Embed Size (px)
Citation preview

Molec. gen. Genet. 128, 1--9 (197~) © by Springer-Verlag 1974
The Ribosomes of Drosophila I . S u b u n i t and Protein Compos i t ion*
E d w a r d Berger
Department of Biology, State University of New York at Albany, Albany, New York
Received July 3, 1973
Summary. Functional ribosomes from Drosophila virilis dissociate to form 40S and 55S subunits. One-dimensional gel electrophoresis in either dodeeyl sulfate-acrylamide gels (Fig. 1) or urea-acrylamide gels (Fig. 2) reveal complex protein patterns. Increased resolution in two- dimensional acrylamide gels (Figs. 3 and 4) indicate the existence of at least 28 proteins in the small subunit, and 39 proteins in the large subunit. Molecular weight estimates indicate a mole- cular weight range (and average) of 16500-40000 (25800) for the 40S subunit proteins, and 12500-45000 (24300) for the 55S subunit proteins. Comparison of larval and adult ribosomal protein patterns did not reveal qualitative differences between them.
1. Introduction
I t is known t h a t in E. coli most, if no t all, of the prote ins conta ined in r ibosomes are genet ica l ly c lus tered (Dekio, T a k a t a and Osawa, 1970) and under coordinate syn the t i c control (Nomura and Engback , 1972). Studies on anucleola te m u t a n t s of Xenopus (Hal lberg and Brown, 1969) reveal t h a t in the ~bsence of r ibosomal R N A genes the normal onset of r ibosomal pro te in synthesis , dur ing gas t ru la t ion , fails to occu r suggest ing t h a t a coordina te mechanism regula t ing r ibosome bio- genesis m a y exist in eucaryotes as well. A repor t has appea red describing non-co- o rd ina te control of r ibosomal R~NA and r ibosomal p ro te in synthesis in h u m a n L- cells (Craig and Per ry , 1971), however the au thors po in t out t h a t because of numerous differences be tween the two exper imenta l systems the results are no t comparable .
I n an effort to fur ther explore the quest ion of coordinate control in higher eucaryotes a de ta i led s t ruc tura l and genet ic analysis of Drosophila r ibosomes was begun. The purpose of this r epor t is to describe the me thodo logy of r ibosome iso- la t ion, dissociat ion, and p ro te in ext rac t ion , for Drosophila, and provide a prel imi- n a r y phys iochemical descr ip t ion of the subuni t pro te ins b y e lec t rophoret ic proce- dures.
2. Methods and Materials
a) Ribosome Isolation and Dissociation
Drosophila virilis adults or third instar larvae grown on banana-agar medium were homo- genized in a buffer containing 50 m ~ Tris-HC1, 250 mM sucrose, i m ~ MgC12, 25 mM KCI, 1 mM dithiothreitol, pH 7.6, at 0 ° C. The slurry was centrifuged at 18000 x g, for 15 minutes at
* This work was supported in part by a grant from the National Institutes of Health (GM- 18910).
1 Molec. gen. Genet. 128

2 E. Berger
4 ° C in a Sorval RC-2B centrifuge and the supernatant filtered through cotton gauze to remove lipid aggregates. The centrifugation was repeated to completely remove nuclei and mite- chondria. The supernatant was then made 0.5% with respect to deoxycholate (sodium salt) and centrifuged at 27 000 × g for 20 minutes and the pellet discarded. The clear supernatant was next centrifuged at 109000 × g for 2 hours at 4 ° C, in an International B-60 ultracentri- fuge. The crude ribosomal pellet was gently resuspended in the homogenizing buffer modified to contain 10 mM MgC1 a and clarified by centrifugation at 27000 × g for 10 minutes. The super- natant was next layered on a discontinuous sucrose gradient (2 ml of 30% sucrose under 3 ml of 10% sucrose) containing the modified homogenizing buffer, and centrifuged at 109000 × g for 3 hours at 4 ° C. The washed ribosomal pellet was finally resuspended in a dissociation buffer containing 20 mM Tris-HC1, 100 mM KC1, pH 7.4, layered on a 35 ml continuous sucrose gradient (10% to 30% ) containing dissociation buffer, and centrifuged at 42000 × g 15.5 hours, at 4 ° C, in an International SB- l i0 swinging bucket rotor.
b) Ribosomal Protein Isolation Following centrifugation gradients were fractionized in an Isco Model 640 instrument and
the appropriate fractions pooled. An equal volume of ice cold 20% trichloroacetic acid (TCA) was added and incubated on ice for an hour. The tubes were then centrifuged at 18000 × g for 20 minutes, and the subunit pellets washed in acetone, 95% ethanol, and anhydrous ether, and finally dehydrated by evacuation.
To extract the ribosomal proteins the pellets were resuspended overnight in a buffer con- taining 10 mM Tris-ttC1, pH 7.4, and next treated overnight with two volumes of glacial acetic acid containing 100 mM MgCle. All operations were carried out at 4 ° C. The preparation was then centrifuged at 18000 × g for 30 minutes and the RNA pellet discarded. Protein was pre- cipitated by the addition of 5 volumes of acetone, and after an hour on ice the suspension was centrifuged at 18000 × g for 30 minutes. The protein pellet was dehydrated by successive washing in acetone, 95% ethanol, and anhydrous ether, and stored at - -20 ° C.
e) Eleetrophoresis Protein pellets were resuspended in 4 M urea (deionized) and the protein concentration
determined by the Folin procedure (Lowry, Rosebrough, Farr, and Randall, 1951) using re- crystallized bovine serum albumin as a standard. For one-dimensional electrophoresis in dodecylsulfate acrylamide slab gels, a modification of the technique of Blattler, Garner, Slyke, and Bradley (1972) was employed. For one-dimensional electrophoresis in urea-acrylamide disc gels the procedure of Lambertsson, Rasmuson, and Bloom (1970) was used. For two-dimen- sional electrophoresis proteins were first separated in urea-acrylamide cylinders, and the disc polymerized on to a dodecylsulfate-acrylamide slab gel prepared according to the procedure of Weber and Osborn (1969). In all cases proteins were visualized, following electrophoresis, by staining in a 0.1% solution of Coomassie Brilliant Blue dissolved in 50 % TCA, and destained in a 5:5:1 mixture of methanol, water, acetic acid. Exact conditions of electrophoresis are described in individual figure legends.
3. Results
a) Properties o] Monosome8 and Subunits
R i b o s o m e s i so la t ed f r o m adu l t Drosophila virilis were f o u n d t o con t a in 50 %
pro te in , b y weigh t . Op t i ca l d e n s i t y d e t e r m i n a t i o n s r e v e a l e d an OD~60/OD~s 0
ra t io of 1.87 a n d OD~0/OD235 ra t io of 1.50, wel l w i t h i n t h e a c c e p t e d l imi t s of
p u r i t y for e u c a r y o t i c r ibosomes (Mart in , Ro l l e s ton , Low, a n d Wool , 1969). Ut i l i -
z ing E. coli 70 S monosomes , and 30 S and 50 S r i bosoma l subun i t s as pa ra l l e l s t anda rds , Drosophila m o n o s o m e s were f o u n d to h a v e a s e d i m e n t a t i o n coeff ic ient of 80 S. S tud ies on t h e b i o s y n t h e t i c a c t i v i t y of our 80 S p r e p a r a t i o n s were ca r r i ed
Drosophila Ribosomes 3
a b c d e f g h i j k 1
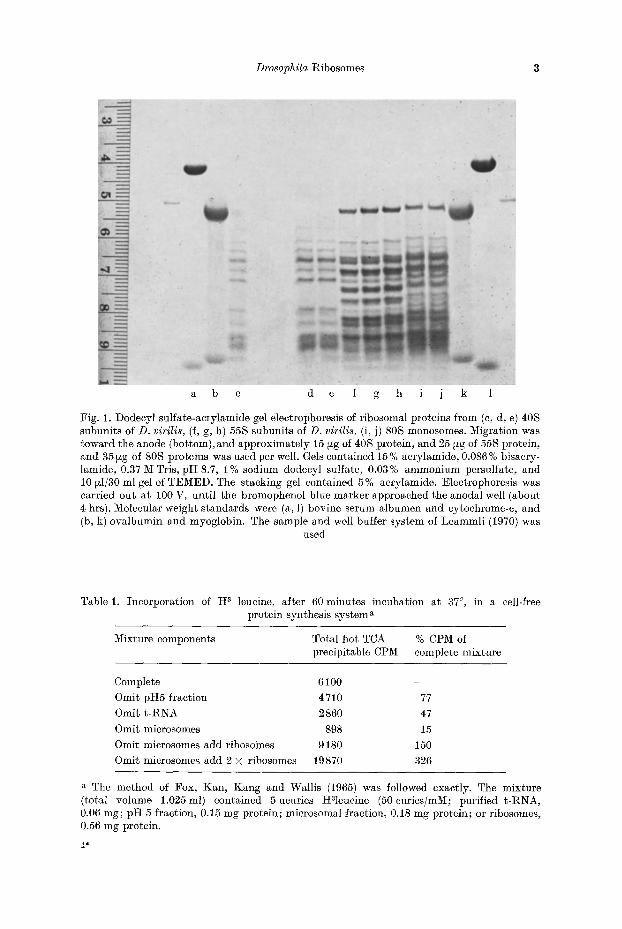
Fig. 1. Dodeeyl sulfate-acrylamide gel electrophoresis of ribosomal proteins from (c, d, e) 40S subunits of D. virilis, (f, g, h) 55S subunits of D. virilis, (i, j) 80S monosomes. Migration was toward the anode (bottom), and approximately 15 ~g of 40S protein, and 25 ~g of 55S protein, and 35 ~zg of 80S proteins was used per well. Gels contained 15 % aerylamide, 0.086% bisacry- lamide, 0.37 M Tris, pH 8.7, 1% sodium dodecyl sulfate, 0.03% ammonium persulfate, and 10 ~I/30 ml gel of TEMED. The stacking gel contained 5% acrylamide. Electrophoresis was carried out at 100 V, until the bromophenol blue marker approached the anodal well (about 4 hrs). Molecular weight standards were (a, 1) bovine serum albumen and cytochrome-e, and (b, k) ovalbumin and myoglobin. The sample and well buffer system of Leammli (1970) was
used
Table 1. Incorporation of H a ]eucine, after 60 minutes incubation at 37 °, in a cell-free protein synthesis system a
Mixture components Total hot TCA % CPM of precipitable CPM complete mixture
Complete 6100 - -
Omit pH5 fraction 4710 77
Omit t -RNA 2860 47
Omit microsomes 898 15
Omit microsomes add ribosomes 9180 150
Omit microsomes add 2 × ribosomes 19870 326
a The method of Fox, Kan, Kang and Wallis (1965) was followed exactly. The mixture (total volume 1.025ml) contained 5ucuries tIZleucine (50curies/raM; purified t-RNA, 0.06 mg; pH 5 fraction, 0.15 mg protein; microsomal fraction, 0.18 mg protein; or ribosomes, 0.56 mg protein.
1"

4 E. Berger
out in a cell free protein synthesizing system (Fox, Kan, Kang, and Wallis, 1965). The results in Table 1 indicate that our preparations are active in stimulating labelled amino acid incorporation into TCA precipitable material. Essentially identical results were obtained when the source of ribosomes was Drosophila virilis larvae, or D. melanogaster adults or larvae.
Ribosomes resuspended in a Tris HC1 buffer containing 100 mM KC1 dissociate into subunits having sedimentation coefficients of 40 S and 55 S, with respect to E. coli standards. The optical density ratios of these subunits were : OD260/OD2s 0 of 1.93 for the 40 S subunit ; OD260/OD2s 0 of 1.79 for the 55 S subunit.
b) Dodeeyl Sul/ate-acrylamide Electrophoresis
Analysis of 40 S subunit proteins in 15% acrylamide gels containing 0.1% SDS revealed the presence of 14 major bands and several minor components (Fig. 1). Electropherograms of 55 S subunit proteins were found to contain 20 major bands and several minor components. The molecular weights of ribosomal proteins ranged from about 12000 to 40000 with the exception of a single large subunit band of about 48000 molecular weight. Protein standards utilized for molecular weight estimations in these gels are in Fig. 1.
When proteins were separated in 20 % acrylamide gels, several changes in the pattern appear. The most prominent of these is the resolution of two sharp bands from the high molecular weight component of the 55 S subunit. Molecular weight estimates for these proteins, determined from protein standards included in the gel, were 48000 and 50000. From the variable staining intensity of bands in either subunit it would appear that many of the bands resolved in one dimension contain a number of different proteins.
c) Urea-acrylamide Gel Electrophoresis
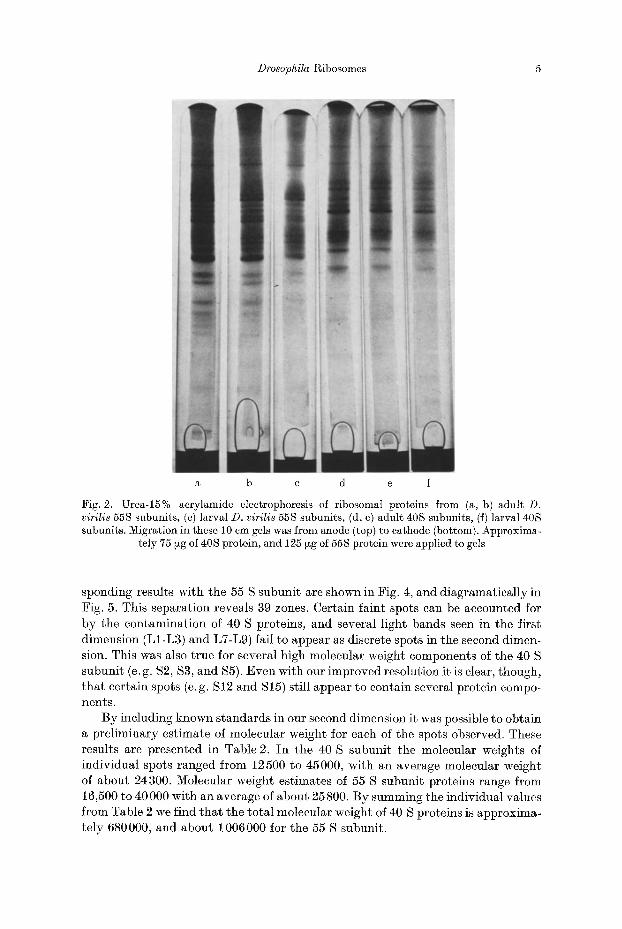
Ribosomal subunit proteins were next studied in 15% acrylamide gels con- taining 2.5 M Urea. Under these conditions electropherograms of 40 S subunit proteins were found to contain 25 major bands (Fig. 2d, e, and diagramatically in Fig. 5), while gels containing 55 S subunit proteins showed 33 major bands (Fig. 2a, b, and diagramatically in Fig. 5). Again the variable staining intensity of bands in both subunits indicates that electrophoresis in one-dimension fails to resolve the total protein complement. Included in Fig. 2 are gels containing 40 S and 55 S proteins extracted from D. virilis larvae. A comparison of the adult and larval ribosomal subunit patterns using our procedure revealed no stage-specific differences.
d) Two-dimensional Electrophoresis
Two-dimensional electrophoresis provided significantly higher resolution than either one-dimensional procedure. In our procedure protein separation is made by charge, in the first dimension, and then by molecular weight in the second dimen- sion. Results obtained with 40 S subunit proteins are shown in Fig. 3, and dia- gramatically in Fig. 5. The pattern consists of 29 zones which can be alligned, in position, with most of the bands resolved in urea-acrylamide disc gels. Corre-
Drosophila Ribosomes 5
a b c d e f
Fig. 2. Urea-15% acrylamide electrophoresis of ribosomal proteins from (a, b) adult D. virilis 55S subunits, (c) larval D. virilis 55S subunits, (d, e) adult 40S subunits, (f) larval 40S subunits. Migration in these 10 cm gels was from anode (top) to cathode (bottom). Approxima-
tely 75 ~zg of 40S protein, and 125 ~g of 55S protein were applied to gels
sponding results with the 55 S subunit are shown in Fig. 4, and diagramatieally in Fig. 5. This separation reveals 39 zones. Certain faint spots can be accounted for by the contaminat ion of 40 S proteins, and several light bands seen in the first dimension (L1-L3) and L7-L9) fail to appear as discrete spots in the second dimen- sion. This was also t rue for several high molecular weight components of the 40 S subunit (e. g. $2, $3, and $5). Even with our improved resolution it is clear, though, tha t certain spots (e. g. S12 and S15) still appear to contain several protein compo- nents.
By including known standards in our second dimension it was possible to obtain a prel iminary estimate of molecular weight for each of the spots observed. These results are presented in Table 2. I n the 40 S subunit the molecular weights of individual spots ranged from 12500 to 45000, with an average molecular weight of about 24300. Molecular weight estimates of 55 S subunit proteins range from 16,500 to 40 000 with an average of about 25 800. By summing the individual values f rom Table 2 we find tha t the total molecular weight of 40 S proteins is approxima- tely 680000, and about 1006000 for the 55 S subunit.
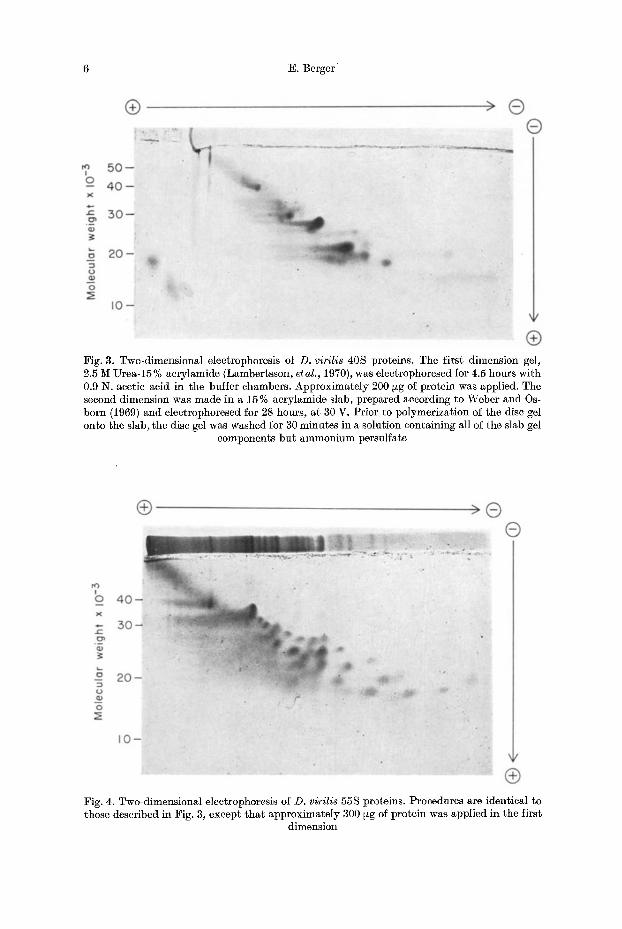
6 E. Berger
Fig. 3. Two-dimensional electrophoresis of D. virilis 40S proteins. The first dimension gel, 2.5 M Urea-15 % acrylamide (Lambertsson, et al., 1970), was electrophoresed for 4.5 hours with 0.9 N. acetic acid in the buffer chambers. Approximately 200 ~zg of protein was applied. The second dimension was made in a 15% acrylamide slab, prepared according to Weber and Os- born (1969) and electrophoresed for 28 hours, at 30 V. Prior to polymerization of the disc gel onto the slab, the disc gel was washed for 30 minutes in a solution containing all of the slab gel
components but ammonium persulfate
Fig. 4. Two-dimensional eleetrophoresis of D. virilis 55S proteins. Procedures are identical to those described in Fig. 3, except that approximately 300 ~g of protein was applied in the first
dimension
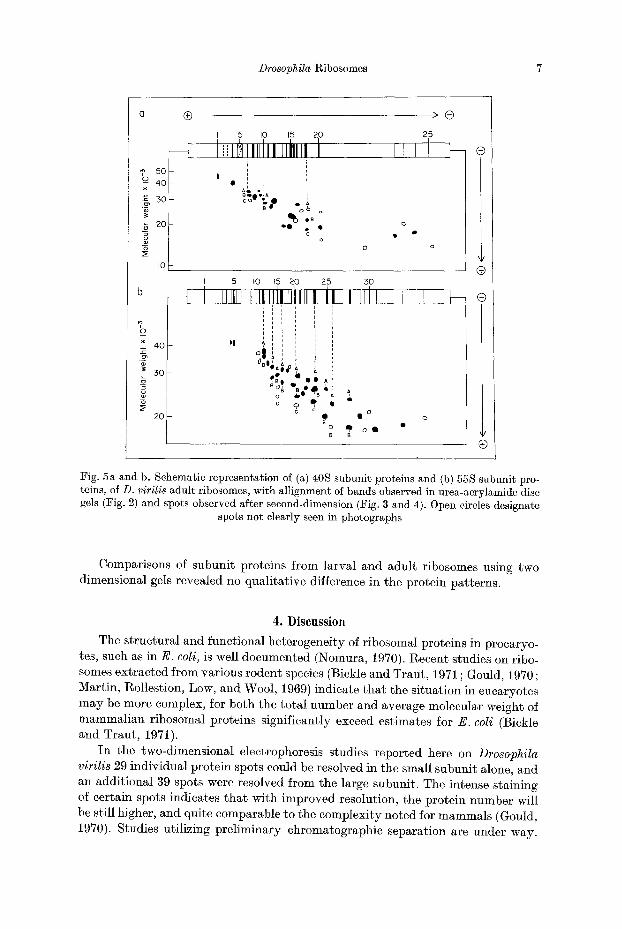
Drosophila Ribosomes 7
5O o 40 7
30
20
io
× 40
~ 3o
2O
®
I 5 io 15 20
l , IIillillllt lll, it I i
B~CC)~ e~l.A i - . - . ,~
o o
' ,o ~ • o
> ®
25
iIi1 >
0
o
5 io 15 20 25 30
IIllli!filtlttllltltltl it i lLJl i I , , r
i i i i t i q p t t
i E , i i i I i i
o : , : , W A e ~ A a F
%-, • , i ~! Bog O e e d l QI A
o ~ $ 'B A ,
o • o l B B
Q
®
®
$ ®
Fig. 5a and b. Schematic representation of (a) 40S subunit proteins and (b) 55S subunit pro- teins, of D. virilis adult ribosomes, with Mlignment o~ bands observed in urea-acrylamide disc gels (Fig. 2) and spots observed after second-dimension (Fig. 3 and 4). Open circles designate
spots not clearly seen in photographs
Comparisons of subunit proteins f rom larval and adult ribosomes using two dimensional gels revealed no qualitative difference in the protein patterns.
4 . D i s c u s s i o n
The structural and functional heterogeneity of ribosomal proteins in procaryo- tes, such as in E. coli, is well documented (Nomura, 1970). Recent studies on ribo- somes extracted from various rodent species (Bickle and Traut , 1971 ; Gould, 1970 ; Martin, Rollestion, Low, and Wool, 1969) indicate tha t the si tuation in eucaryotes m a y be more complex, for both the total number and average molecular weight of mammal ian ribosomal proteins significantly exceed estimates for E. coli (Bickle and Traut , 1971).
I n the two-dimensional electrophoresis studies reported here on Drosophila virilis 29 individual protein spots could be resolved in the small subunit alone, and an additional 39 spots were resolved from the large subunit. The intense staining of certain spots indicates t ha t with improved resolution, the protein number will be still higher, and quite comparable to the complexity noted for mammals (Gould, 1970). Studies utilizing prel iminary chromatographic separation are under way.
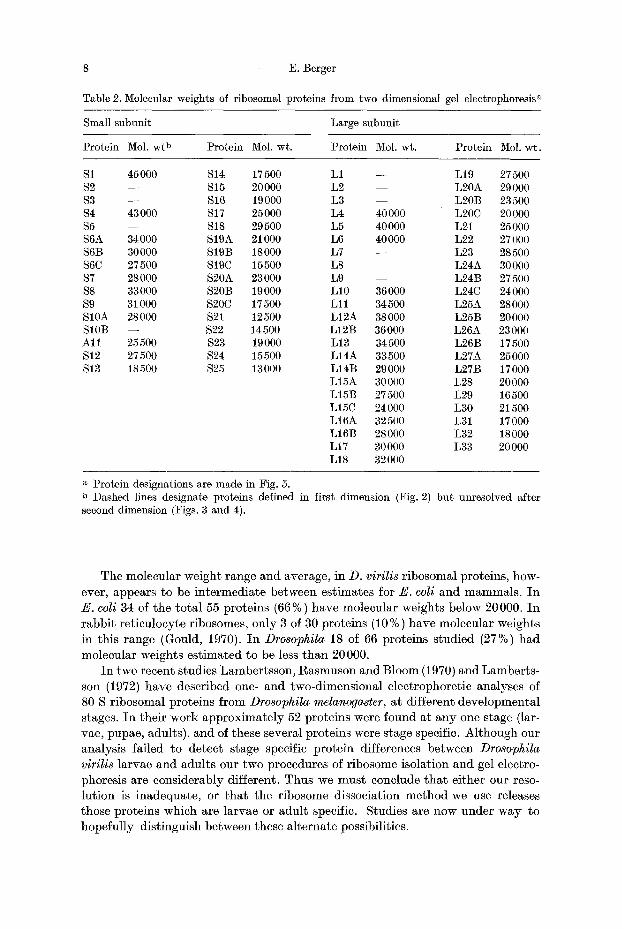
8 E. Berger
Table 2. Molecular weights of ribosomal proteins from two dimensional gel electrophoresis a
Small subunit Large subunit
Protein Mol. wt b Protein Mol. wt. Protein Mol. wt. Protein Mol. wt.
$1 45000 S14 17500 L1 - - L19 27500 $2 - - $15 20000 L2 - - L20A 29000 $3 - - $16 19000 L3 - - L20B 23500 $4 43000 $17 25000 L4 40000 L20C 20000 $5 - - $18 29500 L5 40000 L21 25000 S6A 34000 $19A 21000 L6 40000 L22 27000 S6B 30000 S19B 18000 L7 - - L23 28500 $6C 27500 $19C 15500 L8 - - L24A 30000 $7 28000 S20A 23000 L9 - - L24B 27500 $8 33000 S20B 19000 L10 36000 L24C 24000 $9 31000 $20C 17500 L11 34500 L25A 28000 S10A 28000 $21 12500 L12A 38000 L25B 20000 S10B - - $22 14500 L12B 36000 L26A 23000 Al l 25500 $23 19000 L13 34500 L26B 17500 $12 27500 $24 15500 L14A 33500 L27A 25000 S 13 18500 $25 13000 L14B 29000 L27B 17000
L15A 30000 L28 20000 L15B 27500 L29 16500 L15C 24 000 L30 21500 L16A 32500 L31 17000 L16B 28000 L32 18000 L17 30000 L33 20000 L18 32000
a Protein designations are made in Fig. 5. b Dashed lines designate proteins defined in first dimension (Fig. 2) but unresolved after second dimension (Figs. 3 and 4).
The molecular weight range and average, in D. virilis r ibosomal proteins, how-
ever, appears to be in te rmedia te be tween est imates for E. cell and mammals . I n
E. cell 34 of the to ta l 55 proteins (66%) have molecular weights below 20000. In
rabbi t re t iculocyte ribosomes, only 3 of 30 proteins (10 %) have molecular weights
in this range (Gould, 1970). I n Drosophila 18 of 66 proteins studied (27%) had
molecular weights es t imated to be less t han 20 000.
I n two recent studies Lambertsson, Rasmuson and Bloom (1970) and Lamber ts -
son (1972) have described one- and two-dimensional electrophoret ic analyses of 80 S r ibosomal proteins f rom Drosophila melanogaster, at different deve lopmenta l
stages. I n their work approx imate ly 52 proteins were found at any one stage (lar-
vae, pupae, adults), and of these several proteins were stage specific. Al though our
analysis failed to detec t stage specific prote in differences between Drosophila virilis la rvae and adults our two procedures of r ibosome isolation and gel electro-
phoresis are considerably different. Thus we must conclude t h a t ei ther our reso-
lut ion is inadequate , or t h a t the r ibosome dissociation me thod we use releases those proteins which are larvae or adul t specific. Studies are now under way to
hopeful ly dist inguish between these a l ternate possibilities.

Drosophila Ribosomes 9
Acknowledgements. The author is indebted to Linda Salai and Deirdre Berger for their able technical assistance, and to Dr. David Shub for his valuable advice.
References
Bickle, T. A., Traut, R. 1%. : Differences in size and number of 80 S and 70 S ribosomal proteins by dodecyl sulfate gel electrophoresis. J. biol. Chem. 246, 6828-6834 (1971)
Blattler, D. P., Garner, :F., VanSlyke, K., Bradley, A. : Quantitative electrophoresis in poly- acrylamide gels of 240%. J. Chromatography 64, 147-155 (1972)
Craig, N., Perry, R. P.: Persistent cytoplasmic synthesis of ribosomal proteins during the selective inhibition of ribosomal RNA synthesis. Nature (Lond.)New Biol. 229, 75-80(1971)
Dekio, S., Takata, R., Osawa, S. : Genetic studies of the ribosomal proteins in Escherichia coll. VI. Determination of chromosomal loci for several ribosomal protein components using a hybrid strain between Escherichia coli and Salmonella typhimurium. Molec. gen. Genet. 109, 131-141 (1970)
Fox, A. S., Kan, J., Kang, S. H., Wallis, B. : Protein synthesis in cell-free preparations from Drosophila melanogaster. J. biol. Chem. 240, 2059-2065 (1965)
Gould, H. J.: Proteins of rabbit reticulocyte ribosomal subunits. Nature (Lond.) 227, 1145- 1147 (1970)
Hallberg, R. L., Brown, D. D. : Co-ordinated synthesis of some ribosomal proteins and ribo- somM RNA in embryos of Xenopus laevis. J. molec. Biol. 46, 393 411 (1969)
Lambertsson, A. G. : The ribosomal proteins of Drosophila melanogaster. II. Comparisons of protein patterns of ribosomes from larvae, pupae and adult flies by two-dimensional poly- acrylamide gel electrophoresis. Molee. gen. Genet. 118, 215-222 (1972)
Lambertsson, A. G., Rasmuson, S. B., Bloom, G. D.: The ribosomal proteins of Drosophila melanogaster. I. Characterization in polyacrylamide gels of proteins from larval, adult, and ammonium chloride-treated ribosomes. Molec. gen. Genet. 108, 349-357 (1970)
Leammli, U. K. : Cleavage of structural proteins during the assembly of the head of bacterio- phage T4. Nature (Lond.) 207, 680-685 (1970)
Lowry, O. H., Rosebrough, N. J., Farr, A. L., Randall, R. J. : Protein measurement with the Folin phenol reagent. J. biol. Chem. 193, 265-275 (1951)
martin, T. E., Rolleston, F. S., Low, R. B., Wool, I. G.: Dissociation and reassociation of skeletal muscle ribosomes. J. molec. Biol. 43, 135-149 (1969)
Nomura, M. : Bacterial ribosome. Bact. Rev. 34, 228-277 (1970) Nomura, M., Engback, F. : Expression of ribosomal genes as analyzed by bacteriophage MU-
induced mutations. Proc. nat. Acad. Sci. (Wash.) 69, 1526-1530 (1972) Weber, K., Osborn, M. : The reliability of molecular weight determinations by dodecyl sulfate-
polyacrylamide gel electrophoresis. J. biol. Chem. 244, 4406-4412 (1969)
Communicated by H. G. W i t t m a n n
Dr. Edward Berger Department of Biology State University of New York at Albany Albany, New York 12222 USA