Embed Size (px)
Citation preview

of October 5, 2018.This information is current as
HypersensitivityThe Role for Decorin in Delayed-Type
GötteSchäfers, Renato V. Iozzo, Alexander Zarbock and MartinAnika Stadtmann, Sven Hermann, Klaus Schäfers, Michael Daniela G. Seidler, Negia A. Mohamed, Carla Bocian,
http://www.jimmunol.org/content/187/11/6108doi: 10.4049/jimmunol.1100373October 2011;
2011; 187:6108-6119; Prepublished online 31J Immunol
MaterialSupplementary
3.DC1http://www.jimmunol.org/content/suppl/2011/11/01/jimmunol.110037
Referenceshttp://www.jimmunol.org/content/187/11/6108.full#ref-list-1
, 20 of which you can access for free at: cites 55 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
The Role for Decorin in Delayed-Type Hypersensitivity
Daniela G. Seidler,* Negia A. Mohamed,* Carla Bocian,*,† Anika Stadtmann,‡,x
Sven Hermann,{ Klaus Schafers,{ Michael Schafers,{ Renato V. Iozzo,‖
Alexander Zarbock,‡,x and Martin Gotte†
Decorin, a small leucine-rich proteoglycan, regulates extracellular matrix organization, growth factor-mediated signaling, and cell
growth. Because decorin may directly modulate immune responses, we investigated its role in a mouse model of contact allergy
(oxazolone-mediated delayed-type hypersensitivity [DTH]) in decorin-deficient (Dcn2/2) and wild-type mice. Dcn2/2 mice showed
a reduced ear swelling 24 h after oxazolone treatment with a concurrent attenuation of leukocyte infiltration. These findings were
corroborated by reduced glucose metabolism, as determined by 18fluordeoxyglucose uptake in positron emission tomography
scans. Unexpectedly, polymorphonuclear leukocyte numbers in Dcn2/2 blood vessels were significantly increased and accompa-
nied by large numbers of flattened leukocytes adherent to the endothelium. Intravital microscopy and flow chamber and static
adhesion assays confirmed increased adhesion and reduced transmigration of Dcn2/2 leukocytes. Circulating blood neutrophil
numbers were significantly increased in Dcn2/2 mice 24 h after DTH elicitation, but they were only moderately increased in wild-
type mice. Expression of the proinflammatory cytokine TNF-a was reduced, whereas syndecan-1 and ICAM-1 were overexpressed
in inflamed ears of Dcn2/2 mice, indicating that these adhesion molecules could be responsible for increased leukocyte adhesion.
Decorin treatment of endothelial cells increased tyrosine phosphorylation and reduced syndecan-1 expression. Notably, absence of
syndecan-1 in a genetic background lacking decorin rescued the attenuated DTH phenotype of Dcn2/2 mice. Collectively, these
results implicated a role for decorin in mediating DTH responses by influencing polymorphonuclear leukocyte attachment to the
endothelium. This occurs via two nonmutually exclusive mechanisms that involve a direct antiadhesive effect on polymorphonu-
clear leukocytes and a negative regulation of ICAM-1 and syndecan-1 expression. The Journal of Immunology, 2011, 187: 6108–
6119.
Delayed-type hypersensitivity (DTH) is a common mousemodel for allergic contact dermatitis (1), which allowsthe study of cell-mediated immune responses in vivo (2).
The sensitization phase is characterized by covalent modifica-tion of surface proteins with the hapten oxazolone, followed bythe uptake and processing by dendritic and Langerhans cells that
migrate to lymph nodes and prime oxazolone-specific T cell pop-ulations (1, 2). During the elicitation phase, oxazolone exposureevokes recruitment and activation of primed T cells, followed bysynthesis and local release of chemokines and cytokines, as wellas mast cell degranulation. The release of vasoactive mediatorsleads to a massive leukocytic infiltration of the skin, a process thatdepends on cytokine gradients, and the synthesis and activation ofadhesion molecules of the integrin, selectin, and cell adhesionmolecule families (1, 2).During inflammation, polymorphonuclear leukocytes follow
chemotactic gradients to attach to activated endothelial cells, re-sulting in leukocyte diapedesis, penetration of the subendothelialmatrix, and migration into areas of tissue damage (3, 4). Thisprocess involves coordinated signaling events mediated by pro-inflammatory cytokines and chemokines, as well as sequentialinteractions with multiple adhesion molecules, including selectinsand their carbohydrate ligands, and integrins (3, 4). All of thesesteps are modulated by various types of proteoglycans (4, 5).Biochemical data demonstrated sequence-specific interactions ofglycosaminoglycans with a variety of ligands relevant to inflam-mation (6). For example, mice deficient in syndecan-1 (Sdc12/2)show increased leukocyte recruitment upon i.p. TNF-a stimula-tion, as well as during kidney inflammation, contact allergies, andcolitis (7–9). Although a role for heparan sulfate in inflammationis increasingly recognized, the role of dermatan/chondroitin sul-fate is less well investigated. Several observations indicated thatdermatan sulfate can interact with various cytokines and chemo-kines (10), including fibroblast growth factor family members(11), and heparin cofactor II (6). Moreover, injection of mice withdermatan sulfate increases the soluble levels of circulating ICAM-1 (12). From a clinical perspective, the use of dermatan sulfatemay be advantageous in comparison with heparin, because of the
*Institute of Physiological Chemistry and Pathobiochemistry, D-48149 Munster,Germany; †Department of Gynecology and Obstetrics, Munster University Hospital,D-48149 Munster, Germany; ‡Department of Anesthesiology and Critical Care Med-icine, University of Munster, D-48149 Munster, Germany; xMax-Planck Institute forMolecular Biomedicine, D-48149 Munster, Germany; {European Institute for Molec-ular Imaging, University of Munster, D-48149 Munster, Germany; and ‖Department ofPathology, Anatomy, and Cell Biology, Cancer Cell Biology and Signaling Program,Kimmel Cancer Center, Thomas Jefferson University, Philadelphia, PA 19107
Received for publication February 4, 2011. Accepted for publication September 29,2011.
This work was supported by the Innovative Medizinische Forschung of the MedicalFaculty Munster (I-SE 12 08 11), Deutsche Forschungsgemeinschaft SE 1431/3-1and GRK 1549 from the International Research Training Group “Molecular andCellular GlycoSciences” (to D.G.S. and M.G.), Deutsche ForschungsgemeinschaftAZ 428/3-1 (to A.Z.), German Israeli Foundation I-1004-136.11/2008 (to M.G.), theInterdisciplinary Center of Clinical Research (Munster, Germany; core unit SmAP)(to M.S. and S.H.), and Interdisciplinary Center for Clinical Research Grant Za2/001/10 (to A.Z.).
Address correspondence and reprint requests to Dr. Daniela G. Seidler or Dr. MartinGotte, Institute of Physiological Chemistry and Pathobiochemistry, Waldeyerstrasse15, D-48149 Munster, Germany (D.G.S.) or Department of Gynecology and Obstet-rics, Munster University Hospital, Albert-Schweitzer-Campus 1, Domagkstrasse 11,D-48149 Munster, Germany (M.G.). E-mail addresses: [email protected](D.G.S.) and [email protected] (M.G.)
The online version of this article contains supplemental material.
Abbreviations used in this article: CT, computerized tomography; DTH, delayed-typehypersensitivity; FDG, fluordeoxyglucose; %ID/ml, percentage of infected dose pervolume; KC, keratinocyte chemoattractant; PET, positron-emission tomography.
Copyright� 2011 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1100373
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

lack of anticoagulant side effects. The most prominent dermatansulfate proteoglycan in the skin is decorin, which is involvedin collagen fibrillogenesis and extracellular matrix organization(13–15). Decorin also acts as a key signaling molecule that canmodulate the activity of several tyrosine kinase receptors (16–18)and integrins (19). Furthermore, decorin inhibits growth of dif-ferent tumor cell types in vitro (20) and in vivo (21), via inter-actions with the epithelial growth factor receptor. In vitro, endo-thelial cells synthesize decorin under inflammatory conditions(22) and Dcn2/2 mice (23) show a delayed wound healing withenhanced blood vessel formation (24). In tubulointerstitial kidneyfibrosis, decorin deficiency enhances apoptosis and increases in-flammation (25). Based on these findings and considering thestructural homology of heparan and dermatan sulfate, we hypoth-esized that decorin could play a role in modulating contact al-lergy. Thus, we investigated the role of decorin in delayed-typeinflammation using an in vivo model of contact allergy and in vitromodels of leukocyte recruitment like intravital microscopy andflow chamber assays on P-selectin, ICAM-1, and CXCL-1. To ourknowledge, our results showed for the first time that decorin isexpressed by polymorphonuclear leukocytes and mononuclearcells and that it influences the expression of adhesion moleculeslike ICAM-1 and SDC1. Combined with the antiadhesive propertiesof decorin, this regulation of adhesion molecules promotes leuko-cyte extravasation into the tissue.
Materials and MethodsDecorin-null mice and decorin/syndecan-1 double-deficientmice
Decorin-deficient mice (Dcn2/2) (23) and syndecan-1–deficient mice(Sdc12/2) (26) were bred in the animal facility in accordance with theGerman Animal Protection Act and approved by the responsible EthicsReview Committee. Decorin/syndecan-1 double-deficient mice (Dcn2/2
/Sdc12/2) were generated by breeding and genotyped by genomic PCR, aspreviously described (23, 26).
DTH assay
DTH was carried out with 8–12-wk-old male Dcn2/2 mice and the re-spective controls, as described previously (8). Briefly, mice were sensitizedon abdominal shaved skin with 150 ml 2.5% oxazolone (Sigma, Deisen-hofen, Germany) dissolved in acetone/ethanol (3:1 [vol/vol]). Mice werechallenged 7 d later with 10 ml 1% oxazolone topically administrated tothe ear twice. Thickness of a constant area (1 cm2) of the ear was measuredwith a Mitutoyo engineer’s micrometer, immediately before challenge, aswell as at 24, 48, and 72 h and 7 d. Five mice per time point were used foreach experimental condition. Experiments were carried out four times withsimilar results. In total, we used 32 male wild-type and Dcn2/2 mice andexcluded 3 mice because they did not respond to the oxazolone treatment.
Positron-emission tomography, quantification, andcomputerized tomography scanning
Eight to twelve-week-old male Dcn2/2 mice (n = 9) and the respectivecontrols were subjected to DTH and investigated 24 h later. Animals wereanesthetized with isoflurane (1.8%) and placed on a heating pad tomaintain the body temperature. [18F]-fluordeoxyglucose (FDG) (10 MBqin 100 ml 0.9% saline) was injected i.v. 1 h prior to each positron-emissiontomography (PET) analysis. For PET acquisition, animals were placed on aheat-controlled multimodal scanning bed, and PET list mode data wereacquired for 15 min using the 32-module quadHIDAC scanner (OxfordPositron Systems, Weston-on-the-Green, U.K.) dedicated to small-animalimaging. The scanner has an effective resolution of 0.7 mm full-width athalf-maximum in the transaxial and axial directions when using an itera-tive resolution recovery reconstruction algorithm. Subsequently, the scan-ning bed was transferred to the computerized tomography (CT) scanner(Inveon, Siemens Medical Solutions), and a medium-resolution (25 mm)CT acquisition was performed for each mouse. PET data were recon-structed into a single-image volume for each mouse, with a voxel size of0.4 3 0.4 3 0.4 mm3. CT was reconstructed into a volume data set witha voxel size of 0.007 3 0.007 3 0.007 mm3. Image data sets were coreg-istered using extrinsic markers attached to the multimodal scanning bed
and the image analysis software (Inveon Research Workplace, SiemensMedical Solutions). For quantification of regional FDG uptake, the outerears were segmented three dimensionally using CT data, and the resultingvolume of interest was applied to the coregistered PET data set. Measureddata were corrected for partial volume effects, scatter, and backgroundusing a model-based approach. The FDG uptake was calculated as thepercentage of injected dose per volume (%ID/ml) for each ear, and the netuptake in the inflamed left ear (D%ID/ml) was calculated as (%ID/ml leftear) 2 (%ID/ml right ear).
Decorin purification
Decorin was purified from conditioned cell culture medium of humanfibroblasts by anion-exchange chromatography and analyzed for purity on asilver gel, as described previously (27, 28).
Adhesion assay of polymorphonuclear leukocytes onendothelial cells
The murine endothelial cell line bEnd.3 was used for static leukocyte-adhesion assays, as previously described (9). About 20,000 endothelialcells/96-well plate were cultured overnight, followed by treatment with5 nM TNF-a to stimulate endothelial cells, or were used directly for theexperiment. Under these conditions, TNF-a treatment did not result inincreased cytotoxicity (data not shown). Polymorphonuclear leukocytes ofwild-type and Dcn2/2 mice were prepared from bone marrow of tibias andfemurs, as previously described (8). A total of 2 3 106 cells/ml poly-morphonuclear leukocytes in PBS/1% FCS was incubated with 1 mMfluorescent marker 2979-bis-(2 carboxyethyl)-5 carboxyfluorescein ace-toxymethyl ester (Molecular Probes, Eugene, OR) in DMSO for 20 min at37˚C. Equal labeling efficiency was controlled using standard curves.Labeled samples were centrifuged for 5 min at 1500 rpm, and the pelletwas resuspended in medium. Polymorphonuclear leukocytes were treatedat 37˚C for 30 min with 5 mg/ml decorin and the respective solvent con-trols. Thereafter, endothelial cells were incubated with pretreated poly-morphonuclear leukocytes (2 3 106/ ml, 50 ml/well) for 10 min at 37˚C.Wells were washed two times with PBS, and adherent polymorphonu-clear leukocytes were lysed with lysis buffer (10 mM Tris/HCl, 0.1% SDS[pH 8.5]). The fluorescence signal was quantified in a Spectramax Fluo-rimeter (excitation, 485 nm; emission, 535 nm). The adhesion was reportedas the number of adherent cells/mm2. The results were expressed asmean 6 SEM.
Histology and immunohistochemistry
Paraffin-embedded ears were cut in 5-mm sections, and every 20–30thsection was stained with H&E. Sections were analyzed for polymorpho-nuclear leukocyte distribution in the tissue and in the blood vessels. Theblood vessel size was also evaluated for wild-type and Dcn2/2 mice. Free,round, flattened, and transmigrated polymorphonuclear leukocytes wereanalyzed by scoring adapted from Bixel et al. (29). Briefly, free or non-adherent and adherent polymorphonuclear leukocytes/104 mm2 of bloodvessel surface area were evaluated by light microscopy with a color viewsoft imaging system (Sis, Munster, Germany). Furthermore, extravasatedpolymorphonuclear leukocytes/104 mm2 of inflamed tissue surface in wild-type and Dcn2/2 were counted.
For immunohistochemistry, after rehydration the PFA-fixed sectionswere blocked with 10% BSA for 30 min at room temperature, followingincubation with either rat anti-mouse syndecan-1 mAb (BD Biosciences,Franklin Lakes, NJ) or rat anti-mouse ICAM-1 mAb (BioLegend, Uithoorn,The Netherlands), both 1/100 with PBS containing 1% BSA, overnight at4˚C; omission of the primary Ab served as a negative control. SDC1staining and development were carried out with secondary anti-rat IgGbiotinylated Ab (Vectastain ABC, Vector Labs, Burlingame, MA), dilutedas described by the manufacturer, followed by counterstaining withMayer’s hemalum (Merck, Darmstadt, Germany) and mounting in Kaiser’sglycerol gelatin (Merck) (8). For ICAM-1, a secondary peroxidase-conjugated affinity-purified anti-rat IgG (Dako, Glostrup, Germany), di-luted 1/1000 with PBS containing 1% BSA, was applied for 30 min atroom temperature. After washing with PBS, sections were incubated for 5min with diaminobenzidine reagent (Dako) and counterstained with he-matoxylin. Sections were analyzed with an Olympus photomicroscope.
For immunofluorescence staining, the tissuewas embedded in Tissue TekOCT compound (Sakura Finetek, Tokyo, Japan). Five-micron tissue sec-tions were fixed with methanol for 10 min at 220˚C, blocked with 1%BSA (Serva, Heidelberg, Germany)/PBS for 30 min at room tempera-ture, followed by incubation with rat anti-mouse CD4 (clone L3T4; BDPharmingen, Heidelberg, Germany), anti-mouse CD8 (clone 53–6.7;BD Pharmingen, Heidelberg, Germany), rat-anti human F4/80 (Abcam,
The Journal of Immunology 6109
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Cambridge, U.K.), or rabbit-anti human decorin antiserum LF-113 (kindlyprovided by Dr. Larry Fisher, National Institutes of Health, Bethesda,MD) for 1 h at room temperature. Positive cells were detected usingCy3-labeled donkey anti-rat IgG (H+L) (Dianova, Hamburg, Germany)or Alexa Fluor 488-labeled goat-anti rabbit IgG (Invitrogen, Karlsruhe,Germany).
Autoperfused flow chamber
To investigate neutrophil arrest, we used a previously published flowchamber system (30, 31). Briefly, rectangular glass capillaries (20 3 200mm, VitroCom) were coated with P-selectin (40 mg/ml), ICAM-1 (25 mg/ml), and CXCL1 (25 mg/ml; all from R&D Systems) for 2 h and blockedfor 1 h with casein (Thermo Fisher Scientific). The chamber was connectedat one side to a PE 50 tubing and used to control the wall shear stress. Theother side of the chamber was connected to a syringe filled with isolatedPMNs resuspended in PBS containing 1 mM Mg2+ and 1 mM Ca2+. Weperfused each chamber with PMNs for 6 min before we recorded repre-sentative fields of view for 30 s using an SW40/0.75 objective (AxioScope, Carl Zeiss).
Intravital microscopy
We prepared cremaster muscle for intravital microscopy, as describedpreviously (31, 32), and investigated leukocyte rolling flux fraction, rollingvelocity, adhesion, and extravasation in postcapillary venules (20–40 mmdiameter) 4 h after an intrascrotal injection of 50 ng IL-1b.
Reverse transcription and quantitative TaqMan real-time PCR
After euthanasia of the animals, ears were excised and snap-frozen in liquidnitrogen, followed by preparation of total RNA using the RNeasy kit(Qiagen, Hilden, Germany). One microgram of total RNA was transcribedusing the First Strand cDNA Synthesis Kit (Fermentas, St. Leon Rot, Ger-many). To detect Dcn, Sdc1, and Gapdh expression, conventional PCR wasused with primer pairs for Dcn-sense 59-CCT TCT GGC ACA AGT CTCTTG G-39, Dcn-antisense 59-TCG AAG ATG ACA CTG GCA TCG G-39(24), Sdc1-sense 59-GAC TCT GAC AAC TTC TCT GGC TCT-39, Sdc1-antisense 59-GCT GTG GTG ACT CTG ACT GTT G-93 and Gapdh. cDNAcorresponding to 25 ng total RNA was used as a template in the PCRconsisting of Applied Biosystems MasterMix and predesigned TaqMangene-expression systems (Applied Biosystems, Darmstadt, Germany), ac-cording to the manufacturer’s instructions. For detection of Sdc1, Tnfa,and Icam1 mRNA, primers Mm00448918_m1 (Sdc1 exons 2 and 3),Mm00443258_m1 (Tnfa exons 1 and 2), and Mm00516023_m1 (Icam1exons 2 and 3) were used and normalized to the expression of mammalian18S rRNA (Hs99999901_s1; all primers from Applied Biosystems). Quan-titative real-time PCR was performed with an Applied Biosystems PRISM7300 Sequence Detection System using the default thermal cycling con-ditions (10 min at 95˚C and 40 cycles of 15 s at 95˚C plus 1 min at 60˚C).Relative quantification was performed using the comparative cycle thresholdmethod (33). Three to five biological replicates were used for each timepoint investigated. The Mann–Whitney U test was used for statisticalanalysis. A p value , 0.05 was considered statistically significant.
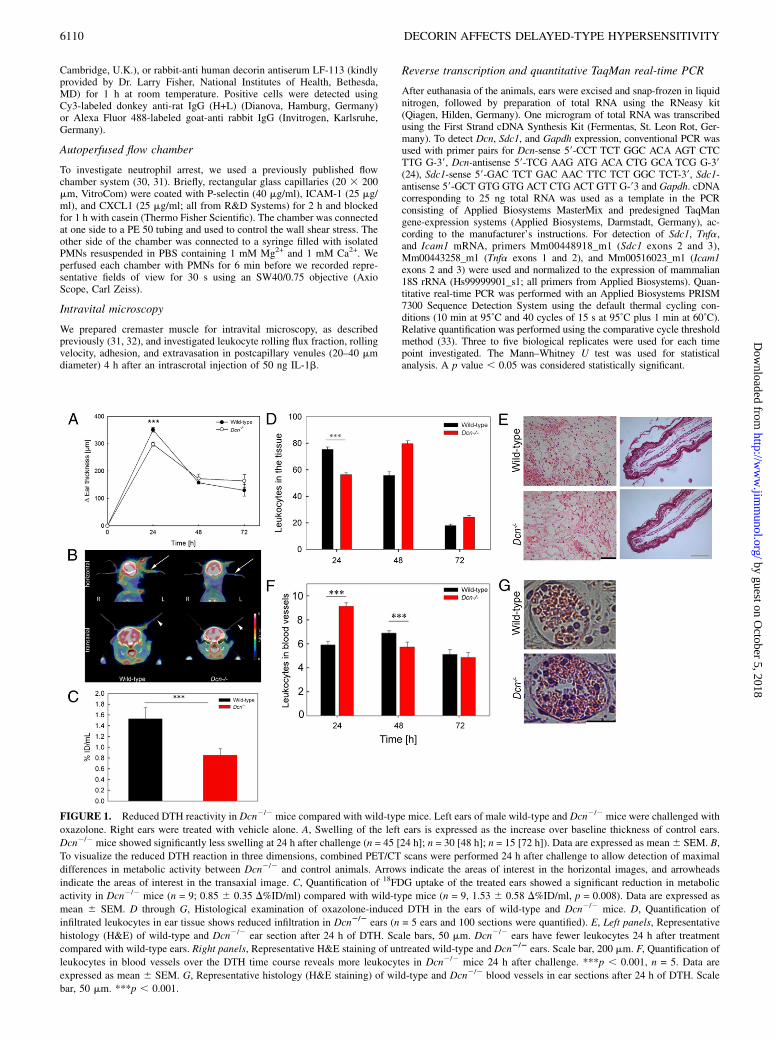
FIGURE 1. Reduced DTH reactivity in Dcn2/2 mice compared with wild-type mice. Left ears of male wild-type and Dcn2/2 mice were challenged with
oxazolone. Right ears were treated with vehicle alone. A, Swelling of the left ears is expressed as the increase over baseline thickness of control ears.
Dcn2/2 mice showed significantly less swelling at 24 h after challenge (n = 45 [24 h]; n = 30 [48 h]; n = 15 [72 h]). Data are expressed as mean6 SEM. B,
To visualize the reduced DTH reaction in three dimensions, combined PET/CT scans were performed 24 h after challenge to allow detection of maximal
differences in metabolic activity between Dcn2/2 and control animals. Arrows indicate the areas of interest in the horizontal images, and arrowheads
indicate the areas of interest in the transaxial image. C, Quantification of 18FDG uptake of the treated ears showed a significant reduction in metabolic
activity in Dcn2/2 mice (n = 9; 0.85 6 0.35 D%ID/ml) compared with wild-type mice (n = 9, 1.53 6 0.58 D%ID/ml, p = 0.008). Data are expressed as
mean 6 SEM. D through G, Histological examination of oxazolone-induced DTH in the ears of wild-type and Dcn2/2 mice. D, Quantification of
infiltrated leukocytes in ear tissue shows reduced infiltration in Dcn2/2 ears (n = 5 ears and 100 sections were quantified). E, Left panels, Representative
histology (H&E) of wild-type and Dcn2/2 ear section after 24 h of DTH. Scale bars, 50 mm. Dcn2/2 ears have fewer leukocytes 24 h after treatment
compared with wild-type ears. Right panels, Representative H&E staining of untreated wild-type and Dcn2/2 ears. Scale bar, 200 mm. F, Quantification of
leukocytes in blood vessels over the DTH time course reveals more leukocytes in Dcn2/2 mice 24 h after challenge. ***p , 0.001, n = 5. Data are
expressed as mean 6 SEM. G, Representative histology (H&E staining) of wild-type and Dcn2/2 blood vessels in ear sections after 24 h of DTH. Scale
bar, 50 mm. ***p , 0.001.
6110 DECORIN AFFECTS DELAYED-TYPE HYPERSENSITIVITY
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Protein extraction, ELISA, and immunoblotting
For protein extraction, excised ears were snap-frozen in liquid nitrogen andhomogenized, as described previously (8). Briefly, ears were homogenizedon ice with 500 ml PBS containing 10 mM EDTA and a mixture of proteaseinhibitors. Samples were centrifuged for 10 min at 12,000 3 g, and su-pernatant was collected. Total protein concentration was quantified byBCA-Lowry assay (Pierce, Rockford, IL). Protein extracts were used forELISA or Western blotting. All protein samples were diluted to 1.5 mg/mlkeratinocyte chemoattractant (KC) or 1 mg/ml TNF-a, and the tissue con-centrations of KC and TNF-a immunoassays were determined exactly asdescribed by the manufacturer (R&D Systems, Wiesbaden, Germany). ForWestern blotting, ∼40 mg protein extracts of ears derived from DTHexperiments or of bEnd.3 cells subjected to 24 h of TNF-a (5 nM) and/ordecorin (5 mg/ml) stimulation were loaded on a 12% SDS-gel under nonre-ducing conditions. After blotting, the nitrocellulose membrane was blockedwith 5% milk in TBST. The membrane was incubated with ICAM-1 Ab ratanti-mouse clone YN1/1.7.4 (BioLegend) or mouse anti P-tyrosine (P-Tyr-100; Cell Signaling) at 4˚C overnight. After washing the sections, HRP-labeled secondary anti-rat (Pierce, Rockland, PA) or anti-mouse (Calbio-chem) Abs were used to detect ICAM-1 or P-tyrosine, respectively. Decorinwas detected analogously following digestion of tissue extracts withChondroitinase ABC (Seikagaku, Kogyo, Japan) for 2 h at 37˚C, using apolyclonal antiserum kindly provided by Dr. Larry Fisher, and HRP-labeledgoat anti-rabbit IgG (Calbiochem) as a secondary Ab. The dot-blot for Sdc-1 was performed, as previously described (34), and analyzed densito-metrically using ImageJ software (National Institutes of Health).
Statistical analysis
Statistical evaluation was performed with GraphPad Prism. If not men-tioned, we used the Student t test, and p, 0.05 was considered significant.
ResultsDecorin deficiency attenuates edema formation and leukocyterecruitment in oxazolone-mediated DTH
To study the role of decorin in a contact allergy model, we usedoxazolone as a hapten and followed ear swelling as a readout ofinflammation-induced edema formation for up to 72 h afterchallenging the mice. Dcn2/2 mice showed a suppressed responseto oxazolone, ∼25% less swelling, compared with wild-type (p ,0.001, Fig. 1A). Next, inflammatory activity of the ears wasassessed noninvasively by PET/CT scanning, a strategy that allowsdirect visualization and quantification of tissue-metabolic activ-ity by administration of a radioactive sugar, 18FDG, as surrogatemarker for cellular glucose metabolism (35). We found that me-tabolism was reduced in Dcn2/2 ears (Fig. 1B). Quantification of18FDG uptake in the treated ears of different animals, normalizedon the signal of the contralateral ear, showed that wild-type micehad an uptake of 18FDG of 1.536 0.58%ID/ml. In contrast, Dcn2/2
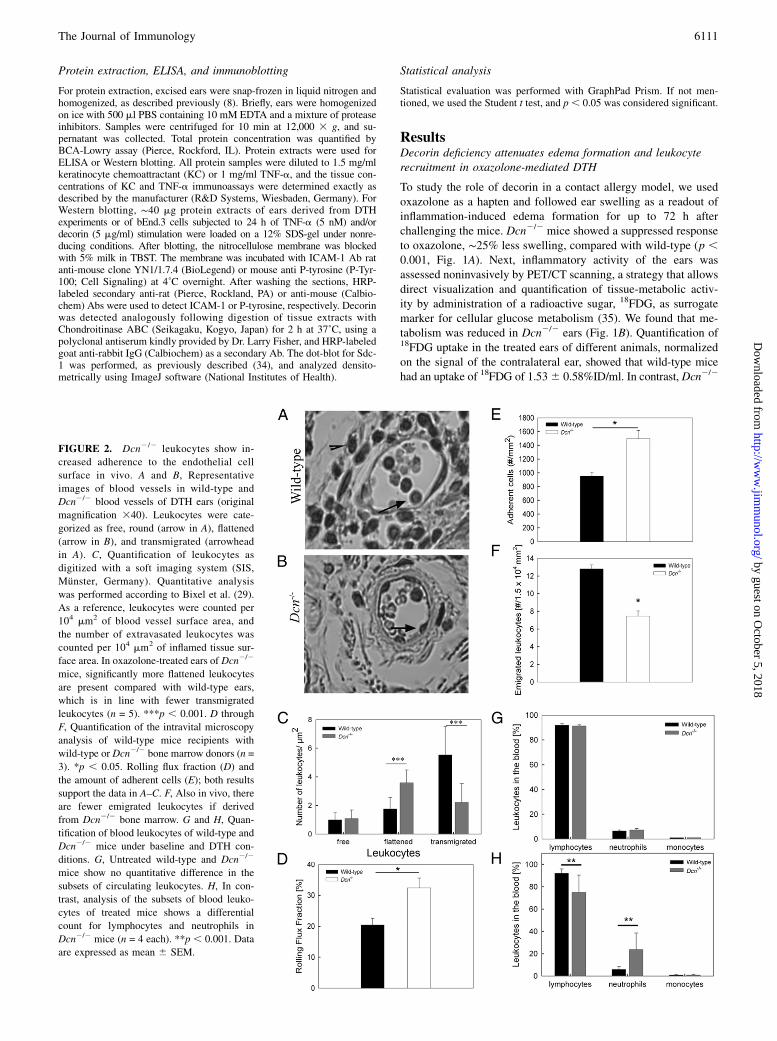
FIGURE 2. Dcn2/2 leukocytes show in-
creased adherence to the endothelial cell
surface in vivo. A and B, Representative
images of blood vessels in wild-type and
Dcn2/2 blood vessels of DTH ears (original
magnification 340). Leukocytes were cate-
gorized as free, round (arrow in A), flattened
(arrow in B), and transmigrated (arrowhead
in A). C, Quantification of leukocytes as
digitized with a soft imaging system (SIS,
Munster, Germany). Quantitative analysis
was performed according to Bixel et al. (29).
As a reference, leukocytes were counted per
104 mm2 of blood vessel surface area, and
the number of extravasated leukocytes was
counted per 104 mm2 of inflamed tissue sur-
face area. In oxazolone-treated ears of Dcn2/2
mice, significantly more flattened leukocytes
are present compared with wild-type ears,
which is in line with fewer transmigrated
leukocytes (n = 5). ***p , 0.001. D through
F, Quantification of the intravital microscopy
analysis of wild-type mice recipients with
wild-type or Dcn2/2 bone marrow donors (n =
3). *p , 0.05. Rolling flux fraction (D) and
the amount of adherent cells (E); both results
support the data in A–C. F, Also in vivo, there
are fewer emigrated leukocytes if derived
from Dcn2/2 bone marrow. G and H, Quan-
tification of blood leukocytes of wild-type and
Dcn2/2 mice under baseline and DTH con-
ditions. G, Untreated wild-type and Dcn2/2
mice show no quantitative difference in the
subsets of circulating leukocytes. H, In con-
trast, analysis of the subsets of blood leuko-
cytes of treated mice shows a differential
count for lymphocytes and neutrophils in
Dcn2/2 mice (n = 4 each). **p , 0.001. Data
are expressed as mean 6 SEM.
The Journal of Immunology 6111
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

mice showed a reduced tracer accumulation of 0.85 6 0.35%ID/ml, equivalent to an ∼40% reduction in metabolic activity (p ,0.001, Fig. 1C). These values are in good agreement with themeasured reduction in ear swelling in Dcn2/2 mice vis-a-viswild-type mice. Additionally, ear volume was identified by CTscanning, confirming the results obtained by caliper measure-ments (data not shown).Next, we examined leukocyte infiltration, because invasion of
leukocytes into the tissue is a pivotal step during DTH (1, 36).Quantification of total tissue leukocytes showed that, at 24 hpostchallenge, Dcn2/2 mice contained significantly less comparedwith wild-type mice (p , 0.001, Fig. 1D, 1E). The untreatedcontrol and Dcn2/2 mouse ear tissues showed no difference (Fig.1D, 1E, right panels). Histological examination of the treated earsrevealed a different leukocyte distribution within the tissue ofwild-type and Dcn2/2 mice, which was particularly prominent inblood vessels (Fig. 1F). At 24 h after oxazolone treatment, theblood vessels of Dcn2/2 treated ears contained significantly moreleukocytes compared with wild-type ones (Fig. 1F, 1G). Analysisof the blood vessel size revealed no differences (data not shown).In contrast, 48 h after elicitation, leukocyte numbers in the bloodvessels of Dcn2/2 mice had increased significantly compared withwild-type mice (Fig. 1F), and leukocyte recruitment into the tissueresumed, suggesting a delayed leukocyte infiltration followingDTH elicitation in the decorin-null background.
Dcn2/2 leukocytes show increased adherence to theendothelial cell surface in vivo
The previous results raised the question about which mechanismswould prevent leukocytes from leaving the blood vessel lumenduring inflammation in the absence of decorin. Therefore, the tissuesections were scored with respect to free luminal and attached andtransmigrated leukocytes (29). Quantification of 35 sections fromwild-type and Dcn2/2 ear sections revealed no significant differ-ence in the number of free leukocytes in the blood vessels (Fig.2A–C). However, we found a significant increase in the number offlattened leukocytes in Dcn2/2 ears (Fig. 2C, p , 0.001, n = 5).Although Dcn2/2 leukocytes adhered more frequently to the en-
dothelial cells, the number of transmigrated leukocytes decreasedcompared with wild-type ones 24 h after DTH elicitation (Fig.2C). To investigate the molecular mechanism of this observation,we performed intravital microscopy of the cremaster muscle usingchimeric mice reconstituted with wild-type or Dcn2/2 bonemarrow cells. We observed no differences in the rolling velocity ofthe leukocytes from both populations (data not shown), but therolling flux fraction was increased in chimeric mice reconstitutedwith bone marrow from Dcn2/2 mice (Fig. 2D, p , 0.05). Fur-thermore, these chimeric mice showed more adherent and trans-migrated leukocytes compared with chimeric mice reconstitutedwith wild-type bone marrow leukocytes (Fig. 2E, 2F; p , 0.05).Moreover, detailed analysis of leukocyte subsets extravasated intothe tissue was performed (Supplemental Fig. 1). Upon measuringperoxidase activity as a readout of neutrophil activity in ear pro-tein extracts (8), no differences between genotypes were observed24 h after DTH elicitation; however, after 48 h, a significant in-crease was observed in Dcn2/2 mouse ears compared with wild-type ones (Supplemental Fig. 1A, 1B). F4/80 staining revealed nodifference in macrophage numbers in the dermis between wild-type and Dcn2/2 treated ears (Supplemental Fig. 1C, 1D). Theimmune fluorescence staining revealed that the numbers of CD4+
leukocytes were not differently affected in wild-type and Dcn2/2
treated ears during DTH (Supplemental Fig. 1E, 1F). Similarly,the number of CD8+ cells was not significantly changed 24 hafter oxazolone treatment (Supplemental Fig. 1G, 1H). Analysisof blood leukocyte numbers did not reveal a significant differencebetween Dcn2/2 and wild-type mice (data not shown), as well asno changes in the differential blood analysis (Fig. 2G; n = 4).Interestingly, 24 h after oxazolone treatment, the number of neu-trophils increased significantly in Dcn2/2 mice compared withwild-type mice, whereas the number of lymphocytes decreasedsignificantly (Fig. 2H, n = 5; p , 0.05).
ICAM-1 and Sdc1 are differentially expressed in Dcn2/2 miceduring DTH
Extravasation is a complex process, which requires sequential stepsof leukocyte adhesion to endothelial cells followed by release of
FIGURE 3. ICAM-1 expression is differentially altered in wild-type and Dcn2/2 mice during DTH activity. A, Real-time PCR for ICAM-1 expression
during the time course of DTH (n = 3–5). The p value was not statistically significant. B and C, Increased Icam1 expression in Dcn2/2 mouse tissues is
confirmed by Western blotting 24 h after DTH induction. B, Representative Western blot. Ponceau S staining was used as loading control (lower panel). C,
Densitometric analysis (n = 4). *p = 0.039. D, Western blot analysis of ICAM-1 expression in plasma of wild-type and Dcn2/2 mice prior to and 24 h after
DTH elicitation; 20 mg of protein/lane was analyzed. GAPDH was used as a loading control.
6112 DECORIN AFFECTS DELAYED-TYPE HYPERSENSITIVITY
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
leukocytes. The cell surface adhesion molecules ICAM-1 andSDC1 are known to be involved in the multistep leukocyte-adhesion cascade (3, 4). Therefore, expression of ICAM-1 wasevaluated by quantitative real-time PCR (Fig. 3A, n = 3–5). Nosignificant differences in ICAM-1 mRNA expression were ob-served between wild-type and Dcn2/2 mice. In unstimulated con-trol ears, ICAM-1 protein was not detectable by immunohisto-chemistry for either genotype; however, 24 h after elicitation,Dcn2/2 ears expressed more ICAM-1 protein (Supplemental Fig.2). This observation was confirmed by Western blotting; althoughICAM-1 expression increased in wild-type mice during the DTHresponse, as expected, oxazolone-treated ears of Dcn2/2 miceshowed an even stronger increase compared with wild-type mice(Fig. 3B, 3C). ICAM-1 levels in the plasma were also increased inDcn2/2 mice compared with wild-type mice (Fig. 3D).Syndecan-1 is an adhesion molecule with anti-inflammatory
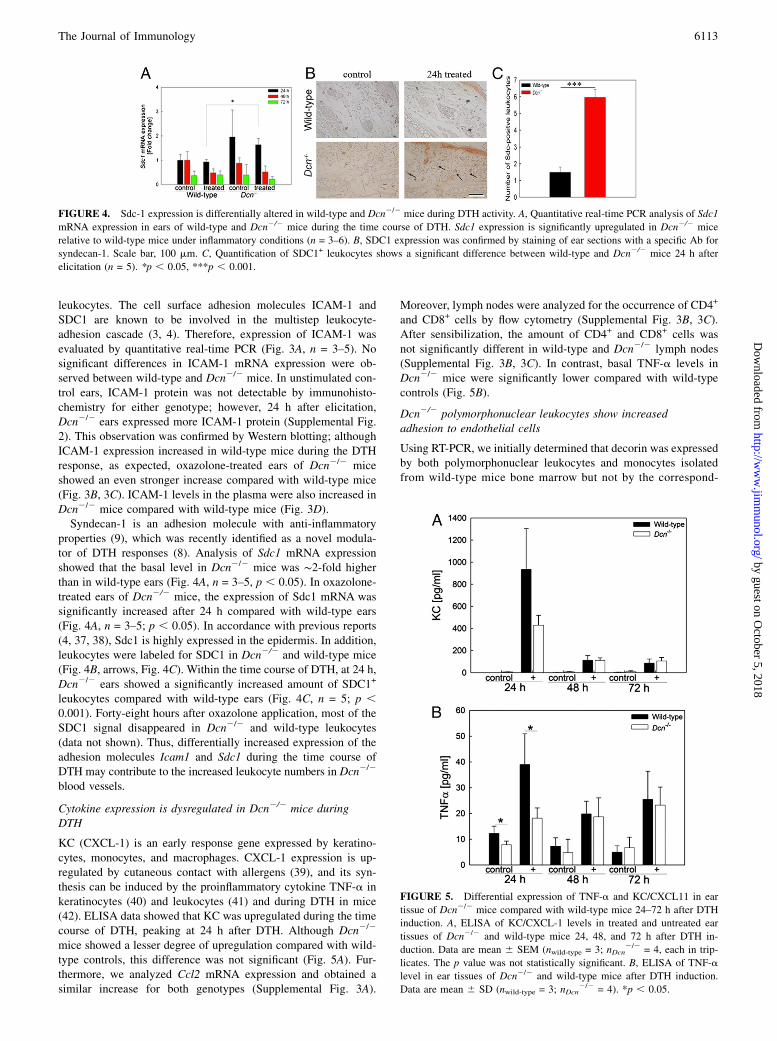
properties (9), which was recently identified as a novel modula-tor of DTH responses (8). Analysis of Sdc1 mRNA expressionshowed that the basal level in Dcn2/2 mice was ∼2-fold higherthan in wild-type ears (Fig. 4A, n = 3–5, p , 0.05). In oxazolone-treated ears of Dcn2/2 mice, the expression of Sdc1 mRNA wassignificantly increased after 24 h compared with wild-type ears(Fig. 4A, n = 3–5; p , 0.05). In accordance with previous reports(4, 37, 38), Sdc1 is highly expressed in the epidermis. In addition,leukocytes were labeled for SDC1 in Dcn2/2 and wild-type mice(Fig. 4B, arrows, Fig. 4C). Within the time course of DTH, at 24 h,Dcn2/2 ears showed a significantly increased amount of SDC1+
leukocytes compared with wild-type ears (Fig. 4C, n = 5; p ,0.001). Forty-eight hours after oxazolone application, most of theSDC1 signal disappeared in Dcn2/2 and wild-type leukocytes(data not shown). Thus, differentially increased expression of theadhesion molecules Icam1 and Sdc1 during the time course ofDTH may contribute to the increased leukocyte numbers in Dcn2/2
blood vessels.
Cytokine expression is dysregulated in Dcn2/2 mice duringDTH
KC (CXCL-1) is an early response gene expressed by keratino-cytes, monocytes, and macrophages. CXCL-1 expression is up-regulated by cutaneous contact with allergens (39), and its syn-thesis can be induced by the proinflammatory cytokine TNF-a inkeratinocytes (40) and leukocytes (41) and during DTH in mice(42). ELISA data showed that KC was upregulated during the timecourse of DTH, peaking at 24 h after DTH. Although Dcn2/2
mice showed a lesser degree of upregulation compared with wild-type controls, this difference was not significant (Fig. 5A). Fur-thermore, we analyzed Ccl2 mRNA expression and obtained asimilar increase for both genotypes (Supplemental Fig. 3A).
Moreover, lymph nodes were analyzed for the occurrence of CD4+
and CD8+ cells by flow cytometry (Supplemental Fig. 3B, 3C).After sensibilization, the amount of CD4+ and CD8+ cells wasnot significantly different in wild-type and Dcn2/2 lymph nodes(Supplemental Fig. 3B, 3C). In contrast, basal TNF-a levels inDcn2/2 mice were significantly lower compared with wild-typecontrols (Fig. 5B).
Dcn2/2 polymorphonuclear leukocytes show increasedadhesion to endothelial cells
Using RT-PCR, we initially determined that decorin was expressedby both polymorphonuclear leukocytes and monocytes isolatedfrom wild-type mice bone marrow but not by the correspond-
FIGURE 5. Differential expression of TNF-a and KC/CXCL11 in ear
tissue of Dcn2/2 mice compared with wild-type mice 24–72 h after DTH
induction. A, ELISA of KC/CXCL-1 levels in treated and untreated ear
tissues of Dcn2/2 and wild-type mice 24, 48, and 72 h after DTH in-
duction. Data are mean 6 SEM (nwild-type = 3; nDcn2/2 = 4, each in trip-
licates. The p value was not statistically significant. B, ELISA of TNF-a
level in ear tissues of Dcn2/2 and wild-type mice after DTH induction.
Data are mean 6 SD (nwild-type = 3; nDcn2/2 = 4). *p , 0.05.
FIGURE 4. Sdc-1 expression is differentially altered in wild-type and Dcn2/2 mice during DTH activity. A, Quantitative real-time PCR analysis of Sdc1
mRNA expression in ears of wild-type and Dcn2/2 mice during the time course of DTH. Sdc1 expression is significantly upregulated in Dcn2/2 mice
relative to wild-type mice under inflammatory conditions (n = 3–6). B, SDC1 expression was confirmed by staining of ear sections with a specific Ab for
syndecan-1. Scale bar, 100 mm. C, Quantification of SDC1+ leukocytes shows a significant difference between wild-type and Dcn2/2 mice 24 h after
elicitation (n = 5). *p , 0.05, ***p , 0.001.
The Journal of Immunology 6113
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
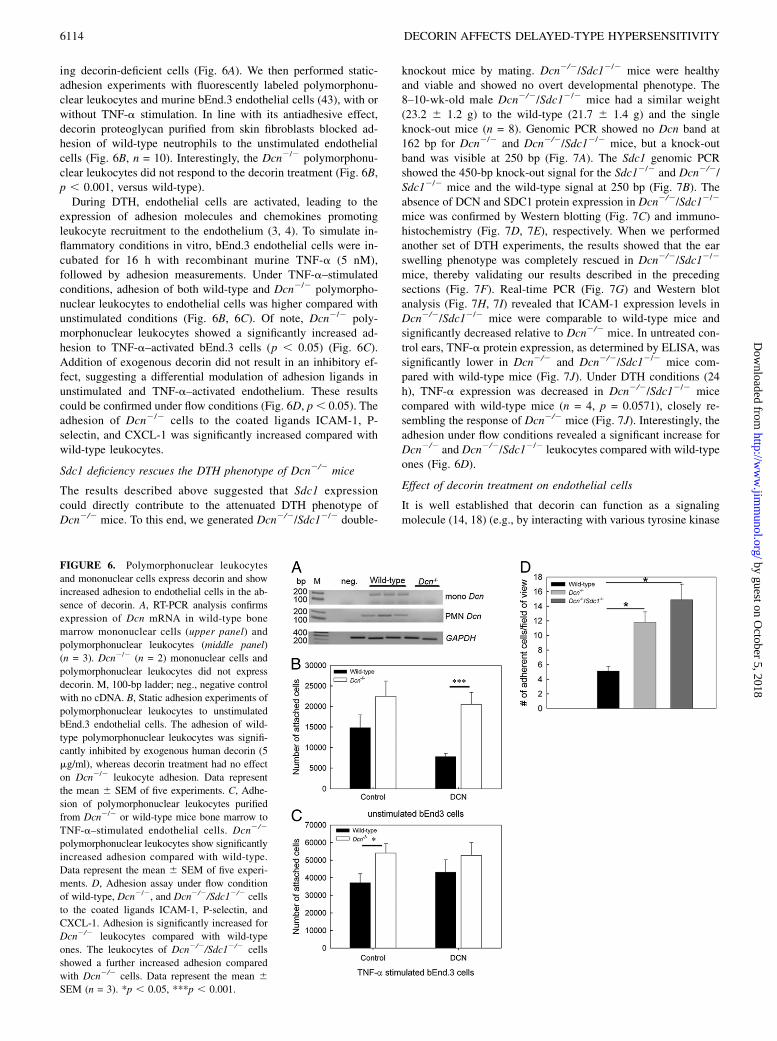
ing decorin-deficient cells (Fig. 6A). We then performed static-adhesion experiments with fluorescently labeled polymorphonu-clear leukocytes and murine bEnd.3 endothelial cells (43), with orwithout TNF-a stimulation. In line with its antiadhesive effect,decorin proteoglycan purified from skin fibroblasts blocked ad-hesion of wild-type neutrophils to the unstimulated endothelialcells (Fig. 6B, n = 10). Interestingly, the Dcn2/2 polymorphonu-clear leukocytes did not respond to the decorin treatment (Fig. 6B,p , 0.001, versus wild-type).During DTH, endothelial cells are activated, leading to the
expression of adhesion molecules and chemokines promotingleukocyte recruitment to the endothelium (3, 4). To simulate in-flammatory conditions in vitro, bEnd.3 endothelial cells were in-cubated for 16 h with recombinant murine TNF-a (5 nM),followed by adhesion measurements. Under TNF-a–stimulatedconditions, adhesion of both wild-type and Dcn2/2 polymorpho-nuclear leukocytes to endothelial cells was higher compared withunstimulated conditions (Fig. 6B, 6C). Of note, Dcn2/2 poly-morphonuclear leukocytes showed a significantly increased ad-hesion to TNF-a–activated bEnd.3 cells (p , 0.05) (Fig. 6C).Addition of exogenous decorin did not result in an inhibitory ef-fect, suggesting a differential modulation of adhesion ligands inunstimulated and TNF-a–activated endothelium. These resultscould be confirmed under flow conditions (Fig. 6D, p, 0.05). Theadhesion of Dcn2/2 cells to the coated ligands ICAM-1, P-selectin, and CXCL-1 was significantly increased compared withwild-type leukocytes.
Sdc1 deficiency rescues the DTH phenotype of Dcn2/2 mice
The results described above suggested that Sdc1 expressioncould directly contribute to the attenuated DTH phenotype ofDcn2/2 mice. To this end, we generated Dcn2/2/Sdc12/2 double-
knockout mice by mating. Dcn2/2/Sdc12/2 mice were healthyand viable and showed no overt developmental phenotype. The8–10-wk-old male Dcn2/2/Sdc12/2 mice had a similar weight(23.2 6 1.2 g) to the wild-type (21.7 6 1.4 g) and the singleknock-out mice (n = 8). Genomic PCR showed no Dcn band at162 bp for Dcn2/2 and Dcn2/2/Sdc12/2 mice, but a knock-outband was visible at 250 bp (Fig. 7A). The Sdc1 genomic PCRshowed the 450-bp knock-out signal for the Sdc12/2 and Dcn2/2/Sdc12/2 mice and the wild-type signal at 250 bp (Fig. 7B). Theabsence of DCN and SDC1 protein expression in Dcn2/2/Sdc12/2
mice was confirmed by Western blotting (Fig. 7C) and immuno-histochemistry (Fig. 7D, 7E), respectively. When we performedanother set of DTH experiments, the results showed that the earswelling phenotype was completely rescued in Dcn2/2/Sdc12/2
mice, thereby validating our results described in the precedingsections (Fig. 7F). Real-time PCR (Fig. 7G) and Western blotanalysis (Fig. 7H, 7I) revealed that ICAM-1 expression levels inDcn2/2/Sdc12/2 mice were comparable to wild-type mice andsignificantly decreased relative to Dcn2/2 mice. In untreated con-trol ears, TNF-a protein expression, as determined by ELISA, wassignificantly lower in Dcn2/2 and Dcn2/2/Sdc12/2 mice com-pared with wild-type mice (Fig. 7J). Under DTH conditions (24h), TNF-a expression was decreased in Dcn2/2/Sdc12/2 micecompared with wild-type mice (n = 4, p = 0.0571), closely re-sembling the response of Dcn2/2 mice (Fig. 7J). Interestingly, theadhesion under flow conditions revealed a significant increase forDcn2/2 and Dcn2/2/Sdc12/2 leukocytes compared with wild-typeones (Fig. 6D).
Effect of decorin treatment on endothelial cells
It is well established that decorin can function as a signalingmolecule (14, 18) (e.g., by interacting with various tyrosine kinase
FIGURE 6. Polymorphonuclear leukocytes
and mononuclear cells express decorin and show
increased adhesion to endothelial cells in the ab-
sence of decorin. A, RT-PCR analysis confirms
expression of Dcn mRNA in wild-type bone
marrow mononuclear cells (upper panel) and
polymorphonuclear leukocytes (middle panel)
(n = 3). Dcn2/2 (n = 2) mononuclear cells and
polymorphonuclear leukocytes did not express
decorin. M, 100-bp ladder; neg., negative control
with no cDNA. B, Static adhesion experiments of
polymorphonuclear leukocytes to unstimulated
bEnd.3 endothelial cells. The adhesion of wild-
type polymorphonuclear leukocytes was signifi-
cantly inhibited by exogenous human decorin (5
mg/ml), whereas decorin treatment had no effect
on Dcn2/2 leukocyte adhesion. Data represent
the mean 6 SEM of five experiments. C, Adhe-
sion of polymorphonuclear leukocytes purified
from Dcn2/2 or wild-type mice bone marrow to
TNF-a–stimulated endothelial cells. Dcn2/2
polymorphonuclear leukocytes show significantly
increased adhesion compared with wild-type.
Data represent the mean 6 SEM of five experi-
ments. D, Adhesion assay under flow condition
of wild-type, Dcn2/2, and Dcn2/2/Sdc12/2 cells
to the coated ligands ICAM-1, P-selectin, and
CXCL-1. Adhesion is significantly increased for
Dcn2/2 leukocytes compared with wild-type
ones. The leukocytes of Dcn2/2/Sdc12/2 cells
showed a further increased adhesion compared
with Dcn2/2 cells. Data represent the mean 6SEM (n = 3). *p , 0.05, ***p , 0.001.
6114 DECORIN AFFECTS DELAYED-TYPE HYPERSENSITIVITY
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 7. Absence of Sdc1 rescues the anti-inflammatory phenotype of Dcn2/2 mice. A and B, Genotyping of Dcn2/2/Sdc12/2 knockout mice. A,
Genomic PCR shows no Dcn signal (162 bp) for Dcn2/2 and Dcn2/2/Sdc12/2 mice, but it reveals the respective knock-out signal of 250 bp. B, Genomic PCR
shows no Sdc1 signal (250 bp) for Sdc12/2 and Dcn2/2/Sdc12/2 mice, but it reveals the respective knock-out signal of 450 bp. C, Western blotting reveals the
expression of DCN in extracts of wild-type (WT) and Sdc2/2 mouse ear tissue and its absence in Dcn2/2 and Dcn2/2/Sdc12/2 mouse ear tissue. D, Im-
munohistochemistry demonstrates the presence of SDC1 expression in the epithelial layer of wild-type mouse ears and the absence of SDC1 expression in
Dcn2/2/Sdc12/2 mouse ears (original magnification320). E, Immunofluorescence microscopy demonstrates the presence of DCN expression in the interstitial
matrix of wild-type mouse ears, and absence of DCN expression in Dcn2/2/Sdc12/2 mouse ears (original magnification320). F, Dcn2/2/Sdc12/2, wild-type,
and Dcn2/2 mice were used in a DTH assay, as described in Figure 1, to evaluate the contribution of Sdc1 to the phenotype of Dcn2/2 mice. Dcn2/2
mice show a reduced response to DTH compared with wild-type mice (compare with Fig. 1). Dcn2/2/Sdc12/2 mice respond (Figure legend continues)
The Journal of Immunology 6115
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

receptors, such as EGFR, the insulin-like growth factor receptor I,and c-Met) (16, 18, 19). Thus, we determined the effects of ex-ogenous decorin and TNF-a treatment on overall tyrosine phos-phorylation in bEnd.3 endothelial cells. Both decorin and TNF-asignificantly increased generalized Tyr phosphorylation (Fig. 8A).Furthermore, TNF-a–evoked phosphorylation was significantlypotentiated by decorin (Fig. 8B, n = 3, p , 0.01). Quantitativereal-time PCR analysis revealed that Sdc1 mRNA expression wassignificantly reduced by TNF-a and decorin treatment (Fig. 8C,n = 3, p , 0.01) but showed no additive effect for TNF-a +decorin treatment. Dot-blot analysis confirmed significant down-regulation of SDC1 protein expression by decorin treatment inTNF-a–stimulated bEnd.3 cells (Fig. 8D, n = 3, p , 0.05). Incontrast to the observed transcriptional downregulation, TNF-amono treatment did not result in significantly altered SDC1 pro-tein levels (Fig. 8D).
DiscussionDecorin is a well-established regulator of matrix assembly andgrowth factor activity (14, 44). The function of decorin is diverseand exerts both pro- and antifibrotic conditions, depending on theexperimental system (14, 23, 24, 44). In this study, we expanded thespectrum of decorin functions to the field of allergic skin inflam-mation, establishing a novel role in leukocyte recruitment duringDTH responses. Successful sensitization and upregulated earlycytokine expression (Ccl2) showed that decorin-deficient mice re-sponded to the allergen. However, compared with wild-type mice,lack of decorin resulted in an attenuated edema and metabolic ac-tivity at the site of inflammation, concomitant with a reductionin polymorphonuclear leukocytes transmigrating into the inflamedtissue. Histological studies showed that the reduced leukocyte di-apedesis in Dcn2/2 mice was associated with increased numbersof flattened, adherent polymorphonuclear leukocytes in the bloodvessels, as well as an increased percentage of circulating poly-morphonuclear leukocytes. Intravital microscopy experiments dem-onstrated that chimeric mice reconstituted with Dcn2/2 bone mar-row cells displayed an increased leukocyte adhesion and a reducednumber of transmigrated leukocytes. Similarly, in vitro, adhesionof Dcn2/2 polymorphonuclear leukocytes to endothelial cells wasincreased compared with wild-type ones, and wild-type adhesioncould be inhibited by addition of human decorin. Overall, our datasuggested that decorin directly modulates leukocyte adhesion to theendothelium and affects diapedesis of leukocytes into the inflamedtissue during DTH. Because the in vivo adhesion phenotype couldbe replicated in vitro using Dcn2/2 polymorphonuclear leukocytes,part of the phenotype appears to depend on decorin expression bypolymorphonuclear leukocytes, in accordance with the antiadhe-sive properties of decorin (45). Our flow chamber assays identifiedincreased adhesion of Dcn2/2 polymorphonuclear leukocytes toICAM-1, P-Selectin, and CXCL-1 as relevant ligands for thisprocess. Interestingly, polymorphonuclear leukocytes synthesizeddecorin but obviously not to a sufficient degree to block adhesioncompletely as we demonstrated with exogenous decorin. Increasednumbers of circulating polymorphonuclear cells in Dcn2/2 micemay be indicative of a lack of inhibitory signals on leukocyte
recruitment triggered by successful diapedesis, which is inhibitedat the 24-h time point.Under fibrotic conditions, genetic deficiency of decorin results in
an increased inflammatory reaction, likely mediated by the com-pensatory upregulation of biglycan (25), a known proinflammatoryproteoglycan that acts by activating TLR4 (46). In our model,biglycan expression did not appreciably change in either wild-typeor decorin-null–treated ears (data not shown), indicating that thereare tissue-specific changes. Notably, biglycan can stimulate thesynthesis of TNF-a and MIP-2 (46), whereas TNF-a was reducedin our Dcn2/2 model. Therefore, we concluded that biglycan isnot compensating for decorin under contact allergy conditions atthe analyzed time points. Moreover, our findings are supported bya recent study on the role of decorin in a mouse model of allergen-induced asthma: Marchica et al. (47) demonstrated reduced al-lergic inflammation in Dcn2/2 mice relative to wild-type mice,which could partially be attributed to differentially reducedamounts of TGF-b in the bronchoalveolar lavage of the allergen-stimulated decorin-deficient mice.The resolution of inflammation depends on the localization and
activation of lymphocytes and macrophages. Apparently, alteredmacrophage recruitment was not the cause of the reduced DTHresponse in Dcn2/2 mice. In contrast, it was shown that lumican,another member of the small leucine-rich proteoglycan family,is necessary for peritoneal polymorphonuclear leukocytes to ex-travasate into inflamed tissue in a b2 integrin-dependent manner(48).Increased adhesion of Dcn2/2 polymorphonuclear leukocytes
resembles previous observations in Dcn2/2 embryonic fibroblasts,for which an involvement of increased a2b1 integrin on the cellsurface was demonstrated (49, 50). Similarly, an increased ex-pression of adhesion molecules was observed in the current study,because the expression of the endothelial cell adhesion moleculeICAM-1, and of SDC1, a novel player in DTH regulation andleukocyte recruitment (8), was upregulated in Dcn2/2 mice vis-a-vis wild-type mice. Of note, dermatan sulfate, the major car-bohydrate moiety of skin-derived decorin, was shown to be in-volved in the induction of soluble ICAM-1 in an animal model(12). Similarly, under DTH conditions, endothelial cells expressedICAM-1, which increased during the course of DTH. In decorin-deficient mice, differential ICAM-1 upregulation during DTH maybe a potential compensatory mechanism activated in an attempt toincrease the amount of polymorphonuclear leukocytes availablefor diapedesis. Dressler et al. (51) showed that ICAM-1 is in-creased in endothelial cells near inflammation infiltration duringwound healing. Importantly, our data identified ICAM-1 as a rel-evant ligand for the increased recruitment of Dcn2/2 neutrophilscompared with wild-type ones. However, in the absence ofdecorin, leukocyte diapedesis, rather than adhesion, appears to beinhibited, suggesting that decorin also modulates downstreamsteps of the leukocyte-adhesion cascade (3, 4). Although specu-lative at this point, the strong increase in soluble ICAM-1 in theserum of Dcn2/2 mice may be linked to this phenomenon, be-cause it was shown that soluble ICAM-1 depletion from serumresults in increased leukocyte diapedesis in vitro (52).
to oxazolone similarly to wild-type mice. Compared with Dcn2/2 mice, the response is significantly different, indicating that Dcn-dependent modulation of
Sdc1 contributes to the DTH response in vivo (n = 5). G, Quantitative real-time PCR analysis for ICAM-1 expression in ear tissue of wild-type, Dcn2/2, and
Dcn2/2/Sdc12/2 mice 24 h after DTH elicitation. Data represent the mean6 SEM (n$ 3). H and I, Western blot analysis of ICAM-1 expression in ear tissue
of wild-type, Dcn2/2, Sdc12/2, and Dcn2/2/Sdc12/2 mice at 24 h after DTH elicitation. H, Semiquantitative densitometric analysis of ICAM-1 normalized to
total protein expression. Data represent the mean6 SEM (n$ 4). I, Representative Western blot. J, TNF-a ELISA of ear tissue extracts of wild-type, Dcn2/2,
and Dcn2/2/Sdc12/2 mice at 24 h after DTH elicitation. Data represent the mean 6 SEM (n $ 4). *p , 0.05, **p , 0.01.
6116 DECORIN AFFECTS DELAYED-TYPE HYPERSENSITIVITY
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Part of the phenotype reported in this study could be linked to theincreased expression of SDC1 in Dcn2/2 mice, because we re-cently demonstrated increased DTH reactions in Sdc12/2 mice
(8). Both Sdc12/2 (9) and Dcn2/2 polymorphonuclear leukocytesshow increased adhesion to endothelial cells in vitro. However,additional mechanisms, such as the coreceptor role of the heparansulfate chains of SDC1 in chemokine signaling (4, 5), or theproposed effect of SDC1 on edema formation during inflammation(reviewed in Ref. 4) may be of relevance in this context. ICAM-1expression was similar to wild-type levels in an Sdc12/2 back-ground, suggesting a potential mechanistic contribution of SDC1to upregulated ICAM-1 expression in the absence of decorin. Animportant contribution of SDC1 to the inflammatory phenotype ofDcn2/2 mice is further suggested by the observation that geneticablation of Sdc1 in the Dcn2/2 background efficiently rescued theear swelling phenotype of Dcn2/2 mice during DTH at the 24-htime point. Because both Sdc1 and Dcn deficiency promote leu-kocyte adhesion to endothelium and ICAM-1, reduced leukocyterecruitment does not appear to be the mechanism behind thisfinding. However, our intravital microscopy data demonstratedthat, although Dcn2/2 showed increased leukocyte adhesion, leu-kocyte transmigration was significantly inhibited. In contrast, aprevious study showed that Sdc12/2 mice exhibit both increasedleukocyte adhesion and transmigration during intravital micros-copy (37). Therefore, our data strongly suggested that the absenceof Dcn on the leukocytes plays a pivotal role in reducing allergicinflammation during DTH and that the absence of Sdc1 relievesthe block on diapedesis imposed on Dcn2/2 leukocytes. Diape-desis phenotypes similar to our findings have been reported inthe case of a loss of PECAM-1 or CD99 function (reviewed inRef. 53). In fact, impaired diapedesis, combined with a lack offeedback regulation indicating the presence of leukocytes in theinflamed tissue, may be a reason for the increased circulatingleukocyte numbers in Dcn2/2 mice, despite leukocyte hyper-adhesiveness. Sdc1-derived heparan sulfate chains may, in fact,bind to PECAM-1, possibly at a heparin-binding domain locatedbetween the IgG-like domains 2 and 3 (54) and, thereby, maycontribute to endothelial cell–endothelial cell integrity (55). Fur-ther work will be required to clarify the exact role of glyco-saminoglycans in PECAM-1 function. Finally, the differentialdownregulation of TNF-a expression observed in Dcn2/2 micewas also present in an Sdc2/2 background, suggesting that decorinmay either act upstream of SDC1 or that its effect on TNF-aexpression may be independent from SDC1.Sdc1 is expressed in polymorphonuclear leukocytes and mono-
nuclear cells, indicating that it might play a role in adhesion ofthese cell types. Consistent with the observed upregulation ofSdc1 in Dcn2/2 mice, bEnd.3 cells showed decreased expressionof Sdc1 mRNA and protein upon decorin treatment. Of note,decorin treatment of bEnd.3 cells resulted in an activation ofgeneral tyrosine phosphorylation, which was enhanced synergis-tically by TNF-a treatment. These findings are in accordance withthe established role for decorin as a modulator of receptor tyrosinekinase activation (16–18) and open up an additional mechanisticlevel of decorin function in contact allergic reactions. Althoughour bone marrow transplantation and in vitro adhesion assayssuggested a major contribution of decorin-deficient leukocytesto the DTH phenotype, its influence on endothelial receptor tyro-sine kinase signaling suggests that an endothelial contributioncannot be fully excluded. TNF-a treatment leads to significantdownregulation of Sdc1 mRNA expression, whereas its proteinexpression was not significantly altered. We can only speculatewhether SDC1 expression is subject to posttranscriptional regu-lation in our experimental system, similar to previous reports oncAMP-dependent modulation of SDC1 levels (56).In addition to altered adhesion molecule expression, cytokine
expression was dysregulated during DTH in Dcn2/2 mice. Ex-
FIGURE 8. Analysis of tyrosine phosphorylation and Sdc1 expression in
bEnd.3 cells stimulated with human decorin in the presence or absence of
TNF-a. A, Immunoblots for P-tyrosine after treatment of bEnd.3 cells with
TNF-a and decorin (5 mg/ml). The migration position of m.w. markers is
indicated at the left margin. B, Quantification of data as in A. Data represent
the mean 6 SEM of three experiments. **p , 0.01. C, Quantitative real-
time PCR analysis of Sdc1 mRNA expression in bEnd.3 cells treated with
TNF-a and decorin as described in A. Data represent the mean6 SEM (n =
3). *p, 0.05, **p, 0.01. D, Dot blot for SDC1 expression in bEnd.3 cells
following TNF-a stimulation with or without decorin treatment. Upper
panel, Representative dot blot. Lower panel, Densitometric quantification of
dot blot result (n = 3). *p , 0.05, TNF-a–treated samples.
The Journal of Immunology 6117
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

pression of proinflammatory TNF-a was reduced in Dcn2/2 ears24 h after DTH elicitation. Cellular adhesion molecules and C-X-C chemokines, such as KC/CXCL-1 and MIP-2, regulate tissueleukocyte accumulation in a multitude of inflammatory states.However, although TNF-a induces KC synthesis in keratinocytesduring DTH reactions (40, 42), the reduction in KC expression inDcn2/2 mice did not reach statistical significance. We concludedthat primarily the altered levels of TNF-a expression rather thanchanges in KC expression may have contributed to the altereddiapedesis, considering its key regulatory role in inflammation.In summary, we identified a novel role for decorin as a modulator
of contact hypersensitivity reactions. Decorin inhibits the tightadhesion of polymorphonuclear leukocytes to the endothelial layerof blood vessels, facilitates diapedesis in an SDC1-dependentmanner, and activates receptor tyrosine kinase-dependent signal-ing pathways. These events, in turn, induce transcriptional andposttranscriptional changes in TNF-a and adhesion molecule ex-pression. The loss of decorin results in increased expression ofICAM-1 and anti-inflammatory SDC1, as well as exerts a protec-tive influence on the allergen oxazolone.
AcknowledgmentsWe thank Margret Bahl, Anne Forsberg, Birgit Pers, and Christine Batza
for expert technical assistance; Dr. Deirdre Coombe for discussions; and
Dr. Larry Fisher for the generous gift of decorin antiserum.
DisclosuresThe authors have no financial conflicts of interest.
References1. Grabbe, S., and T. Schwarz. 1998. Immunoregulatory mechanisms involved in
elicitation of allergic contact hypersensitivity. Immunol. Today 19: 37–44.2. Abbas, A. K. 2005. Cell-mediated (type IV) hypersensitivity. In Pathologic Basis
of Disease, 7th Ed., V. Kumar, A. K. Abbas, and N. Fausto, eds. ElsevierSaunders, Philadelphia, p. 216–217.
3. Vestweber, D. 2007. Adhesion and signaling molecules controlling the trans-migration of leukocytes through endothelium. Immunol. Rev. 218: 178–196.
4. Gotte, M. 2003. Syndecans in inflammation. FASEB J. 17: 575–591.5. Parish, C. R. 2006. The role of heparan sulphate in inflammation. Nat. Rev.
Immunol. 6: 633–643.6. Taylor, K. R., and R. L. Gallo. 2006. Glycosaminoglycans and their proteo-
glycans: host-associated molecular patterns for initiation and modulation of in-flammation. FASEB J. 20: 9–22.
7. Rops, A. L., M. Gotte, M. H. Baselmans, M. J. van den Hoven, E. J. Steenbergen,J. F. Lensen, T. J. Wijnhoven, F. Cevikbas, L. P. van den Heuvel, T. H. vanKuppevelt, et al. 2007. Syndecan-1 deficiency aggravates anti-glomerularbasement membrane nephritis. Kidney Int. 72: 1204–1215.
8. Kharabi Masouleh, B., G. B. Ten Dam, M. K. Wild, R. Seelige, J. van der Vlag,A. L. Rops, F. G. Echtermeyer, D. Vestweber, T. H. van Kuppevelt, L. Kiesel,and M. Gotte. 2009. Role of the heparan sulfate proteoglycan syndecan-1(CD138) in delayed-type hypersensitivity. J. Immunol. 182: 4985–4993.
9. Floer, M., M. Gotte, M. K. Wild, J. Heidemann, E. S. Gassar, W. Domschke,L. Kiesel, A. Luegering, and T. Kucharzik. 2010. Enoxaparin improves thecourse of dextran sodium sulfate-induced colitis in syndecan-1-deficient mice.Am. J. Pathol. 176: 146–157.
10. Kawashima, H., K. Atarashi, M. Hirose, J. Hirose, S. Yamada, K. Sugahara, andM. Miyasaka. 2002. Oversulfated chondroitin/dermatan sulfates containingGlcAbeta1/IdoAalpha1-3GalNAc(4,6-O-disulfate) interact with L- and P-selectin and chemokines. J. Biol. Chem. 277: 12921–12930.
11. Seidler, D. G., J. Peter-Katalinic, and A. D. Zamfir. 2007. Galactosaminoglycanfunction and oligosaccharide structure determination. ScientificWorldJournal 7:233–241.
12. Penc, S. F., B. Pomahac, E. Eriksson, M. Detmar, and R. L. Gallo. 1999. Der-matan sulfate activates nuclear factor-kappab and induces endothelial and cir-culating intercellular adhesion molecule-1. J. Clin. Invest. 103: 1329–1335.
13. Reed, C. C., and R. V. Iozzo. 2002. The role of decorin in collagen fibrillogenesisand skin homeostasis. Glycoconj. J. 19: 249–255.
14. Schaefer, L., and R. V. Iozzo. 2008. Biological functions of the small leucine-rich proteoglycans: from genetics to signal transduction. J. Biol. Chem. 283:21305–21309.
15. Kalamajski, S., and A. Oldberg. 2010. The role of small leucine-rich proteo-glycans in collagen fibrillogenesis. Matrix Biol. 29: 248–253.
16. Schonherr, E., C. Sunderkotter, R. V. Iozzo, and L. Schaefer. 2005. Decorin,a novel player in the insulin-like growth factor system. J. Biol. Chem. 280:15767–15772.
17. Feugaing, D. D., R. Tammi, F. G. Echtermeyer, H. Stenmark, H. Kresse,M. Smollich, E. Schonherr, L. Kiesel, and M. Gotte. 2007. Endocytosis of thedermatan sulfate proteoglycan decorin utilizes multiple pathways and is modu-lated by epidermal growth factor receptor signaling. Biochimie 89: 637–657.
18. Goldoni, S., A. Humphries, A. Nystrom, S. Sattar, R. T. Owens, D. J. McQuillan,K. Ireton, and R. V. Iozzo. 2009. Decorin is a novel antagonistic ligand of theMet receptor. J. Cell Biol. 185: 743–754.
19. Fiedler, L. R., E. Schonherr, R. Waddington, S. Niland, D. G. Seidler,D. Aeschlimann, and J. A. Eble. 2008. Decorin regulates endothelial cell motilityon collagen I through activation of insulin-like growth factor I receptor andmodulation of alpha2beta1 integrin activity. J. Biol. Chem. 283: 17406–17415.
20. Santra, M., D. M. Mann, E. W. Mercer, T. Skorski, B. Calabretta, andR. V. Iozzo. 1997. Ectopic expression of decorin protein core causes a general-ized growth suppression in neoplastic cells of various histogenetic origin andrequires endogenous p21, an inhibitor of cyclin-dependent kinases. J. Clin. In-vest. 100: 149–157.
21. Seidler, D. G., S. Goldoni, C. Agnew, C. Cardi, M. L. Thakur, R. T. Owens,D. J. McQuillan, and R. V. Iozzo. 2006. Decorin protein core inhibits in vivocancer growth and metabolism by hindering epidermal growth factor receptorfunction and triggering apoptosis via caspase-3 activation. J. Biol. Chem. 281:26408–26418.
22. Nelimarkka, L., H. Salminen, T. Kuopio, S. Nikkari, T. Ekfors, J. Laine,L. Pelliniemi, and H. Jarvelainen. 2001. Decorin is produced by capillary endo-thelial cells in inflammation-associated angiogenesis. Am. J. Pathol. 158: 345–353.
23. Danielson, K. G., H. Baribault, D. F. Holmes, H. Graham, K. E. Kadler, andR. V. Iozzo. 1997. Targeted disruption of decorin leads to abnormal collagenfibril morphology and skin fragility. J. Cell Biol. 136: 729–743.
24. Jarvelainen, H., P. Puolakkainen, S. Pakkanen, E. L. Brown, M. Hook, R. V. Iozzo,E. H. Sage, and T. N. Wight. 2006. A role for decorin in cutaneous wound healingand angiogenesis. Wound Repair Regen. 14: 443–452.
25. Schaefer, L., K. Macakova, I. Raslik, M. Micegova, H. J. Grone, E. Schonherr,H. Robenek, F. G. Echtermeyer, S. Grassel, P. Bruckner, et al. 2002. Absence ofdecorin adversely influences tubulointerstitial fibrosis of the obstructed kidneyby enhanced apoptosis and increased inflammatory reaction. Am. J. Pathol. 160:1181–1191.
26. Stepp, M. A., H. E. Gibson, P. H. Gala, D. D. Iglesia, A. Pajoohesh-Ganji, S. Pal-Ghosh, M. Brown, C. Aquino, A. M. Schwartz, O. Goldberger, et al. 2002.Defects in keratinocyte activation during wound healing in the syndecan-1-deficient mouse. J. Cell Sci. 115: 4517–4531.
27. Seidler, D. G., L. Schaefer, H. Robenek, R. V. Iozzo, H. Kresse, and E. Schonherr.2005. A physiologic three-dimensional cell culture system to investigate the roleof decorin in matrix organisation and cell survival. Biochem. Biophys. Res.Commun. 332: 1162–1170.
28. Zamfir, A. D., C. Flangea, E. Sisu, A. F. Serb, N. Dinca, P. Bruckner, andD. G. Seidler. 2009. Analysis of novel over- and under-sulfated glycosamino-glycan sequences by enzyme cleavage and multiple stage MS. Proteomics 9:3435–3444.
29. Bixel, M. G., B. Petri, A. G. Khandoga, A. Khandoga, K. Wolburg-Buchholz,H. Wolburg, S. Marz, F. Krombach, and D. Vestweber. 2007. A CD99-relatedantigen on endothelial cells mediates neutrophil but not lymphocyte extravasa-tion in vivo. Blood 109: 5327–5336.
30. Zarbock, A., T. L. Deem, T. L. Burcin, and K. Ley. 2007. Galphai2 is requiredfor chemokine-induced neutrophil arrest. Blood 110: 3773–3779.
31. Zarbock, A., C. L. Abram, M. Hundt, A. Altman, C. A. Lowell, and K. Ley.2008. PSGL-1 engagement by E-selectin signals through Src kinase Fgr andITAM adapters DAP12 and FcR gamma to induce slow leukocyte rolling. J. Exp.Med. 205: 2339–2347.
32. Mueller, H., A. Stadtmann, H. Van Aken, E. Hirsch, D. Wang, K. Ley, andA. Zarbock. 2010. Tyrosine kinase Btk regulates E-selectin-mediated integrinactivation and neutrophil recruitment by controlling phospholipase C (PLC)gamma2 and PI3Kgamma pathways. Blood 115: 3118–3127.
33. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(2Delta Delta C(T)) Method.Methods 25: 402–408.
34. Vanhoutte, D., M. W. Schellings, M. Gotte, M. Swinnen, V. Herias, M. K. Wild,D. Vestweber, E. Chorianopoulos, V. Cortes, A. Rigotti, et al. 2007. Increasedexpression of syndecan-1 protects against cardiac dilatation and dysfunctionafter myocardial infarction. Circulation 115: 475–482.
35. Stelljes, M., S. Hermann, J. Albring, G. Kohler, M. Loffler, C. Franzius,C. Poremba, V. Schlosser, S. Volkmann, C. Opitz, et al. 2008. Clinical molecularimaging in intestinal graft-versus-host disease: mapping of disease activity,prediction, and monitoring of treatment efficiency by positron emission to-mography. Blood 111: 2909–2918.
36. Ahlfors, E., R. Jonsson, and C. Czerkinsky. 1996. Experimental T cell-mediatedinflammatory reactions in the murine oral mucosa. II. Immunohistochemicalcharacterization of resident and infiltrating cells. Clin. Exp. Immunol. 104: 297–305.
37. Gotte, M., A. M. Joussen, C. Klein, P. Andre, D. D. Wagner, M. T. Hinkes,B. Kirchhof, A. P. Adamis, and M. Bernfield. 2002. Role of syndecan-1 inleukocyte-endothelial interactions in the ocular vasculature. Invest. Ophthalmol.Vis. Sci. 43: 1135–1141.
38. McDermott, S. P., E. A. Ranheim, V. S. Leatherberry, S. S. Khwaja, K. S. Klos,and C. M. Alexander. 2007. Juvenile syndecan-1 null mice are protected fromcarcinogen-induced tumor development. Oncogene 26: 1407–1416.
39. Engeman, T., A. V. Gorbachev, D. D. Kish, and R. L. Fairchild. 2004. The in-tensity of neutrophil infiltration controls the number of antigen-primed CD8T cells recruited into cutaneous antigen challenge sites. J. Leukoc. Biol. 76: 941–949.
6118 DECORIN AFFECTS DELAYED-TYPE HYPERSENSITIVITY
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

40. Yamada, H., M. Matsukura, T. Yudate, J. Chihara, G. Stingl, and T. Tezuka.
1997. Enhanced production of RANTES, an eosinophil chemoattractant factor,
by cytokine-stimulated epidermal keratinocytes. Int. Arch. Allergy Immunol. 114
(Suppl. 1): 28–32.41. Zarbock, A., C. A. Lowell, and K. Ley. 2007. Spleen tyrosine kinase Syk is
necessary for E-selectin-induced alpha(L)beta(2) integrin-mediated rolling on
intercellular adhesion molecule-1. Immunity 26: 773–783.42. Cunha, T. M., W. A. Verri, Jr., D. A. Valerio, A. T. Guerrero, L. G. Nogueira,
S. M. Vieira, D. G. Souza, M. M. Teixeira, S. Poole, S. H. Ferreira, and
F. Q. Cunha. 2008. Role of cytokines in mediating mechanical hypernoci-
ception in a model of delayed-type hypersensitivity in mice. Eur. J. Pain 12:
1059–1068.43. Montesano, R., M. S. Pepper, U. Mohle-Steinlein, W. Risau, E. F. Wagner, and
L. Orci. 1990. Increased proteolytic activity is responsible for the aberrant
morphogenetic behavior of endothelial cells expressing the middle T oncogene.
Cell 62: 435–445.44. Seidler, D. G., and R. Dreier. 2008. Decorin and its galactosaminoglycan chain:
extracellular regulator of cellular function? IUBMB Life 60: 729–733.45. Winnemoller, M., P. Schon, P. Vischer, and H. Kresse. 1992. Interactions be-
tween thrombospondin and the small proteoglycan decorin: interference with
cell attachment. Eur. J. Cell Biol. 59: 47–55.46. Schaefer, L., A. Babelova, E. Kiss, H. J. Hausser, M. Baliova, M. Krzyzankova,
G. Marsche, M. F. Young, D. Mihalik, M. Gotte, et al. 2005. The matrix com-
ponent biglycan is proinflammatory and signals through Toll-like receptors 4 and
2 in macrophages. J. Clin. Invest. 115: 2223–2233.47. Marchica, C. L., V. Pinelli, M. Borges, J. Zummer, V. Narayanan, R. V. Iozzo,
and M. S. Ludwig. 2011. A role for decorin in a murine model of allergen-
induced asthma. Am. J. Physiol. Lung Cell. Mol. Physiol. 300: L863–L873.
48. Lee, S., K. Bowrin, A. R. Hamad, and S. Chakravarti. 2009. Extracellular matrixlumican deposited on the surface of neutrophils promotes migration by bindingto beta2 integrin. J. Biol. Chem. 284: 23662–23669.
49. Ferdous, Z., V. M. Wei, R. V. Iozzo, M. Hook, and K. J. Grande-Allen. 2007.Decorin-transforming growth factor- interaction regulates matrix organizationand mechanical characteristics of three-dimensional collagen matrices. J. Biol.Chem. 282: 35887–35898.
50. Ferdous, Z., S. B. Peterson, H. Tseng, D. K. Anderson, R. V. Iozzo, andK. J. Grande-Allen. 2010. A role for decorin in controlling proliferation, adhe-sion, and migration of murine embryonic fibroblasts. J. Biomed. Mater. Res. A93: 419–428.
51. Dressler, J., L. Bachmann, M. Kasper, J. G. Hauck, and E. Muller. 1997. Timedependence of the expression of ICAM-1 (CD 54) in human skin wounds. Int. J.Legal Med. 110: 299–304.
52. Ohno, N., H. Ichikawa, L. Coe, P. R. Kvietys, D. N. Granger, and J. S. Alexander.1997. Soluble selectins and ICAM-1 modulate neutrophil-endothelial adhesionand diapedesis in vitro. Inflammation 21: 313–324.
53. Muller, W. A. 2011. Mechanisms of leukocyte transendothelial migration. Annu.Rev. Pathol. 6: 323–344.
54. Coombe, D. R., S. M. Stevenson, B. F. Kinnear, N. S. Gandhi, R. L. Mancera,R. I. Osmond, and W. C. Kett. 2008. Platelet endothelial cell adhesion molecule1 (PECAM-1) and its interactions with glycosaminoglycans: 2. Biochemicalanalyses. Biochemistry 47: 4863–4875.
55. DeLisser, H. M., H. C. Van, P. J. Newman, W. A. Muller, C. A. Buck, andS. M. Albelda. 1993. Platelet/endothelial cell adhesion molecule-I (CD31)-mediated cellular aggregation involves cell surface glycosaminoglycans. J.Biol. Chem. 268: 16037–16046.
56. Yeaman, C., and A. C. Rapraeger. 1993. Post-transcriptional regulation ofsyndecan-1 expression by cAMP in peritoneal macrophages. J. Cell Biol. 122:941–950.
The Journal of Immunology 6119
by guest on October 5, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from





























