Embed Size (px)
Citation preview

The Retinoblastoma Tumor Suppressor Promotes Efficient HumanCytomegalovirus Lytic Replication
Halena R. VanDeusen, Robert F. Kalejta
Institute for Molecular Virology and McArdle Laboratory for Cancer Research, University of Wisconsin—Madison, Madison, Wisconsin, USA
ABSTRACT
The retinoblastoma (Rb) tumor suppressor controls cell cycle, DNA damage, apoptotic, and metabolic pathways. DNA tumorvirus oncoproteins reduce Rb function by either inducing Rb degradation or physically disrupting complexes between Rb and itsmyriad binding proteins. Human cytomegalovirus (HCMV), a betaherpesvirus being investigated for potential roles in humancancers, encodes multiple lytic-phase proteins that inactivate Rb in distinct ways, leading to the hypothesis that reduced Rb lev-els and/or activity would benefit HCMV lytic infection. Paradoxically, we found that Rb knockdown prior to infection, whethertransient or constitutive, impaired HCMV lytic infection at multiple stages, notably viral DNA replication, late protein expres-sion, and infectious virion production. The existence of differentially modified forms of Rb, the temporally and functionally dis-tinct means by which HCMV proteins interact with Rb, and the necessity of Rb for efficient HCMV lytic replication combine tohighlight the complex relationship between the virus and this critical tumor suppressor.
IMPORTANCE
Initial work examining viral protein modulation of cell cycle progression and oncogenic transformation revealed that these pro-teins inactivated the function of cellular tumor suppressor proteins. However, subsequent work, including experiments de-scribed here using human cytomegalovirus, demonstrate a more nuanced interaction that includes the necessity of cellular tu-mor suppressors for efficient viral replication. Understanding the positive impacts that cellular tumor suppressors have on viralinfections may reveal new activities of these well-studied yet incompletely understood proteins. The basis for oncolytic viraltherapy is the selective replication of viruses in transformed cells in which tumor suppressor function may be compromised.Understanding how tumor suppressors support viral infections may allow for the generation of modified oncolytic viruses withgreater selective tumor cell replication and killing.
The retinoblastoma (Rb) protein is a tumor suppressor (1, 2).Loss of both Rb alleles predisposes patients to the develop-
ment of cancer (3). Rb, through its association with more than 200other cellular proteins (4), controls pathways that regulate cellcycle progression, DNA repair, apoptosis, and energy metabolism,all of which are intimately involved in oncogenic transformationand tumor cell survival (5–7). Most, if not all, human tumors havedefects (mutations) in one or more components of the pathwayscontrolled by Rb (8).
The unphosphorylated or hypophosphorylated form of Rb isgenerally considered the active form of the protein (9). Hypo-phosphorylated Rb interacts with many cellular proteins, includ-ing a critical association with the E2F family of transcription fac-tors (10). E2F transcription factors control the expression of manygenes required for cell cycle progression, and Rb binding inhibitsE2F-dependent transcription (11). Rb binding to E2F protectscells from untimely progression through the cell cycle and pre-vents E2F-mediated oncogenic transformation (12, 13). Duringnormal cell cycle progression, a series of cellular cyclin-dependentkinases (Cdks) phosphorylate Rb, converting it into a fully phos-phorylated form, termed hyperphosphorylated Rb. This form isconsidered inactive (14), although it may retain some unrecog-nized function (15). Hyperphosphorylated Rb no longer bindsE2F and thus permits E2F-dependent transcription and cell cycleprogression (10). Recently, Cdk-dependent monophosphoryla-tion of Rb has been reported (16), but the physiological relevanceof this is unknown. Rb can also be acetylated, methylated,SUMOylated, ubiquitinated, and phosphorylated on non-Cdk-
mediated sites in response to stimuli that may activate non-cell-cycle-associated functions of Rb (17).
In addition to being a tumor suppressor, Rb might also be a“virus suppressor,” at least for the DNA tumor virus human pap-illomavirus (HPV). The HPV E7 protein binds to Rb and inducesits proteasomal degradation (18, 19). E7 proteins unable to bindor degrade Rb are unable to support productive papillomavirusreplication (20, 21). However, as E7 Rb-binding-deficient mu-tants have other defects (22), it is premature to conclude the in-ability to degrade Rb is the only reason for the observed defects inthe viral life cycle. Unfortunately, the role of Rb during HPV in-fection remains unclear due to difficulties in studying productiveHPV replication in vitro (through organotypic raft cultures) andthe justifiable focus on the essential role of E7-mediated inactiva-tion of Rb during HPV-induced cellular transformation and hu-man cancers.
Received 20 January 2015 Accepted 13 February 2015
Accepted manuscript posted online 18 February 2015
Citation VanDeusen HR, Kalejta RF. 2015. The retinoblastoma tumor suppressorpromotes efficient human cytomegalovirus lytic replication. J Virol 89:5012–5021.doi:10.1128/JVI.00175-15.
Editor: K. Frueh
Address correspondence to Robert F. Kalejta, [email protected].
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00175-15
5012 jvi.asm.org May 2015 Volume 89 Number 9Journal of Virology

Adenovirus, another DNA tumor virus, encodes the E1A pro-tein that binds Rb and disrupts its complexes with E2F (23). E1Amutants unable to bind Rb display only modest defects in viralreplication (24). Therefore, Rb does not appear to be a suppressorof adenovirus. In fact, during adenoviral infection, Rb-E2F1 com-plexes selectively remain intact (25), and recently E1A-Rb com-plexes were demonstrated to suppress the transcription of geneswith antiviral functions in adenovirus-infected cells (26). Thus,Rb might contribute to adenovirus infection in a positive way,although this remains to be determined through knockdown orknockout studies.
Human cytomegalovirus (HCMV) is a DNA virus that is beingexplored as a potential cofactor for human cancers, most notablyglioblastoma brain tumors (27–30). One possible mechanism bywhich HCMV could contribute to cellular transformation is ma-nipulation of the cell cycle through inactivation of Rb (28).HCMV has multiple, potentially semiredundant ways in which tomodulate Rb protein function during lytic infection. The viralpp71 protein is delivered to cells by infecting virions where it bindsto and induces the proteasomal degradation of hypophosphory-lated Rb (31, 32). This pp71-mediated degradation occurs onlywithin the first hours of infection, and Rb reaccumulates laterduring infection (33). The biological relevance of this degradationremains unclear, as a pp71 mutant virus unable to induce Rbdegradation supports wild-type levels of lytic infection in fibro-blasts (34). The viral UL97 protein is a viral cyclin-dependentkinase (v-Cdk) that phosphorylates Rb on several Cdk target sites(33, 35). A mutant virus lacking UL97 fails to phosphorylate Rband has a growth defect, and it should also fail to phosphorylateother known cellular and viral targets (36). Mutants unable tophosphorylate Rb but retaining kinase activity have not been re-ported. The viral IE1 and IE2 proteins have also been reported toactivate E2F-responsive promoters (37–39), though mechanismshave not been explored and evidence for Rb inactivation by IE1and IE2 during viral infection is lacking.
With at least two and perhaps as many as four ways to inacti-vate the Rb protein, we suspected that Rb would suppress HCMVinfection. However, we found that Rb knockdown, either tran-siently by small interfering RNA (siRNA) transfection or consti-tutively through short hairpin RNA (shRNA) expression, im-paired HCMV lytic replication in fibroblasts. HCMV viral DNAreplication and late gene expression were decreased, and as manyas 10-fold-fewer infectious virions were produced. We concludethat the Rb protein is required for efficient HCMV replication andthat the multiple ways in which HCMV interacts with Rb repre-sent viral attempts not to completely eliminate Rb function but tomodulate it in temporally and likely functionally distinct ways.
MATERIALS AND METHODSCells and viruses. Primary normal human dermal fibroblasts (NHDFs)(Clonetics) used for transient experiments and to derive the stable knock-down populations were cultured in Dulbecco’s modified Eagle medium(DMEM) (Invitrogen and Sigma). THP-1 cells (ATCC TIB-202) werecultured in RPMI 1640 medium (Invitrogen). Both DMEM and RPMI1640 medium were supplemented with 10% (vol/vol) fetal bovine serum(FBS) (Sigma), and 100 U/ml penicillin, 100 �g/ml streptomycin, and0.292 mg/ml glutamine (PSG) (Invitrogen) to generate complete media.Transduction with shRNA retroviruses was performed as previously de-scribed (40). Transduced cells were maintained in complete media con-taining 1 �g/ml puromycin (Sigma) and used for experiments until pas-sage 16. Asynchronous cells were plated at a subconfluent density and
cultured in complete media for 24 h before harvesting or infection. Toarrest cells by serum starvation, cells were plated at 1 � 104 cells/cm2, and24 h after plating, cells were washed twice with Dulbecco’s phosphate-buffered saline (DPBS) (Invitrogen) and then maintained in DMEM con-taining 0.1% FBS supplemented with PSG (0.1% FBS DMEM�PSG)(low-serum medium) for 48 h before harvesting or infection. Confluentcells were fed complete media every 2 days for 144 h after plating beforebeginning contact inhibition cell experiments. All infections usedHCMV-AD169 or an AD169 derivative expressing IE2 fused to greenfluorescent protein (AD169 IE2-GFP, 41) or GFP-expressing HCMVstrain FIX (42). Multiplicities of infection (MOIs) were calculated for eachcell line by counting an extra plated well. Cells were infected in minimalvolumes for 60 min, inocula were removed, and media were replaced.Viral titers were measured by standard plaque assay on nontransducedfibroblasts.
RNA interference. The pSUPER retro puro Scr shRNA (retro standsfor retroviral, puro stands for puromycin, and Scr stands for scrambled)was a gift from John Gurdon (Addgene plasmid 30520) (43), and the Rb2and Rb3 plasmids were previously described (RB-sh2 and RB-sh3 [44]).Transient knockdown was achieved by transfecting cells with 80 pg/cell ofsmall interfering oligonucleotides (small interfering RNA [siRNA]) fromDharmacon (scrambled control [Scr], catalog no. D-001810-01-20;siRNA targeting the coding region of the Rb mRNA [siRb1], 5=-GAA CAGGAG UGC ACG GAU AUU-3=) using Lonza nucleofection reagents (VPI-1002; Lonza) according to the manufacturer’s instructions.
Inhibitors and antibodies. Valproic acid (VPA) (1 mM) (Sigma) dis-solved in water was added to THP-1 cells 3 h prior to infection and main-tained throughout the infection. Antibodies used in these experimentsthat are available from commercial sources are as follows: anti-Rb (4H1;Cell Signaling; catalog no. 554136; BD Pharmingen), anti-p107 (C-18;Santa Cruz), anti-cyclin E (C-19; Santa Cruz), anti-tubulin (clone DM 1A;Sigma), and anti-UL44 (10D8; Virusys). Monoclonal antibodies againstpp71 (2H10-9), IE1 (1B12), and pp150 (CMV127) have been describedpreviously (45). Infrared (IR) dye 680- and 800-conjugated secondaryantibodies (Li-Cor) were used for Western blotting, and Alexa Fluor 488-conjugated secondary antibody (catalog no. A-11017; Molecular Probes;Thermo Fisher Scientific) was used for immunofluorescence.
Western blots. Equal cell numbers were lysed in 1% SDS and 2%�-mercaptoethanol and boiled for 20 min prior to separation by SDS-PAGE and transfer to nitrocellulose membranes (Whatman). Five percentbovine serum albumin (BSA) in Tris-buffered saline with Tween 20(TBST [46]) was used to block membranes and incubate primary andsecondary antibodies. Membranes were washed in TBST and imaged withthe Odyssey Fc imager (Li-Cor). Images were generated, and bands werequantitated with the Image Studio version 2.1.10 software (Li-Cor).
Flow cytometry. After trypsinization, cells were harvested by low-speed centrifugation, washed twice with PBS, and fixed with 70% ethanolat �20°C. Cells were then stained with propidium iodide (PI) (Sigma) 1 hprior to analysis on a FACSCalibur flow cytometer (BD Biosciences). Datawere analyzed with ModFit LT software (Verity Software House).
Quantitative real-time PCR. Total DNA was extracted from equalnumbers of cells using a genomic DNA minikit (IB47202; IBI). DNA wasquantitated using iTaq universal SYBR green supermix (catalog no. 172-5124; Bio-Rad) on an Applied Biosystems 7900HT instrument. ViralDNA was amplified with IE1 primers (47), and cellular genomes weremeasured with glyceraldehyde-3-phosphate dehydrogenase (GAPDH)primers (48).
Indirect immunofluorescence. Cells were plated on glass coverslipsand serum starved for 48 h prior to infection. After infection, cells werefixed with 1% paraformaldehyde in PBS and processed as previously de-scribed (46). Stained cells were visualized with a Nikon Ti-Eclipse invertedwide-field microscope, and images were collected with a CoolSnap HQcamera and Nikon NIS Elements software (v 4.00.03). At least 400 nucleiwere counted per condition.
Rb Is Required for HCMV Replication
May 2015 Volume 89 Number 9 jvi.asm.org 5013Journal of Virology
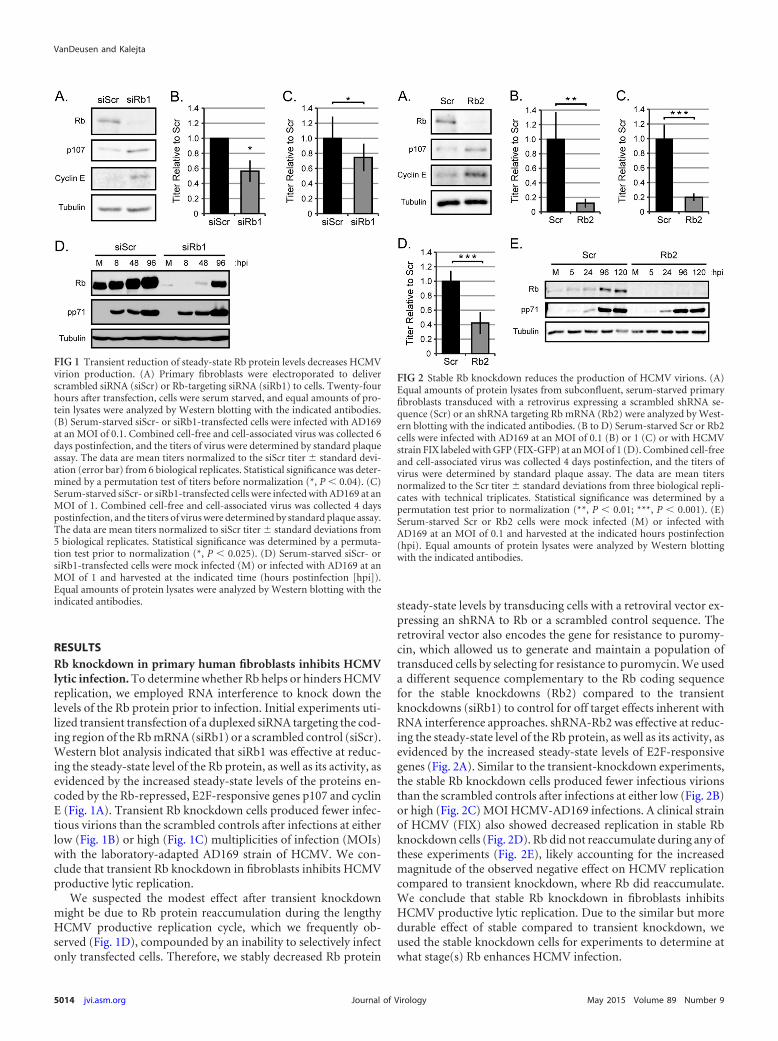
RESULTSRb knockdown in primary human fibroblasts inhibits HCMVlytic infection. To determine whether Rb helps or hinders HCMVreplication, we employed RNA interference to knock down thelevels of the Rb protein prior to infection. Initial experiments uti-lized transient transfection of a duplexed siRNA targeting the cod-ing region of the Rb mRNA (siRb1) or a scrambled control (siScr).Western blot analysis indicated that siRb1 was effective at reduc-ing the steady-state level of the Rb protein, as well as its activity, asevidenced by the increased steady-state levels of the proteins en-coded by the Rb-repressed, E2F-responsive genes p107 and cyclinE (Fig. 1A). Transient Rb knockdown cells produced fewer infec-tious virions than the scrambled controls after infections at eitherlow (Fig. 1B) or high (Fig. 1C) multiplicities of infection (MOIs)with the laboratory-adapted AD169 strain of HCMV. We con-clude that transient Rb knockdown in fibroblasts inhibits HCMVproductive lytic replication.
We suspected the modest effect after transient knockdownmight be due to Rb protein reaccumulation during the lengthyHCMV productive replication cycle, which we frequently ob-served (Fig. 1D), compounded by an inability to selectively infectonly transfected cells. Therefore, we stably decreased Rb protein
steady-state levels by transducing cells with a retroviral vector ex-pressing an shRNA to Rb or a scrambled control sequence. Theretroviral vector also encodes the gene for resistance to puromy-cin, which allowed us to generate and maintain a population oftransduced cells by selecting for resistance to puromycin. We useda different sequence complementary to the Rb coding sequencefor the stable knockdowns (Rb2) compared to the transientknockdowns (siRb1) to control for off target effects inherent withRNA interference approaches. shRNA-Rb2 was effective at reduc-ing the steady-state level of the Rb protein, as well as its activity, asevidenced by the increased steady-state levels of E2F-responsivegenes (Fig. 2A). Similar to the transient-knockdown experiments,the stable Rb knockdown cells produced fewer infectious virionsthan the scrambled controls after infections at either low (Fig. 2B)or high (Fig. 2C) MOI HCMV-AD169 infections. A clinical strainof HCMV (FIX) also showed decreased replication in stable Rbknockdown cells (Fig. 2D). Rb did not reaccumulate during any ofthese experiments (Fig. 2E), likely accounting for the increasedmagnitude of the observed negative effect on HCMV replicationcompared to transient knockdown, where Rb did reaccumulate.We conclude that stable Rb knockdown in fibroblasts inhibitsHCMV productive lytic replication. Due to the similar but moredurable effect of stable compared to transient knockdown, weused the stable knockdown cells for experiments to determine atwhat stage(s) Rb enhances HCMV infection.
FIG 1 Transient reduction of steady-state Rb protein levels decreases HCMVvirion production. (A) Primary fibroblasts were electroporated to deliverscrambled siRNA (siScr) or Rb-targeting siRNA (siRb1) to cells. Twenty-fourhours after transfection, cells were serum starved, and equal amounts of pro-tein lysates were analyzed by Western blotting with the indicated antibodies.(B) Serum-starved siScr- or siRb1-transfected cells were infected with AD169at an MOI of 0.1. Combined cell-free and cell-associated virus was collected 6days postinfection, and the titers of virus were determined by standard plaqueassay. The data are mean titers normalized to the siScr titer � standard devi-ation (error bar) from 6 biological replicates. Statistical significance was deter-mined by a permutation test of titers before normalization (*, P � 0.04). (C)Serum-starved siScr- or siRb1-transfected cells were infected with AD169 at anMOI of 1. Combined cell-free and cell-associated virus was collected 4 dayspostinfection, and the titers of virus were determined by standard plaque assay.The data are mean titers normalized to siScr titer � standard deviations from5 biological replicates. Statistical significance was determined by a permuta-tion test prior to normalization (*, P � 0.025). (D) Serum-starved siScr- orsiRb1-transfected cells were mock infected (M) or infected with AD169 at anMOI of 1 and harvested at the indicated time (hours postinfection [hpi]).Equal amounts of protein lysates were analyzed by Western blotting with theindicated antibodies.
FIG 2 Stable Rb knockdown reduces the production of HCMV virions. (A)Equal amounts of protein lysates from subconfluent, serum-starved primaryfibroblasts transduced with a retrovirus expressing a scrambled shRNA se-quence (Scr) or an shRNA targeting Rb mRNA (Rb2) were analyzed by West-ern blotting with the indicated antibodies. (B to D) Serum-starved Scr or Rb2cells were infected with AD169 at an MOI of 0.1 (B) or 1 (C) or with HCMVstrain FIX labeled with GFP (FIX-GFP) at an MOI of 1 (D). Combined cell-freeand cell-associated virus was collected 4 days postinfection, and the titers ofvirus were determined by standard plaque assay. The data are mean titersnormalized to the Scr titer � standard deviations from three biological repli-cates with technical triplicates. Statistical significance was determined by apermutation test prior to normalization (**, P � 0.01; ***, P � 0.001). (E)Serum-starved Scr or Rb2 cells were mock infected (M) or infected withAD169 at an MOI of 0.1 and harvested at the indicated hours postinfection(hpi). Equal amounts of protein lysates were analyzed by Western blottingwith the indicated antibodies.
VanDeusen and Kalejta
5014 jvi.asm.org May 2015 Volume 89 Number 9Journal of Virology
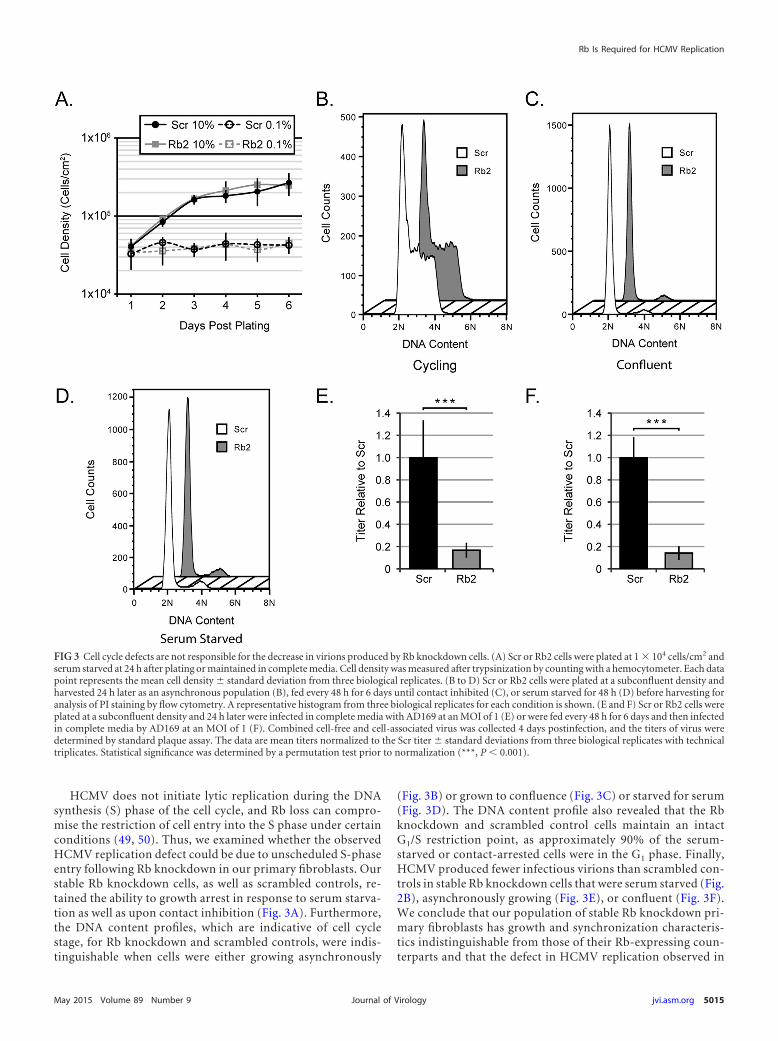
HCMV does not initiate lytic replication during the DNAsynthesis (S) phase of the cell cycle, and Rb loss can compro-mise the restriction of cell entry into the S phase under certainconditions (49, 50). Thus, we examined whether the observedHCMV replication defect could be due to unscheduled S-phaseentry following Rb knockdown in our primary fibroblasts. Ourstable Rb knockdown cells, as well as scrambled controls, re-tained the ability to growth arrest in response to serum starva-tion as well as upon contact inhibition (Fig. 3A). Furthermore,the DNA content profiles, which are indicative of cell cyclestage, for Rb knockdown and scrambled controls, were indis-tinguishable when cells were either growing asynchronously
(Fig. 3B) or grown to confluence (Fig. 3C) or starved for serum(Fig. 3D). The DNA content profile also revealed that the Rbknockdown and scrambled control cells maintain an intactG1/S restriction point, as approximately 90% of the serum-starved or contact-arrested cells were in the G1 phase. Finally,HCMV produced fewer infectious virions than scrambled con-trols in stable Rb knockdown cells that were serum starved (Fig.2B), asynchronously growing (Fig. 3E), or confluent (Fig. 3F).We conclude that our population of stable Rb knockdown pri-mary fibroblasts has growth and synchronization characteris-tics indistinguishable from those of their Rb-expressing coun-terparts and that the defect in HCMV replication observed in
FIG 3 Cell cycle defects are not responsible for the decrease in virions produced by Rb knockdown cells. (A) Scr or Rb2 cells were plated at 1 � 104 cells/cm2 andserum starved at 24 h after plating or maintained in complete media. Cell density was measured after trypsinization by counting with a hemocytometer. Each datapoint represents the mean cell density � standard deviation from three biological replicates. (B to D) Scr or Rb2 cells were plated at a subconfluent density andharvested 24 h later as an asynchronous population (B), fed every 48 h for 6 days until contact inhibited (C), or serum starved for 48 h (D) before harvesting foranalysis of PI staining by flow cytometry. A representative histogram from three biological replicates for each condition is shown. (E and F) Scr or Rb2 cells wereplated at a subconfluent density and 24 h later were infected in complete media with AD169 at an MOI of 1 (E) or were fed every 48 h for 6 days and then infectedin complete media by AD169 at an MOI of 1 (F). Combined cell-free and cell-associated virus was collected 4 days postinfection, and the titers of virus weredetermined by standard plaque assay. The data are mean titers normalized to the Scr titer � standard deviations from three biological replicates with technicaltriplicates. Statistical significance was determined by a permutation test prior to normalization (***, P � 0.001).
Rb Is Required for HCMV Replication
May 2015 Volume 89 Number 9 jvi.asm.org 5015Journal of Virology
the Rb knockdown cells does not result from cell cycle pertur-bations secondary to Rb knockdown.
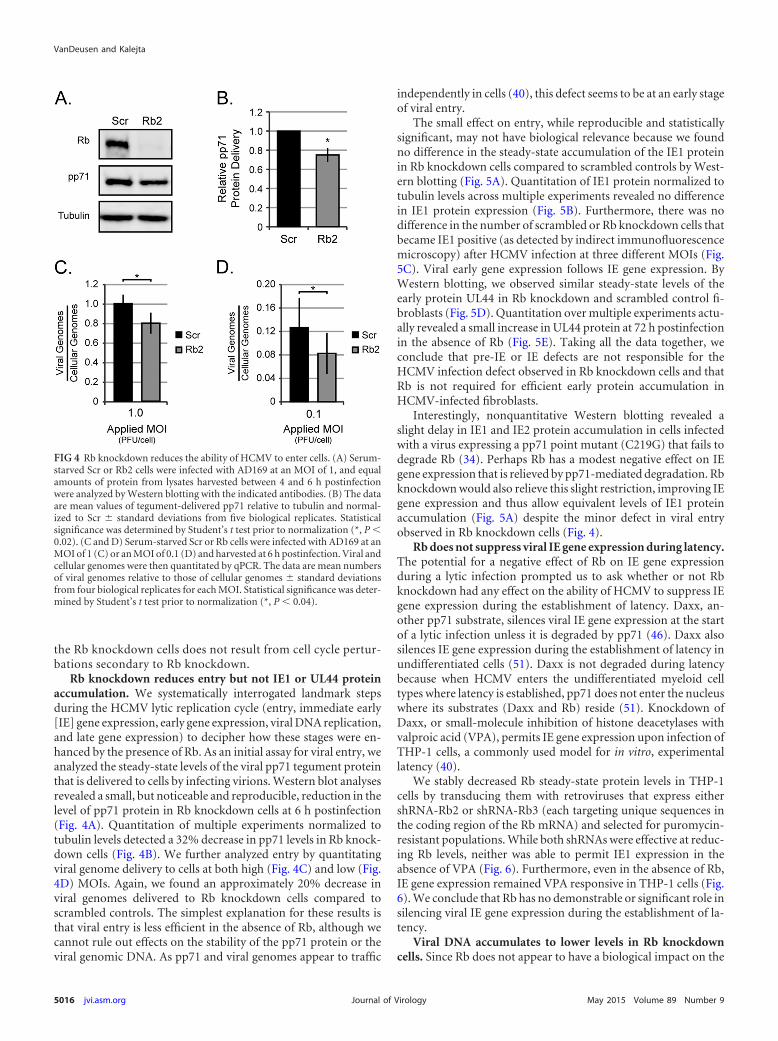
Rb knockdown reduces entry but not IE1 or UL44 proteinaccumulation. We systematically interrogated landmark stepsduring the HCMV lytic replication cycle (entry, immediate early[IE] gene expression, early gene expression, viral DNA replication,and late gene expression) to decipher how these stages were en-hanced by the presence of Rb. As an initial assay for viral entry, weanalyzed the steady-state levels of the viral pp71 tegument proteinthat is delivered to cells by infecting virions. Western blot analysesrevealed a small, but noticeable and reproducible, reduction in thelevel of pp71 protein in Rb knockdown cells at 6 h postinfection(Fig. 4A). Quantitation of multiple experiments normalized totubulin levels detected a 32% decrease in pp71 levels in Rb knock-down cells (Fig. 4B). We further analyzed entry by quantitatingviral genome delivery to cells at both high (Fig. 4C) and low (Fig.4D) MOIs. Again, we found an approximately 20% decrease inviral genomes delivered to Rb knockdown cells compared toscrambled controls. The simplest explanation for these results isthat viral entry is less efficient in the absence of Rb, although wecannot rule out effects on the stability of the pp71 protein or theviral genomic DNA. As pp71 and viral genomes appear to traffic
independently in cells (40), this defect seems to be at an early stageof viral entry.
The small effect on entry, while reproducible and statisticallysignificant, may not have biological relevance because we foundno difference in the steady-state accumulation of the IE1 proteinin Rb knockdown cells compared to scrambled controls by West-ern blotting (Fig. 5A). Quantitation of IE1 protein normalized totubulin levels across multiple experiments revealed no differencein IE1 protein expression (Fig. 5B). Furthermore, there was nodifference in the number of scrambled or Rb knockdown cells thatbecame IE1 positive (as detected by indirect immunofluorescencemicroscopy) after HCMV infection at three different MOIs (Fig.5C). Viral early gene expression follows IE gene expression. ByWestern blotting, we observed similar steady-state levels of theearly protein UL44 in Rb knockdown and scrambled control fi-broblasts (Fig. 5D). Quantitation over multiple experiments actu-ally revealed a small increase in UL44 protein at 72 h postinfectionin the absence of Rb (Fig. 5E). Taking all the data together, weconclude that pre-IE or IE defects are not responsible for theHCMV infection defect observed in Rb knockdown cells and thatRb is not required for efficient early protein accumulation inHCMV-infected fibroblasts.
Interestingly, nonquantitative Western blotting revealed aslight delay in IE1 and IE2 protein accumulation in cells infectedwith a virus expressing a pp71 point mutant (C219G) that fails todegrade Rb (34). Perhaps Rb has a modest negative effect on IEgene expression that is relieved by pp71-mediated degradation. Rbknockdown would also relieve this slight restriction, improving IEgene expression and thus allow equivalent levels of IE1 proteinaccumulation (Fig. 5A) despite the minor defect in viral entryobserved in Rb knockdown cells (Fig. 4).
Rb does not suppress viral IE gene expression during latency.The potential for a negative effect of Rb on IE gene expressionduring a lytic infection prompted us to ask whether or not Rbknockdown had any effect on the ability of HCMV to suppress IEgene expression during the establishment of latency. Daxx, an-other pp71 substrate, silences viral IE gene expression at the startof a lytic infection unless it is degraded by pp71 (46). Daxx alsosilences IE gene expression during the establishment of latency inundifferentiated cells (51). Daxx is not degraded during latencybecause when HCMV enters the undifferentiated myeloid celltypes where latency is established, pp71 does not enter the nucleuswhere its substrates (Daxx and Rb) reside (51). Knockdown ofDaxx, or small-molecule inhibition of histone deacetylases withvalproic acid (VPA), permits IE gene expression upon infection ofTHP-1 cells, a commonly used model for in vitro, experimentallatency (40).
We stably decreased Rb steady-state protein levels in THP-1cells by transducing them with retroviruses that express eithershRNA-Rb2 or shRNA-Rb3 (each targeting unique sequences inthe coding region of the Rb mRNA) and selected for puromycin-resistant populations. While both shRNAs were effective at reduc-ing Rb levels, neither was able to permit IE1 expression in theabsence of VPA (Fig. 6). Furthermore, even in the absence of Rb,IE gene expression remained VPA responsive in THP-1 cells (Fig.6). We conclude that Rb has no demonstrable or significant role insilencing viral IE gene expression during the establishment of la-tency.
Viral DNA accumulates to lower levels in Rb knockdowncells. Since Rb does not appear to have a biological impact on the
FIG 4 Rb knockdown reduces the ability of HCMV to enter cells. (A) Serum-starved Scr or Rb2 cells were infected with AD169 at an MOI of 1, and equalamounts of protein from lysates harvested between 4 and 6 h postinfectionwere analyzed by Western blotting with the indicated antibodies. (B) The dataare mean values of tegument-delivered pp71 relative to tubulin and normal-ized to Scr � standard deviations from five biological replicates. Statisticalsignificance was determined by Student’s t test prior to normalization (*, P �0.02). (C and D) Serum-starved Scr or Rb cells were infected with AD169 at anMOI of 1 (C) or an MOI of 0.1 (D) and harvested at 6 h postinfection. Viral andcellular genomes were then quantitated by qPCR. The data are mean numbersof viral genomes relative to those of cellular genomes � standard deviationsfrom four biological replicates for each MOI. Statistical significance was deter-mined by Student’s t test prior to normalization (*, P � 0.04).
VanDeusen and Kalejta
5016 jvi.asm.org May 2015 Volume 89 Number 9Journal of Virology
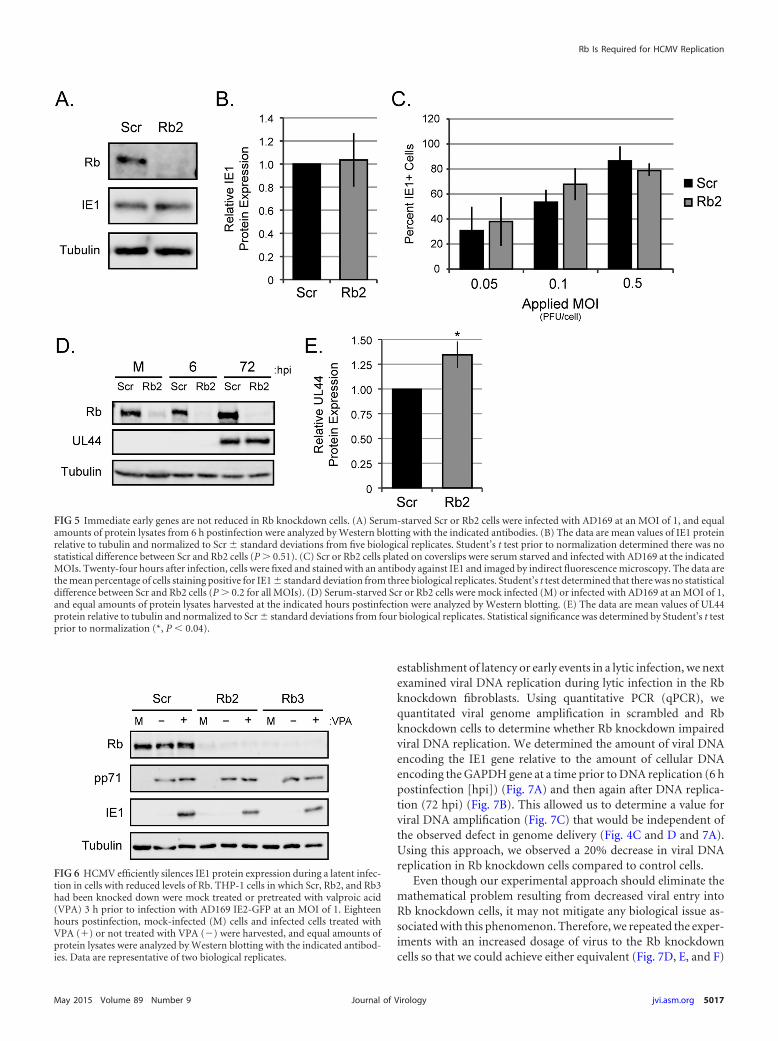
establishment of latency or early events in a lytic infection, we nextexamined viral DNA replication during lytic infection in the Rbknockdown fibroblasts. Using quantitative PCR (qPCR), wequantitated viral genome amplification in scrambled and Rbknockdown cells to determine whether Rb knockdown impairedviral DNA replication. We determined the amount of viral DNAencoding the IE1 gene relative to the amount of cellular DNAencoding the GAPDH gene at a time prior to DNA replication (6 hpostinfection [hpi]) (Fig. 7A) and then again after DNA replica-tion (72 hpi) (Fig. 7B). This allowed us to determine a value forviral DNA amplification (Fig. 7C) that would be independent ofthe observed defect in genome delivery (Fig. 4C and D and 7A).Using this approach, we observed a 20% decrease in viral DNAreplication in Rb knockdown cells compared to control cells.
Even though our experimental approach should eliminate themathematical problem resulting from decreased viral entry intoRb knockdown cells, it may not mitigate any biological issue as-sociated with this phenomenon. Therefore, we repeated the exper-iments with an increased dosage of virus to the Rb knockdowncells so that we could achieve either equivalent (Fig. 7D, E, and F)
FIG 5 Immediate early genes are not reduced in Rb knockdown cells. (A) Serum-starved Scr or Rb2 cells were infected with AD169 at an MOI of 1, and equalamounts of protein lysates from 6 h postinfection were analyzed by Western blotting with the indicated antibodies. (B) The data are mean values of IE1 proteinrelative to tubulin and normalized to Scr � standard deviations from five biological replicates. Student’s t test prior to normalization determined there was nostatistical difference between Scr and Rb2 cells (P 0.51). (C) Scr or Rb2 cells plated on coverslips were serum starved and infected with AD169 at the indicatedMOIs. Twenty-four hours after infection, cells were fixed and stained with an antibody against IE1 and imaged by indirect fluorescence microscopy. The data arethe mean percentage of cells staining positive for IE1 � standard deviation from three biological replicates. Student’s t test determined that there was no statisticaldifference between Scr and Rb2 cells (P 0.2 for all MOIs). (D) Serum-starved Scr or Rb2 cells were mock infected (M) or infected with AD169 at an MOI of 1,and equal amounts of protein lysates harvested at the indicated hours postinfection were analyzed by Western blotting. (E) The data are mean values of UL44protein relative to tubulin and normalized to Scr � standard deviations from four biological replicates. Statistical significance was determined by Student’s t testprior to normalization (*, P � 0.04).
FIG 6 HCMV efficiently silences IE1 protein expression during a latent infec-tion in cells with reduced levels of Rb. THP-1 cells in which Scr, Rb2, and Rb3had been knocked down were mock treated or pretreated with valproic acid(VPA) 3 h prior to infection with AD169 IE2-GFP at an MOI of 1. Eighteenhours postinfection, mock-infected (M) cells and infected cells treated withVPA (�) or not treated with VPA (�) were harvested, and equal amounts ofprotein lysates were analyzed by Western blotting with the indicated antibod-ies. Data are representative of two biological replicates.
Rb Is Required for HCMV Replication
May 2015 Volume 89 Number 9 jvi.asm.org 5017Journal of Virology
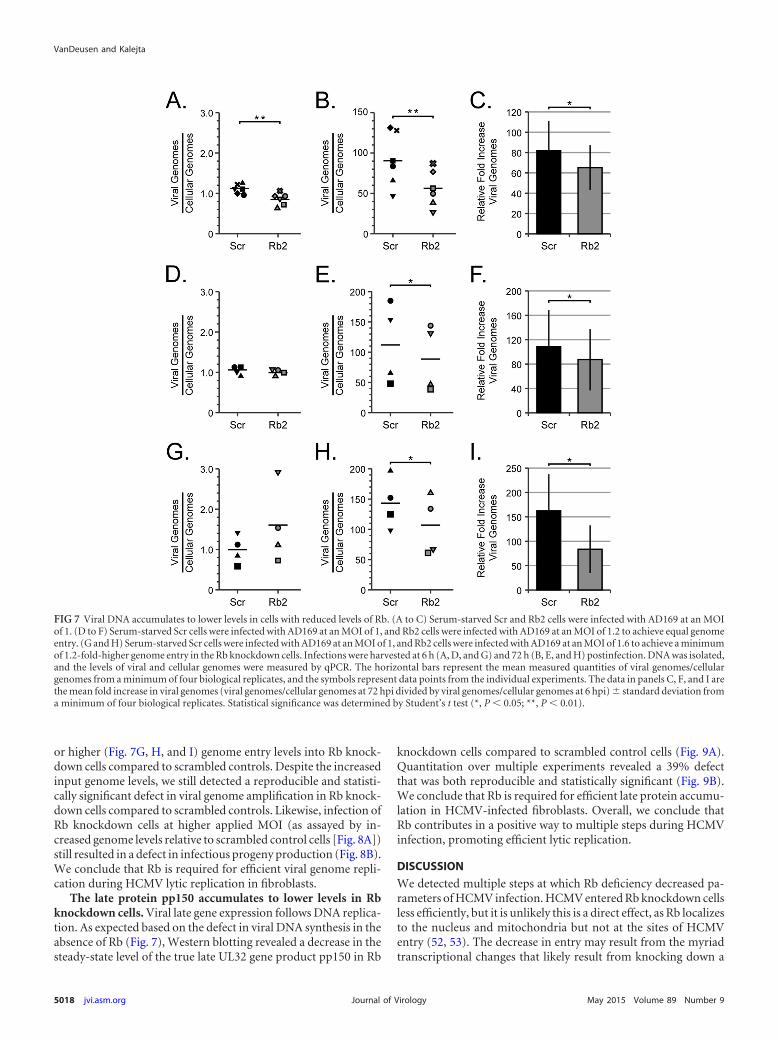
or higher (Fig. 7G, H, and I) genome entry levels into Rb knock-down cells compared to scrambled controls. Despite the increasedinput genome levels, we still detected a reproducible and statisti-cally significant defect in viral genome amplification in Rb knock-down cells compared to scrambled controls. Likewise, infection ofRb knockdown cells at higher applied MOI (as assayed by in-creased genome levels relative to scrambled control cells [Fig. 8A])still resulted in a defect in infectious progeny production (Fig. 8B).We conclude that Rb is required for efficient viral genome repli-cation during HCMV lytic replication in fibroblasts.
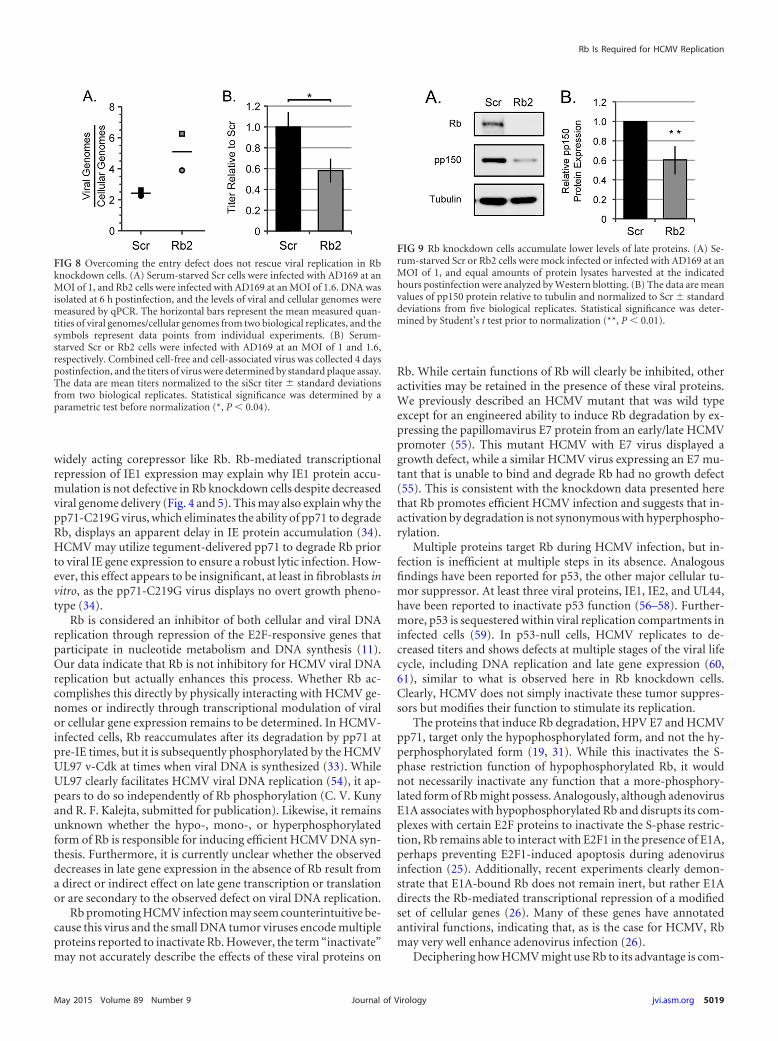
The late protein pp150 accumulates to lower levels in Rbknockdown cells. Viral late gene expression follows DNA replica-tion. As expected based on the defect in viral DNA synthesis in theabsence of Rb (Fig. 7), Western blotting revealed a decrease in thesteady-state level of the true late UL32 gene product pp150 in Rb
knockdown cells compared to scrambled control cells (Fig. 9A).Quantitation over multiple experiments revealed a 39% defectthat was both reproducible and statistically significant (Fig. 9B).We conclude that Rb is required for efficient late protein accumu-lation in HCMV-infected fibroblasts. Overall, we conclude thatRb contributes in a positive way to multiple steps during HCMVinfection, promoting efficient lytic replication.
DISCUSSION
We detected multiple steps at which Rb deficiency decreased pa-rameters of HCMV infection. HCMV entered Rb knockdown cellsless efficiently, but it is unlikely this is a direct effect, as Rb localizesto the nucleus and mitochondria but not at the sites of HCMVentry (52, 53). The decrease in entry may result from the myriadtranscriptional changes that likely result from knocking down a
FIG 7 Viral DNA accumulates to lower levels in cells with reduced levels of Rb. (A to C) Serum-starved Scr and Rb2 cells were infected with AD169 at an MOIof 1. (D to F) Serum-starved Scr cells were infected with AD169 at an MOI of 1, and Rb2 cells were infected with AD169 at an MOI of 1.2 to achieve equal genomeentry. (G and H) Serum-starved Scr cells were infected with AD169 at an MOI of 1, and Rb2 cells were infected with AD169 at an MOI of 1.6 to achieve a minimumof 1.2-fold-higher genome entry in the Rb knockdown cells. Infections were harvested at 6 h (A, D, and G) and 72 h (B, E, and H) postinfection. DNA was isolated,and the levels of viral and cellular genomes were measured by qPCR. The horizontal bars represent the mean measured quantities of viral genomes/cellulargenomes from a minimum of four biological replicates, and the symbols represent data points from the individual experiments. The data in panels C, F, and I arethe mean fold increase in viral genomes (viral genomes/cellular genomes at 72 hpi divided by viral genomes/cellular genomes at 6 hpi) � standard deviation froma minimum of four biological replicates. Statistical significance was determined by Student’s t test (*, P � 0.05; **, P � 0.01).
VanDeusen and Kalejta
5018 jvi.asm.org May 2015 Volume 89 Number 9Journal of Virology
widely acting corepressor like Rb. Rb-mediated transcriptionalrepression of IE1 expression may explain why IE1 protein accu-mulation is not defective in Rb knockdown cells despite decreasedviral genome delivery (Fig. 4 and 5). This may also explain why thepp71-C219G virus, which eliminates the ability of pp71 to degradeRb, displays an apparent delay in IE protein accumulation (34).HCMV may utilize tegument-delivered pp71 to degrade Rb priorto viral IE gene expression to ensure a robust lytic infection. How-ever, this effect appears to be insignificant, at least in fibroblasts invitro, as the pp71-C219G virus displays no overt growth pheno-type (34).
Rb is considered an inhibitor of both cellular and viral DNAreplication through repression of the E2F-responsive genes thatparticipate in nucleotide metabolism and DNA synthesis (11).Our data indicate that Rb is not inhibitory for HCMV viral DNAreplication but actually enhances this process. Whether Rb ac-complishes this directly by physically interacting with HCMV ge-nomes or indirectly through transcriptional modulation of viralor cellular gene expression remains to be determined. In HCMV-infected cells, Rb reaccumulates after its degradation by pp71 atpre-IE times, but it is subsequently phosphorylated by the HCMVUL97 v-Cdk at times when viral DNA is synthesized (33). WhileUL97 clearly facilitates HCMV viral DNA replication (54), it ap-pears to do so independently of Rb phosphorylation (C. V. Kunyand R. F. Kalejta, submitted for publication). Likewise, it remainsunknown whether the hypo-, mono-, or hyperphosphorylatedform of Rb is responsible for inducing efficient HCMV DNA syn-thesis. Furthermore, it is currently unclear whether the observeddecreases in late gene expression in the absence of Rb result froma direct or indirect effect on late gene transcription or translationor are secondary to the observed defect on viral DNA replication.
Rb promoting HCMV infection may seem counterintuitive be-cause this virus and the small DNA tumor viruses encode multipleproteins reported to inactivate Rb. However, the term “inactivate”may not accurately describe the effects of these viral proteins on
Rb. While certain functions of Rb will clearly be inhibited, otheractivities may be retained in the presence of these viral proteins.We previously described an HCMV mutant that was wild typeexcept for an engineered ability to induce Rb degradation by ex-pressing the papillomavirus E7 protein from an early/late HCMVpromoter (55). This mutant HCMV with E7 virus displayed agrowth defect, while a similar HCMV virus expressing an E7 mu-tant that is unable to bind and degrade Rb had no growth defect(55). This is consistent with the knockdown data presented herethat Rb promotes efficient HCMV infection and suggests that in-activation by degradation is not synonymous with hyperphospho-rylation.
Multiple proteins target Rb during HCMV infection, but in-fection is inefficient at multiple steps in its absence. Analogousfindings have been reported for p53, the other major cellular tu-mor suppressor. At least three viral proteins, IE1, IE2, and UL44,have been reported to inactivate p53 function (56–58). Further-more, p53 is sequestered within viral replication compartments ininfected cells (59). In p53-null cells, HCMV replicates to de-creased titers and shows defects at multiple stages of the viral lifecycle, including DNA replication and late gene expression (60,61), similar to what is observed here in Rb knockdown cells.Clearly, HCMV does not simply inactivate these tumor suppres-sors but modifies their function to stimulate its replication.
The proteins that induce Rb degradation, HPV E7 and HCMVpp71, target only the hypophosphorylated form, and not the hy-perphosphorylated form (19, 31). While this inactivates the S-phase restriction function of hypophosphorylated Rb, it wouldnot necessarily inactivate any function that a more-phosphory-lated form of Rb might possess. Analogously, although adenovirusE1A associates with hypophosphorylated Rb and disrupts its com-plexes with certain E2F proteins to inactivate the S-phase restric-tion, Rb remains able to interact with E2F1 in the presence of E1A,perhaps preventing E2F1-induced apoptosis during adenovirusinfection (25). Additionally, recent experiments clearly demon-strate that E1A-bound Rb does not remain inert, but rather E1Adirects the Rb-mediated transcriptional repression of a modifiedset of cellular genes (26). Many of these genes have annotatedantiviral functions, indicating that, as is the case for HCMV, Rbmay very well enhance adenovirus infection (26).
Deciphering how HCMV might use Rb to its advantage is com-
FIG 8 Overcoming the entry defect does not rescue viral replication in Rbknockdown cells. (A) Serum-starved Scr cells were infected with AD169 at anMOI of 1, and Rb2 cells were infected with AD169 at an MOI of 1.6. DNA wasisolated at 6 h postinfection, and the levels of viral and cellular genomes weremeasured by qPCR. The horizontal bars represent the mean measured quan-tities of viral genomes/cellular genomes from two biological replicates, and thesymbols represent data points from individual experiments. (B) Serum-starved Scr or Rb2 cells were infected with AD169 at an MOI of 1 and 1.6,respectively. Combined cell-free and cell-associated virus was collected 4 dayspostinfection, and the titers of virus were determined by standard plaque assay.The data are mean titers normalized to the siScr titer � standard deviationsfrom two biological replicates. Statistical significance was determined by aparametric test before normalization (*, P � 0.04).
FIG 9 Rb knockdown cells accumulate lower levels of late proteins. (A) Se-rum-starved Scr or Rb2 cells were mock infected or infected with AD169 at anMOI of 1, and equal amounts of protein lysates harvested at the indicatedhours postinfection were analyzed by Western blotting. (B) The data are meanvalues of pp150 protein relative to tubulin and normalized to Scr � standarddeviations from five biological replicates. Statistical significance was deter-mined by Student’s t test prior to normalization (**, P � 0.01).
Rb Is Required for HCMV Replication
May 2015 Volume 89 Number 9 jvi.asm.org 5019Journal of Virology

plicated because at least four viral proteins (pp71, IE1, IE2, andUL97) have been demonstrated to physically and/or functionallyinteract with the Rb/E2F pathway (31–33, 37–39). Furthermore,an additional 14 HCMV proteins (US16, US22, US26, US29,UL20, UL23, UL24, UL25, UL29, UL69, UL77, and UL93 in allstrains and UL111a and UL128 in clinical strains) encode a canon-ical Rb-binding LXCX(D/E) motif (62). While any one or more ofthese proteins may be a functional partner with Rb, none areknown to play direct roles in the processes shown here to be defi-cient in the absence of Rb (viral DNA replication and late geneexpression), although genetic ablation of UL69 shows a similarphenotype (63). UL77 and UL93 play roles in DNA encapsidation(64, 65), provocatively suggesting that Rb may participate in an-other viral process critical for HCMV replication not directly as-sayed here. No doubt, the interaction between HCMV and Rb ismultifaceted and complex.
The vast majority of research on the interactions between vi-ruses and Rb has focused on oncogenic transformation events(66). Such explorations have been instrumental in defining theRb-E2F pathway and its role in both normal cell cycle progressionand cancer. However, oncogenicity is not a central part of theinfectious cycle of these viruses, and the actual role of viral target-ing of Rb remains, for the most part, undefined. Thus, despiteintensive study using viral Rb-inactivating proteins to illuminatethe contribution of the Rb pathway to tumor suppression andoncogenesis, we still have a rudimentary comprehension of therole of Rb in productive viral replication. Understanding how vi-ruses manipulate the function of Rb to their benefit may revealnovel functions of this fascinating tumor suppressor.
ACKNOWLEDGMENTS
We thank Phil Balandyk for expert technical assistance, Paul Lambert andHiroyuki Sakai for shRNA retroviral plasmids, Nathan Sherer for sharingequipment, and Emily Albright for comments on the manuscript.
This work was supported by National Institutes of Health grantAI080675 to R.F.K. H.R.V. was supported by training grant T32CA009135.
REFERENCES1. Viatour P, Sage J. 2011. Newly identified aspects of tumor suppression by
RB. Dis Model Mech 4:581–585. http://dx.doi.org/10.1242/dmm.008060.2. Di Fiore R, D’Anneo A, Tesoriere G, Vento R. 2013. RB1 in cancer:
different mechanisms of RB1 inactivation and alterations of pRb pathwayin tumorigenesis. J Cell Physiol 228:1676 –1687. http://dx.doi.org/10.1002/jcp.24329.
3. Horowitz JM, Park SH, Bogenmann E, Cheng JC, Yandell DW, Kaye FJ,Minna JD, Dryja TP, Weinberg RA. 1990. Frequent inactivation of theretinoblastoma anti-oncogene is restricted to a subset of human tumorcells. Proc Natl Acad Sci U S A 87:2775–2779. http://dx.doi.org/10.1073/pnas.87.7.2775.
4. Morris EJ, Dyson NJ. 2001. Retinoblastoma protein partners. Adv Can-cer Res 82:1–54. http://dx.doi.org/10.1016/S0065-230X(01)82001-7.
5. Dick FA, Rubin SM. 2013. Molecular mechanisms underlying RB proteinfunction. Nat Rev Mol Cell Biol 14:297–306. http://dx.doi.org/10.1038/nrm3567.
6. Nicolay BN, Dyson NJ. 2013. The multiple connections between pRB andcell metabolism. Curr Opin Cell Biol 25:735–740. http://dx.doi.org/10.1016/j.ceb.2013.07.012.
7. Hanahan D, Weinberg RA. 2011. Hallmarks of cancer: the next genera-tion. Cell 144:646 – 674. http://dx.doi.org/10.1016/j.cell.2011.02.013.
8. Sherr CJ, McCormick F. 2002. The RB and p53 pathways in cancer. CancerCell 2:103–112. http://dx.doi.org/10.1016/S1535-6108(02)00102-2.
9. Rubin SM. 2013. Deciphering the retinoblastoma protein phosphoryla-tion code. Trends Biochem Sci 38:12–19. http://dx.doi.org/10.1016/j.tibs.2012.10.007.
10. Harbour JW, Luo RX, Dei Santi A, Postigo AA, Dean DC. 1999. Cdkphosphorylation triggers sequential intramolecular interactions that pro-gressively block Rb functions as cells move through G1. Cell 98:859 – 869.http://dx.doi.org/10.1016/S0092-8674(00)81519-6.
11. Trimarchi JM, Lees JA. 2002. Sibling rivalry in the E2F family. Nat RevMol Cell Biol 3:11–20. http://dx.doi.org/10.1038/nrm714.
12. Johnson DG. 2000. The paradox of E2F1: oncogene and tumor suppressorgene. Mol Carcinog 27:151–157. http://dx.doi.org/10.1002/(SICI)1098-2744(200003)27:3�151::AID-MC13.0.CO;2-C.
13. Johnson DG, Cress WD, Jakoi L, Nevins JR. 1994. Oncogenic capacity ofthe E2F1 gene. Proc Natl Acad Sci U S A 91:12823–12827. http://dx.doi.org/10.1073/pnas.91.26.12823.
14. Burke JR, Hura GL, Rubin SM. 2012. Structures of inactive retinoblas-toma protein reveal multiple mechanisms for cell cycle control. Genes Dev26:1156 –1166. http://dx.doi.org/10.1101/gad.189837.112.
15. Barrientes S, Cooke C, Goodrich DW. 2000. Glutamic acid mutagenesis ofretinoblastoma protein phosphorylation sites has diverse effects on function.Oncogene 19:562–570. http://dx.doi.org/10.1038/sj.onc.1203332.
16. Narasimha AM, Kaulich M, Shapiro GS, Choi YJ, Sicinski P, Dowdy SF.2014. Cyclin D activates the Rb tumor suppressor by mono-phosphorylation.eLife 3:e02872. http://dx.doi.org/10.7554/eLife.02872.
17. Macdonald JI, Dick FA. 2012. Posttranslational modifications of theretinoblastoma tumor suppressor protein as determinants of function.Genes Cancer 3:619 – 633. http://dx.doi.org/10.1177/1947601912473305.
18. Huh K, Zhou X, Hayakawa H, Cho J-Y, Libermann TA, Jin J, HarperJW, Munger K. 2007. Human papillomavirus type 16 E7 oncoproteinassociates with the cullin 2 ubiquitin ligase complex, which contributes todegradation of the retinoblastoma tumor suppressor. J Virol 81:9737–9747. http://dx.doi.org/10.1128/JVI.00881-07.
19. Boyer SN, Wazer DE, Band V. 1996. E7 protein of human papillomavirus-16 induces degradation of retinoblastoma protein through the ubiq-uitin-proteasome pathway. Cancer Res 56:4620 – 4624.
20. Collins AS, Nakahara T, Do A, Lambert PF. 2005. Interactions withpocket proteins contribute to the role of human papillomavirus type 16 E7in the papillomavirus life cycle. J Virol 79:14769 –14780. http://dx.doi.org/10.1128/JVI.79.23.14769-14780.2005.
21. Bodily JM, Mehta KPM, Cruz L, Meyers C, Laimins LA. 2011. The E7open reading frame acts in cis and in trans to mediate differentiation-dependent activities in the human papillomavirus type 16 life cycle. J Virol85:8852– 8862. http://dx.doi.org/10.1128/JVI.00664-11.
22. Roman A, Munger K. 2013. The papillomavirus E7 proteins. Virology445:138 –168. http://dx.doi.org/10.1016/j.virol.2013.04.013.
23. Bagchi S, Raychaudhuri P, Nevins JR. 1990. Adenovirus E1A proteinscan dissociate heteromeric complexes involving the E2F transcription fac-tor: a novel mechanism for E1A trans-activation. Cell 62:659 – 669. http://dx.doi.org/10.1016/0092-8674(90)90112-R.
24. Sauthoff H, Pipiya T, Heitner S, Chen S, Bleck B, Reibman J, Chang W,Norman RG, Rom WN, Hay JG. 2004. Impact of E1a modifications ontumor-selective adenoviral replication and toxicity. Mol Ther 10:749 –757. http://dx.doi.org/10.1016/j.ymthe.2004.07.014.
25. Seifried LA, Talluri S, Cecchini M, Julian LM, Mymryk JS, Dick FA.2008. pRB-E2F1 complexes are resistant to adenovirus E1A-mediated dis-ruption. J Virol 82:4511– 4520. http://dx.doi.org/10.1128/JVI.02713-07.
26. Ferrari R, Gou D, Jawdekar G, Johnson SA, Nava M, Su T, Yousef AF,Zemke NR, Pellegrini M, Kurdistani SK, Berk AJ. 2014. Adenovirussmall E1A employs the lysine acetylases p300/CBP and tumor suppressorRb to repress select host genes and promote productive virus infection.Cell Host Microbe 16:663– 676. http://dx.doi.org/10.1016/j.chom.2014.10.004.
27. Cinatl J, Scholz M, Kotchetkov R, Vogel J-U, Doerr HW. 2004. Molec-ular mechanisms of the modulatory effects of HCMV infection in tumorcell biology. Trends Mol Med 10:19 –23. http://dx.doi.org/10.1016/j.molmed.2003.11.002.
28. Soroceanu L, Cobbs CS. 2011. Is HCMV a tumor promoter? Virus Res157:193–203. http://dx.doi.org/10.1016/j.virusres.2010.10.026.
29. Barami K. 2010. Oncomodulatory mechanisms of human cytomegalovi-rus in gliomas. J Clin Neurosci 17:819 – 823. http://dx.doi.org/10.1016/j.jocn.2009.10.040.
30. Ranganathan P, Clark PA, Kuo JS, Salamat MS, Kalejta RF. 2012.Significant association of multiple human cytomegalovirus genomic lociwith glioblastoma multiforme samples. J Virol 86:854 – 864. http://dx.doi.org/10.1128/JVI.06097-11.
31. Kalejta RF, Bechtel JT, Shenk T. 2003. Human cytomegalovirus pp71
VanDeusen and Kalejta
5020 jvi.asm.org May 2015 Volume 89 Number 9Journal of Virology

stimulates cell cycle progression by inducing the proteasome-dependent degradation of the retinoblastoma family of tumor suppres-sors. Mol Cell Biol 23:1885–1895. http://dx.doi.org/10.1128/MCB.23.6.1885-1895.2003.
32. Kalejta RF, Shenk T. 2003. Proteasome-dependent, ubiquitin-independent degradation of the Rb family of tumor suppressors by thehuman cytomegalovirus pp71 protein. Proc Natl Acad Sci U S A 100:3263–3268. http://dx.doi.org/10.1073/pnas.0538058100.
33. Hume AJ, Finkel JS, Kamil JP, Coen DM, Culbertson MR, Kalejta RF.2008. Phosphorylation of retinoblastoma protein by viral protein withcyclin-dependent kinase function. Science 320:797–799. http://dx.doi.org/10.1126/science.1152095.
34. Cantrell SR, Bresnahan WA. 2005. Interaction between the human cyto-megalovirus UL82 gene product (pp71) and hDaxx regulates immediate-early gene expression and viral replication. J Virol 79:7792–7802. http://dx.doi.org/10.1128/JVI.79.12.7792-7802.2005.
35. Prichard MN, Sztul E, Daily SL, Perry AL, Frederick SL, Gill RB,Hartline CB, Streblow DN, Varnum SM, Smith RD, Kern ER. 2008.Human cytomegalovirus UL97 kinase activity is required for the hyper-phosphorylation of retinoblastoma protein and inhibits the formation ofnuclear aggresomes. J Virol 82:5054 –5067. http://dx.doi.org/10.1128/JVI.02174-07.
36. Jacob T, Van den Broeke C, Favoreel HW. 2011. Viral serine/threonineprotein kinases. J Virol 85:1158 –1173. http://dx.doi.org/10.1128/JVI.01369-10.
37. Poma EE, Kowalik TF, Zhu L, Sinclair JH, Huang ES. 1996. The humancytomegalovirus IE1-72 protein interacts with the cellular p107 proteinand relieves p107-mediated transcriptional repression of an E2F-responsive promoter. J Virol 70:7867–7877.
38. Wiebusch L, Asmar J, Uecker R, Hagemeier C. 2003. Human cytomeg-alovirus immediate-early protein 2 (IE2)-mediated activation of cyclin Eis cell-cycle-independent and forces S-phase entry in IE2-arrested cells. JGen Virol 84:51– 60. http://dx.doi.org/10.1099/vir.0.18702-0.
39. Song Y-J, Stinski MF. 2002. Effect of the human cytomegalovirus IE86protein on expression of E2F-responsive genes: a DNA microarray analy-sis. Proc Natl Acad Sci U S A 99:2836 –2841. http://dx.doi.org/10.1073/pnas.052010099.
40. Saffert RT, Kalejta RF. 2007. Human cytomegalovirus gene expression issilenced by Daxx-mediated intrinsic immune defense in model latent in-fections established in vitro. J Virol 81:9109 –9120. http://dx.doi.org/10.1128/JVI.00827-07.
41. Sanchez V, Clark CL, Yen JY, Dwarakanath R, Spector DH. 2002. Viablehuman cytomegalovirus recombinant virus with an internal deletion ofthe IE2 86 gene affects late stages of viral replication. J Virol 76:2973–2989.http://dx.doi.org/10.1128/JVI.76.6.2973-2989.2002.
42. Murphy E, Yu D, Grimwood J, Schmutz J, Dickson M, Jarvis MA, HahnG, Nelson JA, Myers RM, Shenk TE. 2003. Coding potential of labora-tory and clinical strains of human cytomegalovirus. Proc Natl Acad SciU S A 100:14976 –14981. http://dx.doi.org/10.1073/pnas.2136652100.
43. Pasque V, Gillich A, Garrett N, Gurdon JB. 2011. Histone variantmacroH2A confers resistance to nuclear reprogramming. EMBO J 30:2373–2387. http://dx.doi.org/10.1038/emboj.2011.144.
44. Ueno T, Sasaki K, Yoshida S, Kajitani N, Satsuka A, Nakamura H, SakaiH. 2006. Molecular mechanisms of hyperplasia induction by human pap-illomavirus E7. Oncogene 25:4155– 4164. http://dx.doi.org/10.1038/sj.onc.1209445.
45. Nowak B, Sullivan C, Sarnow P, Thomas R, Bricout F, Nicolas JC,Fleckenstein B, Levine AJ. 1984. Characterization of monoclonal anti-bodies and polyclonal immune sera directed against human cytomegalo-virus virion proteins. Virology 132:325–338. http://dx.doi.org/10.1016/0042-6822(84)90039-4.
46. Saffert RT, Kalejta RF. 2006. Inactivating a cellular intrinsic immunedefense mediated by Daxx is the mechanism through which the humancytomegalovirus pp71 protein stimulates viral immediate-early gene ex-pression. J Virol 80:3863–3871. http://dx.doi.org/10.1128/JVI.80.8.3863-3871.2006.
47. Hwang J, Saffert RT, Kalejta RF. 2011. Elongin B-mediated epigeneticalteration of viral chromatin correlates with efficient human cytomegalo-virus gene expression and replication. mBio 2(2):e00023-11. http://dx.doi.org/10.1128/mBio.00023-11.
48. Juckem LK, Boehme KW, Feire AL, Compton T. 2008. Differentialinitiation of innate immune responses induced by human cytomegalovi-
rus entry into fibroblast cells. J Immunol 180:4965– 4977. http://dx.doi.org/10.4049/jimmunol.180.7.4965.
49. Salvant BS, Fortunato EA, Spector DH. 1998. Cell cycle dysregulation byhuman cytomegalovirus: influence of the cell cycle phase at the time ofinfection and effects on cyclin transcription. J Virol 72:3729 –3741.
50. Sage J, Miller AL, Pérez-Mancera PA, Wysocki JM, Jacks T. 2003. Acutemutation of retinoblastoma gene function is sufficient for cell cycle re-entry. Nature 424:223–228. http://dx.doi.org/10.1038/nature01764.
51. Saffert RT, Penkert RR, Kalejta RF. 2010. Cellular and viral controlover the initial events of human cytomegalovirus experimental latencyin CD34� cells. J Virol 84:5594 –5604. http://dx.doi.org/10.1128/JVI.00348-10.
52. Hilgendorf KI, Leshchiner ES, Nedelcu S, Maynard MA, Calo E, IanariA, Walensky LD, Lees JA. 2013. The retinoblastoma protein inducesapoptosis directly at the mitochondria. Genes Dev 27:1003–1015. http://dx.doi.org/10.1101/gad.211326.112.
53. Antonucci LA, Egger JV, Krucher NA. 2014. Phosphorylation of theRetinoblastoma protein (Rb) on serine-807 is required for associationwith Bax. Cell Cycle 13:3611–3617. http://dx.doi.org/10.4161/15384101.2014.964093.
54. Wolf DG, Courcelle CT, Prichard MN, Mocarski ES. 2001. Distinct andseparate roles for herpesvirus-conserved UL97 kinase in cytomegalovirusDNA synthesis and encapsidation. Proc Natl Acad Sci U S A 98:1895–1900. http://dx.doi.org/10.1073/pnas.98.4.1895.
55. Kamil JP, Hume AJ, Jurak I, Münger K, Kalejta RF, Coen DM. 2009.Human papillomavirus 16 E7 inactivator of retinoblastoma family pro-teins complements human cytomegalovirus lacking UL97 protein kinase.Proc Natl Acad Sci U S A 106:16823–16828. http://dx.doi.org/10.1073/pnas.0901521106.
56. Hwang E-S, Zhang Z, Cai H, Huang DY, Huong S-M, Cha C-Y, HuangE-S. 2009. Human cytomegalovirus IE1-72 protein interacts with p53 andinhibits p53-dependent transactivation by a mechanism different fromthat of IE2-86 protein. J Virol 83:12388 –12398. http://dx.doi.org/10.1128/JVI.00304-09.
57. Speir E, Modali R, Huang ES, Leon MB, Shawl F, Finkel T, Epstein SE.1994. Potential role of human cytomegalovirus and p53 interaction incoronary restenosis. Science 265:391–394. http://dx.doi.org/10.1126/science.8023160.
58. Kwon Y, Kim M-N, Young Choi E, Heon Kim J, Hwang E-S, Cha C-Y.2012. Inhibition of p53 transcriptional activity by human cytomegalovirusUL44. Microbiol Immunol 56:324 –331. http://dx.doi.org/10.1111/j.1348-0421.2012.00446.x.
59. Fortunato EA, Spector DH. 1998. p53 and RPA are sequestered in viralreplication centers in the nuclei of cells infected with human cytomegalo-virus. J Virol 72:2033–2039.
60. Casavant NC, Luo MH, Rosenke K, Winegardner T, Zurawska A,Fortunato EA. 2006. Potential role for p53 in the permissive life cycle ofhuman cytomegalovirus. J Virol 80:8390 – 8401. http://dx.doi.org/10.1128/JVI.00505-06.
61. Hannemann H, Rosenke K, O’Dowd JM, Fortunato EA. 2009. Thepresence of p53 influences the expression of multiple human cytomega-lovirus genes at early times postinfection. J Virol 83:4316 – 4325. http://dx.doi.org/10.1128/JVI.02075-08.
62. Chellappan S, Kraus VB, Kroger B, Munger K, Howley PM, Phelps WC,Nevins JR. 1992. Adenovirus E1A, simian virus 40 tumor antigen, andhuman papillomavirus E7 protein share the capacity to disrupt the inter-action between transcription factor E2F and the retinoblastoma geneproduct. Proc Natl Acad Sci U S A 89:4549 – 4553. http://dx.doi.org/10.1073/pnas.89.10.4549.
63. Hayashi ML, Blankenship C, Shenk T. 2000. Human cytomegalovirusUL69 protein is required for efficient accumulation of infected cells in theG1 phase of the cell cycle. Proc Natl Acad Sci U S A 97:2692–2696. http://dx.doi.org/10.1073/pnas.050587597.
64. Toropova K, Huffman JB, Homa FL, Conway JF. 2011. The herpessimplex virus 1 UL17 protein is the second constituent of the capsid ver-tex-specific component required for DNA packaging and retention. J Vi-rol 85:7513–7522. http://dx.doi.org/10.1128/JVI.00837-11.
65. Tandon R, Mocarski ES. 2012. Viral and host control of cytomegalovirusmaturation. Trends Microbiol 20:392– 401. http://dx.doi.org/10.1016/j.tim.2012.04.008.
66. Bellacchio E, Paggi MG. 2013. Understanding the targeting of the RBfamily proteins by viral oncoproteins to defeat their oncogenic machinery.J Cell Physiol 228:285–291. http://dx.doi.org/10.1002/jcp.24137.
Rb Is Required for HCMV Replication
May 2015 Volume 89 Number 9 jvi.asm.org 5021Journal of Virology



















![The Retinoblastoma (RB) Tumor Suppressor: Pushing Back ... · The retinoblastoma (RB) tumor suppressor plays an important role in cell cycle progression [1–3]. The function of RB](https://img.pdfslide.us/doc/110x75/5e4d9b57a62227248b3387f2/the-retinoblastoma-rb-tumor-suppressor-pushing-back-the-retinoblastoma-rb.jpg)









