidine [94], estradiol [95], menadi-one [33], methadone [96] and quinacrine [22] have been reportedto act as efficient inhibitors of AOX1. With respect to this,the drug endowed with the strongest inhibitory activity is rep-resented by the SERM (selective estrogen receptor modula-tor), raloxifene, which is used in the treatment ofosteoporosis [22]. Very little information is available on xeno-biotics inducing the expression or modulating the activity ofAOX1 in a positive fashion. Administration of the alkylatingagents, N-methyl-N’-nitro-N-nitrosoguanidine, in rats, expo-sure to phthalazine in rabbits, as well as dioxin in mice resultsin the induction of AOX1 [97-99]. In addition, fenofibrate, aPPARa agonist used in the treatment of obesity reduces thelevels of rat and human liver AOX1 [19]. Although the dataobtained in animals cannot be directly transferred to thehuman situation, they suggest that inhibition or inductionof AOX1 by foreign chemicals may be at the basis ofunexpected drug/drug interactions.
5. Conclusion
AOX1 is a cytosolic enzyme of increasing interest in the fieldof drug metabolism because of its broad substrate specificityand its high levels of expression in the liver. Although thereis no question that CYP450 is currently the enzymatic systemof major importance for phase I drug metabolism, it is pre-dicted that, in the near future, AOX1 will compete withCYP450 in terms of relevance for the primary metabolic acti-vation/inactivation of certain structural classes of drugs andtoxicants. This prediction stems from the observation thatcurrent synthetic efforts in medicinal chemistry are aimed atminimizing CYP450 involvement in drug clearance with aconsequent enrichment for chemical scaffolds that act asAOX substrates [19]. The emerging importance ofAOX1 will require research efforts in multiple directions.A priority is represented by the development of simplein vitro assays based on the use of the catalytically activerecombinant form of human AOX1. Progress in the directionof developing E. coli bacteria expressing catalytically activeAOXs has been made in the past few years [100,101]. Althoughthese systems are still unsatisfactory for a number ofreasons, they are essential for screening programs aimed atdefining the potential of AOXs to metabolize new drug candi-dates. This must be accompanied by a better definition of themost reliable animal models to be used as proxy of the humansituation in ADME (Absorption, Distribution, Metabolism
E. Garattini & M. Terao
498 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

and Excretion) studies focusing on the role of AOX1.Humanization of mouse or rat models with the use of geneticengineering or other techniques, such as xenografting ofhuman hepatocytes is also a research direction to be pursued.In addition, the current focus on personalized and stratifiedtherapy calls for further studies aimed at the identificationand functional characterization of new AOX1 allelic variants.Finally, it is necessary to increase our knowledge of the endog-enous and exogenous factors regulating the levels and theactivity of human AOX1. This is fundamental to avoid prob-lems associated with drug--drug interactions and to allowappropriate individual dose adjustments for drugs known tobe metabolized by AOX1.
6. Expert opinion
Aldehyde oxidases (AOXs) are a small family of evolutionaryconserved enzymes belonging to the family of molybdo-flavoproteins along with xanthine oxidoreductase, the keyenzyme in the catabolism of purines. AOXs require FADand a molybdo-pterin (MoCo) as cofactors for their catalyticactivity. The complement of AOXs present in different mam-mals varies from one in humans to four in rodents. This is theresult of an evolutionary process involving gene duplicationand gene suppression events. AOXs catalyze the oxidation ofnumerous endogenous and exogenous compounds, as theseenzymes are characterized by broad substrate specificity.AOXs oxidize not only organic molecules endowed with alde-hyde functional groups into the corresponding carboxylicacid, but also aza- and oxo-heterocycles. AOX1, the onlyfunctionally active human aldehyde oxidase is expressed inthe liver at very high levels, although detectable amounts ofthe enzyme are present also in other tissues such as the adrenalgland, lungs, small intestine and pancreas. The physiologicfunction(s) and substrate(s) of human AOX1 have not yetbeen identified, nevertheless, the enzyme is known to play akey role in the metabolism of various compounds of medicalas well as toxicological interest, due to its broad substratespecificity and localization in the hepatic tissue. IndeedAOX1 represents the major phase I drug metabolizing enzymepresent in the cytosol of the hepatocyte, often acting in con-cert with endoplasmic CYP450 on intermediary metabolites.The role of AOX1 as a drug metabolizing enzyme is likelyto increase as the consequence of successful strategies put inplace in the design and synthesis of new drug candidatesthat are poor substrates for the CYP450 system. This trendis likely to select for chemical structures acting as AOX sub-strates in the new generation of drugs under development [24].Thus, it is expected that the relevance of AOX1 in the realmsof drug discovery and drug metabolism will increase in thenear future.
The emerging role of AOX1 in pharmacology calls for thedevelopment of new and appropriate experimental paradigmsto screen for chemicals that are metabolized by this enzymaticsystem. A prerequisite for these types of studies is the
availability of rapid screening tests. At present there is a gen-eral lack of in vitro assays based on the use of the recombinanthuman AOX1 protein. In fact, the production of catalyticallyactive recombinant molybdo-flavoenzymes in prokaryotes hasproven difficult because of the problems associated with theassembly of MoCo into the holoenzyme. This is largely dueto the complexity of the biochemical pathways leading tothe production of MoCo and to the differences in these met-abolic pathways between eukaryotes and prokaryotes [1,4]. Therecent development of novel and promising prokaryotic sys-tems for the expression of active mammalian AOXs is animportant step in this direction [100,101]. Liver cytosolic frac-tions or whole hepatocytes are other viable options. However,because of the problems discussed in the present article, thechoice of these experimental paradigms has been so far limitedto human preparations, with all the associated problems ofavailability and AOX1 enzyme stability [1]. The demonstra-tion that the livers of guinea pigs (EG and MT, manuscriptin preparation) and certain mouse strains [16,60] express a sin-gle AOX1 gene like humans will open up new opportunities.The development of new drugs requires pharmacokineticand pharmacodynamic studies in appropriate preclinicalmodels recapitulating the human situation also in terms ofAOX expression. The evidence available indicates that Rhesusmonkeys, guinea pigs and certain strains of mice are currentlythe best proxy of the human situation. However, these modelsare not yet completely satisfactory. Future efforts must bedevoted to the generation of better animal models. This canbe accomplished by genetic engineering of mice that mayinvolve generation of animals expressing a human AOX1transgene on an AOX knock-out background. Humanizationof the mouse and rat liver by the transplant of human hepato-cytes in immunodepressed animals is another option. Finally,the current emphasis on personalized and stratified therapywill require further efforts aimed at the definition of theimpact that allelic variants exert on the AOX1-dependentmetabolism of drugs.
Acknowledgements
The authors thank N Fisher for the critical reading of themanuscript and F Deceglie and A Soave for their assistancewith the artwork.
Declaration of interest
The authors acknowledge the financial support of theFondazione “Italo Monzino,” the Associazione Italiana perla Ricerca contro il Cancro (AIRC) and the Weizmann-Negri Foundation E Garattini and M Terao are membersof the permanent scientific staff of the Istituto di RicercheFarmacologiche “Mario Negri,” a private non-profit researchinstitute. The two authors have no conflict of interestto declare.
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 499
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

BibliographyPapers of special note have been highlighted as
either of interest (�) or of considerable interest(��) to readers.
1. Garattini E, Terao M. Increasing
recognition of the importance of
aldehyde oxidase in drug development
and discovery. Drug Metab Rev
2011;43:374-86. The review article highlights the
problems associated in drug
metabolism studies focussing on
aldehyde oxidases.
2. Garattini E, Fratelli M, Terao M.
Mammalian aldehyde oxidases: genetics,
evolution and biochemistry. Cell Mol
Life Sci 2008;65:1019-48.. This is an extensive overview of the
general characteristics, genetics,
evolution and function of
aldehyde oxidases.
3. Garattini E, Fratelli M, Terao M. The
mammalian aldehyde oxidase gene
family. Hum Genomics 2009;4:119-30
4. Garattini E, Mendel R, Romao MJ, et al.
Mammalian molybdo-flavoenzymes, an
expanding family of proteins: structure,
genetics, regulation, function and
pathophysiology. Biochem J
2003;372:15-32.. The structural characteristics of
aldehyde oxidases and xanthine
oxidoreductases are the focus of this
review article.
5. Rodriguez-Trelles F, Tarrio R, Ayala FJ.
Convergent neofunctionalization by
positive Darwinian selection after ancient
recurrent duplications of the xanthine
dehydrogenase gene. Proc Natl Acad
Sci USA 2003;100:13413-17
6. Cazzaniga G, Terao M, Lo Schiavo P,
et al. Chromosomal mapping, isolation,
and characterization of the mouse
xanthine dehydrogenase gene. Genomics
1994;23:390-402
7. Kurosaki M, Zanotta S, Li Calzi M,
et al. Expression of xanthine
oxidoreductase in mouse mammary
epithelium during pregnancy and
lactation: regulation of gene expression
by glucocorticoids and prolactin.
Biochem J 1996;319:801-10
8. Terao M, Cazzaniga G, Ghezzi P, et al.
Molecular cloning of a cDNA coding for
mouse liver xanthine dehydrogenase.
Regulation of its transcript by interferons
in vivo. Biochem J 1992;283:863-70
9. Terao M, Kurosaki M, Zanotta S, et al.
The xanthine oxidoreductase gene:
structure and regulation.
Biochem Soc Trans 1997;25:791-6
10. Kuwabara Y, Nishino T, Okamoto K,
et al. Unique amino acids cluster for
switching from the dehydrogenase to
oxidase form of xanthine oxidoreductase.
Proc Natl Acad Sci USA
2003;100:8170-5
11. Enroth C, Eger BT, Okamoto K, et al.
Crystal structures of bovine milk
xanthine dehydrogenase and xanthine
oxidase: structure-based mechanism of
conversion. Proc Natl Acad Sci USA
2000;97:10723-8.. This article describes the crystal
structure of bovine xanthine
oxidoreductase, the only mammalian
molybdo-flavoproteins for which the
crystallographyc coordinates are
available. The structure is a template
for the prediction of the secondary
structure of aldehyde oxidases.
12. Calzi ML, Raviolo C, Ghibaudi E, et al.
Purification, cDNA cloning, and tissue
distribution of bovine liver aldehyde
oxidase. J Biol Chem
1995;270:31037-45. This is the first article reporting on the
amino acid sequence of a mammalian
aldehyde oxidase.
13. Terao M, Kurosaki M, Demontis S,
et al. Isolation and characterization of the
human aldehyde oxidase gene:
conservation of intron/exon boundaries
with the xanthine oxidoreductase gene
indicates a common origin. Biochem J
1998;332:383-93
14. Nishino T, Okamoto K, Eger BT, et al.
Mammalian xanthine oxidoreductase -
mechanism of transition from xanthine
dehydrogenase to xanthine oxidase.
FEBS J 2008;275:3278-89
15. Hille R. Molybdenum and tungsten in
biology. Trends Biochem Sci
2002;27:360-7
16. Terao M, Kurosaki M, Barzago MM,
et al. Role of the molybdoflavoenzyme
aldehyde oxidase homolog 2 in the
biosynthesis of retinoic acid: generation
and characterization of a knockout
mouse. Mol Cell Biol 2009;29:357-77.. The article reports on the first
knock-out animal for an aldehyde
oxidase gene, i.e. AOX4.
17. Neumeier M, Weigert J, Schaffler A,
et al. Aldehyde oxidase 1 is highly
abundant in hepatic steatosis and is
downregulated by adiponectin and
fenofibric acid in hepatocytes in vitro.
Biochem Biophys Res Commun
2006;350:731-5
18. Weigert J, Neumeier M, Bauer S, et al.
Small-interference RNA-mediated knock-
down of aldehyde oxidase 1 in
3T3-L1 cells impairs adipogenesis and
adiponectin release. FEBS Lett
2008;582:2965-72.. The study demonstrates that human
AOX1 is involved in the differentiation
of adipocytes and lipid synthesis/
accumulation.
19. Beedham C, Miceli JJ, Obach RS.
Ziprasidone metabolism, aldehyde
oxidase, and clinical implications.
J Clin Psychopharmacol 2003;23:229-32
20. Dalvie D, Zhang C, Chen W, et al.
Cross-species comparison of the
metabolism and excretion of zoniporide:
contribution of aldehyde oxidase to
interspecies differences.
Drug Metab Dispos 2011;38:641-54
21. Obach RS. Potent inhibition of human
liver aldehyde oxidase by raloxifene.
Drug Metab Dispos 2004;32:89-97
22. Obach RS, Huynh P, Allen MC, et al.
Human liver aldehyde oxidase: inhibition
by 239 drugs. J Clin Pharmacol
2004;44:7-19.. The article contains the most complete
list of aldehyde oxidase inhibitors so
far published.
23. Obach RS, Walsky RL. Drugs that
inhibit oxidation reactions catalyzed by
aldehyde oxidase do not inhibit the
reductive metabolism of ziprasidone to
its major metabolite,
S-methyldihydroziprasidone: an in
vitro study. J Clin Psychopharmacol
2005;25:605-8
24. Pryde DC, Dalvie D, Hu Q, et al.
Aldehyde oxidase: an enzyme of
emerging importance in drug discovery.
J Med Chem 2010;53:8441-60.. A very interesing review article
highlighting the importance of
aldehyde oxidase as an enzyme
involved in the metabolism of new
drug candidates.
25. Zientek M, Jiang Y, Youdim K, et al. In
vitro-in vivo correlation for intrinsic
E. Garattini & M. Terao
500 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

clearance for drugs metabolized by
human aldehyde oxidase.
Drug Metab Dispos 2010;38:1322-7
26. Beedham C. The role of
non-P450 enzymes in drug oxidation.
Pharm World Sci 1997;19:255-63
27. Critchley DJ, Rance DJ, Beedham C.
Biotransformation of carbazeran in
guinea pig: effect of hydralazine
pretreatment. Xenobiotica 1994;24:37-47
28. Panoutsopoulos GI, Beedham C. Kinetics
and specificity of guinea pig liver
aldehyde oxidase and bovine milk
xanthine oxidase towards substituted
benzaldehydes. Acta Biochim Pol
2004;51:649-63
29. Panoutsopoulos GI, Beedham C.
Metabolism of isovanillin by aldehyde
oxidase, xanthine oxidase, aldehyde
dehydrogenase and liver slices.
Pharmacology 2005;73:199-208
30. Panoutsopoulos GI, Kouretas D,
Beedham C. Contribution of aldehyde
oxidase, xanthine oxidase, and aldehyde
dehydrogenase on the oxidation of
aromatic aldehydes. Chem Res Toxicol
2004;17:1368-76
31. Rashidi MR, Beedham C, Smith JS,
et al. In vitro study of 6-mercaptopurine
oxidation catalysed by aldehyde oxidase
and xanthine oxidase.
Drug Metab Pharmacokinet
2007;22:299-306
32. Rashidi MR, Smith JA, Clarke SE, et al.
In vitro oxidation of famciclovir and
6-deoxypenciclovir by aldehyde oxidase
from human, guinea pig, rabbit, and rat
liver. Drug Metab Dispos
1997;25:805-13
33. Sahi J, Khan KK, Black CB. Aldehyde
oxidase activity and inhibition in
hepatocytes and cytosolic fractions from
mouse, rat, monkey and human.
Drug Metab Lett 2008;2:176-83
34. Beedham C. Molybdenum hydroxylases:
biological distribution and
substrate-inhibitor specificity.
Prog Med Chem 1987;24:85-127
35. Ruenitz PC, Bai X. Acidic metabolites of
tamoxifen. Aspects of formation and fate
in the female rat. Drug Metab Dispos
1995;23:993-8
36. Rochat B, Kosel M, Boss G, et al.
Stereoselective biotransformation of the
selective serotonin reuptake inhibitor
citalopram and its demethylated
metabolites by monoamine oxidases in
human liver. Biochem Pharmacol
1998;56:15-23
37. Ambroziak W, Izaguirre G, Pietruszko R.
Metabolism of retinaldehyde and other
aldehydes in soluble extracts of human
liver and kidney. J Biol Chem
1999;274:33366-73
38. Ghafourian T, Rashidi MR. Quantitative
study of the structural requirements of
phthalazine/quinazoline derivatives for
interaction with human liver aldehyde
oxidase. Chem Pharm Bull (Tokyo)
2001;49:1066-71
39. Hodnett CN, McCormack JJ, Sabean JA.
Oxidation of selected pteridine
derivatives by mamalian liver xanthine
oxidase and aldehyde oxidase.
J Pharm Sci 1976;65:1150-4
40. Kuroda T, Namba K, Torimaru T, et al.
Species differences in oral bioavailability
of methotrexate between rats and
monkeys. Biol Pharm Bull
2000;23:334-8
41. Relling MV, Lin JS, Ayers GD, et al.
Racial and gender differences in
N-acetyltransferase, xanthine oxidase, and
CYP1A2 activities. Clin Pharmacol Ther
1992;52:643-58
42. Moriwaki Y, Yamamoto T, Nasako Y,
et al. In vitro oxidation of pyrazinamide
and allopurinol by rat liver aldehyde
oxidase. Biochem Pharmacol
1993;46:975-81
43. Klecker RW, Cysyk RL, Collins JM.
Zebularine metabolism by aldehyde
oxidase in hepatic cytosol from humans,
monkeys, dogs, rats, and mice: influence
of sex and inhibitors. Bioorg Med Chem
2006;14:62-6
44. Robertson IG, Palmer BD, Officer M,
et al. Cytosol mediated metabolism of
the experimental antitumor agent
acridine carboxamide to the 9-acridone
derivative. Biochem Pharmacol
1991;42:1879-84
45. Boinpally RR, Zhou SL, LoRusso PM,
et al. Bioavailability and
pharmacokinetics of the investigational
anticancer agent XK469 (NSC 698215)
in rats following oral and intravenous
administration.
Cancer Chemother Pharmacol
2005;55:404-7
46. Acheampong AA, Chien DS, Lam S,
et al. Characterization of brimonidine
metabolism with rat, rabbit, dog,
monkey and human liver fractions and
rabbit liver aldehyde oxidase. Xenobiotica
1996;26:1035-55
47. Porter DJ, Harrington JA, Almond MR,
et al. 5-ethynyl-2(1H)-pyrimidinone:
aldehyde oxidase-activation to
5-ethynyluracil, a mechanism-based
inactivator of dihydropyrimidine
dehydrogenase. Biochem Pharmacol
1994;47:1165-71
48. Guo X, Lerner-Tung M, Chen HX,
et al. 5-Fluoro-2-pyrimidinone, a liver
aldehyde oxidase-activated prodrug of
5-fluorouracil. Biochem Pharmacol
1995;49:1111-16
49. Brandange S, Lindblom L. The enzyme
"aldehyde oxidase" is an iminium
oxidase. Reaction with nicotine delta
1’(5’) iminium ion. Biochem Biophys
Res Commun 1979;91:991-6
50. Vickers S, Polsky SL. The
biotransformation of nitrogen containing
xenobiotics to lactams. Curr Drug Metab
2000;1:357-89
51. Kitamura S, Sugihara K, Ohta S.
Drug-metabolizing ability of
molybdenum hydroxylases.
Drug Metab Pharmacokinet
2006;21:83-98
52. Kitamura S, Tatsumi K. Reduction of
tertiary amine N-oxides by liver
preparations: function of aldehyde
oxidase as a major N-oxide reductase.
Biochem Biophys Res Commun
1984;121:749-54
53. Kitamura S, Tatsumi K. Involvement of
liver aldehyde oxidase in the reduction of
nicotinamide N-oxide. Biochem Biophys
Res Commun 1984;120:602-6
54. Dick RA, Kanne DB, Casida JE.
Substrate specificity of rabbit aldehyde
oxidase for nitroguanidine and
nitromethylene neonicotinoid
insecticides. Chem Res Toxicol
2006;19:38-43
55. Linton A, Kang P, Ornelas M, et al.
Systematic Structure Modifications of
Imidazo[1,2-a]pyrimidine to Reduce
Metabolism Mediated by Aldehyde
Oxidase (AO). J Med Chem
2011;54:7705-12
56. Terao M, Kurosaki M, Saltini G, et al.
Cloning of the cDNAs coding for two
novel molybdo-flavoproteins showing
high similarity with aldehyde oxidase and
xanthine oxidoreductase. J Biol Chem
2000;275:30690-700
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 501
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

57. Terao M, Kurosaki M, Marini M, et al.
Purification of the aldehyde oxidase
homolog 1 (AOH1) protein and cloning
of the AOH1 and aldehyde oxidase
homolog 2 (AOH2) genes. Identification
of a novel molybdo-flavoprotein gene
cluster on mouse chromosome 1.
J Biol Chem 2001;276:46347-63.. This is the first study reporting the
presence of multiple aldehyde oxidase
isoenzymes in mice. The article
demonstrates that these isoenzymes are
the product of separate and conserved
genes resulting from specific
duplication events.
58. Kurosaki M, Terao M, Barzago MM,
et al. The aldehyde oxidase gene cluster
in mice and rats. Aldehyde oxidase
homologue 3, a novel member of the
molybdo-flavoenzyme family with
selective expression in the olfactory
mucosa. J Biol Chem
2004;279:50482-98
59. Kurosaki M, Demontis S, Barzago MM,
et al. Molecular cloning of the
cDNA coding for mouse aldehyde
oxidase: tissue distribution and regulation
in vivo by testosterone. Biochem J
1999;341:71-80
60. Vila R, Kurosaki M, Barzago MM, et al.
Regulation and biochemistry of mouse
molybdo-flavoenzymes. The DBA/
2 mouse is selectively deficient in the
expression of aldehyde oxidase
homologues 1 and 2 and represents a
unique source for the purification
and characterization of aldehyde
oxidase. J Biol Chem 2004;279:
8668-83. The article demonstrate the existence
of mouse strains characterized by
epigenetic suppression of AOX3 and
are characterized by a complement of
liver oxidases similar to that
of humans.
61. Sanoh S, Nozaki K, Murai H, et al.
Prediction of Human metabolism of
FK3453 by aldehyde oxidase using
chimeric mice transplanted with human
or rat hepatocytes. Drug Metab Dispos
2011; [Epub ahead of print].. This is the first report of a humanized
model of aldehyde oxidase dependent
drug metabolism.
62. Beedham C, Bruce SE, Critchley DJ,
et al. Species variation in hepatic
aldehyde oxidase activity. Eur J Drug
Metab Pharmacokinet 1987;12:307-10
63. Rodrigues AD. Comparison of levels of
aldehyde oxidase with cytochrome
P450 activities in human liver in vitro.
Biochem Pharmacol 1994;48:197-200
64. Rodrigues AD, Ferrero JL, Amann MT,
et al. The in vitro hepatic metabolism of
ABT-418, a cholinergic channel activator,
in rats, dogs, cynomolgus monkeys, and
humans. Drug Metab Dispos
1994;22:788-98
65. O’Connor D, Jones P, Chambers MS,
et al. Aldehyde oxidase and its
contribution to the metabolism of a
structurally novel, functionally selective
GABAA alpha5-subtype inverse agonist.
Xenobiotica 2006;36:315-30
66. Terao M, Kurosaki M, Barzago MM,
et al. Avian and canine aldehyde
oxidases. Novel insights into the biology
and evolution of molybdo-flavoenzymes.
J Biol Chem 2006;281:19748-61
67. Diaz GJ, Squires EJ. Role of aldehyde
oxidase in the hepatic in vitro
metabolism of 3-methylindole in pigs.
J Agric Food Chem 2000;48:833-7
68. Beedham C, al-Tayib Y, Smith JA. Role
of guinea pig and rabbit hepatic aldehyde
oxidase in oxidative in vitro metabolism
of cinchona antimalarials.
Drug Metab Dispos 1992;20:889-95
69. Beedham C, Critchley DJ, Rance DJ.
Substrate specificity of human liver
aldehyde oxidase toward substituted
quinazolines and phthalazines:
a comparison with hepatic enzyme from
guinea pig, rabbit, and baboon.
Arch Biochem Biophys 1995;319:481-90
70. Jordan CG, Rashidi MR, Laljee H, et al.
Aldehyde oxidase-catalysed oxidation of
methotrexate in the liver of guinea-pig,
rabbit and man. J Pharm Pharmacol
1999;51:411-18
71. Ardan T, Kovaceva J, Cejkova J.
Comparative histochemical and
immunohistochemical study on xanthine
oxidoreductase/xanthine oxidase in
mammalian corneal epithelium.
Acta Histochem 2004;106:69-75
72. Panoutsopoulos GI. Metabolism of
2-phenylethylamine to phenylacetic acid,
via the intermediate phenylacetaldehyde,
by freshly prepared and cryopreserved
guinea pig liver slices. In Vivo
2004;18:779-86
73. Panoutsopoulos G. Metabolism of
homovanillamine to homovanillic acid in
guinea pig liver slices.
Cell Physiol Biochem 2005;15:225-32
74. Panoutsopoulos GI. Phenylacetaldehyde
oxidation by freshly prepared and
cryopreserved guinea pig liver slices: the
role of aldehyde oxidase. Int J Toxicol
2005;24:103-9
75. Panoutsopoulos GI. Contribution of
aldehyde oxidizing enzymes on the
metabolism of 3,4-dimethoxy-2-
phenylethylamine to
3,4-dimethoxyphenylacetic acid by guinea
pig liver slices. Cell Physiol Biochem
2006;17:47-56
76. Pirouzpanah S, Rashidi MR, Delazar A,
et al. Inhibitory effects of Ruta
graveolens L. extract on guinea pig liver
aldehyde oxidase. Chem Pharm Bull
(Tokyo) 2006;54:9-13
77. Veskoukis AS, Kouretas D,
Panoutsopoulos GI. Substrate specificity
of guinea pig liver aldehyde oxidase and
bovine milk xanthine oxidase for methyl-
and nitrobenzaldehydes. Eur J Drug
Metab Pharmacokinet 2006;31:11-16
78. Kadam RS, Iyer KR. Isolation of liver
aldehyde oxidase containing fractions
from different animals and determination
of kinetic parameters for benzaldehyde.
Indian J Pharm Sci 2008;70:85-8
79. Pirouzpanah S, Hanaee J, Razavieh SV,
et al. Inhibitory effects of flavonoids on
aldehyde oxidase activity. J Enzyme Inhib
Med Chem 2009;24:14-21
80. Sugihara K, Kitamura S, Tatsumi K,
et al. Differences in aldehyde oxidase
activity in cytosolic preparations of
human and monkey liver. Biochem Mol
Biol Int 1997;41:1153-60
81. Kitamura S, Sugihara K, Nakatani K,
et al. Variation of hepatic methotrexate
7-hydroxylase activity in animals and
humans. IUBMB Life 1999;48:607-11
82. Kinsella TJ, Schupp JE, Davis TW, et al.
Preclinical study of the systemic toxicity
and pharmacokinetics of 5-iodo-2-
deoxypyrimidinone-2’-deoxyribose as a
radiosensitizing prodrug in two,
non-rodent animal species: implications
for phase I study design.
Clin Cancer Res 2000;6:3670-9
83. Mutlib AE, Shockcor J, Chen SY, et al.
Disposition of 1-[3-(aminomethyl)
phenyl]-N-[3-fluoro-2’- (methylsulfonyl)-
[1,1’-biphenyl]-4-yl]-3-(trifluoromethyl)-
1H-pyrazole-5-carboxamide (DPC 423)
by novel metabolic pathways.
E. Garattini & M. Terao
502 Expert Opin. Drug Metab. Toxicol. (2012) 8(4)
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.

Characterization of unusual metabolites
by liquid chromatography/mass
spectrometry and NMR.
Chem Res Toxicol 2002;15:48-62
84. Jones P, Atack JR, Braun MP, et al.
Pharmacokinetics and metabolism studies
on (3-tert-butyl-7-(5-methylisoxazol-3-
yl)-2-(1-methyl-1H-1,2,4-triazol-5-ylm
ethoxy) pyrazolo[1,5-d][1,2,4]triazine, a
functionally selective GABA(A)
alpha5 inverse agonist for cognitive
dysfunction. Bioorg Med Chem Lett
2006;16:872-5
85. Steinberg KK, Relling MV,
Gallagher ML, et al. Genetic studies of a
cluster of acute lymphoblastic leukemia
cases in Churchill County, Nevada.
Environ Health Perspect
2007;115:158-64.. This is the first report on the existence
of non-sense allelic variants of
aldehyde oxidase in the human
population with definition of the
relative allelic frequency.
86. Adachi M, Itoh K, Masubuchi A, et al.
Construction and expression of mutant
cDNAs responsible for genetic
polymorphism in aldehyde oxidase in
Donryu strain rats. J Biochem Mol Biol
2007;40:1021-7
87. Itoh K, Maruyama H, Adachi M, et al.
Lack of formation of aldehyde oxidase
dimer possibly due to
377G > A nucleotide substitution.
Drug Metab Dispos 2007;35:1860-4
88. Itoh K, Maruyama H, Adachi M, et al.
Lack of dimer formation ability in rat
strains with low aldehyde oxidase activity.
Xenobiotica 2007;37:709-16
89. Itoh K, Masubuchi A, Sasaki T, et al.
Genetic polymorphism of aldehyde
oxidase in Donryu rats.
Drug Metab Dispos 2007;35:734-9
90. Yoo CB, Chuang JC, Byun HM, et al.
Long-term epigenetic therapy with oral
zebularine has minimal side effects and
prevents intestinal tumors in mice.
Cancer Prev Res (Phila) 2008;1:233-40
91. Tayama Y, Moriyasu A, Sugihara K,
et al. Developmental changes of aldehyde
oxidase in postnatal rat liver.
Drug Metab Pharmacokinet
2007;22:119-24
92. Tayama Y, Miyake K, Sugihara K, et al.
Developmental changes of aldehyde
oxidase activity in young Japanese
children. Clin Pharmacol Ther
2007;81:567-72
93. Gormley PE, Rossitch E, D’Anna ME,
et al. An extremely potent anilinoacridine
inhibitor of aldehyde oxidase.
Biochem Biophys Res Commun
1983;116:759-64
94. Renwick AB, Ball SE, Tredger JM, et al.
Inhibition of zaleplon metabolism by
cimetidine in the human liver: in vitro
studies with subcellular fractions and
precision-cut liver slices. Xenobiotica
2002;32:849-62
95. Johns DG, Spector T, Robins RK.
Studies on the mode of oxidation of
pyrazolo(3,4-d)pyrimidine by aldehyde
oxidase and xanthine oxidase.
Biochem Pharmacol 1969;18:2371-83
96. Robertson IG, Gamage RS. Methadone:
a potent inhibitor of rat liver aldehyde
oxidase. Biochem Pharmacol
1994;47:584-7
97. Johnson C, Stubley-Beedham C,
Stell JG. Elevation of molybdenum
hydroxylase levels in rabbit liver after
ingestion of phthalazine or its
hydroxylated metabolite.
Biochem Pharmacol 1984;33:3699-705
98. Ohkubo M, Sakiyama S, Fujimura S.
Increase of nicotinamide
methyltransferase and
N1-methyl-nicotinamide oxidase
activities in the livers of the rats
administered alkylating agents.
Cancer Lett 1983;21:175-81
99. Rivera SP, Choi HH, Chapman B, et al.
Identification of aldehyde oxidase 1 and
aldehyde oxidase homologue 1 as
dioxin-inducible genes. Toxicology
2005;207:401-9
100. Alfaro JF, Joswig-Jones CA, Ouyang W,
et al. Purification and mechanism of
human aldehyde oxidase expressed in
Escherichia coli. Drug Metab Dispos
2009;37:2393-8
101. Schumann S, Terao M, Garattini E,
et al. Site directed mutagenesis of amino
acid residues at the active site of mouse
aldehyde oxidase AOX1. PLoS One
2009;4:e5348
AffiliationEnrico Garattini† & Mineko Terao†Author for correspondence
Laboratory of Molecular Biology, Istituto di
Ricerche Farmacologiche “Mario Negri”, via La
Masa 19, 20156 Milano, Italy
Tel: +39 02 39014533; Fax: +39 02 39014744;
E-mail: [email protected]
The role of aldehyde oxidase in drug metabolism
Expert Opin. Drug Metab. Toxicol. (2012) 8(4) 503
Exp
ert O
pin.
Dru
g M
etab
. Tox
icol
. Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f W
ashi
ngto
n on
04/
04/1
3Fo
r pe
rson
al u
se o
nly.





























